БИОХИМИЯ, 2021, том 86, вып. 6, с. 845 - 856
УДК 577.25
НЕОНАТАЛЬНЫЙ ПРОВОСПАЛИТЕЛЬНЫЙ СТРЕСС
И ЭКСПРЕССИЯ ГЕНОВ, АССОЦИИРОВАННЫХ
С НЕЙРОВОСПАЛЕНИЕМ, В ГИППОКАМПЕ КРЫС
© 2021
А.А. Квичанский*, Л.В. Третьякова, М.Н. Волобуева,
А.О. Манолова, М.Ю. Степаничев, М.В. Онуфриев,
Ю.В. Моисеева, Н.А. Лазарева, А.П. Большаков, Н.В. Гуляева
Институт высшей нервной деятельности и нейрофизиологии РАН,
117485 Москва, Россия; электронная почта: al.kvichans@gmail.com
Поступила в редакцию 24.02.2021
После доработки 26.03.2021
Принята к публикации 27.03.2021
Исследовали дифференциальное влияние неонатального провоспалительного стресса (НПС) на развитие
нейровоспаления в гиппокампе и депрессивно подобного поведения у ювенильных и взрослых самцов и са
мок крыс. Индукция НПС при помощи введения бактериального липополисахарида в неонатальном перио
де вызывает повышение экспрессии мРНК генов Il6 и Tnf на фоне формирования депрессивно подобного
поведения у взрослых самцов крыс. У ювенильных самцов крыс НПС повышает экспрессию мРНК фрак
талкина и его рецептора в вентральном гиппокампе, однако этот эффект не сопровождается повышением
экспрессии провоспалительных цитокинов и растворимой формы белка фракталкина. В дорсальном гиппо
кампе ювенильных самцов НПС снижает экспрессию мРНК фракталкина. У самок не было обнаружено
достоверного влияния НПС на исследованные показатели. Таким образом, НПС индуцирует сложные дол
говременные изменения экспрессии генов, ассоциированных с нейровоспалением в разных областях гип
покампа, приводящие в конечном счёте к индукции нейровоспаления, которое, по видимому, ассоцииро
вано с развитим депрессивно подобного поведения у самцов крыс.
КЛЮЧЕВЫЕ СЛОВА: депрессия, нейровоспаление, цитокины, гиппокамп, межполовые различия.
DOI: 10.31857/S0320972521060075
ВВЕДЕНИЕ
программирует развитие тревожно подобных и
депрессивно подобных нарушений поведения
В настоящее время большое депрессивное
во взрослом возрасте [3, 4].
расстройство является одним из наиболее рас
Механизмы программирования депрессив
пространённых в мире заболеваний [1]. Во мно
но подобного состояния у взрослых животных
гих экспериментальных исследованиях, посвя
при помощи НПС остаются не до конца понят
щённых механизмам патогенеза депрессии, экс
ными [5]. Известно, что бактериальный липо
периментальных животных помещают в условия
полисахарид (ЛПС) активирует рецептор TLR
неизбегаемого стресса различной модальности,
4, запуская неспецифический иммунный ответ.
что приводит к индукции депрессивно подоб
Этот же рецептор участвует в запуске нейровос
ных состояний [2]. Модель неонатального про
паления в мозге крыс, подвергшихся стрессу в
воспалительного стресса (НПС) является при
модели выученной беспомощности [6]. НПС
мером альтернативного подхода. В этой модели
приводит к развитию периферического воспа
животных подвергают действию провоспали
ления и нейровоспаления и вызывает ответ ор
тельных факторов в раннем постнатальном воз
ганов гипоталамо гипофизарно надпочечнико
расте (на 3 и 5 постнатальные дни (ПНД)), что
вой оси у новорождённых животных [7]. В нас
тоящее время известно, что НПС может приво
Принятые сокращения: ВГ - вентральный гиппо
дить к нарушению функционирования иммун
камп; ГЭБ - гематоэнцефалический барьер; ДГ - дорсаль
ной системы у подопытных животных [8, 9].
ный гиппокамп; ЗФ - зубчатая фасция; ЛПС - бактери
Можно предположить, что в условиях несфор
альный липополисахарид; НПС - неонатальный провос
мированного гематоэнцефалического барье
палительный стресс; ПНД - постнатальные дни; ПС - по
ра (ГЭБ) воспаление, перенесённое в раннем
веденческий стресс; ТВП - тест вынужденного плавания;
ТПС - тест предпочтения сахарозы; ФСБ - фосфатно со
постнатальном онтогенезе, индуцирует склон
левой буфер.
ность к нейровоспалению у взрослеющих и
* Адресат для корреспонденции.
взрослых особей.
845
846
КВИЧАНСКИЙ и др.
Нейровоспаление является специфической
состояниях [22, 23]. При этом гиппокамп и осо
для нервной системы реакцией, сходной с клас
бенно зубчатая фасция (ЗФ), неоднородны
сическим воспалением по набору участвующих
вдоль продольной (септо темпоральной - у лю
молекулярно клеточных механизмов. Данную
дей и дорсо вентральной - у мелких млекопита
реакцию характеризует повышение экспрессии
ющих) оси. В результате воздействия кортико
провоспалительных медиаторов (ИЛ 1β, ИЛ 6,
стероидов происходит изменение синаптичес
ФНОα) в тканях ЦНС и функциональная акти
кой пластичности нейронов гиппокампа, при
вация клеток микроглии. Такая реакция впер
этом дорсальный гиппокамп (ДГ) и вентраль
вые была описана как ответ на различные пов
ный гиппокамп (ВГ) демонстрируют различную
реждения мозга, такие как травмы, токсические
чувствительность к стрессу [24, 25]. При моде
повреждения, гипоксия и инфекции, однако
лировании депрессии у животных, помимо на
позже было показано, что нейровоспалительная
рушения синаптической пластичности нейро
реакция может запускаться и внутренними моз
нов, в ЗФ гиппокампа нарушается нейроге
говыми стимулами [10]. В частности, нейровос
нез [22, 26, 27]. Так, в работе Zhang et al. [28] бы
паление наблюдают как в моделях депрессии на
ло показано, что в условиях обогащенной среды
животных [11], так и у пациентов, страдающих
дорсальная и вентральная части гиппокампа ре
большим депрессивным расстройством [12].
агируют неодинаково - в дорсальной части ней
Предполагается, что индукция нейровоспале
рогенез активнее и транскрипционный профиль
ния приводит к формированию депрессивно
также значительно отличается от вентральной.
подобного поведения у подопытных живот
Транскриптомные и протеомные исследования
ных [13].
показали, что реакция на стресс в дорсальной и
За последние 10 лет накоплено большое чис
вентральной частях гиппокампа различают
ло данных о регуляции нейровоспаления. Уста
ся [29]. В предыдущих работах мы также показа
новлена важность баланса концентрации в тка
ли, что НПС может вызывать изменения экс
нях мозга провоспалительных (ИЛ 1β, ИЛ 6,
прессии генов, специфичные для ДГ и ВГ
ФНОα) и противовоспалительных (ИЛ 10 и
[30, 31].
другие) цитокинов, вырабатываемых клетками
Самцы и самки по разному реагируют на
как иммунной, так и нервной систем [10]. ИЛ
стресс. В частности, Tenk et al. [32] показали, что
1β и ФНОα могут экспрессироваться большин
самцы, подвергнутые НПС, сильнее теряют
ством типов клеток, представленных в мозге, в
массу после повторной инъекции ЛПС во взрос
зависимости от условий их функционального
лом возрасте, чем самки после аналогичного
статуса и микроокружения. Ключевыми проду
воздействия. Между самцами и самками крыс в
центами ИЛ 6 в ЦНС интактных здоровых мы
этой парадигме не было выявлено разницы по
шей являются активированные клетки микро
концентрации медиаторов воспаления и глюко
глии в головном мозге и клетки эпендимы спин
кортикостероидов (ГКС), но наблюдали зависи
ного мозга [14]. Наличие мРНК Il10 в тка
мую от пола реакцию микроглии - снижение
нях ЦНС является предметом для дискуссии, но
количества микроглиоцитов в гиппокампе сам
секретируемый белок представлен в мозге и яв
цов и повышение - в гиппокампе самок [7].
ляется мощным противовоспалительным меди
Можно предположить, что этот эффект лежит в
атором [15]. Особая роль принадлежит системе
основе дифференциальной предрасположен
фракталкина (CX3CL1). Этот сигнальный белок
ности самцов и самок к развитию депрессивно
синтезируется главным образом в нейронах, но
подобных нарушений во взрослом возрасте [33].
показан его синтез и в других клетках ЦНС [14].
Ранее мы показали, что введение ЛПС в ран
Рецептор фракталкина экспрессируется в мозге
нем постнатальном периоде самцам крыс при
в основном клетками микроглии [14]. Фрактал
водит к повышению тревожности у самцов юве
кин регулирует формирование зрелой микро
нильных крыс (ПНД 30-36) в тестах открытого
глии в ходе формирования мозга (в т.ч. гиппо
поля и приподнятого крестообразного лабирин
кампа и соматосенсорной коры) в эмбриональ
та. У взрослых самцов крыс (ПНД 101-107), но
ном и раннем постнатальном периоде, а также
не у ювенильных животных, наблюдали прояв
миграцию макрофагов в ЦНС у взрослых жи
ление депрессивно подобного поведения [4].
вотных [16-20]. Кроме того, в зрелом мозге
Исходя из этих данных, мы посвятили работу
фракталкин выполняет функции противовоспа
изучению влияния НПС на экспрессию генов,
лительного медиатора, обеспечивающего отри
ассоциированных с нейровоспалением (Il1b, Il6,
цательную обратную связь от нейронов к мик
Tnf, Cx3cl1, Cx3cr1), в гиппокампе взрослых жи
роглии [21].
вотных обоего пола. Также мы изучили влияние
Гиппокамп является одной из ключевых
НПС на морфологию микроглии в ДГ и ВГ юве
структур мозга, страдающих при депрессивных
нильных крыс обоего пола.
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНЫЙ СТРЕСС И ЭКСПРЕССИЯ ГЕНОВ В ГИППОКАМПЕ
847
МАТЕРИАЛЫ И МЕТОДЫ
экспрессии генов, из левого - для иммунофер
ментного анализа.
Животные. В эксперименте использовали
Крыс второй серии в возрасте 1 мес. выводи
потомство крыс линии Wistar, полученных из
ли из эксперимента путём декапитации под хло
питомника «Столбовая». Животных содержали
ралгидратным наркозом. У части крыс брали
в индивидуальных клетках со свободным досту
мозг для исследования реакции микроглии в
пом к пище и воде при 12 часовом световом
гиппокампе, а мозг остальных животных брали
цикле (свет 8:00-20:00). День родов считали
для изучения экспрессии генов (правое полуша
постнатальным днём 0 (ПНД 0). После родов ог
рие) и проведения иммуноферментного анали
раничивали численность выводка 9 особями.
за (левое полушарие).
В возрасте ПНД 21 отсаживали родившую сам
Исследование поведения. Тест предпочтения
ку, выводок разделяли на самцов и самок и со
сахарозы (ТПС). Процедура состояла из двух сеан
держали по пометам. Для экспериментов брали
сов. В ходе первого сеанса крыс приучали к пот
как самцов, так и самок.
реблению раствора сахарозы в условиях, сходных
Экспериментальные процедуры. Выводки слу
с условиями теста. Для этого животных помеща
чайным образом делили на экспериментальные
ли в индивидуальные клетки, где после 2 мин
и контрольные. На ПНД 3 и ПНД 5 животных из
привыкания им предъявляли две поилки. В од
экспериментальных выводков подвергали нео
ной из поилок была вода, в другой - 20% ный
натальному провоспалительному стрессу путём
раствор сахарозы
[34]. Приучение длилось
введения подкожно стерильного раствора ЛПС
15 мин. На следующий день животных помеща
в изотоническом растворе NaCl (10 мкл/г массы
ли в те же клетки, где после 2 мин привыкания
тела; 0,05 мг/кг ЛПС из Escherichia coli; серотип
им предъявляли предварительно взвешенные
O26:B6, «Sigma Aldrich», США). Крысам из по
поилки, а через 1 ч фиксировали изменение мас
метов контрольной группы (Контроль) вводили
сы поилок. Предпочтение сахарозы оценивали,
соответствующий объём изотонического сте
исходя из индекса, рассчитываемого по форму
рильного раствора NaCl. Выбранная доза ЛПС
ле: индекс = (масса выпитого раствора сахаро
не вызывала гибели подопытных животных. Бы
зы/масса выпитого раствора сахарозы + масса
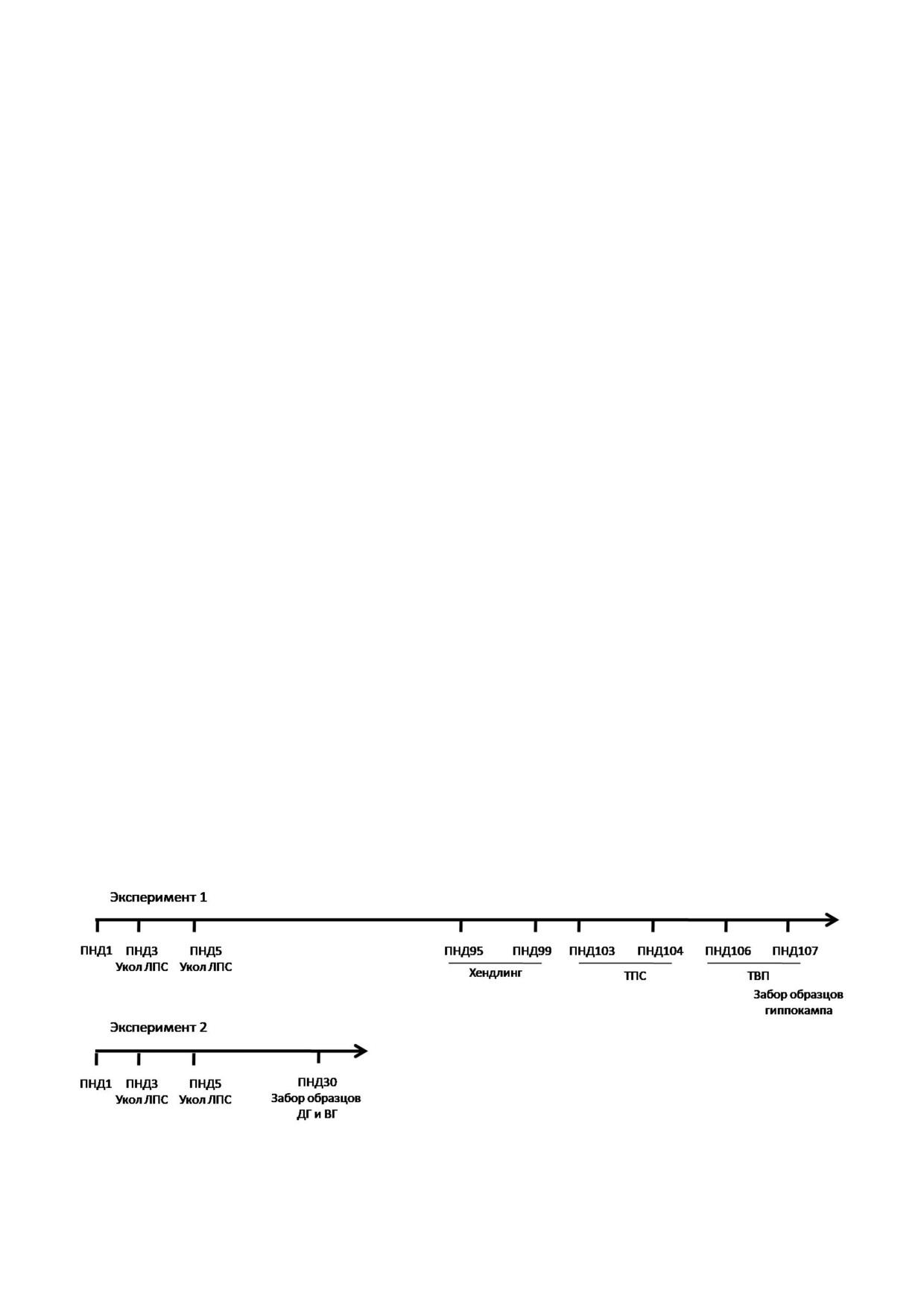
ло проведено две серии экспериментов (рис. 1).
выпитого раствора воды) × 100%. Оба сеанса
В первой серии экспериментов животных,
проводили в вечернее время (17:00-20:00).
достигших возраста 3 мес., подвергали хендлин
Тест вынужденного плавания (ТВП). Тестиро
гу в течение 4 х дней, после чего делили на две
вание поведения проводили в прозрачных ци
подгруппы. С животными одной подгруппы
линдрах высотой 40 см и диаметром 20 см («От
проводили тесты предпочтения сахарозы (ТПС)
крытая наука», Россия), наполненных во
и вынужденного плавания (ТВП). Другую под
дой (23 ± 1 °С) через 2 дня после ТПС. В первый
группу крыс оставляли в домашних клетках. Че
день ТВП крыс запускали в цилиндры на
рез 30 мин после завершения ТВП животных
15 мин, после чего вынимали, вытирали поло
выводили из эксперимента путем декапитации
тенцем, высушивали под струёй тёплого воздуха
под хлоралгидратным наркозом. Животных, не
и после этого возвращали в домашние клетки.
задействованных в поведенческих тестах, дека
На следующий день крыс помещали в цилиндры
питировали в это же время. Образцы тканей из
на 5 мин. Поведение крыс записывали с по
правого полушария использовали для изучения
мощью видеокамеры. Анализ поведения прово
Рис. 1. Схема экспериментов
БИОХИМИЯ том 86 вып. 6 2021
848
КВИЧАНСКИЙ и др.
дили по видеозаписям, оценивая время пассив
Количественная ПЦР «в реальном времени».
ного плавания (иммобильности). Под иммо
1 мкг РНК подвергали обработке ДНКазной при
бильностью понимали такое положение живот
помощи набора DNAaseI
(«Thermo Fisher
ных, при котором они не совершали плаватель
Scientific», США) в соответствии с рекомендаци
ных движений, за исключением тех, что были
ями производителя. Половину обработанной
необходимы для поддержания головы над поверх
РНК использовали для проведения реакции об
ностью воды. Стоит отметить, что ТВП, сам по
ратной транскрипции при помощи набора реак
себе, является стрессирующей парадигмой для
тивов MMLV RT kit («Евроген»), используя инги
подопытных животных. Перенесённый поведен
битор РНКаз RNase Inhibitor («New England
ческий стресс (ПС) может вызывать изменения
Biolabs», США) в соответствии с рекомендация
экспрессии генов и морфологии микроглии [35].
ми производителей. Использовали эквимоляр
Биохимический и молекулярно@биологический
ную смесь случайного декапраймера («Евроген»,
анализ. Подготовка ткани мозга для изучения
SB002) и олиго(dT)15 праймера
(«Евроген»,
экспрессии генов. Через 30 мин после последнего
SB001), концентрация каждого праймера в реак
поведенческого теста крыс декапитировали под
ционной системе составляла 1 мкМ. После об
хлоралгидратным наркозом, мозг вынимали,
ратной транскрипции полученный продукт раз
промывали в ледяном изотоническом раство
водили в 8 раз деионизованной водой. Вторую
ре NaCl, выделяли гиппокампы (цельные, или
половину РНК, обработанной ДНКазой, ис
ДГ и ВГ по отдельности) из правого полушария
пользовали в качестве отрицательного контроля
и замораживали их в жидком азоте. ДГ считали
«без обратной транскрипции». Экспрессию
дорсальную половину гиппокампа вдоль дор
целевых генов анализировали при помощи набо
сально вентральной оси, ВГ - вентральную чет
ра
«Готовая смесь для ПЦР qPCRmix HS
верть. В случае выделения ДГ и ВГ остатки тка
SYBR+LowROX» («Евроген», PK156L) при по
ни гиппокампа не анализировали. Образцы го
мощи станции для количественной ПЦР CFX384
могенизировали в реактиве для выделения
Touch («Bio Rad», США). Праймеры подбирали
РНК (ExtractRNA, «Евроген», Россия) в соответ
на основе последовательностей из базы данных
ствии с рекомендациями производителя, и вы
NCBI в программном пакете Lasergene
деляли фракцию тотальной РНК. РНК в ходе
PrimerSelect. Последовательности праймеров,
работы хранили в виде раствора в деионизован
использованных в работе, приведены в таблице.
ной воде и долговременно - в виде осадка
В качестве нормировочного гена использо
в 80% этаноле.
вали праймеры к кДНК гена Hprt, выбранного
Последовательности праймеров, использованных в работе
Последовательности праймеров (5′-3′)
Ген
Прямой
Обратный
Il1b
TCTGTGACTCGTGGGATGAT
CACTTGTTGGCTTATGTTCTGTC
NM_031512.2
Il6
GCCACTGCCTTCCCTACTTCAC
GACAGTGCATCATCGCTGTTCATAC
NM_012589.2
Tnf
GTCCAACTCCGGGCTCAGAAT
ACTCCCCCGATCCACTCAG
NM_012675.3
Il10
GACAATAACTGCACCCACTTCC
GCATCACTTCTACCAGGTAAAACTTG
NM_012854.2
Cx3cl1
ATCACCACCATCACCACCAAC
GAGGAACACTTTAAACCCTCACAGA
NM_134455.2
Cx3cr1
GGACCTCACCATGCCTACCT
CACCAACAGATTCCCCACCAG
NM_133534.2
Hprt
CGTCGTGATTAGTGATGATGAAC
CAAGTCTTTCAGTCCTGTCCATA
NM_012583.2
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНЫЙ СТРЕСС И ЭКСПРЕССИЯ ГЕНОВ В ГИППОКАМПЕ
849
по результатам анализа транскриптома гиппо
кубировали с первичными поликлональными
кампа крысы [36]. Относительную представлен
антителами Rabbit anti Iba1 IgG («Wako», Япо
ность транскриптов оценивали по методу EΔΔCt.
ния) в разведении 1/1000 при температуре 4 °С.
Эффективность реакции измеряли для каждого
На следующий день после трех отмывок в ФСБТ
рабочего разведения праймеров методом серий
срезы инкубировали (2 ч при комнатной темпе
ных разведений. Во всех экспериментах эффек
ратуре) со вторичными антителами Goat anti
тивность реакции находилась в диапазоне
Rabbit IgG (H+L), конъюгированными с
1,8-2,0. Каждую пробу дублировали, кроме того,
Alexa488 («Thermo Fisher Scientific»). После трех
для каждой пробы и каждого гена ставили конт
отмывок в ФСБ срезы заключали под покров
роль «без обратной транскрипции». Для каждой
ные стекла в среду для заливки препаратов
лунки контролировали температуру плавления
ProLong Gold («Thermo Fisher Scientific»), пре
продукта ПЦР. Данные на графиках представле
пятствующую выгоранию, с подкрашиванием
ны в виде относительного количества:
клеточных ядер DAPI («Invitrogen», США) и
хранили при -20 °C до использования.
Для проведения морфологического анализа
микроглиальных клеток получали z стеки с по
мощью микроскопа Zeiss Axio Imager Z2, осна
щённого объективом EC Plan NEOFLUAR 40×
Иммуноферментный анализ. Отделы мозга
(NA = 0,75) и цифровой камерой MRm («Carl
гомогенизировали на льду в буфере для экстрак
Zeiss», Германия). Количество изображений в
ции растворимых белков
(1% ный NP 40
z стеке было фиксированным (17), расстояние
(«Sigma Aldrich», США) в фосфатно солевом
между ними было установлено оптималь
буфере (ФСБ, «ПанЭко», Россия), pH 7,5) при
ным (0,61 мкм). Фотографировали поля CA1
помощи гомогенизатора Поттера 10 ударами
и CA3 гиппокампа и хилус зубчатой фасции, ис
пестика при 1000 об./мин. Гомогенат центрифу
пользуя следующие координаты: для дорсально
гировали 20 мин при 4 °С и 13 200 g. Суперна
го гиппокампа AP 2,80-3,80 мм и для вентраль
тант отбирали и хранили при -20 °С. Концент
ного - AP 4,80-5,30 мм, согласно атласу моз
рацию белка в пробах измеряли при помощи на
га [37]. Для каждого животного было получено
бора Pierce™ BCA Protein Assay Kit («Thermo
по два изображения из каждой области.
Fisher Scientific»). Концентрацию растворимой
Для последующей обработки изображений,
фракции фракталкина, ИЛ 6 и ИЛ 1β в пробах
подсчёта числа клеток и оценки степени актива
определяли при помощи наборов Rat
ции микроглии использовали программное
CX3CL1/Fractalkine DuoSet ELISA, Rat IL 6
обеспечение Fiji (NIH, США). Для подсчёта
Quantikine ELISA Kit и Rat IL 1 beta/IL 1F2
клеток микроглии из z стека получали проек
DuoSet ELISA («R&D Systems», США) в соответ
цию максимальной интенсивности. Подсчиты
ствии с рекомендациями производителя. Каж
вали количество Iba1 позитивных клеток, со
дую пробу дублировали, сигнал детектировали
держащих чётко очерченную сому клетки и не
при помощи планшетного ридера Hidex Sense
менее двух отростков. Клетки, сома которых пе
(«Hidex», Финляндия).
ресекала левую или нижнюю границы изобра
Изучение реакции микроглии в гиппокампе.
жения, не учитывали. Для оценки активации
Мозг крысы фиксировали погружением в
микроглии рассчитывали фрактальную размер
50 мл 4% ного раствора параформальдегида в
ность контура отдельных клеток с использова
ФСБ, pH 7,5 на 1 ч. Затем заменяли фиксатор на
нием плагина FracLac для Fiji [33]. Чтобы полу
свежий и инкубировали в нем следующие 24 ч.
чить контур микроглиальной клетки использо
Для иммуногистохимического окрашивания
вали последовательность преобразования изо
готовили фронтальные срезы толщиной 50 мкм
бражений, предложенную Young и Morrison [38],
на вибрационном микротоме Leica VT1200 S
с некоторыми модификациями [39]. Вкратце,
(«Leica Biosystems», Германия). Срезы помещали
эта последовательность включала в себя следую
в криопротекторную среду (ФСБ/глицерин/
щие шаги: 1 - получение проекции максималь
этиленгликоль = 1/0,75/0,75 (v/v)) и хранили
ной интенсивности; 2 - преобразования, усили
при -20 °C до обработки. Перед окрашиванием
вающие контрастность изображения и оптими
срезы трижды промывали в 0,3% ном (w/v)
зирующие соотношение сигнал/шум; 3 - выре
Triton X 100 («Serva», Германия) в ФСБ (ФСБТ),
зание из изображения отдельных клеток мик
и блокировали неспецифичное связывание ан
роглии (с каждого изображения брали по три
тител, для чего инкубировали в 5% ной нор
клетки с чётко видимой сомой и отростками);
мальной козьей сыворотке («Sigma Aldrich»,
4 - бинаризация изображений отдельных кле
США) в ФСБТ. Затем срезы в течение ночи ин
ток; 5 - получение контура отдельных клеток из
7 БИОХИМИЯ том 86 вып. 6 2021
850
КВИЧАНСКИЙ и др.
бинаризованного изображения. Всего было
проанализировано по шесть клеток у каждого
животного в каждом из исследованных отделов
гиппокампа. Фрактальная размерность контура
объекта тем выше, чем сильнее его извилис
тость. Так, например, для такого простого объ
екта, как окружность, фрактальная размерность
равна 1. Для амебоидной микроглии фракталь
ная размерность, как правило, составляет ~1,2, а
для разветвлённой рамифицированной микро
глии - 1,5. Таким образом, оценку фрактальной
размерности можно использовать для оценки
степени активированности микроглии.
Статистическая обработка данных. Данные о
поведении крыс и о морфологии клеток гиппо
кампа на графиках представлены в виде наложе
ния столбчатых диаграмм (среднее ± SE) и то
чечных диаграмм, во всех группах n ≥ 5. Данные
об экспрессии генов и концентрации их белко
Рис. 2. Индукция депрессивно подобного поведения НПС
вых продуктов на графиках представлены в виде
у взрослых самцов и самок крыс. * p < 0,05, тест Тьюки
наложения диаграмм размаха и точечных диаг
рамм, во всех группах n ≥ 5. Распределение пере
менных в выборке оценивали на соответствие
ровали развитие так называемого «поведения
нормальному с помощью критерия Шапиро-
отчаяния», которое зачастую интерпретируют
Уилка. Достоверность различий в исследовании
как депрессивно подобное, тогда как самки де
поведения и морфологии клеток гиппокампа
монстрировали относительную устойчивость к
определяли двухфакторным дисперсионным
развитию депрессивно подобного поведения.
анализом с последующим апостериорным ана
Не было выявлено достоверного влияния
лизом по методу Фишера или Тьюки в зависи
пола, НПС и взаимодействия факторов на пот
мости от применимости методов в программном
ребление сахарозы взрослыми крысами (данные
пакете Statistica 12. В качестве факторов для не
не представлены).
зависимых переменных использовали «пол» и
Экспрессия генов, ассоциированных с нейро@
«НПС». В исследованиях экспрессии мРНК и
воспалением, в гиппокампе взрослых крыс. Влия
концентрации белков распределение перемен
ние перенесённого НПС на экспрессию
ных не соответствовало нормальному по крите
мРНК Il1b, Il6, Tnf, Cx3cl1 и Cx3cr1, а также на
рию Шапиро-Уилка, поэтому достоверность
концентрацию белка ИЛ 6 было исследовано в
различий между ДГ и ВГ определяли по методу
гиппокампе взрослых самцов и самок крыс, ко
Уилкоксона, достоверность различий между
торые либо оставались в домашних клетках, ли
экспериментальными группами выявляли по
бо принимали участие в ТВП. Мы обнаружили,
методу Манна-Уитни с поправкой Бонферро
что в гиппокампе взрослых самцов крыс, пере
ни (3 гипотезы, α = 0,017) в программном паке
нёсших НПС, экспрессия мРНК Il6 была выше,
те Statistica 12.
чем у контрольных животных независимо от то
го, были они подвергнуты испытанию в ТВП
или нет (p = 0,008 и p = 0,003 соответственно;
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
тест Манна-Уитни, рис. 3, а). Однако этот эф
фект не сопровождался изменением концентра
Исследование поведения взрослых крыс. Пе
ции белка ИЛ 6 в гиппокампе взрослых сам
ренесённый НПС приводил к появлению приз
цов (рис. 3, в). Не было выявлено влияния НПС,
наков депрессивно подобного поведения у
ПС и их сочетания на экспрессию мРНК этого
взрослых крыс в тесте вынужденного плавания.
цитокина у самок (рис. 3, б), однако, парадок
Было выявлено взаимодействие факторов «пол»
сально, ПС приводил к снижению концентра
и «НПС» F (1, 35) = 4,81, p = 0,034. Апостериор
ции ИЛ 6 у самок, подвергнутых НПС
ное сравнение средних выявило повышение вре
(p = 0,014, рис. 3, г). Кроме того, НПС не приво
мени пассивного плавания у самцов, подверг
дил к достоверному повышению экспрессии
нутых НПС (тест Тьюки, p = 0,036), но не выя
мРНК Tnf, хотя была выявлена тенденция к
вил значимых изменений поведения у са
снижению этого параметра под действием ПС у
мок (рис. 2). Таким образом, самцы демонстри
самцов, подвергнутых НПС (p = 0,027, тест
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНЫЙ СТРЕСС И ЭКСПРЕССИЯ ГЕНОВ В ГИППОКАМПЕ
851
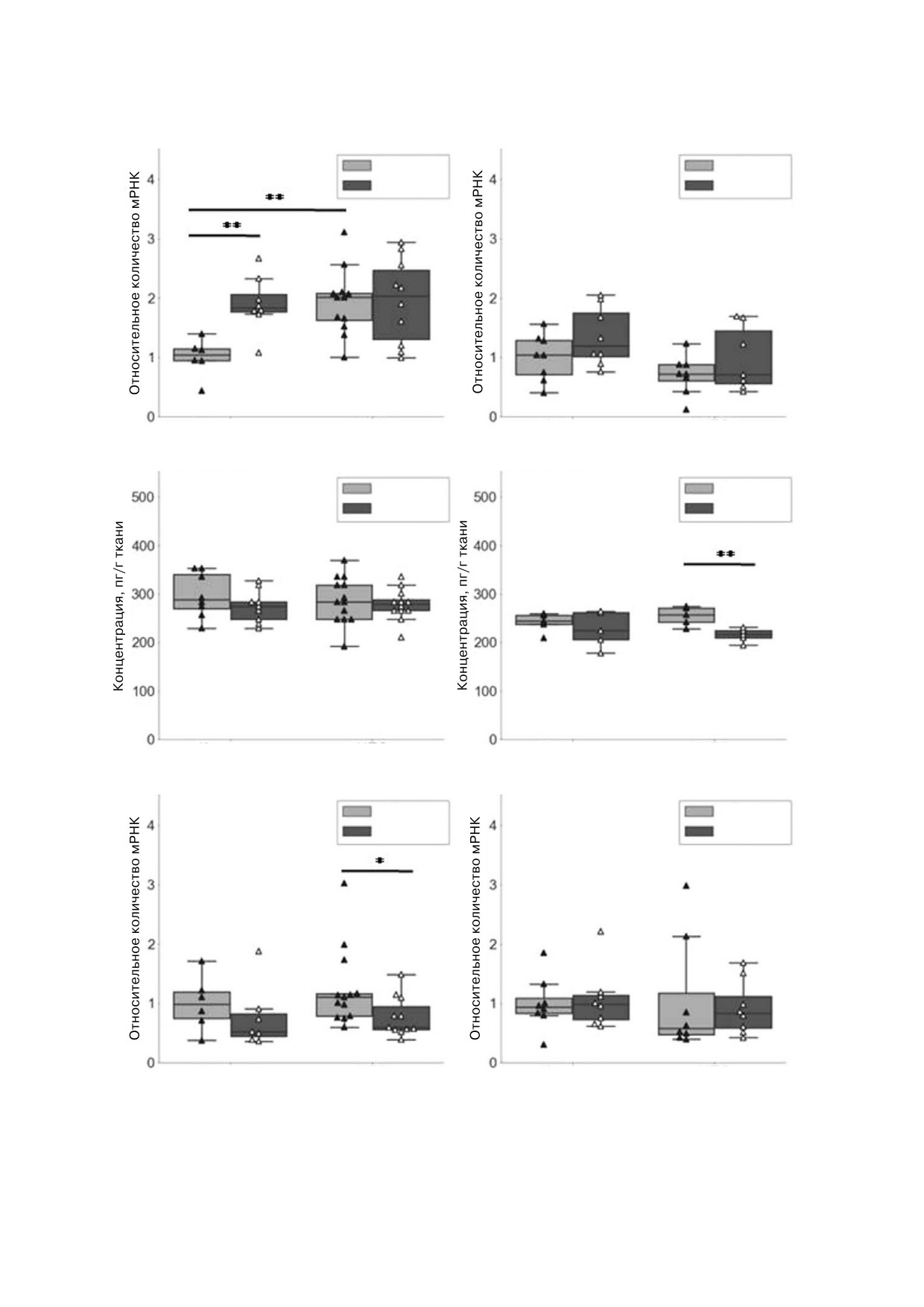
а
Il6, самцы
б
Il6, самки
Контроль
Контроль
ПС
ПС
Контроль
НПС
Контроль
НПС
в
ИЛ 6, самцы
г
ИЛ 6, самки
Контроль
Контроль
ПС
ПС
Контроль
НПС
Контроль
НПС
д
Tnf, самцы
е
Tnf, самки
Контроль
Контроль
ПС
ПС
Контроль
НПС
Контроль
НПС
Рис. 3. Экспрессия генов, ассоциированных с нейровоспалением в гиппокампе взрослых крыс. а - Экспрессия мРНК Il6
в гиппокампе самцов, б - экспрессия мРНК Il6 в гиппокампе самок, в - концентрация ИЛ 6 в гиппокампе самцов,
г - концентрация ИЛ 6 в гиппокампе самок, д - экспрессия мРНК Tnf в гиппокампе самцов, е - экспрессия мРНК Tnf в
гиппокампе самок. * p < 0,05, ** p < 0,017, тест Манна-Уитни
БИОХИМИЯ том 86 вып. 6 2021
7*
852
КВИЧАНСКИЙ и др.
Манна-Уитни, рис. 3, д). Не было выявлено
ванных с нейровоспалением в ДГ и ВГ самцов и
влияния НПС и ПС на экспрессию мРНК Tnf в
самок ювенильных крыс.
гиппокампе самок (рис. 3, е). Таким образом,
Мы обнаружили, что во всех эксперимен
мы обнаружили, что НПС и ПС индуцируют
тальных группах наблюдается больший уровень
экспрессию мРНК Il6 в гиппокампе самцов
экспрессии мРНК Il1b в ВГ по сравнению с
крыс на фоне развития депрессивно подобного
ДГ (в 4,7 раза у самцов и в 3,4 раза у самок) и
поведения.
меньший уровень экспрессии мРНК Tnf (в
Не было выявлено влияния НПС и ПС на
1,9 раза у крыс обоего пола) в этих же структурах
экспрессию мРНК Il1b, Cx3cl1 и его рецептора
(тест Вилкоксона, p < 0,05). Было выявлено сни
Cx3cr1 в гиппокампе взрослых крыс (данные не
жение экспрессии мРНК Cx3cl1 в ДГ ювениль
представлены).
ных самцов под действием НПС (p = 0,005, тест
Экспрессия генов, ассоциированных с нейро@
Манна-Уитни). Кроме того, было выявлено по
воспалением, в ДГ и ВГ ювенильных крыс. Для
вышение экспрессии мРНК этого хемокина
изучения возможных механизмов, участвующих
(p = 0,005, тест Манна-Уитни) и тенденция к
в формировании предрасположенности к деп
повышению экспрессии мРНК его рецептора
рессивно подобному поведению, мы изучили
Cx3Cr1 (p = 0,03, тест Манна-Уитни) в ВГ юве
влияние НПС на экспрессию генов, ассоцииро
нильных самцов под действием НПС. Однако
а
Cx3cl1, самцы
б
Cx3cl1, самки
Дорсальный гиппокамп
Дорсальный гиппокамп
Вентральный гиппокамп
Вентральный гиппокамп
Контроль
НПС
Контроль
НПС
в
Cx3cr1, самцы
г
Cx3cr1, самки
Dorsal hippocampus
Dorsal hippocampus
Ventral hippocampus
Ventral hippocampus
Контроль
НПС
Контроль
НПС
Рис. 4. Влияние НПС на экспрессию генов, ассоциированных с нейровоспалением в ДГ и ВГ ювенильных крыс.
а - Экспрессия мРНК Cx3cl1 в ДГ и ВГ самцов, б - экспрессия мРНК Cx3cl1 в ДГ и ВГ самок, в - экспрессия мРНК Cx3cr1
в ДГ и ВГ самцов, г - экспрессия мРНК Cx3cr1 в ДГ и ВГ самок. * p < 0,05, ** p < 0,017, тест Манна-Уитни
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНЫЙ СТРЕСС И ЭКСПРЕССИЯ ГЕНОВ В ГИППОКАМПЕ
853
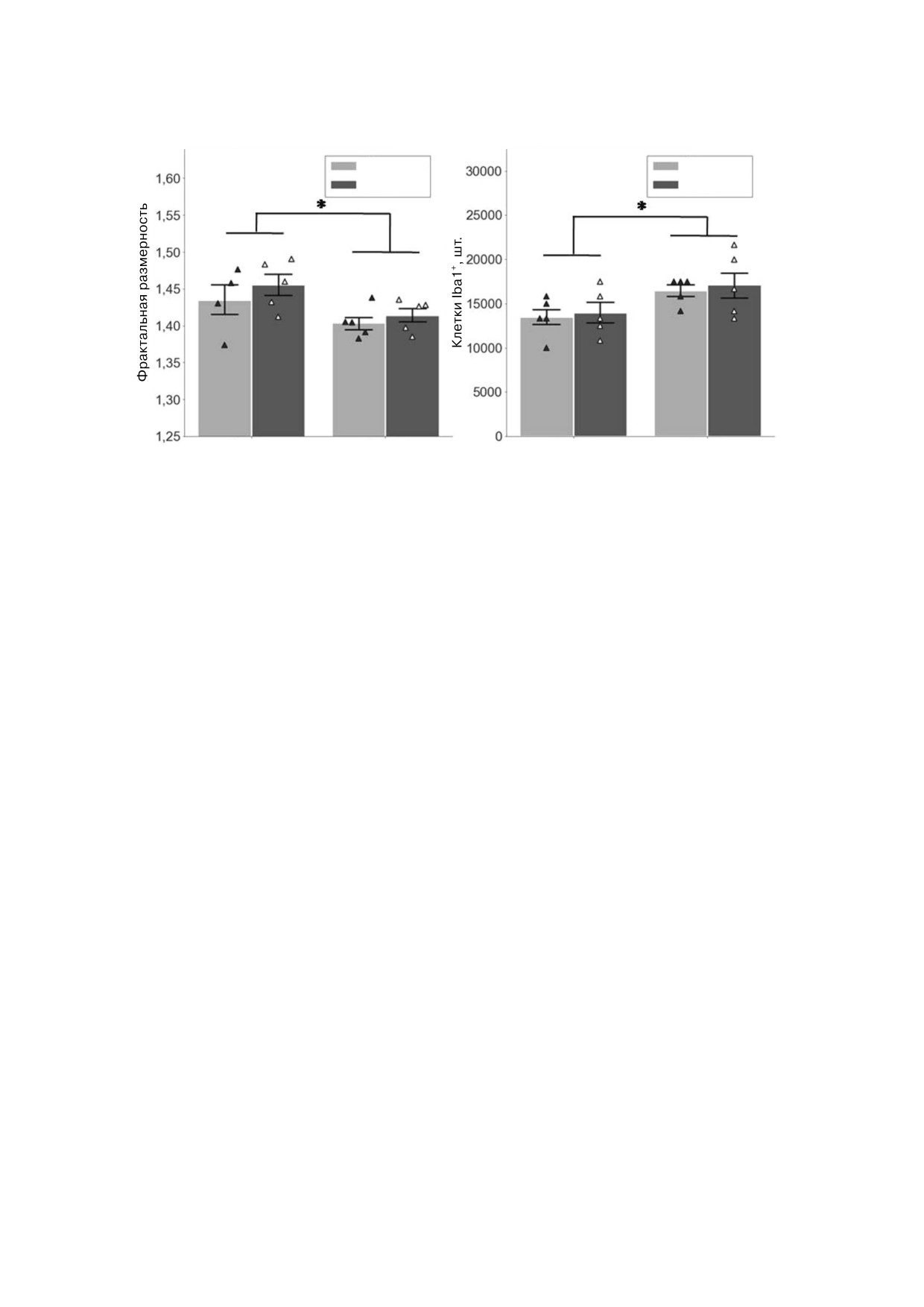
а
Вентральный гиппокамп, CA1
б
Дорсальный гиппокамп, ЗФ
Контроль
Контроль
НПС
НПС
Самцы
Самки
Самцы
Самки
Рис. 5. Влияние НПС на морфологию клеток микроглии в ДГ и ВГ ювенильных крыс. а - Фрактальная размерность Iba1
позитивных клеток в области CA1 ВГ, б - количество клеток Iba1+ в ЗФ ДГ. * p < 0,05, тест Фишера
эти эффекты не сопровождались изменениями
влияния НПС на потребление сахарозы экспе
экспрессии провоспалительных цитокинов и
риментальными животными. Можно предполо
растворимой формы белка фракталкина. У са
жить, что нарушения, развившиеся у подопыт
мок не было обнаружено достоверных измене
ных животных, затрагивают системы мозга, от
ний, вызванных НПС (рис. 4). Уровень экспрес
ветственные за реакцию на стресс, но не ответ
сии мРНК Il10 был ниже порога детекции (дан
ственные за гедонистическое поведение.
ные не представлены). Таким образом, не было
Экспрессия мРНК Il6 была повышена в гип
обнаружено признаков нейровоспалительной
покампе взрослых самцов, подвергнутых НПС,
реакции в ДГ и ВГ ювенильных крыс, подверг
у самок аналогичного эффекта выявлено не бы
нутых НПС. Однако были выявлены специфич
ло. Это может указывать на развитие хроничес
ные для отделов гиппокампа изменения экс
кого нейровоспаления у самцов, приводящего к
прессии мРНК фракталкина и его рецептора.
депрессивно подобному состоянию. ПС вызы
Исследование морфологии микроглии в ДГ и
вал повышение экспрессии мРНК Il6 в гиппо
ВГ ювенильных крыс. Выявлена достоверно
кампе самцов, не подвергнутых НПС, до уров
большая фрактальная размерность клеток мик
ня, наблюдаемого у животных, подвергнутых
роглии в СА1 ВГ самцов по сравнению с самка
НПС, но не подвергнутых ПС. Эти данные хо
ми (F (1, 15) = 6,36, p = 0,016, критерий Фише
рошо согласуются с данными о повышении
ра, рис. 5, а). Кроме того, в ЗФ ДГ самцов наб
экспрессии мРНК данного цитокина в микро
людали большее число Iba1 позитивных клеток
глии под действием хронического стресса [40].
(F (1, 16) = 6,95, p = 0,015, критерий Фишера,
В то же время не было выявлено влияния НПС
рис. 5, б). Влияния НПС, взаимодействия пола и
и ПС на концентрацию белка ИЛ 6 в гиппокам
НПС и достоверных различий в CA3 не выявле
пе взрослых крыс. Можно предположить, что
но (данные не представлены). Таким образом,
синтез этого цитокина происходит локально с
нельзя заключить, что НПС влияет на морфоло
быстрой секрецией и утилизацией комплекса
гию клеток микроглии в гиппокампе ювениль
ИЛ 6 и его рецепторов клетками мишенями.
ных крыс.
Кроме того, для данного цитокина известно ак
тивное вторичное использование клетками им
мунной системы, что не требует повышения
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
средней концентрации ИЛ 6 в ткани [41]. Важ
ным свойством ИЛ 6 может оказаться его спо
Мы продемонстрировали формирование
собность сдвигать соотношение субпопуляций
депрессивно подобного поведения в ТВП у
Treg/Th17 лимфоцитов в сторону Th17, которые
взрослых самцов, но не у самок крыс, подверг
осуществляют провоспалительную функ
нутых НПС. Вместе с тем не было выявлено
цию [42]. Показана связь накопления в мозге
БИОХИМИЯ том 86 вып. 6 2021
854
КВИЧАНСКИЙ и др.
Th17 лимфоцитов с развитием депрессии, выз
цесса снижает интенсивность нейровоспале
ванной хроническим стрессом [43].
ния [16-20]. В прямом эксперименте введение в
Не было выявлено влияния собственно НПС
гиппокамп растворимого фракталкина вызывает
на экспрессию мРНК Tnf у взрослых животных
активацию микроглии [45]. Таким образом, наб
обоего пола, а дополнительный стресс, связан
людается двойственный характер регуляции
ный с ТВП, парадоксально приводил к сниже
нейровоспалительной реакции фракталкином.
нию экспрессии мРНК Tnf в гиппокампе взрос
Можно предположить, что растворимая форма
лых самцов, подвергнутых НПС. Можно пред
фракталкина может выступать в качестве ло
положить, что острый стресс, ассоциированный
кального провоспалительного агента, привлека
с вынужденным плаванием, вызывал активацию
ющего клетки иммунной системы в очаг воспа
неизвестных противовоспалительных механиз
ления через ГЭБ, в то время как трансмембран
мов, подавлявших экспрессию мРНК Tnf у этих
ная форма может выступать в качестве негатив
животных.
ного иммунного регулятора. Можно предполо
Мы обнаружили повышение экспрессии
жить, что повышение экспрессии мРНК Cx3cr1
мРНК фракталкина и его рецептора у ювениль
является признаком изменения свойств микро
ных самцов в ВГ через месяц после НПС, одна
глии, а повышение экспрессии Cx3cl1 - попыт
ко фрактальная размерность и количество кле
кой ткани мозга скомпенсировать нарастающие
ток микроглии у этих животных не отличались
нарушения. Сходные эффекты детектируют в ДГ
от контрольных. Можно предположить, что у
крыс, подвергнутых стрессу иммобилизации,
этих животных усиленно работает отрицатель
также индуцирующему депрессивно подобное
ная обратная связь от нейронов к микроглии,
поведение [46]. Небольшое по амплитуде, но
направленная на подавление провоспалитель
достоверное снижение экспрессии мРНК фрак
ной активности [21]. Отсутствие изменения кон
талкина в ДГ может указывать на неизвестные
центрации растворимого фракталкина не явля
механизмы регуляции нейровоспаления, отли
ется строгим доказательством отсутствия повы
чающиеся в ДГ и ВГ. Можно предположить, что
шения синтеза белкового продукта гена Cx3cl1.
в дальнейшем изменения свойств микроглии
Этот белок экспонируется на поверхности кле
приводят к программированию склонности к
ток в виде функционально активного трансмем
развитию нейровоспаления у взрослых крыс.
бранного белка, который может подвергаться
протеолитическому расщеплению с образовани
ем растворимой формы, вероятно, без измене
Финансирование. Исследование поведения
ния способности активировать его рецеп
животных поддержано средствами государ
тор [44]. Трансмембранная форма фракталкина
ственного бюджета в соответствии с государ
недоступна для иммуноферментного анализа и
ственным заданием Министерства образования
не была исследована в нашей работе. Предпола
и науки Российской Федерации. Исследование
гают, что в зрелом мозге фракталкин является
морфологии микроглии поддержано Российс
противовоспалительным медиатором, обеспе
ким научным фондом (грант № 19 75 00063).
чивающим отрицательную обратную связь от
Конфликт интересов. Авторы заявляют об от
нейронов к микроглии. В то же время известно,
сутствии конфликта интересов.
что рецептор фракталкина регулирует миграцию
Соблюдение этических норм. Все принятые
моноцитов через ГЭБ и их активацию в ходе раз
международные, национальные и/или институ
вития нейровоспалительной реакции в некото
циональные принципы ухода и использования
рых моделях патологий, и подавление этого про
животных были соблюдены.
СПИСОК ЛИТЕРАТУРЫ
1.
Kupfer, D. J., Frank, E., and Phillips, M. L. (2012) Major
4.
Tishkina, A., Stepanichev, M., Kudryashova, I.,
depressive disorder: new clinical, neurobiological, and
Freiman, S., Onufriev, M., et al. (2016) Neonatal proin
treatment perspectives, Lancet,
379,
10451055,
flammatory challenge in male Wistar rats: effects on behav
doi: 10.1016/S0140 6736(11)60602 8.
ior, synaptic plasticity, and adrenocortical stres sresponse,
2.
Stepanichev, M., Dygalo, N. N., Grigoryan, G.,
Behav. Brain Res., 304, 1 10, doi: 10.1016/j.bbr.2016.
Shishkina, G. T., and Gulyaeva, N. (2014) Rodent models
02.001.
of depression: Neurotrophic and neuroinflammatory bio
5.
Spencer, S. J., and Meyer, U. (2017) Perinatal program
markers, Biomed Res. Int., 2014, 932757, doi: 10.1155/
ming by inflammation, Brain Behav. Immun., 63, 1 7,
2014/932757.
doi: 10.1016/j.bbi.2017.02.007.
3.
Bilbo, S., and Schwarz, J. (2009) Early life programming
6.
Cheng, Y., Pardo, M., Armini, R., Martinez, A.,
of later life brain and behavior: a critical role for the
Mouhsine, H., et al. (2016) Stress induced neuroinflam
immune system, Front. Behav. Neurosci.,
3,
14,
mation is mediated by GSK3 dependent TLR4 signaling
doi: 10.3389/neuro.08.014.2009.
that promotes susceptibility to depression like behavior,
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНЫЙ СТРЕСС И ЭКСПРЕССИЯ ГЕНОВ В ГИППОКАМПЕ
855
Brain. Behav. Immun., 53, 207 222, doi: 10.1016/j.bbi.
logical inhibition of the chemokine receptor CX3CR1
2015.12.012.
attenuates disease in a chronic relapsing rat model for
7.
Claypoole, L. D., Zimmerberg, B., and Williamson, L. L.
multiple sclerosis, Proc. Natl. Acad. Sci. USA, 111, 5409
(2017) Neonatal lipopolysaccharide treatment alters hip
5414, doi: 10.1073/pnas.1316510111.
pocampal neuroinflammation, microglia morphology and
21.
Rogers, J. T., Morganti, J. M., Bachstetter, A. D., Hudson,
anxiety like behavior in rats selectively bred for an infantile
C. E., Peters, M. M., et al. (2011) CX3CR1 deficiency
trait, Brain Behav. Immun., 59, 135 146, doi: 10.1016/j.bbi.
leads to impairment of hippocampal cognitive function and
2016.08.017.
synaptic plasticity, J. Neurosci.,
31,
1624116250,
8.
Barth, C. R., Luft, C., Funchal, G. A., Oliveira, J. R., de
doi: 10.1523/JNEUROSCI.3667 11.2011.
Porto, B. N., and Donadio, M. V. F. (2016) LPS induced
22.
Boldrini, M., Santiago, A. N., Hen, R., Dwork, A. J.,
neonatal stress in mice affects the response profile to an
Rosoklija, G. B., et al. (2013) Hippocampal granule neuron
inflammatory stimulus in an age and sex dependent man
number and dentate gyrus volume in antidepressant treated
ner, Dev. Psychobiol., 58, 600 613, doi: 10.1002/dev.21404.
and untreated major depression, Neuropsychopharmacology,
9.
Walker, F. R., Hodyl, N. A., and Hodgson, D. M. (2009)
38, 1068 1077, doi: 10.1038/npp.2013.5.
Neonatal bacterial endotoxin challenge interacts with
23.
Gulyaeva, N. V. (2015) Ventral hippocampus, stress and
stress in the adult male rat to modify KLH specific anti
phychopathology: translational implications, Neurochem. J.,
body production but not KLH stimulated ex vivo cytokine
9, 85 94, doi: 10.1134/S1819712415020075.
release, J. Neuroimmunol., 207, 5765, doi: 10.1016/
24.
Maggio, N., and Segal, M. (2012) Steroid modulation of
j.jneuroim.2008.11.012.
hippocampal plasticity: switching between cognitive and
10.
Estes, M. L., and McAllister, A. K. (2014) Alterations in
emotional memories, Front. Cell Neurosci.,
6,
12,
immune cells and mediators in the brain: it’s not always
doi: 10.3389/fncel.2012.00012.
neuroinflammation! Brain Pathol.,
24,
623630,
25.
Gulyaeva, N. V. (2019) Functional neurochemistry of the
doi: 10.1111/bpa.12198.
ventral and dorsal hippocampus: stress, depression,
11.
Kubera, M., Obuchowicz, E., Goehler, L., Brzeszcz, J.,
dementia and remote hippocampal damage, Neurochem.
and Maes, M. (2011) In animal models, psychosocial
Res., 44, 1306 1322, doi: 10.1007/s11064 018 2662 0.
stress induced (neuro)inflammation, apoptosis and
26.
Mahar, I., Bambico, F. R., Mechawar, N., and Nobrega,
reduced neurogenesis are associated to the onset of depres
J. N. (2014) Stress, serotonin, and hippocampal neurogen
sion, Prog. Neuropsychopharmacol. Biol. Psychiatry, 35,
esis in relation to depression and antidepressant effects,
744 759, doi: 10.1016/j.pnpbp.2010.08.026.
Neurosci. Biobehav. Rev., 38, 173192, doi: 10.1016/
12.
Setiawan, E., Wilson, A. A., Mizrahi, R., Rusjan, P. M.,
j.neubiorev.2013.11.009.
Miler, L., et al. (2015) Role of translocator protein density,
27.
Podgorny, O. V., and Gulyaeva, N. V.
(2020)
a marker of neuroinflammation, in the brain during major
Glucocorticoid mediated mechanisms of hippocampal
depressive episodes, JAMA Psychiatry, 72, 268, doi: 10.1001/
damage: contribution of subgranular neurogenesis,
jamapsychiatry.2014.2427.
J. Neurochem., doi: 10.1111/jnc.15265.
13.
Wang, Y., Cui, X. L., Liu, Y. F., Gao, F., Wei, D., et al.
28.
Zhang, T. Y., Keown, C. L., Wen, X., Li, J., Vousden,
(2011) LPS inhibits the effects of fluoxetine on depression
D. A., Anacker, C., et al. (2018) Environmental enrich
like behavior and hippocampal neurogenesis in rats, Prog.
ment increases transcriptional and epigenetic differentia
Neuropsychopharmacol. Biol. Psychiatry, 35, 1831 1835,
tion between mouse dorsal and ventral dentate gyrus, Nat.
doi: 10.1016/j.pnpbp.2011.07.004.
Commun., 9, 1 11, doi: 10.1038/s41467 017 02748 x.
14.
Zeisel, A., Hochgerner, H., Lönnerberg, P., Johnsson, A.,
29.
Floriou Servou, A., Ziegler, L., von Stalder, L.,
Memic, F., et al. (2018) Molecular architecture of the
Sturman, O., Privitera, M., et al. (2018) Distinct proteom
mouse nervous system, Cell,
174,
9991014.e22,
ic, transcriptomic, and epigenetic stress responses in dorsal
doi: 10.1016/j.cell.2018.06.021.
and ventral hippocampus, Biol. Psychiatry, 84, 531 541,
15.
Pisanu, A., Lecca, D., Mulas, G., Wardas, J., Simbula, G.,
doi: 10.1016/j.biopsych.2018.02.003.
et al. (2014) Dynamic changes in pro and anti inflamma
30.
Kvichansky, A. A., Volobueva, M. N., Manolova, A. O.,
tory cytokines in microglia after PPAR γ agonist neuropro
Bolshakov, A. P., and Gulyaeva, N. V. (2017) Neonatal
tective treatment in the MPTPp mouse model of progres
proinflammatory stress alters the expression of genes of
sive Parkinson’s disease, Neurobiol. Dis., 71, 280 291,
corticosteroid receptors in the rat hippocampus: septo
doi: 10.1016/j.nbd.2014.08.011.
temporal differences, Neurochem. J.,
11,
255258,
16.
Tremblay, M. È., and Sierra, A. (2014) Microglia in Health
doi: 10.1134/S1819712417030059.
and Disease. Chapter 1. Introduction, doi: 10.1007/978 1
31.
Kvichansky, A. A., Volobueva, M. N., Manolova, A. O.,
4939 1429 6.
Bolshakov, A. P., and Gulyaeva, N. V. (2018) The influence
17.
Tang, Z., Gan, Y., Liu, Q., Yin, J. X., Liu, Q., Shi, J., and
of neonatal pro inflammatory stress on the expression of
Shi, F. D. (2014) CX3CR1 deficiency suppresses activation
genes associated with stress in the brains of juvenile rats:
and neurotoxicity of microglia/macrophage in experimen
septo temporal specificity, Neurochem. J., 12, 180 183,
tal ischemic stroke, J. Neuroinflammation,
11,
26,
doi: 10.1134/s1819712418020083.
doi: 10.1186/1742 2094 11 26.
32.
Tenk, C. M., Kavaliers, M., and Ossenkopp, K. P. (2008)
18.
Lee, S., Varvel, N. H., Konerth, M. E., Xu, G., Cardona,
Sexually dimorphic effects of neonatal immune system acti
A. E., et al. (2010) CX3CR1 deficiency alters microglial
vation with lipopolysaccharide on the behavioural response
activation and reduces beta amyloid deposition in two
to a homotypic adult immune challenge, Int. J. Dev.
Alzheimer’s disease mouse models, Am. J. Pathol., 177,
Neurosci., 26, 331 338, doi: 10.1016/j.ijdevneu.2008.01.001.
2549 2562, doi: 10.2353/ajpath.2010.100265.
33.
Walker, A. K., Nakamura, T., Byrne, R. J., Naicker, S.,
19.
Hellwig, S., Brioschi, S., Dieni, S., Frings, L., Masuch, A.,
Tynan, R. J., et al. (2009) Neonatal lipopolysaccharide and
et al. (2016) Altered microglia morphology and higher
adult stress exposure predisposes rats to anxiety like behav
resilience to stress induced depression like behavior in
iour and blunted corticosterone responses: implications for
CX3CR1 deficient mice, Brain. Behav. Immun., 55, 126
the double hit hypothesis, Psychoneuroendocrinology, 34,
137, doi: 10.1016/j.bbi.2015.11.008.
1515 1525, doi: 10.1016/j.psyneuen.2009.05.010.
20.
Ridderstad Wollberg, A., Ericsson Dahlstrand, A.,
34.
Sarkisova, K. Y., Kulikov, M. A., Kudrin, V. S.,
Juréus, A., Ekerot, P., Simon, S., et al. (2014) Pharmaco
Midzyanovskaya, I. S., and Birioukova, L. M. (2014) Age
БИОХИМИЯ том 86 вып. 6 2021
856
КВИЧАНСКИЙ и др.
related changes in behavior, in monoamines and their
40. Ramirez, K., Shea, D. T., McKim, D. B., Reader, B. F.,
metabolites content, and in density of Dl and D2 dopamine
and Sheridan, J. F. (2015) Imipramine attenuates neuroin
receptors in the brain structures of WAG/Rij rats with depres
flammatory signaling and reverses stress induced social
sion like pathology, Zhurn. Vyss. Nervn. Deyatelnosti Im. I. P.
avoidance, Brain. Behav. Immun.,
46,
212220,
Pavlova, 64, 668 685, doi: 10.7868/S0044467714060094.
doi: 10.1016/j.bbi.2015.01.016.
35.
Ma, L., Xu, Y., Wang, G., and Li, R. (2019) What do we
41. Verboogen, D. R. J., Revelo, N. H., Ter Beest, M., and van
know about sex differences in depression: a review of ani
der Bogaart, G. (2019) Interleukin 6 secretion is limited by
mal models and potential mechanisms, Prog.
self signaling in endosomes, J. Mol. Cell Biol., 11, 144 157,
Neuropsychopharmacol. Biol. Psychiatry,
89,
4856,
doi: 10.1093/jmcb/mjy038.
doi: 10.1016/j.pnpbp.2018.08.026.
42. Kimura, A., and Kishimoto, T. (2010) IL 6: regulator of
36.
Dobryakova, Y. V., Kasianov, A., Zaichenko, M. I.,
Treg/Th17 balance, Eur. J. Immunol., 40, 1830 1835,
Stepanichev, M. Y., Chesnokova, E. A., et al.
(2018)
doi: 10.1002/eji.201040391.
Intracerebroventricular administration of 192IgG saporin
43. Hong, M., Zheng, J., Ding, Z. Y., Chen, J. H., Yu, L.,
alters expression of microglia associated genes in the dor
et al. (2013) Imbalance between Th17 and Treg cells may
sal but not ventral hippocampus, Front. Mol. Neurosci., 10,
play an important role in the development of chronic
doi: 10.3389/fnmol.2017.00429.
unpredictable mild stress induced depression in mice,
37.
Paxinos, G., Watson, C., Diego, S., Boston, L., and
Neuroimmunomodulation,
20,
3950, doi:
10.1159/
York, N. (1997) The Rat Brainin Stereotaxic Coordinates
000343100.
44. Sheridan, G. K., and Murphy, K. J. (2013) Neuron glia
(accessed on 9 February 2021).
crosstalk in health and disease: fractalkine and CX3CR1
38.
Young, K., and Morrison, H. (2018) Quantifying microglia
take centre stage, Open Biol., 3, 130181, doi: 10.1098/
morphology from photomicrographs of immunohisto
rsob.130181.
chemistry prepared tissue using imagej, J. Vis. Exp., 2018,
45. Hughes, P. M., Botham, M. S., Frentzel, S., Mir, A., and
57648, doi: 10.3791/57648.
Perry, V. H. (2002) Expression of fractalkine (CX3CL1)
39.
Stepanichev, M. Y., Goryakina, T., Manolova, A.,
and its receptor, CX3CR1, during acute and chronic
Lazareva, N., Kvichanskii, A., et al. (2021) Neonatal
inflammation in the rodent CNS, Glia, 37, 314 327,
proinflammatory challenge evokes a microglial response
doi: 10.1002/glia.10037.
and affects the ratio between subtypes of GABAergic
46. Bollinger, J. L., Collins, K. E., Patel, R., and Wellman,
interneurons in the hippocampus of juvenile rats: sex
C. L.
(2017) Behavioral stress alters corticolimbic
dependent and sex independent effects, Brain Struct.
microglia in a sex and brain region specific manner, PLoS
Funct., 1, 3, doi: 10.1007/s00429 020 02199 z.
One, 12, doi: 10.1371/journal.pone.0187631.
NEONATAL PROINFLAMMATORY STRESS AND EXPRESSION
OF GENES ASSOCIATED WITH NEUROINFLAMMATION
IN THE HIPPOCAMPUS OF RATS
A. A. Kvichansky*, L. V. Tret’yakova, M. N. Volobueva, A. O. Manolova, M. Yu. Stepanichev,
M. V. Onufriev, Y. V. Moiseeva, N. A. Lazareva, A. P. Bolshakov, and N. V. Gulyaeva
Institute of Higher Nervous Activity and Neurophysiology, Russian Academy of Sciences,
117485 Moscow, Russia; e)mail: al.kvichans@gmail.com
Differential effect of the neonatal proinflammatory stress (NPS) on the development of neuroinflammation in the
hippocampus and induction of the depressive like behavior in juvenile and adult male and female rats was studied.
NPS induction by bacterial lipopolysaccharide in the neonatal period upregulated expression of the Il6 and Tnf
mRNAs accompanied by the development of depressive like behavior in the adult male rats. NPS increased expres
sion of the mRNAs for fractalkine and its receptor in the ventral hippocampus of the juvenile male rats, but did not
affect expression of mRNAs for the proinflammatory cytokines and soluble form of fractalkine. NPS downregulated
expression of fractalkine mRNA in the dorsal hippocampus of juvenile males. No significant effects of NPS were
found in the female rats. Therefore, the NPS induces long term changes in the expression of neuroinflammation
associated genes in different regions of the hippocampus, which ultimately leads to the induction of neuroinflamma
tion and development of depressive like behavior in male rats.
Keywords: depression, neuroinflammation, cytokines, hippocampus, sex differences
БИОХИМИЯ том 86 вып. 6 2021