БИОХИМИЯ, 2021, том 86, вып. 2, с. 274 - 287
УДК 578.32;578.865;539.26
СТРУКТУРА ВИРИОНОВ A ВИРУСА КАРТОФЕЛЯ ПО ДАННЫМ
МАЛОУГЛОВОГО РЕНТГЕНОВСКОГО РАССЕЯНИЯ
И КОМПЛЕМЕНТАРНЫХ МЕТОДОВ*
© 2021
Э.В. Штыкова1, М.В. Петухов1, Н.В. Федорова2, А.М. Арутюнян2,
Е.В. Скурат3, Л.В. Кордюкова2, А.В. Моисеенко3, А.Л. Ксенофонтов2**
1 Институт кристаллографии им. А.В. Шубникова Федерального научно%исследовательского центра
«Кристаллография и фотоника» Российской академии наук, 119333 Москва, Россия;
электронная почта: shtykova@ns.crys.ras.ru
2 НИИ физико%химической биологии им. А.Н. Белозерского, Московский государственный университет
им. М.В. Ломоносова, 119991 Москва, Россия; электронная почта: ksenofon@belozersky.msu.ru
3 Московский государственный университет им. М.В. Ломоносова,
биологический факультет, 119991 Москва, Россия
Поступила в редакцию 26.08.2020
После доработки 27.09.2020
Принята к публикации 27.09.2020
Белки оболочки вирионов A вируса картофеля содержат на своей поверхности частично неупорядоченные
N концевые домены, структурные и функциональные характеристики которых важны для понимания ме
ханизма инфицирования растений этим вирусом. В настоящей работе исследованы свойства и строение ин
тактных A вирусов картофеля и частично деградированных трипсином вирионов (AВКΔ32) с помощью ма
лоуглового рентгеновского рассеяния и комплементарных методов. Было показано, что при удалении 32
N концевых аминокислот белка оболочки вирион не разрушается и остается компактным, но изменяется
шаг спиральной упаковки его белковой оболочки. Для определения природы этих изменений по данным
малоуглового рентгеновского рассеяния, было проведено ab initio, в том числе многофазное, моделирование
геометрическими телами (спиралями) и восстановление структуры A вируса картофеля в растворе с ис
пользованием доступных структур атомного разрешения, схожего с белками оболочки A вируса картофе
ля - белка потивируса Y вируса картофеля. В результате впервые была получена структура низкого разре
шения нитевидного A вируса картофеля, интактного и частично деградированного, в условиях, близких к
естественным. Дополнительные исследования показали, что спектры кругового дихроизма в дальнем УФ
образцов A вируса картофеля и AВКΔ32 значительно различались по амплитуде и по положению основно
го отрицательного максимума. Степень термической денатурации данных образцов в диапазоне температур
20-55 °C также была различной. Данные просвечивающей электронной микроскопии показали, что вири
оны AВКΔ32 представляли собой преимущественно палочковидные частицы, в отличие от гибких, харак
терных для интактного вируса, что хорошо коррелирует с результатами малоуглового рассеяния. В целом,
проведенный структурный анализ свидетельствует о значении N концевых доменов белка оболочки для вы
полнения жизненно важных функций A вируса картофеля, что следует учитывать для выработки стратегии
по борьбе с этими растительными патогенами.
КЛЮЧЕВЫЕ СЛОВА: потивирусы, белок оболочки, A вирус картофеля, N концевые неупорядоченные до
мены, малоугловое рентгеновское рассеяние, структурное моделирование.
DOI: 10.31857/S0320972521020111
ВВЕДЕНИЕ
частиц 700 нм и диаметром 11-14 нм. С по
мощью различных предсказательных и экспери
Потивирусы представляют собой экономи
ментальных подходов было обнаружено, что бе
чески важную группу вирусов растений, прино
лок оболочки (БО) A вируса картофеля (AВК),
сящих значительный урон сельскому хозяйству.
типичного представителя потивирусов, имел
Гибкие нитевидные вирионы потивирусов име
значительное содержание неупорядоченных
ют спиральную симметрию со средней длиной
участков [1]. Позднее методом криоэлектрон
Принятые сокращения: AВК - A вирус картофеля; БО - белок оболочки; МУРР - малоугловое рентгеновское рас
сеяние; ПЭМ - просвечивающая электронная микроскопия.
biokhimiya, в рубрике «Papers in Press», BM20 293, 11.01.2021.
** Адресат для корреспонденции.
274
СТРУКТУРА ВИРИОНОВ А ВИРУСА КАРТОФЕЛЯ
275
ной микроскопии это было прямо подтвержде
риона вируса табачной мозаики (ВТМ) и ста
но для других представителей потивирусов: ви
бильных реполимеров БО ВТМ, так называе
руса мозаики арбуза (ВМА) [2], Y вируса карто
мых «стопок дисков» [14]. В настоящей работе
феля (YВК) [3] и вируса мозаики турнепса
обнаруженные нами различия в структурных
(ВМТу) [4]. Было показано, что неупорядочен
характеристиках интактных AВК и частично
ные фрагменты N концевых доменов БО поти
деградированных вирионов AВКΔ32 были соот
вирусов расположены на поверхности вирионов
несены с найденными различиями их оптичес
и взаимодействуют с соседними субъединица
ких свойств. Полученные результаты свиде
ми. В настоящее время общепринята концеп
тельствуют не только о значении N концевых
ция, что неструктурированные области белко
доменов БО для выполнения жизненно важных
вой молекулы играют ключевую роль в различ
функций AВК, но могут также способствовать
ных биохимических процессах [5]. Ранее мы
выработке стратегии по борьбе с вирусными за
наблюдали участие N доменов белка AВК в про
болеваниями растений, поскольку разупорядо
цессе сборки вирусоподобных частиц (ВПЧ):
ченная N концевая область белка оболочки яв
было показано, что субъединицы этого белка не
ляется возможной мишенью для дезактивации
существуют в индивидуальном виде в растворе,
вируса.
а часть его неупорядоченных сегментов транс
формируется в β структуры [6, 7]. Мягкая трип
синовая обработка интактных вирионов поти
МАТЕРИАЛЫ И МЕТОДЫ
вирусов приводит к образованию устойчивых
деградированных частиц, лишенных N конце
Получение препаратов интактных и дегради
вого домена [8]. Для прояснения роли N доме
рованных вирионов AВК. Изолят В11 вириона
нов потивирусов в ряде работ изучали процесс
AВК очищен, как описано ранее [8]. Коротко,
сборки ВПЧ из укороченных мутантных БО.
молодые растения N. benthamiana были зараже
Обнаружили, что для некоторых представителей
ны препаратом вируса AВК. На 18-21 день пос
потивирусов удаление 20-50 N концевых остат
ле заражения листья с вирусными симптомами
ков препятствовало процессу сборки ВПЧ [9,
были гомогенизированы в блендере Waring в
10], тогда как для белка YВК этого не происхо
трех объемах 50 мМ фосфатного буфера pH 7,8 с
дит [3]. В то же время у БО вируса гравировки
добавлением 0,01% ного β меркаптоэтанола.
табака (ВГТ) сборка нарушалась лишь при уда
После низкоскоростного центрифугирования
лении 120 аминокислотных остатков [11]. Пока
(НСЦ) (8 000 g, 8 °C, 20 мин) супернатант пере
зана также роль N доменов потивирусов в
мешивали в течение 1 ч при 4 °C с Тритоном
межклеточном транспорте вирионов и в процес
Х 100 (1% по объему); после НСЦ к суперна
се переноса их тлями [10].
танту добавляли 1/5 объема хлороформа. Вод
В настоящее время отсутствует структура
ную фазу отделяли НСЦ в тех же условиях. К
AВК высокого разрешения. В данной работе мы
смеси добавляли полиэтиленгликоль 6000 до
впервые детально охарактеризовали структуру
5% с добавлением 1,2% NaCl, смесь перемеши
вириона AВК, выполнив моделирование интакт
вали при температуре 4 °C в течение 1,5 ч. Оса
ных и деградированных трипсином вирионов
док осаждали с помощью НСЦ (12 000 g, 8 °C,
по данным синхротронного малоуглового рент
20 мин). Осадок, содержащий вирус, растворя
геновского рассеяния (МУРР). Этот структур
ли в 50 мМ фосфатном буфере pH 7,8 с добавле
ный метод с разрешением порядка 1-2 нм, то
нием 0,1 мМ фенилметилсульфонил фторида.
есть метод низкого разрешения, используется
Далее препарат подвергали высокоскоростному
для изучения строения биологических макро
центрифугированию на ультрацентрифуге
молекул в растворе, в условиях близких к естест
(100 000 g, 5 °C, 1,5 ч). Осадок вируса ресуспен
венным [12]. В последние десятилетия МУРР
дировали в 50 мМ фосфатном буфере при
активно развивался благодаря появлению но
pH 7,8.
вых методов анализа и интерпретации данных
Для получения деградированных вирионов
малоуглового рассеяния и компьютерного мо
AВКΔ32 препарат интактного вируса обрабаты
делирования структуры [13]. Ранее нами с по
вали трипсином в соотношении фермент/
мощью МУРР были охарактеризованы вирусо
субстрат 1 : 500, 15 мин инкубировали при ком
подобные частицы AВК, собранные из выде
натной температуре, для остановки реакции до
ленного свободного белка, и изучена их струк
бавляли фенилметилсульфонил фторид до кон
тура и диссоциация в различных водно солевых
центрации 1 мМ. Контроль чистоты образцов
растворах [7, 8]. Недавно данным методом мы
осуществляли с помощью электрофореза с доде
исследовали и определили структуры с низким
цилсульфатом натрия (SDS) по методу Лэммли
разрешением палочковидного спирального ви
[15], используя 15% ный полиакриламидный
БИОХИМИЯ том 86 вып. 2 2021
9*
276
ШТЫКОВА и др.
гель (ПААГ) в Mini PROTEAN 3 Cell («Bio
на синхротроне Petra III («DESY», Германия) на
Rad», США). Гели окрашивали 0,22% ным Ку
линии P12. Эта линия оснащена оборудованием
масси G 250 («Serva», Германия).
для автоматической смены образцов и двумер
Определение концентрации препаратов. Кон
ным детектором Pilatus 2M («DECTRIS», Швей
центрацию определяли на спектрофотометре
цария). Интенсивность рассеяния I(s) была из
UV 2600 («Hitachi», Япония), используя спектры
мерена в области значений волновых векторов
поглощения в УФ диапазоне 240-340 нм. В свя
0,08 < s < 7 нм-1, где s = (4πsinθ)/λ - вектор рас
зи с большим вкладом рассеяния в поглощение
сеяния, 2θ - угол рассеяния, λ = 0,124 нм - дли
вирионов (30-40%), истинные спектры погло
на волны излучения [17]. Для каждого образца
щения (E) суспензий частиц вычисляли по мето
было снято по 50 экспериментальных кривых
ду экстраполяции [16] в области 320-338 нм. Ко
рассеяния для контроля возможных радиацион
эффициенты поглощения препаратов вируса
ных повреждений. Радиационного повреждения
принимали равными E0
1% = 2,4.
обнаружено не было.
Измерение спектров кругового дихроизма.
Измерения проводили в 15 мМ фосфатном
Спектры кругового дихроизма (КД) измеряли в
буфере pH 7,8 с 0,05 М NaCl при температуре
5 мМ фосфатном буфере pH 7,8 при 20 °C в
10 °C при нескольких различных концентрациях
1-2
мм кюветах на дихрографе Chirascan
в диапазоне 2,0-5,0 мг/мл для учета возможной
(«Applied Photophysics», Англия). Концентрация
концентрационной зависимости. Зависимость
образцов составляла 50-100 мкг/мл. Спектры
кривых МУРР от концентрации измеряемых об
КД записывали на скорости 0,5-1,0 нм/с в диа
разцов не наблюдалась.
пазоне 185-250 нм с вычетом базовой линии.
Первичная обработка кривых рассеяния бы
Измеренные спектры обрабатывали с использо
ла проведена с использованием программы
ванием стандартного пакета программного обес
PRIMUS [18]. С помощью этой программы бы
печения, поставляемого с прибором. Величины
ло проведено усреднение экспериментальных
КД выражали в величинах молярной эллиптич
кривых рассеяния от образцов и соответствую
ности [Θ] ([Θ] = 3 300 × Δε). Δε - Молярный ко
щего буфера, с последующим вычитанием рас
эффициент дихроичного поглощения для препа
сеяния буфера из кривых образцов МУРР. Далее
ратов белка, рассчитывали на моль аминокис
для структурного анализа и моделирования ис
лотных остатков. Расчет проводили по формуле
пользовалась наиболее информативная часть
Δε = ΔD/(c × l), где ΔD - измеренная величина
малоугловых кривых в интервале волновых век
дихроизма, c - концентрация белка (в молях
торов 0,08 < s < 2,7 нм-1.
аминокислотных остатков), l - длина оптичес
Оценка неоднозначности восстановления
кого пути в сантиметрах. Среднюю молярную
формы для заданного профиля рассеяния про
массу аминокислотного остатка принимали рав
водилась с помощью программы AMBIMETER
ной 110 г/моль. Для термического анализа каж
[19].
дый препарат нагревали в кюветном отделении
Анализ брэгговских пиков на кривых мало
дихрографа от 20 до 70 °C со скоростью 1 °C в ми
углового рассеяния выполняли с помощью
нуту. Регистрацию каждого нового спектра начи
программы PEAK [18]. Размер области кристал
нали после увеличения температуры на 5 °C. Из
личности L определялся из полуширины макси
меренные спектры обрабатывали как описано
мума интенсивности первого брэгговского пика
выше.
в угол 2θ1:
Просвечивающая электронная микроскопия.
Исследования проводились с помощью элект
,
(1)
ронного микроскопа JEM 2100, 200 кВ («JEOL»,
Япония), оснащенного источником электронов
LaB6 на оборудовании Уникальной научной ус
и степень разупорядочения в образце Δ/d1 опре
тановки
«3D ЭМС» МГУ (идентификатор
делялась как:
RFMEFI61919X0014). Снимки были сделаны с
помощью CCD камеры Gatan Ultrascan 1000XP
,
(2)
2k («Gatan», США) при параллельном освеще
нии и расфокусировке в диапазоне 1-2 мкм.
Изображения были получены и обработаны с
где d1 = 2π/s1 - период структуры, межплоскост
помощью программного обеспечения Gatan
ное расстояние, βs - полная ширина на полувы
DigitalMicrograph.
соте максимума интенсивности рассеяния (в ра
Эксперимент и анализ данных МУРР. Иссле
дианах), наблюдаемого на угле рассеяния 2θ1,
дования структуры белка с помощью малоугло
соответствующего волновому вектору s1, а Δ -
вого рентгеновского рассеяния были проведены
среднеквадратичное отклонение от расстояния
БИОХИМИЯ том 86 вып. 2 2021
СТРУКТУРА ВИРИОНОВ А ВИРУСА КАРТОФЕЛЯ
277
между ближайшими регулярно упакованными
моделирования служил фрагмент YВКΔ43 Y ви
структурными мотивами.
руса картофеля (PDB ID: 6HXX), поскольку
Для построения функций распределения по
первичная последовательность БО AВК и YВК
расстояниям p(r), которые необходимы для вос
имеют значительное сходство - 64% идентич
становления формы вириона в растворе по дан
ных и 91% подобных аминокислотных остатков
ным МУРР, использовалась компьютерная
(со схожими физико химическими свойства
программа GNOM [20]. Функции распределе
ми), за исключением N концевых фрагментов.
ния по расстояниям p(r) определяются с по
Теоретическая интенсивность рассеяния пост
мощью косвенного Фурье преобразования ин
роенных моделей рассчитывалась программой
тенсивности рассеяния в соответствии с интег
CRYSOL [23].
ральным уравнением:
Выравнивание аминокислотных последова
тельностей. Выравнивание последовательностей
представителей рода потивирусов выполнено на
,
(3)
сервере Expasy с использованием алгоритма
LALIGN [24]. Количественную оценку сходства
где I(s) - интенсивность рассеяния, при этом
последовательностей проводили попарным вы
максимальный размер частицы (Dmax) находится
равниванием структуры «коровой части» без
из условия p(r) = 0 при r > Dmax.
N концевых 30-32 остатков.
Ab initio метод восстановления формы низко
го разрешения вириона AВК и был реализован с
помощью программы DAMMIN [21], которая
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
использует алгоритм имитации отжига путем
минимизации невязки χ2 с экспериментальны
Электрофоретической анализ препаратов. Бе
ми данными
лок оболочки AВК обладает аномально низкой
электрофоретической подвижностью [1], соот
ветствующей примерно 35 кДа (рис. 1, дорож
,
(4)
ки 2-4), в то время как его молекулярная масса
(M) составляет 30,4 кДа. Возможным объясне
где N - число экспериментальных точек, Iexp(sj)
нием этого феномена является наличие в струк
и σ(sj) - экспериментальные интенсивности и
туре белка неупорядоченных сегментов. Мягкая
их ошибки, Icalc(sj) - интенсивность, вычислен
обработка трипсином интактных вирионов (в
ная от модели, c - шкалирующий множитель.
соотношении фермент/субстрат 1 : 500, 15 мин
В более универсальном подходе ab initio мо
инкубации) приводила к исчезновению полосы
дель вируса была получена путем одновремен
исходного белка, и основная доля частиц в этом
ного построения как общей формы, так и струк
случае (около 95% общей массы) приходилась
туры отдельных компонентов вириона (белок и
РНК). Построение двухкомпонентной (двух
фазной) модели проводилось путем приближе
ния кривой МУРР от вирусной частицы с ис
пользованием программы MONSA [21]. Прог
рамма представляет частицу как коллекцию
N >> 1 плотно упакованных шариков внутри
объема поиска, заданного пользователем. Для
описания общей и внутренней структуры слож
ной частицы каждый шарик может быть назна
чен либо принадлежащим растворителю (ин
декс = 0), либо одной из компонент (в нашем
случае индекс = 1 соответствует белку оболочки,
индекс = 2 соответствует вирусной РНК). Комп
лекс между белком и РНК поэтому представлен
в низком разрешении двумя «фазами», а струк
тура описывается строкой длиной N, содержа
щей фазовый индекс для каждого шарика (0, 1
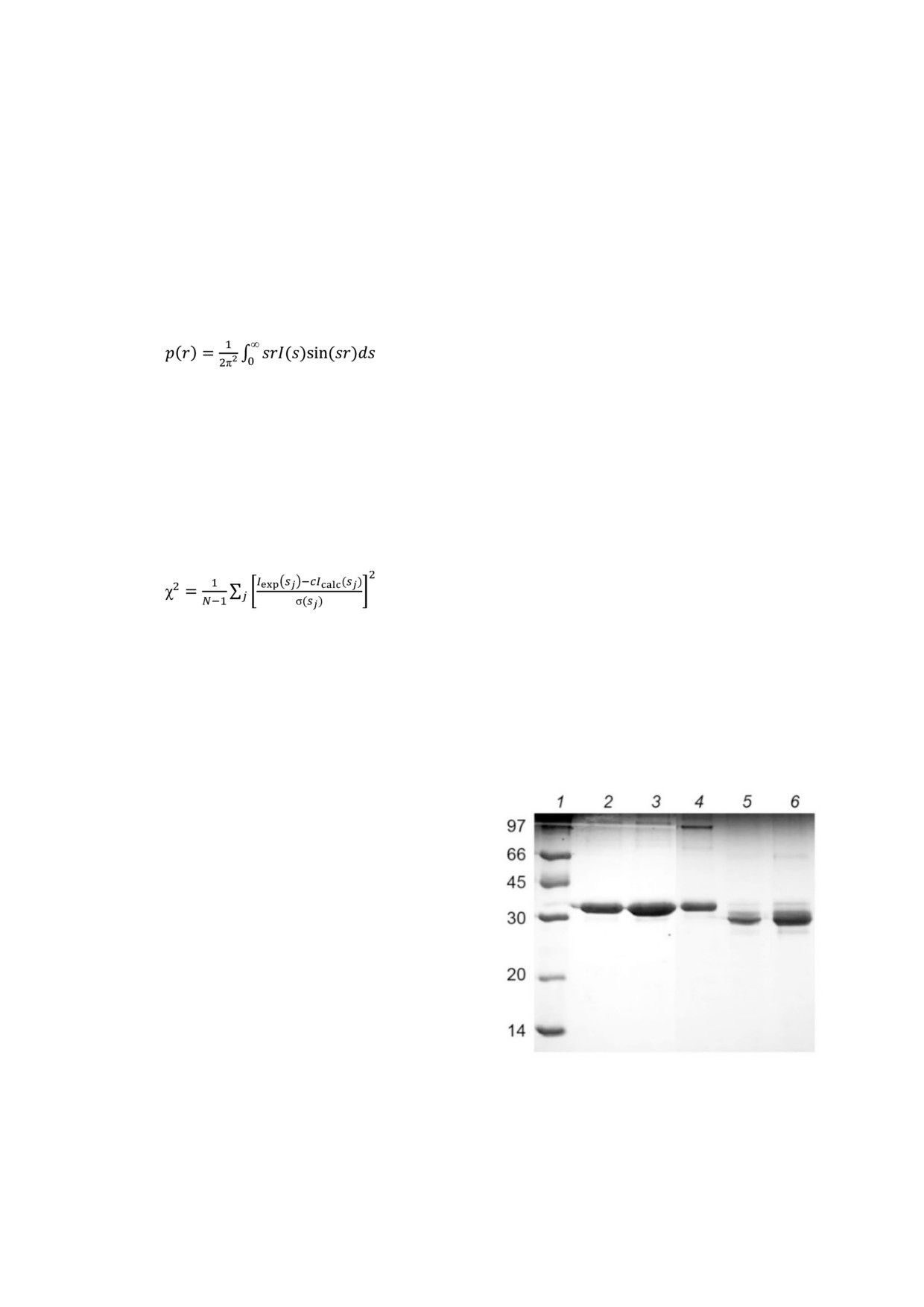
Рис. 1. Электрофоретический анализ интактных вирионов
или 2).
(дорожки 2-4) AВК и продуктов ограниченного трипсино
Моделирование жесткими телами структуры
лиза (дорожки 5, 6). Были использованы аликвоты по 1 мкг
вирионов AВК проводилось с помощью прог
(4, 5), 5 мкг (2) и 10 мкг (3, 6) на полосу. Молекулярная
раммы MASSHA [22]. Единичным элементом
масса указана рядом с контрольными белками (дорожка 1)
БИОХИМИЯ том 86 вып. 2 2021
278
ШТЫКОВА и др.
на полосу с M ~ 27 кДа (рис. 1, дорожки 5 и 6),
тельно ниже, чем в спектре интактных вирионов
что по своему значению соответствует молеку
( 10 600° и
25 000° соответственно). Предполо
лярной массе БО с укороченными N концевы
жительно, с удалением N доменов в БО AВКΔ32
ми фрагментами. Недавно с помощью масс
снижается содержание неупорядоченных сег
спектроскопии было показано почти полное ис
ментов (рис. 2, а, кривая 1).
чезновение пика m/z = 30,4 кДа (исходный БО)
Степень термической денатурации также
и появление мажорного пика 27,12 кДа, соответ
различалась для интактного и деградированно
ствующего белку без 32 N концевых остатков
го трипсином вирионов AВК (рис. 2, б). Ранее
(AВКΔ32) [8]. То есть деградированный трипси
было показано, что плавление интактных вири
ном белок AВКΔ32 относительно контрольных
онов AВК происходит при температуре
белков действительно имеет электрофоретичес
Тпл ≈ 55 °C [1]. В данной работе следили за изме
кую подвижность, соответствующую молеку
нением интенсивности отрицательных макси
лярной массе вириона с укороченными N пос
мумов [θ]max при нагревании. Для препаратов
ледовательностями, что косвенным образом
AВК в диапазоне 20-55 °C наблюдали посте
подтверждает их удаление.
пенный рост интенсивности [θ]max до -45 000° и
Анализ вторичной структуры вирионов по
-29 600° у интактного и деградированного
КД спектрам. Изменения во вторичной структу
AВКΔ32 соответственно (рис. 2, б). Вероятно,
ре белка оболочки AВК после обработки трип
белок оболочки AВКΔ32 денатурировал при
сином отслеживались по спектрам кругового
нагревании в меньшей степени, и вирусные
дихроизма. Спектр КД препарата интактных ви
частицы были более упорядоченными, чем ин
рионов AВК в дальнем УФ характеризуется от
тактные вирионы.
рицательным максимумом [θ]max при 203 нм
Анализ данных МУРР и структурное модели
(рис. 2, а, кривая 2), хотя известно, что для боль
рование. Экспериментальные кривые малоугло
шинства растительных вирусов этот максимум
вого рассеяния от интактных вирионов и от дег
расположен ~208 нм. Предположительно, эта
радированных трипсином вирионов AВКΔ32,
аномалия объясняется необычно высоким со
представленные на рис. 3 (кривые 1 и 2 соответ
держанием разупорядоченных фрагментов в БО
ственно), характеризуются двумя основными
[1]. Насколько нам известно, AВК - это един
особенностями: во первых, хорошо выражен
ственный растительный вирус с таким смеще
ной зависимостью рассеяния от формы вирио
нием главного отрицательного максимума
нов и, во вторых, присутствием на каждой кри
КД спектра в дальнем УФ спектре. В препарате
вой малоуглового рассеяния брэгговского пика
деградированных трипсином вирионов AВКΔ32
в области s от 1,5 до 2,0 нм-1.
мы обнаружили некоторое смещение отрица
Форма вирионов определяется по начальной
тельного максимума [θ]max в сторону к 205 нм, и
части кривых МУРР на интервале волновых
интенсивность этого максимума была значи
векторов s от 0,08 до 1,25 нм-1. В этой области
Рис. 2. а - Спектры кругового дихроизма препаратов, деградированных трипсином (1) и интактных (2) вирионов AВК;
б - термическая денатурация препаратов AВК, деградированных трипсином (1) и интактных (2), по данным КД спект
ров. Приведена температурная зависимость интенсивности отрицательных максимумов [θ]max
БИОХИМИЯ том 86 вып. 2 2021
СТРУКТУРА ВИРИОНОВ А ВИРУСА КАРТОФЕЛЯ
279
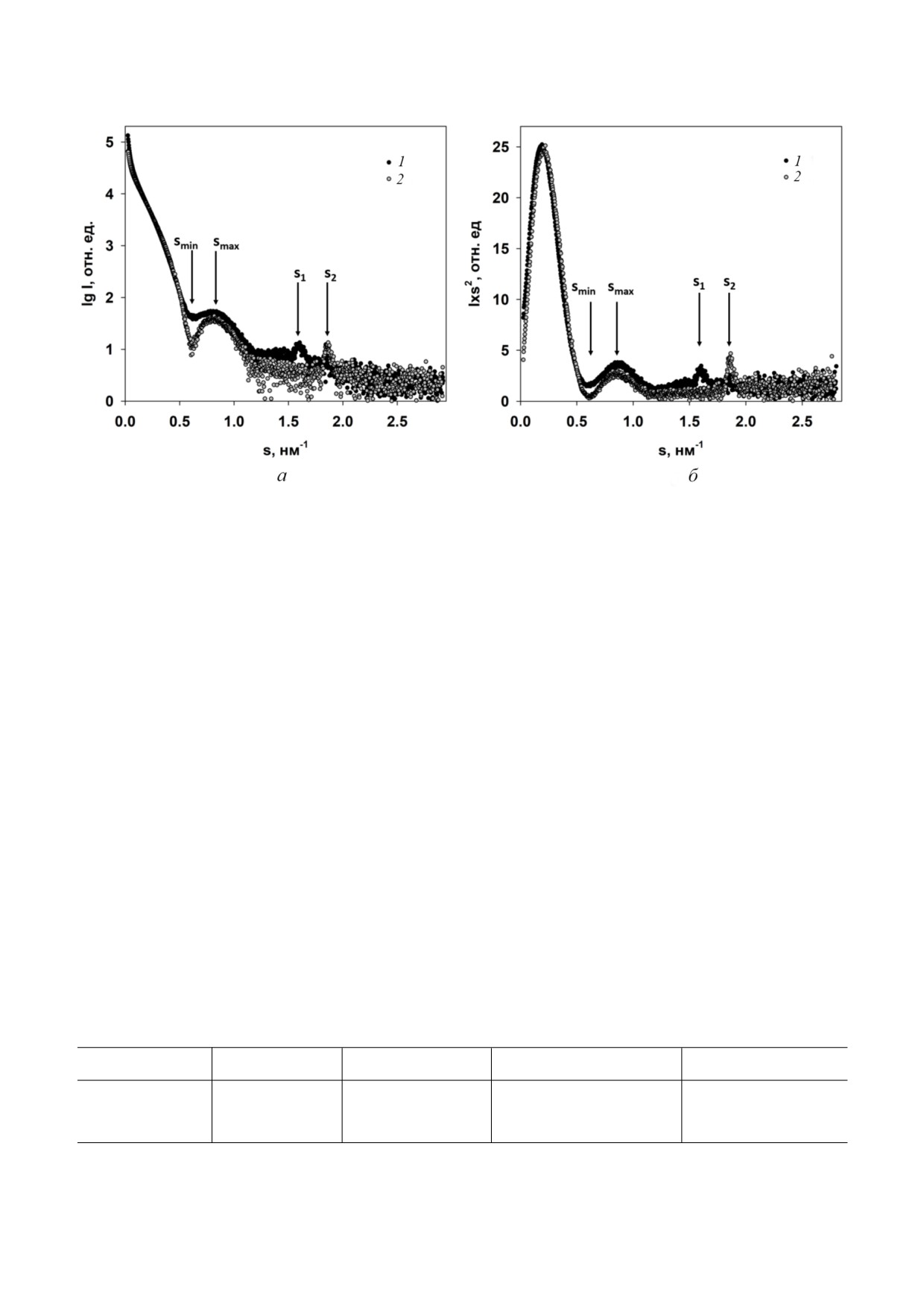
Рис. 3. а - Экспериментальные кривые МУРР от интактных (1) и деградированных AВКΔ32 (2) вирионов; б - графики
Кратки для интактных (1) и деградированных AВКΔ32 (2) вирионов
положение минимума (smin = 0,61 ± 0,01 нм-1) и
Появление брэгговских пиков свидетель
максимума (smax = 0,81 ± 0,01 нм-1) обеих кривых
ствует об определенном упорядочении в образ
совпадает, что свидетельствует о схожести форм
це. Поскольку рассеяние Брэгга характерно для
интактных вирионов и вирионов AВКΔ32. Од
кристаллов, упорядоченные области называют
нако для интактных вирионов кривая МУРР
ся квази кристаллическими, и их структурные
несколько более сглажена, то есть минимум и
характеристики определяются из уравнений (1)
максимум менее выражены по сравнению с кри
и (2) (см. «Материалы и методы»). Рассчитан
вой рассеяния от AВКΔ32. Это объясняется вли
ные по этим уравнениям период структуры
янием на кривые МУРР структурного полимор
d = 2π/s, размеры области кристалличности L и
физма и/или большей полидисперсностью ин
степень разупорядочения в образце Δ/d пред
тактных вирионов [25].
ставлены в таблице.
В самых малых углах, на интервале волновых
Из таблицы следует, что вирион с 32 удален
векторов s от 0,08 до 0,25 нм-1, также наблюдает
ными аминокислотными остатками, AВКΔ32,
ся некоторое расхождение кривых МУРР: интакт
заметно более структурирован: у него больший
ные вирионы имеют несколько большую ампли
размер упорядоченных областей L и меньшая
туду интенсивности рассеяния, что говорит о
степень разупорядочения Δ/d. Для анализа
большем количестве рассеивающего вещества и
структуры также важно, что период d, то есть
косвенно подтверждает наличие N концевых
расстояние между периодически повторяющи
доменов в БО интактных вирионов и отсутствие
мися элементами структуры, уменьшился на
таковых у AВКΔ32. И, наконец, важным отли
0,5 нм при удалении 32 аминокислотных остат
чием профилей рассеяния является различное
ков из N концевых доменов БО. Эти изменения
положение брэгговских пиков на кривых МУРР
относятся к малым, межмолекулярным разме
от AВК и от AВКΔ32 (рис. 3).
рам, то есть к изменениям внутри белковой обо
Структурные характеристики интактных вирионов и вирионов AВКΔ32
Образец
s, нм-1
d, нм
L, нм
Δ/d, нм
Интактный вирион
1,60 ± 0,02
3,92 ± 0,02
125 ± 10
0,06 ± 0,01
AВКΔ32
1,86 ± 0,01
3,37 ± 0,01
180 ± 5
0,04 ± 0,01
БИОХИМИЯ том 86 вып. 2 2021
280
ШТЫКОВА и др.
лочки, и не могут быть интерпретированы как
функции распределения по расстояниям p(r)
взаимодействие между самими вирионами, на
были рассчитаны, во первых, для ab initio опре
пример, при образовании упорядоченных клас
деления формы вирионов в целом, а во вторых,
теров этих крупных частиц. В этом случае пери
для анализа размеров их поперечного сечения.
од системы d был бы не меньше диаметра вири
Последнее необходимо в связи с возможным из
онов даже при самой плотной упаковке. Кроме
менением диаметра нитей при удалении 32 ами
того, при измерении МУРР не наблюдались ни
нокислот из N концевой области БО, предполо
концентрационные зависимости, ни появление
жительно находящихся на внешней поверхнос
пика структурного фактора в самых малых уг
ти вириона. Режим анализа поперечного сече
лах, что подтверждает отсутствие межчастично
ния для удлиненных частиц доступен в качестве
го взаимодействия и формирования класте
одной из опций программы GNOM [20].
ров [26].
Как видно из рис. 4, б, функции распределе
О том, что при удалении N концевых доме
ния по расстояниям p(r), отражающие общую
нов БО не происходит разрушение вириона и в
форму вирионов, характерны для удлиненных
основном сохраняется внешняя форма, но име
рассеивающих объектов с поперечным сечени
ют место внутренние перестройки в белковой
ем порядка 10-16 нм [27]. Более точный анализ
оболочке, свидетельствуют также кривые МУРР
распределения по размерам поперечного сече
в координатах Кратки (рис. 3, б). Они демон
ния (рис. 4, б, вставка) показывает, что макси
стрируют почти полное совпадение профилей
мальный размер поперечного сечения интакт
рассеяния и имеют характерный колоколооб
ного вириона может достигать 18 нм, в то время
разный вид, что указывает на компактность и
как для AВКΔ32 он не превышает 16 нм. Макси
структурированность образцов в целом [12], и в
мум p(r) для AВК расположен на R = 6 нм, то
то же время из за сдвига брэгговских пиков эти
есть в основном диаметр интактного вириона
кривые отражают различия в квази кристалли
равен 12 нм, а для AВКΔ32 R = 5,4 нм и, таким
ческой структуре, то есть изменения на молеку
образом, диаметр вириона действительно стано
лярном уровне.
вится несколько меньше после удаления 32
Определить форму вирионов AВК и AВКΔ32
N концевых аминокислот.
с разрешением 1-2 нм можно, используя ab initio
Восстановленные с помощью программы
протокол и программу DAMMIN [21]. Постро
DAMMIN структуры низкого разрешения AВК
енные с этой целью функции распределения по
и AВКΔ32 представлены на рис. 4, в, пронуме
расстояниям p(r) представлены на рис. 4, б.
рованы 1 и 2 соответственно. Поскольку вирио
Поскольку известно, что вирионы AВК
ны представляют собой длинные цилиндры (ни
представляют собой длинные гибкие нити, ти), поиск решений проводился в цилиндричес
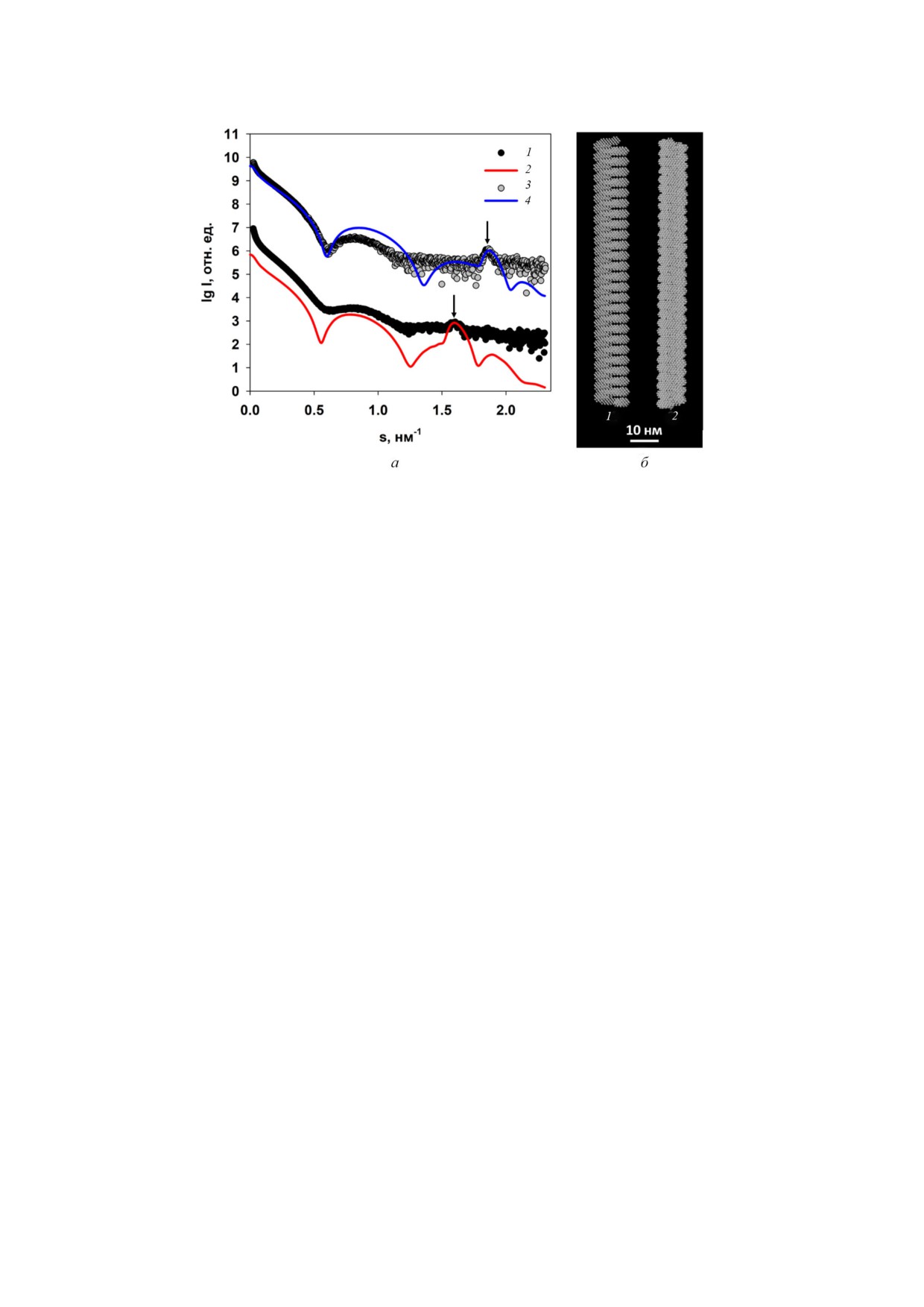
Рис. 4. а - Экспериментальные кривые рассеяния (1 и 5); кривые, рассчитанные программой GNOM от функций расп
ределения по расстояниям p(r) (2 и 6); кривые рассеяния от форм, восстановленных ab initio программой DAMMIN (3 и
7); кривые рассеяния от форм, восстановленных ab initio программой MONSA (4 и 8) для AВК и AВКΔ32 соответственно.
Группы кривых рассеяния для AВК и AВКΔ32 смещены по вертикали для лучшей визуализации; б - функции распреде
ления по расстояниям для AВК (1) и AВКΔ32 (2). Вставка: функции распределения по расстояниям p(r) в режиме анали
за поперечного сечения для AВК (1) и AВКΔ32 (2); в - восстановленные программой DAMMIN формы вирионов AВК (1)
и AВКΔ32 (2); г - восстановленные программой MONSA формы вирионов AВК (1) и AВКΔ32 (2). Белковая оболочка по
казана серыми виртуальными атомами (шариками), вирусная РНК - красными. (С цветными вариантами рис. 4, 5 и 7
БИОХИМИЯ том 86 вып. 2 2021
СТРУКТУРА ВИРИОНОВ А ВИРУСА КАРТОФЕЛЯ
281
кой области с длиной цилиндров Dmax = 105 нм
брэгговских пиков отражает регулярную спи
(рис. 4, б, основная панель) и диаметрами, по
ральную упаковку белков оболочки, которая
лученными при анализе размеров поперечного
присутствует в полученных моделях. При этом
сечения нитей вирионов (рис. 4, б, вставка).
периодичность структуры вирионов более за
Кривые рассеяния от моделей (рис. 4, а, кривые
метна для AВКΔ32.
3 и 7) демонстрируют хорошее совпадение с
Поскольку в МУРР решение обратных за
экспериментальными данными с χ2 = 2,3 для
дач, то есть определение трехмерной структуры
AВК и 1,8 - для частично деградированного ви
рассеивающего объекта по одномерной кривой
риона. В структуре AВКΔ32 заметна определен
интенсивности рассеяния, является принципи
ная периодичность, в то время как интактный
ально неоднозначным, необходима оценка ши
вирион менее упорядочен, и на его общую фор
рины коридора возможных решений. Это про
му оказывает существенное влияние гибкость
изводится с помощью программы AMBIME
нитей.
TER [19]. Этот инструмент быстро определяет
В более универсальном подходе, ab initio мо
степень неоднозначности произвольного про
дель вируса была получена путем одновремен
филя рассеяния от монодисперсного раствора
ного построения как общей формы, так и
однородных частиц и основан на обширной
структуры отдельных компонентов вируса, то
библиотеке диаграмм рассеяния от каркасов
есть белка и РНК. Двухфазная модель, разгра
формы (шаблонов), задающих многообразие
ничивающая белковую и РНК части, была
топологий структуры частиц низкого разреше
построена программой MONSA [21]. В качестве
ния. Количество шаблонов, похожих на задан
объема поиска были использованы два цилинд
ные экспериментальные данные МУРР, обеспе
ра. Первый, для моделирования РНК, пред
чивают меру неоднозначности, связанной с
ставлял собой внутренний сплошной цилиндр с
этими данными, и логарифм этого значения
радиусом 3,2 нм и длиной 105 нм как для AВК,
выражается в виде количественной оценки в
так и для частично деградированного вириона.
программе AMBIMETER. Как правило, оценка
Второй цилиндр являлся объемом поиска для
ниже 1,5 предполагает, что найдено уникаль
восстановления структуры белковой оболочки
ное, то есть практически единственное реше
и был полым и внешним по отношению к пер
ние, а оценка выше 2,5 говорит о неоднознач
вому цилиндру с внешним радиусом 8,0 нм для
ном восстановлении формы.
интактного вириона и 7,0 нм - для AВКΔ32, в
В настоящей работе для AВКΔ32 нашлась
соответствии с результатами анализа попереч
только одна совместимая топология, что свиде
ного сечения. Внутренний радиус второго ци
тельствует об узком интервале близких форм
линдра был равен 3,5 нм для обоих вирионов. В
структур низкого разрешения. В случае с инта
процессе моделирования виртуальные атомы
ктным AВК совместимыми оказались 88 карка
(шарики) во внешнем цилиндре могли прини
сов формы, и соответствующая оценка неодноз
мать фазовые индексы 1 и 0 (т.е. белок или раст
начности составляет 1,9, что говорит о возмож
воритель), а во внутреннем 2 и 0 (т.е. РНК или
ной неоднозначности восстановлении формы
растворитель). Таким образом, была обеспечена
этого образца, обусловленной гибкостью интакт
разумность модели, где белок оболочки оказы
ного вириона.
вался на внешней поверхности, а РНК в глуби
В целом, ab initio восстановление формы ви
не. Построенные двухфазные модели, показан
рионов по данным малоуглового рассеяния
ные на рис. 4, г, аппроксимируются искажен
продемонстрировало заметные изменения в
ными цилиндрами соответствующих размеров
структуре вириона после удаления 32 амино
и хорошо согласуются с моделями, построен
кислот N концевых доменов белка оболочки.
ными программой DAMMIN. Искажения полу
Эти изменения касаются как размера попереч
ченных форм (неидеальность) свидетельствуют
ного сечения нитей вириона, так и периодич
как о влиянии гибкости вирионов, так и о час
ности структуры БО. Уменьшение периода d
тичной разупорядоченности структуры в целом
предположительно связано с уменьшением
и белков оболочки в частности. Характерно, что
шага спиральной упаковки белка в оболочке
шарики, моделирующие РНК, простираются
вируса.
вдоль всей оси цилиндра, что соответствует из
Для того, чтобы проверить правильность
вестным представлениям о структуре вируса.
этого предположения было проведено модели
Двухфазные модели хорошо приближают экс
рование структуры AВК и AВКΔ32 геометричес
периментальные кривые МУРР (рис. 4, а, кри
кими телами. Для этого были построены шари
вые 4 и 8) с χ2 = 3,4. Следует отметить, что брэг
ковые модели спиральных тел, внешние разме
говские пики в районе s = 1,6 и 1,9 нм-1 также
ры которых определялись из анализа функций
очень хорошо воспроизведены. Присутствие
распределений по расстояниям p(r). От полу
БИОХИМИЯ том 86 вып. 2 2021
282
ШТЫКОВА и др.
Рис. 5. Моделирование строения интактного и частично деградированного вирионов: а - экспериментальные кривые
МУРР (1 и 3) и рассеяние от спиральных тел (2 и 4) для AВК и AВКΔ32 соответственно. Стрелками показаны моделируе
мые брэгговские пики; б - спиральные модели, имитирующие интактный вирион (1) и AВКΔ32 (2)
ченных спиралей были рассчитаны интенсив
жестким и поэтому лучше аппроксимируется
ности малоуглового рассеяния и проведено
цилиндрической формой (рис. 4).
сравнение с экспериментальными данными
Из полученных результатов можно также сде
МУРР от AВК и AВКΔ32 (рис. 5, а).
лать предположение о том, что N концевой до
Моделирование геометрическими телами
мен частично расположен между витками спи
представляет собой метод грубого приближе
ральной упаковки белка оболочки, осуществляя
ния, который не может учитывать все особен
взаимодействие с соседними белковыми субъ
ности реальных структур, подвижность и гиб
единицами и удерживая витки спирали на опре
кость их отдельных фрагментов, но позволяет
деленном расстоянии друг от друга. С другой сто
построить модели выделенных, особенно важ
роны, часть белковой цепи N концевого домена
ных для анализа элементов. В данном случае
выступает над поверхностью вируса, о чем свиде
требовалось доказать, что брэгговские пики на
тельствует больший диаметр интактного вириона
экспериментальных кривых рассеяния отража
по сравнению с частично деградированным.
ют шаг спиральной упаковки белка в оболочке
Важно отметить, что неструктурированные об
вируса. Для этого расстояние между витками
ласти белковых молекул являются гибким связу
модельных спиралей было задано в соответ
ющим звеном при взаимодействии между раз
ствии с характеристическим периодом d = 2π/s,
личными белками и другими компонентами кле
то есть d = 3,9 нм для интактного вириона и
ток [5]. В то же время межклеточный транспорт
d = 3,4 нм для частично деградированного (таб
необходимых веществ для жизнедеятельности
лица). Толщина нитей спиралей была выбрана
растений происходит через плазмодесмы - и че
равной 3,4 нм в предположении, что эта величи
рез эти же плазмодесмы в клетки могут прони
на не может быть больше шага спирали для
кать растительные патогены, несмотря на свои
AВКΔ32. Небольшие изменения в меньшую сто
большие размеры. Для этого вирусы избиратель
рону не влияли на результат моделирования.
но используют, например, некоторые миозины
Как видно из рис. 5, а, максимумы интенсив
клеток [28]. Поэтому можно предположить, что
ности рассеяния от модельных структур хорошо
разупорядоченные, гибкие фрагменты N конце
описывают брэгговские пики на эксперимен
вого домена AВК могут служить инструментом
тальных данных. Отсюда можно сделать вывод,
связывания с определенными клеточными бел
что при удалении 32 аминокислот N концевого
ками для обеспечения межклеточного транспор
домена AВК действительно происходит измене
та. В таком случае прямое воздействие на эти
ние шага спирали белковой оболочки - спираль
фрагменты вириона соответствующими агента
сжимается, и вирион в целом становится более
ми может привести к дезактивации патогена.
БИОХИМИЯ том 86 вып. 2 2021
СТРУКТУРА ВИРИОНОВ А ВИРУСА КАРТОФЕЛЯ
283
С другой стороны, большое значение для
парным выравниванием первичных структур
проникновения вируса в клетки имеет гибкость
определяла 64% идентичных и 91% подобных
вириона. Известно, что гибкость возрастает с
остатков из 237 остатков БО (без 32 N концевых
увеличением длины вирионов в ряду: тобамови
остатков) [24] (рис. 6).
русы (300 нм), потексвирусы (500-1000 нм),
Таким образом, высокая идентичность пер
клостеровирусы (1400-2000 нм), что облегчает
вичной структуры белков оболочки этих двух
их межклеточный транспорт через плазмодес
вирусов позволила использовать модель с раз
мы. То есть, чем длиннее вирион, тем более гиб
решением 3,4 Å БО YВК (PDB ID: 6HXX) для
ким он должен быть для проникновения в клет
моделирования строения белковой оболочки
ку. Возможно также, что именно N концевой
A вируса картофеля. БО YВК характеризуется
домен определяет гибкость вирионов. Получен
спиральной симметрией, а использованный для
ные в данной работе результаты свидетельству
моделирования фрагмент состоял из 35 белко
ют в пользу такого предположения.
вых субъединиц, включая в себя атомные коор
В настоящее время не существует структуры
динаты аминокислот с 44 по 267 остаток, то есть
высокого разрешения для AВК, но недавно ме
без 43 аминокислотных остатков N концевого
тодом криоэлектронной микроскопии была по
домена, атомные координаты которых не были
лучена структура с разрешением 3,4 Å БО друго
определены из за низкой электронной плот
го потивируса, YВК, с максимально близкой к
ности этих разупорядоченных фрагментов.
БО AВК гомологией [3]. Поэтому нами была
Фрагмент YВК был многократно воспроизве
предпринята попытка сравнения структуры вы
ден с помощью программы MASSHA [22] вдоль
сокого разрешения YВК со структурой A вируса
винтовой оси для построения модели, содержа
картофеля, используя данные малоуглового рас
щей 30 витков (260 субъединиц) с максималь
сеяния и метод моделирования жесткими тела
ной длиной 105 нм. Число витков подбиралось
ми. Первичные структуры БО AВК и YВК име
таким образом, чтобы получить наилучшее со
ют значительное сходство, за исключением
ответствие между экспериментальными данны
N концевых остатков. Программа LALIGN по
ми и кривой рассеяния. Сравнение профилей
Рис. 6. Выравнивание первичных структур AВК (англ. абб. PVA) и YВК (англ. абб. PVY) с помощью программы LALIGN
(Expasy). Звездочками (*) отмечены идентичные остатки, точками (.) - полуконсервативные замены. Подчеркнуты по
следовательности, удаленные трипсином (для AВК) или отсутствующие в PDB ID: 6HXX (для YВК). Представлены струк
туры БО AВК № GI:25013598 и YВК (штамм NTN) № GI:728041063
БИОХИМИЯ том 86 вып. 2 2021
284
ШТЫКОВА и др.
Рис. 7. Результат моделирования строения белковой оболочки A вируса картофеля с использованием структурных блоков
высокого разрешения БО YВК (PDB ID: 6HXX): а - (1) экспериментальная кривая малоуглового рассеяния от AВКΔ32;
(2) рассчитанный программой CRYSOL профиль рассеяния от полученной структурной модели; б - структурная модель
белковой оболочки, имитирующая A вирус картофеля
Рис. 8. Результаты электронной микроскопии интактных (а) и деградированных AВКΔ32 (б) вирионов
экспериментальной и модельной кривых демон
отклонения свидетельствуют о различии струк
стрирует некоторое расхождение кривых рассе
тур белковых оболочек вирионов AВК и YВК,
яния как в интервале 0,08 < s < 1,2 нм1, так и
несмотря на то, что они являются ближайшими
небольшой сдвиг брэгговского пика с максиму
гомологами. При этом следует учесть, что полу
мом на s = 1,9 нм1 (d = 3,4 нм) в сторону боль
ченная модель более близка не к интактному
ших размеров с s = 1,8 нм1 (d = 3,5 нм). Эти
вириону AВК, а к его частично деградирован
БИОХИМИЯ том 86 вып. 2 2021
СТРУКТУРА ВИРИОНОВ А ВИРУСА КАРТОФЕЛЯ
285
ной форме. Кроме того, расхождение между
вого рассеяния с учетом разного состояния ви
экспериментальной и модельной кривой χ2 ~ 25
русов при измерениях малоуглового рассеяния и
(рис.
7) отражает неидеальность реальной
ПЭМ: в МУРР вирионы находятся в растворе,
структуры этого вириона по сравнению с его
то есть в состоянии, наиболее близком к естест
жесткой моделью на атомном уровне, не спо
венному, в то время как для исследования с по
собной изменять свою конформацию. В то же
мощью электронной микроскопии образцы ви
время биологические измерения в малоугловом
рионов были высушены на подложке.
рассеянии происходят в растворе, где рассеива
Впервые структура низкого разрешения ни
ющие частицы сохраняют свои основные свой
тевидного растительного вируса AВК, интакт
ства, в том числе гибкость.
ного и частично деградированного, была полу
Тем не менее полученная модельная форма
чена в условиях, близких к естественным. Ре
AВК высокого разрешения показывает несом
зультаты малоуглового рассеяния показали, что
ненное структурное сходство белковых оболо
удаление 32 N концевых аминокислот привело
чек двух разных потивирусов.
к изменению шага спиральной упаковки белко
Основной вывод этой части работы состоит в
вой оболочки. Дополнительные исследования
том, что обработка трипсином и частичная дег
показали, что деградированные трипсином ви
радация AВК привела к изменению шага спи
рионы имели более упорядоченную структуру и
ральной упаковки белковой оболочки: после
представляли собой преимущественно палочко
удаления 32 аминокислот расстояние между
видные частицы меньшего диаметра в сравне
витками спирали уменьшилось с 3,9 нм до
нии с гибкими частицами интактного вируса,
3,4 нм. Такое сжатие спирали неизбежно долж
что хорошо коррелировало с результатами
но было привести к изменению гибкости час
МУРР. В целом, проведенный структурный ана
тично деградированного вириона: сжатая пру
лиз свидетельствует о значении N концевых до
жина всегда значительно более жесткая, чем
менов белка оболочки для выполнения жизнен
свободная спираль. Подтверждение этому выво
но важных функций AВК, что следует учитывать
ду было найдено с помощью просвечивающей
для выработки стратегии по борьбе с этими рас
электронной микроскопии.
тительными патогенами.
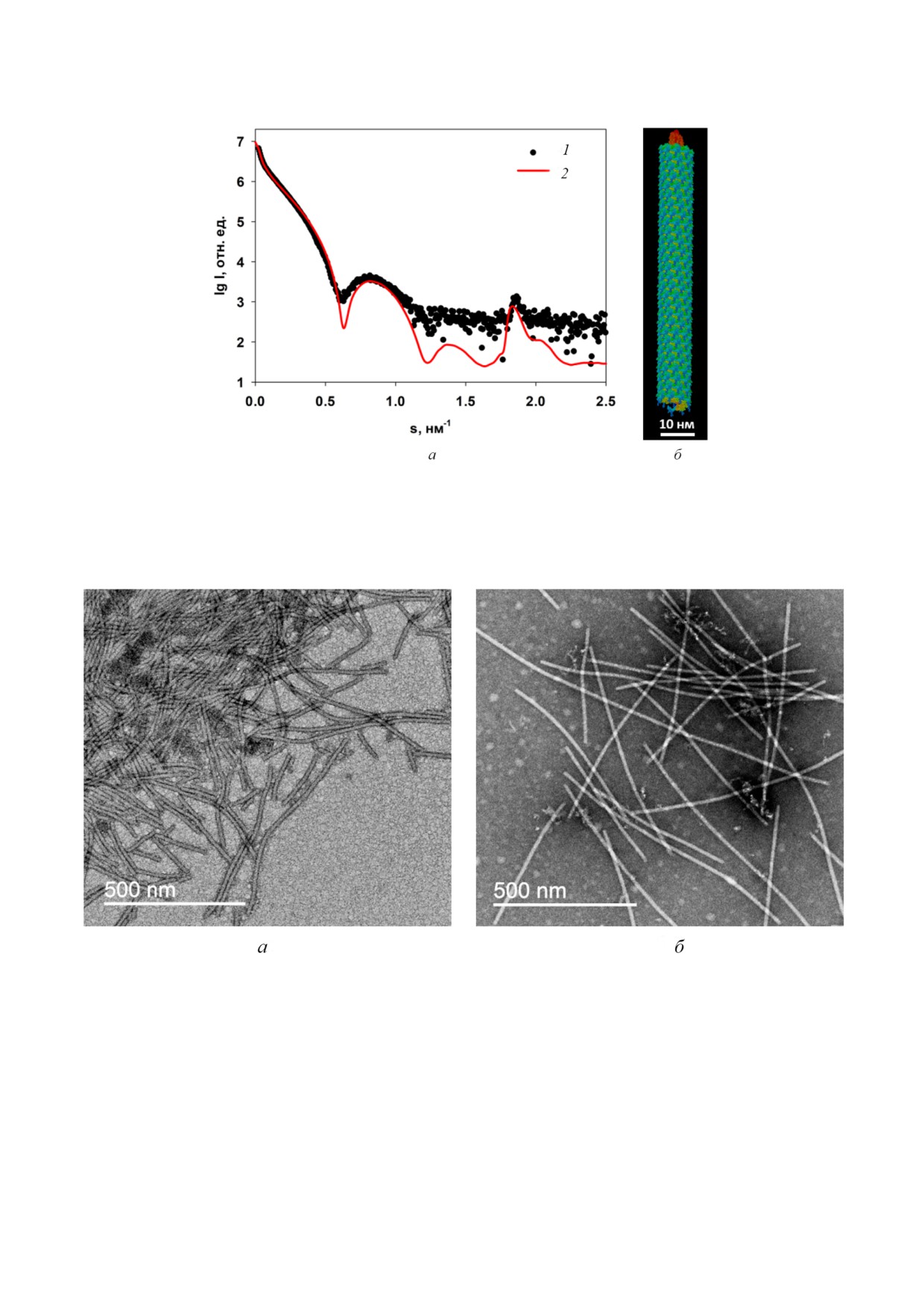
Электронно микроскопический анализ вирио
нов AВК. Согласно данным ПЭМ, интактные
вирионы представляли собой типичные гибкие
Финансирование. Работа выполнена при под
нитевидные частицы (рис. 8, а), тогда как дегра
держке Российского фонда фундаментальных
дированные трипсином вирионы AВКΔ32 в ос
исследований (грант № 18 04 00525а) и Минис
новном представляли собой палочковидные
терства науки и высшего образования РФ: в
частицы с меньшим диаметром (рис. 8, б), похо
рамках выполнения работ по Государственному
жие на «сухие спагетти». С помощью комплекса
заданию ФНИЦ «Кристаллография и фотони
«Gatan» (DigitalMicrograph software) диаметры
ка» РАН в части проведения экспериментов
этих частиц были измерены и оказались равны
МУРР; в рамках соглашения №075 15 2019
ми 14,4 ± 0,2 нм и 12,1 ± 0,3 нм, для интактных
1653 в части экспериментов ПЭМ.
и деградированных вирионов соответственно,
Конфликт интересов. Авторы заявляют об от
что также косвенно указывало на удаление
сутствии конфликта интересов.
N концевых остатков с поверхности вирионов
Соблюдение этических норм. Настоящая
AВК.
статья не содержит описания каких либо иссле
В целом, полученные методом ПЭМ данные
дований с участием людей или использованием
хорошо коррелируют с результатами малоугло
животных в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Ksenofontov, A. L., Paalme, V., Arutyunyan, A. M.,
ssRNA viruses, Sci. Adv., 3, eaao2182, doi: 10.1126/
Semenyuk, P. I., Fedorova, N. V., et al. (2013) Partially dis
sciadv.aao2182.
ordered structure in intravirus coat protein of potyvirus
3.
Kezar, A., Kavcic, L., Polak, M., Novacek, J.,
potato virus A, PLoS One, 8, e67830, doi: 10.1371/journal.
Gutierrez Aguirre, I., et al. (2019) Structural basis for
pone.0067830.
the multitasking nature of the potato virus Y coat pro
2.
Zamora, M., Mendez Lopez, E., Agirrezabala, X.,
tein, Sci. Adv.,
5, eaaw3808, doi:
10.1126/sciadv.
Cuesta, R., Lavin, J. L., Sanchez Pina, M. A., Aranda,
aaw3808.
M. A., and Valle, M. (2017) Potyvirus virion structure
4.
Cuesta, R., Yuste Calvo, C., Gil Carton, D., Sanchez, F.,
shows conserved protein fold and RNA binding site in
Ponz, F., and Valle, M. (2019) Structure of Turnip mosaic
БИОХИМИЯ том 86 вып. 2 2021
286
ШТЫКОВА и др.
virus and its viral like particles, Sci. Rep., 9,
15396,
(2006) Determination of concentration and aggregate size
doi: 10.1038/s41598 019 51823 4.
in influenza virus preparations using the true UV absorp
5.
Uversky, V. N. (2013) Unusual biophysics of intrinsically
tion spectra, Mol. Biol., 40, 152158, doi: 10.1134/
disordered proteins, Biochim. Biophys. Acta, 1834, 932
S0026893306010201.
951, doi: 10.1016/j.bbapap.2012.12.008.
17.
Blanchet, C. E., Spilotros, A., Schwemmer, F., Graewert,
6.
Ksenofontov, A. L., Parshina, E. Y., Fedorova, N. V.,
M. A., Kikhney, A., et al. (2015) Versatile sample environ
Arutyunyan, A. M., Rumvolt, R., Paalme, V., Baratova,
ments and automation for biological solution X ray scat
L. A., Jarvekulg, L., and Dobrov, E. N. (2016) Heating
tering experiments at the P12 beamline (PETRA III,
induced transition of Potyvirus Potato Virus A coat pro
DESY), J. Appl. Crystallogr., 48, 431 443, doi: 10.1107/
tein into beta structure, J. Biomol. Struct. Dyn., 34, 250
S160057671500254X.
258, doi: 10.1080/07391102.2015.1022604.
18.
Konarev, P. V., Volkov, V. V., Sokolova, A. V., Koch, M. H.
7.
Ksenofontov, A. L., Dobrov, E. N., Fedorova, N. V.,
J., and Svergun, D. I. (2003) PRIMUS: a Windows PC
Arutyunyan, A. M., Golanikov, A. E., Järvekülg, L., and
based system for small angle scattering data analysis, J.
Shtykova, E. V. (2018) Structure of Potato Virus A coat
Appl. Crystallogr.,
36,
12771282, doi:
10.1107/
protein particles and their dissociation, Mol. Biol. (Mosk.),
s0021889803012779.
52, 1055 1065, doi: 10.1134/S0026898418060101.
19.
Petoukhov, M. V., and Svergun, D. I. (2015) Ambiguity
8.
Ksenofontov, A. L., Dobrov, E. N., Fedorova, N. V.,
assessment of small angle scattering curves from monodis
Serebryakova, M. V., Prusov, A. N., Baratova, L. A.,
perse systems, Acta Crystallogr. D Biol. Crystallogr., 71,
Paalme, V., Jarvekulg, L., and Shtykova, E. V. (2018) Isolated
1051 1058, doi: 10.1107/s1399004715002576.
Potato Virus A coat protein possesses unusual properties and
20.
Svergun, D. I. (1992) Determination of the regularization
forms different short virus like particles, J. Biomol. Struct.
parameter in indirect transform methods using perceptual
Dyn., 36, 1 11, doi: 10.1080/07391102.2017.1333457.
criteria, J. Appl. Crystallogr., 25, 495 503, doi: 10.1107/
9.
Anindya, R., and Savithri, H. S. (2003) Surface exposed
s0021889892001663.
amino and carboxy terminal residues are crucial for the
21.
Svergun, D. I. (1999) Restoring low resolution structure of
initiation of assembly in Pepper vein banding virus: a flex
biological macromolecules from solution scattering using
uous rod shaped virus, Virology,
316,
325336,
simulated annealing, Biophys. J.,
76,
28792886,
doi: 10.1016/s0042 6822(03)00593 2.
doi: 10.1016/S0006 3495(99)77443 6.
10.
Tatineni, S., McMechan, A. J., and Hein, G. L. (2018)
22.
Konarev, P. V., Petoukhov, M. V., and Svergun, D. I. (2001)
Wheat streak mosaic virus coat protein is a determinant for
MASSHA- a graphics system for rigid body modelling of
vector transmission by the wheat curl mite, Virology, 514,
macromolecular complexes against solution scattering
42 49, doi: 10.1016/j.virol.2017.10.018.
data, J. Appl. Crystallogr., 34, 527 532, doi: 10.1107/
11.
Voloudakis, A. E., Malpica, C. A., Aleman Verdaguer, M.
s0021889801006100.
E., Stark, D. M., Fauquet, C. M., and Beachy, R. N.
23.
Svergun, D., Barberato, C., and Koch, M. H. J. (1995)
(2004) Structural characterization of Tobacco etch virus
CRYSOL - a program to evaluate x ray solution scattering
coat protein mutants, Arch. Virol.,
149,
699712,
of biological macromolecules from atomic coordinates, J.
doi: 10.1007/s00705 003 0247 x.
Appl. Crystallogr.,
28,
768773, doi:
10.1107/
12.
Svergun, D. I., Koch, M. H. J., Timmins, P. A., and May,
s0021889895007047.
R. P. (2013) Small angle x ray and neutron scattering from
24.
Artimo, P., Jonnalagedda, M., Arnold, K., Baratin, D.,
solutions of biological macromolecules, First Ed., Oxford
Csardi, G., et al. (2012) ExPASy: SIB bioinformatics
University Press, Oxford, doi:
10.1093/acprof:oso/
resource portal, Nucleic Acids Res,, 40, W597 603,
9780199639533.001.0001.
doi: 10.1093/nar/gks400.
13.
Franke, D., Petoukhov, M. V., Konarev, P. V., Panjkovich,
25.
Shtykova, E. V. (2015) Shape determination of polydis
A., Tuukkanen, A., et al. (2017) ATSAS 2.8: a comprehen
perse and polymorphic nanoobjects from small angle X
sive data analysis suite for small angle scattering from
ray scattering data (computer simulation). Nanotechnol.
macromolecular solutions, J. Appl. Crystallogr., 50, 1212
Russia, 10, 408 419, doi: 10.1134/S1995078015030155.
1225, doi: 10.1107/S1600576717007786.
26.
Tardieu, A. (1994) Neutron and synchrotron radiation for
14.
Ksenofontov, A. L., Petoukhov, M. V., Prusov, A. N.,
condensed matter studies. Applications to Soft Condensed
Fedorova, N. V., and Shtykova, E. V. (2020) Characteriza
Matter and Biology, vol III (Les editions de Physique
tion of tobacco mosaic virus virions and repolymerized
(France)) (Berlin: Springer) pp 145 160.
coat protein aggregates in solution by small angle x ray
27.
Feigin, L. A., and Svergun, D. I. (1987) Structure analysis
scattering,
Biochemistry (Mosc.),
85,
310317,
by small angle x ray and neutron scattering, Plenum Press,
doi: 10.1134/s0006297920030062.
New York, p. 335.
15.
Laemmli, U. K. (1970) Cleavage of structural proteins dur
28.
Amari, K., Lerich, A., Schmitt Keichinger, C., Dolja, V.
ing the assembly of the head of bacteriophage T4, Nature,
V., and Ritzenthaler, C. (2011) Tubule guided cell to cell
227, 680 685, doi: 10.1038/227680a0.
movement of a plant virus requires class XI myosin motors,
16.
Ksenofontov, A. L., Kozlovskii, V. S., Kordiukova, L. V.,
PloS Pathog., 7, e1002327, doi: 10.1371/journal.ppat.
Radiukhin, V. A., Timofeeva, A. V., and Dobrov, E. N.
1002327.
БИОХИМИЯ том 86 вып. 2 2021
СТРУКТУРА ВИРИОНОВ А ВИРУСА КАРТОФЕЛЯ
287
STRUCTURE OF POTATO VIRUS A VIRIONS ACCORDING TO SMALL
ANGLE X RAY SCATTERING DATA AND COMPLEMENTARY METHODS*
E. V. Shtykova1, M. V. Petukhov1, N. V. Fedorova2, A. M. Arutyunyan2, E. V. Skurat3,
L. V. Kordyukova2, А. V. Moiseenko3, and A. L. Ksenofontov2**
1 Shubnikov Institute of Crystallography of Federal Scientific Research Centre «Crystallography and Photonics»,
Russian Academy of Sciences, 119333 Moscow, Russia; E%mail: shtykova@ns.crys.ras.ru
2 Belozersky Institute of Physico%Сhemical Biology, Lomonosov Moscow State University,
119991 Moscow, Russia; E%mail: ksenofon@belozersky.msu.ru
3 Faculty of Biology, Lomonosov Moscow State University, 119991 Moscow, Russia
Potato virus A (PVA) protein coat contains on its surface partially unstructured N terminal domain of the viral coat
protein (CP), whose structural and functional characteristics are important for understanding the mechanism of plant
infection with this virus. In this work, we investigated the properties and the structure of intact PVA and partially
trypsinized PVAΔ32 virions using small angle X ray scattering (SAXS) and complimentary methods. It was shown
that after the removal of 32 N terminal amino acids of the CP, the virion did not disintegrate and remained compact,
but the helical pitch of the CP packing changed. To determine the nature of these changes, we performed ab initio
modeling, including the multiphase procedure, with the geometric bodies (helices) and restoration of the PVA struc
ture in solution using available high resolution structures of the homologous CP from the PVY potyvirus, based on
the SAXS data. As a result, for the first time, a low resolution structure of the filamentous PVA virus, both intact and
partially degraded, was elucidated under conditions close to natural. The far UV circular dichroism spectra of the
PVA and PVAΔ32 samples differed significantly in the amplitude and position of the main negative maximum. The
extent of thermal denaturation of these samples in the temperature range of 20 55°C was also different. The data of
transmission electron microscopy showed that the PVAΔ32 virions were mostly rod shaped, in contrast to the flexible
filamentous particles typical of the intact virus, which correlated well with the SAXS results. In general, structural
analysis indicates an importance of the CP N terminal domain for the vital functions of PVA, which can be used to
develop a strategy for combating this plant pathogen.
Keywords: potyviruses, coat protein, potato virus A, N terminal disordered domains, small angle X ray scattering,
structural modeling
БИОХИМИЯ том 86 вып. 2 2021