БИОХИМИЯ, 2021, том 86, вып. 9, с. 1254 - 1264
УДК 577.1
ПУТЕШЕСТВИЕ ИНФОРМОСОМ ВО ВРЕМЕНИ:
РАННЯЯ КОНЦЕПЦИЯ мРНК В КОНТЕКСТЕ
СОВРЕМЕННЫХ ПРЕДСТАВЛЕНИЙ ОБ мРНП
Обзор
© 2021
П.В. Иванов1,2,3
1 Division of Rheumatology, Inflammation, and Immunity, Brigham and Women’s Hospital,
Boston, Massachusetts, 02115 USA; e mail: pivanov@rics.bwh.harvard.edu
2 Department of Medicine, Harvard Medical School, Boston, Massachusetts, 02115 USA
3 Harvard Medical School Initiative for RNA Medicine, Harvard University, Boston, Massachusetts, 02115 USA
Поступила в редакцию 27.05.2021
После доработки 09.07.2021
Принята к публикации 12.07.2021
На всём протяжении своего жизненного цикла матричная РНК (мРНК) существует в составе комплексов с
белками. Впервые мРНК"содержащие частицы нерибосомной природы, называемые информосомами, бы"
ли обнаружены в цитоплазматическом экстракте эмбрионов рыб сотрудниками лаборатории Александра
Спирина, а позднее эти частицы были описаны и в живых клетках. Со временем были открыты и охаракте"
ризованы и другие цитоплазматические, а также ядерные, мРНК"содержащие рибонуклеопротеины
(мРНП). Несмотря на различия во внутриклеточной локализации, структуре и функциях, эти мРНП тем не
менее обладают многими общими чертами, характерными также для информосом. В настоящем мини"
обзоре рассматривается история открытия информосом, их характеристики и предполагаемые функции, а
также их возможное родство с другими мРНП.
КЛЮЧЕВЫЕ СЛОВА: информосомы, мРНК, рибонуклеопротеины, мРНП, РНК"гранулы, биосинтез
белка.
DOI: 10.31857/S0320972521090025
ВВЕДЕНИЕ: ТИПИЧНЫЙ
жённой нетранслируемыми участками (UTRs,
ЖИЗНЕННЫЙ ЦИКЛ мРНК
untranslated regions) - 5′" и 3′"UTRs соответ"
ственно. В свою очередь, UTRs имеют характер"
Эукариотические информационные (мат"
ные признаки, такие как присутствие 7"метил"
ричные) РНК (мРНК) транскрибируются в ядре
гуанозина на 5′"конце мРНК (кэп) и последова"
с помощью РНК"полимеразы II. В процессе
тельность из остатков аденина (A) на 3′"конце,
транскрипции эти насцентные (растущие) тран"
образующая поли(A)"хвост. Созревание молеку"
скрипты окружаются набором разнообразных
лы мРНК происходит в результате скоордини"
РНК"связывающих белков (RBPs, RNA"binding
рованного действия множества ферментов и ас"
proteins), которые активно участвуют в превра"
социированных с ними ядерных белков, многие
щении молекул"предшественников мРНК (пре"
из которых являются РНК"связывающими бел"
мРНК) в молекулы зрелых мРНК. Созревание
ками [2]. Кроме того, другие RBPs, не участвую"
мРНК включает три основных этапа, а именно:
щие в созревании ядерной мРНК, распознают
сплайсинг РНК, кэпирование 5′"конца и от"
последовательности и/или структуры, входящие
щепление/полиаденилирование 3′"конца (см.
в UTRs (cis"элементы) и участвуют в ряде про"
обзор [1]). По завершении этих процессов стан"
цессов, определяющих судьбу мРНК [3]. В ре"
дартная молекула мРНК состоит из открытой
зультате формируются различные нуклеопроте"
рамки считывания (ORF, open reading frame),
иновые частицы на основе матричной РНК
кодирующей последовательность белка, окру"
(мРНП), отличающиеся по белковому составу и
общей структуре [4].
Принятые сокращения: мРНП - нуклеопротеино"
По завершении процесса формирования
вые частицы на основе матричной (информационной)
мРНП в зависимости от состава частицы, мРНП
РНК; PBs - процессирующие тельца (processing bodies);
RBPs - РНК"связывающие белки (RNA"binding proteins);
могут быстро экспортироваться из ядра в цито"
SGs - стрессовые гранулы (stress granules).
плазму, где могут быть сразу использованы для
1254
ИНФОРМОСОМЫ В МЕТАБОЛИЗМЕ мРНК
1255
трансляции мРНК, или же они могут оставаться
менённый к цитоплазматическим экстрактам из
в ядре как «молчащие» мРНК"содержащие час"
эмбрионов, преинкубированных с радиоактив"
тицы [5]. Некоторые мРНП транспортируются
но мечеными аминокислотами, показал очень
для трансляции в определенные участки клетки,
сходную картину седиментации. Принимая во
например, в синапсы нейронов для продукции
внимание тот факт, что в эмбрионах M. fossilis
нейромедиаторов. Эти процессы сопровожда"
новые рибосомы (или рибосомные РНК) не
ются активным ремоделингом мРНП путем свя"
синтезируются вплоть до завершения гаструля"
зывания с цитоплазматическими РНК"связыва"
ции [8], было высказано предположено, что но"
ющими белками [6]. Следует отметить, что в ак"
восинтезированные молекулы РНК, оседающие
тивно растущих клетках наблюдается более вы"
в пострибосомной зоне, представляют собой
сокий уровень трансляции в сравнении с неко"
молекулы мРНК в комплексах с белками [9].
торыми специализированными клетками (таки"
В продолжение этого исследования Спирин
ми как ооциты), в которых обычно происходит
с соавт. использовали метод, основанный на
накопление мРНП, которые позднее использу"
фиксации РНП в формальдегиде, который спо"
ются для трансляции мРНК только на специфи"
собствует эффективной перекрестной сшивке
ческих стадиях развития этих клеток. Как актив"
РНК"белковых комплексов [10]. Благодаря та"
но транслируемые, так и нетранслируемые
кой стабилизации стало возможным проанали"
мРНК в итоге подвергаются процессу деграда"
зировать распределение перекрестно"сшитых
ции, который завершает их жизненный цикл [7].
компонентов в градиенте плотности CsCl [11].
Общей особенностью метаболизма всех мРНК
Было очевидно, что эти фиксированные фор"
является невозможность их существования без
мальдегидом пострибосомные комплексы со"
взаимодействия с белковыми партнерами.
держат радиоактивно меченые аминокислоты и
РНК, седиментирующие в один регион плот"
ности. Исследуемые РНП также определенно
ОТКРЫТИЕ ИНФОРМОСОМ
отличались от рибосом/их субъединиц и сво"
бодной РНК, т.к. обладали низким значением
Несмотря на то что в настоящее время обще"
плавучей плотности ~1,40-1,45 г/см3, находя"
известно, что молекулы мРНК не могут сущест"
щимся между значениями, характерными для
вовать в «обнаженном» виде, и на всех этапах
свободного белка и РНК [12]. Кроме того, зна"
своего жизненного цикла они взаимодействуют
чения плавучей плотности указывали на то, что
с белками, образуя различные мРНП, в начале
в этих комплексах белок доминирует над РНК в
1960"х гг. предположение, что новосинтезиро"
соотношении ~3 : 1. В пострибосомной зоне
ванные молекулы мРНК находятся в комплексе
было обнаружено семь таких РНК"белковых
с белками и могут существовать в свободном от
комплексов с коэффициентами седиментации,
рибосом состоянии, было довольно провокаци"
равными 20S, 30S, 40S, 50S, 55S, 65S и 75S, что
онным. Эта идея казалась тогда не имеющей
указывало на воспроизводимость полученного
смысла, т.к. основной функцией мРНК счита"
результата [12]. Кроме того, эти комплексы мог"
лось их участие в синтезе белка в ассоциирован"
ли быть количественно абсорбированы на нит"
ном с рибосомами состоянии.
роцеллюлозных мембранах, что было невоз"
В 1964 г. Спирин и его коллеги предположи"
можно для свободной РНК или белков. Изучае"
ли, что матричные РНК эукариотических кле"
мые комплексы также подвергались деградации
ток, находящиеся в цитоплазме и временно не
проназой (неспецифическая протеаза из Strepto
подвергающиеся трансляции, существуют в
myces griseus) и рибонуклеазой, что исключало
комплексе с неизвестными белками, не имею"
возможность существования ДНК в их составе
щими отношения к рибосомам [8, 9]. Такие вы"
[12]. Более того, высвобожденные в результате
воды были основаны на результатах исследова"
депротеинизации и центрифугирования в гра"
ния эмбрионов Misgurnus fossilis (вьюн обыкно"
диенте плотности сахарозы радиоактивно мече"
венный) на поздних стадиях развития бластулы,
ные РНК"компоненты пострибосомных РНП
которые подвергались предварительной инкуба"
имели коэффициенты седиментации, значи"
ции с [14C]"аденином или [14C]"уридином, спо"
тельно отличающиеся от значений 18S и 28S,
собными эффективно включаться в насцентные
характерных для рибосомной РНК [12]. Данный
транскрипты [9]. С помощью центрифугирова"
факт говорит о том, что молекулы РНК в
ния экстрактов цитоплазмы эмбрионов в гради"
этих РНП не имели отношения к рибосомной
енте сахарозы было обнаружено, что вновь син"
РНК [13].
тезированные транскрипты осаждаются после
Важно отметить, что схожие РНП были так"
80S рибосом (в промежутке между 20S и 75S). К
же обнаружены в эмбрионах морского ежа [14] и
удивлению исследователей, тот же подход, при"
в клетках HeLa, инфицированных вирусом ос"
БИОХИМИЯ том 86 вып. 9 2021
1256
ИВАНОВ
повакцины [15]. В эмбрионах морского ежа ра"
зоной - они также могут быть обнаружены и в
диоактивно меченые РНК, выделенные из пост"
прерибосомной зоне [12, 26]. При центрифуги"
рибосомных РНП, количественно гибридизова"
ровании в градиенте сахарозы цитоплазмати"
лись с ДНК, что указывало на их мРНК"подоб"
ческих экстрактов эмбрионов вьюна новосинте"
ную природу [14, 16]. В клетках HeLa, инфици"
зированные радиоактивно меченые мРНК об"
рованных вирусом осповакцины, в которых вы"
наруживались еще до пика, характерного для
соко экспрессируется только один транскрипт, а
80S рибосом, а именно в составе РНП с коэф"
именно вирусная мРНК, также обнаруживались
фициентами седиментации, равными 90S, 100S
пострибосомные РНП [17, 18], содержащие ново"
и 110S [12]. Эти мРНК не могли быть составны"
синтезированную вирусную мРНК [15].
ми частями полисом, т.к. даже димер 80S рибо"
В целом приведенные выше данные позво"
сомы имел бы коэффициент седиментации, рав"
лили предположить, что исследуемые пострибо"
ный ~120S. Принимая во внимание тот факт,
сомные рибонуклеопротеины с характерным
что плавучая плотность прерибосомных 90S и
низким значением плавучей плотности состоят
110S компонентов схожа с плавучей плотностью
из мРНК и белков, и при этом не содержат ни
пострибосомных
50S-75S компонентов
целых рибосом, ни их субъединиц. Эти мРНП
(~1,39 г/см3), был сделан вывод, что пострибо"
получили название информосомы, отражающее
сомные мРНП в действительности являются
тот факт, что РНК"составляющая этих частиц
информосомами [12]. Таким образом, стало воз"
несет информацию (причём имеется в виду не
можным выделить две крупные субпопуляции
только матрица, кодирующая последователь"
информосом: пострибосомные (20S-75S) и пре"
ность белка), а также то, что белковый компо"
рибосомные (90S-110S). Поскольку обнару"
нент частицы принимает активное участие в оп"
женные информосомы различались по размеру,
ределении судьбы мРНК.
было логично предположить, что они также от"
личаются друг от друга по составу белков и
мРНК [12].
РАЗВИТИЕ КОНЦЕПЦИИ
Один из аспектов исследований, не удосто"
ИНФОРМОСОМЫ
енный достаточным вниманием, заключается в
существовании информосом, обладающих вы"
Вскоре после открытия информосом они
соким коэффициентом седиментации, хоть и не
были обнаружены и в ряде других эксперимен"
таких многочисленных, как описанные ранее
тальных моделей, таких как неинфицированные
прерибосомные информосомы. Возможно, они
клетки HeLa [19], пролиферирующие клетки
представляют собой отдельные крупные части"
эпидермиса гигантского шелкопряда [20], клет"
цы или же агрегацию меньших по размеру ин"
ки печени крыс [21], культивированные фиб"
формосом, ведущую к формированию более
робласты мыши (L"клетки) [22], L"клетки, ин"
крупного мРНП [13]. Биологическое значение
фицированные вирусом Менго [23], а также
таких высокомолекулярных частиц пока остаёт"
клетки асцитной опухоли Эрлиха, инфициро"
ся под вопросом, однако их функция может
ванные вирусом Сендай [24]. Все обнаруженные
быть связана с процессами сборки РНК"содер"
информосомы отличались друг от друга по раз"
жащих гранул (см. ниже).
меру, однако сохраняли ключевые характерис"
После обнаружения информосом в цито"
тики пострибосомных информосом. Интерес"
плазме было высказано предположение о воз"
но, что в двух ранних работах по анализу мРНП
можном существовании их аналогов в клеточ"
из пострибосомной зоны в нормальных клетках
ном ядре. И действительно, несколькими иссле"
HeLa [19] и клетках печени крысы [21] было
довательскими группами было продемонстри"
продемонстрировано образование 45S частиц,
ровано существование мРНП, аналогичных ци"
которые могли быть как информосомами, так и
топлазматическим информосомам, в ядерных
комплексами мРНК с 40S субъединицами рибо"
экстрактах [27-32]. Ядерные мРНП отличаются
сомы [21, 25]. Важно отметить, что анализ ин"
по размеру, однако имеют сходные с цитоплаз"
фицированных вирусом клеток позволил иден"
матическими коэффициенты плавучей плот"
тифицировать специфические типы информо"
ности (~1,4 г/см3) [30, 31]. Белковый состав
сом, целиком ассоциированных с вирусной
ядерных мРНП отличается от цитоплазматичес"
мРНК. Такие вирус"специфичные 45S инфор"
кого, что свидетельствует в пользу того, что во
мосомы были выделены и охарактеризованы
время экспорта из ядра молекулы мРНК могут
[23, 24].
менять своих белковых партнеров.
Еще одно важное наблюдение заключается в
И все же оставался один важный вопрос, су"
том, что при седиментации расположение ин"
ществуют ли на самом деле информосомы in vivo
формосом не ограничивается пострибосомной
или же их формирование является лишь арте"
БИОХИМИЯ том 86 вып. 9 2021
ИНФОРМОСОМЫ В МЕТАБОЛИЗМЕ мРНК
1257
фактом, сопровождающим разрушение и/или
Способность различных экзогенных РНК
гомогенизацию клеток. Вначале было обнару"
индуцировать образование информосомопо"
жено, что добавление свободных, не связанных
добных частиц подняла другие важные вопросы.
с белками, молекул РНК в цитоплазматические
Что такое «фактор загрузки»? Это один белок
экстракты, полученные из клеток различных
или группа белков, которые кооперативно свя"
моделей, приводило к образованию искусствен"
зываются с РНК? Обладает ли «фактор загруз"
ных информосомоподобных частиц, о чём мож"
ки» какой"либо специфичностью связывания,
но было судить по их поведению в градиенте
зависимой от последовательности или структу"
плотности CsCl [33, 34]. Эти частицы были го"
ры РНК? Несмотря на то что ответы на часть
могенными, и их формирование не зависело от
этих вопросов до сих пор отсутствуют, в ходе по"
природы добавляемых свободных РНК (ни от
пыток идентифицировать/выделить «фактор
длины, ни от происхождения РНК, в частности,
загрузки» был получен важный результат, свиде"
в большинстве экспериментов использовали
тельствующий о различной стабильности есте"
рРНК из E. coli). Единственным ограничением
ственных и искусственных информосом. Фрак"
при формировании таких комплексов было ко"
ционирование этих комплексов в градиенте
личество добавляемой РНК - её должно было
плотности сахарозы в присутствии или в отсут"
быть немного, избыток молекул РНК не вклю"
ствие формальдегида (со сшивками или без них)
чался в состав РНП [34]. Данный факт свиде"
и последующий седиментационный анализ в
тельствовал о том, что в клеточных экстрактах
градиенте плотности CsCl однозначно продемон"
содержится титруемый и ограниченный по ко"
стрировали эти различия. В отличие от искус"
личеству «фактор загрузки», доступный для ас"
ственных информосомоподобных РНП, кото"
социации с добавляемой свободной РНК [34].
рые характеризовались нестабильностью и чьё
На основе результатов других экспериментов
образование было обратимо, процесс формиро"
[35, 36, 38] был сделан вывод, что этот «фактор
вания естественных информосом был необра"
загрузки» имеет белковую природу (является
тим и приводил к образованию стабильных во
белком или группой белков), большой молеку"
всех протестированных условиях частиц [36].
лярный вес, составляет примерно 0,3% от общей
Еще одна интересная находка заключалась в
растворимой фракции экстракта и легко вступа"
том, что добавление эндогенных изолирован"
ет в реакцию с введенной экзогенной РНК. Сог"
ных из эмбрионов вьюна РНК приводило к
ласно результатам этих экспериментов, всё ещё
формированию значительно более стабильных
оставалась вероятность того, что свободные эн"
информосомоподобных частиц по сравнению с
догенные мРНК взаимодействуют с высвобо"
добавлением экзогенных РНК [36].
дившимися «факторами загрузки» с образова"
Поиск «фактора загрузки» оказал важное
нием искусственных информосом в процессе
влияние на развитие биологии РНК, в частнос"
разрушения/гомогенизации клеток, что свиде"
ти в области биосинтеза белка и регуляции
тельствовало бы против существования естест"
трансляции. Анализ различных цитоплазмати"
венных информосом, формирующихся in vivo.
ческих информосом (например, «маскирован"
В целях поиска ответа на этот вопрос Спи"
ных» и свободных), выделенных из спящих или
рин и его коллеги провели несколько экспери"
активно делящихся клеток, показал, что в этих
ментов по гомогенизации эмбрионов вьюна в
мРНК"содержащих комплексах присутствуют
присутствии избытка свободной экзогенной
два основных белка и большое количество раз"
РНК [35]. Полагали, что если информосомы об"
личных минорных белков. В отличие от рибо"
разуются в результате ассоциации свободных
сомных частиц, эти комплексы демонстрирова"
эндогенных мРНК с «фактором загрузки» в про"
ли характерную устойчивость к удалению Mg2+
цессе гомогенизации, то следовало бы ожидать,
(например, при добавлении ЭДТА), который всё
что экзогенная РНК может выступать в качестве
же мог быть удален из препарата информосом
эффективного конкурента, снижая уровень
путём последовательного повышения концент"
формирования информосом. Тем не менее ре"
рации хелатирующей соли [37]. Основные белки
зультаты такого эксперимента по конкуренции
комплексов характеризовались значениями мо"
между РНК ясно свидетельствовали в пользу то"
лекулярного веса, равными 50-55 кДа (p50) и
го, что информосомы действительно существу"
70-80 кДа (p70), которые были определены по
ют в живых клетках - до их разрушения, гомоге"
их электрофоретической подвижности в денату"
низации и фракционирования, т.к. добавление
рирующих гелях [38-43]. Результаты проведен"
избытка экзогенной РНК не влияло на профили
ного позднее биохимического анализа позволи"
седиментации/распределения информосом и
ли предположить, что белок p70 обладает повы"
характерные для них значения плавучей плот"
шенным сродством к поли(A)"последователь"
ности [35].
ности; впоследствии он получил название по"
БИОХИМИЯ том 86 вып. 9 2021
1258
ИВАНОВ
ли(А)"связывающий белок (PABP, poly(A)"bind"
рыб и морского ежа, клетки которых значитель"
ing protein) [39, 43]. Белки p50 были позднее оха"
но отличаются от активно пролиферирующих
рактеризованы как ДНК"связывающие факто"
клеток, в которых биосинтез белка и его регуля"
ры транскрипции, стимулирующие синтез
ция находятся под сильным влиянием экзоген"
мРНК с промоторов Y"бокса, при этом Y"бокс"
ных стимулов, таких как доступность питатель"
связывающий белок 1 (YB"1 или YBX1) являлся
ных веществ и кислорода. Эмбриональное раз"
универсальным для всех комплексов и был оха"
витие, напротив, контролируется внутренними
рактеризован лучше других [44]. В состав этого
факторами, которые совместно именуют «часо"
белка входит большое число остатков пролина и
вым» механизмом. Одной из особенностей регу"
аланина, он демонстрирует аномальную в срав"
ляции трансляции в раннем эмбриогенезе явля"
нении с предсказанной электрофоретическую
ется то, что уникальные паттерны развития от"
подвижность (36 кДа), характеризуется высоким
ражены в пространственно"временной регуля"
значением изоэлектрической точки (pI > 9) и
ции трансляции мРНК [48, 49]. После тран"
универсальным сродством к различным гетеро"
скрипции новосинтезированные мРНК экспор"
генным последовательностям молекул мРНК
тируются из ядра в цитоплазму, где они «хранят"
[45]. Оба белка, YB"1 и PABP, широко распрост"
ся» в молчащей
(«маскированной») форме
ранены и обладают характеристиками, подходя"
вплоть до определенного времени на «часах раз"
щими для «фактора загрузки», такими как спо"
вития», когда в нужный момент они станут дос"
собность PABP связываться с поли(A)"хвостами
тупными для трансляции. Существует значи"
и способность белка YB"1 связываться с боль"
тельный временной разрыв между моментом их
шим количеством разнообразных последова"
экспорта в цитоплазму и действительным вовле"
тельностей и структур РНК.
чением в процесс биосинтеза белка. Например,
Следует отметить, что в процессе поиска
такой временной разрыв ярко продемонстриро"
«фактора загрузки» было также сделано не"
ван в ходе развития эмбриона вьюна, у которого
сколько других важных открытий. Так, было по"
транскрипция ядерной информации происхо"
казано, что многие РНК"связывающие белки,
дит на стадии поздней гаструлы, а ее реализа"
обнаруженные в свободных информосомах,
ция - только после гаструляции [50].
присутствуют в их составе лишь в качестве ми"
Спирин и его коллеги предположили, что су"
норных белков в ассоциированных с полисома"
ществование «маскированных» форм молекул
ми информосомах (см. ниже и рисунок). Данное
мРНК можно объяснить существованием ин"
наблюдение позволяет предположить, что во
формосом (см. подробное обсуждение в [13]), а
время ассоциации информосом с полисомами
именно тем, что белковый компонент этих
происходит их активный ремоделинг [37]. Позд"
мРНП предположительно играет регуляторную
нее многие из этих РНК"связывающих белков
роль в принятии решения о том, будут ли мРНК
были охарактеризованы как факторы инициа"
накапливаться и «молчать», или же они будут
ции и элонгации трансляции. Аналогичным об"
доступны для ассоциации с рибосомами. Ис"
разом был обнаружен ряд новых РНК"связыва"
пользуя для этой цели эмбрионы вьюна, преин"
ющих белков, специфически связывающихся с
кубированные с радиоактивно мечеными ами"
5′" или 3′"UTR, распознавая при этом опреде"
нокислотами (для детекции de novo транслиро"
ленные cis"элементы, такие как мотивы TOP
ванных полипептидов) или [14C]"уридином (для
(5′"концевые олигопиримидиновые моти"
детекции новосинтезированных РНК), они по"
вы [46]) или ARE (AU"rich elements [47], обога"
лучили цитоплазматические экстракты клеток
щенные остатками аденина и урацила элемен"
эмбрионов одной и той же стадии развития
ты). Таким образом, физиологическая роль та"
(поздняя гаструла). Затем экстракты смешали
ких взаимодействий между РНК"связываю"
друг с другом и подвергли фракционированию
щими белками и cis"элементами была продемо"
путем центрифугирования в градиенте сахаро"
нстрирована в контексте регуляции трансляции,
зы, что привело к выявлению дифференциаль"
метаболизма и определения локализации тран"
ного распределения меченых мРНК и полипеп"
скриптов, а также их роли в развитии патологи"
тидов между пострибосомными фракциями,
ческих состояний.
моносомами и полисомами [25, 51]. Результаты
этого эксперимента продемонстрировали, что
биосинтез белка в полисомах идет на ранее син"
СВЯЗЬ МЕЖДУ ИНФОРМОСОМАМИ
тезированных мРНК, в то время как новосинте"
И БИОСИНТЕЗОМ БЕЛКА
зированные мРНК локализуются в трансляци"
онно неактивных фракциях. На основе резуль"
Многие из ранних работ в области исследо"
татов центрифугирования в градиенте плотнос"
вания информосом были сделаны на эмбрионах
ти CsCl было показано, что почти вся новосин"
БИОХИМИЯ том 86 вып. 9 2021
ИНФОРМОСОМЫ В МЕТАБОЛИЗМЕ мРНК
1259
тезированная мРНК (> 80%) присутствует в ин"
из ядра в цитоплазму происходит в виде инфор"
формосомах (плотность ~1,40 г/см3), в полисо"
мосом, в составе которых они могут быть немед"
мах находится лишь небольшая часть мРНК
ленно транспортированы к транслирующим по"
(~1,51 г/см3), а в 80S моносомах ничего обнару"
лисомам или же поступить на хранение в виде
жено не было (~1,55 г/см3) [25]. Эти биохими"
«маскированных» мРНП. Некоторые белки ин"
ческие данные согласуются с наблюдаемой за"
формосом могут содействовать такому транс"
держкой трансляции новосинтезированных
порту, при этом неясно, меняется ли белковый
мРНК в эмбрионах ввиду образования «маски"
состав информосом в процессе их транспорта из
рованных» мРНП [13].
ядра в цитоплазму. «Прикрепление» информо"
Неожиданный результат показал анализ
сом к транслирующим рибосомам ассоциирова"
распределения мРНК информосом, 80S рибо"
но с рядом изменений в информосомах или вхо"
сом и полисом - значение плавучей плотности
дящих в их состав белках [13]. Во"вторых, ин"
мРНК"фракции полисом было ниже соответ"
формосомы представляют собой форму мРНП,
ствующего значения для моносом, что указыва"
которая служит для защиты и стабилизации
ет на присутствие в полисомах компонентов,
мРНК от дальнейшего процессинга, например,
обладающих меньшей по сравнению с РНК
от действия таких ферментов как нуклеазы.
плотностью, таких как дополнительные связан"
Действительно, информосомы намного более
ные белки [25]. Более того, диссоциация выде"
устойчивы к действию рибонуклеаз в сравнении
ленных полисом на моносомы (при добавлении
со свободной РНК, и белки информосом играют
ЭДТА) сопровождалась высвобождением гете"
«защитную роль» [13]. В"третьих, информосомы
рогенных по коэффициенту седиментации
являются составной частью механизма регуля"
мРНП, не свободных от мРНК, что также поз"
ции биосинтеза белка. Белковые компоненты
волило предположить, что информосомы ста"
информосом могут служить в качестве модуля"
бильны даже в присутствии ЭДТА [21, 25]. По"
торов трансляции, например, как репрессор
скольку комплексы обладали сходной плавучей
трансляции [13]. Еще более интересным являет"
плотностью (~ 1,4 г/см3), эти результаты свиде"
ся то, что белки в составе информосом могут
тельствовали о том, что высвобожденные мРНП
напрямую регулировать динамику ассоциации и
являются информосомами [51]. Таким образом,
диссоциации информосомы с транслирующими
было показано, что информосомы способны к
рибосомами.
ассоциации с транслирующими рибосомами, и
Такие предположения перекликаются с бо"
мРНК в живой клетке не существует в «обна"
лее ранними концептуальными идеями, в кото"
женном» виде, независимо от того, транслирует"
рых термин «информационная РНК» использо"
ся она в данный момент или нет.
вался для описания промежуточных состояний
РНК различного размера, не являющихся ни
рибосомной, ни транспортной РНК, и обладаю"
ИНФОРМОСОМЫ
щих способностью к гибридизации с ДНК
В СЛОЖНОМ МИРЕ мРНП
(«комплементарная РНК») [53, 54]. Одним из
важных следствий такой концепции является
Основываясь на доступных эксперименталь"
то, что каждая «комплементарная РНК» все еще
ных данных, Спирин с соавт. предложили следу"
содержит последовательность"специфичную
ющую концепцию взаимосвязей между мРНК и
информацию, необходимую для гибридизации с
информосомами [52]:
определенной ДНК. Таким образом, в то время
1. эукариотические мРНК не существуют в
как любая мРНК может рассматриваться как
свободном виде и в условиях in vivo всегда нахо"
«информационная» (матричная; имеется в виду
дятся в комплексе с белками;
её непосредственная роль в биосинтезе белка),
2. мРНП нерибосомной природы являются
было также высказано предположение о суще"
информосомами;
ствовании других типов «информационных»
3. многие эукариотические белки, участвую"
РНК, которые не служат непосредственно в ка"
щие в метаболизме РНК, обладают РНК"связы"
честве источников информации для синтеза
вающей активностью;
белка, но могут иметь, например, регуляторную
4. отдельные белковые компоненты инфор"
функцию. Открытие информосом привело к
мосом (и других РНП) являются РНК"связыва"
значительному расширению знаний в этой об"
ющими белками.
ласти, продемонстрировав, что белковый ком"
Еще в 1969 г. было сделано несколько прог"
понент различных рибонуклеопротеиновых час"
нозов, основанных на предположениях, вытека"
тиц, не содержащих рибосомы (вирусные
ющих из этой концепции [13]. Во"первых, было
РНП/мРНП, ядерные РНП/мРНП, цитоплаз"
высказано предположение, что экспорт мРНК
матические «маскированные» мРНП), может
БИОХИМИЯ том 86 вып. 9 2021
1260
ИВАНОВ
активно влиять на их судьбу в клетке, в том чис"
рые находятся в комплексе с мРНК внутри
ле на их внутриклеточную локализацию, ста"
мРНП. Утверждение, что RBPs играют значи"
бильность или ассоциацию с рибосомами. В
тельную роль в определении судьбы и функций
этом плане термин «информосомы» значитель"
РНП, основанное на результатах изучения ин"
но шире, чем «мРНП», и он скорее служит для
формосом, является фундаментальной основой
объединения различных РНП, которые могут
для понимания структуры и функций рибонук"
содержать мРНК, а также РНП, в которых
леопротеинов [55]. В ранних работах по инфор"
мРНК отсутствует (например, комплексы пре"
мосомам было постулировано несколько прин"
мРНК или компоненты РНК"гранул).
ципов, которые могут быть применимы и к дру"
Жесткий контроль определенных стадий ме"
гим мРНП, многие из которых хорошо изучены
таболизма мРНК, таких как транспорт, деграда"
(рисунок). Например, мРНП часто локализуют"
ция и трансляция мРНК, жизненно необходим
ся в определенных внутриклеточных компарт"
для регуляции экспрессии генов. Все указанные
ментах [56]. Такая компартментализация облег"
процессы модулируются путем взаимодействия
чает активацию определенных процессов биоге"
с РНК"связывающими белками (RBPs), кото"
неза мРНК (процессинг мРНК и др.) как сос"
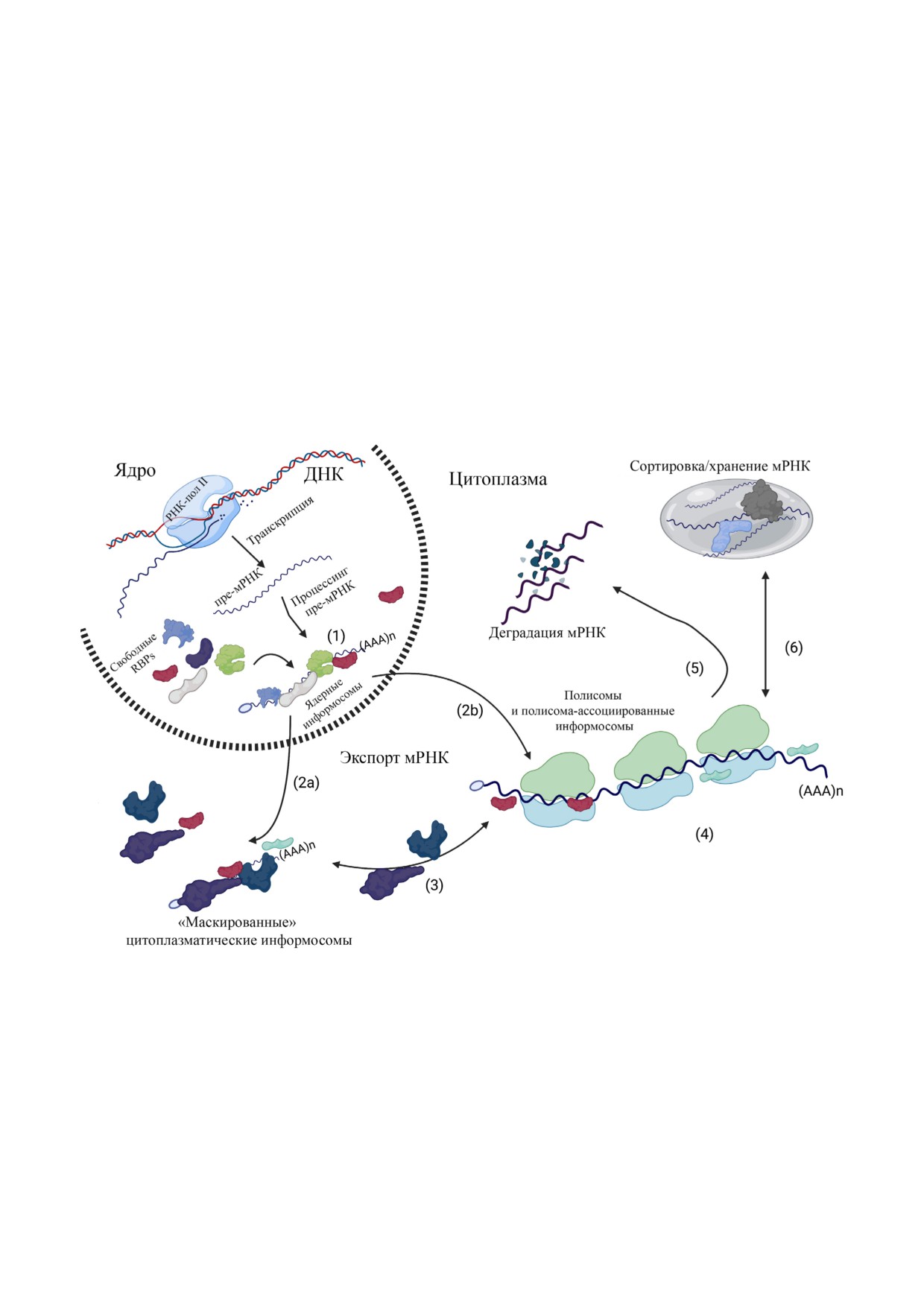
Информосомы и метаболизм мРНК. С помощью РНК"полимеразы II в ядре в результате транскрипции образуются пре"
мРНК, которые затем подвергаются процессингу с образованием зрелых мРНК. Затем процессированные мРНК связы"
ваются с РНК"связывающими белками с формированием ядерных информосом (1). Эти ядерные комплексы экспорти"
руются в цитоплазму, в которой они либо находятся в нетранслируемой форме («маскированные» цитоплазматические
информосомы) (2a), либо ассоциируются с транслирующими полисомами с образованием полисома"ассоциированных
информосом (2b). В процессе экспорта мРНК из ядра некоторые ядерные РНК"связывающие белки (RBPs) заменяются
на свои цитоплазматические аналоги. В цитоплазме «маскированные» информосомы могут активироваться под действи"
ем внешних или внутренних стимулов и ассоциироваться с рибосомой с образованием полисома"ассоциированных ин"
формосом (3). В ходе трансляции мРНК (4) ассоциированные с полисомами информосомы могут отделиться от трансли"
рующих рибосом и вернуться в «маскированную» форму (3), могут быть направлены на деградацию (5) или сортировку с
последующим запасанием РНК"гранул, таких как стрессовые гранулы (stress granules) и процессирующие тельца (pro"
cessing bodies) (6). Этот рисунок был создан на сайте BioRender.com
БИОХИМИЯ том 86 вып. 9 2021
ИНФОРМОСОМЫ В МЕТАБОЛИЗМЕ мРНК
1261
тавной части программы развития (классичес"
мРНК, а также обеспечению её правильного
кие «маскированные» информосомы эмбрионов
расположения относительно стартового AUG
вьюна) или в ответ на изменения условий (нап"
кодона. В этой связи следует отметить, что в от"
ример, стресс) [57]. Многие мРНП являются ди"
дельных работах высказывалось предположе"
намичными, что означает возможность их ассо"
ние, что информосомы возможно регистриро"
циации с другими РНП (например, обратимое
вать в виде комплексов мРНК с 40S субъедини"
взаимодействие информосом с транслирующи"
цами рибосомы (45S информосомы), напоми"
ми рибосомами), а также могут менять свой сос"
нающими рибосомные комплексы преинициа"
тав в зависимости от изменений микроокруже"
ции. При распознавании старт"кодона большая
ния (например, превращение ядерных инфор"
60S субъединица рибосомы объединяется с т.н.
мосом в цитоплазматические в процессе экс"
преинициационным 48S комплексом с дальней"
порта мРНК).
шим образованием функциональной 80S рибо"
Важным следствием концепции информосо"
сомы [59]. В дальнейшем 80S рибосома участву"
мы стало сопряжение различных связанных с
ет в процессе элонгации трансляции и синтезе
мРНК процессов через взаимодействие мРНП с
насцентных полипептидов, закодированных в
другими РНК"связывающими белками (рису"
открытой рамке считывания молекул мРНК.
нок). В настоящее время известно, что после за"
После завершения трансляции 80S рибосома
вершения транскрипции и процессинга зрелые
снова распадается на 60S и 40S субъединицы.
мРНК связываются со множеством различных
Следует, однако, отметить, что эффективно
белков (в частности, из семейства гетерогенных
транслируемые мРНК задействуют для
ядерных рибонуклеопротеинов (hnРНП), кото"
собственной трансляции сразу множество рибо"
рые оказывают влияние на различных стадиях
сом, а после стадии терминации 40S субъедини"
метаболизма мРНК (транспорт, определение
цы подвергаются рециклизации для участия в
локализации, стабильность)) [55]. По крайней
следующем раунде инициации трансляции на
мере некоторые из этих ядерных мРНП являют"
той же молекуле мРНК [60]. Этому процессу
ся классическими ядерными информосомами
способствует циркуляризация мРНК, опосредо"
(рисунок). После экспорта мРНК из ядра неко"
ванная взаимодействием фактора инициации
торые ядерные РНК"связывающие белки заме"
трансляции eIF4G, связанным с кэп"структурой
няются на другие, участвующие в регуляции ме"
мРНК (в составе комплекса eIF4F), с поли(A)"
таболизма мРНК в цитоплазме. Некоторые из
связывающим белком (PABP), расположенным
этих цитоплазматических белков обладают мно"
на поли(A)"хвосте [61]. Заманчиво предполо"
жественными функциями, т.е. они участвуют в
жить, что наблюдаемые информосомы, ассоции"
определении локализации, поддержании ста"
рованные с полисомами, или высвобождаю"
бильности или/и трансляции мРНК [58]. Мно"
щиеся из полирибосом, являются, по крайней
жество мРНП, обнаруженных в специализиро"
мере, одним из представителей мРНП, ассоции"
ванных клетках (например, ооциты), по поведе"
рованных с фактором трансляции.
нию напоминают «маскированные» информо"
Недавно было показано, что самоорганиза"
сомы из эмбрионов вьюна, т.к. содержат белки,
ция мРНП в различные не окруженные мембра"
участвующие в определении их внутриклеточ"
ной органеллы, называемые РНК"гранулами,
ной локализации и стабилизации [57] (рису"
представляет собой эволюционно консерватив"
нок).
ный феномен, лежащий в основе различных ас"
Обнаружение факта взаимодействия инфор"
пектов метаболизма мРНК [62, 63]. Были иден"
мосом с транслирующими рибосомами открыло
тифицированы высокомолекулярные «гигант"
путь к изучению молекулярных механизмов ре"
ские» информосомы, которые могут быть про"
гуляции трансляции. Изменение значения пла"
дуктом самоагрегации. Таким образом, возмож"
вучей плотности, которое наблюдается у транс"
но предположить, что информосомы могут быть
лирующих полисом относительно 80S рибосом,
частью других мРНП"содержащих РНК"гранул.
свидетельствует о том, что произошло связыва"
В контексте метаболизма мРНК лучше других
ние дополнительных белков с полирибосома"
были изучены два класса таких цитоплазмати"
ми [51]. В настоящее время стало совершенно
ческих РНК"гранул [64]. В первый класс входят
ясным, что группы факторов трансляции, в осо"
т.н. процессирующие тельца, или РНК"гранулы,
бенности факторы инициации трансляции, в
обогащенные деаденилированными мРНК и
первую очередь взаимодействуют с мРНК, и
специфическими факторами деградации мРНК,
лишь потом - с рибосомой [59]. Факторы ини"
которые, как предполагается, участвуют в рас"
циации трансляции локализуются вокруг кэп"
паде и трансляции мРНК [65]. Второй класс
структуры и способствуют взаимодействию
мРНК"содержащих гранул составляют т.н.
между малой 40S субъединицей рибосомы и
стрессовые гранулы, в которых содержится
БИОХИМИЯ том 86 вып. 9 2021
1262
ИВАНОВ
мРНК, трансляция которой была приостанов"
мосомами/мРНП, но также затрагивает другие
лена на уровне инициации [66, 67]. Стрессовые
РНП. Было высказано предположение, что в
гранулы формируются при различных видах
процессе эволюции РНП было достигнуто опти"
стресса. Полагают, что они оказывают влияние
мальное стехиометрическое и структурное соот"
на выживаемость клеток через участие в репро"
ветствие между РНК и их белковыми партнера"
граммировании трансляции [68]. Поскольку эти
ми, определяющее их совместное функциони"
два типа гранул содержат множество молекул
рование в качестве рибонуклеопротеиновой
мРНК, связанных с различными РНК"связыва"
частицы [69]. Эти биофизические принципы
ющими белками, то классические информосо"
сборки РНК"содержащих комплексов в настоя"
мы могут входить в состав этих РНК"содержа"
щее время интенсивно исследуются, т.к. многие
щих гранул.
из таких комплексов вносят значительный
вклад в поддержание здоровья человека, а также
играют роль в развитии заболеваний.
ВЫВОДЫ
Гипотеза о том, что мРНК всегда находятся в
Благодарности. Павел Иванов выражает бла"
связанном с белками состоянии и существуют в
годарность сотрудникам его лаборатории и док"
виде информосомы, более полувека назад носи"
тору Claire Riggs за помощь в редактировании
ла революционный характер. Информосомы яв"
статьи.
ляются динамичными структурами, и их сборка
Финансирование. Работа не была финанси"
зависит и от мРНК, и от белков. Согласно моде"
рована какими"либо фондами или проектами.
ли информосомы, взаимодействующие с мРНК
Конфликт интересов. Автор заявляет об отсут"
белки играют не только структурную роль при
ствии конфликта интересов в финансовой или
формировании мРНП, но также участвуют в ре"
какой"либо иной сфере.
гуляции метаболизма мРНК. Следует отметить,
Соблюдение этических норм. Настоящая
что провозглашенный Спириным принцип, ко"
статья не содержит описания каких"либо иссле"
торый звучит как «omnia mea mecum porto» («всё
дований с участием людей или животных в каче"
своё ношу с собой») [52], не ограничен инфор" стве объектов, выполненных автором.
СПИСОК ЛИТЕРАТУРЫ
1.
Bentley, D. L. (2014) Coupling mRNA processing with
10. Spirin, A. S., Belitsina, N. V., and Lerman, M. I. (1965)
transcription in time and space, Nat. Rev. Genet., 15, 163"
Use of formaldehyde fixation for studies of ribonucleopro"
175, doi: 10.1038/nrg3662.
tein particles by caesium chloride density"gradient cen"
2.
Luna, R., Gaillard, H., Gonzalez"Aguilera, C., and
trifugation, J. Mol. Biol., 14, 611"615, doi: 10.1016/s0022"
Aguilera, A. (2008) Biogenesis of mRNPs: integrating dif"
2836(65)80213"3.
ferent processes in the eukaryotic nucleus, Chromosoma,
11. Spirin, A. S., Belitsina, N. V., and Aitkhozhin, M. A.
117, 319"331, doi: 10.1007/s00412"008"0158"4.
(1964) Messenger RNA in early embryogenesis, Zhurn.
3.
Corley, M., Burns, M. C., and Yeo, G. W. (2020) How
Obshch. Biol., 25, 321"338.
RNA"binding proteins interact with RNA: molecules and
12. Ovchinnikov, L. P., Ajtkhozhin, M. A., Bystrova, T. F., and
mechanisms, Mol. Cell, 78, 9"29, doi: 10.1016/j.molcel.
Spirin, A. S. (1969) Newt embryo informosomes: 1. Sediment"
2020.03.011.
ation and density parameters, Mol. Biol. (USSR), 3, 449"464.
4.
Khong, A., and Parker, R. (2020) The landscape of eukary"
13. Spirin, A. S. (1969) The Second Sir Hans Krebs Lecture.
otic mRNPs, RNA, 26, 229"239, doi: 10.1261/rna.
Informosomes, Eur. J. Biochem., 10, 20"35.
073601.119.
14. Spirin, A. S., and Nemer, M. (1965) Messenger RNA in
5.
Hershey, J. W. B., Sonenberg, N., and Mathews, M. B.
early sea"urchin embryos: cytoplasmic particles, Science,
(2019) Principles of translational control, Cold Spring Harb.
150, 214"217, doi: 10.1126/science.150.3693.214.
Perspect. Biol., 11, doi: 10.1101/cshperspect.a032607.
15. Belitsina, N. V., Ovchinnikov, L. P., Spirin, A. S., Gendon,
6.
Mateu"Regue, A., Nielsen, F. C., and Christiansen, J.
Yu. Z., and Cheros, V. I. (1968) Informosomes of HeLa
(2020) Cytoplasmic mRNPs revisited: singletons and con"
cells infected with vaccinia virus, Mol. Biol. (U.S.S.R.), 2.
densates, Bioessays, 42, e2000097, doi: 10.1002/bies.
16. Infante, A. A., and Nemer, M. (1968) Heterogeneous
202000097.
ribonucleoprotein particles in the cytoplasm of sea urchin
7.
Keene, J. D. (2007) RNA regulons: coordination of post"
embryos, J. Mol. Biol., 32, 543"565, doi: 10.1016/0022"
transcriptional events, Nat. Rev. Genet., 8, 533"543,
2836(68)90342"2.
doi: 10.1038/nrg2111.
17. Joklik, W. K., and Becker, Y. (1965) Studies on the genesis
8.
Aitkhozhin, M. A., Belitsina, N. V., and Spirin, A. S.
of polyribosomes. II. The association of nascent messenger
(1964) Nucleic Acids in the early stages of development of
RNA with the 40 S subribosomal particle, J. Mol. Biol., 13,
fish embryos (based on the loach Misgurnus Fossilis),
511"520, doi: 10.1016/s0022"2836(65)80113"9.
Biokhimiia, 29, 169"175.
18. Shatkin, A. J., Sebring, E. D., and Salzman, N. P. (1965)
9.
Belitsina, N. V., Aitkhozhin, M. A., Gavrilova, L. P., and
Vaccinia virus directed RNA: its fate in the presence of
Spirin, A. S. (1964) The messenger ribonucleic acids of dif"
actinomycin, Science, 148, 87"90, doi: 10.1126/science.
ferentiating animal cells, Biokhimiia, 29, 363"374.
148.3666.87.
БИОХИМИЯ том 86 вып. 9 2021
ИНФОРМОСОМЫ В МЕТАБОЛИЗМЕ мРНК
1263
19.
McConkey, E. H., and Hopkins, J. W.
(1965)
36.
Ovchinnikov, L. P. and Avanesov, A. C.
(1969)
Subribosomal particles and the transport of messenger
Informosomes of loach embryos. 3. Specificity of interac"
RNA in HeLa cells, J. Mol. Biol.,
14,
257"270,
tion of “informosome"forming” protein with RNA, Mol.
doi: 10.1016/s0022"2836(65)80245"5.
Biol. (U.S.S.R.), 3, 5"12.
20.
Kafatos, F. C. (1968) Cytoplasmic particles carrying rapid"
37.
Spirin, A. S. (1994) Storage of messenger RNA in eukary"
ly labeled RNA in developing insect epidermis, Proc. Natl.
otes: envelopment with protein, translational barrier at 5′
Acad. Sci. USA, 59, 1251"1258, doi: 10.1073/pnas.59.4.
side, or conformational masking by 3′ side? Mol. Reprod.
1251.
Dev., 38, 107"117, doi: 10.1002/mrd.1080380117.
21.
Henshaw, E. C. (1968) Messenger RNA in rat liver polyri"
38.
Blobel, G. (1972) Protein tightly bound to globin mRNA,
bosomes: evidence that it exists as ribonucleoprotein parti"
Biochem. Biophys. Res. Commun., 47, 88"95, doi: 10.1016/
cles, J. Mol. Biol., 36, 401"411, doi: 10.1016/0022"
s0006"291x(72)80014"7.
2836(68)90164"2.
39.
Blobel, G. (1973) A protein of molecular weight 78,000
22.
Perry, R. P., and Kelley, D. E. (1968) Messenger RNA"pro"
bound to the polyadenylate region of eukaryotic messenger
tein complexes and newly synthesized ribosomal subunits:
RNAs, Proc. Natl. Acad. Sci. USA,
70,
924"928,
analysis of free particles and components of polyribosomes,
doi: 10.1073/pnas.70.3.924.
J. Mol. Biol.,
35,
37"59, doi:
10.1016/s0022"
40.
Jain, S. K., Pluskal, M. G., and Sarkar, S. (1979) Thermal
2836(68)80035"x.
chromatography of eukaryotic messenger ribonucleopro"
23.
Levy, H. B., and Carter, W. A. (1968) Molecular basis of the
tein particles on oligo (dT)"cellulose. Evidence for com"
action of interferon, J. Mol. Biol.,
31,
561"577,
mon mRNA"associated proteins in various cell types,
doi: 10.1016/0022"2836(68)90428"2.
FEBS Lett., 97, 84"90, doi: 10.1016/0014"5793(79)80058"7.
24.
Volkova, M. Y., Zaides, V. M., and Zaslavsky, V. G. (1969)
41.
Kumar, A., and Pederson, T. (1975) Comparison of pro"
Slowly sedimenting particles present in cytoplasmic extract
teins bound to heterogeneous nuclear RNA and messenger
of Ehrlich ascites cells infected by Sendai virus, Mol. Biol.
RNA in HeLa cells, J. Mol. Biol.,
96,
353"365,
(U.S.S.R.), 3, 4"9.
doi: 10.1016/0022"2836(75)90165"5.
25.
Ovchinnikov, L. P., Belitsina, N. V., Avanesov, A., and
42.
Morel, C., Kayibanda, B., and Scherrer, K. (1971) Proteins
Spirin, A. S. (1969) Postribosomal RNA"containing parti"
associated with globin messenger RNA in avian erythrob"
cles of cytoplasm of animal cells according to CsCl density
lasts: isolation and comparison with the proteins bound to
gradient centrifugation data, Dokl. Akad. Nauk SSSR, 186,
nuclear messenger"likie RNA, FEBS Lett., 18, 84"88,
1202"1205.
doi: 10.1016/0014"5793(71)80413"1.
26.
Neifakh, A. A. (1959) Method of inactivation of nuclei by
43.
van Venrooij, W. J., van Eekelen, C. A., Jansen, R. T., and
radiation and its possible applications for the investigation
Princen, J. M. (1977) Specific poly"A"binding protein of
of nuclei functions during early development of fish,
76,000 molecular weight in polyribosomes is not present on
Zhurn. Obshch. Biol. (Russian), 20, 202"207.
poly A of free cytoplasmic mRNP, Nature, 270, 189"191,
27.
Samarina, O. P., Asriian, I. S., and Georgiev, G. P. (1965)
doi: 10.1038/270189a0.
Isolation of nuclear nucleoproteins containing informa"
44.
Minich, W. B., Maidebura, I. P., and Ovchinnikov, L. P.
tional ribonucleic acid, Dokl. Akad. Nauk SSSR, 163,
(1993) Purification and characterization of the major 50"
1510"1513.
kDa repressor protein from cytoplasmic mRNP of rabbit
28.
Samarina, O. P., Krichevskaya, A. A., and Georgiev, G. P.
reticulocytes, Eur. J. Biochem., 212, 633"638, doi: 10.1111/
(1966) Nuclear ribonucleoprotein particles containing
j.1432"1033.1993.tb17701.x.
messenger ribonucleic acid, Nature, 210, 1319"1322,
45.
Mordovkina, D., Lyabin, D. N., Smolin, E. A., Sogorina,
doi: 10.1038/2101319a0.
E. M., Ovchinnikov, L. P., and Eliseeva, I. (2020) Y"box
29.
Samarina, O. P., Lerman, M. I., Tumanian, V. D.,
binding proteins in mRNP assembly, translation, and stabil"
Anan’eva, L. N., and Georgiev, G. P. (1965) Character"
ity control, Biomolecules, 10, doi: 10.3390/biom10040591.
istics of chromosomal information RNA, Biokhimiia, 30,
46.
Cockman, E., Anderson, P., and Ivanov, P. (2020) TOP
880"893.
mRNPs: molecular mechanisms and principles of regula"
30.
Samarina, O. P., Lukanidin, E. M., and Georgiev, G. P.
tion, Biomolecules, 10, doi: 10.3390/biom10070969.
(1967) On the structural organization of the nuclear com"
47.
Ivanov, P., and Anderson, P. (2013) Post"transcriptional
plexes containing messenger RNA, Biochim. Biophys. Acta,
regulatory networks in immunity, Immunol. Rev., 253, 253"
142, 561"564, doi: 10.1016/0005"2787(67)90642"9.
272, doi: 10.1111/imr.12051.
31.
Samarina, O. P., Lukanidin, E. M., Molnar, J., and
48.
Stebbins"Boaz, B., and Richter, J. D. (1997) Translational
Georgiev, G. P. (1968) Structural organization of nuclear
control during early development, Crit. Rev. Eukaryot. Gene
complexes containing DNA"like RNA, J. Mol. Biol., 33,
Expr., 7, 73"94, doi: 10.1615/critreveukargeneexpr.v7.i1"2.50.
251"263, doi: 10.1016/0022"2836(68)90292"1.
49.
Teixeira, F. K., and Lehmann, R. (2019) Translational con"
32.
Samarina, O. P., Molnar, J., Lukanidin, E. M., Bruskov,
trol during developmental transitions, Cold Spring Harb.
V. I., Krichevskaya, A. A., and Georgiev, G. P. (1967)
Perspect. Biol., 11, doi: 10.1101/cshperspect.a032987.
Reversible dissociation of nuclear ribonucleoprotein parti"
50.
Neyfakh, A. A. (1964) Radiation investigation of nucleo"
cle containing mRNA into RNA and protein, J. Mol. Biol.,
cytoplasmic interrelations in morphogenesis and biochem"
27, 187"191, doi: 10.1016/0022"2836(67)90359"2.
ical differentiation, Nature, 201, 880"884, doi: 10.1038/
33.
Girard, M., and Baltimore, D. (1966) The effect of HeLa
201880a0.
cell cytoplasm on the rate of sedimentation of RNA, Proc.
51.
Ovchinnikov, L. P., Bystrova, T. F., and Spirin, A. S. (1969)
Natl. Acad. Sci. USA, 56, 999"1002, doi: 10.1073/pnas.56.
Sedimentation and density characteristics of ribosomes
3.999.
and their subunits from embryonic groundlings, Dokl.
34.
Ovchinnikov, L. P., Voronina, A. S., Stepanov, A. S.,
Akad. Nauk SSSR, 185, 210"213.
Belitsina, N. V., and Spirin, A. S. (1968) Informosome"like
52.
Spirin, A. S. (1978) Eukaryotic messenger RNA and infor"
complexes were formed by RNA adding to animal cell
mosomes. Omnia mea mecum porto, FEBS Lett., 88, 15"
homogenates, Mol. Biol. (U.S.S.R.), 2, 752"761.
17, doi: 10.1016/0014"5793(78)80596"1.
35.
Ovchinnikov, L. P., Avanesov, A. C., and Spirin, A. S.
53.
Spiegelman, S. (1961) The relation of informational RNA
(1969) Informosomes from loach embryos, Molek. Biol.
to DNA, Cold Spring Harb. Symp. Quant. Biol., 26, 75"90,
(U.S.S.R.), 3, 465471.
doi: 10.1101/sqb.1961.026.01.013.
БИОХИМИЯ том 86 вып. 9 2021
1264
ИВАНОВ
54.
Spiegelman, S., Hall, B. D., and Storck, R. (1961) The
62. Zhang, H., Ji, X., Li, P., Liu, C., Lou, J., et al. (2020)
occurrence of natural DNA-RNA complexes in E. coli
Liquid-liquid phase separation in biology: mechanisms,
infected with T2, Proc. Natl. Acad. Sci. USA, 47, 1135"
physiological functions and human diseases, Sci. China
1141, doi: 10.1073/pnas.47.8.1135.
Life Sci.,
63,
953"985, doi:
10.1007/s11427"020"
55.
Bjork, P., and Wieslander, L. (2017) Integration of mRNP
1702"x.
formation and export, Cell. Mol. Life Sci., 74, 2875"2897,
63. Hyman, A. A., Weber, C. A., and Julicher, F. (2014)
doi: 10.1007/s00018"017"2503"3.
Liquid"liquid phase separation in biology, Annu. Rev. Cell
56.
Fazal, F. M., Han, S., Parker, K. R., Kaewsapsak, P.,
Dev. Biol., 30, 39"58, doi: 10.1146/annurev"cellbio"
Xu, J., et al. (2019) Atlas of subcellular RNA localization
100913"013325.
revealed by APEX"Seq, Cell,
178,
473"490.e426,
64. Ivanov, P., Kedersha, N., and Anderson, P. (2019) Stress
doi: 10.1016/j.cell.2019.05.027.
granules and processing bodies in translational control,
57.
Kong, J., and Lasko, P. (2012) Translational control in cel"
Cold Spring Harb. Perspect. Biol., 11, doi: 10.1101/cshper"
lular and developmental processes, Nat. Rev. Genet., 13,
spect.a032813.
383"394, doi: 10.1038/nrg3184.
65. Luo, Y., Na, Z., and Slavoff, S. A. (2018) P"Bodies: com"
58.
Singh, G., Pratt, G., Yeo, G. W., and Moore, M. J. (2015)
position, properties, and functions, Biochemistry, 57, 2424"
The clothes make the mRNA: past and present trends in
2431, doi: 10.1021/acs.biochem.7b01162.
mRNP fashion, Annu. Rev. Biochem., 84, 325"354,
66. Hofmann, S., Kedersha, N., Anderson, P., and Ivanov, P.
doi: 10.1146/annurev"biochem"080111"092106.
(2021) Molecular mechanisms of stress granule assembly
59.
Jackson, R. J., Hellen, C. U., and Pestova, T. V. (2010) The
and disassembly, Biochim. Biophys. Acta Mol. Cell Res.,
mechanism of eukaryotic translation initiation and princi"
1868, 118876, doi: 10.1016/j.bbamcr.2020.118876.
ples of its regulation, Nat. Rev. Mol. Cell Biol., 11, 113"127,
67. Riggs, C. L., Kedersha, N., Ivanov, P., and Anderson, P.
doi: 10.1038/nrm2838.
(2020) Mammalian stress granules and P bodies at a
60.
Hellen, C. U. T. (2018) Translation termination and ribo"
glance, J. Cell Sci., 133, doi: 10.1242/jcs.242487.
some recycling in eukaryotes, Cold Spring Harb. Perspect.
68. Advani, V. M., and Ivanov, P. (2019) Translational control
Biol., 10, doi: 10.1101/cshperspect.a032656.
under stress: reshaping the translatome, Bioessays, 41,
61.
Wells, S. E., Hillner, P. E., Vale, R. D., and Sachs, A. B.
e1900009, doi: 10.1002/bies.201900009.
(1998) Circularization of mRNA by eukaryotic translation
69. Mitchell, S. F., and Parker, R. (2014) Principles and prop"
initiation factors, Mol. Cell, 2, 135"140, doi: 10.1016/
erties of eukaryotic mRNPs, Mol. Cell, 54, 547"558,
s1097"2765(00)80122"7.
doi: 10.1016/j.molcel.2014.04.033.
INFORMOSOMES TRAVEL IN TIME: AN EARLY mRNA CONCEPT
IN THE CURRENT mRNP LANDSCAPE
Review
P. Ivanov1,2,3
1 Division of Rheumatology, Inflammation, and Immunity, Brigham and Women’s Hospital,
Boston, Massachusetts, 02115 USA; e mail: pivanov@rics.bwh.harvard.edu
2 Department of Medicine, Harvard Medical School, Boston, Massachusetts, 02115 USA
3 Harvard Medical School Initiative for RNA Medicine, Harvard University, Boston, Massachusetts, 02115 USA
Messenger RNA is complexed with proteins throughout its life cycle. The first mRNA"containing particles of non"
ribosomal nature, named informosomes, were discovered in cytoplasmic extracts of fish embryos by the laboratory of
Alexander Spirin, and later described in live cells. Over time, various other nuclear and cytoplasmic mRNA"contain"
ing ribonucleoproteins (mRNPs) have been found and characterized. Although these mRNPs are very diverse in their
subcellular localization, structure and functions, they share many common characteristics with informosomes. In this
mini"review, I will discuss the discovery of informosomes, their characteristics and proposed functions, and their
potential relationship to other mRNPs.
Keywords: informosomes, mRNA, ribonucleoproteins, mRNP, RNA granules, protein biosynthesis
БИОХИМИЯ том 86 вып. 9 2021