БИОХИМИЯ, 2021, том 86, вып. 9, с. 1366 - 1379
УДК 577.21
МАЛ, ДА УДАЛ. РАЗНООБРАЗИЕ ФУНКЦИЙ МАЛЫХ БЕЛКОВ
И ПЕПТИДОВ - ПРОДУКТОВ ТРАНСЛЯЦИИ
КОРОТКИХ РАМОК СЧИТЫВАНИЯ
Обзор
© 2021
П.В. Сергиев1,2,3*, М.П. Рубцова1*
1 Московский государственный университет им. М.В. Ломоносова, химический факультет, 119991 Москва,
Россия; электронная почта: petya@genebee.msu.ru, mprubtsova@gmail.com
2 Центр наук о жизни Сколковского института науки и технологий, 143025 дер. Сколково, Россия
3 Институт функциональной геномики Московского государственного университета им. М.В. Ломоносова,
119991 Москва, Россия
Поступила в редакцию 27.04.2021
После доработки 27.05.2021
Принята к публикации 27.05.2021
Функционирование клетки является сложно регулируемым процессом. Долгие годы исследования в облас
тях протеомики и функциональной геномики были сфокусированы на роли белков в жизнедеятельности
клетки. Развитие науки привело к пониманию того, что короткие открытые рамки считывания, ранее счи
тавшиеся нефункциональными, в действительности выполняют разнообразные функции. В составе поли
цистронных мРНК короткие рамки считывания часто регулируют их стабильность и эффективность транс
ляции основной рамки считывания. Совершенствование методов протеомного анализа позволило иденти
фицировать продукты трансляции коротких рамок считывания в количествах, позволяющих предположить
существование функциональной роли у синтезируемых пептидов и коротких белков. Их изучение открыва
ет новый уровень регуляции функционирования клетки и ее адаптации к изменяющимся условиям. Насто
ящий обзор посвящен анализу функций недавно открытых пептидов и коротких белков.
КЛЮЧЕВЫЕ СЛОВА: пептиды, малые рамки считывания, длинные некодирующие РНК, альтернативные
белки.
DOI: 10.31857/S0320972521090098
ВВЕДЕНИЕ
скриптах, на первых порах считали случай
ностью и игнорировали. С развитием методов
Разнообразие белков предопределяет функ
системного исследования биосинтеза белка в
ционирование живых существ, а для таких
клетках, в первую очередь рибосомного профи
сложных организмов, как млекопитающие, да
лирования [6], а также в результате совершен
же простая каталогизация протеома является
ствования методов протеомного анализа [7-10]
нетривиальным и далеко не законченным про
научное сообщество с удивлением обнаружило,
цессом. В результате развития системной биоло
что и трансляция транскриптома не ограничена
гии стало очевидным, что транскрипции под
каноническими длинными рамками считыва
вергается много больше участков генома, чем
ния, начинающимися непременно с инициатор
предполагалось ранее [1, 2]. С возникновением
ного AUG кодона. Результатом такой «всеоб
методов высокопроизводительного секвениро
щей» трансляции являются короткие белки и
вания РНК [3] описание транскриптов достигло
пептиды (sPEP), продукты трансляции т.н. ко
небывалых ранее масштабов. Поскольку многие
ротких открытых рамок считывания (кОРС), а
из выявленных транскриптов не содержали про
также длинные белки, необычные продукты
тяженных рамок считывания, их в большинстве
трансляции известных мРНК, часто называе
своем аннотировали как длинные некодирую
мые альтернативными белками (alt PROT), или
щие РНК (см. [4, 5] для обзора). Короткие рам
некодирующих РНК.
ки считывания, обнаруживаемые в этих тран
Не акцентируя чрезмерно внимание на сов
ременных методах выявления кодирующих пос
Принятые сокращения: НТО - нетранслируемая
ледовательностей в геномах и транскриптомах,
область; ОРС - открытая рамка считывания.
авторы рекомендуют заинтересованным в мето
* Адресат для корреспонденции.
дологической стороне вопроса читателям обра
1366
РАЗНООБРАЗИЕ ФУНКЦИЙ МАЛЫХ БЕЛКОВ И ПЕПТИДОВ
1367
титься к нескольким обзорным статьям на эту
может служить механизмом регуляции трансля
тему [11, 12]. В данном обзоре мы остановимся
ции «основной» рамки считывания. Этот про
на результатах исследований последних лет,
цесс хорошо изучен для мРНК GCN4 дрожжей,
посвященных поиску функциональной роли
содержащей несколько 5′ ОРС, наличие кото
малых белков.
рых обеспечивает селективную трансляцию
этой мРНК в условиях аминокислотного голо
дания [15]. В дальнейших исследованиях было
ГДЕ НАХОДЯТ НОВЫЕ
показано, что подобный тип регуляции очень
ТРАНСЛИРУЕМЫЕ ОРС?
распространен [16]. В случае 5′ ОРС MIEF1,
DDIT3, FBXO9, HMGA2 и HAUS6 обнаружено
Где же закодированы неизвестные ранее бел
взаимодействие продуктов трансляции «основ
ки, обнаруживаемые с помощью современных
ной» рамки считывания и ОРС, расположенной
компьютерных и экспериментальных методов,
ближе к 5′ концу соответствующих мРНК [13].
и в чем их особенности, позволившие им оста
В этих случаях, возможно, расположение двух
ваться в тени столько времени? Во первых,
рамок на одной мРНК необходимо для коорди
бóльшая часть этих белков, хотя и не все, имеет
нированного синтеза соответствующих белков,
небольшие размеры, до 100 аминокислотных
как это происходит в случае бактериальных опе
остатков (а.о.). Поскольку в любой некодирую
ронов. При этом остается открытым вопрос о
щей последовательности можно встретить слу
механизмах эффективной трансляции обеих ра
чайно присутствующие нефункциональные ко
мок считывания в системе трансляции эукариот.
роткие рамки считывания, исследователю дос
Можно предположить, что повышенная час
таточно сложно выявить ту небольшую долю ра
тота использования отличающихся от AUG ко
мок считывания, которые транслируются с об
донов в качестве инициаторных для 5′ ОРС
разованием функциональных коротких белков и
имеет «механистическое» объяснение. Исполь
пептидов. Во вторых, новые белки образуются
зование AUG кодона в данном случае приведет
при трансляции, которая начинается с некано
к стабильному ингибированию трансляции ос
нических инициаторных кодонов. Так, помимо
новной рамки. Против такого варианта, если он
AUG, который остается наиболее распростра
возникнет в ходе случайной мутации, будет
ненным инициаторным кодоном (с него начи
действовать отбор, связанный с функциониро
нается трансляция в 64% случаев, по дан
ванием основной ОРС. Напротив, использова
ным [13], в 43% - по данным [9]), у эукариот
ние инициаторного кодона, который может с
трансляция часто начинается с CUG (в 3% - по
некоторой вероятностью пропускаться сканиру
данным [9]; в 23% - [13]), GUG (в 6% - [9]; в
ющим комплексом, будет закрепляться отбо
9% - [13]) и UUG (в 4% - [13]) кодонов. До
ром, поскольку способствует синтезу обоих бел
вольно большой разброс частот использования
ковых продуктов, закодированных в мРНК [17].
различных кодонов в качестве инициаторных
Значительная доля выявленных функцио
объясняется использованием разных выборок
нальных ОРС, по некоторым данным
-
ОРС, содержащих разные соотношения кано
24% [13], находится в транскриптах, аннотиро
нических и альтернативных белков, а также,
ванных ранее как некодирующие. При этом но
возможно, различным процентом включения
вые транслируемые ОРС обнаруживаются как в
ложноположительных результатов. Подробное
транскриптах, функция которых ранее не была
сопоставление обсуждаемых частот использова
известной [18-24], так и в РНК, имеющих дру
ния кодонов для инициации трансляции можно
гую охарактеризованную функцию, например,
найти в отдельной работе [14].
белок TERP, закодированный в теломеразной
В третьих, неизвестные ранее белки и пеп
РНК [25], Humanin - в митохондриальной 16S
тиды могут быть закодированы в мРНК ранее
рРНК [26], E пептид - в бактериальной 23S
аннотированных генов так, что их рамки считы
рРНК [27]. Описаны случаи, когда новые функ
вания располагаются в 5′ нетранслируемой об
ции имеет и длинная некодирующая РНК, и ко
ласти (5′ НТО) или 3′ НТО, а также в рамке счи
дируемый ею белок. Например, РНК AW112010
тывания, частично или полностью перекрываю
кодирует небольшой белок, стимулирующий
щейся, но не совпадающей с ранее аннотиро
синтез IL 12p40. Мутантные мыши с нарушени
ванной. В этой группе наиболее часто встреча
ем синтеза пептида, закодированного в
ются ОРС в 5′ НТО - по разным данным, от
AW112010, демонстрируют бóльшую скорость
40 [9] до 68% [13] всех неаннотированных ранее
распространения сальмонелл и бóльшую бакте
ОРС. Такой высокий процент может объяснять
риемию, а также ускоренную смертность от
ся тем, что сам факт считывания 5′ ОРС, даже
сальмонеллеза [22]. В последующей работе об
если продукт этого процесса нефункционален,
наружена самостоятельная роль РНК AW112010,
БИОХИМИЯ том 86 вып. 9 2021
1368
СЕРГИЕВ, РУБЦОВА
заключающаяся в привлечении KDM5A и пос
ной литературе имеются сведения о других, бо
ледующем деметилировании остатков H3K4me3
лее тщательно исследованных малых белках, чья
гена, кодирующего IL 10 [28]. Сходным обра
функция в ядре связана с репликацией и репара
зом, после обнаружения функциональной роли
цией ДНК, а также с процессом транскрипции
пептида SPAR (другое название - SPAAR), зако
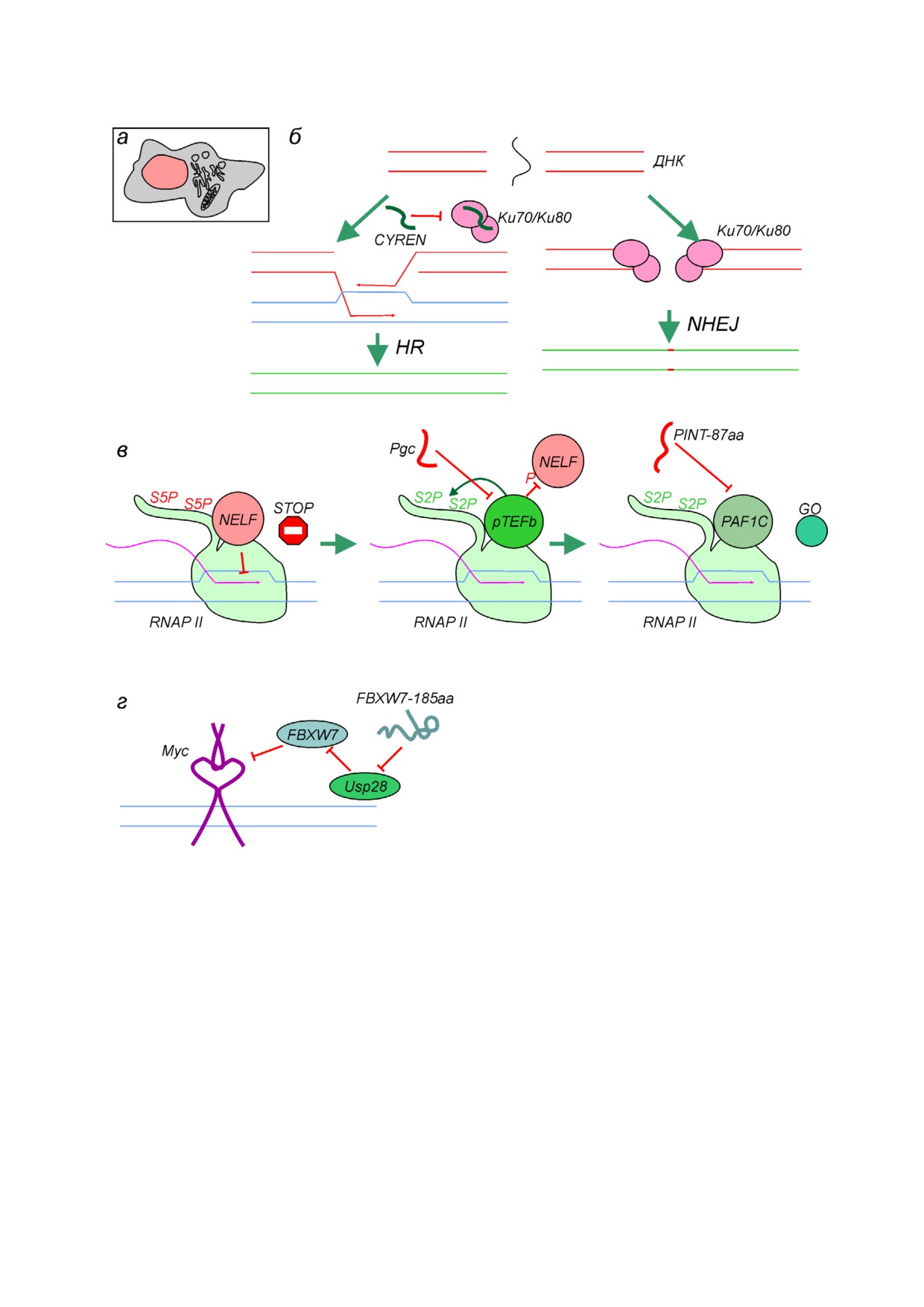
(рис. 1, а).
дированного в LINC00961, заключающейся в
Белок SHPRH 146aa, закодированный в
подавлении активности mTORC1 киназного
кольцевой РНК, проявляет онкосупрессорные
комплекса [24] (см. более подробное описание
свойства, способствуя деградации необходимо
ниже), выявлено, что LINC00961 как РНК имеет
го для пролиферации ядерного белка PCNA,
отдельную функцию в ангиогенезе, причем ее
обеспечивающего процессивность репликатив
активность противоположна таковой для пепти
ных ДНК полимераз [30]. AltMRVI1 длиной
да SPAR [29].
134 а.о. закодирован в 3′ НТО гена MRVI1. Про
Транслируемые ОРС находят в таких нео
дукт трансляции этой рамки имеет ядерную ло
бычных РНК, как кольцевые (circRNA), образу
кализацию и взаимодействует с BRCA1 [36],
емые в результате ошибочного сплайсин
белком онкосупрессором, участвующим в репа
га [30-32] (см. обзор [33]), и примикроРНК (pri
рации разрывов ДНК. Интересно, что ранее это
miRNA) [34]. В случае кольцевых РНК встает
взаимодействие определили с помощью дрож
вопрос о механизме инициации трансляции,
жевой двугибридной системы, но посчитали
поскольку классическая модель сканирования,
ложноположительным [37]. Кроме AltMRVI1, в
очевидно, для них не подходит. Авторы соответ
регуляции репарации двуцепочечных разрывов
ствующих работ объясняют взаимодействие ри
принимает участие пептид CYREN 2 [38], по су
босом с кольцевыми РНК участками внутрен
ти, являющийся короткой изоформой ранее ан
ней посадки рибосом (IRES, internal ribosome
нотированного белка CYREN/MRI (модулятор
entry site) и привлечением инициаторных фак
ретровирусной инфекции) [39], образующейся в
торов модифицированными остатками m6A;
результате альтернативного сплайсинга. Обе
предложенные модели требуют дальнейшей
формы CYREN взаимодействуют с Ku70/Ku80
тщательной проверки (см. рассуждения на эту
субъединицами ДНК зависимой протеинкина
тему в [35]). Таким образом, разнообразие коди
зы, участвующей в репарации двуцепочечных
рующих РНК значительно выходит за рамки на
разрывов ДНК по механизму негомологичного
ших обычных представлений. Это, с одной сто
соединения концов (NHEJ, nonhomologous end
роны, ошеломляет, а с другой стороны, вызыва
joining). Известно, что работа NHEJ может вы
ет определенную долю скепсиса.
зывать мутации, но не имеет альтернативы в
В чем же может заключаться функциональ
G1 фазе клеточного цикла, когда ДНК еще не
ная роль продуктов трансляции недавно откры
реплицирована с образованием сестринских
тых ОРС, малых и альтернативных белков? Хотя
хроматид. В поздней S и G2 фазах, когда дву
общего ответа на этот вопрос нет, мы рассмот
цепочечные разрывы ДНК могут быть исправ
рим несколько интересных частных случаев,
лены с помощью более точного механизма го
описанных в современной научной литературе.
мологичной рекомбинации (HR, homologous
recombination), активность NHEJ подавляется, в
том числе и с помощью прямого взаимодей
РОЛЬ МАЛЫХ БЕЛКОВ
ствия CYREN и Ku70/Ku80 (рис. 1, б). При
В БИОСИНТЕЗЕ ДНК И РНК
инактивации CYREN повышается частота слия
ния теломерных участков хромосом [38], а инак
Практически во всех экспериментах по ис
тивация CYREN/MRI вместе с одним из основ
следованию функциональной роли вновь от
ных участников системы NHEJ - XLF - приво
крываемых малых белков определяют их внут
дит к эмбриональной летальности мышей из за
риклеточную локализацию. Например, в работе
полного блокирования системы NHEJ [40].
Chen et al. [13] локализацию малых белков изу
Согласно нескольким недавним публикаци
чали с помощь их гибридов с пептидным «довес
ям, продукты трансляции малых рамок считы
ком» зеленого флуоресцентного белка GFP11,
вания участвуют в регуляции транскрипции.
который при взаимодействии с оставшейся
Например, пептид Pgc, состоящий из 71 а.о., по
частью GFP1 10 образует функциональный
давляет транскрипцию в предшественниках за
флуорофор, детекцию которого можно осущест
родышевых клеток дрозофилы, взаимодействуя
вить при помощи микроскопии. Так, например,
с киназой pTEFb (рис. 1, в). pTEFb осуществля
обнаружена ядерная локализация продукта
ет фосфорилирование белка NELF, а также ос
трансляции 5′ ОРС HMGA2. Хотя функциональ
татка Ser2 С концевого домена РНК полимера
ная роль этого пептида не определена, в науч
зы II [41]. Подобное фосфорилирование приво
БИОХИМИЯ том 86 вып. 9 2021
РАЗНООБРАЗИЕ ФУНКЦИЙ МАЛЫХ БЕЛКОВ И ПЕПТИДОВ
1369
Рис. 1. Функционирование малых белков в ядре, участие в процессах репарации ДНК и транскрипции. а - Схема строе
ния эукариотической клетки. Цветом выделено ядро, в котором осуществляется функционирование показанных на ри
сунке белков. б - Пептид CYREN ингибирует репарацию двуцепочечных разрывов ДНК по пути негомологичного соеди
нения концов ДНК (NHEJ, nonhomologous end joining), подавляя активность Ku70/Ku80 (на рисунке HR - homologous
recombination). в - Остановка РНК полимеразы II, вызванная белком NELF и обусловленная фосфорилированием остат
ков Ser5 (S5P) гептааминокислотных повторов С концевого домена (CTD) RNAP II (слева). Киназный комплекс pTEFb
фосфорилирует NELF и остатки Ser2 (S2P) гептааминокислотных повторов CTD RNAP II, что обусловливает связывание
PAF1C и переход к процессивной элонгации (по центру). Пептид Pgc ингибирует pTEFb, а пептид PINT 87aa подавляет
активность PAF1C, что приводит к репрессии транскрипции (справа). г - Влияние белка FBXW7 185 aa на стабильность
транскрипционного фактора Myc. FBXW7 185 aa конкурирует с FBXW7 за взаимодействие с Usp28 деубиквитиназой, что
приводит к повышению активности FBXW7 и дестабилизации Myc
дит к снятию ингибирования и увеличивает эф
ванный в кольцевой РНК, проявляет онкосуп
фективность транскрипции. После действия
рессорные свойства, взаимодействуя с PAF1c
pTEFb активность РНК полимеразы II увели
(рис. 1, в) и замедляя таким образом транскрип
чивается под действием другого фактора элонга
цию онкогенов [31]. Еще одним малым белком,
ции, PAF1c [42]. Пептид PINT 87aa, закодиро
взаимодействующим с транскрипционным фак
БИОХИМИЯ том 86 вып. 9 2021
1370
СЕРГИЕВ, РУБЦОВА
тором и проявляющим онкосупрессорные свой
Помимо участия в созревании мРНК, малые
ства, является FBXW7 185 aa [43]. FBXW7 185 aa
белки принимают участие в созревании рибо
конкурирует с полноразмерным FBXW7 за взаи
сом, одним из них является пептид MIEF1 MP/
модействие с Usp28 деубиквитиназой, стабили
L0R8F8 длиной 70 а.о. Этот пептид закодирован
зирующей транскрипционный фактор Myc. Та
в 5′ ОРС гена белка MID51. «Основной» про
ким образом, действие FBXW7 185 aa противо
дукт трансляции этой мРНК, MID51, локализу
положно активности FBXW7 и приводит к де
ется во внешней мембране митохондрий и регу
стабилизации Myc (рис. 1, г).
лирует их слияние [48]. Функциональность со
ответствующей
5′ ОРС, MIEF1/MP/L0R8F8,
предсказана на основе данных рибосомного
РОЛЬ МАЛЫХ БЕЛКОВ
профилирования и анализа консервативности
В СОЗРЕВАНИИ РНК, СИНТЕЗЕ
последовательности [16]. Дальнейшее исследо
И СВОРАЧИВАНИИ БЕЛКОВ
вание показало, что MIEF1 MP взаимодействует
с миторибосомами и способствует митохондри
В научной литературе можно найти множест
альной трансляции
[49]. Суперэкспрессия
во примеров регуляции созревания мРНК и
MIEF1 MP повышает, а нокдаун - снижает эф
трансляции с помощью малых белков (рис. 2, а).
фективность митохондриальной трансляции
Некоторые из этих исследований демонстриру
практически всех белков, синтезируемых мито
ют пока довольно поверхностные данные. Так,
рибосомами. Примечательно, что уровень
описано взаимодействие белков altSETD1B и
трансляции MIEF1 превышает таковой для ос
altLINC00624 с аминоацил тРНК синтетазами
новной аннотированной рамки этого гена, бел
[44], однако функциональные последствия этих
ка MID51 [50]. Пространственное расположе
взаимодействий исследованы не были. Значи
ние пептида MIEF1 MP/L0R8F8 в структуре
тельно более подробно исследованы функции
интермедиата сборки миторибосомы определе
других малых белков в процессинге РНК и био
но в лаборатории В. Рамакришнана с помощью
синтезе белка. Пептид, состоящий из 53 а.о., зако
криоэлектронной микроскопии [51]. В этой
дированный в транскрипте HOXB AS3, взаимо
структуре MIEF1 MP соединяет гомолог бакте
действует с hnRNP A1 и рядом других факторов
риального фактора сборки рибосом RsfS,
процессинга РНК [45]. Экспрессия HOXB AS3
MALSU1, и митохондриальный носитель жир
падает в клетках раковых опухолей, например,
ных кислот mt ACP. Предположительно, эта
при раке прямой кишки. Подобное снижение
белковая пирамидка предотвращает преждевре
экспрессии предвещает плохой прогноз для па
менное взаимодействие интермедиата сборки
циентов. Инактивация рамки считывания при
большой субъединицы миторибосомы с малой
сохранении большей части РНК HOXB AS3 при
субъединицей (рис. 2, в).
водит к повышению колониеобразования, сти
Взаимодействие белка altATAD2 длиной
муляции роста и миграции клеток. Эктопичес
139 а.о. с цитоплазматическим рибосомным
кая экспрессия данного гена в клетках рака пря
белком RPL10 обнаружено с помощью приши
мой кишки, соответственно, наоборот, замедля
вок с последующей масс спектрометрической
ет пролиферацию клеток. Онкосупрессорная
идентификацией [52]. Помимо взаимодействия
природа HOXB AS3 объясняется его антагониз
с рибосомным белком, обнаружен контакт
мом с hnRNP A1. Пептид HOXB AS3 «отменяет»
altATAD2 с AUF1/hnRNP D, узнающим AU
включение экзона 10 в мРНК пируваткиназы,
богатые последовательности РНК. Как предпо
обусловленное активностью hnRNP A1. Феталь
лагают исследователи, altATAD2 может служить
ная сплайс форма пируваткиназы содержит эк
посредником в привлечении рибосом на мРНК,
зон 10 и участвует в аэробном гликолизе, в то
имеющие AU богатые нуклеотидные последо
время как сплайс форма, содержащая экзон 9,
вательности, хотя молекулярный механизм
экспрессируется в тканях взрослого организма и
функционирования altATAD2 пока так и остает
участвует в окислительном фосфорилировании.
ся гипотетическим (рис. 2, г).
Раковые клетки, возвращаясь к гликолитичес
Деградация мРНК, несущих AU богатые
кому метаболизму в результате эффекта Варбур
последовательности и другие дестабилизирую
га, начинают экспрессировать фетальную фор
щие сигналы, происходит в специализирован
му пируваткиназы [46], что обусловлено актив
ных компартментах цитоплазмы, называемых
ностью hnRNP A1 [47]. Пептид HOXB AS3 по
P тельцами (P bodies). В этих структурах лока
давляет эффект Варбурга и рост клеток опухоли,
лизован продукт трансляции LINC01420 - пеп
являясь антагонистом hnRNP A1 и способствуя
тид Nobody длиной 68 а.о. Анализ белок белко
экспрессии пируваткиназы с включенным экзо
вых взаимодействий выявил его ассоциацию с
ном 9 [45] (рис. 2, б).
аппаратом декэпирования, причем прямое взаи
БИОХИМИЯ том 86 вып. 9 2021
РАЗНООБРАЗИЕ ФУНКЦИЙ МАЛЫХ БЕЛКОВ И ПЕПТИДОВ
1371
Рис. 2. Функционирование малых белков в процессах созревания, трансляции и деградации РНК. а - Схема локализации
процессов созревания, трансляции и деградации РНК в клетке. б - Регулируемый альтернативный сплайсинг пре мРНК
пируваткиназы. В эмбриональных и раковых клетках повышенная концентрация hnRNP A1 подавляет включение экзо
на 9 и способствует включению экзона 10, что приводит к синтезу PKM2 изоформы. В большинстве клеток взрослого ор
ганизма пептид HOXB AS3 подавляет активность hnRNP A1, что приводит к включению экзона 9, но не 10 - и появле
нию изоформы PKM1, способствующей окислительному фосфорилированию. в - Пептид MIEF1 вместе с MALSU1 и
mt ACP взаимодействует с интермедиатом сборки большой субчастицы (39S) миторибосомы, чтобы не допустить преж
девременного взаимодействия с малой (28S) субчастицей. г - Белок altATAD2 взаимодействует с AUF1, узнающим мРНК
с AU богатыми последовательностями (ARE), и рибосомным белком RPL10 (на рисунке m7G - 5′ кэп). д - Пептид
Nobody напрямую взаимодействует с EDC4 и опосредованно - с другими компонентами комплекса, декэпирующего
мРНК, в результате чего подавляет их активность
модействие Nobody с EDC4 важно для опосре
которые исчезают при его суперэкспрессии
дованного сближения этого пептида с EDC3,
(рис. 2, д). Одновременно с исчезновением Р телец
Dcp1A, Dcp1B и Dcp2. При этом Nobody являет
при суперэкспрессии Nobody в клетке возраста
ся отрицательным регулятором сборки P телец,
ет количество мРНК, являющихся субстратом
БИОХИМИЯ том 86 вып. 9 2021
1372
СЕРГИЕВ, РУБЦОВА
нонсенс опосредованной деградации (NMD,
ся в различные мембранные структуры клетки
nonsense mediated mRNA decay) [53].
и/или взаимодействует с мембранными белко
После синтеза белки часто становятся
выми комплексами (рис. 3, а). Например, в ходе
субстратами шаперонов, которые способствуют
скринингового исследования функциональной
формированию правильной пространственной
роли и белковых партнеров малых белков выяв
структуры. Роль малых белков в этом процессе
лено, что пептид, закодированный в LINC00998,
также имеет существенное значение. Так, не
совыделяется с группой белков эндоплазмати
давно определена функциональная роль пепти
ческого ретикулума, а инактивация этого гена
да, закодированного в 5′ ОРС мРНК ASNSD1,
приводит к изменению, по видимому, компен
названного ASDURF [54]. Как оказалось, этот
саторному, экспрессии генов, имеющих отно
пептид длиной 96 а.о. является компонентом
шение к процессу гликозилирования, также
комплекса шаперонов PAQсомы, похожего на
происходящему в данном клеточном компарт
комплекс префолдинов. Наличие ASDURF и
менте [13]. Согласно той же работе [13], пептид
его взаимодействие с другими компонентами
длиной 70 а.о., закодированный в гене RP11
PAQсомы объясняет существовавшее ранее
84A1.3, локализуется в цитоплазматической
противоречие в предполагаемом белковом сос
мембране и взаимодействует с группой белков
таве PAQсомы - тогда были известны лишь
той же локализации.
5 субъединиц этого комплекса, при том, что
Пептид длиной 54 а.о. закодирован в РНК,
сходные структуры содержат, как правило,
транскрибирующейся с цепи ДНК, комплемен
6
субъединиц. Как компонент PAQсомы,
тарной гену PIBG, в связи с чем получил курьез
ASDURF принимает участие в формировании
ное название PIGBOS [21]. Исследование этого
структур мяРНП и компонентов сигнального
пептида показало, что он локализуется на внеш
пути mTOR.
ней мембране митохондрии и взаимодействует с
белком CLCC1, расположенном в эндоплазма
тическом ретикулуме (ЭР), в местах контакта
МАЛЫЕ БЕЛКИ В СИСТЕМЕ
ЭР с митохондриями. Взаимодействие с CLCC1
ВЕЗИКУЛЯРНОГО ТРАНСПОРТА
важно для функционирования PIGBOS. Нокаут
PIGBOS, как и нокдаун CLCC1, приводит к усу
Значительная доля малых белков и пептидов
гублению стресса ЭР, ответу развернутого белка
имеет трансмембранные сегменты и встраивает (UPR, unfolded protein response) при обработке
Рис. 3. Функционирование малых белков в системе везикулярного транспорта. а - Схема клетки с выделением органелл,
показанных на следующих панелях. б - Взаимодействие пептида PIGBOS, локализованного во внешней митохондриаль
ной мембране в местах контакта эндоплазматического ретикулума (ЭР) и митохондрии с белком ЭР CLCC1. Данное взаи
модействие предотвращает стресс ЭР (UPR, unfolded protein response). в - Белок аltRPL36, локализованный в местах кон
такта ЭР и цитоплазматической мембраны, взаимодействует с TMEM24 и ингибирует перенос PI(4,5)P2 из ЭР на внеш
нюю мембрану. Таким образом, это ингибирование приводит к активации mTOR киназы через образование PI(3,4,5)P3
(фосфатидилинозитолтрифосфата) киназой PI3K. г - Расположенный в мембране эндосом пептид CASIMO1 стимулиру
ет сквален эпоксидазу SQLE, способствуя повышенному синтезу различных стероидных соединений. д - Пептиды фос
фоламбан (PLN), сарколипин (SLN) и миорегулин (MLN), локализованные в саркоплазматическом ретикулуме (СР) раз
личных типов мышц, подавляют активность ATP зависимой кальциевой помпы SERCA
БИОХИМИЯ том 86 вып. 9 2021
РАЗНООБРАЗИЕ ФУНКЦИЙ МАЛЫХ БЕЛКОВ И ПЕПТИДОВ
1373
туникамицином и усилению апоптоза, вызван
способствовать такого рода термогенезу [60].
ного этим стрессом (рис. 3, б).
Миорегулин (MLN), пептид длиной 46 а.о., взаи
Белок altRPL36 закодирован в мРНК рибо
модействует с SERCA в мышцах всех типов. Так
сомного белка RPL36, причем область, кодиру
же как сарколипин и фосфоламбан, миорегулин
ющая альтернативный белок, начинается рань
содержит трансмембранную альфа спираль в
ше, а заканчивается позже «основной» ОРС.
N концевой части молекулы. Взаимодействие
Поскольку эти кодирующие области сдвинуты
миорегулина с насосом SERCA отрицательно
друг относительно друга со сдвигом рамки счи
регулирует активность последнего. Инактива
тывания (на некратное трем число нуклеотидов),
ция гена миорегулина приводит к более эффек
соответствующие аминокислотные последова
тивному закачиванию ионов кальция в саркоп
тельности не похожи друг на друга. В отличие от
лазматический ретикулум, что стимулирует ра
RPL36, являющегося компонентом рибосомы,
боту мышц [23] (рис. 3, д).
altRPL36 взаимодействует с трансмембранным
Помимо функционирования пептидов и ма
белком ЭР TMEM24, локализованным в местах
лых белков в различных компартментах систе
контактов ЭР и цитоплазматической мембраны.
мы везикулярного транспорта, известны приме
AltRPL36 ингибирует активность TMEM24, нап
ры пептидов/малых белков, экспортируемых че
равленную на транспорт фосфатидилинозитол
рез эту систему во внеклеточное пространство.
дифосфата, PI(4,5)P2, из ЭР во внешнюю мемб
Так, пептид Humanin длиной 24 а.о. закодиро
рану. Функция как altRPL36, так и TMEM24 со
ван в гене 16S митохондриальной рРНК. Учиты
стоит в регулировании фосфорилирования.
вая, что митохондриальная рРНК содержит
Инактивация аltRPL36 приводит к активации
множество гипотетических рамок считывания,
TMEM24, увеличению концентрации субстрата
предшествующих рамке, кодирующей Humanin,
киназы PI3K, PI(4,5)P2, и, таким образом, к ак
маловероятно, чтобы считывание этой рамки
тивации mTOR сигнализации (рис. 3, в).
считывания происходило в ходе трансляции в
В эндосомах локализован пептид длиной
митохондриях. Возможно, экспрессия этого ге
83 а.о., CASIMO1, который является продуктом
на может осуществляться с копий митохондри
трансляции рамки считывания, находящейся в
альной ДНК, присутствующих в ядерном гено
небольшой РНК NR_029455. Экспрессия этого
ме в виде псевдогенов. Согласно оригинальной
гена возрастает в клетках рака молочной железы
статье [26], продукт экспрессии этого гена сек
с высоким уровнем экспрессии ядерных рецеп
ретируется в межклеточную среду и подавляет
торов стероидных гормонов ER/PR [55]. Инак
токсичность бета амилоида, основного агента
тивация кодирующего CASIMO1 гена замедля
патогенеза болезни Альцгеймера, в отношении
ла, а повышенная экспрессия - ускоряла рост
культуры нейронов. В последующие годы, не
клеток. Эксперименты по коиммунопреципита
смотря на открытые вопросы о механизме
ции продемонстрировали, что CASIMO1 взаи
экспрессии, у пептида обнаружили ряд других
модействует с другим онкогенным белком, сква
«чудодейственных» свойств в отношении остео
лен эпоксидазой SQLE, и стимулирует ее актив
пороза, сердечно сосудистых заболеваний, диа
ность. Повышенную экспрессию SQLE также
бета и рака, наряду с первоначально обнаружен
наблюдают в клетках рака молочной железы. По
ной активностью в отношении нейродегенера
всей видимости, функция CASIMO1 заключает
тивных заболеваний (для обзора см. [61]).
ся в стимуляции активности сквален эпоксида
зы (рис. 3, г).
Группа малых белков встречается в специа
МАЛЫЕ БЕЛКИ МИТОХОНДРИЙ
лизированном типе ЭР, саркоплазматическом
ретикулуме, клеток различных типов мышц.
В протеоме митохондрий (рис. 4, а) значи
Пептиды фосфоламбан (PLN) [56, 57] длиной
тельную долю (до 5%) составляют малые белки
52 а.о. и сарколипин (SLN) [58] длиной 31 а.о.
длиной до 100 а.о. [62]. Выше мы уже обсуждали
присутствуют в сердечной мышце и медленных
функциональную роль недавно открытого ми
мышцах соответственно. Их активность направ
тохондриального белка MIEF1/L0R8F8, участ
лена на ингибирование ATP зависимой кальци
вующего в сборке митохондриальных рибо
евой помпы саркоплазматического ретикулума
сом [49, 51]. Более широко в научной литерату
SERCA (для обзора см., например, [59]), работа
ре представлены недавно открытые пептиды,
которой способствует расслаблению мышц пос
влияющие на работу комплексов дыхательной
ле сокращения. SERCA также может генериро
цепи митохондрий. Например, выполненное в
вать тепло при ингибировании транспорта ио
лаборатории Дж. Вейсмана исследование мно
нов кальция без снижения ее ATPазной актив
жества малых ОРС выявило митохондриальную
ности. SLN, взаимодействуя с SERCA, может
локализацию продукта трансляции РНК
БИОХИМИЯ том 86 вып. 9 2021
1374
СЕРГИЕВ, РУБЦОВА
Рис. 4. Функционирование малых белков в мембранах митохондрий. а - Схема клетки с указанием митохондрий и схема
работы окислительного фосфорилирования. Показаны комплексы дыхательной цепи I-IV (СI-CIV), комплекс V (CV)
ATP синтазы, цепь переноса восстановительных эквивалентов и направление переноса протонов. б - Функционирова
ние недавно открытых митохондриальных пептидов. Показано взаимодействие RP11_469A15.2 с цитохром С оксидазой,
стимуляция цитохром С редуктазы пептидом BRAWNIN и взаимодействие пептида MTLN с цитохром B5 редуктазой 3
(CYB5R3) и компонентами трифункционального комплекса окисления жирных кислот HADHA/B. Активность MTLN
влияет на липидный состав мембран и эффективность дыхания
RP11_469A15.2, ранее считавшейся некодирую
ма, соответствует дефектам функционирования
щей [13]. Соответствующий пептид длиной
CIII (рис. 4б).
62 а.о. взаимодействует с комплексом IV дыха
В нашей лаборатории [18], как и в несколь
тельной цепи, цитохром С оксидазой. Молеку
ких других [19, 20], была исследована функция
лярный механизм функционирования данного
малого белка митохондрий миторегулина
пептида остается пока неизвестным (рис. 4, б).
(MTLN), закодированного в транскрипте, ра
Более подробно исследована функция еще
нее аннотированном как длинная некодирую
одного недавно открытого митохондриального
щая РНК LINC00116. Присутствие MTLN про
пептида, получившего название BRAWNIN.
демонстрировано в клетках человека [19, 20],
Этот пептид состоит из 71 а.о., консервативен у
мыши [18, 64] и рыбы D. rerio [65]. Последова
позвоночных и локализуется во внутренней
тельность этого белка длиной 56 а.о. весьма
мембране митохондрий [63]. Экспрессия гена
консервативна у позвоночных и содержит
этого пептида находится под контролем основ
трансмембранный сегмент, заякоривающий
ного транскрипционного фактора, отвечающего
MTLN во внутренней митохондриальной мемб
за биогенез митохондрий, PGC1a, и регулирует
ране [20]. Данные о партнерах MTLN, получен
ся киназой AMPK, активирующейся в условиях
ные в разных лабораториях, несколько отлича
нехватки ATP. У рыб Danio rerio с инактивиро
ются друг от друга. Так, в работах Makarewich
ванным геном, кодирующим BRAWNIN, наб
et al. [20] и Friesen et al. [66] продемонстрирова
людается лактазный ацидоз, характерный для
но взаимодействие MTLN с компонентами
многих митохондриальных заболеваний. Как
комплекса β окисления жирных кислот
показали опыты по коиммунопреципитации,
HADHA и HADHB, а в последней работе -
BRAWNIN взаимодействует с комплексом III
также и взаимодействие с ATP синтазой. Наши
(СIII) дыхательной цепи, цитохром С редукта
эксперименты по коиммунопреципитации вы
зой. Фенотипическое проявление инактивации
явили взаимодействие MTLN с цитохром B5 ре
гена этого пептида, согласно анализу метаболо
дуктазой, CYB5R3. Практически все исследова
БИОХИМИЯ том 86 вып. 9 2021
РАЗНООБРАЗИЕ ФУНКЦИЙ МАЛЫХ БЕЛКОВ И ПЕПТИДОВ
1375
тели сходятся в том, что MTLN способствует
В РНК LINC00961, изначально аннотиро
работе комплексов респираторной цепи, одна
ванной как длинная некодирующая, обнаруже
ко, опять же, нюансы этого влияния видятся
на ОРС, соответствующая пептиду длиной
ими по разному. В нашей работе [18] и работе
75-90 а.о. в зависимости от используемого
Lin et al. [64] было продемонстрировано сниже
старт кодона. Этот пептид, названный SPAR,
ние эффективности работы комплекса I (CI)
имеет трансмембранный домен, с помощью ко
дыхательной цепи, NADH дегидрогеназы. По
торого он взаимодействует с мембраной позд
результатам других исследований было выдви
них эндосом и лизосом [24]. Аффинное совыде
нуто предположение, что наблюдаемый при
ление показало, что SPAR взаимодействует с
инактивации MTLN дефект функционирова
v ATPазой, протонным насосом, отвечающим
ния дыхательной цепи проявляется только при
за понижение pH в эндосомах и лизосомах. Ока
окислении пальмитоилкарнитина, субстрата
залось, что SPAR ингибирует активацию
системы β окисления жирных кислот
[19,
mTORC1 киназного комплекса, привлекаемого
20, 66]. Хотя мы и наблюдали снижение актив
к мембране лизосомы комплексом v ATPазы,
ности CI комплекса дыхательной цепи, это яв
Ragulator и GTPаз семейства Rag при экспорте
ление проявлялось только на уровне пермеаби
аминокислот из лизосомы в цитозоль. Этот эф
лизованных клеток и выделенных митохонд
фект проявлялся при увеличении концентрации
рий, но не очищенного CI. Поэтому мы сделали
аминокислот после голодания. В клетках с из
вывод о том, что MTLN не является ни компо
быточной экспрессией LINC00961, кодирующей
нентом CI, ни фактором его сборки. Поскольку
SPAR, mTORC1 киназный комплекс не мог свя
известно, что работа CI сильно зависит от ли
заться с мембраной лизосом и оставался диф
пидного состава мембран, а партнер MTLN,
фузно локализованным в цитозоле. Активация
CYB5R3, участвует в метаболизме липидов, мы
mTORC1 при помощи внешнего сигнального
изучили изменения липидного состава клеток с
пути от рецепторных тирозинкиназ через PI3K
инактивированным геном, кодирующим
не зависит от SPAR [24]. Поскольку известно,
MTLN. Оказалось, что отсутствие функцио
что активация mTOR пути способствует восста
нального миторегулина действительно снижало
новлению мышц после повреждения, у мышей с
количество необходимого для работы CI фос
инактивированным геном 5430416O09Rik, коди
фатидилхолина и повышало содержание в клет
рующим SPAR, была протестирована способ
ке триглицеридов. Таким образом, функцио
ность мышц к регенерации после повреждения
нальная роль MTLN связана, на наш взгляд, с
кардиотоксином. Оказалось, что мышцы мы
формированием липидного состава клеточных
шей без SPAR восстанавливаются быстрее, что
мембран посредством взаимодействия с
вполне укладывается в модель гиперстимуляции
CYB5R3. В свою очередь, правильная работа
mTOR пути в отсутствие данного ингибиторно
данной системы обеспечивает работу комплек
го пептида [24]. Интересно отметить, что в пос
са CI дыхательной цепи за счет благоприятного
ледующих работах также обнаружены отдель
липидного окружения (рис. 4, б).
ные, не связанные с кодированием SPAR, функ
ции РНК LINC00961 [29] (рис. 5, а).
Регулятором метаболического ответа оказал
МАЛЫЕ БЕЛКИ В РЕГУЛЯЦИИ
ся малый белок hTERP (human TERP), продукт
МЕТАБОЛИЗМА
трансляции предшественника теломеразной
РНК человека [25]. В процессе транскрипции
Несколько малых белков вовлечено в сиг
гена теломеразной РНК, регулируемой мульти
нальные пути, регулирующие метаболизм.
субъединичным комплексом Integrator [68],
Ключевая роль в этих сигнальных путях отво
синтезируется удлиненный предшественник,
дится киназным комплексам mTOR, стимуля
содержащий открытую рамку считывания,
ция которых приводит к более активному био
трансляция которой приводит к образованию
синтезу белков, а ингибирование - к усилению
белка длиной 121 а.о. Оказалось, что белок
аутофагии, регулируемому распаду компонен
hTERP способствует выживанию клеток в усло
тов клетки (см. [67] в качестве актуального об
виях индукции апоптоза, а мутации в нем при
зора). Выше упомянуто, что белок altRPL36 яв
водят к нарушениям протекания аутофагии
ляется негативным регулятором mTOR пути,
(рис. 5, б). В отсутствие белка hTERP происхо
поскольку ингибирует белок TMEM24, кото
дит активация киназы p70S6K1 (неопублико
рый транспортирует PI(4,5)P2. В свою очередь,
ванные данные), что согласуется с данными, по
PI(4,5)P2 является субстратом киназы PI3K, ак
лученными при изучении мышей, в организме
тивирующей mTOR через несколько белков
которых произведена делеция гена теломераз
посредников [7].
ной РНК [69].
БИОХИМИЯ том 86 вып. 9 2021
1376
СЕРГИЕВ, РУБЦОВА
Рис. 5. Функционирование малых белков в регуляции аутофагии и биосинтеза белка. а - Пептид SPAR, взаимодействую
щий с протонной помпой лизосомы, подавляет активацию киназы mTOR аминокислотами, экспортируемыми из лизосо
мы. При этом SPAR не влияет на активацию mTOR через сигнальный путь от рецепторных тирозинкиназ. б - Белок
hTERP, закодированный в теломеразной РНК, участвует в регуляции переключения метаболизма клетки, воздействуя на
аутофагию и биосинтез белка, а также подавляет апоптоз (подробнее - в тексте)
Участие белка hTERP в модуляции киназных
ства продуктов трансляции коротких рамок счи
каскадов, регулирующих метаболизм клеток,
тывания затрудняют их идентификацию и изу
приоткрывает неизведанный уровень стимуля
чение функциональной роли. Развитие методов
ции пролиферации, ассоциированный как пра
исследования позволило открыть новый уро
вило с активацией теломеразного комплекса, од
вень регуляции внутриклеточных процессов при
ним из основных компонентов которого являет
помощи пептидов и малых белков. Исследова
ся теломеразная РНК. Известно, что стимуляция
ния последних лет демонстрируют участие про
клеток иммунной системы, онкотрансформация
дуктов трансляции малых рамок считывания во
клеток и другие процессы, требующие повышен
всех процессах функционирования клеток, а ис
ной скорости пролиферации и, следовательно,
тинное количество кодируемых ими белковых
ресурсов, сопровождаются переключением ме
продуктов трудно оценить. Постоянно появля
таболической программы, а также повышением
ющиеся публикации, демонстрирующие разно
активности теломеразы. Для активации теломе
образные функции продуктов трансляции ко
разного комплекса требуется повышенная
ротких рамок считывания, приоткрывают до сих
экспрессия гена обратной транскриптазы, а так
пор нераскрытый мощный регуляторный по
же эффективный процессинг теломеразной
тенциал «серой материи» протеома. Исследова
РНК с образованием формы, ассоциированной с
ния, направленные на идентификацию и выяс
теломеразным комплексом. Можно предполо
нение роли компонентов «альтернативного»
жить, что выбор одного из двух путей экспрес
протеома, абсолютно необходимы для получе
сии гена теломеразной РНК должен зависеть от
ния полной картины регуляторных событий, ле
метаболического состояния клеток и скорости
жащих в основе как поддержания гомеостаза
их пролиферации, а также влиять на эти процес
клетки, так и развития патологических процес
сы. Таким образом, белок hTERP может функ
сов.
ционировать либо как регулятор метаболичес
кой программы клетки, либо как сенсор пере
ключения окислительного фосфорилирования и
Финансирование. Работа выполнена при фи
гликолиза. Роль белка TERP в регуляции мета
нансовой поддержке Российского научного
болизма клеток требует дальнейшего изучения.
фонда (грант № 21 64 00006).
Конфликт интересов. Авторы заявляют об от
сутствии конфликта интересов.
ЗАКЛЮЧЕНИЕ
Соблюдение этических норм. Настоящая
статья не содержит описания каких либо иссле
Малый размер, низкая консервативность
дований с участием людей или животных в каче
последовательностей и незначительные количе стве объектов.
БИОХИМИЯ том 86 вып. 9 2021
РАЗНООБРАЗИЕ ФУНКЦИЙ МАЛЫХ БЕЛКОВ И ПЕПТИДОВ
1377
СПИСОК ЛИТЕРАТУРЫ
1.
Numata, K., Kanai, A., Saito, R., Kondo, S., Adachi, J.,
complexes and respiratory efficiency, Cell Rep., 23, 3710
et al. (2003) Identification of putative noncoding RNAs
3720.e8.
among the RIKEN mouse full length cDNA collection,
20.
Makarewich, C. A., Baskin, K. K., Munir, A. Z.,
Genome Res., 13, 1301 1306.
Bezprozvannaya, S., Sharma, G., et al. (2018) MOXI is a
2.
Okazaki, Y., Furuno, M., Kasukawa, T., Adachi, J.,
mitochondrial micropeptide that enhances fatty acid β
Bono, H., et al. (2002) Analysis of the mouse transcrip
oxidation, Cell Rep., 23, 3701 3709.
tome based on functional annotation of 60,770 full length
21.
Chu, Q., Martinez, T. F., Novak, S. W., Donaldson, C. J.,
cDNAs, Nature, 420, 563 573.
Tan, D., et al. (2019) Regulation of the ER stress response
3.
Wilhelm, B. T., Marguerat, S., Watt, S., Schubert, F.,
by a mitochondrial microprotein, Nat. Commun., 10, 4883.
Wood, V., et al. (2008) Dynamic repertoire of a eukaryotic
22.
Jackson, R., Kroehling, L., Khitun, A., Bailis, W.,
transcriptome surveyed at single nucleotide resolution,
Jarret, A., et al. (2018) The translation of non canonical
Nature, 453, 1239 1243.
open reading frames controls mucosal immunity, Nature,
4.
Ulitsky, I., and Bartel, D. P. (2013) lincRNAs: Genomics,
564, 434 438.
evolution, and mechanisms, Cell, 154, 26 46.
23.
Anderson, D. M., Anderson, K. M., Chang, C. L.,
5.
Kopp, F., and Mendell, J. T. (2018) Functional classifica
Makarewich, C. A., Nelson, B. R., et al. (2015) A
tion and experimental dissection of long noncoding RNAs,
micropeptide encoded by a putative long noncoding rna
Cell, 172, 393 407.
regulates muscle performance, Cell, 160, 595 606.
6.
Ingolia, N. T., Ghaemmaghami, S., Newman, J. R. S., and
24.
Matsumoto, A., Pasut, A., Matsumoto, M., Yamashita, R.,
Weissman, J. S. (2009) Genome wide analysis in vivo of
Fung, J., et al. (2017) mTORC1 and muscle regeneration
translation with nucleotide resolution using ribosome pro
are regulated by the LINC00961 encoded SPAR polypep
filing, Science, 324, 218 223.
tide, Nature, 541, 228 232.
7.
Cao, X., Khitun, A., Na, Z., Dumitrescu, D. G.,
25.
Rubtsova, M., Naraykina, Y., Vasilkova, D., Meerson, M.,
Kubica, M., et al. (2020) Comparative proteomic profiling
Zvereva, M., et al. (2018) Protein encoded in human
of unannotated microproteins and alternative proteins in
telomerase RNA is involved in cell protective pathways,
human cell lines, J. Proteome Res., 19, 3418 3426.
Nucleic Acids Res., 46, 8966 8977.
8.
Ma, J., Ward, C. C., Jungreis, I., Slavoff, S. A., Schwaid,
26.
Hashimoto, Y., Niikura, T., Tajima, H., Yasukawa, T.,
A. G., et al. (2014) Discovery of human sORF encoded
Sudo, H., et al. (2001) A rescue factor abolishing neuronal
polypeptides (SEPs) in cell lines and tissue, J. Proteome
cell death by a wide spectrum of familial Alzheimer’s dis
Res., 13, 1757 1765.
ease genes and A, Proc. Natl. Acad. Sci. USA, 98, 6336
9.
Slavoff, S. A., Mitchell, A. J., Schwaid, A. G., Cabili,
6341.
M. N., Ma, J., et al. (2013) Peptidomic discovery of short
27.
Tenson, T., DeBlasio, A., and Mankin, A. (1996) A func
open reading frame encoded peptides in human cells, Nat.
tional peptide encoded in the Escherichia coli 23S rRNA,
Chem. Biol., 9, 59 64.
Proc. Natl. Acad. Sci. USA, 93, 5641 5646.
10.
Cardon, T., Hervé, F., Delcourt, V., Roucou, X.,
28.
Yang, X., Bam, M., Becker, W., Nagarkatti, P. S., and
Salzet, M., et al. (2020) Optimized sample preparation
Nagarkatti, M. (2020) Long Noncoding RNA AW112010
workflow for improved identification of ghost proteins,
Promotes the differentiation of inflammatory T Cells by
Anal. Chem., 92, 1122 1129.
suppressing IL 10 expression through histone demethyla
11.
Cardon, T., Fournier, I., and Salzet, M. (2021) Shedding
tion, J. Immunol., 205, 987 993.
light on the ghost proteome, Trends Biochem. Sci., 46, 239
29.
Spencer, H. L., Sanders, R., Boulberdaa, M., Meloni, M.,
250.
Cochrane, A., et al. (2020) The LINC00961 transcript and
12.
Chugunova, A., Navalayeu, T., Dontsova, O., and
its encoded micropeptide, small regulatory polypeptide of
Sergiev, P. (2018) Mining for small translated ORFs,
amino acid response, regulate endothelial cell function,
J. Proteome Res., 17, 1 11.
Cardiovasc. Res., 116, 1981 1994.
13.
Chen, J., Brunner, A. D., Cogan, J. Z., Nuñez, J. K., et al.
30.
Zhang, M., Huang, N., Yang, X., Luo, J., Yan, S., et al.
(2020) Pervasive functional translation of noncanonical
(2018) A novel protein encoded by the circular form of the
human open reading frames, Science, 367, 1140 1146.
SHPRH gene suppresses glioma tumorigenesis, Oncogene,
14.
Cao, X., and Slavoff, S. A. (2020) Non AUG start codons:
37, 1805 1814.
expanding and regulating the small and alternative
31.
Zhang, M., Zhao, K., Xu, X., Yang, Y., Yan, S., et al.
ORFeome, Exper. Cell Res., 391, 111973.
(2018) A peptide encoded by circular form of LINC PINT
15.
Hinnebusch, A. G. (1997) Translational regulation of yeast
suppresses oncogenic transcriptional elongation in
GCN4, J. Biol. Chem., 272, 21661 21664.
glioblastoma, Nat. Commun., 9, 4475.
16.
Andreev, D. E., O’Connor, P. B., Fahey, C., Kenny, E. M.,
32.
Yin, H., Shen, X., Zhao, J., Cao, X., He, H., et al. (2020)
Terenin, I. M., et al. (2015) Translation of 5′ leaders is per
Circular RNA CircFAM188B encodes a protein that regu
vasive in genes resistant to eIF2 repression, eLife, 4,
lates proliferation and differentiation of chicken skeletal
e03971.
muscle satellite cells, Front. Cell Dev. Biol., 8, 522588.
17.
Michel, A. M., Andreev, D. E., and Baranov, P. V. (2014)
33.
Wang, J., Zhu, S., Meng, N., He, Y., Lu, R., and Yan, G.
Computational approach for calculating the probability of
R. (2019) ncRNA encoded peptides or proteins and can
eukaryotic translation initiation from ribo seq data that
cer, Mol. Ther., 27, 1718 1725.
takes into account leaky scanning, BMC Bioinformatics, 15,
34.
Fang, J., Morsalin, S., Rao, V. N., and Reddy, E. S. P.
380.
(2017) Decoding of non coding DNA and non coding
18.
Chugunova, A., Loseva, E., Mazin, P., Mitina, A.,
RNA: Pri micro RNA encoded novel peptides regulate
Navalayeu, T., et al. (2019) LINC00116 codes for a mito
migration of cancer cells, J. Pharmaceut. Sci. Pharmacol.,
chondrial peptide linking respiration and lipid metabolism,
3, 23 27.
Proc. Natl. Acad. Sci. USA, 116, 4940 4945.
35.
Shi, Y., Jia, X., and Xu, J. (2020) The new function of
19.
Stein, C. S., Jadiya, P., Zhang, X., McLendon, J. M.,
circRNA: translation, Clin. Transl. Oncol., 22, 2162 2169.
Abouassaly, G. M., et al. (2018) Mitoregulin: a lncRNA
36.
Vanderperre, B., Lucier, J. F., Bissonnette, C., Motard, J.,
encoded microprotein that supports Mitochondrial super
Tremblay, G., et al. (2013) Direct detection of alternative
9 БИОХИМИЯ том 86 вып. 9 2021
1378
СЕРГИЕВ, РУБЦОВА
open reading frames translation products in human signif
ghost proteins involved in proteins translation. Biochim.
icantly expands the proteome, PLoS One, 8, e70698.
Biophys. Acta Gen. Subjects, 1863, 1458 1470.
37.
Liu, Y., Woods, N. T., Kim, D., Sweet, M., Monteiro,
53.
D’Lima, N. G., Ma, J., Winkler, L., Chu, Q., Loh, K. H.,
A. N. A., and Karchin, R. (2011) Yeast two hybrid junk
et al. (2017) A human microprotein that interacts with the
sequences contain selected linear motifs, Nucleic Acids
mRNA decapping complex, Nat. Chem. Biol., 13, 174 180.
Res., 39, e128 e128.
54.
Cloutier, P., Poitras, C., Faubert, D., Bouchard, A.,
38.
Arnoult, N., Correia, A., Ma, J., Merlo, A., Garcia
Blanchette, M., et al. (2020) Upstream ORF encoded
Gomez, S., et al. (2017) Regulation of DNA repair path
ASDURF is a novel prefoldin like subunit of the
way choice in S and G2 phases by the NHEJ inhibitor
PAQosome, J. Proteome Res., 19, 18 27.
CYREN, Nature, 549, 548 552.
55.
Polycarpou Schwarz, M., Groß, M., Mestdagh, P.,
39.
Agarwal, S., Harada, J., Schreifels, J., Lech, P.,
Schott, J., Grund, S. E., et al. (2018) The cancer associat
Nikolai, B., et al. (2006) Isolation, characterization, and
ed microprotein CASIMO1 controls cell proliferation and
genetic complementation of a cellular mutant resistant to
interacts with squalene epoxidase modulating lipid droplet
retroviral infection, Proc. Natl. Acad. Sci. USA, 103, 15933
formation, Oncogene, 37, 4750 4768.
15938.
56.
Kranias, E. G., and Hajjar, R. J. (2012) Modulation of car
40.
Hung, P. J., Johnson, B., Chen, B. R., Byrum, A. K.,
diac contractility by the phopholamban/SERCA2a regu
Bredemeyer, A. L., et al. (2018) MRI is a DNA damage
latome, Circ. Res., 110, 1646 1660.
response adaptor during classical non homologous end
57.
MacLennan, D. H., and Kranias, E. G. (2003) Phospho
joining, Mol. Cell, 71, 332 342.e8.
lamban: a crucial regulator of cardiac contractility, Nat.
41.
Hanyu Nakamura, K., Sonobe Nojima, H., Tanigawa, A.,
Rev. Mol. Cell Biol., 4, 566 577.
Lasko, P., and Nakamura, A. (2008) Drosophila Pgc pro
58.
Tupling, A. R., Bombardier, E., Gupta, S. C., Hussain, D.,
tein inhibits P TEFb recruitment to chromatin in primor
Vigna, C., et al. (2011) Enhanced Ca2+ transport and mus
dial germ cells, Nature, 451, 730 733.
cle relaxation in skeletal muscle from sarcolipin null mice,
42.
Hou, L., Wang, Y., Liu, Y., Zhang, N., Shamovsky, I., et al.
Am. J. Physiol. Cell Physiol., 301, C841 C849.
(2019) Paf1C regulates RNA polymerase II progression by
59.
Gorski, P. A., Ceholski, D. K., and Young, H. S. (2017)
modulating elongation rate, Proc. Natl. Acad. Sci. USA,
Structure function relationship of the SERCA Pump and
116, 14583 14592.
its regulation by phospholamban and sarcolipin, in
43.
Yang, Y., Gao, X., Zhang, M., Yan, S., Sun, C., et al.
Membrane Dynamics and Calcium Signaling, vol. 981,
(2018) Novel role of FBXW7 circular RNA in repressing
Springer International Publishing, Cham, pp. 77 119.
glioma tumorigenesis, JNCI: J. Natl. Cancer Institute, 110,
60.
Bal, N. C., and Periasamy, M. (2020) Uncoupling of sar
304 315.
coendoplasmic reticulum calcium ATPase pump activity
44.
Cardon, T., Franck, J., Coyaud, E., Laurent, E. M. N.,
by sarcolipin as the basis for muscle non shivering thermo
Damato, M., et al. (2020) Alternative proteins are func
genesis, Phil. Trans. R. Soc. B, 375, 20190135.
tional regulators in cell reprogramming by PKA activation,
61.
Hazafa, A., Batool, A., Ahmad, S., Amjad, M., Chaudhry,
Nucleic Acids Res., 48, 7864 7882.
S. N., et al. (2021) Humanin: a mitochondrial derived
45.
Huang, J. Z., Chen, M., Chen, D., Gao, X. C., Zhu, S.,
peptide in the treatment of apoptosis related diseases, Life
et al. (2017) A peptide encoded by a putative lncRNA
Sci., 264, 118679.
HOXB AS3 suppresses colon cancer growth, Mol. Cell, 68,
62.
Calvo, S. E., Clauser, K. R., and Mootha, V. K. (2016)
171 184.e6.
MitoCarta2.0: an updated inventory of mammalian mito
46.
Christofk, H. R., Vander Heiden, M. G., Harris, M. H.,
chondrial proteins, Nucleic Acids Res., 44, D1251 D1257.
Ramanathan, A., Gerszten, R. E., et al. (2008) The
63.
Zhang, S., Reljić, B., Liang, C., Kerouanton, B.,
M2 splice isoform of pyruvate kinase is important for
Francisco, J. C., et al. (2020) Mitochondrial peptide
cancer metabolism and tumour growth, Nature, 452, 230
BRAWNIN is essential for vertebrate respiratory complex
233.
III assembly, Nat. Commun., 11, 1312.
47.
Chen, M., Zhang, J., and Manley, J. L. (2010) Turning on
64.
Lin, Y. F., Xiao, M. H., Chen, H. X., Meng, Y., Zhao, N.,
a fuel switch of cancer: hnRNP proteins regulate alterna
et al. (2019) A novel mitochondrial micropeptide MPM
tive splicing of pyruvate kinase mRNA: Figure 1. Cancer
enhances mitochondrial respiratory activity and promotes
Res., 70, 8977 8980.
myogenic differentiation, Cell Death Dis., 10, 528,
48.
Palmer, C. S., Osellame, L. D., Laine, D., Koutsopoulos,
doi: 10.1038/s41419 019 1767 y.
O. S., Frazier, A. E., and Ryan, M. T. (2011) MiD49 and
65.
Wang, L., Fan, J., Han, L., Qi, H., Wang, Y., et al. (2020)
MiD51, new components of the mitochondrial fission
The micropeptide LEMP plays an evolutionarily conserved
machinery, EMBO Rep., 12, 565 573.
role in myogenesis, Cell Death Dis.,
11,
357,
49.
Rathore, A., Chu, Q., Tan, D., Martinez, T. F.,
doi: 10.1038/s41419 020 2570 5.
Donaldson, C. J., et al. (2018) MIEF1 microprotein regu
66.
Friesen, M., Warren, C. R., Yu, H., Toyohara, T.,
lates mitochondrial translation, Biochemistry, 57, 5564
Ding, Q., et al. (2020) Mitoregulin controls β oxidation in
5575.
human and mouse adipocytes, Stem Cell Rep., 14, 590 602.
50.
Delcourt, V., Brunelle, M., Roy, A. V., Jacques, J. F.,
67.
Liu, G. Y., and Sabatini, D. M. (2020) mTOR at the nexus
Salzet, M., et al. (2018) The protein coded by a short open
of nutrition, growth, ageing and disease, Nat. Rev. Mol. Cell
reading frame, not by the annotated coding sequence, is
Biol., 21, 183 203.
the main gene product of the dual coding gene MIEF1,
68.
Rubtsova, M. P., Vasilkova, D. P., Moshareva, M. A.,
Mol. Cell. Proteomics, 17, 2402 2411.
Malyavko, A. N., Meerson, M. B., et al. (2019) Integrator
51.
Brown, A., Rathore, S., Kimanius, D., Aibara, S., Bai, X.,
is a key component of human telomerase RNA biogenesis,
et al. (2017) Structures of the human mitochondrial ribo
Sci. Rep., 9, 1701.
some in native states of assembly, Nat. Struct. Mol. Biol., 24,
69.
Ferrara Romeo, I., Martinez, P., Saraswati, S.,
866 869.
Whittemore, K., Graña Castro, O., et al. (2020) The
52.
Cardon, T., Salzet, M., Franck, J., and Fournier, I. (2019)
mTOR pathway is necessary for survival of mice with short
Nuclei of HeLa cells interactomes unravel a network of
telomeres, Nat. Commun., 11, 1168.
БИОХИМИЯ том 86 вып. 9 2021
РАЗНООБРАЗИЕ ФУНКЦИЙ МАЛЫХ БЕЛКОВ И ПЕПТИДОВ
1379
LITTLE BUT LOUD. THE DIVERSITY OF FUNCTIONS
OF SMALL PROTEINS AND PEPTIDES -
TRANSLATIONAL PRODUCTS OF SHORT READING FRAMES
Review
P. V. Sergiev1,2,3* and M. P. Rubtsova1*
1 Lomonosov Moscow State University, Faculty of Chemistry, 119991 Moscow, Russia;
e/mail: petya@genebee.msu.ru, mprubtsova@gmail.com
2 Center of Life Sciences, Skolkovo Institute of Sciences and Technology, 143025 Skolkovo, Russia
3 Institute of Functional Genomics, Lomonosov Moscow State University, Moscow, 119991 Moscow, Russia
Cell functioning is a tightly regulated process. For many years, research in the fields of proteomics and functional
genomics has been focused on the role of proteins in cell functioning. The advances in science have led to the uncov
ering that short open reading frames, previously considered non functional, serve a variety of functions. Short read
ing frames in polycistronic mRNAs often regulate their stability and translation efficiency of the main reading frame.
The improvement of proteomic analysis methods has made it possible to identify the products of translation of short
reading frames in quantities that suggest the existence of functional role of those peptides and short proteins. Studies
demonstrating their role unravel a new level of the regulation of cell functioning and its adaptation to changing con
ditions. This review is devoted to the analysis of functions of recently discovered peptides and short proteins.
Keywords: peptides, short open reading frame, long noncoding RNAs, alternative proteins
БИОХИМИЯ том 86 вып. 9 2021
9*