БИОХИМИЯ, 2022, том 87, вып. 1, с. 45 - 67
УДК 573.554
ПРОИСХОЖДЕНИЕ ГЕНЕТИЧЕСКОГО КОДА И ТРАНСЛЯЦИИ
В РАМКАХ СОВРЕМЕННЫХ КОНЦЕПЦИЙ
ПРОИСХОЖДЕНИЯ ЖИЗНИ
Обзор
© 2022
Л.Г. Кондратьева1, М.С. Дьячкова2, А.В. Гальченко3*
1 Институт биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова РАН,
117997 Москва, Россия
2 Институт общей генетики им. Н.И. Вавилова РАН, 119991 Москва, Россия
3 Российский университет дружбы народов (РУДН), 117198 Москва, Россия;
электронная почта: gav.jina@gmail.com
Поступила в редакцию 26.05.2021
После доработки 10.11.2021
Принята к публикации 08.12.2021
Происхождение генетического кода и системы трансляции, возможно, является центральной и самой труд
ной проблемой в изучении происхождения жизни и одной из самых трудных во всей эволюционной биоло
гии. Существует большое количество гипотез возникновения и развития современных генетических систем,
затрагивающих происхождение и раннюю эволюцию генетического кода, а также возникновение реплика
ции и трансляции. Наиболее широко известные гипотезы рассмотрены в данном обзоре. Однако ни одна из
этих гипотез не описывает без пробелов и допущений все этапы ранней эволюции генетических систем. Ги
потеза РНК мира является главенствующей на сегодняшний день научной идеей о ранней эволюции био
логических и пребиологических объектов. Главное её преимущество заключается в том, что она предлагает
в качестве первых живых систем РНК как самодостаточные, с точки зрения воспроизведения, молекулы,
которые способны функционировать как каталитический компонент системы и в то же время - как матрич
ный. Однако есть и существенные недостатки. В частности, до сих пор не открыта и не получена экспери
ментально рибозимная процессивная полимераза. Учитывая взаимную потребность белков и нуклеиновых
кислот в современном мире, многие авторы предлагают сценарии ранней эволюции на основе коэволюции
этих двух классов органических молекул. Подобные гипотезы постулируют, что для репликации нуклеино
вых кислот было необходимо возникновение трансляции, в отличие от мира РНК, где появлению трансля
ции предшествовала эра самореплицирующихся РНК. И хотя такие сценарии менее экономичны, с эволю
ционной точки зрения, так как требуют одномоментного появления и эволюции сразу двух классов органи
ческих молекул, а также синхронизации по времени появления репликации и трансляции, большим их пре
имуществом является то, что они предлагают развитие сразу гораздо более точной и процессивной белко
вой репликации.
КЛЮЧЕВЫЕ СЛОВА: эволюция, зарождение жизни, трансляция, генетический код, РНК мир, проген,
белковый мир, липидный мир.
DOI: 10.31857/S0320972522010043
ВВЕДЕНИЕ
вели к появлению особого состояния материи,
которое мы называем жизнью. По определению
Земля сформировалась ~4,5 миллиарда лет
NASA, жизнь - это «самоподдерживающаяся
назад, и вскоре после этого, ~4 миллиарда лет
химическая система, способная к дарвиновской
назад [1], сложившиеся на планете условия при
эволюции» [2]. Для реализации данной способ
ности появился первобытный генетический
код, который обеспечил трансляцию нуклеино
Принятые сокращения: aaRS - аминоацил тРНК
синтетаза; DRT - прямое кодирование аминокислот с
вых кислот в белки. Для объяснения потенци
РНК матрицы; C DRT - цепь, комплементарная DRT
альных механизмов зарождения генетических
матрице; GADV - модель эволюции белкового мира; систем было предложено большое количество
GARD - модель репликации мицеллярного автокатализа; различных гипотез, каждая из которых делает
PR - шкала растворимости аминокислот в пиридине
акцент на различных событиях, приводящих к
(PolarRequirement); Self rARS - самоаминоацилирующие
рибозимы; SART - самоаминоацилирующаяся рибозим
возникновению функциональной трансляции и
ная матрица.
самоподдерживающейся системы. Они описы
* Адресат для корреспонденции.
вают как происхождение генетического кода,
45
46
КОНДРАТЬЕВА и др.
т.е. возникновение соответствия между амино
довательности без коррекции ошибок, лимити
кислотами и триплетами нуклеотидов, так и
руемый порогом Эйгена, крайне незначителен.
происхождение трансляции, т.е. полимериза
В рамках теории Эйгена увеличение информа
ции аминокислот на основе закодированной в
ционной ёмкости и расширение генома могло
нуклеиновых кислотах информации, и репли
быть достигнуто благодаря увеличению точнос
кации - воспроизведения генетической инфор
ти репликации. Однако эволюция высокоточ
мации.
ной системы репликации, обеспечивающей
Трансляционная система даже в самых прос
коррекцию ошибок, требовала бы значительно
тых современных клетках (например, паразити
большего объёма информации, чем та, которая
ческих и эндосимбиотических бактериях и архе
могла быть закодирована в последовательности
ях, таких как Candidatus Carsonella ruddii,
самореплицирующейся системы в данный мо
Mycoplasma или Nanoarchaeon) чрезвычайно
мент эволюционной траектории. Это создаёт, по
сложна [3]. В основе трансляции лежит большой
меньшей мере, видимость парадокса возникно
комплекс (рибосома), состоящий по меньшей
вения и эволюции живых систем, описываемого
мере из трёх молекул РНК и 60-80 белков (хотя
следующим образом: для кодирования древнего
у облигатных эндосимбионтов может быть
механизма репликации была необходима неко
меньше 35 рибосомных белков [4]), взаимодей
торая минимальная сложность генома, в то вре
ствующих с другими компонентами системы
мя как концепция предела ошибок существенно
трансляции: полным набором тРНК для 20 ами
ограничивала его размер. Таким образом, на
нокислот (~40 видов тРНК с учётом присут
первый взгляд усложнение генома не представ
ствия изоакцепторных тРНК у всех видов),
лялось возможным благодаря существованию
20 аминоацил тРНК синтетазами (aaRS), а так
порога Эйгена [6]. Одним из возможных вари
же факторами трансляции.
антов разрешения кажущегося парадокса явля
При этом ключевой проблемой в определе
ется модель гиперциклов, предложенная Eigen и
нии того, как возникла современная трансля
Schuster [7]. В гиперцикле РНК и ферменты ко
ционная система, является следующий пара
оперируются следующим образом: РНК Ii коди
докс: высокая точность трансляции вряд ли
рует фермент Ei (i = 1, 2, …, n); ферменты цик
могла быть достигнута без сложных, высокораз
лически катализируют репликацию РНК таким
витых молекул РНК и белков, при этом слож
образом, что Ei катализирует репликацию Ii+1, а
ный белковый механизм не мог бы развиваться
En катализирует репликацию I1. Таким образом,
без точной системы трансляции. Этот парадокс
в гиперцикле пептиды совместно с РНК фор
рассматривается в цикле Дарвина-Эйгена
мируют целостную систему кооперативно взаи
(Darwin-Eigen cycle) [3].
модействующих макромолекул. Согласно дан
ной модели, ферменты могли способствовать
повышению точности репликации, в результате
ЦИКЛ ДАРВИНА-ЭЙГЕНА
чего количество наследуемой информации в
примитивной системе возрастало. Модель ги
Теория Eigen [5] постулирует существование
перциклов имеет и недостатки: как было пока
фундаментального предела, ограничивающего
зано в ряде работ [8-10], гиперциклы демон
размер генома и точность репликации в само
стрируют слабую устойчивость по отношению к
реплицирующихся системах (порог Эйгена): ес
мутациям, которые могут приводить к разруше
ли произведение частоты ошибок и информаци
нию структуры гиперцикла. Другим потенци
онной ёмкости системы ниже порога Эйгена,
альным решением парадокса Эйгена является
происходит стабильное наследование; если дан
комбинация естественного отбора и генетичес
ная величина выше пороговой, мутационный
кого дрейфа. Случайные события рекомбина
процесс неизбежно приводит к катастрофичес
ции или дупликации частей генома, приводя
кому вымиранию системы [5]. Иными словами,
щие к умеренному увеличению его размера, мо
уровень ошибок репликационного аппарата ли
гут закрепляться в малых популяциях благодаря
митирует максимальный размер генома (коли
генетическому дрейфу. При условии достаточ
чество закодированной информации). Для ста
ной точности механизма репликации подобные
бильного поддержания репликации, что являет
небольшие изменения в размере генома не
ся необходимым начальным условием биологи
уменьшают значительным образом приспособ
ческой эволюции, система обязана непрерывно
ленность системы, однако предоставляют гене
демонстрировать значения, не выходящие за
тический материал для эволюции новых адап
пределы указанного интервала, в том числе на
тивных функций. Среди таких функций могут
самых ранних этапах своего существования.
быть механизмы, повышающие точность реп
Максимальный размер реплицируемой после
ликации, что отодвигает систему от «обрыва
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
47
РНК-МИР
Гипотеза РНК мира является наиболее ши
роко принятой идеей возникновения жизни на
Земле. Rich [13] в 1962 г. впервые предложил на
личие энзиматической активности у РНК, затем
Crick [14] и Orgel [15] в 1968 г. предположили,
что исходной молекулой для возникновения
жизни могла являться РНК, а Gilbert [16] ввёл
удачный термин «мир РНК» в 1986 г. после того,
как была доказана энзиматическая активность
РНК [15]. Классическая версия зарождения
жизни в рамках РНК мира заключается в перво
начальном синтезе и случайной полимеризации
нуклеотидов, которые привели к образованию
пулов олигомеров реплицирующихся нуклеино
вых кислот; рекомбинационные события приве
ли к образованию более длинных олигомеров,
которые складывались в структуры различной
Рис. 1. Цикл Дарвина-Эйгена (Darwin-Eigen cycle). Мо
сложности и образовывали ферментативно ак
дифицировано по [3]
тивные рибозимы. По мере увеличения слож
ности полинуклеотидов возникали первые
РНК репликазы, образовывались оболочки, и
Эйгена» и обеспечивает необходимый потенци
это привело к появлению протоклеток со спо
ал к дальнейшему увеличению объёма кодируе
собностью к эволюции [17].
мой информации. Таким образом, цикл повто
Свидетельством в пользу РНК мира стало
ряет себя в спиральной прогрессии, приводя к
открытие разнообразных рибозимов, которые
непрерывному увеличению размера генома [6].
могли осуществлять всю основную «работу» на
Данная система с положительной обратной
заре зарождения жизни [18-21]. Обнаруженные
связью получила название цикла Дарвина-Эй
рибозимные активности могли бы свидетель
гена [3] (рис. 1).
ствовать о том, что именно РНК играли роль био
Основной проблемой рассматриваемой тео
логических катализаторов в рамках гипотезы
рии остаётся вопрос о первоначальной инициа
РНК мира. И несмотря на то что природные
ции цикла Дарвина-Эйгена, т.е. как именно бы
РНК репликазы до сих пор не были найдены,
ла достигнута наименьшая сложность генома,
существуют искусственно созданные рибозимы
необходимая для приемлемой репликации. Да
со свойствами РНК полимеразы дистрибутив
же в наименее точных современных системах,
ного типа [22, 23].
таких как вироиды (точность механизмов реп
Известно, что самыми консервативными су
ликации составляет всего ~10-2 [11], т.е. в сред
ществующими генами являются гены трансля
нем одна ошибка на 100 нуклеотидов), реплика
ции [24]. Рибосомные белки, тРНК и aaRS, а
ция катализируется сложными белковыми по
также белковые факторы, которые участвуют в
лимеразами, которые, в свою очередь, синтези
инициации, элонгации и терминации, обладают
руются в результате трансляции при участии
удивительно консервативными структурами и
сложного и многокомпонентного рибосомного
универсальны для всех видов [25, 26]. Общая ко
аппарата. Koonin и Novozhilov [12] предложили
ровая рРНК, которая является главным функ
следующее решение кажущегося парадокса пер
циональным компонентом рибосомы, консер
воначальной инициации цикла Дарвина-Эйге
вативна на протяжении всей филогенетической
на: трансляция в древних самореплицирующих
истории [27, 28], при этом сохраняется как пос
ся системах могла обеспечиваться компонента
ледовательность, так и вторичная, и третичная
ми небелкового происхождения, а инициация
структуры [24].
цикла произошла в условиях мира РНК. При
При этом в теории РНК мира есть ряд нере
этом, помимо гипотезы РНК мира, существуют
шённых проблем [29-31]. Так, аргументом про
другие теории, описывающие в качестве родо
тив теории является то, что РНК слишком слож
начальных ДНК, нуклеопротеиды, белки или
на, чтобы возникать de novo в пребиотической
предполагающие их коэволюцию. Рассмотрим
среде. В частности, есть сложности с абиотичес
возникновение аппарата трансляции и реплика
ким синтезом пиримидиновых рибонуклеоти
ции в рамках этих теорий.
дов [30, 32, 33], нестабильностью молекул РНК
БИОХИМИЯ том 87 вып. 1 2022
48
КОНДРАТЬЕВА и др.
и их высокой чувствительностью к pH и нали
продемонстрировать надёжную и непрерывную
чию ионов магния [29]. Кроме того, для сущест
саморепликацию РНК из исходных молекул.
вования и эволюции РНК мира существует ряд
Изолированная РНК может оказаться недоста
ограничений по физическим условиям: необхо
точным условием для того, чтобы катализиро
димо наличие РНК адсорбирующих поверхнос
вать собственную репликацию, и может нуж
тей, регулярных циклов подсушивания/увлаж
даться в наличии либо других молекул, либо оп
нения, нагревания/охлаждения, заморажива
ределённых условий окружающей среды [11].
ния/оттаивания, а также механизмов защиты
Тем не менее теория РНК мира является наи
РНК от космического излучения [34]. Сущест
более принятой гипотезой, в рамках которой
вует и проблема хиральности при репликации
предложены различные модели эволюции клю
РНК [35]. Нуклеотиды и их предшественники в
чевых компонентов репликации и трансляции:
пребиотическом мире существовали, вероятно,
возникновения генетического кода, образова
как рацемическая смесь право и левовращаю
ния молекул мРНК, тРНК, aaRS, рибосом, а
щих молекул, и включение противоположно
также самого механизма репликации РНК.
вращающего нуклеотида в цепь нарушало бы
Происхождение репликации в рамках гипоте-
репликацию.
зы РНК-мира. Неферментативная матричная
Надёжность репликации является ещё одной
полимеризация нуклеотидов. Согласно гипотезам
проблемой РНК мира: поскольку точность са
неферментативной репликации, удвоение моле
мосборки РНК низкая, количество ошибок уве
кулы РНК могло проходить с помощью достраи
личивается по мере удлинения цепей, и циклы
вания праймера активированными (например,
повторяются. Накопление ошибочных молекул
имидазолом) мономерами [37, 38] или оли
в конечном итоге могло бы привести к гибели
гомерами [39]. Механизм, использующий акти
всей системы [30]. Большой проблемой являет
вированные мономеры, имеет много общего с
ся также низкая процессивность предполагае
современными универсальными биологически
мой древней репликации молекул РНК. Кроме
ми процессами ферментативной репликации
того, при рассмотрении самопроизвольной реп
РНК и ДНК. Экспериментально показано, что
ликации матриц скорость их образования со
скорость присоединения активированных мо
поставима со скоростью их деградации [36].
номеров к праймеру значительно выше, чем ли
Часть парадоксов, связанных с накоплением
гирование праймера к олигонуклеотиду [40].
ошибок, может быть разрешена, если предполо
При этом олигонуклеотиды могут долгое время
жить, что РНКовая РНК полимераза состояла
находиться на матрице, и низкая скорость лиги
из нескольких частей, каждая из которых отно
рования в таком случае не будет проблемой.
сительно короткая, а сборка осуществлялась за
Считается, что эффективное копирование
счёт спаривания. Это могло бы повысить веро
длинных матриц РНК может быть достигнуто за
ятность безошибочной репликации фрагмен
счёт гибридного процесса, в котором олигонук
тов, а значит и образования полноценных комп
леотиды выступают в роли праймеров на не
лексов и, по видимому, могло сделать более эф
скольких сайтах на матрице, а присоединяющие
фективным отбор. Однако подобные полимера
ся мономеры достраивают места пробелов,
зы не были обнаружены или получены экспери
после чего происходит лигирование всех фраг
ментально.
ментов [38, 41]. При этом при рассмотрении не
Появление смысловых цепочек большого
ферментативной репликации необходимо отме
размера также сложно объяснить в рамках РНК
тить, что при таком механизме могут образовы
мира [30]. Этот момент можно проиллюстриро
ваться не только правильные связи между нук
вать на примере случайного происхождения ри
леотидами (3′-5′), но и связи между «непра
бозимной РНК полимеразы: появление не
вильными» гидроксилами рибозы (2′-5′). Кро
скольких РНК полимеразных рибозимов по
ме того, низкая скорость такой репликации со
тенциально возможно, однако они будут зани
поставима со скоростью деградации синтезиро
мать лишь незначительную долю от огромного
ванной РНК, что накладывает значительное ко
количества вероятных последовательностей
личество ограничений на эту концепцию.
РНК [36]. Таким образом, формирование значи
РНК катализируемая репликация. Концеп
мых функциональных полинуклеотидов случай
ция РНК мира подразумевает существование
ным образом кажется маловероятным [30].
рибозима, обладающего свойствами РНК зави
Кроме того, катализ является относительно
симой РНК полимеразы, которая синтезирова
редким свойством РНК, а также РНК имеет
ла бы комплементарные РНК для воспроизвод
слишком ограниченный каталитический репер
ства дополнительных копий самой себя. Образо
туар [29]. Несмотря на достижения в области
вание способной к такой активности молекулы
пребиотической химии, до сих пор не удалось
РНК могло происходить по следующему сцена
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
49
рию: благодаря неферментативной репликации
да Khorana и Nirenberg вместе с Holley в 1968 г.
образовывались популяции реплицирующихся
получили Нобелевскую премию по физиологии
РНК. Среди них некоторые РНК получили бы
и медицине. С момента завершения процесса
большее распространение в случае, если бы они
расшифровки стандартного генетического кода,
обладали более высокой скоростью воспроизво
в котором приняли участие несколько исследо
дства. Таким образом, мог произойти отбор наи
вательских групп [46-49], множество исследо
более подходящих матричных последователь
вателей были вовлечены в изучение его структу
ностей и РНК, способствующий увеличению
ры и свойств. Существует несколько теорий
эффективности процесса репликации. При про
происхождения этой системы строгого соответ
должающемся селективном давлении этот про
ствия кодируемых аминокислот триплетам нук
цесс химической репликации будет вытеснен
леотидов, которые не являются взаимоисключаю
биохимическим процессом репликации [38].
щими, а, вероятно, описывают разные этапы
Возможность существования рибозимной
формирования структуры генетического кода.
РНК полимеразы была показана в работе Bartell
Стереохимическая гипотеза. Согласно стерео
и Szostak [42], которые in vitro воспроизвели эво
химической теории, генетический код должен
люцию большой популяции РНК со случайны
был возникнуть благодаря физико химическим
ми последовательностями и в результате отобра
взаимодействиям между нуклеотидами антико
ли рибозим РНК лигазы размером ~100 нуклео
донов или кодонов и соответствующими амино
тидов, катализирующий соединение двух свя
кислотами.
занных с матрицей олигонуклеотидов [38]. Да
Для экспериментальной проверки гипотезы
лее была проведена серия экспериментов по
стереохимического взаимодействия была разрабо
эволюции in vitro для преобразования этой лига
тана специальная шкала (англ. PolarRequirement,
зы в настоящую РНК полимеразу, которая
PR), отражающая растворимость аминокислот в
действует на отдельной матрице РНК. В резуль
пиридине и его производных (аналогах пирими
тате дальнейшей оптимизации рибозимной по
дина) [50]. Используя эту шкалу, Woese [51] по
лимеразы была увеличена длина синтезируемо
казал, что аминокислоты со схожими значения
го фрагмента, скорость полимеризации, эффек
ми PR, как правило, кодируются родственными
тивность работы на сложных матрицах и точ
кодонами, обычно - кодонами с одинаковым
ность рибозима [38]. Полученный в итоге поли
вторым основанием [50, 51]. С развитием техно
меразный рибозим был способен достаточно
логий получения аптамеров возникли новые ар
быстро синтезировать множество сложнострук
гументы в поддержку стереохимической теории.
турированных РНК [43]. Таким образом, эта оп
В экспериментах SELEX было показано, что
тимизированная РНК полимераза могла ката
фракции случайных аптамеров РНК, отобран
лизировать реципрокный синтез как РНК, так и
ных на основе их способности связываться с оп
её комплементарной цепи, обеспечивая экспо
ределенными аминокислотными остатками,
ненциальную амплификацию коротких матриц
значительно обогащены соответствующими
РНК в форме полимеразной цепной реакции, и
триплетами. Критика экспериментов SELEX
кроме того, была способна синтезировать поли
обычно связана с тем, что специфические ста
меры ДНК на матрице РНК [44]. Несмотря на то
тистически значимые корреляции были обнару
что наиболее оптимизированная форма такой
жены для незначительного количества амино
полимеразы не может реплицировать длинные
кислот, при этом в некоторых случаях последо
последовательности РНК, исследователи дан
вательности были обогащены кодонами, а в дру
ного направления эволюции репликации Joyce
гих - антикодонами [51-55]. Кроме того, срод
и Szostak полагают, что в конечном итоге в лабо
ство было показано для аминокислот, большин
ратории будет разработана процессивная РНК
ство из которых имело сложное строение и ко
репликаза (молекулярная машина, которая
торые могли быть недоступны для формирова
продвигается по полинуклеотидной матрице и
ния структуры генетического кода на ранних
включает мономеры в растущую комплементар
этапах его эволюции [12, 45, 56].
ную цепь) [38].
В серии недавно опубликованных протеом
Происхождение генетического кода в рамках
ных исследований было установлено, что про
гипотезы мира РНК. Генетический код представ
фили распределения плотности пиримидина в
ляет собой систему, определяющую трансляцию
последовательностях мРНК коррелируют с про
нуклеотидной последовательности нуклеино
филями распределения значений PR, кодируе
вых кислот в аминокислотную последователь
мых белков. Иными словами, участки мРНК,
ность полипептидов, и является практически
богатые пиримидинами, кодируют участки бел
универсальной системой живых организ
ка, которые проявляют высокую склонность к
мов [12, 45]. За расшифровку генетического ко
взаимодействию с миметиками пиримидина, и
4 БИОХИМИЯ том 87 вып. 1 2022
50
КОНДРАТЬЕВА и др.
наоборот [51, 57-60]. Неясно, однако, могли ли
щей способности путём последовательного наз
предполагаемые стереохимические взаимодей
начения специфичности второму, затем первому
ствия между нуклеиновыми кислотами и пепти
и, в некоторых случаях, третьему нуклеотиду ко
дами, а не свободными аминокислотами, играть
дона.
значимую роль в происхождении кода [12].
Метаболическая теория. Теория коэволюции
Для объяснения наблюдаемых корреляций в
(метаболическая теория) Wong [67, 68] утвержда
рамках стереохимической гипотезы различны
ет, что структура стандартного кода отражает пу
ми авторами было предложено множество моде
ти биосинтеза аминокислот. Он разделил все
лей стереохимического взаимодействия между
протеиногенные аминокислоты на аминокисло
аминокислотами и нуклеотидами нуклеиновых
ты пребиотического происхождения - амино
кислот [61-65]. Следует отметить, что только
кислоты Фазы 1 (Gly, Ala, Ser, Asp, Glu, Val, Leu,
некоторые из предложенных механизмов (на
Ile, Pro и Thr) и аминокислоты, возникшие в ре
пример, предложенный Woese et al. [50]) потен
зультате биосинтеза из пребиотических - ами
циально способны объяснить процесс прямого
нокислоты Фазы 2 (Phe, Tyr, Arg, His, Trp, Asn,
формирования полипептидной цепи на матрице
Gln, Lys, Cys и Met) [67, 68]. Основываясь на су
нуклеотидной последовательности, так как для
ществующих представлениях о биосинтезе ами
возникновения пептидной связи было необхо
нокислот, Wong предложил следующую схему
димо в первую очередь обеспечить сближение
эволюции биохимических путей синтеза амино
аминокислот в пространстве. Существенная
кислот Фазы 2 из путей синтеза их предшествен
разница в размерах между нуклеотидными
ников, т.е. аминокислот Фазы 1: Ser - Trp,
триплетами и аминокислотами затрудняет
Ser - Cys, Val - Leu, Thr - Ile, Gln - His,
пространственное сближение аминокислот в
Phe - Tyr, Glu - Gln и Asp - Asn [68]. По его
процессе прямого матричного синтеза. Тем не
предположению, возникающая в системе гене
менее, несмотря на свои недостатки, стереохи
тического кода аминокислота наследовала ко
мическая гипотеза может значительно облег
дон своего предшественника. Критика теории
чить понимание того, как происходило форми
коэволюции «код путь» Wong отражена в рабо
рование и ранняя эволюция генетического ко
тах Amirnovin [69] и Ronneberget et al. [70].
да [56].
Теория «замороженной случайности». Соглас
Гипотеза расширения кода. Сценарий эволю
но современным представлениям, расширение
ции кода, предложенный ещё Crick [14], вклю
системы генетического кода по описанному вы
чал примитивную легко реплицирующуюся
ше сценарию было возможно только в случае
нуклеиновую кислоту, несколько наиболее дос
кодирования незначительного количества бел
тупных аминокислот и первичный код, базирую
ков, характеризующихся относительно простой
щийся на двух основаниях. Crick полагал, что
структурой. По мере эволюционного развития
двух оснований могло быть достаточно для ко
организмов усложнялись как структура и специ
дирования ограниченного репертуара амино
фичность функций кодируемых белков, так и их
кислот в составе примитивных белков. При
репертуар. В конечном итоге должен был насту
этом кодон должен был оставаться триплетным
пить момент, когда любое незначительное изме
на протяжении всей эволюции генетического
нение в существующей системе кодирования и
кода, так как нарушение принципа непрерыв
введение новой аминокислоты приводило бы к
ности кода с высокой вероятностью оказалось
критическому нарушению первичной структу
бы летальным даже для примитивного организ
ры значимого количества кодируемых бел
ма. Woese et al. [50], основываясь на паттернах
ков [14]. Таким образом, наблюдаемая высокая
кластеризации аминокислот, наблюдаемых в
консервативность структуры генетического ко
структуре стандартного генетического кода,
да обусловлена давлением отрицательного отбо
выдвинул гипотезу о том, что примитивный код
ра, сила которого увеличивалась по мере услож
мог обладать низкой специфичностью: отдель
нения системы [14, 45]. Соответственно, в ка
ные триплеты кодировали группы химически
кой то момент код перестал накапливать изме
родственных аминокислот, при этом система
нения, и, обсуждая эту проблему, Crick [14]
распознавания базировалась на одном основа
предложил термин «замороженной случайнос
нии. Таким образом, примитивный код, вероят
ти» (англ. «frozen accident»).
но, обладал гораздо более высокой степенью
Теория минимизации ошибок. Crick [14] и
вырожденности и характеризовался относитель
Woese [71] полагали, что формирование наблю
но низким потенциалом кодирования. Данные
даемой структуры стандартного генетического
представления были развиты в модели «2 1 3»
кода происходило случайным образом и не
[66], которая предлагала сценарий расширения
включало какие либо механизмы оптимизации.
генетического кода и увеличения его разрешаю
Однако это плохо согласуется с тем фактом, что
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
51
стандартный генетический код характеризуется
гут углубить наше понимание механизмов эво
высокой степенью устойчивости к ошибкам (в
люции канонического кода.
частности, синонимичные замены, как прави
Возникновение разнообразия молекул РНК.
ло, не приводят к нарушению структуры коди
Молекулы РНК являются центральным звеном
руемого белка; кроме того, замена первого нук
процесса передачи информации от генов к бел
леотида в кодоне часто приводит к включению
кам (мРНК), функционирования самого меха
аминокислоты со схожими физико химически
низма трансляции (рРНК и тРНК) и осущест
ми свойствами). Устойчивость кода к ошибкам
вления различных регуляторных функций (раз
была подтверждена в многочисленных экспери
личные классы некодирующих РНК). Для фор
ментах in silico [56, 72, 73]. Было показано, на
мирования современной трансляторной систе
пример, что вероятность достичь аналогичного
мы в процессе эволюции должны были возник
уровня минимизации ошибок путём случайных
нуть предшественники этих молекул. Так, в ряде
перестановок кодонов стандартного генетичес
гипотез предлагаются модели последовательных
кого кода составляет менее 10-6 [56]. Высокая
преобразований молекул РНК, в ходе которых
устойчивость стандартного генетического кода
сформировались структуры, выполняющие
к трансляционным ошибкам стала главным ар
функции мРНК, тРНК и рибосом.
гументом в пользу теории формирования кода
Теория рекомбинирующихся РНК. В рамках
под давлением естественного отбора. Теория по
РНК мира существует гипотеза, выдвинутая
лучила название теории минимизации оши
Спириным [75], в которой предполагается, что
бок [12, 45]. В исследовании адаптивного ланд
первичный трансляционный аппарат возник до
шафта генетического кода авторы изучили воз
появления аппарата энзиматической реплика
можную эволюцию устойчивости генетического
ции генетического материала и базировался на
кода к трансляционным ошибкам и подтверди
нескольких видах специализированных РНК.
ли гипотезу о том, что эволюция стандартного
Согласно этой гипотезе, многообразие молекул
генетического кода могла включать в себя ста
РНК, в том числе молекул с рибозимной актив
дию его частичной оптимизации с целью мини
ностью, достигалось благодаря рекомбинации
мизировать эффект мутаций, возникающих в
вследствие спонтанной неэнзиматической
процессе трансляции [73].
трансэстерификации абиогенно синтезирован
Таким образом, сценарий возникновения
ных олигорибонуклеотидов. Благодаря такой
современной системы кодирования, называе
рекомбинации коротких молекул РНК, компле
мой стандартным генетическим кодом, мог
ментарно связывающихся с полинуклеотидной
включать следующие стадии: на этапе, когда пул
матрицей, происходило объединение фрагмен
протеиногенных аминокислот характеризовал
тов и первичное размножение молекул РНК.
ся невысоким разнообразием, назначение кодо
А с появлением рибозимов с полимеразной ак
нов могло определяться одним или двумя нуклео
тивностью эффективность репликации должна
тидами, согласно стереохимическим взаимо
была значительно возрастать [75, 76]. Согласно
действиям; затем по мере увеличения доступ
данной гипотезе, рекомбинация молекул РНК
ности аминокислот эволюция происходила по
приводила к образованию первичного трансля
пути расширения кода и увеличения специфич
ционного аппарата: прото мРНК, набора прото
ности кодирования, при этом биохимически
тРНК, каталитически активной проторибосом
родственные аминокислоты кодировались
ной РНК с пептидилтрансферазной актив
родственными кодонами; по мере усложнения
ностью и проторибосомной РНК, способной
кодируемых белков код был оптимизирован та
взаимодействовать одновременно с каталити
ким образом, чтобы минимизировать возмож
ческой проторибосомной РНК, прото мРНК и
ные ошибки трансляции; и, в конце концов,
прото тРНК. Эта система уже могла осущест
когда любые изменения в системе устоявшегося
влять первичную трансляцию [75]. В дальней
соответствия аминокислот кодонам могли стать
шем эта гипотеза развилась в предположение,
летальными для организма, код был «заморо
что только рекомбинаций полирибонуклеоти
жен» в его локальном состоянии, а его дальней
дов недостаточно для появления и эволюции ка
шая эволюция стала невозможной за редкими
талитических РНК, и сначала должны возник
исключениями, что обеспечило наблюдаемую
нуть нуклеозидтрифосфатзависимые хеликазы
универсальность генетического кода. На сегод
на основе РНК [76]. Такие рибозимные хелика
няшний день быстроразвивающаяся синтети
зы могли быть способны раскручивать стабиль
ческая биология уже обладает инструментами и
ные двухспиральные РНК, неизбежно образую
технологиями, которые позволяют модифици
щиеся в процессе синтеза РНК на комплемен
ровать стандартный генетический код [74].
тарных матрицах. Благодаря этому мог возник
Дальнейшие исследования в этой области помо
нуть механизм репликации РНК с использова
БИОХИМИЯ том 87 вып. 1 2022
4*
52
КОНДРАТЬЕВА и др.
нием двухспиральных РНК, что положило нача
тельного прямого кодирования - спиралевид
ло быстрой эволюции древнего мира РНК [76].
ное закручивание нуклеотидной нити вокруг
Можно предположить, что образующиеся пеп
аминокислотной. Однако в этом случае могли
тиды, не так уж и необходимые для такого мира
бы возникать различные межнуклеотидные взаи
рекомбинирующихся РНК, могли стабилизиро
модействия, которые многократно повышали
вать РНК, защищая от гидролиза и потери пра
бы вероятность образования РНКовых шпилек
вильной пространственной структуры [77].
и подобных структур. Большим вопросом оста
Теория прямого кодирования РНК матрицей.
ётся и процесс извлечения пептидного стержня
Самый простой способ кодирования пептидов
из РНКовой спирали. Единственное решение
по шаблону РНК заключается в непосредствен
такой проблемы - непоследовательное кодиро
ном последовательном связывании активиро
вание аминокислот, т.е. расположение кодирую
ванных аминокислот на матрице РНК. Такой
щих триплетов не подряд (рис. 2, а). Переход же
механизм предложен Yarus et al. [52, 53] в теории
к сплошному кодированию мог произойти толь
прямого кодирования РНК (англ. Direct RNA
ко после подключения тРНК к процессу транс
Template, DRT) и основан на стереохимической
ляции.
гипотезе возникновения генетического кода.
Идея напрямую кодирующей РНК была раз
Согласно этой модели, активированные амино
вита также в работе Ma [80] в гипотезу возник
кислоты непоследовательно связывались непос
новения механизма трансляции DRT с учётом
редственно с РНК матрицей (рис. 2, а). Взаимо
принципа репликативной экономии (англ.
действие аминокислоты при этом могло быть
Direct RNA Template with replication parsimony).
как с последовательностью кодона, так и с пос
Под «репликативной экономией» в данной ги
ледовательностью антикодона. Матрица РНК
потезе подразумевается тот факт, что под давле
представляла собой аптамероподобные структу
нием отбора из за сложности репликации суще
ры, распознающие активированные аминокис
ствовала тенденция к «скупому», «экономному»
лоты (рис. 2, а; б, часть 1). Стереохимическое
использованию генетического материала, по
взаимодействие определённых аминокислот с
этому на ранних этапах не могли возникнуть
матрицей РНК сопровождалось образованием
сложные, но «бессмысленные» в данный мо
пептидной связи либо химическим, либо рибо
мент структуры, которые потом обрели бы
зим опосредованным катализом, который осу
функциональность. Данная гипотеза DRT с учё
ществлялся или за счёт активности самой мат
том принципа репликативной экономии пыта
рицы DRT, или реализовывался сторонним ри
ется последовательно объяснить эволюционные
бозимом [58]. Так, активированные аминокис
события появления таких промежуточных
лоты полимеризовались, и формировался коди
звеньев, как тРНК, мРНК или рРНК [80].
руемый DRT пептид. На следующем этапе сис
Гипотеза подразумевает, что из за нестабиль
тема DRT могла эволюционировать в сторону
ности РНК некоторые молекулы DRT могли
современного непрямого кодирования, исполь
деградировать или быть недореплицированны
зующего промежуточное звено - тРНК. Такая
ми из за низкой эффективности рибозимных
промежуточная аминоацил РНК образовыва
РНК полимераз. Эти события могли привести к
лась через присоединение антикодонного фраг
образованию фрагментов, содержащих основ
мента DRT к активированной рибозой группе
ной домен аминокислотного аптамера, окру
аминокислоты (рис. 2, б, часть 2). Таким обра
жённого последовательностями спейсеров. При
зом, происходило преобразование исходной
этом полная репликация DRT производила це
функции связывания аминокислоты в функцию
пи, комплементарные DRT - C DRT (рис 3, а).
кодирования нуклеиновой кислоты [55]. На бо
Фрагменты DRT «распознавали» C DRT сог
лее позднем этапе произошёл переход к едино
ласно принципу комплементарности. Аптамер
образной версии непрямого кодирования
ная область фрагментов трансформировалась в
(рис. 2, б, часть 3), включающего отдельную
конформацию с большой «петлёй распознава
мРНК, активированные аминокислоты, связан
ния», и они могли превратиться в адаптеры, ис
ные с их антикодонами (аминоацил тРНК), а
пользующие C DRT в качестве матрицы
также РНК с пептидилтрансферазной актив
(рис. 3, б). При этом самоаминоацилирование
ностью - риборибосому [52, 78].
могло происходить благодаря рибозимной ак
Основная проблема гипотезы прямого коди
тивности при условии, что фрагмент РНК со
рования аминокислот с РНК матрицы - это вы
держал сайт связывания аминокислоты и один
раженное пространственное несоответствие
из концов цепи находился вблизи сайта связы
размеров аминокислот и нуклеотидных трипле
вания аминокислоты. Это могло быть возмож
тов (рис. 2, в). Такое положение дел оставляет
ным при формировании L образной тРНК по
только один возможный вариант последова
добной структуры, т.е. прото тРНК, имеющей
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
53
a
б
в
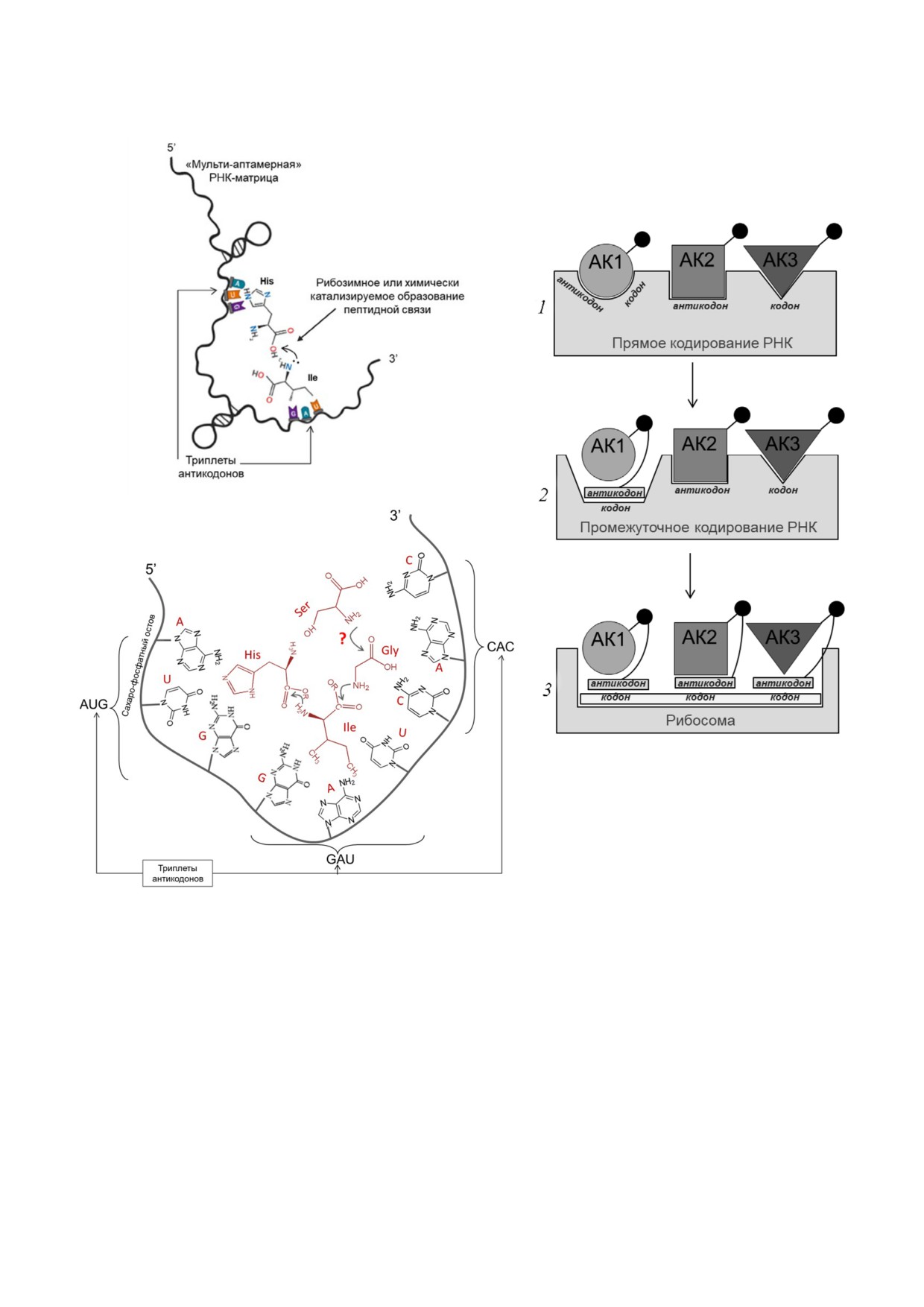
Рис. 2. Модель прямого кодирования на матрице РНК (англ. Direct RNA Template, DRT). а - Схематическое изображение
матрицы DRT на примере фрагмента РНК, непоследовательно связывающего гистидин и изолейцин. Модифицировано
по [79]. б, часть 1 - Первичная трансляция, при которой активированные аминокислоты связываются для полимеризации
с сайтами (кодонами или антикодонами) на матрице РНК (DRT). Активирующие карбоксильные группы (*) участвуют в
полимеризации аминокислот с получением закодированного пептида. б, часть 2 - Промежуточное кодирование РНК. Об
разуется аминоацил РНК (подобие тРНК) путём объединения антикодона кодирующей РНК с аминокислотой. б, часть 3 -
Переход к единой версии современного непрямого кодирования: появление отдельной прото мРНК, примитивных амино
ацил тРНК, состоящих из активированных форм аминокислот, связанных с их антикодонами, и риборибосомы. Модифи
цировано по [52, 53]. в - Пространственное несоответствие триплетов и аминоксилот. При прямом сплошном кодировании
аминокислоты либо «не дотягиваются» друг до друга, либо РНКовая нить закручивается, формируя кольца или спирали
«ногу», вертикально связывающуюся с C DRT,
ностей C DRT, находящихся между прото
и «руку», горизонтально доставляющую амино
тРНК связывающими сайтами (рис. 3, в) [80].
кислоты. Прото мРНК могла бы возникнуть в
Возникшая прото тРНК распознавала прото
результате утраты спейсерных последователь
мРНК за счёт связывания довольно большого
БИОХИМИЯ том 87 вып. 1 2022
54
КОНДРАТЬЕВА и др.
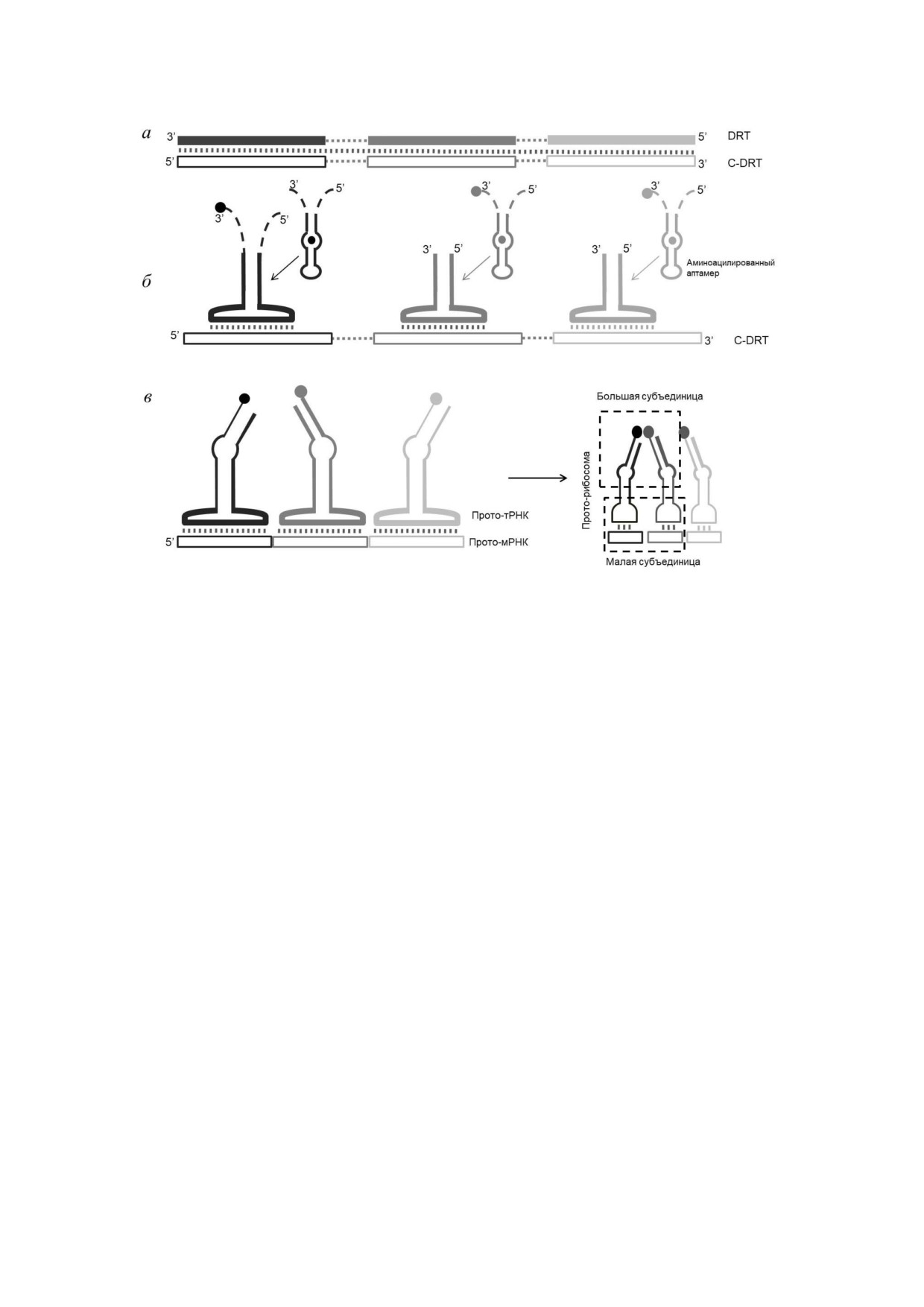
Рис. 3. Модель прямого кодирования на матрице РНК с учётом принципа репликативной экономии (англ. Direct RNA
Template with replication parsimony). а - Репликация DRT приводит к образованию комплементарной цепи - C DRT.
б - В результате деградации или неполной репликации DRT могут образоваться сегменты с аминоацил аптамерным до
меном, фланкированным некоторыми спейсерными последовательностями. Они могут трансформироваться в конфор
мацию с большой «петлёй распознавания», и, таким образом, возникают адаптеры, использующие C DRT в качестве мат
рицы. в - Возникновение мРНК: C DRT теряет последовательности между сайтами связывания аминоацилированного
аптамера, появление малой субъединицы прото рРНК способствует стабилизации кодон антикодонного взаимодействия
триплетных пар прото тРНК и прото мРНК. Появление большой субъединицы прото рРНК способствует сближению
аминокислот, что приводит к образованию пептидных связей. Модифицировано по [80]
количества нуклеотидных остатков, в результате
посредством сложных третичных взаимодей
чего образовывался стабильный комплекс. При
ствий, в том числе между D и Т плечами, обра
сокращении количества «антикодонных» осно
зованными акцепторным стержнем плюс T
ваний до трёх ослаблялось связывание между за
стержнем с Т петлёй и D стержнем с D петлёй
ряженной аминокислотой прото тРНК и прото
вместе с антикодонным стержнем и петлёй. Не
мРНК, при этом появление малой субъединицы
большой размер молекулы тРНК делает её прив
прото рРНК могло обеспечить большую ста
лекательным кандидатом на роль первого воз
бильность комплекса. Большая субъединица
никшего компонента генетической системы.
прото рибосомы возникала в рамках этой гипо
Это предположение основывается на том, что до
тезы для обеспечения эффективности реакции
возникновения гиперциклической самовоспро
пептидилпереноса и удерживала заряженные
изводящейся системы (см. гиперцикл
аминокислоты рядом друг с другом, работая как
Eigen-Schuster [7]) длина нуклеотидных цепей,
рибозим, таким же образом, как и большая
которые могли быть точно реплицированы при
субъединица рРНК в современной рибосо
дарвиновском отборе, составляла не более
ме [80].
100 нуклеотидов, что сопоставимо с размером
Возникновение тРНК. Современные транс
молекулы тРНК.
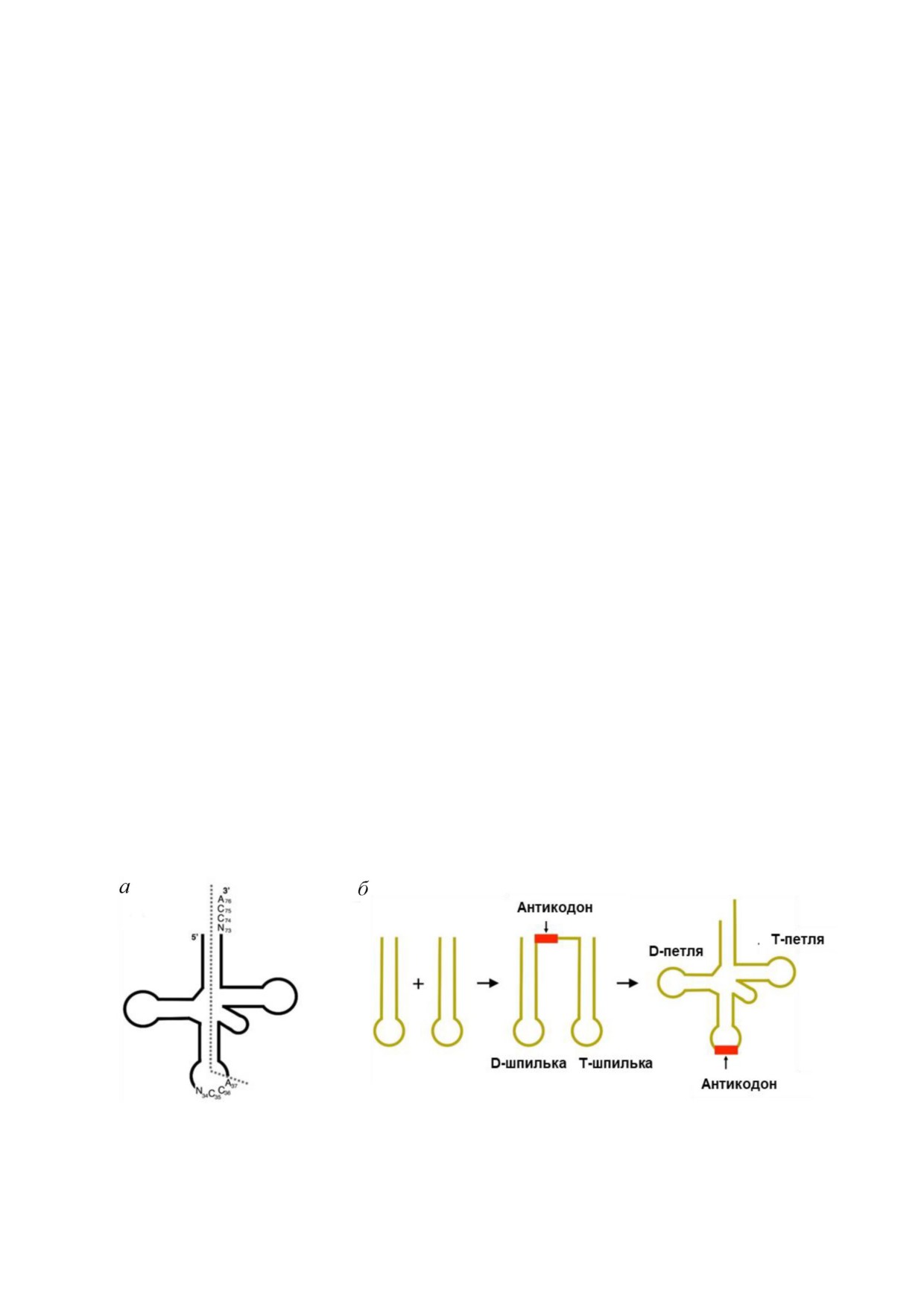
портные РНК состоят в среднем из 76 нуклеоти
В процессе выявления возможного механиз
дов (73-93), обладают универсальной последо
ма возникновения тРНК, проанализировав бо
вательностью ССА на 3′ конце и вторичной
лее тысячи последовательностей тРНК,
структурой клеверного листа. Вторичная струк
Rodin et al. [81] реконструировали консенсус
тура клеверного листа молекулы тРНК склады
ную структуру акцепторных доменов тРНК.
вается в L образную трёхмерную конформацию
Консенсусная последовательность представляла
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
55
собой двухцепочечный палиндром длиной
зывает на то, что большинство последователь
11 нуклеотидов с комплементарными триплета
ностей тРНК возникли в результате двойной
ми в центре, каждый из которых был окружен
дупликации шпильки [83, 84]. Комплементар
мотивами 3′ ACCD и NGGU 5′ на каждой нити,
ное спаривание между нуклеотидами акцептор
где D - это дискриминаторное основание. Та
ного ствола и основаниями антикодонных ство
кой палиндром мог реплицироваться в анало
ла/петли в 5′ половине тРНК и 3′ половине
гичный палиндром длиной 25 пар нуклеотидов с
тРНК соответствует двойному складыванию
комплементарным триплетом в середине, и да
шпильки (рис. 4, б). Таким образом, это может
лее самопроизвольная элонгация обоих палинд
свидетельствовать о том, что образование двой
ромов должна была привести к образованию
ной шпильки в древнем пребиотическом мире
двух спиралей с внутренними комплементарны
лежит в основе эволюции современной структу
ми триплетами. Наконец, удаление 5′ UGGN с
ры тРНК [82].
5′ конца привело к образованию современного
Возникновение рРНК. Существует предполо
тРНК подобного клеверного листа (рис. 4, а).
жение, также основанное на моделях объедине
В исследовании Rodin et al. [81] также была об
ния фрагментов молекул тРНК, которое описы
наружена следующая закономерность: в парах
вает эволюцию рибосом путём слияния (аккре
тРНК с комплементарными антикодонами их
ции) тРНК подобных РНК [85-88]. Димерная
основания во второй позиции акцепторного
центральная часть рибосомной РНК вокруг
ствола также были комплементарными. За
пептидилтрансферазного центра сходна с диме
единственным исключением (пара тРНК
рами тРНК, связанными комплементарными
Phe/тРНК Glu), такой параллелизм особенно
антикодонами. Отметим, что в современных
впечатляет для пар тРНК, распознаваемых ами
митохондриальных рибосомах позвоночных ре
ноацил тРНК синтетазами (aaRS) из противо
гулярные митохондриальные тРНК конститу
положных классов.
тивно выполняют функции 5S рРНК [89], что
Возможным доказательством того, что тРНК
может свидетельствовать в пользу такого сцена
возникла путём лигирования двух шпилечных
рия эволюции рРНК. Кластеризация вторичных
молекул РНК, является следующее наблюдение.
структур РНК [90] обнаружила две основные
Число возможных последовательностей РНК,
группы вторичных структур РНК, одна из кото
состоящих из четырёх нуклеотидных основа
рых характеризуется небольшими, предположи
ний, равно 4n (где n - число нуклеотидов). По
тельно, древними тРНК подобными вторичны
математической оценке Tamura [82], не учиты
ми структурами, и предполагаемая более позд
вающей функциональную частоту, общая масса,
няя группа, состоящая из более крупных рРНК
необходимая для образования нуклеотидной це
подобных вторичных структур.
пи длиной, аналогичной современной тРНК
Существует так называемая аккреционная
(~76 нуклеотидов), составила бы ~1/25 от общей
модель эволюции рибосомы, основанная на ис
массы Земли. Однако для полу тРНК (~35 нук
следовании тРНК рРНК эволюционной оси
леотидов) требуемая масса составляла бы не бо
[87]. Модель предполагает возникновение пред
лее 100 г, что говорит о вероятности образования
ковых сегментов РНК и их объединение в струк
современных тРНК путём, похожим на шпиль
туры большой и малой прото рРНК. Авторы ак
ки [82]. Анализ, проведённый в ряде работ, ука
креционной модели делят эволюцию системы
Рис. 4. Образование молекулы тРНК путём лигирования двух олигомерных молекул на примере глициновой тРНК (а).
Модель образования тРНК из двух шпилечных структур (б). Модифицировано по [82]
БИОХИМИЯ том 87 вып. 1 2022
56
КОНДРАТЬЕВА и др.
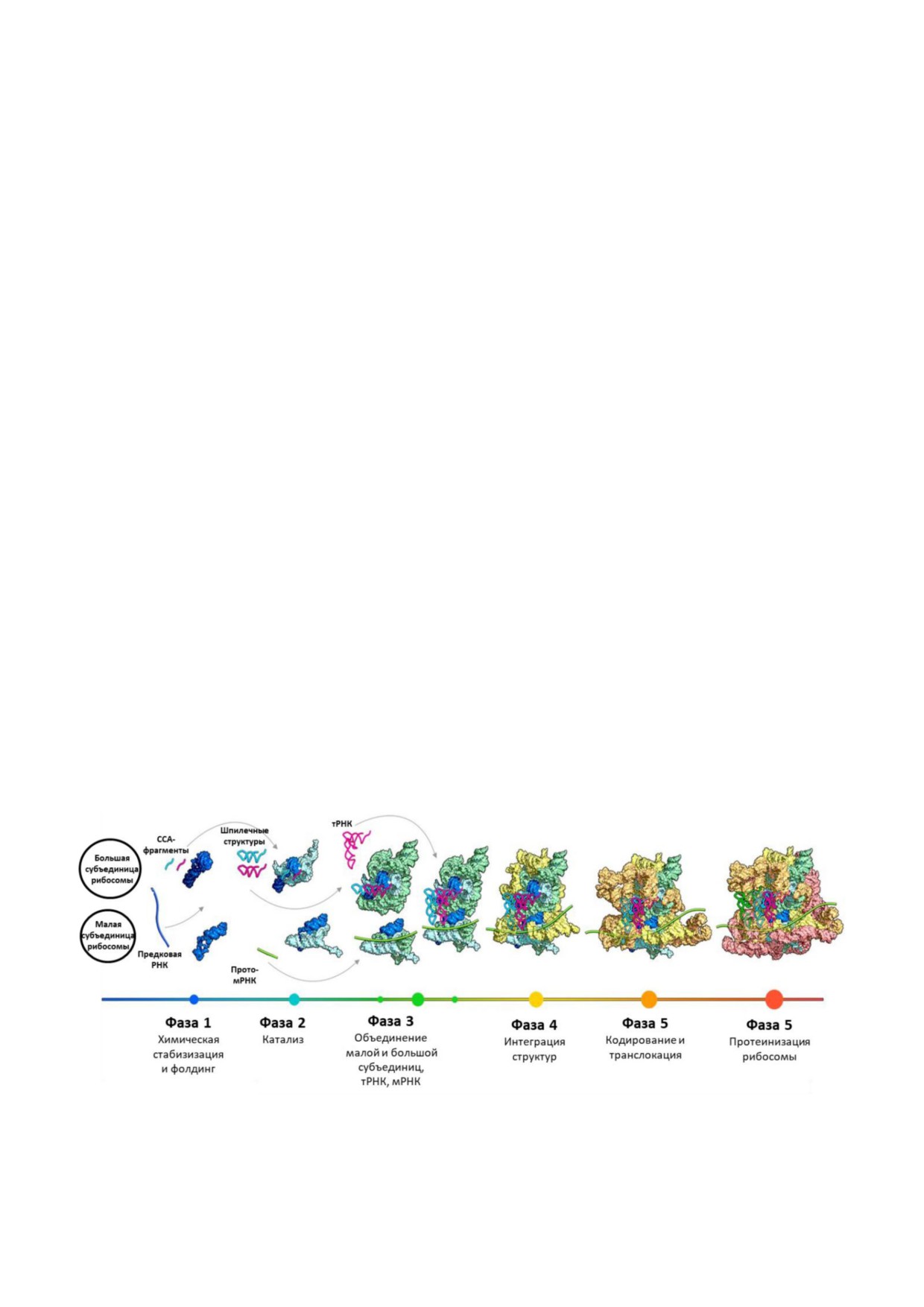
трансляции на шесть фаз (рис. 5). В первой фазе
Р сайты рибозима. В пятой фазе рибосома при
формировались шпилечные структуры РНК, со
обрела примитивную способность к декодиро
держащие ССА последовательность на 3′ конце
ванию, стала энергозависимой и транслоцирую
и стабилизировавшиеся взаимодействиями с ка
щей, возник механизм храповика. Шестая фаза
тионами металлов. Они служили мономерами
знаменует завершение формирования общего
как для будущих прото тРНК, так и прото
ядра рРНК, взаимодействий с рибосомными
рРНК. Во второй фазе шпилечные фрагменты
белками и оптимизацию генетического кода.
неспецифически объединялись в олигомеры бу
Переход от синтеза некодированных гетероген
дущих большой прото рРНК и малой прото
ных аминокислотных олигомеров к белкам с по
рРНК. Большая прото рРНК могла представ
мощью рибосомы давал преимущества, по
лять собой рибозим с пептидилтрансферазным
скольку некоторые продукты реакции связыва
центром, катализирующим неспецифическую
лись с рибосомой. Таким образом, рибосома
полимеризацию аминокислот. Шпилечные
последовательно приобретала способность к
структуры с заряженными аминокислотами
структурированию РНК, катализу, ассоциации
ССА хвостами могли доставлять аминокислоты
субъединиц, коэволюции субъединиц, декоди
в пептидилтрансферазный центр прото рРНК.
рованию и передаче энергии и породила суще
Функция малой прото рРНК могла заключаться
ствующие симбиотические отношения белка и
в формировании ассоциации с одноцепочечной
нуклеиновой кислоты [87].
РНК (прото мРНК). В третьей фазе происходи
Гипотеза de Farias et al. [91] подчёркивает
ло объединение шпилечных структур прото
центральную роль молекул тРНК в возникнове
тРНК в современную L структуру тРНК. За
нии и развитии матричных процессов. Согласно
рождающаяся ассоциация между субъединица
этой гипотезе, тРНК дали начало первым ге
ми прото рРНК опосредовалась прото мРНК и
нам (мРНК) и пептидилтрансферазному центру
тРНК. Малая прото рРНК и прото мРНК рек
рибосомы (рРНК). Эволюционные превраще
рутировались сформировавшейся тРНК: тРНК
ния генетической системы в рамках этой гипо
на одном конце связывалась с пептидилтранс
тезы отражены на рис. 5. Первые гены образова
феразным центром большой прото рРНК, а на
лись из тРНК путём структурных изменений
другом - с прото мРНК, которая, в свою оче
(структура тРНК подобной мРНК) и связывали
редь, связывалась с одноцепочечной областью
другие тРНК (каноническая структура тРНК
малой прото рРНК. В четвертой фазе происхо
клеверного листа) с помощью петли прото ан
дило дальнейшее объединение шпилечных
тикодона. Аминокислоты и небольшие пептиды
структур субъединиц прото рРНК, сформиро
при этом работали подобно кофакторам, стаби
вались А и Р сайты, большая прото рРНК не
лизируя альтернативные конформации прото
кодируемо полимеризовала аминокислоты, а
тРНК. Связывание двух или более тРНК повы
прото мРНК и малая прото рРНК действовали
шало стабильность этих молекул. Связывающие
как кофакторы, позиционирующие тРНК в А и
ся с аминокислотами тРНК могли взаимодей
Рис. 5. Шесть фаз аккреционной модели эволюции рибосом. По работе Petrov, A. S., et al. (2015) History of the ribosome and
the origin of translation, Proc. Natl. Acad. Sci. USA, 112, 15396 15401, doi: 10.1073/pnas.1509761112 [87]. (PNAS не несёт ответ
ственность за точность перевода.)
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
57
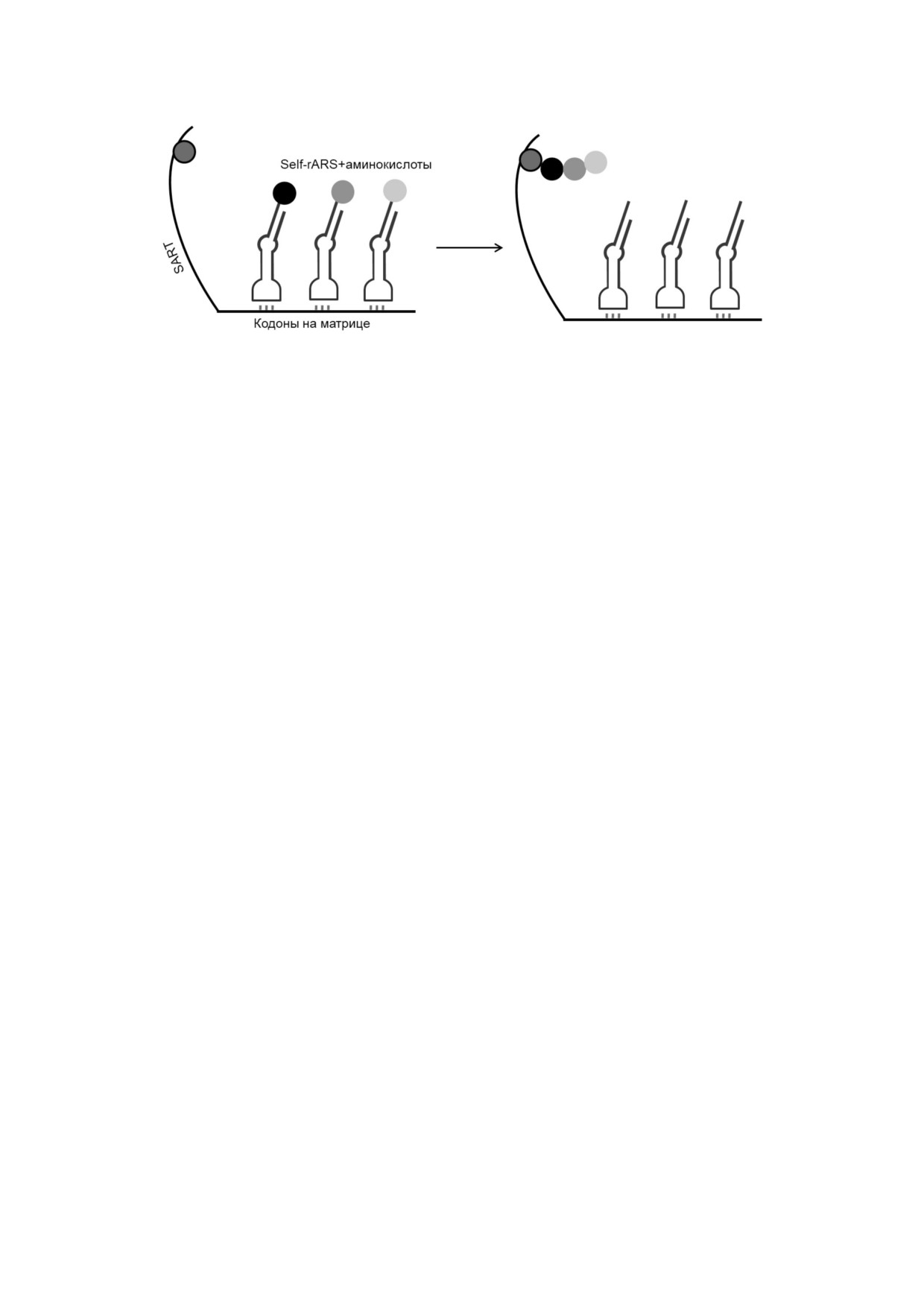
Рис. 6. Модель самоаминоацилирующихся рибозимных матриц (англ. Self rARS Template, SART). Синтез боковой цепи
из трёх аминокислот на нуклеотиде мишени X (темно серый круг) из конъюгатов аминокислота Self rARS, связанных со
специфичными аминокислотам кодонными сайтами на матрице SART. Модифицировано по [94]
ствовать с другими тРНК в открытой конформа
нию самоаминоацилирующих рибозимов (англ.
ции (тРНК подобная структура мРНК). Это
Self rARS) с различной структурой и аминокис
взаимодействие стабилизировало сложную кле
лотной специфичностью [95-98]. Это открытие
верную тРНК каноническую структуру. Кроме
позволяет предположить, что такие Self rARS
того, образование комплекса прото тРНК/
были отнюдь не редким явлением в гипотетичес
мРНК стабилизировало взаимодействие с прото
ком мире РНК. Эти Self rARS осуществляли бы
рРНК (прото пептидилтрансферазный центр),
различные типы реакций. Помимо катализа цис
образуя тройную структуру прото тРНК/мРНК/
аминоацилирования их собственных 2′(3′) OH,
рРНК [91]. Предполагается, что первые пепти
5′ OH или внутреннего 2′ OH, некоторые из
ды, которые были зафиксированы в биологи
них могли быть способны трансаминоацилиро
ческой системе, обладали способностью связы
вать субстратную РНК. Конъюгаты аминокис
вать и стабилизировать тРНК в различных
лот и rARS, образованные Self rARS посред
структурных конформациях, что, в свою оче
ством цис аминоацилирования, могли связы
редь, увеличивало стабильность и эффектив
ваться с кодонными сайтами на матрице функ
ность процесса, тем самым устанавливая первую
циональной РНК и включать их аминоациль
положительную обратную связь между пептида
ные фрагменты в пептид, связанный с ней. При
ми и нуклеиновыми кислотами [92]. С увеличе
этом могла использоваться либо собственная
нием стабильности системы синтеза примитив
трансаминоацилирующая активность, либо
ного белка её эффективность возросла, а ско
принимал участие специализированный транс
рость включения аминокислот в образующиеся
аминоацилирующий рибозим, например, пяти
прото белки увеличилась [91].
нуклеотидный rARS, который работает с раз
Возникновение аминоацилирования РНК. Ги
личными аминокислотами [97]. В любом случае,
потеза самоаминоацилирующихся рибозимных
порядок кодонов сайтов для различных
матриц. Для образования системы кодирова
собственных rARS на РНК матрице определял
ния, трансляции и репликации необходимо,
аминокислотную последовательность получаю
чтобы сформировался процесс, избирательно
щегося в результате пептида [94].
вызывающий обогащение только функциональ
Надо отметить, что существование в рамках
ными РНК (фРНК). Для этого необходим меха
РНК мира рибозимных aaRS, функционирую
низм, позволяющий разделять молекулы на
щих столь же эффективно, что и современные
фРНК, способные связывать лиганды метабо
белковые аминоацилирующие ферменты, при
литов, для которых селективно перезапускается
вело бы к ситуации, когда отсутствовала бы не
репликация, и нефункциональные РНК, не свя
обходимость в этих компонентах системы
зывающие лиганды и разрушающиеся [93]. Для
трансляции именно белковой природы.
иллюстрации такого решения Wong et al. [94]
Гипотеза РНК коферментов - кодирующих
была предложена модель самоаминоацилирую
коферментных «ручек». Существует другая гипо
щихся рибозимных матриц (англ. Self rARS
теза, связывающая РНК мир и аминокисло
Template, SART) (рис. 6).
ты, - The Coding Coenzyme Handle Hypothesis
Изучение рибозимов, обладающих амино
(CCH) [99], в которой аминокислоты действуют
ацилирующей активностью, привело к обнаруже
как коферменты для рибозимов. Данная теория
БИОХИМИЯ том 87 вып. 1 2022
58
КОНДРАТЬЕВА и др.
базируется на том, что многие современные ко
мощью более реакционноспособных сложно
ферменты обладают нуклеотидоподобными
эфирных связей.
фрагментами, что может свидетельствовать об
их возможном происхождении от мира РНК. В
рибо организмах нуклеотидоподобные части
НУКЛЕОПЕПТИДНЫЙ МИР
коферментов могли быть использованы в каче
стве «ручек» (возможно, несущих другие мелкие
Некоторые исследователи [103-105] предпо
молекулы), благодаря чему рибозимы могли
ложили, что полноценного мира РНК на самом
легко захватывать их и управлять ими.
деле никогда не существовало: вместо этого они
В рамках данной гипотезы ранние адаптеры,
предполагают очень раннюю коэволюцию РНК
предки современных антикодонов, были заря
и пептидов в «рибонуклеопептидном» мире.
жены аминокислотами, действующими как ко
Свидетельством в пользу такого сценария явля
ферменты рибозимов в метаболически сложном
ется предполагаемое постепенное добавление
мире РНК. Предковые аминоацил адаптерные
рибосомных белков к рибосомным РНК [106].
синтетазы могли быть подобны современным
Таким образом, возможно, образовавшиеся ри
самосплайсинговым интронам тРНК. Ко
босомные РНК начали взаимодействовать с
дон/антикодон дискриминаторный базовый
аминокислотами или пептидами, что повысило
комплекс, встроенный в эти синтетазы, мог бы
стабильность, эффективность и специфичность
сыграть важную роль в распознавании амино
примитивной рибосомы [104]. На более поздней
кислот.
стадии эволюции генетически кодируемые бел
CCH гипотеза также пытается объяснить,
ки могли заменить эти пептиды. Следовательно,
каково было адаптивное преимущество генети
строгое сохранение структуры рибосомных бел
ческого кода. Специфичность однозначного
ков и их повсеместное присутствие во всех трёх
связывания аминокислот с «ручками» определя
доменах жизни отражают их важный вклад в
ла то, какая аминокислота должна была быть
поддержании структуры и функции рибосо
присоединена, а рибозимы были первыми ката
мы [104]. Кроме того, рибосомы не могут функ
лизаторами этого процесса установления соот
ционировать без своих белковых частей, что мо
ветствия между триплетами и аминокислотами.
жет свидетельствовать об одновременном воз
Позже в ходе эволюции «ручки» превратились в
никновении белковой части и РНК [107]. Соот
адаптеры (тРНК), рибозимы были заменены
ветственно, другие рибонуклеопротеиды, такие
белковыми аминоацил тРНК синтетазами, и
как сплайсосома и РНКаза P, могли столкнуться
многие рибозимы стали молекулами мРНК, по
с той же эволюционной судьбой. И хотя сплай
теряв свою первоначальную ферментативную
сосома, как предполагается, возникла из само
активность. Таким образом, этот механизм мог
сплайсирующихся интронов группы II, которые
являться преадаптацией к кодированию и
не нуждаются в белках для собственного сплай
трансляции [100].
синга, белки им всё же необходимы для фолдин
Изначально предполагалось, что «ручки»
га и стабилизации РНК, а также обратной тран
могли быть моно или динуклеотидными [99],
скрипции [108].
однако только тринуклеотидные соединения
В целом большинство рибозимов имеют ог
могли обеспечить стабильное связывание [100].
раниченные каталитические ресурсы и не демон
В работе Kazakov и Altman [101] показано, что
стрируют каких либо доказательств значитель
тринуклеотид может катализировать металл -
ной сложности [29]. Взаимодействие между
ион зависимое расщепление специфического
пептидами и рибозимами могло существовать
мотива последовательности РНК GAAA.
на ранних этапах эволюции и эффективно спо
Согласно теории коферментных «ручек»
собствовать увеличению каталитического раз
[100], предполагается, что сначала возникла ан
нообразия рибозимов, придавая новые функции
тикодоновая шпилька тРНК, а акцепторный
рибозимам и ускоряя эволюционный про
ствол тРНК был эволюционным следствием.
цесс [104].
Свидетельством этого являются данные [102],
Система из двух полимераз. К коэволюцион
благодаря которым была обнаружена эволюци
ным теориям развития ранних событий эволю
онная связь между антикодоном и пер
ции можно отнести теорию о двух полимеразах
вым-третьим триплетом (и иногда семидеся
Kunin [109]. Он предложил теорию зарождения
тым-семьдесят вторым) в акцепторном стволе.
репликаторно трансляционной системы, за
Далее шпилька антикодона могла стать более
ключающуюся в возникновении тандемно рабо
длинной, несущей повтор антикодона на 3′
тающих молекул: РНК, катализирующей обра
конце молекулы благодаря появлению адапте
зование пептидных связей, и олигопептида, ка
ров, которые связывают аминокислоту с по
тализирующего репликацию РНК. Благодаря
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
59
способности молекулы РНК катализировать об
чии только одной молекулы РНК? [109]. Опи
разование пептидных связей синтезировались
санная двухполимеразная система может быть
олигопептиды из аминокислот, доступных в
инициирована появлением одного компонен
«первичном бульоне». По такому механизму
та - одна каталитическая РНК может синтези
могло происходить присоединение аминокис
ровать пептид с РНК полимеразной актив
лот как к коротким пептидам, так и непосред
ностью, тем самым создавая всю систему, а пеп
ственно к аминокислотам. Короткие пептиды
тидная РНК полимераза, не имеющая матрицы,
могли также образовывать пептидные связи
могла бы способствовать случайному синтезу
друг с другом вследствие неспецифического
молекул РНК с периодическим образованием
пептидилтрансферазного катализа молекулой
функциональных рибозимов. Это означает, что
РНК. Рост цепи прекращался из за простран
появление любого компонента системы могло
ственных помех, поскольку пептидная цепь ста
запустить автокаталитический каскад [109]. При
новилась длиннее и приобретала трёхмерную
этом данная гипотеза не описывает появление
структуру. Среди таких синтезированных пепти
системы кодирования, и обе полимеразы фор
дов могли быть и пептиды, обладающие актив
мируются случайным образом среди пула не
ностью РНК зависимой РНК полимеразы. Эта
функциональных РНК и олигопептидов. Такая
активность олигопептидов позволила бы репли
гипотетическая пептидная полимераза не коди
цировать молекулы РНК. Реакция полимериза
рована, а значит, не сможет воспроизводиться, а
ции использовала РНК в качестве матрицы и
полимераза из РНК не сможет вести процессив
рибонуклеотиды, доступные в окружающей сре
ный синтез пептида, обладающего полимераз
де, и могла протекать в две температурно зави
ной активностью.
симые стадии. При повышенных температурах
Гипотеза прогенов. Существует альтернатив
водородные связи между участками молекул
ная РНК миру гипотеза о возникновении гене
РНК разрушались, делая молекулы доступными
тической системы клетки - гипотеза прогенов,
для РНК зависимой РНК полимеразы, катали
предложенная Альтштейном [107, 110, 111].
зировавшей полимеризацию РНК. Когда темпе
Данная гипотеза объяснила возникновение од
ратура снижалась, молекулы РНК укладывались
новременного синтеза полипептидов и поли
во вторичную структуру и снова становились
нуклеотидов, которые эволюционируют не пос
способны катализировать образование пептид
ледовательно, а взаимосвязанно. Согласно этой
ных связей. Смена дня и ночи могла быть при
гипотезе, нуклеотиды сначала объединялись в
чиной таких температурных колебаний [109].
аминоацилированные тринуклеотиды (проге
Последовательность молекул РНК в рамках
ны), которые являлись единственным субстра
данной гипотезы зависела от процесса естест
том для одновременного синтеза полинуклеоти
венного отбора. Наследственность в системе
да и полипептида, при этом последовательность
поддерживалась через сохранение нуклеотид
аминокислот кодировалась в полинуклеотиде, а
ной последовательности молекул РНК. Неточ
образующийся полипептид обладал свойствами
ная репликация и, возможно, другие типы мута
процессивной полимеразы (прогенлигазы). Об
ций могли давать материал для отбора. Улучше
щая формула прогена NpNpNp~pХ~aa, где N -
ние каталитической активности РНК привело
нуклеозид, p - фосфат, Х - бифункциональный
бы к увеличению концентрации каталитических
агент, например рибоза или другой углевод, аа -
пептидов в ближайшем микроокружении, а ка
аминокислота, ~ - макроэргическая связь. При
талитические пептиды, в свою очередь, репли
этом аминокислота специфична по отношению
цировали бы ближайшую доступную РНК,
к своему триплету.
т.е. в большинстве случаев ту, которая послужи
В рамках данной гипотезы прогены образу
ла матрицей для их синтеза. В результате луч
ются путём соединения динуклеотида NpNp и
ший РНК катализатор имел больше шансов
аминоацилированного нуклеотида Np~pX~aa,
быть воспроизведённым. Таким образом, такая
специфически обусловленного взаимодействи
двухполимеразная система может рассматри
ем динуклеотида и аминокислоты третьего нук
ваться как примитивный дарвиновский орга
леотида. Механизм образования прогенов
низм [109].
представлен на рис. 7. Этот механизм позволяет
При кажущейся простоте теории сам автор
объяснить первичное возникновение генетичес
теории указывает на её проблемы. 1. Достаточно
кого кода и отбор веществ для возникающей ге
ли специфичности системы для успешного син
нетической системы, включая гомохираль
теза РНК полимераз? 2. Будут ли неизбежно об
ность [95, 96]. Фосфат в нуклеотидах прогена
разованы случайные ферменты, расходующие
находится на 3′ , а не на 5′ конце (это повыша
«впустую» активированные мономеры? 3. Явля
ет вероятность образования фосфодиэфирной
ется ли синтез пептидов возможным при нали
связи), и третий нуклеотид должен обязательно
БИОХИМИЯ том 87 вып. 1 2022
60
КОНДРАТЬЕВА и др.
быть дезоксирибонуклеотидом, второй - чаще
событие, формируется связанная пара «ген-
дезоксирибонуклеотидом, а первый - чаще ри
процессивная полимераза (прогенлигаза)», при
бонуклеотидом, т.е. полинуклеотид является по
чём полимераза имеет тропность к 3′ концу по
лирибодезоксирибонуклеотидом.
линуклеотида.
Такие прогены являются мономерами для
Полимераза, расположенная на 3′ конце ге
одновременного синтеза полинуклеотида и по
на, перемещается к 5′ концу, соединяя прогены,
липептида (рис. 7). Два прогена за счёт слабого
комплементарные матрице. Одновременно син
стэкинг взаимодействия сближаются друг с дру
тез комплементарного полинуклеотида проис
гом, и второй проген удерживается рядом с пер
ходил посредством репликации и транскрип
вым взаимодействием с аминокислотой перво
ции, поскольку исходный ген был представлен
го. Стабилизируется комплекс комплементар
(-) цепью, и сопровождается синтезом «непра
ным олигонуклеотидом, состоящим из 6 нуклео
вильного» белка, кодируемого (-) цепью. По
тидов. N Конец аминокислоты второго прогена
лимераза перемещается к 3′ концу вновь синте
сближается с активированным С концом ами
зированной (+) цепи и реплицирует её, одно
нокислоты первого прогена; образуется дипеп
временно формируя новую молекулу полимера
тид, связанный со вторым прогеном. N Конце
зы и новую (-) цепь. Две молекулы полимеразы
вая аминокислота дипептида, Glu или Asp (из за
дополнительно создают две новые (+) цепи. За
их кислотных боковых групп), влияет на образо
тем синтезируются две новые (-) цепи и две до
вание фосфодиэфирной связи между первым и
полнительные молекулы полимеразы (всего
вторым прогенами. В результате образуется гек
4 молекулы полимеразы).
сануклеотид, связанный с дипептидом. Дипеп
Таким образом, гипотеза прогенов предлага
тид обладает повышенной (по сравнению с од
ет единый механизм, который позволяет объяс
ной аминокислотой) способностью удерживать
нить: 1) возникновение генетического кода;
следующий проген, и весь процесс повторяется,
2) механизм отбора компонентов для образова
при этом дипептид переносится на третий про
ния генетической системы, в том числе возни
ген (транспептидация) и помогает последнему
кающей гомохиральности; и 3) возникновение и
присоединяться к гексануклеотиду. Образуется
размножение бимолекулярной генетической
нонануклеотид с трипептидом. Таким образом,
системы, состоящей из гена полинуклеотида и
при присоединении прогенов каждый триплет
белковой процессивной полимеразы, кодируе
запоминает кодируемую аминокислоту и уста
мой этим геном.
навливается связь «генотип (порядок триплетов
Согласно данной гипотезе, возникшая би
в полинуклеотиде)-фенотип (порядок амино
молекулярная система - это первый вирусопо
кислот в полипептиде)» [107].
добный организм (Protoviroidum primum), кото
Растущий пептид постоянно участвует в
рый размножается и эволюционирует путём
формировании системы: если последняя амино
единого процесса репликации, транскрипции и
кислота, присоединяющаяся к пептиду, увели
трансляции и является прародителем жизни на
чивает вероятность включения следующего про
Земле [107]. Экспериментальная химическая
гена, рост всей системы продолжается; если
проверка механизма образования прогенов
уменьшает - рост системы останавливается. Та
принципиально возможна, но пока не осущест
ким образом, возникающий полипептид фер
влена.
мент растёт в тесном взаимодействии с его буду
Гипотеза урзимов. Согласно другой гипотезе,
щим субстратом (прогеном) и, как очень редкое
описанной Li et al. [112], катализ и кодирование
Рис. 7. Формирование бимолекулярной генетической системы из прогенов. Модифицировано по [107]
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
61
совместно развились благодаря одновременно
комплекс пептид-РНК; комплексы росли в
му образованию пептидов и РНК.
длину, возможно, до размера предполагаемого
В центре этой гипотезы лежат «урзимы»,
гена, кодирующего 23 аминокислотные смыс
представляющие собой небольшие (120-130
ловые/антисмысловые продукты. 23 аминокис
аминокислот), высококонсервативные фраг
лотные смысловые/антисмысловые пептиды
менты доменов двух суперсемейств аминоацил
стали родоначальниками aaRS ферментов клас
тРНК синтетаз (aaRS). Два этих суперсемей
са I и II; 23 аминокислотные пептиды были ли
ства - aaRS I и aaRS II - были когда то закоди
гированы в 46 аминокислотный белок, обладав
рованы противоположными цепями одного и
ший большей эффективностью аминоацилиро
того же гена [113]. В отличие от рибозимов, ко
вания. Некоторые из образовывавшихся пепти
торые лежат в основе гипотезы о мире РНК, па
дов всё более специализировались как полиме
ры урзимов I и II классов представляют собой
разы нуклеиновых кислот, другие аминоацили
консенсусные предковые формы, достаточные
ровали двухцепочечную РНК, двухцепочечная
для кодон направленного синтеза неслучайных
РНК сохранила свою роль шаблона репликации
пептидов. Наличие механизма
«активации»
и развила пептидилтрансферазную активность,
аминокислот экспоненциально увеличивает эф
став большой рибосомальной субъединицей.
фективность их полимеризации, а возникнове
Одноцепочечная РНК стала всё чаще брать на
ние урзимов решает эту проблему. Урзимы зна
себя доминирующую роль матрицы и эволюцио
чительно снижают структурную сложность, не
нировала в современную мРНК [115].
обходимую для осуществления трансляции ру
Авторы гипотезы об урзимах, Li et al. [112],
диментарного генетического кода. Таким обра
указывают на два недостатка РНК мира: 1) от
зом, урзимы проявляют каталитические свой
сутствие естественных рибозимов, реплицирую
ства задолго до полного формирования генети
щих РНК, активирующих аминокислоты и ами
ческого кода.
ноацилирующих тРНК, и 2) зависимость рибо
Урзимная модель коэволюции предполагает
зимного пути от раннего появления точной реп
стереохимическое «комплементарное» взаимо
ликации. Такой путь не предлагает механизма
действие между РНК и пептидами, основываю
для преодоления «обрыва Эйгена» [116]. Оба
щееся на примитивном генетическом коде, в ко
этих слабых места, согласно Li et al. [112], объяс
тором каждый нуклеотид в дуплексе РНК соот
няются совместной эволюцией пептидных ката
ветствует двум аминокислотам, а каждый ди
лизаторов и РНК.
пептид определяет соответствующее основание.
Данная гипотеза интересна для понимания
Подобное взаимодействие описано в работе
эволюции трансляции, но не даёт чёткого про
Carter и Kraut [114]. Стереохимическое кодиро
веряемого ответа на вопрос о кодировании дру
вание могло генерировать пептиды и соответ
гих пептидов. Непонятно, как может эволюцио
ствующие им РНК гены. При этом такие гены
нировать такая система, в которой белки для
будут иметь длину гена тРНК (~72 основания).
репликации не кодируются РНК.
Таким образом, мир, основанный на коэволю
ции РНК и белков, состоял из коротких РНК
«генов» и кодируемых ими небольших белков,
БЕЛКОВЫЙ МИР
специфически аминоацилирующих молекулы
РНК, благодаря которым осуществлялась рибо
Гипотеза GADV-белкового мира. Гипотеза
зимная трансляция.
GADV белкового мира основана на наблюде
Автор такой гипотезы о коэволюции РНК и
нии, что белки, содержащие примерно одинако
белков, Carter [115], предлагает следующий сце
вое количество четырёх аминокислот (глици
нарий пребиотической эволюции: в течение
на - G (gly), аланина - А (ala), аспарагиновой
длительного периода химической эволюции мо
кислоты - D (asp) и валина - V (val), могут об
номеры аминокислот и нуклеиновых кислот на
разовывать основные вторичные структуры бел
чали собираться в ковалентные комплексы,
ка - α спирали и β слои [117, 118]. В рамках
включающие структурно «комплементарные»
этой теории предполагается, что основные
олигонуклеотиды и дипептиды; далее могли
GADV белки могли полимеризоваться случайно
происходить синтез и лигирование пептидов и
из этих четырёх аминокислот, формируя глобу
олигонуклеотидов, специфичность которых бы
лы. Согласно этой гипотезе, пребиотический
ла бы ограничена спариванием нуклеотидных
мир до появления генов включал стадию, на ко
оснований и грубым стереохимическим кодиро
торой GADV белки были способны к псевдо
ванием между двумя типами полимеров, присое
репликации. Возникновение примитивных био
диняющих новые мономеры таким образом,
логических систем в таком случае должно было
чтобы стабилизировать двойной спиральный
проходить следующие стадии: 1) пребиотичес
БИОХИМИЯ том 87 вып. 1 2022
62
КОНДРАТЬЕВА и др.
кий синтез и накопление gly, ala, asp и val; 2) слу
нии, что воспроизведение состава компонентов
чайное формирование белков из этих четырёх
пребиотического мира предшествует реплика
аминокислот; 3) формирование первичной ме
ции непосредственно информационных после
таболитической системы за счёт активности
довательностей. Согласно этому взгляду на про
GADV белков, синтез нуклеотидов и олигонук
тобиологию, первые репликаторы были скопле
леотидов РНК; 4) формирование первичного ге
ниями спонтанно объединяющихся, гетероген
нетического кода и образование генов; 5) обра
ных и в основном неканонических (не образую
зование двойной спирали РНК, возникновение
щих бислойные структуры) амфифильных ве
наследственности и передачи генетического ма
ществ. Это предположение подтверждается
териала поколениям - начало биологической
построением химической кинетики модели реп
эволюции; 6) эволюция генетического кода и ге
ликации мицеллярного автокатализа (англ.
нетических систем.
Graded autocatalysis replication domain, GARD)
Согласно этой гипотезе, примитивный гене
[122]. Согласно GARD, в пребиотическом мире
тический код состоял из предварительного
существовал отбор неравновесных агрегаций ве
GNC кода, включавшего четыре кодона
-
ществ (композомов), которые могли быть спо
GGC, GCC, GAC и GUC, кодирующих 4 основ
собны к автокаталическому гомеостатическому
ные аминокислоты GADV мира: gly, ala, asp и
росту [124]. Такой процесс, наряду со случай
val. Затем произошло расширение генетическо
ным делением композома, воплощал в себе при
го кода до 16 кодонов (GGC, GGG, GCC, GCG,
митивное клеточное размножение. При нали
GAC, GAG, GUC, GUG, CUC, GUG, CCC,
чии в среде амфифильных веществ из них само
CCG, CAC, CAG, CGC и CGG), кодировавших
формируются мицеллы, этот факт может слу
10 аминокислот (gly, ala, asp, val, glu, leu, pro, his,
жить предпосылкой для возникновения само
gln и arg) [119]. Порядок появления аминокис
поддерживающихся автокаталитических липид
лот в пребиотическом GADV мире примерно
ных систем. В таком случае GARD эволюция
совпадает с установленным Trifonov [120] эво
мира до РНК проявлялась в отборе различных
люционным порядком образования аминокис
композомов в пределах разреженного ландшаф
лот, основанном на 60 различных критериях.
та пригодности в ответ на химические измене
Примитивные одноцепочечные нуклеино
ния окружающей среды. По модели GARD «ге
вые кислоты состояли из GNC повторяющихся
нотипом» являлся сам композом, т.е. состав ав
последовательностей и были получены путём
токаталитических мицелл, и он же определял
образования фосфодиэфирной связи между ко
способность к репликации, т.е. «фенотип» сис
донами GNC. Двуцепочечные молекулы обра
темы. «Геном» композома в такой системе ока
зовывались при комплементарном спаривании
зывает прямое кинетическое влияние на эффек
молекул (GNC)n или кодонов GNC, и, таким
тивность и точность гомеостатического роста,
образом, была сформирована первичная систе
следовательно, на его репликацию. Таким обра
ма репликации [121].
зом, авторы гипотезы указывают на корреляцию
Проблемой гипотезы «белкового мира» яв
между мутациями в системе и её репродукцией
ляется то, что стабильная и функциональная
без необходимости в посреднике, таком как ко
структура возможна только для коротких пребио
дируемый белок или функциональная
тических пептидов, а случайное формирование
РНК [124]. Гипотеза предполагает раннее объ
правильной пространственной укладки слож
единение ключевых компонентов биологии в
ных пептидов/белков в таких условиях практи
липидной протоклетке: амфифильные композо
чески невозможно. Кроме того, пробелом дан
мы могли включать аминокислоты и пептиды,
ной теории является отсутствие чёткого пути
азотистые основания РНК и олигонуклеотиды,
для репликации и передачи информации на
способные к комплементарному спариванию,
ранних этапах эволюции, основанных на пепти
кофакторы, хелаторы металлов, тиолы, олиго
дах.
фосфаты и многие другие соединения, между
которыми формировались метаболические свя
зи. Данная модель не даёт ответов на вопросы о
ЛИПИДНЫЙ МИР
возникновении именно генетических систем,
но связывает при этом между собой разные
Гипотезы липидного мира [122-124] предпо
классы молекул. Кроме того, моделирование
лагают, что простые гидрофобные молекулы ли
GARD демонстрирует в образующихся системах
пидов могут сохранять и распространять инфор
те же термодинамические и кинетические осо
мацию, таким образом, претерпевая эволюцию.
бенности, которые соответствуют предполагае
Модель GARD. Модель липидного мира
мому предшественнику современных живых
Lancet et al. [124] основывается на предположе
клеток. GARD композомы характеризуются ав
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
63
торами как диссипативные системы, что также
ной реплицировать длинные полинуклеотидные
является отличительными признаками жиз
последовательности, решительно укрепило бы
ни [124]. При этом сами авторы гипотезы указы
позиции гипотезы мира РНК, но такого дости
вают на скудость экспериментальных данных
жения ещё нет [38]. В настоящее время не изве
для взаимно каталитических сетей.
стно ни одной построенной только из нуклеоти
РНК-липидный мир. Идея возникновения
дов процессивной полимеразы. Яркими приме
жизни из амфифильных композомов [122] была
рами таких молекулярных машин являются раз
развита в гипотезе РНК липидного мира Mallik
нообразные современные ДНК и РНК поли
и Kundu [123]. Гипотеза о коэволюции липидов
меразы, которые имеют чисто белковую или
и РНК основана на предположении, что совре
нуклеопротеидную природу [125]. Очень выра
менные стабильные взаимодействия ли
зительна ситуация с рибосомой, которая, по су
пид-РНК - это молекулярные «окаменелости»
ти, является молекулярной машиной, полиме
древней стадии эволюции, когда мир РНК воз
ризующей аминокислоты при движении по по
ник из мира липидов. Предполагается, что древ
линуклеотидной матрице (мРНК). Известно,
ние рибозимы были «защищены» липидами, что
что рибосомная РНК является рибозимом, спо
обеспечило их выживание в жёстких условиях
собным соединять аминокислоты пептидной
пребиотического океана. Присутствующие в ок
связью
[125]. Однако рибосома содержит
ружающей среде ионы металлов могли катали
30-40% белка, без которого не может выпол
зировать репликацию РНК внутри защищённых
нять свою функцию. После 1990 г. были искус
липидных мицелл. Авторами представлен воз
ственно получены рибозимы со свойствами
можный в рамках этой модели механизм транс
РНК полимеразы [22, 23], однако это полиме
ляции и перехода от РНК липидного мира к
разы дистрибутивного типа, не способные дви
РНК белковому: РНК-прото рибосомы, при
гаться по матрице. Таким образом, рибозимы
креплённые к внутреннему слою липидного пу
могут образовывать и фосфодиэфирные связи
зырька, случайно полимеризовали аминокисло
между нуклеотидами, и пептидные связи между
ты. Синтез случайных полипептидов в большом
аминокислотами, но, вероятно, не способны без
пуле везикул мог привести к образованию функ
белка выполнять функцию молекулярных ма
циональных белков, которые могли сыграть
шин. Если это действительно так, гипотеза ми
важную роль в ранней эволюции [123].
ра РНК является таким же тупиковым направ
лением для решения проблемы происхождения
жизни, как и гипотеза Опарина [126], поскольку
ЗАКЛЮЧЕНИЕ
не может предложить конкретного механизма
возникновения генетической системы.
Наши знания и представления о современ
Кроме того, несмотря на то что РНК облада
ных организмах и молекулах, которые они содер
ет способностью выполнять многие функции
жат, обеспечивают формирование предпосылок
белковых ферментов, нет данных об участии ри
для предположения о том, что развитие биологи
бозимов в функционировании клеточных сис
ческих систем началось с эволюции семейств мо
тем, которые должны были возникнуть на са
лекул, которые могли бы катализировать их
мых ранних этапах биологической эволюции.
собственную репликацию. Существует большое
Так, рибозимы не принимают участия в синтезе
количество гипотез, которые описывают подоб
нуклеотидов, в работе энергетических систем, в
ные сценарии, наиболее широко известные из
аминоацилировании тРНК. При этом искус
них мы постарались представить в данном обзо
ственно получены рибозимы, способные эф
ре. Однако практически ни одна из этих гипотез
фективно и специфично аминоацилировать
не описывает без пробелов и допущений все эта
тРНК [95-98], но все естественные aaRS явля
пы ранней эволюции генетических систем.
ются белками. Эти факты трудно совместимы с
Гипотеза РНК мира сегодня является главен
гипотезой мира РНК и позволяют думать о том,
ствующей научной идеей, и существует немало
что кодированный синтез белка имел место на
доводов в её пользу, которые мы описали и кото
самых ранних этапах эволюционного развития
рым посвящено множество работ, в том числе
живого мира.
рассмотренных в настоящем обзоре. В рамках
Представляется разумной идея о коэволю
этой теории разработано много вероятных меха
ции двух типов макромолекул - нуклеиновых
низмов эволюции компонентов трансляции.
кислот и белков. Такой сценарий не столь эко
При этом главная проблема мира РНК до сих
номичен в эволюционном отношении, как жи
пор не решена - не достигнуто полное самовос
вой мир на основе одной только РНК, однако,
произведение РНК. Открытие процессивной
вероятно, что пептиды (и полипептиды) играли
рибозимной полимеразы (репликазы), способ
важную роль в раннем биологическом мире. Ги
БИОХИМИЯ том 87 вып. 1 2022
64
КОНДРАТЬЕВА и др.
потезой, которая хорошо дополняет мир РНК
ного генетического кода и первой бимолекуляр
вероятными ранними белковыми молекулами,
ной генетической системы, реплицирующейся
требовавшимися для осуществления трансля
на основе единого процесса репликации тран
ции, является гипотеза урзимов. Эволюционно
скрипции трансляции. Автором предлагается
консервативные aaRS двух классов могли способ
уникальная концепция возможного первого жи
ствовать более точному установлению соответ
вого неклеточного организма - протовироида и
ствия тРНК и аминокислот в трансляции. Комп
описывается принципиальный механизм его
лементарные цепи РНК кодировали каждая свой
биологической (дарвиновской) эволюции в
пептид, невысокая точность репликации могла
направлении к клетке. Главный недостаток ги
привести к образованию новой молекулы, при
потезы заключается в отсутствии её экспери
этом её функциональность могла не нарушиться
ментальной проверки, которая, однако, прин
при сохранении пространственной структуры.
ципиально возможна.
И таким образом, гипотеза урзимов избегает
Критика рассмотренных представлений о
«обрыва Эйгена», потому что наиболее важным
возникновении и эволюции трансляции не озна
её ограничением является не точность последо
чает, что важные экспериментальные данные, по
вательности, а сохранение структурных элемен
лученные на основе гипотезы РНК мира, и идеи,
тов и каталитических активностей коротких по
выдвинутые в этой и других гипотезах, не могут
липептидных шпилек. При этом маловероятным
быть использованы для понимания важных ас
совпадением кажется сохранение комплемен
пектов эволюции живого мира и не могут быть
тарности цепочек РНК, кодирующих такие пеп
положены в основу будущих экспериментов.
тиды, при низкой точности репликации.
Идея о том, что генетические системы сфор
мировались на основе двух коэволюционирую
Финансирование. Работа выполнена при под
щих макромолекул разных классов, реализована
держке Программы стратегического академи
в гипотезе двух полимераз, а также в гипотезе
ческого лидерства РУДН.
прогенов. В последней на основе одного прин
Конфликт интересов. Авторы заявляют об от
ципа (образование аминоацилированных нук
сутствии конфликта интересов.
леотидных триплетов) теоретически решаются
Соблюдение этических норм. Настоящая
несколько важнейших проблем теории возник
статья не содержит описания каких либо иссле
новения жизни: отбор веществ, включая гомо
дований с участием людей или животных в каче
хиральность, механизм возникновения первич стве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Dodd, M. S., Papineau, D., Grenne, T., Slack, J. F.,
10. Zintzaras, E., Santos, M., and Szathmáry, E.
(2002)
Rittner, M., et al. (2017) Evidence for early life in Earth’s
“Living” under the challenge of information decay: the
oldest hydrothermal vent precipitates, Nature, 543, 60 64,
stochastic corrector model vs. hypercycles, J. Theor. Biol.,
doi: 10.1038/nature21377.
217, 167 181.
2.
Benner, S. A. (2010) Defining life, Astrobiology, 10, 1021
11. Gago, S., Elena, S. F., Flores, R., and Sanjuan, R. (2009)
1030, doi: 10.1089/ast.2010.0524.
Extremely high mutation rate of a hammerhead viroid,
3.
Wolf, Y. I., and Koonin, E. V. (2007) On the origin of the
Science, 323, 1308, doi: 10.1126/science.1169202.
translation system and the genetic code in the RNA world
12. Koonin, E. V., and Novozhilov, A. S. (2017) Origin and
by means of natural selection, exaptation, and subfunction
evolution of the universal genetic code, Annu. Rev. Genet.,
alization, Biol. Direct, 2, 14, doi: 10.1186/1745 6150 2 14.
51, 45 62, doi: 10.1146/annurev genet 120116 024713.
4.
Nikolaeva, D. D., Gelfand, M. S., and Garushyants, S. K.
13. Rich, A. (1962) Horizons in Biochemistry (Kasha, M., and
(2021) Simplification of ribosomes in bacteria with tiny
Pullman, B., eds.) Academic Press, New York.
genomes, Mol. Biol. Evol., 38, 58 66, doi: 10.1093/molbev/
14. Crick, F. H. (1968) The origin of the genetic code, J. Mol.
msaa184.
Biol., 38, 367 379, doi: 10.1016/0022 2836(68)90392 6.
5.
Eigen, M. (1971) Selforganization of matter and the evolu
15. Orgel, L. E. (1968) Evolution of the genetic apparatus,
tion of biological macromolecules, Naturwissenschaften,
J. Mol. Biol.,
38,
381393, doi:
10.1016/0022
58, 465 523, doi: 10.1007/bf00623322.
2836(68)90393 8.
6.
Penny, D. (2005) An interpretive review of the origin of life
16. Gilbert, W. (1986) Origin of life: The RNA world, Nature,
research, Biol. Philos., 20, 633 671, doi: 10.1007/s10539
319, 618.
004 7342 6.
17. Le Vay, K., and Mutschler, H. (2019) The difficult case of
7.
Eigen, M., and Schuster, P. (1977) The hypercycle. A prin
an RNA only origin of life, Emerg. Top. Life Sci., 3, 469
ciple of natural self organization. Part A: Emergence of the
475, doi: 10.1042/etls20190024.
hypercycle,
Naturwissenschaften,
64,
541565,
18. Kruger, K., Grabowski, P. J., Zaug, A. J., Sands, J.,
doi: 10.1007/BF00450633.
Gottschling, D. E., et al. (1982) Self splicing RNA:
8.
Szathmáry, E. (1986) Some remarks on hypercycles and the
Autoexcision and autocyclization of the ribosomal RNA
stochastic corrector model, Endocyt. Cell Res., 3, 337 339.
intervening sequence of Tetrahymena, Cell, 31, 147 157,
9.
Szathmáry, E., and Demeter, L. (1987) Group selection of
doi: 10.1016/0092 8674(82)90414 7.
early replicators and the origin of life, J. Theor. Biol., 128,
19. Robertson, H. D., Altman, S., and Smith, J. D. (1972)
463 486.
Purification and properties of a specific Escherichia coli
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
65
ribonuclease which cleaves a tyrosine transfer ribonucleic
inevitability of polymerase function, Orig. Life Evol.
acid presursor, J. Biol. Chem., 247, 5243 5251.
Biosph., 29, 375 390, doi: 10.1023/a:1006544611320.
20.
Klemm, B. P., Wu, N., Chen, Y., Liu, X., Kaitany, K. J.,
40.
Prywes, N., Blain, J. C., Del Frate, F., and Szostak, J. W.
et al. (2016) The diversity of ribonuclease P: Protein and
(2016) Nonenzymatic copying of RNA templates contain
RNA catalysts with analogous biological functions,
ing all four letters is catalyzed by activated oligonu
Biomolecules, 6, doi: 10.3390/biom6020027.
cleotides, eLife, 5, doi: 10.7554/eLife.17756.
21.
Walter, N. G., and Engelke, D. R. (2002) Ribozymes: cat
41.
Szostak, J. W. (2011) An optimal degree of physical and
alytic RNAs that cut things, make things, and do odd and
chemical heterogeneity for the origin of life? Philos. Trans.
useful jobs, Biologist, 49, 199 203.
R. Soc. Lond. B Biol. Sci., 366, 2894 2901, doi: 10.1098/
22.
Johnston, W. K., Unrau, P. J., Lawrence, M. S., Glasner,
rstb.2011.0140.
M. E., and Bartel, D. P. (2001) RNA catalyzed RNA poly
42.
Bartel, D. P., and Szostak, J. W. (1993) Isolation of new
merization: accurate and general RNA templated primer
ribozymes from a large pool of random sequences [see
extension, Science, 292, 1319 1325, doi: 10.1126/science.
comment], Science, 261, 1411 1418, doi: 10.1126/science.
1060786.
7690155.
23.
Wochner, A., Attwater, J., Coulson, A., and Holliger, P.
43.
Horning, D. P., and Joyce, G. F. (2016) Amplification of
(2011) Ribozyme catalyzed transcription of an active
RNA by an RNA polymerase ribozyme, Proc. Natl. Acad.
ribozyme, Science, 332, 209 212, doi: 10.1126/science.
Sci. USA, 113, 9786 9791, doi: 10.1073/pnas.1610103113.
1200752.
44.
Samanta, B., and Joyce, G. F. (2017) A reverse transcrip
24.
Bowman, J. C., Hud, N. V., and Williams, L. D. (2015)
tase ribozyme, eLife, 6, doi: 10.7554/eLife.31153.
The ribosome challenge to the RNA world, J. Mol. Evol.,
45.
Koonin, E. V. (2017) Frozen accident pushing 50:
80, 143 161, doi: 10.1007/s00239 015 9669 9.
Stereochemistry, expansion, and chance in the evolution of
25.
Ouzounis, C., and Kyrpides, N. (1996) The emergence of
the genetic code, Life (Basel), 7, doi: 10.3390/life7020022.
major cellular processes in evolution, FEBS Lett., 390,
46.
Nirenberg, M. W., and Matthaei, J. H. (1961) The depen
119 123, doi: 10.1016/0014 5793(96)00631 x.
dence of cell free protein synthesis in E. coli upon natural
26.
Ganoza, M. C., Kiel, M. C., and Aoki, H. (2002)
ly occurring or synthetic polyribonucleotides, Proc. Nat.
Evolutionary conservation of reactions in translation,
Acad. Sci. USA, 47, 1588 1602.
Microbiol. Mol. Biol. Rev., 66, 460 485, doi: 10.1128/
47.
Gardner, R. S., Wahba, A. J., Basilio, C., Miller, R. S.,
mmbr.66.3.460 485.2002.
et al. (1962) Synthetic polynucleotides and the amino acid
27.
Melnikov, S., Ben Shem, A., Garreau de Loubresse, N.,
code, VII, Proc. Natl. Acad. Sci. USA, 48, 2087.
Jenner, L., Yusupova, G., et al. (2012) One core, two shells:
48.
Wahba, A. J., Gardner, R. S., Basilio, C., Miller, R. S.,
bacterial and eukaryotic ribosomes, Nat. Struct. Mol. Biol.,
Speyer, J. F., et al. (1963) Synthetic polynucleotides and the
19, 560 567, doi: 10.1038/nsmb.2313.
amino acid code, VIII, Proc. Natl. Acad. Sci. USA, 49, 116.
28.
Hsiao, C., Lenz, T. K., Peters, J. K., Fang, P. Y., Schneider,
49.
Söll, D., Ohtsuka, E., Jones, D., Lohrmann, R.,
D. M., et al. (2013) Molecular paleontology: a biochemical
Hayatsu, H., et al. (1965) Studies on polynucleotides,
model of the ancestral ribosome, Nucleic Acids Res., 41,
XLIX. Stimulation of the binding of aminoacyl sRNA’s to
3373 3385, doi: 10.1093/nar/gkt023.
ribosomes by ribotrinucleotides and a survey of codon
29.
Bernhardt, H. S. (2012) The RNA world hypothesis: the
assignments for 20 amino acids, Proc. Natl. Acad. Sci. USA,
worst theory of the early evolution of life (except for all the
54, 1378.
others)(a), Biol. Direct, 7, 23, doi: 10.1186/1745 6150 7
50.
Woese, C. R., Dugre, D. H., Dugre, S. A., Kondo, M., and
23.
Saxinger, W. C. (1966) On the fundamental nature and
30.
Bregestovski, P. D. (2015) “RNA World”, a highly improb
evolution of the genetic code, Cold Spring Harb. Symp.
able scenario of the origin and early evolution of life on
Quant. Biol., 31, 723 736, doi: 10.1101/sqb.1966.031.
earth, J. Evol. Biochem. Physiol.,
51,
7284,
01.093.
doi: 10.1134/S0022093015010111.
51.
Woese, C. R. (1968) The fundamental nature of the genet
31.
Wills, P. R., and Carter, C. W., Jr. (2018) Insuperable prob
ic code: prebiotic interactions between polynucleotides and
lems of the genetic code initially emerging in an RNA
polyamino acids or their derivatives, Proc. Natl. Acad. Sci.
world, Biosystems, 164, 155 166, doi: 10.1016/j.biosystems.
USA, 59, 110 117, doi: 10.1073/pnas.59.1.110.
2017.09.006.
52.
Yarus, M., Widmann, J. J., and Knight, R. (2009) RNA
32.
Orgel, L. E. (2004) Prebiotic chemistry and the origin of
amino acid binding: a stereochemical era for the genetic
the RNA world, Crit. Rev. Biochem. Mol. Biol., 39, 99 123,
code, J. Mol. Evol., 69, 406 429, doi: 10.1007/s00239 009
doi: 10.1080/10409230490460765.
9270 1.
33.
Kim, H. J., Ricardo, A., Illangkoon, H. I., Kim, M. J.,
53.
Yarus, M. (1998) Amino acids as RNA ligands: A direct
Carrigan, M. A., et al. (2011) Synthesis of carbohydrates in
RNA template theory for the code’s origin, J. Mol. Evol.,
mineral guided prebiotic cycles, J. Am. Chem. Soc., 133,
47, 109 117, doi: 10.1007/pl00006357.
9457 9468, doi: 10.1021/ja201769f.
54.
Yarus, M. (2000) RNA-ligand chemistry: a testable source
34.
Спирин А. С. (2007) Когда, где и в каких условиях мог
for the genetic code, RNA, 6, 475 484, doi: 10.1017/
возникнуть и эволюционировать мир РНК? Палеон>
s1355838200002569.
тологический журнал, 41, 11 19.
55.
Yarus, M., Caporaso, J. G., and Knight, R. (2005) Origins
35.
Tupper, A. S., Shi, K., and Higgs, P. G. (2017) The role of
of the genetic code: the escaped triplet theory, Annu. Rev.
templating in the emergence of RNA from the prebiotic
Biochem., 74, 179 198, doi: 10.1146/annurev.biochem.74.
chemical mixture, Life, 7, 41, doi: 10.3390/life7040041.
082803.133119.
36.
Szostak, J. W. (2012) The eightfold path to non enzymatic
56.
Koonin, E. V., and Novozhilov, A. S. (2009) Origin and
RNA replication, J. Syst. Chem., 3, 2, doi: 10.1186/1759
evolution of the genetic code: the universal enigma,
2208 3 2.
IUBMB Life, 61, 99 111, doi: 10.1002/iub.146.
37.
Joyce, G. F., and Orgel, L. E. (1986) Non enzymic tem
57.
Hajnic, M., Osorio, J. I., and Zagrovic, B.
(2014)
plate directed synthesis on RNA random copolymers.
Computational analysis of amino acids and their sidechain
Poly(C, G) templates, J. Mol. Biol., 188, 433441,
analogs in crowded solutions of RNA nucleobases with
doi: 10.1016/0022 2836(86)90166 x.
implications for the mRNA-protein complementarity
38.
Joyce, G. F., and Szostak, J. W. (2018) Protocells and RNA
hypothesis, Nucleic Acids Res., 42, 12984 12994.
Self Replication, Cold Spring Harb. Perspect. Biol., 10,
58.
Hlevnjak, M., Polyansky, A. A., and Zagrovic, B. (2012)
doi: 10.1101/cshperspect.a034801.
Sequence signatures of direct complementarity between
39.
James, K. D., and Ellington, A. D. (1999) The fidelity of
mRNAs and cognate proteins on multiple levels, Nucleic
template directed oligonucleotide ligation and the
Acids Res., 40, 8874 8882, doi: 10.1093/nar/gks679.
5 БИОХИМИЯ том 87 вып. 1 2022
66
КОНДРАТЬЕВА и др.
59.
Polyansky, A. A., Hlevnjak, M., and Zagrovic, B. (2013)
81. Rodin, S., Rodin, A., and Ohno, S. (1996) The presence of
Analogue encoding of physicochemical properties of pro
codon anticodon pairs in the acceptor stem of tRNAs,
teins in their cognate messenger RNAs, Nat. Commun., 4,
Proc. Natl. Acad. Sci. USA, 93, 4537 4542, doi: 10.1073/
2784, doi: 10.1038/ncomms3784.
pnas.93.10.4537.
60.
Polyansky, A. A., Hlevnjak, M., and Zagrovic, B. (2013)
82. Tamura, K. (2015) Origins and early evolution of the tRNA
Proteome wide analysis reveals clues of complementary
molecule, Life (Basel), 5, 16871699, doi: 10.3390/
interactions between mRNAs and their cognate proteins as
life5041687.
the physicochemical foundation of the genetic code, RNA
83. Di Giulio, M. (1992) On the origin of the transfer RNA
Biol., 10, 1248 1254, doi: 10.4161/rna.25977.
molecule, J. Theor. Biol., 159, 199 214, doi: 10.1016/
61.
Gamow, G. (1954) Possible relation between deoxyribonu
s0022 5193(05)80702 7.
cleic acid and protein structures, Nature, 173, 318.
84. Widmann, J., Di Giulio, M., Yarus, M., and Knight, R.
62.
Melcher, G. (1974) Stereospecificity of the genetic code,
(2005) tRNA creation by hairpin duplication, J. Mol. Evol.,
J. Mol. Evol., 3, 121 140, doi: 10.1007/bf01796558.
61, 524 530, doi: 10.1007/s00239 004 0315 1.
63.
Hendry, L., Bransome, E., Hutson, M., and Campbell, L.
85. Caetano Anolles, D., and Caetano Anolles, G. (2016)
(1981) First approximation of a stereochemical rationale
Piecemeal buildup of the genetic code, ribosomes, and
for the genetic code based on the topography and physico
genomes from primordial tRNA building blocks, Life
chemical properties of “cavities” constructed from models
(Basel), 6, doi: 10.3390/life6040043.
of DNA, Proc. Nat. Acad. Sci. USA, 78, 7440 7444, doi:
86. De Farias, S. T., Rêgo, T. G., and José, M. V. (2021) Origin
10.1073/pnas.78.12.7440.
of the 16S ribosomal molecule from ancestor tRNAs,
64.
Balasubramanian, R., Seetharamulu, P., and
J. Mol. Evol., 89, 249 256, doi: 10.1007/s00239 021
Raghunathan, G. (1980) A conformational rationale for
10002 8.
the origin of the mechanism of nucleic acid directed pro
87. Petrov, A. S., Gulen, B., Norris, A. M., Kovacs, N. A.,
tein synthesis of “living” organisms, Orig. Life, 10, 15 30,
Bernier, C. R., et al. (2015) History of the ribosome and the
doi: 10.1007/bf00928940.
origin of translation, Proc. Natl. Acad. Sci. USA, 112,
65.
Shimizu, M. (1982) Molecular basis for the genetic code,
15396 15401, doi: 10.1073/pnas.1509761112.
J. Mol. Evol., 18, 297 303, doi: 10.1007/bf01733895.
88. Demongeot, J., and Seligmann, H. (2020) Comparisons
66.
Massey, S. E. (2006) A sequential “2 1 3” model of genet
between small ribosomal RNA and theoretical minimal
ic code evolution that explains codon constraints, J. Mol.
RNA ring secondary structures confirm phylogenetic and
Evol., 62, 809 810, doi: 10.1007/s00239 005 0222 0.
structural accretion histories, Sci. Rep., 10, 1 14.
67.
Wong, J. T. (1975) A co evolution theory of the genetic
89. Amunts, A., Brown, A., Toots, J., Scheres, S. H., and
code, Proc. Natl. Acad. Sci. USA,
72,
19091912,
Ramakrishnan, V. (2015) The structure of the human mito
doi: 10.1073/pnas.72.5.1909.
chondrial ribosome, Science, 348, 95 98.
68.
Wong, J. T. (2005) Coevolution theory of the genetic code at
90. Seligmann, H., and Raoult, D. (2018) Stem loop RNA
age thirty, Bioessays, 27, 416 425, doi: 10.1002/bies.20208.
hairpins in giant viruses: invading rRNA like repeats and a
69.
Amirnovin, R. (1997) An analysis of the metabolic theory
template free RNA, Front. Microbiol., 9, 101.
of the origin of the genetic code, J. Mol. Evol., 44, 473 476,
91. De Farias, S. T., Rego, T. G., and Jose, M. V. (2016) tRNA
doi: 10.1007/pl00006170.
core hypothesis for the transition from the RNA world to
70.
Ronneberg, T. A., Landweber, L. F., and Freeland, S. J.
the ribonucleoprotein world, Life (Basel),
6,
(2000) Testing a biosynthetic theory of the genetic code:
doi: 10.3390/life6020015.
fact or artifact? Proc. Natl. Acad. Sci. USA, 97, 13690
92. Noller, H. F. (2012) Evolution of protein synthesis from an
13695, doi: 10.1073/pnas.250403097.
RNA world, Cold Spring Harb. Perspect. Biol., 4, a003681,
71.
Woese, C. R. (1965) On the evolution of the genetic code,
doi: 10.1101/cshperspect.a003681.
Proc. Natl. Acad. Sci. USA, 54, 1546 1552, doi: 10.1073/
93. Wong, J. T. (2014) Emergence of life: From functional
pnas.54.6.1546.
RNA selection to natural selection and beyond, Front.
72.
Haig, D., and Hurst, L. D. (1991) A quantitative measure
Biosci. (Landmark Ed), 19, 1117 1150, doi: 10.2741/4271.
of error minimization in the genetic code, J. Mol. Evol., 33,
94. Wong, J. T., Ng, S. K., Mat, W. K., Hu, T., and Xue, H.
412 417, doi: 10.1007/BF02103132.
(2016) Coevolution theory of the genetic code at age forty:
73.
Novozhilov, A. S., Wolf, Y. I., and Koonin, E. V. (2007)
Pathway to translation and synthetic life, Life (Basel), 6,
Evolution of the genetic code: partial optimization of a
doi: 10.3390/life6010012.
random code for robustness to translation error in a rugged
95. Illangasekare, M., Sanchez, G., Nickles, T., and Yarus, M.
fitness landscape, Biol. Direct, 2, 24, doi: 10.1186/1745
(1995) Aminoacyl RNA synthesis catalyzed by an RNA,
6150 2 24.
Science, 267, 643 647, doi: 10.1126/science.7530860.
74.
Mukai, T., Lajoie, M. J., Englert, M., and Söll, D. (2017)
96. Lee, N., Bessho, Y., Wei, K., Szostak, J. W., and Suga, H.
Rewriting the genetic code, Annu. Rev. Microbiol., 71, 557 577.
(2000) Ribozyme catalyzed tRNA aminoacylation, Nat.
75.
Спирин А. (2001) Биосинтез белков, мир РНК и про
Struct. Biol., 7, 28 33, doi: 10.1038/71225.
исхождение жизни, Вестник Российской академии на>
97. Suga, H., Hayashi, G., and Terasaka, N. (2011) The RNA
ук, 71, 320 328.
origin of transfer RNA aminoacylation and beyond, Philos.
76.
Spirin, A. (2013) The emergence of molecular machines as
Trans. R. Soc. B, 366, 2959 2964, doi: 10.1098/rstb.2011.
a prerequisite of the ancient RNA world evolution,
0137.
Paleontol. J., 47, 1016 1029.
98. Turk, R. M., Chumachenko, N. V., and Yarus, M. (2010)
77.
Schimmel, P. (2011) The RNP bridge between two worlds,
Multiple translational products from a five nucleotide
Nat. Rev. Mol. Cell. Biol., 12, 135, doi: 10.1038/nrm3061.
ribozyme, Proc. Natl. Acad. Sci. USA, 107, 4585 4589,
78.
Nissen, P., Hansen, J., Ban, N., Moore, P. B., and Steitz,
doi: 10.1073/pnas.0912895107.
T. A. (2000) The structural basis of ribosome activity in
99. Szathmary, E. (1993) Coding coenzyme handles: a hypoth
peptide bond synthesis, Science,
289,
920930,
esis for the origin of the genetic code, Proc. Natl. Acad. Sci.
doi: 10.1126/science.289.5481.920.
USA, 90, 9916 9920, doi: 10.1073/pnas.90.21.9916.
79.
Dunn, I. S. (2011) RNA templating of molecular assembly
100. Szathmary, E. (1999) The origin of the genetic code:
and covalent modification patterning in early molecular
Amino acids as cofactors in an RNA world, Trends Genet.,
evolution and modern biosystems, J. Theor. Biol., 284, 32
15, 223 229, doi: 10.1016/s0168 9525(99)01730 8.
41, doi: 10.1016/j.jtbi.2011.06.009.
101. Kazakov, S., and Altman, S. (1992) A trinucleotide can
80.
Ma, W. (2010) The scenario on the origin of translation in
promote metal ion dependent specific cleavage of RNA,
the RNA world: in principle of replication parsimony, Biol.
Proc. Natl. Acad. Sci. USA, 89, 7939 7943, doi: 10.1073/
Direct, 5, 65, doi: 10.1186/1745 6150 5 65.
pnas.89.17.7939.
БИОХИМИЯ том 87 вып. 1 2022
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ГЕНЕТИЧЕСКОГО КОДА
67
102. Rodin, S. N., and Ohno, S. (1997) Four primordial modes
114. Carter, C. W., Jr., and Kraut, J. (1974) A proposed model
of tRNA synthetase recognition, determined by the (G,C)
for interaction of polypeptides with RNA, Proc. Natl. Acad.
operational code, Proc. Natl. Acad. Sci. USA, 94, 5183
Sci. USA, 71, 283 287, doi: 10.1073/pnas.71.2.283.
5188, doi: 10.1073/pnas.94.10.5183.
115. Carter, C. W. (2015) What RNA world? Why a peptide/RNA
103. Di Giulio, M. (2008) An extension of the coevolution the
partnership merits renewed experimental attention, Life
ory of the origin of the genetic code, Biol. Direct, 3, 37,
(Basel), 5, 294 320, doi: 10.3390/life5010294.
doi: 10.1186/1745 6150 3 37.
116. Koonin, E. V. (2011) The Logic of Chance: The Nature and
104. Saad, N. Y. (2018) A ribonucleopeptide world at the origin
Origin of Biological Evolution, FT Press.
of life, J. Syst. Evol., 56, 1 13, doi: 10.1111/jse.12287.
117. Ikehara, K. (2002) Origins of gene, genetic code, protein
105. Altstein, A. D., and Efimov, A. V. (1988) Physico chemical
and life: comprehensive view of life systems from a GNC
basis of the genetic code origin: stereochemical analysis of
SNS primitive genetic code hypothesis, J. Biosci., 27, 165
interactions of amino acids and nucleotides based on the
186, doi: 10.1007/BF02703773.
progene hypothesis, Mol. Biol. (Mosk), 22, 1411 1429.
118. Ikehara, K. (2005) Possible steps to the emergence of life:
106. Fox, G. E. (2010) Origin and evolution of the ribosome,
the [GADV] protein world hypothesis, Chem. Record, 5,
Cold Spring Harb. Perspect. Biol.,
2, a003483,
107 118, doi: 10.1002/tcr.20037.
doi: 10.1101/cshperspect.a003483.
119. Ikehara, K., Omori, Y., Arai, R., and Hirose, A. (2002)
107. Altstein, A. D. (2015) The progene hypothesis: the nucleo
A novel theory on the origin of the genetic code: a GNC
protein world and how life began, Biol. Direct, 10, 67,
SNS hypothesis, J. Mol. Evol.,
54,
530538,
doi: 10.1186/s13062 015 0096 z.
doi: 10.1007/s00239 001 0053 6.
108. Lambowitz, A. M., and Zimmerly, S. (2011) Group II
120. Trifonov, E. N. (2004) The triplet code from first princi
introns: mobile ribozymes that invade DNA, Cold Spring
ples, J. Biomol. Struct. Dyn., 22, 1 11, doi: 10.1080/
Harb. Perspect. Biol., 3, a003616, doi: 10.1101/cshperspect.
07391102.2004.10506975.
a003616.
121. Ikehara, K. (2014) [GADV] protein world hypothesis on
109. Kunin, V. (2000) A system of two polymerases - a model
the origin of life, Orig. Life Evol.,
44,
299302,
for the origin of life, Orig. Life. Evol. Biosph., 30, 459 466,
doi: 10.1007/s11084 014 9383 4.
doi: 10.1023/a:1006672126867.
122. Segre, D., Ben Eli, D., Deamer, D. W., and Lancet, D.
110. Альтштейн А. Д., Каверин Н. Н. (1980) О происхожде
(2001) The lipid world, Orig. Life Evol., 31, 119 145,
нии вирусных генетических систем, Журн. Всесоюз.
doi: 10.1023/a:1006746807104.
хим. общ.им. Д.И. Менделеева, 25, 383 390.
123. Mallik, S., and Kundu, S. (2012) The lipid-RNA world,
111.
Альтштейн А. Д. (1987) Происхождение генетической
arXiv preprint arXiv:1211.0413.
системы: гипотеза прогенов, Мол. Биол., 21, 309 321.
124. Lancet, D., Zidovetzki, R., and Markovitch, O. (2018)
112. Li, L., Francklyn, C., and Carter, C. W., Jr.
(2013)
Systems protobiology: origin of life in lipid catalytic net
Aminoacylating urzymes challenge the RNA world
works, J. R. Soc. Interface, 15, doi: 10.1098/rsif.2018.0159.
hypothesis, J. Biol. Chem.,
288,
2685626863,
125. Krebs, J. E., Goldstein, E. S., and Kilpatrick, S. T. (2017)
doi: 10.1074/jbc.M113.496125.
Lewin’s genes XII, Jones & Bartlett Learning.
113. Rodin, A. S., Rodin, S. N., and Carter, C. W., Jr. (2009)
126. Miller, S. L., Schopf, J. W., and Lazcano, A.
(1997)
On primordial sense antisense coding, J. Mol. Evol., 69,
Oparin’s “Origin of Life”: sixty years later, J. Mol. Evol.,
555 567, doi: 10.1007/s00239 009 9288 4.
44, 351 353.
THE ORIGIN OF THE GENETIC CODE AND TRANSLATION
WITHIN CURRENT CONCEPTS OF THE ORIGIN OF LIFE
Review
L. G. Kondratyeva1, М. S. Dyachkova2, and А. V. Galchenko3*
1 Shemyakin-Ovchinnikov Institute of bioorganic chemistry, Russian Academy of Sciences, 117997 Moscow, Russia
2 Vavilov Institute of General Genetics, 119991 Moscow, Russia
3 Peoples’ Friendship University of Russia (RUDN University), 117198 Moscow, Russia; e>mail: gav.jina@gmail.com
The origin of the genetic code and translation system is perhaps the central and the most difficult problem in study
ing the origin of life and one of the most difficult in evolutionary biology at all. There are plenty of hypotheses for the
emergence and development of present genetic systems, considering the origin and early evolution of the genetic
code, as well as the emergence of replication and translation. The most famous hypotheses are discussed in this review.
However, none of these hypotheses describes without gaps and assumptions all stages of the early evolution of genet
ic systems. The RNA world hypothesis is the dominant scientific idea of the early evolution of biological and pre bio
logical objects for now. Its` main advantage is that it offers RNA as the first living systems to be self sufficient. Namely,
RNA are capable of functioning as a catalytic component of the system, and at the same time as a template. However,
there are also significant limitations. In particular, ribozyme processive polymerase has not yet been discovered or syn
thesized. Taking into account the proteins and nucleic acids mutual need in each other in the present world, many
authors propose scenarios of early evolution based on the co evolution of these two classes of organic molecules.
Hypotheses of this kind postulate that the emergence of translation was necessary for the replication of nucleic acids,
in contrast to the world of RNA, where the emergence of translation was preceded by the era of self replicating RNA.
And although such scenarios are less economical from the point of view of the evolution, since they require the simul
taneous emergence and evolution of two classes of organic molecules, as well as synchronization of replication and
translation emergence, their great advantage is that they offer the development of a processive and much more accu
rate protein dependent replication.
Keywords: evolution, origin of life, translation, genetic code, RNA world, progene, protein world, lipid world
БИОХИМИЯ том 87 вып. 1 2022
5*