БИОХИМИЯ, 2021, том 86, вып. 6, с. 904 - 916
УДК 616 092.9
ДЕЙСТВИЕ НЕОНАТАЛЬНЫХ ВВЕДЕНИЙ БАКТЕРИАЛЬНОГО
ЭНДОТОКСИНА НА ПОВЕДЕНИЕ И ЭКСПРЕССИЮ ГЕНОВ
ИОНОТРОПНЫХ РЕЦЕПТОРОВ ГЛУТАМАТА В ГИППОКАМПЕ
ВЗРОСЛЫХ КРЫС ПОСЛЕ ПСИХОГЕННОЙ ТРАВМЫ
© 2021
В.А. Никитина1, М.В. Захарова2, А.Н. Трофимов1, А.П. Шварц2,
Г.В. Безнин1, С.Г. Цикунов1, О.Е. Зубарева2*
1 ФГБНУ «Институт экспериментальной медицины», 197376 Санкт Петербург, Россия
2 ФГБУН Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН,
194223 Санкт Петербург, Россия; электронная почта: zubarevaoe@mail.ru
Поступила в редакцию 06.12.2020
После доработки 26.04.2021
Принята к публикации 28.04.2021
Согласно «двухударной» гипотезе формирования психонейропатологии, инфекционные заболевания, пе
ренесённые в критические периоды раннего онтогенеза, нарушают нормальное развитие мозга, повышая
чувствительность к стрессам в последующей жизни. Эти нарушения могут быть связаны с изменениями
функциональной активности глутаматергической системы гиппокампа. В данной работе изучены особен
ности экспрессии генов ионотропных глутаматных NMDA (GluN1, GluN2a, GluN2b) и AMPA (GluA1,
GluA2) рецепторов, а также транспортёра глутамата EAAT2 в вентральном и дорзальном отделах гиппокам
па крыс, получавших бактериальный липополисахарид (ЛПС) в течение третьей недели жизни и пережив
ших витальный стресс (контакт с хищником - питоном) во взрослом возрасте. Тестирование производили
через 25 дней после стресса. Нарушения были обнаружены в вентральном, но не дорзальном отделе гиппо
кампа. Нестрессированные крысы, которым неонатально вводили ЛПС, имели более низкий уровень бел
ка GluN2b по сравнению с нестрессированным контролем, однако после стресса у них (но не у контроль
ных животных) уровень GluN2b значимо увеличивался. Различные реакции на стресс у экспериментальных
и контрольных крыс выявлены также по показателям исследовательского поведения: в первую минуту пос
ле помещения в «Отрытое поле» у стрессированных контрольных крыс активность увеличивалась, а у стрес
сированных экспериментальных - снижалась. Кроме того, стрессированные и нестрессированные крысы,
которым в раннем возрасте вводили ЛПС, отличались бóльшим временем, проведённым в открытых рука
вах «Приподнятого крестообразного лабиринта» и пониженным уровнем кортикостерона в крови. Таким
образом, воздействие бактериальным ЛПС в раннем постнатальном онтогенезе влияет на характер стресс
индуцированных изменений поведения и на особенности экспрессии генов субъединиц ионотропных ре
цепторов глутамата в гиппокампе после психогенной травмы, перенесённой во взрослом возрасте.
КЛЮЧЕВЫЕ СЛОВА: липополисахарид, раннее развитие, гиппокамп, стресс реактивность, NMDA ре
цептор, AMPA рецептор, кортикостерон, поведение.
DOI: 10.31857/S0320972521060129
ВВЕДЕНИЕ
сматривается, в частности, в рамках «двухудар
ной» гипотезы, согласно которой стрессы, ин
В последние годы активно обсуждается роль
фекционные заболевания и другие патологичес
раннего опыта в развитии повышенной уязви
кие состояния, имевшие место в критические
мости к стресс индуцированным психопатоло
периоды раннего онтогенеза, нарушают нор
гическим состояниям [1, 2]. Эта проблема рас
мальное развитие мозга, повышая чувствитель
ность к стрессам, перенесённым в подростко
вом и взрослом возрасте [3-5]. В качестве одно
Принятые сокращения: ЛПС - липополисахарид,
эндотоксин; ОП
- тест
«Открытое поле»; ПКЛ
-
го из повреждающих факторов могут выступать
тест «Приподнятый крестообразный лабиринт»; ПТСР -
бактериальные инфекции, экспериментальной
посттравматическое стрессовое расстройство; AMPA - моделью которых является введение элемента
α amino 3 hydroxy 5 methyl 4 isoxazolepropionic acid (α
клеточной стенки грамотрицательных бакте
амино 3 гидрокси 5 метил 4 изоксазолпропионовая кис
лота); EAAT2 - excitatory amino acid transporter 2 (возбуж
рий - липополисахарида (ЛПС, эндотоксин).
дающий аминокислотный транспортер 2); NMDA - N
Ранее было показано, что введение ЛПС в ран
methyl D aspartate (N метил D аспартат).
нем онтогенезе может оказывать долговремен
* Адресат для корреспонденции.
ное повреждающее действие на функции ЦНС,
904
НЕОНАТАЛЬНОЕ ВОСПАЛЕНИЕ И СТРЕСС РЕАКТИВНОСТЬ
905
вызывая нарушение когнитивных функций и
делью депрессии [34], и у взрослых крыс, пере
эмоционального поведения [6-11]. Ряд данных
живших социальный стресс (разлучение с ма
указывает на изменение стресс реактивности
терью) в неонатальном периоде [35]. Особен
животных, которым вводили ЛПС в раннем воз
ности экспрессии гена, кодирующего EAAT2, в
расте [12, 13].
мозге взрослых стрессированных животных, ко
Механизмы ЛПС индуцированных наруше
торым в раннем возрасте вводили ЛПС, ранее не
ний поведения и стресс реактивности остаются
изучались.
малоизученными. В качестве одного из возмож
Целью данной работы явилось исследование
ных механизмов рассматривается нарушение
сочетанных влияний неонатальных введе
функциональной активности глутаматергичес
ний ЛПС и витального стресса, перенесённого
кой системы мозга, связанное, в частности, с
во взрослом возрасте, на поведение, уровень
нарушением формирования ионотропных
кортикостерона, а также продукцию EAAT2 и
NMDA и AMPA глутаматных рецепторов [14].
субъединиц NMDA и AMPA рецепторов в дор
Эти рецепторы имеют сложную субъединичную
зальной и вентральной областях гиппокампа
структуру. NMDA рецепторный комплекс
крыс. Модель стресса (контакт крыс с хищни
представляет собой гетеротетрамер, состоящий
ком - питоном) и сроки тестирования были
из облигатной GluN1 субъединицы и вариант
выбраны на основе исследований, проведённых
ных субъединиц GluN2 (a-d) либо GluN3 (a, b),
нами ранее, так как было показано, что данный
благодаря которым обеспечивается функцио
вид стресса влияет на экспрессию изучаемых ге
нальная и региональная вариативность NMDA
нов [36]. Необходимость дифференциального
рецепторов [15]. AMPA рецепторы состоят из
анализа вентрального и дорзального отделов
4 субъединиц GluA (1-4). AMPA рецепторы, со
гиппокампа была обусловлена функциональны
держащие субъединицу GluA2, непроницаемы
ми различиями этих областей, а также тем, что
для ионов кальция [16]. Формирование харак
стресс по разному регулирует экспрессию генов
терного для взрослых особей субъединичного
рецепторов глутамата и EAAT2 в его дорзальном
состава NMDA и AMPA рецепторов в мозге
и вентральном отделах [27, 37].
крыс происходит в первые недели жиз
ни [17-21]. Введение ЛПС в эти сроки приводит
к краткосрочным и долговременным изменени
МАТЕРИАЛЫ И МЕТОДЫ
ям экспрессии генов субъединиц NMDA и
AMPA рецепторов в гиппокампе и коре моз
Объект исследования. Общий план экспери
га [10, 11, 14].
ментов. Исследование выполнено на самцах
Хорошо известно, что NMDA и AMPA ре
крыс Вистар с соблюдением принципов гуман
цепторы принимают участие в реализации ког
ности (Директивы Европейского Сообщества
нитивных функций [22-25] и психоэмоцио
№ 86/609 ЕС), одобренных Локальным этичес
нальных реакций [26]. Экспрессия генов этих
ким комитетом ФГБНУ «ИЭМ». Схема экспе
рецепторов в гиппокампе изменяется при стрес
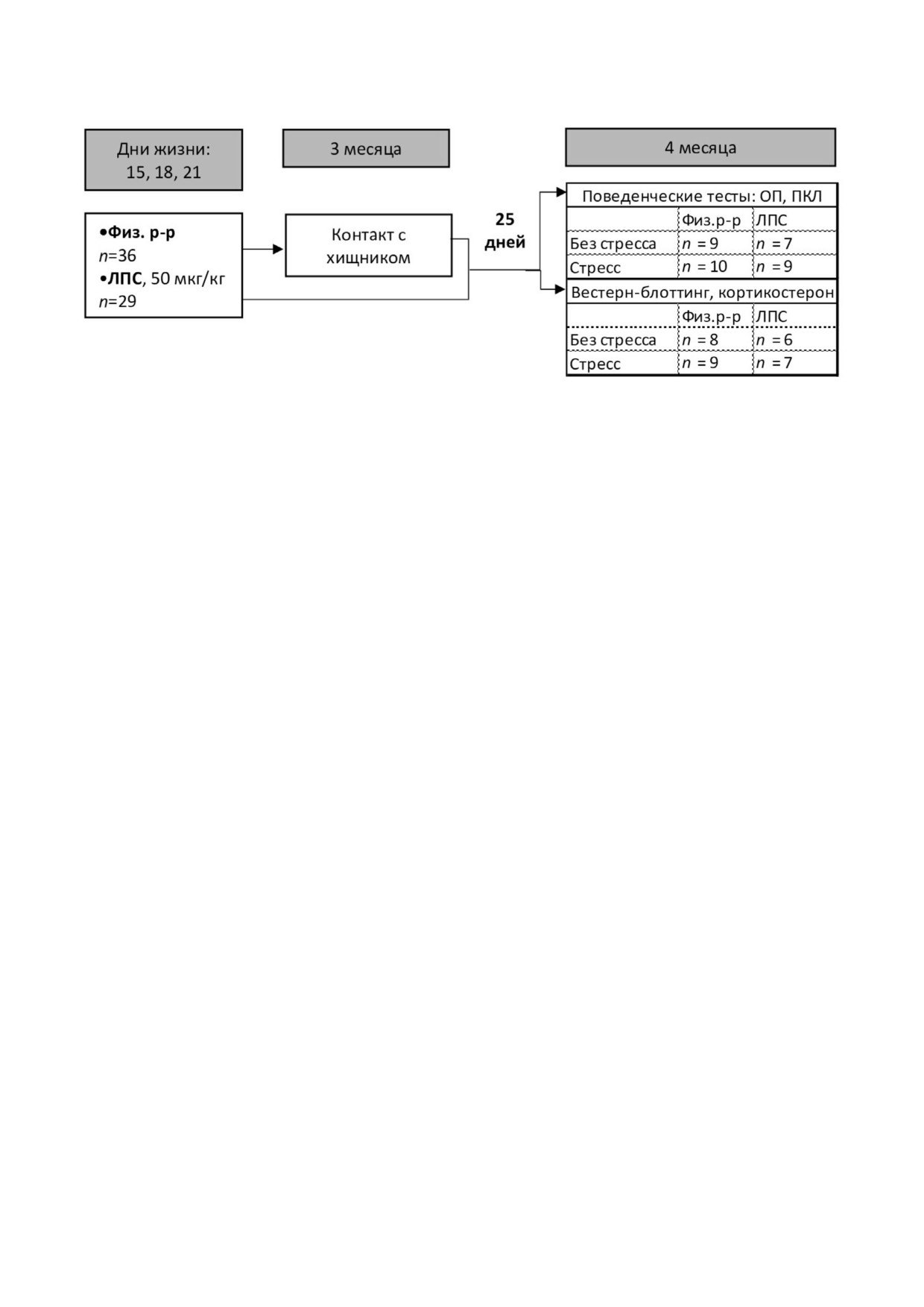
римента представлена на рис. 1. Крысят содер
се [27, 28]. Предполагается, что ионотропные
жали с матерями, по одному помёту в клет
глутаматные рецепторы опосредуют угасание
ке (всего 14 помётов). Количество крысят в по
страха; нарушение этой реакции играет ключе
мете выравнивали (в помёте оставляли не более
вую роль в патофизиологии посттравматическо
7-8 крысят), часть самок при необходимости
го стрессового расстройства (ПТСР) [29, 30].
оставляли, но в экспериментах не использовали.
Использование антагонистов NMDA рецепто
Бактериальный липополисахарид (серотип
ров позволяет предотвратить стресс индуциро
055:B5 Escherichia coli,
50
мкг/кг;
«Sigma
ванные гормональные и поведенческие наруше
Aldrich», США) либо апирогенный физиологи
ния [31, 32], что доказывает их вовлеченность в
ческий раствор вводили внутрибрюшинно 1 раз
регуляцию этих реакций.
в сутки, на 15, 18 и 21 дни жизни. Крысят разлу
Ещё одним белком, влияющим на актив
чали с матерями не более чем на одну минуту.
ность глутаматергической системы, является
В каждом помёте были и контрольные, и экспе
транспортёр глутамата EAAT2 (GLT 1). Он про
риментальные животные. В возрасте 1 мес. кры
дуцируется в основном астроцитами и обеспе
сят отсаживали от самки. Дозировка ЛПС была
чивает обратный захват этими клетками до
выбрана на основе ранее проведённых исследо
90% глутамата из синаптической щели, являясь,
ваний как умеренно пирогенная и влияющая на
таким образом, основным регулятором уровня
экспрессию генов субъединиц NMDA и
внеклеточного глутамата [33]. Изменение про
AMPA рецепторов [10].
дукции EAAT2 в гиппокампе отмечается при
Стрессирование. В возрасте 3 мес. половину
выученной беспомощности, являющейся мо
экспериментальных (n = 16) и контрольных
БИОХИМИЯ том 86 вып. 6 2021
906
НИКИТИНА и др.
Рис. 1. Схема эксперимента
(n = 19) животных подвергали стрессу, заключав
на рис. S1 в Приложении. Выделенные структу
шемуся в переживании обстоятельств гибели со
ры мозга гомогенизировали на льду в оптими
родича и угрозы собственной жизни от действий
зированном лизирующем буфере, предложен
хищника - питона [38]. Для этого группу, состо
ном Kopec et al. [40], c добавлением 1× коктей
ящую из опытных и контрольных крыс (в общей
ля ингибиторов протеаз (Pierce Protease
сложности 17-20 животных), помещали в терра
Inhibitor Tablets, «Thermo Fisher Scientific»,
риум к голодному питону, одна из них станови
США), инкубировали в течение 60 мин при
лась жертвой пищевых потребностей хищника.
комнатной температуре, нерастворённые остат
Остальные крысы, находившиеся в террариуме,
ки удаляли центрифугированием
(15
мин,
подвергались переживанию стрессовой ситуа
14 000 g, 20 °C, «Thermo Scientific»). Концентра
ции: они находились в установке ещё в течение
цию белка в пробе определяли с помощью мо
20-25 мин. Затем крыс изымали из террариума и
дифицированного метода Лоури [41]. Белковый
возвращали в домашние клетки, в которых они
супернатант был разбавлен в соотношении 1/1
содержались до проведения поведенческого тес
2× буфером для нанесения (125 мМ Tris HCl,
тирования или забора биологического материала
рН 6,8; 40% (v/v) глицерин; 4% додецилсульфат
для дальнейшего анализа. Для сравнения ис
натрия; 2,5% β меркаптоэтанол; 0,02% бромфе
пользовали нестрессированных эксперимен
ноловый синий) и инкубировался в термоста
тальных и контрольных животных.
те («BioSan», Латвия) в течение 15 мин при
Поскольку нас интересовали долговремен
70 °C. Электрофоретическое разделение прово
ные стресс индуцированные изменения, экспе
дили в восстанавливающих и денатурирующих
рименты проводили через 25 суток после стрес
условиях [42] в 7% ном полиакриламидном геле
са и в аналогичном возрасте у нестрессирован
вместе со стандартом молекулярного веса
ных животных. Разные группы контрольных и
(Thermo Scientific PAGE Ruler Prestained Protein
экспериментальных крыс были использованы
Ladder 10-170 кДа; «Thermo Fisher Scientific»,
для проведения биохимических (физ. р р,
США) при 125 В. В одинаковом объёме на до
n = 17; ЛПС, n = 13) и поведенческих (физ. р р,
рожки наносили по 6 мкг белка, т.к. это количе
n = 19; ЛПС, n = 16) исследований.
ство позволяло попасть в линейную область
Определение уровня белка методом вестерн;
денситометрического анализа для всех исполь
блоттинга. Исследование уровня белков субъ
зуемых антител в наших условиях. Вместе с ана
единиц NMDA (GluN2a, GluN2b) и AMPA
лизируемыми образцами на каждый гель нано
(GluA1, GluA2) рецепторов проводили методом
сили образец калибратор, полученный при
вестерн блоттинга. Животных декапитировали,
смешивании нескольких образцов от животных
мозг выделяли целиком, немедленно замора
из разных групп.
живали и хранили при температуре -70 °С. Вен
Белок переносили на нитроцеллюлозную
тральный и дорзальный гиппокамп выделяли
мембрану (диаметр пор 45 мкм) полусухим пере
на срезах, производимых на микротоме криос
носом с 1× буфером для переноса Invitrogen
тате Thermo Scientific MICROM HM («Thermo
Power Blotter 1 Step Transfer Buffer («Thermo
Scientific», США), при -20 °C, согласно атла
Fisher Scientific»), согласно инструкции произ
су [39]. Схема выделения структур представлена
водителя. После переноса мембрану окрашива
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНОЕ ВОСПАЛЕНИЕ И СТРЕСС РЕАКТИВНОСТЬ
907
ли 0,1% ным раствором красителя Ponсeau S в
рессии. Все эксперименты проводились в двух
5% ной уксусной кислоте
(«Merck KGaA»,
независимых повторах.
ФРГ), результаты документировали с помощью
Поведенческое тестирование. Ориентировоч
системы гель визуализации ChemiDoc MP
но исследовательское поведение и уровень тре
(«Bio Rad», США). В качестве блокирующего
вожности крыс оценивали в тестах «Открытое
агента использовали обезжиренное сухое моло
поле» (ОП) и «Приподнятый крестообразный
ко («Sigmaaldrich», Швейцария) (5% ный раст
лабиринт» (ПКЛ). Использовали круглое от
вор, 1,5 ч при комнатной температуре). Затем
крытое поле диаметром 1 м, освещённость 10 лк.
мембрану трижды промывали буфером PBS T
Крысу помещали в центр поля, длительность
(0,01 M фосфатный буфер, рН 7,4; 137 мМ NaCl;
теста составляла 3 мин. Анализировали общую
2,7 мМ КCl, содержащий 0,1% (v/v) Tween 20) и
длину пройденного пути (как показатель двига
инкубировали в течение ночи при +4 °С в раст
тельной и исследовательской активности), дис
воре первичных антител против GluN2a
танцию, пройденную за первую минуту (как по
(ab169873, кроличьи поликлональные); GluN2b
казатель ориентировочно исследовательского
(ab65783, кроличьи поликлональные), GluA1
поведения), время нахождения в центральной и
(ab109450, кроличьи моноклональные); GluA2
периферической областях (показатель тревож
(ab106515, мышиные моноклональные); EAAT2
ности), а также проводили анализ общей страте
(ab205248, кроличьи моноклональные). В работе
гии поведения (характер трека, подробно описа
использовали первичные антитела фирмы
но в результатах).
«Abcam», Великобритания в разведении 1/1000.
Тест ПКЛ использовали для оценки уровня
Для детекции первичных антител (для
тревожности. Установка состояла из централь
GluN2a/2b, GluA1 и EAAT2) к мембране добав
ной платформы (10 × 10 см) с двумя открытыми
ляли антитела против кроличьих иммуноглобу
и двумя закрытыми «рукавами» (50 × 10 см),
линов G (31460, 1/60 000, Pierce Goat anti rabit
приподнятыми на 40 см над полом. Закрытые
IgG HRP, «Thermo Fisher Scientific»), в осталь
«рукава» имели 30 сантиметровые стены и ос
ных случаях к мембране добавляли антитела
вещённость 5 лк. На открытых «рукавах» осве
против мышиных иммуноглобулинов G (ab6808,
щённость составляла 10 лк. Тест длился 5 мин.
1/40 000, Sheep Anti Mouse IgG H&L (HRP),
Крысу помещали в один из закрытых «рукавов»
(«Abcam»)). Все растворы антител и блокирую
лабиринта. Измеряли время нахождения в за
щего агента были приготовлены в фосфатно со
крытом и открытом рукавах, время выглядыва
левом буферном растворе PBS T; в этом же бу
ния из закрытых рукавов, число заходов в от
фере проводили 3-6 кратную промывку между
крытые и закрытые рукава, процент времени,
шагами. Хемилюминесцентный сигнал получа
проведённого в открытых рукавах, и время гру
ли с хемилюминесцентного субстрата Super
минга в закрытых «рукавах». Все поведенческие
Signal™ West Pico PLUS
(«Thermo Fisher
эксперименты регистрировали с помощью веб
Scientific») и документировали с помощью сис
камер, размещённых над установками. Анализ
темы ChemiDoc MP («Bio Rad», США). Денси
видеозаписей производился с использованием
тометрический анализ проводили с помощью
пользовательского программного обеспечения
программы Image Lab 6.0.1 software (Bio Rad»):
(Field 4W, Pole 7, Pole_Krest), разработанного в
оптическую плотность сигнала нормировали по
Физиологическом отделе им. И.П. Павлова
отношению к образцу калибратору по сигналу
ФГБНУ «ИЭМ».
Ponсeau S (тотальный белок) соответствующей
Статистическую обработку производили в
дорожки.
программе SPSS Statistics 22 («IBM Corp.», США)
Определение содержания кортикостерона в
с использованием критерия Колмогорова-
крови. Уровень кортикостерона определяли в
Смирнова для оценки нормальности распределе
сыворотке периферической крови, взятой при
ния, критерия Ливиня (проверка равенства дис
декапитации животных, методом конкурентно
персий), двухфакторного дисперсионного анали
го иммуноферментного анализа с использова
за, t критерия Стьюдента c поправкой Бонфер
нием набора реагентов для количественного оп
рони в качестве апостериорного теста. Критерий
ределения кортикостерона (Corticosterone
χ2 Пирсона был использован для анализа меж
(Human, Rat, Mouse) ELISA (RE52211),
групповых различий в частотах встречаемости
«TECAN Trading», Швейцария), согласно реко
разных типов треков в тесте ОП. Различия счита
мендациям производителя. Спектрофотометри
ли достоверными при р < 0,05. Данные на графи
ческий анализ производили на Microplate
ках представлены в виде среднего значе
Reader Immunochem 2100 («HTI Diagnostics»,
ния ± стандартная ошибка. Диаграммы построе
США). Расчёт значений концентраций корти
ны с применением программы GraphPad Prism 8
костерона производили методом линейной рег
(«GraphPad Software, Inc.», США).
БИОХИМИЯ том 86 вып. 6 2021
908
НИКИТИНА и др.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
ролем (n = 6) (рис. 2, t = 4,75; p = 0,002, досто
верно с учётом поправки Бонферрони).
Использованная доза ЛПС 50 мкг/кг не вли
Выявлено также влияние введений ЛПС на
яла значимо на развитие животных - динамику
соотношение содержания субъединиц GluN2a/
веса тела (количественные данные представле
GluN2b: оно было выше у крыс с введением эн
ны на рис. S2 и в табл. S1 в Приложении).
дотоксина (F(1,17) = 6,97; p = 0,02). Особенно это
Экспрессия генов субъединиц инотропных
проявлялось в обычных (без стресса) услови
глутаматных рецепторов была исследована на
ях (рис. 2).
уровне белка. На рис. S3 в Приложении пред
Различные реакции на стресс у эксперимен
ставлены все проанализированные изображе
тальных и контрольных крыс выявлены по по
ния мембран. Более выраженные межгрупповые
казателям экспрессии GluN2b субъединицы
различия выявлены в вентральном (по сравне
NMDA рецепторов и GluA1 субъединицы
нию с дорзальным) отделе гиппокампа. В груп
AMPA рецепторов (рис. 2 и 3; взаимодействие
пе нестрессированных крыс более низкий уро
факторов группы и стресса, соответственно,
вень GluN2b субъединицы NMDA рецепторов
F(1,20) = 4,71; p = 0,04 и F(1,19) = 4,87; p = 0,04).
обнаружен у животных, которым в раннем воз
Уровень белка GluN2b субъединицы у экспери
расте вводили ЛПС (n = 4) по сравнению с конт
ментальных крыс увеличивался после стресса
Рис. 2. Экспрессия генов субъединиц NMDA рецепторов в гиппокампе экспериментальных и контрольных крыс. Коли
чество животных в группах n = 4-8. Каждая точка означает одно животное. F - Критерий Фишера, двухфакторный
ANOVA; * - достоверные отличия от контрольной группы, # достоверные отличия от группы без стресса; p < 0,05, t кри
терий Стьюдента с поправкой Бонферрони. Приведены примеры репрезентативных бендов для животных из всех групп
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНОЕ ВОСПАЛЕНИЕ И СТРЕСС РЕАКТИВНОСТЬ
909
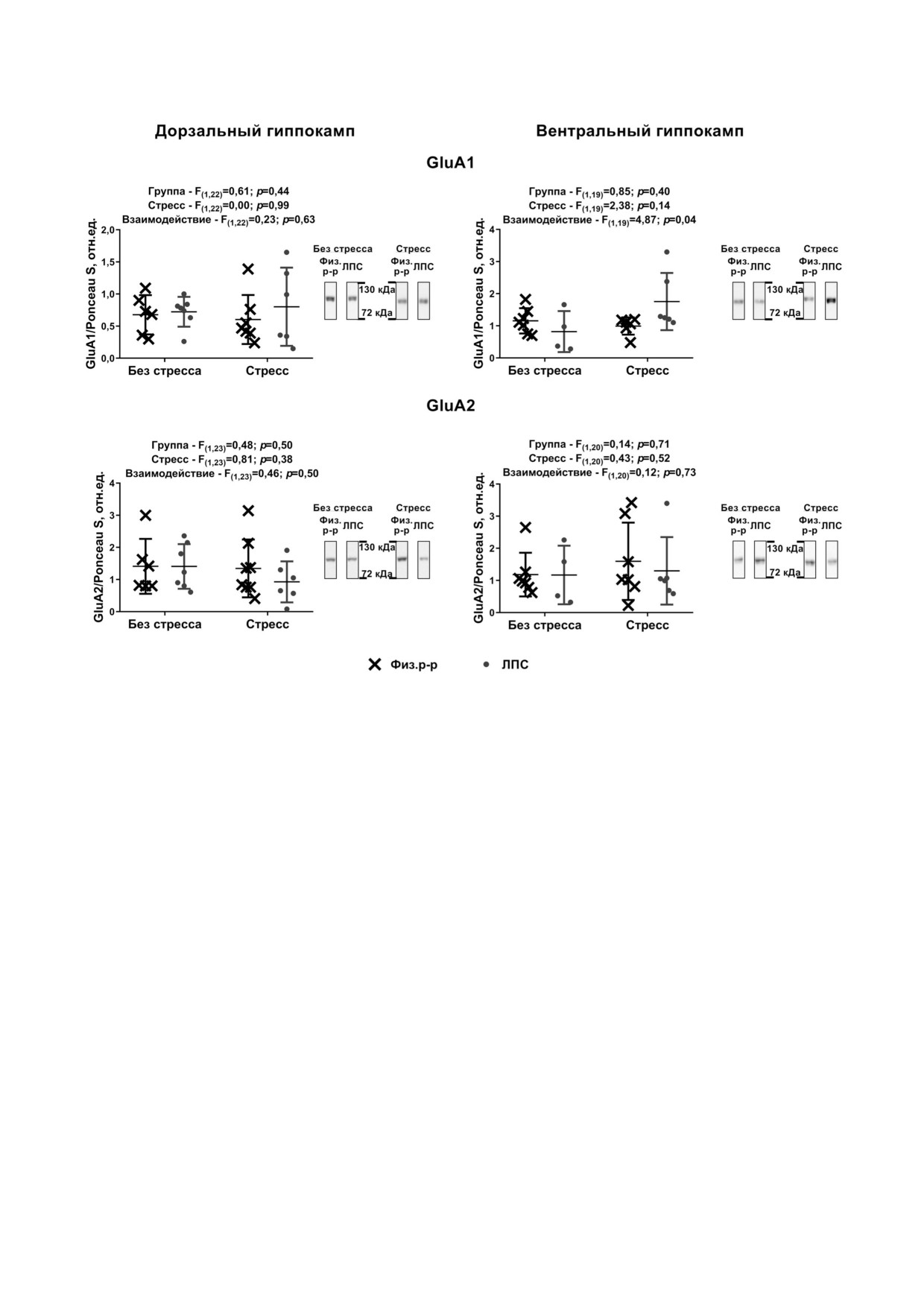
Рис. 3. Экспрессия генов субъединиц AMPA рецепторов в гиппокампе экспериментальных и контрольных крыс. Коли
чество животных в группах n = 4-8. Каждая точка означает одно животное. F - Критерий Фишера, двухфакторный
ANOVA. Приведены примеры репрезентативных бендов для животных из всех групп
(t = 3,04; p = 0,025), у контрольных - имел тен
от активности GluN2b содержащих NMDA ре
денцию к снижению в сравнении с соответству
цепторов [43, 44].
ющей нестрессированной группой.
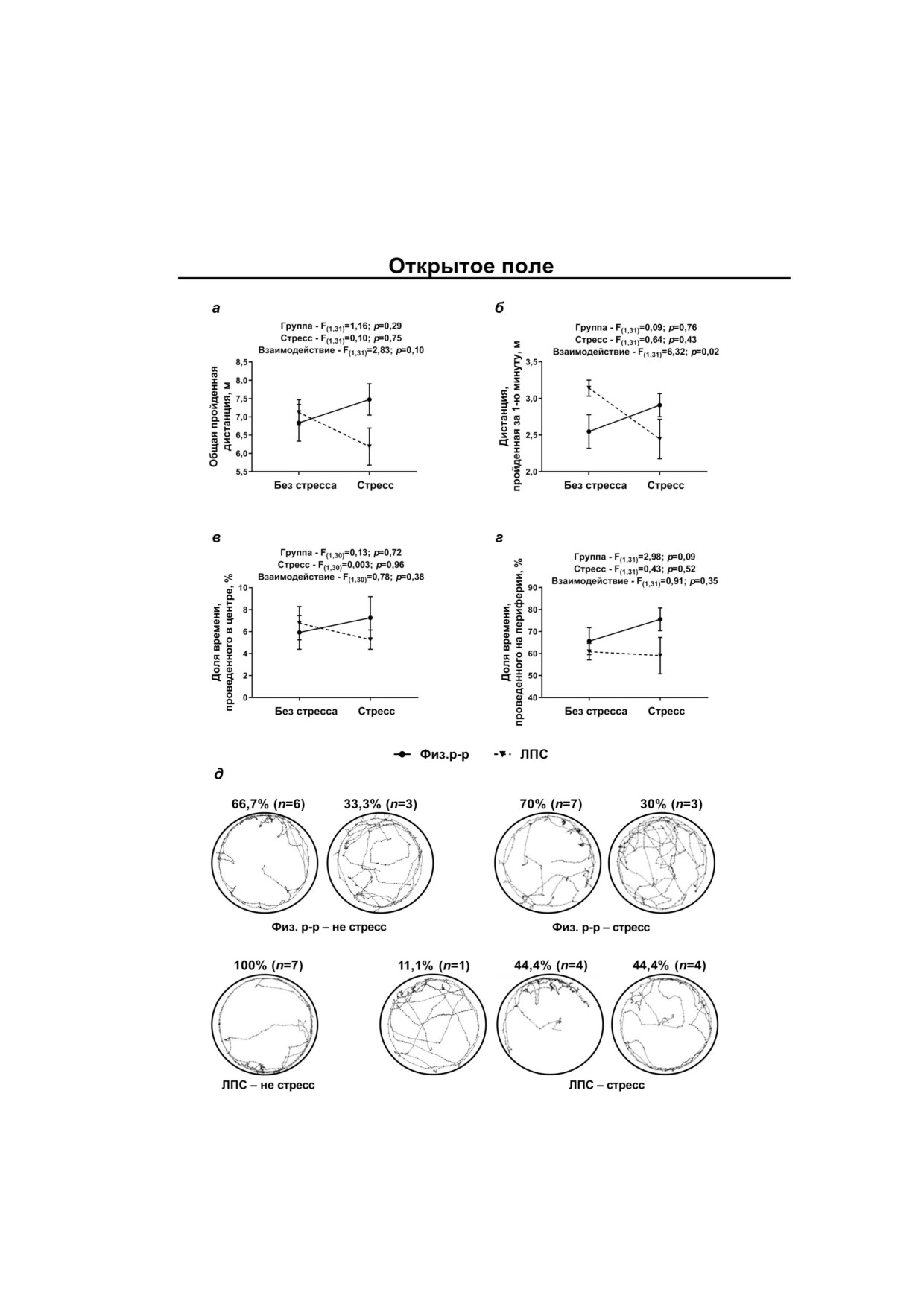
Результаты тестирования поведения показа
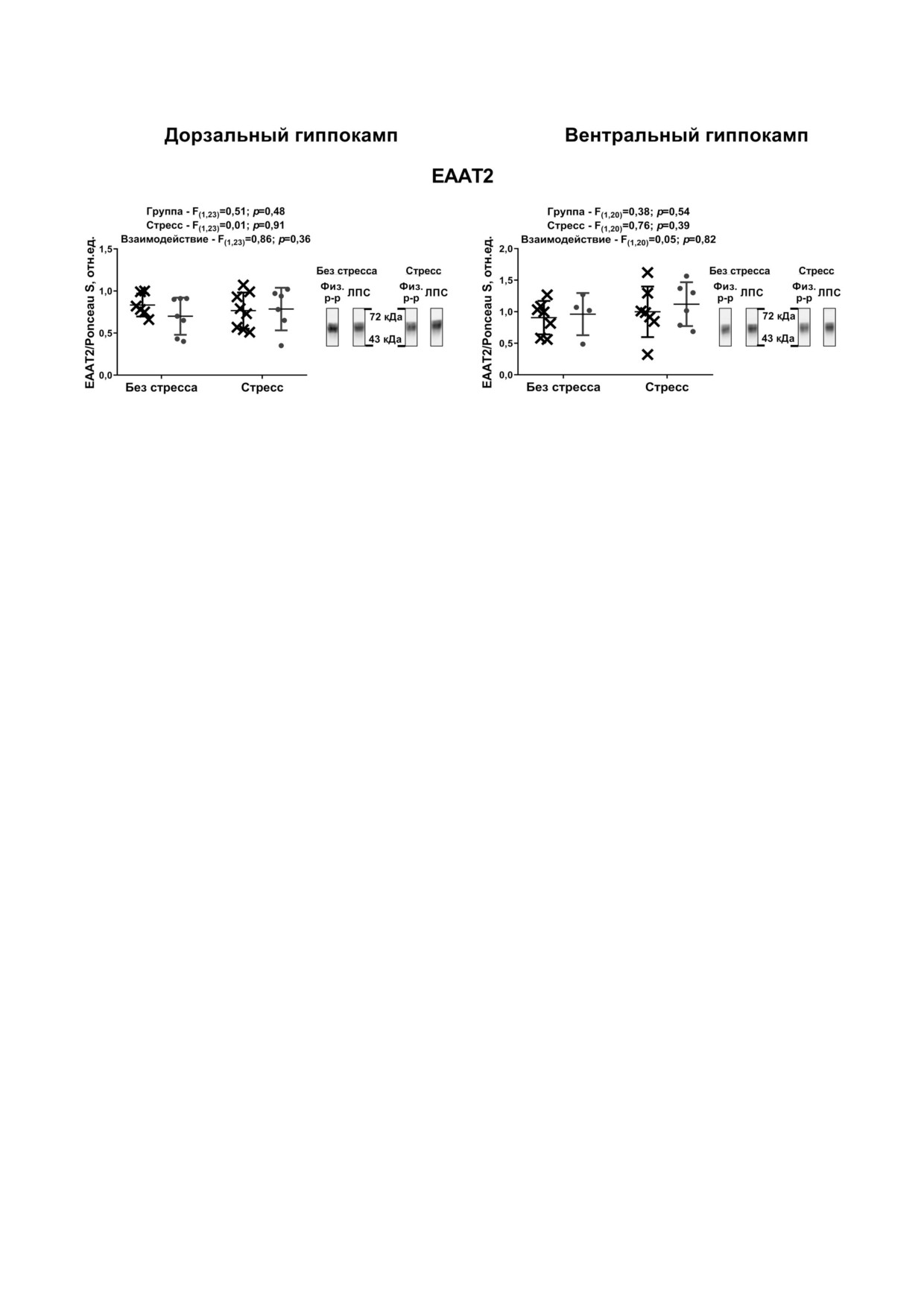
Никаких достоверных изменений не показа
ли, что в тесте ОП (рис. 5) общая длина прой
но в отношении продукции EAAT2 как в вент
денной дистанции достоверно не отличалась в
ральном, так и дорзальном отделе гиппокам
группах, однако активность в течение первой
па (рис. 4).
минуты (показатель ориентировочно исследо
Таким образом, эффекты неонатального вве
вательского поведения) была различной: у конт
дения ЛПС на экспрессию генов субъединиц
рольных крыс, перенёсших стресс, она увеличи
ионотропных рецепторов глутамата выявлены в
валась, у экспериментальных - снижалась (взаи
вентральном, но не дорзальном отделе гиппо
модействие факторов стресса и группы:
кампа. Наиболее выраженные нарушения в
F(1,31) = 6,32; p = 0,017). Достоверных различий
обычных условиях и стресс индуцированные
по времени нахождения в центральной и пери
изменения обнаружены в отношении GluN2b
ферической зоне поля не выявлено, однако ха
субъединицы NMDA рецепторов. Для того что
рактер треков в группах различался. Было выде
бы проверить, могут ли эти изменения влиять на
лено три типа трека. Первый, с преимуществен
поведение после психогенной травмы, нами в
ным нахождением на периферии (обследование
той же экспериментальной парадигме было
всего поля), отмечался у большинства нестрес
проведено тестирование крыс в «Открытом по
сированных контрольных и экспериментальных
ле» и «Приподнятом крестообразном лабирин
животных (соответственно 66,7% и 70%), а так
те», поскольку поведение в этих тестах зависит
же у 100% контрольных стрессированных крыс.
БИОХИМИЯ том 86 вып. 6 2021
910
НИКИТИНА и др.
Рис. 4. Экспрессия генов, кодирующих EAAT2, в гиппокампе экспериментальных и контрольных крыс. Количество жи
вотных в группах n = 4-8. Каждая точка означает одно животное. F - Критерий Фишера, двухфакторный ANOVA. При
ведены примеры репрезентативных бендов для животных из всех групп
Второй, с частыми заходами в центральную об
Для проверки предположения о том, что эти
ласть поля, был характерен для трети контроль
особенности могут быть связаны с изменением
ных и экспериментальных крыс без стресса.
уровня кортикостерона, мы проанализировали
Третий тип трека - обследование только части
его содержание в крови контрольных и экспери
поля - был описан только у 44,4% эксперимен
ментальных крыс, имевших и не имевших пси
тальных крыс, переживших стресс. Очевидно,
хогенную травму. Было показано, что введение
он свидетельствует о пониженной исследова
эндотоксина в раннем возрасте влияет на уро
тельской активности, а также, возможно, повы
вень кортикостерона в крови. Он был ниже у
шенной тревожности. Частота встречаемости
экспериментальных крыс, как подвергавшихся,
разных типов треков достоверно различается в
так и не подвергнутых стрессу (рис. 7, фактор
четырёх группах (χ2 = 16,4; p = 0,012).
введения ЛПС: F(1,23) = 6,22; p = 0,02).
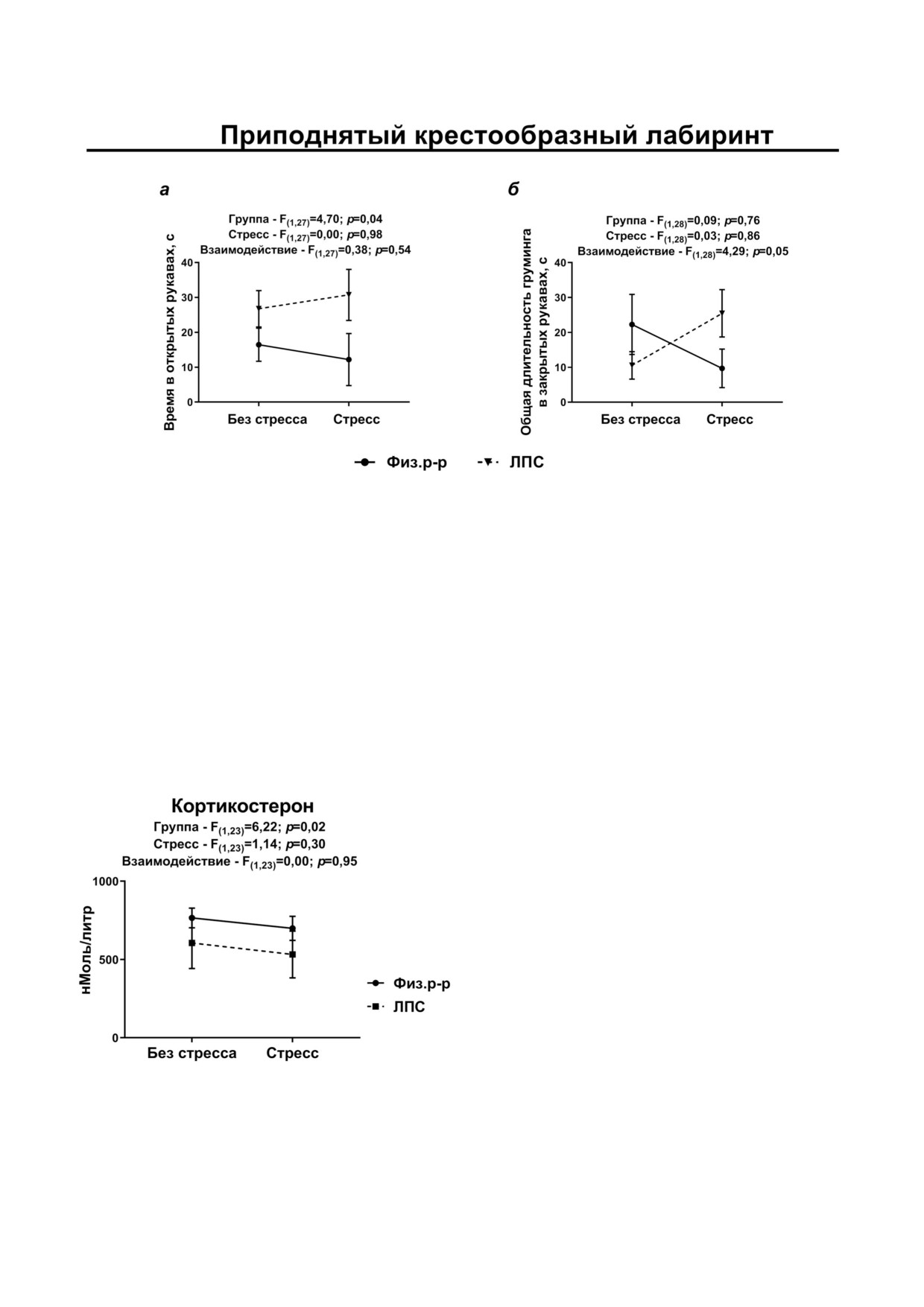
В тесте ПКЛ длительность нахождения на
Таким образом, проведённое исследование
открытых «рукавах» была выше у стрессирован
показало, что введение бактериального ЛПС в
ных и нестрессированных экспериментальных
критический для формирования ионотропных
крыс (рис. 6, а; фактор группы: F(1,27) = 4,70;
глутаматных рецепторов период раннего пост
p = 0,04), аналогичные результаты получены для
натального онтогенеза (фактор группы) влияет
процента времени, которое животные проводят
на соотношение экспрессии генов, кодирующих
в открытых рукавах (фактор группы: F(1,27) =
субъединицы GluN2a/GluN2b в клетках вент
= 4,56; p = 0,04; рис. S4 и табл. S2 в Приложе
рального гиппокампа, на поведение в тес
нии), этот результат соответствовал тенденции
те ПКЛ и на уровень кортикостерона в крови.
меньшего времени, проведённого эксперимен
Различные реакции на стресс у эксперименталь
тальными крысами на периферии «Открытого
ных и контрольных крыс (сочетанное влияние
поля» (F(1,31) = 2,98; p = 0,09). Однако эти резуль
факторов группы и стресса) выявлены в отно
таты, по видимому, нельзя однозначно тракто
шении экспрессии генов GluN2b субъединицы
вать как пониженную тревожность, так как у
NMDA рецепторов и GluA1 субъединицы
стрессированных экспериментальных крыс при
AMPA рецепторов, а также для показателей ис
этом выявляется усиление груминга в закрытых
следовательского и эмоционального поведения.
«рукавах» (рис. 6, б; взаимодействие факторов
стресса и группы: F(1,28) = 4,29; p = 0,048). По
длительности нахождения в закрытых «рукавах»,
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
длительности актов выглядывания из них, а так
же числу заходов в открытые и закрытые «рука
В работе впервые исследованы особенности
ва» различий не было выявлено (данные предс
экспрессии генов субъединиц ионотропных ре
тавлены на рис. 5 и в табл. S2 в Приложении).
цепторов глутамата в гиппокампе крыс после
Таким образом, поведенческие реакции на
сочетанного воздействия неонатальных введе
стресс были различными у крыс с неонатальным
ний ЛПС и психогенной травмы, перенесённой
введением ЛПС и физиологического раствора.
во взрослом возрасте.
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНОЕ ВОСПАЛЕНИЕ И СТРЕСС РЕАКТИВНОСТЬ
911
Значимое влияние фактора группы (введе
в вентральном отделе гиппокампа. Увеличение
ний ЛПС) показано для поведения в тесте ПКЛ
соотношения GluN2a/GluN2b ранее было пока
и для соотношения GluN2a/GluN2b в вентраль
зано у взрослых крыс, перенёсших вирусную ин
ном гиппокампе. Нами выявлено также, что
фекцию пренатально (после введения беремен
нестрессированные экспериментальные крысы
ным самкам синтетической молекулы polyI:C),
отличаются пониженным уровнем белка GluN2b
однако в этом случае соотношение увеличива
Рис. 5. Поведение в тесте «Открытое поле»: а - общая дистанция, б - дистанция, пройденная в течение первой минуты,
в - время в центральной области, г - время на периферии, д - типы треков. F - Критерий Фишера, двухфакторный
ANOVA; n - количество животных
БИОХИМИЯ том 86 вып. 6 2021
912
НИКИТИНА и др.
Рис. 6. Поведение в тесте «Приподнятый крестообразный лабиринт»: а - время в открытых рукавах, б - время груминга
в закрытых рукавах. F - Критерий Фишера, двухфакторный ANOVA
лось за счёт усиления продукции GluN2a субъ
ном отделах гиппокампа [11], однако у взрослых
единицы [45]. Аналогично повышение продук
животных после аналогичных введений ЛПС,
ции мРНК субъединицы GluN2a отмечено у
напротив, нами было обнаружено увеличение
взрослых крыс, которым на 14 день жизни од
уровня мРНК GluN2b [10]. Активность NMDA
нократно вводили достаточно высокую дозу
и AMPA рецепторов напрямую связана с долго
ЛПС (100 мкг/кг) [14]. Соотношение GluN2a/
временной потенциацией нейронов гиппокам
GluN2b в этом случае не оценивали. В исследо
па; неонатальные введения ЛПС приводят к её
ваниях, проведённых нами ранее на подростках
нарушению у неполовозрелых животных [11,
крыс, которым вводили ЛПС в течение третьей
46-48]. Нужно отметить, что более выраженные
недели жизни, выявлено снижение уровня белка
изменения обнаружены нами в вентральном от
GluN2b субъединицы и увеличение соотноше
деле гиппокампа. Ранее Onufriev et al. [49] было
ния GluN2a/GluN2b в дорзальном и вентраль
показано, что введение ЛПС взрослым крысам
вызывает различную динамику изменений ак
тивности клеток вентрального и дорзального от
делов гиппокампа (по показателям долговре
менной потенциации и экспрессии генов про
воспалительных цитокинов): максимальная ре
акция в дорзальном отделе гиппокампа отмеча
лась раньше, в вентральном - позднее.
ЛПС индуцированные перестройки, проис
ходящие в субъединичном составе ионотропных
рецепторов глутамата, могут влиять на поведе
ние [43, 44]. Нарушение исследовательского по
ведения и изменение уровня тревожности после
неонатальных введений ЛПС у взрослых живот
ных ранее были показаны во многих исследова
ниях [46, 50-52]. В частности, введение ЛПС на
третьей неделе жизни приводило у взрослых
крыс к нарушению обследования новых объек
Рис. 7. Уровень кортикостерона в сыворотке крови конт
тов [53] и снижению количества стоек с упором
рольных (сплошная линия) и экспериментальных (пунк
в тесте ОП [10]. Признаки тревожно депрессив
тирная линия) крыс в обычных условиях и после стресса.
F - Критерий Фишера, двухфакторный ANOVA. Количе
ного состояния отмечаются у взрослых крыс и
ство животных в группах n = 6-8
мышей, которым вводили ЛПС на 1 либо на 3 и
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНОЕ ВОСПАЛЕНИЕ И СТРЕСС РЕАКТИВНОСТЬ
913
5 дни жизни [46, 50, 54]. Однако другие авто
тамата подобное исследование проведено впер
ры [55] отмечают пониженную тревожность (по
вые. При этом исследования особенностей
времени пребывания на открытых рука
стресс реактивности у животных, которым нео
вах ПКЛ) у подростков крыс, которым вводили
натально вводили ЛПС, по поведенческим и
100 мкг/кг ЛПС на 5 сутки жизни. Снижение
гормональным показателям проводились и ра
времени нахождения в открытых рукавах ПКЛ
нее. Так, Shanks et al. [60] обнаружили повышен
отмечалось нами ранее у взрослых крыс, кото
ный выброс адренокортикотропного гормо
рым вводили ЛПС в течение третьей недели
на (АКТГ) и кортикостерона в ответ на ограни
жизни [10].
чение движений у взрослых крыс, которым вво
Основная идея настоящего исследования
дили эндотоксин на 3 и 5 сутки жизни. Кроме
заключалась в изучении особенностей реакции
того, у экспериментальных животных отмеча
на психогенную травму у взрослых крыс, кото
лось снижение чувствительности к глюкокорти
рым вводили ЛПС в раннем возрасте. Экспери
коидам, то есть нарушались механизмы отри
ментальные ситуации угрозы жизни, в частнос
цательной обратной связи регуляции стресс ре
ти контакта лабораторных грызунов с хищни
акции. Аналогичные результаты - более силь
ком (или его запахом), часто используются для
ная или пролонгированная реакция на
моделирования ПТСР [56]. Для данного рас
стресс (по уровню кортикостерона) при введе
стройства характерно отсроченное развитие
нии ЛПС в раннем возрасте - получены и в дру
психоневрологических нарушений [57]. Ранее
гих работах [13, 46]. При этом Walker et al. [12],
модель, использованная нами в данном исследо
вводившие ЛПС в те же сроки, отмечают у
вании, была применена для анализа динамики
взрослых экспериментальных крыс ослабление
постстрессовых изменений экспрессии генов
гормонального ответа на трёхдневный комби
субъединиц NMDA и AMPA рецепторов в гип
нированный стресс, хотя на поведенческом
покампе, медиальной префронтальной коре и
уровне стресс реактивность эксперименталь
миндалевидном теле крыс. Наиболее выражен
ных животных была выше, чем в контроле.
ные изменения были выявлены в гиппокампе
В настоящем исследовании крысы, которым
через 25 суток после стресса [36]. Поэтому этот
вводили эндотоксин в течение третьей недели
срок был использован в данной работе для оцен
жизни, имели более низкий уровень кортико
ки особенностей животных, которым неона
стерона как в обычных условиях, так и через
тально вводили ЛПС. В данной работе, в отли
25 дней после психогенной травмы. Самые низ
чие от ранее проведённых исследований, мы не
кие концентрации кортикостерона отмечены у
обнаружили статистически значимого влияния
крыс с введением ЛПС, перенёсших стресс.
фактора стресса на изучаемые показатели. Воз
В ещё одном независимом исследовании, про
можно, это связано с индивидуальными особен
ведённом нами ранее [9], также был выявлен
ностями животных. Данный феномен описан в
низкий уровень кортикостерона у нестрессиро
клинических и экспериментальных работах:
ванных крыс, которым в течение третьей недели
ПТСР развивается только у части стрессирован
жизни трижды вводили ЛПС в дозировках 25
ных людей и животных [56, 58]. Тем не менее в
либо 50 мкг/кг. Возможно, различия в получен
данной работе крысы, которым вводили ЛПС,
ных результатах связаны с различными дозиров
показали бóльшую стресс реактивность по из
ками эндотоксина и различными сроками его
менению уровня белка GluN2b. Нами не выяв
введения. Нужно также отметить, что снижение
лено изменений продукции белка EAAT2 в гип
уровня глюкокортикоидов в крови и нарушение
покампе крыс через 25 дней после психогенной
механизмов отрицательной обратной связи ха
травмы. Ранее было показано снижение
рактерно для посттравматических стрессовых
экспрессии данного белка в гиппокампе хрони
расстройств [61, 62], моделью которых предпо
чески стрессированных мышей [59]. Очевидно,
ложительно является использованное нами
использованный нами стресс был недостаточно
стрессорное воздействие.
сильным для изменения этого показателя.
В целом, проведённое нами исследование по
Основной результат проведённого нами ис
казывает, что воздействие бактериальным ЛПС в
следовании заключается в выявлении сочетан
раннем постнатальном онтогенезе влияет на ха
ных влияний неонатальных введений ЛПС и
рактер стресс индуцированных изменений пове
психогенной травмы на экспрессию генов
дения и на особенности экспрессии генов субъ
GluN2b субъединицы NMDA рецепторов и
единиц ионотропных рецепторов глутамата в
GluA1 субъединицы AMPA рецепторов, а также
гиппокампе после психогенной травмы, перене
на показатели исследовательского и эмоцио
сённой во взрослом возрасте. На возможную взаи
нального поведения. В отношении экспрессии
мосвязь этих нарушений указывают, в частности,
генов субъединиц ионотропных рецепторов глу
результаты Lei et al. [43], показавшие изменение
11 БИОХИМИЯ том 86 вып. 6 2021
914
НИКИТИНА и др.
активности животных в «Открытом поле» после
ких, биохимических и молекулярно биологи
введений антагониста GluN2b содержащих ре
ческих исследований ИЭФБ им. И.М. Сечено
цепторов рисленемдаза. Окончательный вывод о
ва РАН.
связи выявленных нами нейрохимических и по
Конфликт интересов. Авторы заявляют об от
веденческих изменений может быть сделан толь
сутствии конфликта интересов.
ко после проведения дополнительных нейрофар
Соблюдение этических норм. Все примени
макологических исследований.
мые международные, национальные и/или инс
титуциональные принципы ухода и использова
ния животных были соблюдены. Настоящая
Финансирование. Работа выполнена при фи
статья не содержит описания каких либо иссле
нансовой поддержке Российского фонда фунда
дований с участием людей в качестве объектов.
ментальных исследований (грант № 17 04
Дополнительные материалы. Приложение к
02116).
статье на английском языке опубликовано на
Благодарности. Спектрофотометрические
сайте журнала «Biochemistry» (Moscow) (http://
исследования и визуализация мембран выпол
protein.bio.msu.ru/biokhimiya) и на сайте изда
нены на базе Центра коллективного пользова
ния научного оборудования для физиологичес
10541), том 86, вып. 6, 2021.
СПИСОК ЛИТЕРАТУРЫ
1.
Lee, R. S., Oswald, L. M., and Wand, G. S. (2018) Early
neonatal administration of bacterial lipopolysaccharide,
life stress as a predictor of co occurring alcohol use disor
Neurosci. Behav. Physiol., 50, 1239 1248, doi: 10.1007/
der and post traumatic stress disorder, Alcohol. Res., 39,
s11055 020 01025 7.
147 159.
11.
Zubareva, O. E., Postnikova, T. Y., Grifluk, A. V., Schwarz,
2.
Fisher, P. A., Beauchamp, K. G., Roos, L. E., Noll, L. K.,
A. P., Smolensky, I. V., et al. (2020) Exposure to bacterial
Flannery, J., and Delker, B. C. (2016) The neurobiology of
lipopolysaccharide in early life affects the expression of
intervention and prevention in early adversity, Annu. Rev.
ionotropic glutamate receptor genes and is accompanied
Clin. Psychol., 12, 331 357, doi: 10.1146/annurev clinpsy
by disturbances in long term potentiation and cognitive
032814 112855.
functions in young rats, Brain. Behav. Immun., 90, 3 15,
3.
Van Camp, G., Cigalotti, J., Bouwalerh, H., Mairesse, J.,
doi: 10.1016/j.bbi.2020.07.034.
Gatta, E., et al. (2018) Consequences of a double hit of
12.
Walker, A. K., Nakamura, T., Byrne, R. J., Naicker, S.,
stress during the perinatal period and midlife in female rats:
Tynan, R. J., et al. (2009) Neonatal lipopolysaccharide and
mismatch or cumulative effect? Psychoneuroendocrinology,
adult stress exposure predisposes rats to anxiety like behav
93, 45 55, doi: 10.1016/j.psyneuen.2018.04.004.
iour and blunted corticosterone responses: implications for
4.
Jaric I., Rocks, D., Cham, H., Herchek, A., and
the double hit hypothesis, Psychoneuroendocrinology, 34,
Kundakovic, M. (2019) Sex and estrous cycle effects on
1515 1525, doi: 10.1016/j.psyneuen.2009.05.010.
anxiety and depression related phenotypes in a two hit
13.
Walker, A. K., Nakamura, T., and Hodgson, D. M. (2010)
developmental stress model, Front. Mol. Neurosci., 12, 74,
Neonatal lipopolysaccharide exposure alters central
doi: 10.3389/fnmol.2019.00074.
cytokine responses to stress in adulthood in Wistar rats,
5.
Koss, K. J., and Gunnar, M. R. (2018) Annual research
Stress, 13, 506 515, doi: 10.3109/10253890.2010.489977.
review: early adversity, the hypothalamic pituitary adreno
14.
Harré, E. M., Galic, M. A., Mouihate, A.,
cortical axis, and child psychopathology, J. Child Psychol.
Noorbakhsh, F., and Pittman, Q. J. (2008) Neonatal
Psychiatry, 59, 327 346, doi: 10.1111/jcpp.12784.
inflammation produces selective behavioural deficits and
6.
Dinel, A. L., Joffre, C., Trifilieff, P., Aubert, A., Foury, A.,
alters N methyl D aspartate receptor subunit mRNA in
Le Ruyet, P., and Layé, S. (2014) Inflammation early in life
the adult rat brain, Eur. J. Neurosci., 27, 644653,
is a vulnerability factor for emotional behavior at adolescence
doi: 10.1111/j.1460 9568.2008.06031.x.
and for lipopolysaccharide induced spatial memory and neu
15.
Hansen, K. B., Yi, F., Perszyk, R. E., Furukawa, H.,
rogenesis alteration at adulthood, J. Neuroinflammation, 11,
Wollmuth, L. P., et al. (2018) Structure, function, and
155, doi: 10.1186/s12974 014 0155 x.
allosteric modulation of NMDA receptors, J. Gen. Physiol.,
7.
Doenni, V. M., Song, C. M., Hill, M. N., and Pittman,
150, 1081 1105, doi: 10.1085/jgp.201812032.
Q. J. (2017) Early life inflammation with LPS delays fear
16.
Liu, S., Lau, L., Wei, J., Zhu, D., Zou, S., et al. (2004)
extinction in adult rodents, Brain. Behav. Immun., 63, 176
Expression of Ca2+ permeable AMPA receptor channels
185, doi: 10.1016/j.bbi.2016.11.022.
primes cell death in transient forebrain ischemia, Neuron,
8.
Lei, Y., Chen, C. J., Yan, X. X., Li, Z., and Deng, X. H.
43, 43 55, doi: 10.1016/j.neuron.2004.06.017.
(2017) Early life lipopolysaccharide exposure potentiates
17.
Wenzel, A., Fritschy, J. M., Mohler, H., and Benke, D.
forebrain expression of NLRP3 inflammasome proteins
(1997) NMDA receptor heterogeneity during postnatal
and anxiety like behavior in adolescent rats, Brain Res.,
development of the rat brain: differential expression of the
1671, 43 54, doi: 10.1016/j.brainres.2017.06.014.
NR2A, NR2B, and NR2C subunit proteins,
9.
Trofimov, A., Strekalova, T., Mortimer, N., Zubareva, O.,
J. Neurochem.,
68,
469478, doi:
10.1046/j.1471
Schwarz, A., et al. (2017) Postnatal LPS challenge impacts
4159.1997.68020469.x.
escape learning and expression of plasticity factors Mmp9
18.
Babb, T. L., Mikuni, N., Najm, I., Wylie, C., Olive, M.,
and Timp1 in rats: effects of repeated training, Neurotox.
et al. (2005) Pre and postnatal expressions of NMDA
Res., 32, 175 186, doi: 10.1007/s12640 017 9720 2.
receptors 1 and 2B subunit proteins in the normal rat cor
10.
Trofimov, A. N., Rotov, A. Y., Veniaminova, E. A.,
tex, Epilepsy Res., 64, 23 30, doi: 10.1016/j.eplepsyres.
Fomalont, K., Schwarz, A. P., and Zubareva, O. E. (2020)
2005.02.008.
Changes in behavior and the expression of ionotropic glu
19.
Du Bois, T. M., and Huang, X. F. (2007) Early brain devel
tamate receptor genes in the brains of adult rats after
opment disruption from NMDA receptor hypofunction:
БИОХИМИЯ том 86 вып. 6 2021
НЕОНАТАЛЬНОЕ ВОСПАЛЕНИЕ И СТРЕСС РЕАКТИВНОСТЬ
915
relevance to schizophrenia, Brain Res. Rev., 53, 260 270,
34.
Zink, M., Vollmayr, B., Gebicke Haerter, P. J., and Henn,
doi: 10.1016/j.brainresrev.2006.09.001.
F. A. (2010) Reduced expression of glutamate transporters
20.
Lippman Bell, J. J., Zhou, C., Sun, H., Feske, J. S., and
vGluT1, EAAT2 and EAAT4 in learned helpless rats, an
Jensen, F. E. (2016) Early life seizures alter synaptic calci
animal model of depression, Neuropharmacology, 58, 465
um permeable AMPA receptor function and plasticity,
473, doi: 10.1016/j.neuropharm.2009.09.005.
Mol. Cell. Neurosci., 76, 11 20, doi: 10.1016/j.mcn.2016.
35.
Zhang, X. H., Jia, N., Zhao, X. Y., Tang, G. K.,
08.002.
Guan, L. X., et al. (2013) Involvement of pGluR1, EAAT2
21.
Yuan, T., and Bellone, C. (2013) Glutamatergic receptors
and EAAT3 in offspring depression induced by prenatal
at developing synapses: the role of GluN3A containing
stress, Neuroscience,
250,
333341, doi:
10.1016/
NMDA receptors and GluA2 lacking AMPA receptors,
j.neuroscience.2013.04.031.
Eur. J. Pharmacol., 719, 107 111, doi: 10.1016/j.ejphar.
36.
Kovalenko, A. A., Zakharova, M. V., Nikitina, V. A.,
2013.04.056.
Schwarz, A. P., Karyakin, V. B., et al. (2018) Alterations in
22.
Sweatt, J. D. (2016) Neural plasticity and behavior - sixty
the expression of genes that encode subunits of ionotropic
years of conceptual advances, J. Neurochem., 139 Suppl,
glutamate receptors and the glutamate transporter in brain
179 199, doi: 10.1111/jnc.13580.
structures of rats after psychogenic stress, Neurochem. J.,
23.
Diering, G. H., and Huganir, R. L. (2018) The AMPA
12, 135 141, doi: 10.1134/S181971241802006X.
receptor code of synaptic plasticity, Neuron, 100, 314 329,
37.
Nasca, C., Bigio, B., Zelli, D., de Angelis, P., Lau, T., et al.
doi: 10.1016/j.neuron.2018.10.018.
(2017) Role of the Astroglial glutamate exchanger xCT in
24.
Lisman, J. (2017) Glutamatergic synapses are structurally
ventral hippocampus in resilience to stress, Neuron, 96,
and biochemically complex because of multiple plasticity
402 413.e5, doi: 10.1016/j.neuron.2017.09.020.
processes: long term potentiation, long term depression,
38.
Beznin, G. V., Pshenichnaya, A. G., Kusov, A. G., and
short term potentiation and scaling, Philos. Trans. R. Soc.
Tsikunov, S. G. (2012) Morphological and functional bases
Lond. B Biol. Sci., 372, doi: 10.1098/rstb.2016.0260.
of behavioral deviations in the model of acute vital stress in
25.
Sarantis, K., Antoniou, K., Matsokis, N., and
rats, Med. Akadem. Zhurn., 12, 37 39.
Angelatou, F. (2012) Exposure to novel environment is
39.
Paxinos, G., and Watson, C. (2007) The Rat Brain in
characterized by an interaction of D1/NMDA receptors
Stereotaxic Coordinates, 6th Edition, Academic Press.
underlined by phosphorylation of the NMDA and AMPA
40.
Kopec, A. M., Rivera, P. D., Lacagnina, M. J.,
receptor subunits and activation of ERK1/2 signaling,
Hanamsagar, R., and Bilbo, S. D. (2017) Optimized solu
leading to epigenetic changes and gene expression in rat
bilization of TRIzol precipitated protein permits Western
hippocampus, Neurochem. Int., 60, 55 67, doi: 10.1016/
blotting analysis to maximize data available from brain tis
j.neuint.2011.10.018.
sue, J. Neurosci. Methods, 280, 64 76, doi: 10.1016/
26.
Barkus, C., McHugh, S. B., Sprengel, R., Seeburg, P. H.,
j.jneumeth.2017.02.002.
Rawlins, J. N. P., and Bannerman, D. M.
(2010)
41.
Harrington, C. R. (1990) Lowry protein assay containing
Hippocampal NMDA receptors and anxiety: at the inter
sodium dodecyl sulfate in microtiter plates for protein
face between cognition and emotion, Eur. J. Pharmacol.,
determinations on fractions from brain tissue, Anal.
626, 49 56, doi: 10.1016/j.ejphar.2009.10.014.
Biochem., 186, 285 7, doi: 10.1016/0003 2697(90)90081 j.
27.
Pacheco, A., Aguayo, F. I., Aliaga, E., Muñoz, M., Garc a
42.
Laemmli, U. K. (1970) Cleavage of structural proteins dur
Rojo, G., et al. (2017) Chronic stress triggers expression of
ing the assembly of the head of bacteriophage T4, Nature,
immediate early genes and differentially affects the expres
227, 680 685, doi: 10.1038/227680a0.
sion of AMPA and NMDA subunits in dorsal and ventral
43.
Lei, T., Dong, D., Song, M., Sun, Y., Liu, X., and
hippocampus of rats, Front. Mol. Neurosci., 10, 244,
Zhao, H. (2020) Rislenemdaz treatment in the lateral
doi: 10.3389/fnmol.2017.00244.
habenula improves despair like behavior in mice,
28.
Costa Nunes, J., Zubareva, O., Araújo Correia, M.,
Neuropsychopharmacology, 45, 1717 1724, doi: 10.1038/
Valença, A., Schroeter, C. A., et al. (2014) Altered emo
s41386 020 0652 9.
tionality, hippocampus dependent performance and
44.
Delawary, M., Tezuka, T., Kiyama, Y., Yokoyama, K.,
expression of NMDA receptor subunit mRNAs in chroni
Inoue, T., et al. (2010) NMDAR2B tyrosine phosphoryla
cally stressed mice, Stress, 17, 108 116, doi: 10.3109/
tion regulates anxiety like behavior and CRF expression in
10253890.2013.872619.
the amygdala, Mol. Brain, 3, 37, doi: 10.1186/1756 6606
29.
Furini, C., Myskiw, J., and Izquierdo, I. (2014) The learn
3 37.
ing of fear extinction, Neurosci. Biobehav. Rev., 47, 670
45.
Rahman, T., Zavitsanou, K., Purves Tyson, T., Harms,
683, doi: 10.1016/j.neubiorev.2014.10.016.
L. R., Meehan, C., et al. (2017) Effects of immune activa
30.
Matsumoto, Y., Morinobu, S., Yamamoto, S.,
tion during early or late gestation on N methyl d aspartate
Matsumoto, T., Takei, S., et al. (2013) Vorinostat amelio
receptor measures in adult rat offspring, Front. Psychiatry,
rates impaired fear extinction possibly via the hippocampal
8, 77, doi: 10.3389/fpsyt.2017.00077.
NMDA CaMKII pathway in an animal model of posttrau
46.
Tishkina, A., Stepanichev, M., Kudryashova, I.,
matic stress disorder, Psychopharmacology (Berlin), 229,
Freiman, S., Onufriev, M., et al. (2016) Neonatal proin
51 62, doi: 10.1007/s00213 013 3078 9.
flammatory challenge in male Wistar rats: effects on behav
31.
Réus, G. Z., Abelaira, H. M., Stringari, R. B., Fries, G. R.,
ior, synaptic plasticity, and adrenocortical stress response,
Kapczinski, F., and Quevedo, J. (2012) Memantine treat
Behav. Brain Res., 304, 1 10, doi: 10.1016/j.bbr.2016.02.001.
ment reverses anhedonia, normalizes corticosterone levels
47.
Onufriev, M. V., Freiman, S. V., Peregud, D. I.,
and increases BDNF levels in the prefrontal cortex induced
Kudryashova, I. V, Tishkina, A. O., et al. (2017) Neonatal
by chronic mild stress in rats, Metab. Brain Dis., 27, 175
proinflammatory stress induces accumulation of corticos
182, doi: 10.1007/s11011 012 9281 2.
terone and interleukin 6 in the hippocampus of juvenile
32.
Padovan, C. M., and Guimarães, F. S.
(2004)
rats: potential mechanism of synaptic plasticity impair
Antidepressant like effects of NMDA receptor antagonist
ments, Biochemistry (Moscow), 82, 275 281, doi: 10.1134/
injected into the dorsal hippocampus of rats, Pharmacol.
S0006297917030051.
Biochem. Behav., 77, 1519, doi: 10.1016/j.pbb.2003.
48.
Postnikova, T. Y., Griflyuk, A. V, Ergina, J. L., Zubareva,
09.015.
O. E., and Zaitsev, A. V. (2020) Administration of bacteri
33.
Blacker, C. J., Millischer, V., Webb, L. M., Ho, A. M. C.,
al lipopolysaccharide during early postnatal ontogenesis
Schalling, M., et al. (2020) EAAT2 as a research target in
induces transient impairment of long term synaptic plas
bipolar disorder and unipolar depression: a systematic
ticity associated with behavioral abnormalities in young
review, Mol. Neuropsychiatry, 5, 44 59, doi: 10.1159/
rats, Pharmaceuticals (Basel),
13, doi:
10.3390/
000501885.
ph13030048.
БИОХИМИЯ том 86 вып. 6 2021
11*
916
НИКИТИНА и др.
49.
Onufriev, M. V., Uzakov, S. S., Freiman, S. V.,
lipopolysaccharide, Eur. J. Neurosci., 27, 14751484,
Stepanichev, M. Y., Moiseeva, Y. V., et al. (2018) The
doi: 10.1111/j.1460 9568.2008.06121.x.
Dorsal and ventral hippocampus have different reactiv
56. Zoladz, P. R., and Diamond, D. M. (2016) Predator based
ities to proinflammatory stress: corticosterone levels,
psychosocial stress animal model of PTSD: preclinical
cytokine expression, and synaptic plasticity, Neurosci.
assessment of traumatic stress at cognitive, hormonal,
Behav. Physiol., 48, 1024 1031, doi: 10.1007/s11055 018
pharmacological, cardiovascular and epigenetic levels of
0665 6.
analysis, Exp. Neurol., 284, 211 219, doi: 10.1016/j.expneurol.
50.
Benmhammed, H., El Hayek, S., Nassiri, A.,
2016.06.003.
Bousalham, R., Mesfioui, A., et al. (2019) Effects of
57. Zoladz, P. R., Park, C. R., Fleshner, M., and Diamond,
lipopolysaccharide administration and maternal depriva
D. M. (2015) Psychosocial predator based animal model
tion on anxiety and depressive symptoms in male and
of PTSD produces physiological and behavioral sequelae
female Wistar rats: Neurobehavioral and biochemical
and a traumatic memory four months following stress
assessments, Behav. Brain Res., 362, 46 55, doi: 10.1016/
onset, Physiol. Behav., 147, 183 192, doi: 10.1016/j.physbeh.
j.bbr.2019.01.005.
2015.04.032.
51.
Sulakhiya, K., Keshavlal, G. P., Bezbaruah, B. B.,
58. Auxéméry, Y. (2012) Posttraumatic stress disorder (PTSD)
Dwivedi, S., Gurjar, S. S., et al. (2016) Lipopolysaccharide
as a consequence of the interaction between an individual
induced anxiety and depressive like behaviour in mice are
genetic susceptibility, a traumatogenic event and a social
prevented by chronic pre treatment of esculetin, Neurosci.
context, Encephale, 38, 373 380, doi: 10.1016/j.encep.
Lett., 611, 106 111, doi: 10.1016/j.neulet.2015.11.031.
2011.12.003.
52.
Walker, F. R., March, J., and Hodgson, D. M. (2004)
59. Ding, X. F., Li, Y. H., Chen, J. X., Sun, L. J., Jiao, H. Y.,
Endotoxin exposure in early life alters the development of
et al. (2017) Involvement of the glutamate/glutamine cycle
anxiety like behaviour in the Fischer 344 rat, Behav. Brain
and glutamate transporter GLT 1 in antidepressant like
Res., 154, 63 69, doi: 10.1016/j.bbr.2004.01.019.
effects of Xiao Yao san on chronically stressed mice, BMC
53.
Spencer, S. J., Heida, J. G., and Pittman, Q. J. (2005)
Complement. Altern. Med., 17, 326, doi: 10.1186/s12906
Early life immune challenge - effects on behavioural
017 1830 0.
indices of adult rat fear and anxiety, Behav. Brain Res., 164,
60. Shanks, N., Larocque, S., and Meaney, M. J. (1995)
231 238, doi: 10.1016/j.bbr.2005.06.032.
Neonatal endotoxin exposure alters the development of the
54.
Custódio, C. S., Mello, B. S. F., Filho, A. J. M. C., de
hypothalamic pituitary adrenal axis: early illness and later
Carvalho Lima, C. N., Cordeiro, R. C., et al. (2018)
responsivity to stress, J. Neurosci., 15, 376 384.
Neonatal immune challenge with lipopolysaccharide trig
61. Zoladz, P. R., Fleshner, M., and Diamond, D. M. (2012)
gers long lasting sex and age related behavioral and
Psychosocial animal model of PTSD produces a long lasting
immune/neurotrophic alterations in mice: relevance to
traumatic memory, an increase in general anxiety and PTSD
autism spectrum disorders, Mol. Neurobiol., 55,
3775
like glucocorticoid abnormalities, Psychoneuroendocrinology,
3788, doi: 10.1007/s12035 017 0616 1.
37, 1531 1545, doi: 10.1016/j.psyneuen.2012.02.007.
55.
Fan, L. W., Tien, L. T., Mitchell, H. J., Rhodes, P. G., and
62. Olff, M., Güzelcan, Y., de Vries, G. J., Assies, J., and Gersons,
Cai, Z. (2008) Alpha phenyl n tert butyl nitrone amelio
B. P. R. (2006) HPA and HPT axis alterations in chronic
rates hippocampal injury and improves learning and mem
posttraumatic stress disorder, Psychoneuroendocrinology,
ory in juvenile rats following neonatal exposure to
31, 1220 1230, doi: 10.1016/j.psyneuen.2006.09.003.
NEONATAL EXPOSURE TO BACTERIAL LIPOPOLYSACCHARIDE ALTERS
BEHAVIOR AND HIPPOCAMPAL IONOTROPIC GLUTAMATE RECEPTOR
EXPRESSION IN THE ADULT RATS AFTER PSYCHOGENIC TRAUMA
V. A. Nikitina1, M. V. Zakharova2, A. N. Trofimov1, A. P. Schwarz2,
G. V. Beznin1, S. G. Tsikunov1, and O. E. Zubareva2*
1 Institute of Experimental Medicine, 197376 Saint Petersburg, Russia
2 Sechenov Institute of Evolutionary Physiology and Biochemistry, Russian Academy of Sciences,
194223 Saint Petersburg, Russia; E mail: zubarevaoe@mail.ru
According to the “two hit hypothesis” of formation of psychoneuropathology, infectious diseases and other patho
logical conditions during critical periods of early ontogenesis disrupt the normal development of the brain, increasing
its vulnerability to stresses experienced in the adolescence and adulthood. It is assumed that these disorders may be
associated with changes in the functional activity of the hippocampal glutamatergic system. In the present work, we
studied the protein level of gene expression of NMDA (GluN1, GluN2a, GluN2b) and AMPA (GluA1, GluA2) glu
tamate receptor subunits, as well as glutamate transporter EAAT2 in the ventral and dorsal hippocampus of rats which
were injected with LPS during the 3rd week of the life and experienced a vital stress (a contact with a predator, a
python) in the adulthood. The testing was performed 25 days after the stress. Abnormalities were found in the ventral,
but not in the dorsal hippocampus. Non stressed LPS treated rats had a lower level of GluN2b protein in the ventral
hippocampus as compared to the control animals, while after the stress the level of GluN2b was significantly increased
in the LPS treated, but not in the control animals. Different responses to stress in the experimental and control rats
were also revealed in terms of the exploratory behavior: when compared with non stressed animals, the stressed con
trol rats demonstrated a higher locomotor activity in the 1st min of the “Open Field” test, while this parameter was
lower in the stressed experimental rats. In addition, the stressed and non stressed rats which were injected with LPS
in the early age spent a longer time in the open arms when tested in the “Elevated Plus Maze” and had a reduced blood
level of corticosterone. To summarize, our study shows that exposure to bacterial LPS in the early postnatal ontoge
nesis affects the pattern of stress induced changes in the behavior and expression of genes encoding subunits of
ionotropic glutamate receptors in the hippocampus after a psychogenic trauma suffered in the adulthood.
Keywords: lipopolysaccharide, early development, brain, stress reactivity, NMDA receptor, AMPA receptor, behavior
БИОХИМИЯ том 86 вып. 6 2021