БИОХИМИЯ, 2021, том 86, вып. 3, с. 431 - 442
УДК 612.744.24 612.018;577.218
ВЛИЯНИЕ ДИНАМИЧЕСКИХ И СТАТИЧЕСКИХ НАГРУЗОК
НА КОНЦЕНТРАЦИЮ МИОКИНОВ В ПЛАЗМЕ И НА СОДЕРЖАНИЕ
НАТРИЯ И КАЛИЯ В СКЕЛЕТНЫХ МЫШЦАХ МЫШЕЙ
© 2021
Т.А. Кироненко1, К.Г. Милованова1, А.Н. Захарова1, С.В. Сидоренко2,
Е.А. Климанова1,2, Е.Ю. Дьякова1, А.А. Орлова1, Е.С. Негоденко1,
Ю.Г. Калинникова1,
С.Н. Орлов1,2
, Л.В. Капилевич1*
1 Национальный исследовательский Томский государственный университет,
634050 Томск, Россия, электронная почта: kapil@yandex.ru
2 Московский государственный университет имени М.В.Ломоносова, биологический факультет,
119234 Москва, Россия; электронная почта: klimanova.ea@yandex.ru
Поступила в редакцию 09.07.2020
После доработки 15.09.2020
Принята к публикации 29.10.2020
Модификация продукции цитокинов под влиянием физических нагрузок представляет значительный инте
рес, так как является перспективным путем коррекции процессов метаболизма как на клеточном, так и на
системном уровнях. В работе изучено содержание IL 6, IL 8 и IL 15 в плазме и одновалентных катионов в
скелетных мышцах у мышей спустя разное время после статических и динамических нагрузок с учетом
предварительной тренировки. Динамические упражнения вызывают увеличение содержания IL 6 и сниже
ние IL 15 в плазме нетренированных мышей, не влияя на концентрацию IL 8. У тренированных мышей эф
фекты однократной нагрузки на концентрацию IL 6 и IL 15 в плазме усиливались, также отмечалось сни
жение концентрации IL 8. В отличие от динамических, статические нагрузки оказывают аналогичное, но
более выраженное влияние на концентрацию IL 6 и IL 15 в плазме. Однако концентрация IL 8 в ответ на
статическую нагрузку существенно увеличивалась. Предварительные тренировки усиливали описанные ре
акции для всех изученных миокинов. Показано разнонаправленное влияние динамической нагрузки (пла
вание) на содержания внутриклеточного натрия (повышение) и калия (снижение) в musculus soleus мыши.
Впервые обнаружено, что аналогичное явление наблюдается при статической нагрузке (вис на сетке) в mus
culus biceps мыши и достоверно не зависит от предварительной тренировки экспериментальных животных.
Обсуждаются возможные механизмы, обеспечивающие регуляцию секреции цитокинов после физической
нагрузки, в том числе [Na+]i/[K+]i зависимый механизм запуска транскрипции генов.
КЛЮЧЕВЫЕ СЛОВА: физические нагрузки, миокины, ионы, Na+/K+ ATPаза, транскрипция, трансляция.
DOI: 10.31857/S0320972521030118
ВВЕДЕНИЕ
опухоли α (TNF α) и лейкемия ингибирующе
го фактора (LIF). К настоящему времени сфор
Клетки скелетных мышц составляют важную
мировалась концепция об эндокринной функ
часть опорно двигательного аппарата, обеспе
ции скелетных мышц, а за веществами, проду
чивая движение или сохранение позы организ
цируемыми мышечными клетками при сокра
ма. Эта их функция тесно сопряжена с процесса
щении, закрепилось название «миокины». Уста
ми потребления и производства энергии [1]. Од
новлено, что скелетные мышцы являются ос
нако в последнее десятилетие внимание иссле
новным (хотя и не единственным) источником
дователей привлекла способность скелетных
продукции IL 6, с чем связано возрастание его
мышц продуцировать биологически активные
содержания в плазме крови после физической
вещества. Выяснилось [2-5], что на фоне физи
нагрузки [6]. Показано увеличение транскрип
ческой активности происходит повышение со
ции матричной RNA (mRNA) IL 6 в ядрах мы
держания в плазме крови ряда цитокинов, в том
шечных клеток, выделенных из биоптатов
числе IL 1β, IL 6, IL 8, IL 15, фактора некроза
мышц людей после выполнения однократного
упражнения [7]. Кроме того, для изучения про
дукции миокинов мышечными клетками ис
Принятые сокращения: EPS - электростимуляция;
HIF - фактор, индуцируемый гипоксией; IL - интерлей
пользовали культуры миобластов мышей ли
кин; mRNA - матричная RNA.
нии C2C12 и первичные человеческие миотубу
* Адресат для корреспонденции.
лы, которые подвергали электростимуля
431
432
КИРОНЕНКО и др.
ции (EPS) [8-10]. При использовании такого
но. Эти результаты согласуются с уменьшени
подхода было показано, что через 24 ч после EPS
ем [K+]i и увеличением [Na+]i, наблюдавшимися
в миотубулах человека происходит достоверное
при тренировке скелетных мышц in vivo [27].
изменение транскрипции 183 генов, а также уве
Увеличение внутриклеточного соотноше
личение секреции IL 6, IL 8, хемокина CXCL1 и
ния [Na+]i/[K+]i является достаточным условием
лейкемия ингибирующего фактора [11]. Таким
изменения транскрипции десятков универсаль
образом, скелетные мышцы как эндокринный
ных и сотен тканеспецифических генов [28].
орган способны продуцировать цитокины и дру
В ряде случаев эти изменения транскрипции не
гие пептиды [12]. По мнению многих исследова
связаны с возможным приростом [Ca2+]i. Было
телей, эти соединения могут быть классифици
показано, что в условиях гипоксии наблюдается
рованы как миокины, которые оказывают раз
увеличение соотношения [Na+]i/[K+]i, что мо
личные физиологические эффекты на орга
жет быть достаточным условием для изменения
низм [13-15]. В то же время в литературе прак
транскриптома этих клеток [28]. Было обнару
тически не описана зависимость продукции мио
жено, что диссипация трансмембранного гради
кинов от характера (статическая и динамичес
ента одновалентных катионов, вызванная инги
кая) и от интенсивности физических нагрузок.
бированием Na+/K+ ATPазы, приводит к увели
В связи с многочисленными подтверждения
чению транскрипции генов PTGS2 и NR4A1 с
ми того факта, что именно сократительная ак
помощью Ca2+/кальмодулин опосредованного
тивность мышечных клеток является пусковым
фосфорилирования транскрипционного факто
механизмом продукции миокинов, внимание ис
ра, способного связывать CRE последователь
следователей привлекла проблема механохими
ности ДНК (CREB), и опосредованного кальци
ческого сопряжения, то есть механизма, опосре
нейрином дефосфорилирования ядерного фак
дующего образование и/или высвобождение этих
тора активированных Т клеток (NFAT) соответ
молекул при возбуждении/сокращении миоци
ственно [28]. В наших предыдущих работах
тов [16-18]. Существуют различные точки зре
[29-31] доказано влияние диссипации градиен
ния, объясняющие возможные механизмы, одна
тов одновалентных катионов на экспрессию ге
ко наше внимание привлекает гипотеза, предпо
нов и продукцию миокинов.
лагающая ключевую роль изменения внутрикле
Таким образом, можно считать установлен
точных концентраций одновалентных катионов
ным, что сократительная активность мышечных
в запуске продукции миокинов [17, 18].
клеток сопровождается диссипацией градиен
В многочисленных исследованиях было по
тов одновалентных катионов. В свою очередь,
казано, что как у человека, так и у эксперимен
изменение внутриклеточного содержания Nai+ и
тальных животных интенсивные физические
K+ служит пусковым фактором изменения тран
упражнения способствуют увеличению [Na+]i
скрипции генов [32-35].
в 3-4 раза и уменьшению [K+]i до 50% в скелет
Однако в литературе не описаны закономер
ных мышцах за счет активации ионных каналов,
ности диссипации ионных градиентов в зависи
а также путем частичного ингибирования
мости от характера и интенсивности физичес
Na+/K+ ATPазы. Было также продемонстриро
кой нагрузки, а также от предварительной тре
вано, что выход K+ из мышечных клеток во вре
нировки. В связи с этим целью исследования
мя упражнений приводит к повышению [K+] в
было выяснение характера влияния динамичес
интерстициальной жидкости скелетных мышц
ких и статических нагрузок на концентрацию
от 4-5 до 11-15 мМ. У людей интенсивные ди
миокинов в плазме крови и на содержание Nai+ и
намические и статические упражнения приво
K+ в скелетных мышцах мышей.
дят к 2× повышению уровня [K+] в венозной
крови из за его выхода из скелетных мышц, ко
торые являются основным источником внут
МАТЕРИАЛЫ И МЕТОДЫ
риклеточного K+ в организме [19-25].
В исследованиях Danilov et al. [26] было по
В качестве экспериментальных животных ис
казано, что EPS клеток скелетной мускулатуры
пользовались половозрелые (8-12 недель) мыши
мышей линии C2C12, широко используемой в
самцы линии C57Bl/6 массой 25-30 г (виварий
качестве модели мышечных сокращений in vitro,
НИИ фармакологии и регенеративной медицины
приводила к диссипации трансмембранного
имени Е.Д. Гольдберга СО РАН, Томск, Россия).
градиента одновалентных катионов. Через 2 и
Для проведения исследования были сформирова
4 ч EPS внутриклеточное содержание Nai+ увели
ны группы для острого (нетренированные мыши)
чилось со 130 до 330 и 500 нмоль/мг белка, тогда
и хронического (тренированные мыши) экспери
как внутриклеточное содержание K+ снизилось
ментов. Схема экспериментов и количество жи
с 1150 до 922 и 790 нмоль/мг белка соответствен
вотных в каждой группе представлены на рис. 1.
БИОХИМИЯ том 86 вып. 3 2021
ВЛИЯНИЕ НАГРУЗОК НА СОДЕРЖАНИЕ МИОКИНОВ, Na+ И K+
433
Схема эксперимента и количество животных в
нию, а плавание с грузом менее 5% способству
группах. В остром эксперименте мышей конт
ет обучению животных.
рольной группы (n = 5) не подвергали физичес
Умерщвление животных (методом декапита
кой нагрузке, мышей экспериментальной груп
ции) производили непосредственно после на
пы разделяли на две подгруппы, каждую из кото
грузки, а также через 1, 5 и 24 ч. Кровь собирали
рых подвергали воздействию однократной физи
непосредственно после декапитации в пробир
ческой нагрузки одного из двух видов: динами
ки с гепарином (20 ед./мл). Центрифугирование
ческая нагрузка в виде принудительного плава
образцов проводили в течение 11 мин при
ния [36, 37] с отягощениями 5, 7,5 и 10% от мас
2000
об./мин на Microfuge
16 Centrifuge
сы тела (n = 60) и статическая нагрузка в виде ви
(«Beckman Coulter», Германия).
са на сетке [37] с аналогичными отягощения
Определение концентрации белков в плазме
ми (n = 60). Время нагрузки с 5% ным грузом от
производили методом иммуноферментного ана
массы тела составляло 60 мин, с 7,5% ным гру
лиза (ИФА). Были использованы наборы для
зом - 20 мин и с 10% ным - 10 мин. В хроничес
ИФА с антителами к соответствующим белкам:
ком эксперименте всех мышей подвергали регу
IL 6 Platinum ELISA Kit, IL 8 Platinum ELISA
лярной физической нагрузке (тренировке) в ви
Kit («eBioscience», Австрия), а также RayBio®
де плавания [38] (n = 65) или виса на сет
IL 15 ELISA Kit («RayBio®», США). Выбор бел
ке (n = 65) (соответственно группе) в течение
ков для исследования был обусловлен литера
4 х недель по 1 ч в день без отягощения. После
турными данными о продукции миокинов при
этого мышей делили на две группы - контроль
физических нагрузках [2-8, 12], при стимуля
ную (n = 10) и экспериментальную (n = 120).
ции клеточной культуры миобластов [10, 11], а
Исследование выполняли по той же схеме, что и
также результатами наших исследований на
в первом (остром) эксперименте.
спортсменах [17].
Режимы хронических нагрузок подбирались
Определения концентрации ионов Na+ и K+.
на основании литературных данных, в частнос
Материалом для определения концентрации ио
ти работы Каркищенко с соавт. [39], в которой
нов служили мышцы, извлекаемые сразу же пос
показано, что наиболее оптимальной модифи
ле декапитации; в зависимости от характера на
кацией теста Портсолта для оценки физической
грузки извлекали 2 вида мышц: камбаловидную
выносливости крыс является плавание живот
мышцу (musculus soleus) - у мышей, подвергав
ного с грузом 10% от массы тела. Использование
шихся нагрузке в виде плавания, и двуглавую
груза более 15% приводит к быстрому утомле
мышцу (musculus biceps) - у мышей, подвергав
Рис. 1. Схема эксперимента и количество животных в группах
10 БИОХИМИЯ том 86 вып. 3 2021
434
КИРОНЕНКО и др.
шихся нагрузке в виде виса на сетке. Образцы
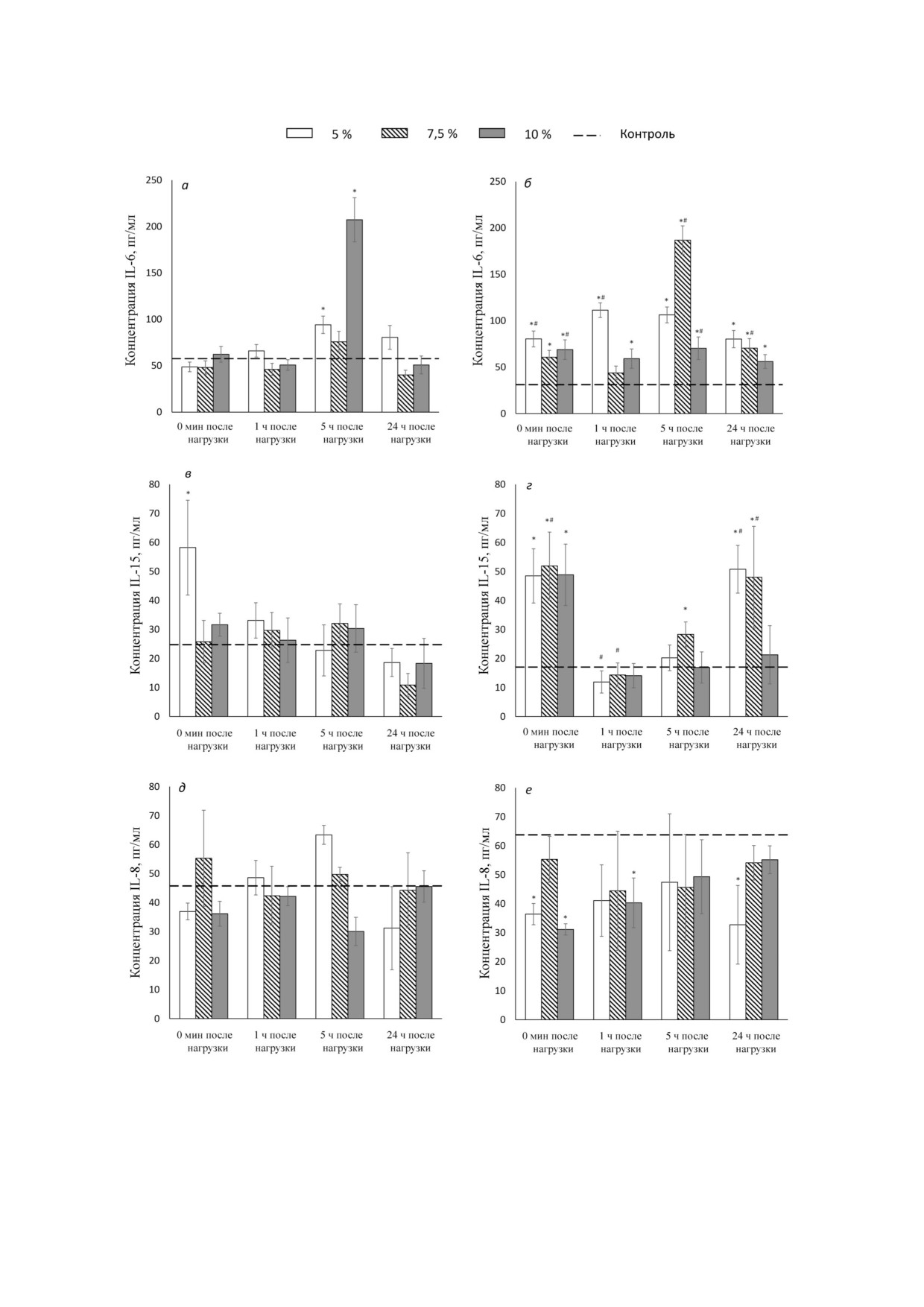
нированных мышей в различные сроки после
мышц для определения ионов забирали только
принудительного плавания. Продукция IL 6 у
непосредственно после нагрузки. Препарирова
нетренированных животных усиливается через
ние мышц проводили в PBS буфере на льду, об
5 ч после плавания (до 207,3 ± 23,7 пг/мл по срав
разцы помещали в промывочный раствор
нению с 57,64 ± 9,36 пг/мл в контроле, р < 0,01) и
(100 мМ MgCl2, 10 мМ Tris HCl, pH 7,4) на льду
пропорциональна степени утяжеления. Через
и инкубировали в течение 15 мин, затем раствор
24 ч после плавания концентрация IL 6 в плазме
меняли на свежий; данную процедуру повторяли
возвращалась к исходным значениям. У трениро
4 раза. Извлеченную мышцу взвешивали и раз
ванных животных исходный уровень данного ци
резали на мелкие части, после чего помещали в
токина в плазме ниже (33,8 ± 8,1 пг/мл против
микропробирку, содержащую 5% ный раствор
57,6 ± 9,3 пг/мл у нетренированных), однако
ТХУ, для полного извлечения ионов из ткани.
прирост после плавания происходит практичес
Через сутки содержимое микропробирки го
ки сразу и достигает 111,5 ± 7,9 пг/мл (р < 0,05),
могенизировали в стеклянном гомогенизаторе
хотя второй пик его концентрации через 5 ч пос
Поттера с тефлоновым пестиком, после чего до
ле плавания (106,3 ± 8,5 пг/мл, р < 0,05) сохраня
полнительно проводили ультразвуковую дезин
ется и в этом случае. Известно, что IL 6 индуци
теграцию образцов с использованием «Bradson
рует расщепление и окисление жиров, он также
450 Digital Sonifier» («Marshall Scientific», США) в
принимает участие в поддержании гомеостаза
течение 30 с при мощности 15% от максималь
глюкозы во время упражнений, поэтому его
ной амплитуды, процедуру повторяли 3 раза до
можно рассматривать как фактор, усиливающий
полной гомогенности суспензии. Полученный
энергообеспечение. Оказывая противовоспали
образец центрифугировали при 18 000 g на лабо
тельный эффект, IL 6 может ингибировать инсу
раторной центрифуге «ELMI CM 50» («ELMI»,
линорезистентность, индуцированную фактором
Латвия) в течение 10 мин, супернатант перено
некроза опухоли α (TNF α) [40].
сили в пробирку 1,5 мл для последующего изме
Концентрация IL 15 в плазме после принуди
рения концентрации ионов. К осадкам добавля
тельного плавания у нетренированных животных
ли лизирующий буфер, RIPA, содержащий кок
изменялась не столь значительно, снижение
тейль ингибиторов протеаз («Sigma Aldrich»,
фиксировалось только через
24
ч (до
USA). При определении концентрации белка ис
10,8 ± 4,0 пг/мл, р < 0,01) (рис. 2). Данное явле
пользовали модифицированный метод Лоури.
ние можно связать с эффектом компенсации, ко
Содержание Na+ и K+ в экстрактах ТХУ оп
торый наиболее выражен при первой нагрузке.
ределяли методом пламенной атомно абсорб
В группе мышей, подвергавшихся ежеднев
ционной спектрометрии с использованием
ному плаванию в течение 4 х недель, на фоне
спектрометра «Квант 2м1» («Кортек», Россия)
исходно сниженной до 17,1 ± 4,6 пг/мл (р < 0,01)
со смесью пропан воздух. Для калибровки ис
концентрации IL 15 в плазме ее увеличение не
пользовали растворы KCl (0,5-4 мг/литр K+)
посредственно после плавания (до
51,9
±
и NaCl (0,05-2 мг/литр Na+), содержащие
± 11,7 пг/мл, р < 0,01) сменялось выраженным
5% ТХУ. Содержание Na+ и K+ каждой пробы
падением до значений 11,9 ± 3,8 пг/мл (р < 0,01)
нормализовали по мокрому весу соответствую
через 1 ч и характеризовалось длительным про
щей мышцы. Все растворы готовили с исполь
цессом восстановления. IL 15 рассматривается
зованием дважды деионизованной воды.
как анаболический фактор, так как способен
Статистическую обработку данных проводи
стимулировать рост мышц. Кроме того, IL 15
ли с использованием пакета статистического
участвует в метаболизме липидов [36].
анализа STATISTICA 8.0. Уровень значимости
Концентрация IL 8 в плазме у нетренирован
при проверке гипотезы принадлежности двух
ных мышей не изменялась на фоне динамических
выборок к одной генеральной совокупности
нагрузок независимо от утяжеления (рис. 2). Пос
оценивали по Kruskal-Wallis ANOVA test. Дан
ле 4 х недель ежедневных плавательных нагрузок
ные представлены в виде среднего значе
фоновая концентрация данного интерлейкина
ния ± стандартная ошибка среднего.
возрастала до 63,75 ± 7,43 пг/мл (р < 0,05). После
однократной физической нагрузки у всех трени
рованных мышей мы наблюдали снижение
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
концентрации IL 8 в плазме. Непосредственно
после нагрузки это снижение было более вы
Динамика содержания интерлейкинов в плазме
ражено в группе с максимальным отягоще
у тренированных и нетренированных мышей. На
нием (63,75 ± 7,43 пг/мл в контроле (1),
рис. 2 представлена динамика содержания ин
36,42 ± 3,65 пг/мл с отягощением 5% (2),
терлейкинов в плазме у тренированных и нетре
55,30 ± 7,91 пг/мл с отягощением 7,5% (3),
БИОХИМИЯ том 86 вып. 3 2021
ВЛИЯНИЕ НАГРУЗОК НА СОДЕРЖАНИЕ МИОКИНОВ, Na+ И K+
435
Рис. 2. Концентрация интерлейкинов в плазме крови мышей после динамической нагрузки. а, в и д - Нетренированные
животные, б, г и е - тренированные животные. Сверху указаны величины дополнительной нагрузки (утяжеления) в про
центах от массы тела. Представлены средние значения ± SEM (пг/мл). Обозначения: * р1 < 0,05 - достоверность разли
чий соответствующего показателя по сравнению с контролем; # р2 < 0,05 - достоверность различий между тренированны
ми и нетренированными животными
БИОХИМИЯ том 86 вып. 3 2021
10*
436
КИРОНЕНКО и др.
31,16 ± 1,93 пг/мл с отягощением 10% (4);
раженный прирост концентрации данного ци
(1-4) - здесь и далее обозначены номера групп,
токина в плазме у всех групп мышей, однако в
для которых ниже указан уровень значимости
группах с утяжелением он был выражен в мень
p); p12 < 0,01; p13 < 0,05; p14 < 0,01; p23 < 0,01;
шей степени. Максимум достигался сразу после
p24 < 0,05; p34 < 0,01).
нагрузок, и хотя через 5 ч отмечалось снижение
На рис. 3 представлены данные об изменении
концентрации, она оставалась существенно вы
содержания интерлейкинов в плазме у трениро
ше фоновых значений до 24 ч эксперимен
ванных и нетренированных мышей в различные
та (рис. 2). После 4 х недель ежедневных стати
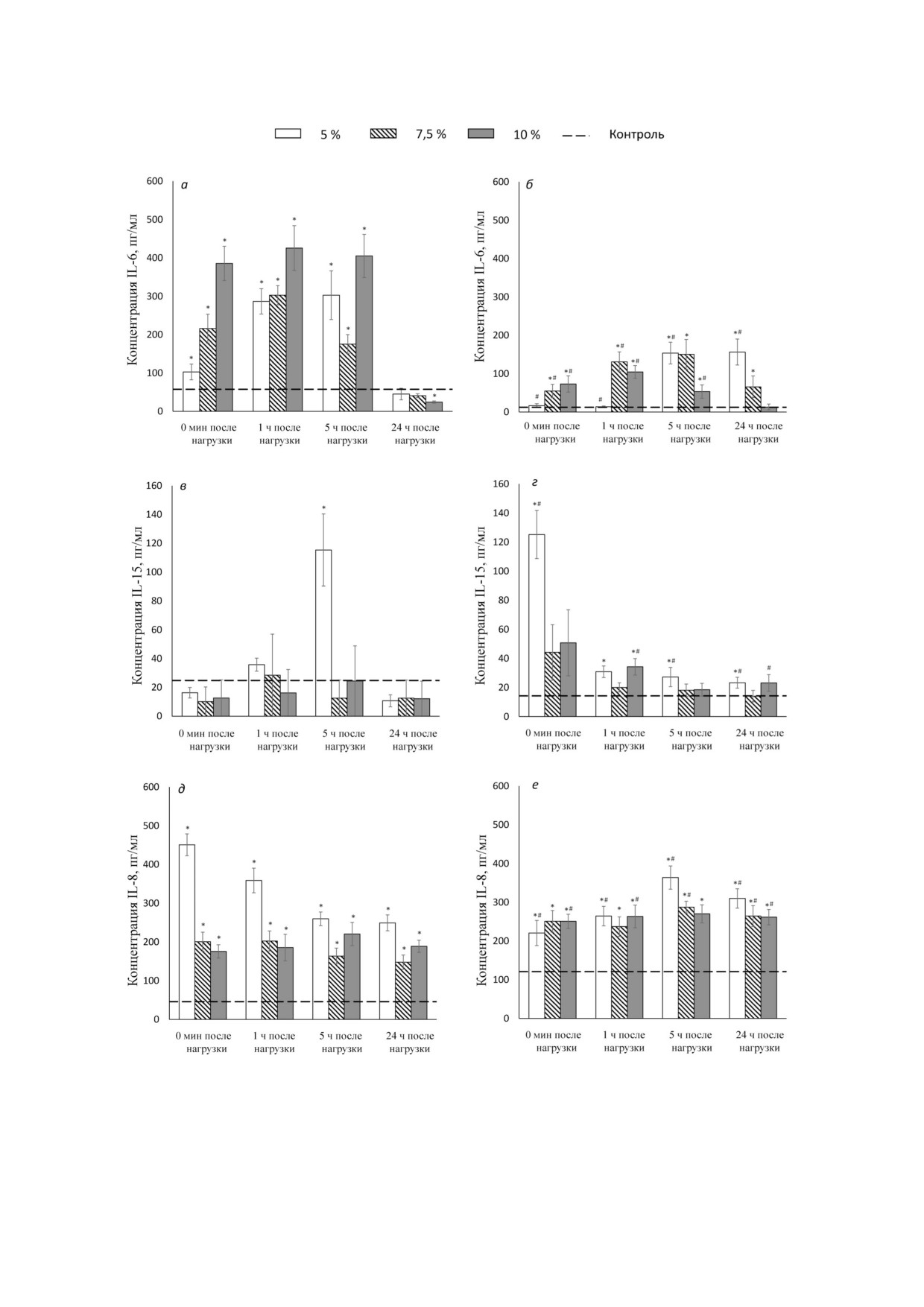
сроки после принудительной статической нагруз
ческих нагрузок фоновая концентрация данно
ки (вис на сетке). Продукция IL 6 у нетрениро
го интерлейкина в плазме возрастала до
ванных животных в этом случае достоверно уси
120,95 ± 30,62 пг/мл (р < 0,001) (рис. 3). После
ливалась сразу после нагрузки, причем увеличе
однократной статической нагрузки у всех тре
ние было пропорционально степени утяжеления.
нированных мышей мы наблюдали существен
Максимум концентрации достигался через
ный прирост концентрации IL 8 в плазме сразу
1-5 ч после нагрузки, к 24 ч наблюдалось значи
после нагрузки, который не зависел от степени
тельное снижение, однако уровень IL 6 оставал
утяжеления. Повышенная концентрация сохра
ся несколько выше исходных значений. У тре
нялась на прежнем уровне в течение 24 ч, в груп
нированных животных исходный уровень дан
пе без утяжеления дополнительный прирост
ного цитокина в плазме был значительно ниже
концентрации был зарегистрирован через 5 ч, в
(12,58 ± 4,33 пг/мл против 57,6 ± 9,3 пг/мл - у
других группах такого не отмечалось.
нетренированных, р < 0,01). Прирост после ста
Исследование действия физической нагрузки
тических нагрузок в группе тренированных жи
на концентрацию одновалентных ионов в мышцах
вотных был выражен в меньшей степени.
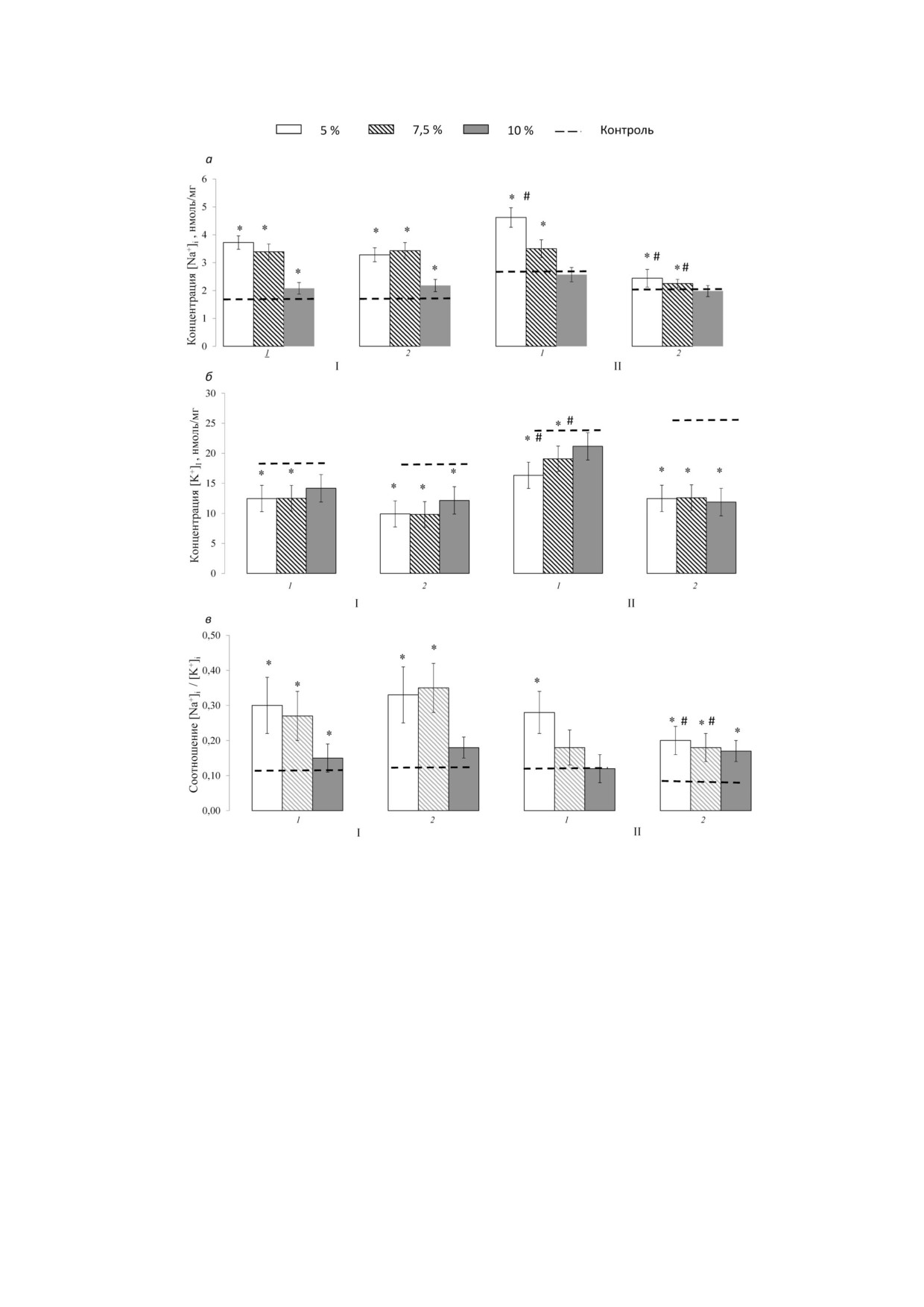
до и после нагрузки. Проведя исследование
Без утяжеления он регистрировался через 5 ч
действия физической нагрузки на концентра
после нагрузки, с утяжелением - сразу после
цию одновалентных ионов в мышцах до и после
нагрузки. Максимум концентрации IL 6 в груп
нагрузки, мы не обнаружили различий в содер
пе без утяжеления регистрировался через 5 ч
жании внутриклеточного Na+ и К+ в m. soleus и
после нагрузки и сохранялся на этом уровне до
m. biceps животных контрольной группы. На
24 ч, в группе с утяжелением 7,5% - через 1-5 ч,
рис. 4, а представлены значения внутриклеточ
в группе с утяжелением 10% - через 5 ч. Через
ного содержания Na+ в зависимости от характе
24 ч после нагрузки концентрация IL 6 в плазме
ра и интенсивности нагрузки у тренированных и
снижалась, а в группе с утяжелением 10% дости
нетренированных мышей. Содержание внут
гала фоновых значений (рис 2).
риклеточного Na+ и K+ в группах нетренирован
Концентрация IL 15 в плазме после стати
ных животных в состоянии покоя было ниже,
ческих нагрузок у нетренированных животных
чем у тренированных мышей. Хронические на
изменялась не столь значительно, сразу после
грузки динамического характера сопровожда
нагрузки отмечалась тенденция к снижению
лись возрастанием концентрации внутрикле
концентрации данного цитокина, однако через
точного Na+ в большей степени, чем статичес
час в группе без утяжеления она сменялась рос
кие. Влияние двух типов хронических нагрузок
том - максимум достигался через 5 ч, после че
на концентрацию внутриклеточного К+ в клет
го опять регистрировалось снижение - через
ках скелетных мышц не различалось.
24 ч концентрация во всех группах была вдвое
Концентрация внутриклеточного Na+ в
ниже исходного уровня (рис. 3).
мышцах в группе тренированных животных,
В группе мышей, подвергавшихся ежеднев
подвергавшихся динамической нагрузке с ма
ным тренировочным статическим нагрузкам в
лыми утяжелениями, была выше, чем у трени
течение 4 х недель, на фоне исходно сниженной
рованных животных. В то же время после на
до 14,27 ± 3,62 пг/мл (р < 0,05) концентрации IL
грузки с утяжелением в 10% содержание ионов
15 в плазме отмечался ее прирост непосредствен
Na+ практически не изменялось. Сходные ре
но после однократной нагрузки у всех мышей, но
зультаты были получены и в группе животных,
максимальным он был в группе без утяжеления.
подвергавшихся статическим нагрузкам.
Однако уже через 5 ч после нагрузки концентра
Необходимо отметить, что содержание ио
ция IL 15 в плазме значительно снижалась, пос
нов Na+ у тренированных мышей было выше,
тепенно достигая фоновых значений (рис. 3).
чем у нетренированных. При этом содержание
В отличие от динамических нагрузок, кото
ионов Na+ у группы, подвергавшейся динами
рые практически не влияли на концентрацию
ческой нагрузке, численно выше по всем под
IL 8 в плазме у нетренированных мышей, сразу
группам, чем у группы, подвергавшейся стати
после статических нагрузок мы наблюдали вы
ческой нагрузке.
БИОХИМИЯ том 86 вып. 3 2021
ВЛИЯНИЕ НАГРУЗОК НА СОДЕРЖАНИЕ МИОКИНОВ, Na+ И K+
437
Рис. 3. Концентрация интерлейкинов в плазме крови мышей после статической нагрузки. а, в и д - Нетренированные жи
вотные, б, г и е - тренированные животные. Сверху указаны величины дополнительной нагрузки (утяжеления) в процен
тах от массы тела. Представлены средние значения ± SEM (пг/мл). Обозначения: * р1 < 0,05 - достоверность различий со
ответствующего показателя по сравнению с контролем; # р2 < 0,05 - достоверность различий между тренированными и
нетренированными животными
БИОХИМИЯ том 86 вып. 3 2021
438
КИРОНЕНКО и др.
Содержание внутриклеточного К+ в мышцах
ганизме животных, вызванными регулярными
в группах нетренированных животных, подверг
физическими нагрузками [13, 42].
шихся однократной динамической или стати
В третьих, как динамические, так и статичес
ческой нагрузке, было ниже, чем у животных из
кие нагрузки влияют на содержание одновалент
контрольной группы (не подвергавшихся ка
ных катионов в клетках скелетных мышц, вызы
кой либо нагрузке) (рис. 4, б). Существеннее
вая увеличение внутриклеточного соотноше
всего уровень К+ в мышцах снизился в подгруп
ния Nai+/K+, однако при динамической нагрузке
пе после нагрузки без утяжеления, в то время
это эффект выражен в большей степени. При
как наименьшее снижение произошло в под
этом содержание внутриклеточного Na+ и K+ в
группе после нагрузки в 10%. Уменьшение содер
группах нетренированных животных в состоянии
жания К+ в мышцах после статической нагрузки
покоя было ниже, чем у тренированных мышей.
было выражено в большей степени, чем после
Особенности влияния динамической и стати
динамической. Были выявлены статистически
ческой нагрузки на продукцию миокинов у нетре
значимые различия в содержании ионов К+ в
нированных и тренированных животных могут
мышцах между группами нетренированных и
быть связаны с различиями в клеточном составе
тренированных животных, подвергшихся как
мышц. Скелетная мышца весьма гетерогенна -
динамической, так и статической нагрузке. Не
кроме миоцитов, она содержит целый ряд кле
обходимо отметить, что содержание ионов К+ в
ток - фибробласты, перициты, адипоциты, чей
мышцах у нетренированной группы снизилось
вклад в общий объем производства цитокинов
сильнее, чем у тренированной.
изучен недостаточно [42]. Относительное содер
Выраженное увеличение внутриклеточного
жание этих клеток, а также их воздействие на выз
соотношения Nai+/K+ в мышцах было зарегист
ванную физической нагрузкой продукцию цито
рировано как после динамических, так и после
кинов может изменяться в результате регулярных
статических нагрузок, причем при высоких вели
физических нагрузок. В дополнение к гетероген
чинах утяжеления оно было выражено в меньшей
ности ткани было показано, что скелетные мио
степени (рис. 4, в). Уровень внутриклеточного
циты могут быть подразделены на 3 различных
соотношения Nai+/K+ в мышцах в группе, подвер
фенотипа, каждый из которых имеет свои особен
гавшейся динамической нагрузке, выше, чем у
ности биоэнергетических механизмов [43, 44].
группы, подвергавшейся статической нагрузке.
Еще один фактор, обеспечивающий описан
ные различия, может быть связан с особенностя
ми транскрипционных механизмов. Показано,
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
что транскрипционные изменения при сокраще
нии мышц наиболее выражены в быстро сокра
Полученные нами результаты позволяют
щающихся мышечных волокнах типа IIa [45],
сделать три основных вывода. Во первых, влия
что также, несомненно, отражается на интен
ние статических и динамических нагрузок на
сивности продукции миокинов. Фактор, инду
содержание в плазме цитокинов значительно
цируемый гипоксией 1 альфа (HIF 1α), сAMP и
различается. Длительные динамические упраж
Ca2+ зависимые протеинкиназы, а также соот
нения вызывают увеличение содержания IL 6 в
ношение концентраций одновалентных катио
плазме нетренированных мышей. Это согласу
нов ([Na+]i/[K+]i) обеспечивают передачу сигна
ется с данными ряда публикаций [5, 6, 41], в ко
лов и участвуют в образовании транскрипцион
торых исследовалось содержание IL 6 в плазме у
ных связей в клетках скелетных мышц [31, 39, 46,
спортсменов - бегунов. В отличие от динами
47]. HIF 1α транслоцируется в ядро, где образу
ческих, статические нагрузки оказывают выра
ет комплекс HIF 1α/HIF 1β и триггеры тран
женное влияние на концентрацию всех исследо
скрипции десятков генов, в том числе фактора
ванных миокинов.
роста эндотелия сосудов (VEGF) и эндотелиаль
Во вторых, как динамические, так и стати
ной синтазы окиси азота (eNOS) [48]. Важно от
ческие нагрузки по разному влияют на содер
метить, что в отличие от динамической нагруз
жание цитокинов в плазме крови нетренирован
ки, статические нагрузки приводят к окклюзии
ных и тренированных животных. Так, в плазме у
кровеносных сосудов и локальной гипоксии, что
тренированных животных после динамической
сопровождается накоплением mRNA фактора
нагрузки наблюдался значительный прирост
роста эндотелия сосудов (VEGF) и эндотелиаль
концентрации IL 15 и снижение концентрации
ной синтазы окиси азота в скелетной мышце
IL 8, тогда как у нетренированных животных
крысы [49]. Однако роль этого пути в регулиро
подобная реакция на однократную динамичес
вании продукции цитокинов еще не изучена.
кую нагрузку отсутствовала. Эти факты можно
Возбуждение миоцитов сопровождается из
объяснить адаптационными изменениями в ор
менением трансмембранного градиента однова
БИОХИМИЯ том 86 вып. 3 2021
ВЛИЯНИЕ НАГРУЗОК НА СОДЕРЖАНИЕ МИОКИНОВ, Na+ И K+
439
Рис. 4. Величины внутриклеточных концентраций Na+ (а), К+ (б) и соотношения Na+/К+ (в) после динамической (1) и
статической (2) нагрузки у нетренированных (I) и тренированных (II) мышей. Вверху указаны величины дополнительной
нагрузки (утяжеления) в процентах от массы тела. Концентрация нормирована на влажный вес мышцы. Представлены
средние значения ± SEM. Обозначения: * р1 < 0,05 - достоверность различий соответствующего показателя по сравнению
с контролем; # р2 < 0,05 - достоверность различий между тренированными и нетренированными животными
лентных катионов вследствие притока Na+ и от
тенциал чувствительных Са2+ каналов и/или
тока K+ через потенциал зависимые и Са2+
Na+/Ca2+ обмена.
чувствительные ионные каналы. В некоторых
Ранее для оценки относительного вклада
типах клеток повышение соотношения
Ca2+ опосредованного и Ca2+ независимого сиг
[Na+]i/[K+]i приводило к экспрессии mRNA ря
нальных путей мы сравнили транскриптомные
да цитокинов, в том числе IL 6 [50, 51]. Это поз
изменения при увеличении соотноше
воляет предположить, что увеличение соотно
ния [Na+]i/[K+]i в клетках, обеднённых Ca2+, и
шения [Na+]i/[K+]i является фактором, стимули
обнаружили увеличение количества специфи
рующим продукцию миокинов. Этот процесс
ческих [Na+]i/[K+]i чувствительных генов [52].
может опосредоваться возрастанием концентра
Среди [Na+]i/[K+]i чувствительных генов, акти
ции [Ca2+]i в цитоплазме за счёт активации по
вируемых независимо от наличия хелато
БИОХИМИЯ том 86 вып. 3 2021
440
КИРОНЕНКО и др.
ров Са2+, был обнаружен ген миокина, IL 6. Не
миокинов. Однако обнаруженное влияние фи
давно также было выявлено, что внеклеточные
зических нагрузок на диссипацию трансмемб
хелаторы Ca2+ значительно увеличивают прони
ранных градиентов одновалентных катионов
цаемость мембраны для одновалентных ионов,
позволяет предположить, что определенную
что приводит к повышению внутриклеточного
роль в этом могут играть [Na+]i/[K+]i зависимые
соотношения [Na+]i/[K+]i [53]. Ранее мы также
механизмы запуска транскрипции генов.
показали, что в клетках гладких мышц сосудов
Модификация этих механизмов под влияни
транскриптомные изменения, индуцированные
ем физических нагрузок различного характера и
гипоксией, по меньшей мере частично вызваны
интенсивности представляет значительный ин
HIF 1α независимыми, [Na+]i/[K+]i опосредо
терес, так как является перспективным путем
ванными, транскрипционными связями [54].
воздействия на процессы метаболизма как на
Однако молекулярная природа сенсоров,
клеточном, так и на системном уровнях, что
посредством которых осуществляется регуляция
весьма актуально как для повышения спортив
процессов транскрипции и трансляции внут
ных результатов, так и для коррекции метаболи
риклеточными Na+ и K+, до сих пор остаётся
ческих расстройств при целом ряде социально
предметом дискуссии [55]. В данном аспекте ин
значимых заболеваний.
тересен тот факт, что увеличение соотноше
ния [Na+]i/[K+]i в цитоплазме клеток скелетных
мышц при сокращении сопровождается увели
Финансирование. Исследование выполнено
чением активности α2 Na+/K+ ATPазы [56], ко
при финансовой поддержке Российского науч
торая способна частично нивелировать этот эф
ного фонда (грант № 16 15 10026 П).
фект благодаря высокому сродству этой изофор
Благодарности. Авторы приносят благодар
мы фермента к [Na+]i [57].
ность профессору Александру Чибалину (Каролин
ский Институт, Стокгольм, Швеция) за высказан
Подводя итог сказанному, следует отметить,
ные ценные идеи при обсуждении результатов.
что спектр вырабатываемых миокинов зависит
Конфликт интересов. Авторы заявляют об от
от характера нагрузок и уровня предваритель
сутствии конфликта интересов.
ной тренировки. Динамические и статические
Соблюдение этических норм. Все процедуры,
нагрузки по разному влияют на содержание мио
выполненные в исследованиях с участием жи
кинов в плазме крови нетренированных и пред
вотных, соответствовали этическим стандартам
варительно тренированных экспериментальных
учреждения, в котором проводились исследова
животных. Обнаруженные различия могут
ния, и утвержденным правовым актам РФ и
иметь в своей основе целый ряд механизмов.
международных организаций. Исследование
Клеточный состав скелетных мышц и феноти
было одобрено локальным этическим комите
пические особенности мышечных волокон, из
том биологического института Томского государ
меняясь в результате регулярных тренировок,
ственного университета (протокол № 32 от
могут модифицировать процессы продукции
19 декабря 2019 года).
СПИСОК ЛИТЕРАТУРЫ
1.
Frontera, W. R., and Ochala, J. (2015) Skeletal muscle: a
6. Steensberg, A., van Hall, G., Osada, T., Sacchetti, M.,
brief review of structure and function, Calcif. Tissue Int.,
Saltin, B., and Klarlund Pedersen, B. (2000) Production of
96, 183 195, doi: 10.1007/s00223 014 9915 y.
interleukin 6 in contracting human skeletal muscles can
2.
Amano, Y., Nonaka, Y., Takeda, R., Kano, Y., and
account for the exercise induced increase in plasma inter
Hoshino, D. (2020) Effects of electrical stimulation
leukin 6, J. Physiol., 529, 237 242, doi: 10.1111/j.1469
induced resistance exercise training on white and brown
7793.2000.00237.x.
adipose tissues and plasma meteorin like concentration in
7. Keller, C., Steensberg, A., Pilegaard, H., Osada, T.,
rats, Physiol. Rep., 16, e14540, doi: 10.14814/phy2.14540.
Saltin, B., et al. (2001) Transcriptional activation of the IL
3.
Drenth, J. P., Van Uum, S. M, Van Deuren, M., Pesman,
6 gene in human contracting skeletal muscle: influence of
G., Van der Ven Jongekrijg, J., and Van der Meer, J. M.
muscle glycogen content, FASEB J., 14, 27482750,
(1985) Endurance run increases circulation IL 6 and IL
doi: 10.1096/fj.01 0507fje.
1ra but downregulates ex vivo TNF alpha and IL 1beta
8. Nedachi, T., Fujita, H., and Kanzaki, M. (2008) Contractile
production, J. Appl. Physiol., 79, 1497 1503, doi: 10.1152/
C2C12 myotube model for studying exercise inducible
JAPPL.1995.79.5.1497.
responses in skeletal muscle, Am. J. Physiol. Endocrinol.
4.
Romagnoli, C., Zonefrati, R., and Sharma, P. (2020)
Metab., 295, E1191 E1204, doi: 10.1152/ajpendo.90280.2008.
Characterization of skeletal muscle endocrine control in an
9. Lambernd, S., Taube, A., Schober, A., Platzbecker, B.,
in vitro model of myogen, Calcif. Tissue Int., 107, 18 30,
Görgens, S. W., et al. (2012) Contractile activity of human
doi: 10.1007/s00223 020 00678 3.
skeletal muscle cells prevents insulin resistance by inhibit
5.
Barbalho, S. M., Flato, U. A. P., Tofano, R. J., de Alvares
ing pro inflammatory signaling pathways, Diabetologia, 55,
Goulart, R., Guiguer, E. L., et al. (2020) Physical exercise
1128 1139, doi: 10.1007/s00125 012 2454 z.
and myokines, Int. J. Molec. Sci.,
21,
3607,
10. Nikolic, N., Bakke, S. S., Kase, E.T., Rudberg, I.,
doi: 10.3390/ijms21103607.
Halle, I. F., et al. (2012) Electrical pulse stimulation of cul
БИОХИМИЯ том 86 вып. 3 2021
ВЛИЯНИЕ НАГРУЗОК НА СОДЕРЖАНИЕ МИОКИНОВ, Na+ И K+
441
tured human skeletal muscle cells as an in vitro model of
contraction induced IL 6 up regulation using contractile
exercise, PLoS One, 7, e33203, doi: 10.1371/journal.
C2C12 myotubes, Endocr. J., 60, 137 147, doi: 10.1507/
pone.0033203.
endocrj.ej12 0316.
11.
Scheler, M., Irmler, M., Lehr, S., Hartwig, S., Staiger, H.,
28.
Smolyaninova, L.V., Koltsova, S. V., Sidorenko, S. V., and
et al. (2013) Cytokine response of primary human myotubes
Orlov, S. N. (2017) Augmented gene expression triggered
in an in vitro exercise model, Am. J. Physiol. Cell. Physiol.,
by Na+,K+ ATPase inhibition: role of Ca2+ mediated and
305, C877 C886, doi: 10.1152/ajpcell.00043.2013.
independent excitation transcription coupling, Cell
12.
Ahn, N., and Kim, K. (2020) Effects of aerobic and resis
Calcium., 68, 5 13, doi: 10.1016/j.ceca.2017.10.002.
tance exercise on myokines in high fat diet induced mid
29.
Sidorenko, S., Klimanova, E., Milovanova, K., Lopina,
dle aged obese rats, Int. J. Environ. Res. Public Health, 17,
O. D., Kapilevich, L. V., et al. (2018) Transcriptomic
2685, doi: 10.3390/ijerph17082685.
changes in C2C12 myotubes triggered by electrical stimu
13.
Pedersen, B. K., and Febbraio, M. A. (2008) Muscle as an
lation: role of Ca2+ mediated and Ca2+ independent sig
endocrine organ: focus on muscle derived interleukin 6,
naling and elevated [Na+]i/[K+]i ratio, Cell Calcium, 76,
Physiol. Rev.,
88,
13791406, doi:
10.1152/physrev.
72 86, doi: 10.1016/j.ceca.2018.09.007.
90100.2007.
30.
Kapilevich, L. V., Kironenko, T. A., Zaharova, A. N.,
14.
Iizuka, K., Machida, T., and Hirafuji, M. (2014) Skeletal
Kotelevtsev, Y. V., Dulin, N. O., and Orlov, S. N. (2015)
muscle is an endocrine organ, J. Pharmacol. Sci., 125, 125
Skeletal muscle as an endocrine organ: role of
131, doi: 10.1254/jphs.14r02cp.
[Na+]i/[K+]i mediated excitation transcription coupling,
15.
Pedersen, B. K., and Febbraio, M. A. (2012) Muscles,
Genes Dis., 2, 328 336, doi: 10.1016/j.gendis.2015.10.001.
exercise and obesity: skeletal muscle as a secetory organ,
31.
Lauritzen, H. P., Brandauer, J., Schjerling, P., Koh, H. J.,
Nat. Rev. Endocrinol., 8, 457 465, doi: 10.1038/nrendo.
Treebak, J. T., et al. (2013) Contraction and AICAR stim
2012.49.
ulate IL 6 vesicle depletion from skeletal muscle fibers
16.
Laurens, C., Bergouignan, A., and Moro, C. (2020)
in vivo, Diabetes, 62, 3081 3092, doi: 10.2337/db12 1261.
Exercise released myokines in the control of energy
32.
Kapilevich, L. V., Milovanova, K. G., Sidorenko, S. V.,
metabolism, Front. Physiol.,
11,
91, doi:
10.3389/
Fedorov, D. A., Kironenko, T. A., et al. (2020) Effect of
fphys.2020.00091.
dynamic and static loads on the concentration of sodium
17.
Kapilevich, L. V., Zakharova, A. N., Kabachkova, A. V.,
and potassium in murine skeletal muscles, Bull. Exp. Biol.
Kironenko, T. A., and Orlov, S. N. (2017) Dynamic and
Med., 169, 1 4, doi: 10.1007/s10517 020 04811 y.
static exercises differentially affect plasma cytokine con
33.
Klimanova, E. A., Sidorenko, S. V., Tverskoi, A. M.,
tent in elite endurance and strength trained athletes and
Shiyan, A. A., Smolyaninova, L. V., et al. (2019) Search for
untrained volunteers, Front. Physiol., 8, 35, doi: 10.3389/
intracellular sensors involved in the functioning of monova
fphys.2017.00035.
lent cations as secondary messengers, Biochemistry (Moscow),
18.
Kapilevich, L. V., Kironenko, T. A., Zakharova, A. N.,
84, 1280 1295, doi: 10.1134/S0006297919110063.
Kabachkova, A. V., and Orlov, S. N. (2017) Level of inter
34.
Shiyan, A. A., Sidorenko, S. V., Fedorov, D., Klimanova,
leukins IL 6 and IL 15 in blood plasma of mice after
E. A., Smolyaninova, L. V., et al. (2019) Elevation of intra
forced swimming test, Bull. Exp. Biol. Med., 163, 10 13,
cellular Na+ contributes to expression of early response
doi: 10.1007/s10517 017 3725 y.
genes triggered by endothelial cell shrinkage, Cell. Physiol.
19.
Jurkat Rott, K., Fauler, M., and Lehmann Horn, F.
Biochem., 53, 638 647, doi: 10.33594/000000162.
(2006) Ion channels and ion transporters of the transverse
35.
Smolyaninova, L. V., Shiyan, A. A., Kapilevich, L. V.,
tubular system of skeletal muscle, J. Muscle Res. Cell Motil.,
Lopachev, A. V., Fedorova, T. N., et al.
(2019)
27, 275 290, doi: 10.1007/s10974 006 9088 z.
Transcriptomic changes triggered by ouabain in rat cere
20.
Sejersted, O. M., and Sjogaard, G. (2000) Dynamics and
bellum granule cells: role of α3 and α1 Na+,K+ ATPase
consequences of potassium shifts in skeletal muscle and
mediated signaling, PLoS One, 14, e0222767, doi: 10.1371/
heart during exercise, Physiol. Rev., 80, 14111481,
journal.pone.0222767.
doi: 10.1152/physrev.2000.80.4.1411.
36.
Lukaszuk, B., Bialuk, I., Górski, J., Zaj czkiewicz, M.,
21.
McDonough, A. A., Thompson, C. B., and Youn, J. H.
Winnicka, M. M., and Chabowski, A. (2012) A single bout
(2002) Skeletal muscle regulates extracellular potassium,
of exercise increases the expression of glucose but not fatty
Am. J. Physio.l Ren. Physiol.,
282, F967 F974,
acid transporters in skeletal muscle of IL 6 KO mice,
doi: 10.1152/ajprenal.00360.2001.
Lipids, 47, 763 772, doi: 10.1007/s11745 012 3678 x.
22.
McKenna, M. J., Bangsbo, J., and Renaud, J. M. (2008)
37.
Bagmetova, V. V., Krivitskaya, A. N., Tyurenkov, I. N.,
Muscle K+, Na+, and Cl- disturbances and Na+ K+ pump
Berestovitskaya, V. M., and Vasilyeva, O. S. (2012) The
inactivation: implications for fatigue, J. Appl. Phys., 104,
influence of fenibut and its salt with succinic acid on ani
288 295, doi: 10.1152/japplphysiol.01037.2007.
mals’ resistance to forced dynamic and static physical
23.
Murphy, K. T., Nielsen, O. B., and Clausen, T. (2008)
loads, Fundament. Res., 4, 243 246.
Analysis of exerciseinduced Na+ K+ exchange in rat skele
38.
Wasinski, F., Bacurau, R. F., Moraes, M. R., Haro, A. S.,
tal muscle, Exp. Physiol., 93, 1249 1262, doi: 10.1113/
Moraes Vieira, P. M., et al. (2013) Exercise and caloric
expphysiol.2008.042457.
restriction alter the immune system of mice submitted
24.
Cairns, S. P., and Lindinger, M. I. (2008) Do multiple
to a high fat diet, Mediators Inflam., 2013, 395672,
ionic interactions contribute to skeletal muscle fatigue?
doi: 10.1155/2013/395672.
J. Physiol.,
586,
40394054, doi:
10.1113/jphysiol.
39.
Karkishchenko, V. N., Kapanadze, G. D., Dengina, S. E.,
2008.155424.
and Stankova, N. V. (2011) Development of technique of
25.
Orlov, S. N., Koltsova, S. V., Kapilevich, L. V., Gusakova,
assessment of physical endurance of small laboratory ani
S. V., and Dulin, N. O. (2015) KCC1 and NKCC2: the
mals for investigation of adaptogenic activity of some ther
pathogenetic role of cation chloride cotransporters in
apeutic drugs, Biomeditsina, 1, 72 74.
hypertension, Genes Dis., 2, 186196, doi: 10.1016/
40.
Gundersen, K. (2011) Excitation transcription coupling in
j.gendis.2015.02.007.
skeletal muscle: the molecular pathways of exercise, Biol.
26.
Danilov, K., Sidorenko, S., Milovanova, K., Klimanova, E.,
Rev. Camb. Philos. Soc., 86, 564 600, doi: 10.1111/j.1469
Kapilevich, L., and Orlov, S. (2017) Electrical pulse stimu
185X.2010.00161.x.
lation decreases electrochemical Na+ and K+ gradients in
41.
Fisher, C. P. (2006) Interleikin 6 in acute exercise and
C2C12 myotubes, Biochem. Biophys. Res. Commun., 493,
training: what is the biological relevance? Exercise
875 878, doi: 10.1016/j.bbrc.2017.09.133.
Immunol. Rev., 12, 6 33.
27.
Farmawati, A., Kitajima, Y., Nedachi, T., Sato, M.,
42.
Peake, J. M., Gatta, P. D., Suzuki, K., and Nieman, D. C.
Kanzaki, M., and Nagatomi, R. (2013) Characterization of
(2015) Cytokine expression and secretion by skeletal mus
БИОХИМИЯ том 86 вып. 3 2021
442
КИРОНЕНКО и др.
cle cells: regulatory mechanisms and exercise effects,
50. Koltsova, S. V., Trushina, Y., Haloui, M., Akimova, O. A.,
Exercise Immunol. Rev., 21, 8 25.
Tremblay, J., et al. (2012) Ubiquitous [Na+]i/[K+]i sensi
43.
Fitts, R. H., and Widrick, J. J. (1996) Muscle mechanics:
tive transcriptome in mammalian cells: evidence for
adaptations with exercise training, Exerc. Sport. Sci. Rev.,
[Ca2+]i independent excitation transcription coupling,
24, 427 473.
PLoS One, 7, e38032, doi: 10.1371/journal.pone.0038032.
44.
Dyakova, E. Y., Kapilevich, L. V., Shylko, V. G., Popov, S.
51. Popov, D. V., Lysenko, E. A., Bokov, R. O., Volodina, M.
V., and Anfinogenova, Y. (2015) Physical exercise associat
A., Kurochkina, N. S., et al. (2018) Effect of aerobic train
ed with NO production: signaling pathways and signifi
ing on baseline expression of signaling and respiratory pro
cance in health and disease, Front. Cell Dev. Biol., 3, 19,
teins in human skeletal muscle, Physiol. Rep., 6, e13868,
doi: 10.3389/fcell.2015.00019.
doi: 10.14814/phy2.13868.
45.
Raue, U., Trappe, T. A., Estrem, S. T., Qian, H. R.,
52. Popov, D. V., Makhnovskii, P. A., Shagimardanova, E. I.,
Helvering, L. M., et al. (2012) Transcriptomic signature of
and Volodina, M. A. (2019) Contactile activity specific
resistance exercise adaptations: mixed muscle and fiber
transcriptome response to acute endurance exercise and
type specific profiles in young and old adults, J. Appl.
training in human skeletal muscle, Am. J. Physiol.
Physiol.,
112,
16251636, doi:
10.1152/japplphysiol.
Endocrinol. Metab., 316, E605 E614, doi: 10.1152/ajpendo.
00435.2011.
00449.2018.
46.
Kapilevich, L. V., D’Yakova, E. Yu., Nosarev, A. V.,
53. Koltsova, S. V., Tremblay, J., Hamet, P., and Orlov, S. N.
Zaitseva, T. N., Petlina, Z. R., et al. (2010) Effect of nano
(2015) Transcriptomic changes in Ca2+ depleted cells: role
disperse ferrite cobalt (CoFe2O4) particles on contractile
of elevated intracellular [Na+]/[K+] ratio, Cell Calcium, 58,
reactions in guinea pigs airways, Bull. Exp. Biol. Med., 149,
317 324, doi: 10.1016/j.ceca.2015.06.009.
70 72.
54. Koltsova, S. V., Shilov, B., Burulina, J. G., Akimova, O. A.,
47.
Kovalev, I. V., Baskakov, M. B., Medvedev, M. A.,
Mounsif, H., et al. (2014) Transcriptomic changes trig
Minochenko, I. L., Kilin, A. A., et al.
(2007)
gered by hypoxia: evidence for HIF 1α independent,
Na+,K+,2Cl(-) cotransport and chloride permeability of
[Na+]i/[K+]i mediated excitation transcription coupling,
the cell membrane in mezaton and histamine regulation of
PLoS One,
9, e110597, doi:
10.1371/journal.pone.
electrical and contractile activity in smooth muscle cells
0110597.
from the guinea pig ureter, Ros. Fiziol. Zh. Im.
55. Orlov, S. N., and Hamet, P. (2015) Salt and gene expres
I.M. Sechenova, 93, 306 317.
sion: evidence for [Na+]i/[K+]i mediated signaling path
48.
Ke, Q., and Costa, M. (2006) Hypoxia inducible factor 1
ways, Pflugers Arch., 467, 489 498, doi: 10.1007/s00424
(HIF 1), Mol. Pharmacol., 70, 1469 1480, doi: 10.1124/
014 1650 8.
mol.106.027029.
56. Kravtsova, V. V., Petrov, A. M., Matchkov, V. V., Timonina,
49.
Rodriguez Miguelez, P., Lima Cabello, E., Martinez
N. A., Zakyrjanova, G. F., et al. (2016) Distinct α2 Na,K
Florez, S., Almar, M., Cuevas, M. J., and González
ATPase membrane pool are differently involved in early
Gallego, J. (2015) Hypoxia inducible factor 1 modulates
skeletal muscle remodeling during disuse, J. Gen. Physiol.,
the expression of vascular endothelial growth factor and
147, 175 188, doi: 10.1085/jgp.201511494.
endothelial nitric oxide synthase induced by eccentric
57. Matchkov, V. V., and Krivoi, I. I. (2016) Specialized func
exercise, J. Appl. Physiol., 118, 1075 1083, doi: 10.1152/
tional diversity and interactions of Na,K ATPase, Front.
japplphysiol.00780.2014.
Physiol., 7, 179, doi: 10.3389/fphys.2016.00179.
INFLUENCE OF DYNAMIC AND STATIC LOADS
ON THE CONCENTRATION OF MYOKINES IN PLASMA
AND ON THE CONTENT OF SODIUM AND POTASSIUM
IN THE SKELETAL MUSCLES OF MICE
T. A. Kironenko1, K. G. Milovanova1, A. N. Zakharova1, S. V. Sidorenko2,
E. A. Klimanova1,2, E. Yu. Dyakova1, A. A. Orlova1, E. S. Negodenko1,
Yu. G. Kalinnikova1, S. N. Orlov1,2
, and L. V. Kapilevich1*
1 National Research Tomsk State University, 634050 Tomsk, Russia, E mail: kapil@yandex.ru
2 Faculty of Biology, Lomonosov Moscow State University, 119234 Moscow, Russia; E mail: klimanova.ea@yandex.ru
Modification of the production of cytokines under the influence of physical activity is of considerable interest, since
it is a promising way of correcting metabolic processes at both the cellular and systemic levels. The content of IL 6,
IL 8, and IL 15 in plasma and monovalent cations in skeletal muscle in mice was studied at different times after sta
tic and dynamic loads, taking into account their weighting and preliminary training. Long term dynamic exercises
cause an increase in the content of IL 6 and a decrease in IL 15 in the plasma of intact mice, without affecting the
concentration. In trained mice, the effects of a single load on the concentration of IL 6 and IL 15 in plasma were
enhanced, and a decrease in the concentration of IL 8 was also noted. Unlike dynamic, static loads have a similar,
but more pronounced effect on the concentration of IL 6 and IL 15 in plasma. However, the concentration of IL 8
in response to static load increased significantly. Preliminary workouts reinforced the described reactions for all
myokines studied. The multidirectional effect of the dynamic load (swimming) on the contents of intracellular sodi
um (increase) and potassium (decrease) in the mouse soleus was shown. It was found for the first time that a similar
phenomenon is observed under static load (hanging on a grid) in mouse biceps mice and does not reliably depend on
preliminary training of experimental animals. Possible mechanisms providing the regulation of cytokine secretion
after exercise are discussed, including the [Na+]i/[K+]i dependent mechanism for triggering gene transcription.
Keywords: physical activity, myokines, ions, Na+/K+ ATPase, transcription, translation
БИОХИМИЯ том 86 вып. 3 2021