БИОХИМИЯ, 2022, том 87, вып. 1, с. 68 - 85
УДК 575.112
КОМПЬЮТЕРНОЕ МОДЕЛИРОВАНИЕ ЛИГАНД РЕЦЕПТОРНЫХ
ВЗАИМОДЕЙСТВИЙ МАННОЗНОГО РЕЦЕПТОРА CD206
В СРАВНЕНИИ С МОДЕЛЬНЫМ ЛЕКТИНОМ КОНКАНАВАЛИНОМ А
© 2022
И.Д. Злотников, Е.В. Кудряшова*
Московский государственный университет имени М.В. Ломоносова, химический факультет,
119991 Москва, Россия; электронная почта: helenakoudriachova@yandex.ru
Поступила в редакцию 08.08.2021
После доработки 07.12.2021
Принята к публикации 16.12.2021
С использованием методов молекулярной динамики и нейросетевого анализа впервые проведено компью
терное моделирование комплексообразования моно и олигосахаридных лигандов с основным (четвёртым)
углеводсвязывающим доменом маннозного рецептора CD206 (CRD4), а также с модельным рецептором
конканавалином А (ConA). Показано, что ConA является релевантной моделью CD206 (CRD4) вследствие
сходства структурной организации сайтов связывания и высокой корреляции значений свободных энергий
комплексообразования r > 0,9, согласно литературным данным и компьютерному моделированию. Обсуж
дается роль основных факторов, влияющих на аффинность взаимодействий лиганд-рецептор: количество
и природа углеводных остатков, наличие Me группы в O1 положении, тип гликозидной связи в диманнозе.
Показано, что комплексообразование ConA и CD206 с лигандами энергетически обусловлено электроста
тическими взаимодействиями заряженных остатков (Asn, Asp, Arg) с атомами кислорода и водорода в угле
водах; меньший вклад вносит гидрофобная и ван дер ваальсова составляющая. Рассмотрен возможный ва
риант дополнительной стабилизации комплексов за счёт CH π стекинг взаимодействий Tyr с плоскостью
Man остатка. Изучена роль ионов кальция и марганца в процессах связывания лигандов. Рассчитанные в
ходе молекулярной динамики свободные энергии комплексообразования коррелируют с эксперименталь
ными данными (опубликованными для модельного ConA): коэффициент корреляции r = 0,68. Проведено
обучение нейросети Pafnucy на основе набора комплексов лиганд-рецептор PDBbind2020, что позволило
увеличить точность предсказаний энергий до r = 0,8 и 0,82 для рецепторов CD206 и ConA соответственно.
Предложена модель нормирования значений энергий комплексообразования для вычисления релевантных
значений ΔGbind и Kd. На основе разработанной методики определены значения констант диссоциации се
рии комплексов CD206 с 9 углеводными лигандами различной структуры, ранее не охарактеризованными.
Полученные данные открывают перспективы применения компьютерного моделирования для разработки
оптимальных носителей лекарств с функцией активного нацеливания на макрофаги, а также определяют
границы применимости использования ConA в качестве релевантной модели для исследования параметров
связывания CD206 с различными углеводными лигандами.
КЛЮЧЕВЫЕ СЛОВА: молекулярная динамика, искусственная нейросеть, CD206, конканавалин А,
макрофаги.
DOI: 10.31857/S0320972522010055
ВВЕДЕНИЕ
маннозного (Man) рецептора макрофагов (МФ)
с углеводными лигандами, что необходимо для
В представленной работе исследованы воз
конструирования носителей лекарств с функци
можности метода молекулярной динамики и ис
ей адресного нацеленного на маннозные рецеп
кусственной нейросети Pafnucy [1] для выявле
торы (МР) макрофага.
ния закономерностей в комплексообразовании
Важной задачей для создания эффективных
методик лечения различных заболеваний, в том
числе болезней дыхательных путей, является не
Принятые сокращения: МД - молекулярная дина
мика; МР - маннозный рецептор; МФ - макрофаги; НС -
только разработка новых лекарственных
нейросеть; ConA - конканавалин А; CRD - углеводрас
средств, но и их направленная доставка к пора
познающий домен; diMan - α D маннопиранозил α D
женным тканям и клеткам [2]. Целевым объек
манноза; E - электростатические взаимодействия; Man - том могут служить МФ, которые играют важную
манноза; MeMan - метил α D маннопиранозид; NP - не
роль в иммунном ответе за счёт распознавания и
полярная сольватация; P
- полярная сольватация;
triMan - триманнозид; vdW - ван дер ваальсовы взаимо
уничтожения патогенов [3]. В то же время МФ
действия.
могут служить резервуаром для роста и размно
* Адресат для корреспонденции.
жения бактерий и вирусов, чем сильно ограни
68
МОДЕЛИРОВАНИЕ ConA, CD206 - МАННОЗНЫЕ ЛИГАНДЫ
69
чивают эффективность антибиотико и антиви
домена (CRD4) [13-14]. Поэтому в данной ра
русной терапии [4-5]. Антигенпрезентующие
боте для моделирования свойств МР и выявле
клетки, в том числе МФ, вносят главный вклад в
ния наиболее специфичных маннозных лиган
создание иммуносупрессивного микроокруже
дов использовался CRD4. Следует отметить,
ния опухолей, при этом снижают иммунный от
что, по результатам экспериментальных дан
вет [6]. Перспективный путь решения этой
ных [15], параметры связывания олигосахари
проблемы - реполяризация иммуносупрессор
дов парой доменов CRD(4-5) во многом совпа
ных опухоль ассоциированных макрофагов
дают с таковыми для целого МР CD206, что по
(М2) в провоспалительные (М1) [7]. Напротив,
казано на примере олигосахарида Man23
избыточная провоспалительная активность
(23 маннозных остатка в молекуле).
макрофагов является причиной аутоиммунных
Однако, поскольку сами МР труднодоступ
заболеваний (ревматоидный артрит, остеоарт
ны, а экспериментальных данных по комплек
рит, рассеянный склероз). Специфическая ак
сам CD206 в литературе крайне мало, для опре
тивация макрофагов предполагается эффектив
деления основных тенденций в связывании
ным инструментом для лечения нейродегенера
маннозных лигандов часто используются мо
тивных заболеваний [8]. Исходя из вышепере
дельные белки рецепторы. Предполагается, что
численного, адресная доставка в макрофаги ан
CD206 демонстрирует высокое сходство с кон
тибактериальных препаратов или веществ, спо
канавалином А (ConA) в закономерностях свя
собных регулировать статус активации МФ в за
зывания углеводов [3]. Одной из задач данной
висимости от задач лечения [9], представляется
работы является сравнение указанных рецепто
перспективным подходом для повышения эф
ров для выявления границ применимости ConA
фективности лекарственной терапии различных
в качестве модели МР. ConA состоит из четырёх
заболеваний.
субъединиц (26,5 кДа, 237 а.о. каждая), сильно
Воздействовать на макрофаги потенциально
гликирован, связывает углеводы в присутствии
возможно с помощью связывания биоактивных
катионов металлов (обычно Mn2+ и Ca2+)
молекул с рецепторами распознавания образов
[16-17]. Катион Ca2+ координирует аминокис
(pattern recognition receptors), включающими
лотные остатки и «подготавливает» рецептор к
toll подобные рецепторы (связывают гликоли
распознаванию углеводов: Ca2+ в CD206, в отли
пиды, липополисахариды, липотейхоевые кис
чие от конканавалина А, участвует в непосред
лоты, пептидогликаны), рецепторы комплемен
ственном комплексообразовании с лигандом.
та (распознают опсонизирующие фрагменты
В сайте ConA присутствует второй катион, в
С3b, iC3b и C3dg), рецепторы мусорщики (рас
данном случае Mn2+ (иногда встречается Co2+,
познают глико и липопротеины) и лектиновые
Ni2+, Zn2+), который фиксирует положение Ca2+,
рецепторы [3]. Воздействие на последние при
снижая подвижность аминокислотных остатков
таргетировании макрофагов позволяет активи
в сайте связывания. Включение обоих катионов
ровать целевые ветви T хелперных лимфоцитов,
в ConA необходимо для активирования связы
Th1, Th2 или Th17 [10-11].
вающей способности белка [18-19].
Примечательно, что нацеливание на МФ че
Поиск высокоаффинных взаимодействий
рез задействование маннозных рецепторов, ко
лиганд-рецептор может быть выполнен с по
торые распознают Man , Fuc и GlcNAc остат
мощью различных физико химических мето
ки олигосахаридов клеточной стенки патоген
дов [20]: изотермическая калориметрия титро
ных микроорганизмов [12], имеет ряд преиму
вания [21-22], флуоресцентные методы [23-24],
ществ в сравнении со стратегией таргетирова
ИК спектроскопия Фурье [25], метод ингиби
ния иных рецепторов: воздействие только на бо
рования Ландштейнера [26]. Применяются так
лезнетворные макрофаги, снижение риска ре
же проточная цитометрия для определения
зистентности и осложнений и повышенная эф
экспрессии антигенов МФ и путей их актива
ции [27], конфокальная микроскопия для визуа
Основным маннозным рецептором МФ являет
лизации клеточного поглощения носителей ле
ся CD206 - лектин С типа, который представ
карств [28]. Активно развивающимся методом
ляет собой трансмембранный белок (175 кДа).
является компьютерное моделирование. Моле
C Концевая часть CD206 содержит корот
кулярная динамика (МД) и искусственные ней
кую (45 а.о.) цитоплазматическую часть [3].
росети (НС) позволяют моделировать разнооб
Связывание углеводов МР происходит за счёт
разные химические процессы, включая лиганд
лектиноподобного участка С типа, состоящего
рецепторные взаимодействия. Методы in silico
из 8 доменов. Однако способность связывать уг
делают возможным изучение широкого круга
леводы изолированно от других частей доказана
соединений с вариабельной структурой и прост
только для четвёртого углеводраспознающего
ранственной организацией, синтез которых в
БИОХИМИЯ том 87 вып. 1 2022
70
ЗЛОТНИКОВ, КУДРЯШОВА
лаборатории является трудоёмким, требует зна
поставимой с квантово химическими расчёта
чительных временных затрат и, зачастую, не
ми, при значительном снижении вычислитель
всегда возможен.
ных затрат симуляции лиганд рецепторных взаи
МД позволяет изучать механизмы взаимо
модействий (например, фосфодиэстераза 5А с
действия белков рецепторов с лигандами: пути
тадалафилом). НС способна предсказывать по
протекания [29-31] и кинетику процессов дис
ложение рецептора и лиганда при связывании, а
социации комплексов [32], а также связывание
также величины энергии Гиббса (с большей кор
лигандов с рецептором [33-35]. МД даёт воз
реляцией с экспериментальными данными) для
можность оценить параметры комплексообразо
ряда биомолекул, например, биотин карбокси
вания: свободную энергию взаимодействия,
лаза, дегидрофолат редуктаза [42]. С помощью
вклад водородных связей, ван дер ваальсо
НС и технологий искусственного интеллекта
вых (vdW) и электростатических (Е) взаимодей
выполнен поиск потенциальных лекарств среди
ствий, энергии растворения и десольватации [33,
используемых в медицине или проходящих кли
36-37]. МД является хорошим инструментом
нические испытания: фалдапревир - гепатит C,
для изучения и уточнения кристаллических
иммунодепрессант циклоспорин, ремдисевир,
структур белков и их комплексов, а именно: мо
мефупариб и другие. Данные терапевтические
делирование выявляет значимые взаимодей
агенты могут быть перепрофилированы для
ствия лиганд-рецептор, которые отсутствуют в
снижения вирусной нагрузки, облегчения симп
кристалле [34]. МД выявляет взаимосвязь гидро
томов COVID 19 и эффективного лечения ви
фобности/гидрофильности лиганда, его формы
русной инфекции [43].
(вытянутая, Т образная, изогнутая) со стабиль
Однако методы in silico, в том числе техноло
ностью комплекса и кинетикой связывания [38].
гия нейросетей, для исследований непосред
Однако МД подход требует значительных вы
ственно биосистем только начинают использо
числительных мощностей и времени моделиро
ваться учёными. В то же время значительный
вания, что обоснованно в случаях, когда важно
потенциал компьютерного моделирования уже
выявление механизма взаимодействия исследуе
продемонстрирован в ряде работ [44-46]. По
мых молекул. В то же время для проведения вы
этому одной из задач данной работы является
сокопродуктивного скрининга широкого круга
демонстрация перспективности использова
лигандов комплементарным методом (или заме
ния НС, а также МД для изучения лиганд ре
ной МД) оценки сродства лигандов к рецептору
цепторных взаимодействий моно и олигосаха
является искусственная нейросеть.
ридов с CRD4 маннозного рецептора CD206, а
Известно несколько нейросетей, предсказы
также с модельным лектином конканавали
вающих константы диссоциации (ассоциации)
ном А. Для ряда комплексов ConA-лиганд в ли
комплексов лиганд-рецептор или константы
тературе определены экспериментальные конс
ингибирования на основе базы данных
танты, что позволило валидировать и оптимизи
PDBbind: SE OnionNet [39], Pafnucy [1], Nnscore
ровать методику моделирования и на этой осно
2.0 [40]. Главное преимущество нейросетей пе
ве изучить взаимодействия серии лигандов с
ред МД - малое время работы (несколько минут
четвёртым углеводсвязывающим доменом
против дней недель моделирования), а также
(CRD4) маннозного рецептора CD206, для ко
альтернативный принцип работы НС, который
торого параметры комплексов ранее не были
заключается в «сравнении» данной пары ли
описаны.
ганд-рецептор с известными из литературных
данных (PDBbind) по целому ряду (в данной ра
боте по 19) параметров, включая типы составляю
МАТЕРИАЛЫ И МЕТОДЫ
щих атомов, валентности, гибридизации, заря
ды. Точность предсказания аффинности зависит
Структуры комплексов. Пространственные
от «обученности» сети, количества слоёв и ней
структуры молекул рецептора ConA и 15 угле
ронов в сети и сложности алгоритма. Стоит от
водных лигандов были построены с использова
метить, что компьютерные методы активно со
нием кристаллографических данных PDB (таб
вершенствуются, что открывает перспективы
лица). Варьировали структуру и пространствен
использования МД и НС как значимых инстру
ную организацию лигандов 1-15: тип углевод
ментов в биохимии.
ных остатков, количество Man и GlcNAc ос
В последнее время в литературе появился
татков в углеводе (от 1 до 5), наличие Me груп
ряд работ, ярко демонстрирующих потенциал
пы в O1 положении углеводов, тип гликозидной
нейросетей. Так, в работе Lahey и Rowley [41] с
связи (α1→2, 1→3, 1→6). Рассмотрены 4 субъ
использованием НС предсказана стабильность
единицы белка по 237 а.о. каждая, катионы Ca2+
молекулярных конформаций с точностью, со
и Mn2+ в сайтах связывания лигандов, кристал
БИОХИМИЯ том 87 вып. 1 2022
МОДЕЛИРОВАНИЕ ConA, CD206 - МАННОЗНЫЕ ЛИГАНДЫ
71
лографическая вода. Структура четвёртого угле
ектории 3-5 нс при концентрации соли 1,0 М
водсвязывающего домена маннозного рецепто
для минимизации неспецифических электро
ра CD206 построена на основе кристаллографи
статических взаимодействий с использованием
ческих данных PDB (Acc. No. 7JUF). Рецептор
моделей generalized born и Пуассона-Больцмана.
состоит из 135 а.о. и катиона Ca2+. Для Ca2+
Нейросетевой анализ комплексов. Выбор ней
и Mn2+ использовали несвязанную модель 12 6 4
росети Pafnucy [1] обусловлен её лидирующей
потенциала Леннарда-Джонса. Структуры ли
позицией в рейтинге алгоритмов по предсказы
гандов к CD206 сгенерированы и подведены к
ванию аффинностей рецептор-лиганд (CASF
сайту связывания вручную с применением сред
2016). С помощью НС Pafnucy анализировались
все пространственные структуры комплексов
(таблица).
рецептор-лиганд, полученные после проведе
Структуры подготовлены с использованием
ния МД. В качестве входных файлов использо
tLeaP из AmberTools20, 21 [47]: добавлены не
вались структуры, усреднённые по 30 фреймам
достающие атомы водорода, ионы Na+ для нейт
заключительной части траектории (ConA), а
рализации общего заряда комплекса. Система
также единичные структуры из траектории, ми
была сольватирована молекулами воды
нимизированные с использованием 10 000 ша
TIP3P [48] с минимальным расстоянием между
гов алгоритма наискорейшего спуска с последую
границей ячейки и белком 10 Å. В среднем раз
щими 10 000 шагами сопряжённого градиентно
мер ячейки с тетрамером ConA составлял
го алгоритма (CD206). С помощью prepare.py
90 × 95 × 90 Å3 с доменом CD206 60 × 65 × 60 Å3.
подготовлен набор из 30 комплексов (по 15 на
Силовые поля Amber ff14SBonlysc, GLYCAM_06j1
рецептор) в формате hdf.
использовались для описания взаимодействий
Использованы 2 конфигурации нейросети
атомов.
Pafnucy: 1) исходная НС, обученная на осно
Проведение молекулярной динамики. Моде
лирование проводилось с использованием паке
та молекулярной динамики Amber20. Системы
2) нейросеть на основе п.1, с обучающим набо
(комплексы рецепторов с лигандами) в водном
ром на основе PDBbind2020, включая 14 107
растворе сначала были минимизированы с ис
комплексов в general set, 5109 - validation, 226 -
пользованием 5000 шагов алгоритма наискорей
core set. Подготовлена авторами настоящей
шего спуска с последующими 5000 шагами соп
статьи с использованием pdbbind data.ipynb и
ряжённого градиентного алгоритма. Систему
скриптов split_dataset.py, training.py. Обучение
нагревали от 0 до 300 К в течение 0,1 нс по моде
проводилось с параметрами, установленными
ли Ланжевена с использованием частоты столк
по умолчанию, кроме числа циклов (эпох) рав
новений 2,0 пс-1 и при периодических гранич
ного 25 (рис. S1, табл. S1 в Приложении).
ных условиях постоянного объёма. Затем была
Ошибка проверки (validation error) снижается
проведена симуляция 100 пс с постоянным дав
примерно до пятой эпохи, далее оптимизирует
лением 1 атм. Далее - уравновешивание систе
ся точность только для обучающей выборки.
мы в течение 100 пс и непосредственно МД (pro
НС предсказывает значение Kd или Ki на ос
duction). Для каждой модели выполнено три не
нове анализа 19 характеристик атомов: тип ато
зависимых цикла с поддержанием температуры
ма (B, C, N, O, P, S, Se, галоген или металл (9 ва
на уровне
300
К термостатом Ланжевена
риантов)), гибридизация и валентность (3 вари
(ntt = 3). В случае тетрамера ConA проводилась
анта), связи с тяжёлыми или гетероатомами,
МД 10 нс (cut off 8 Å, шаг 2 фс). Моделирование
дополнительные параметры (гидрофобность,
четвёртого домена МР CD206 проводили
ароматичность, акцепторные/донорные свой
50-250 нс (cut off 8 Å, шаг 2 фс). Увеличение
ства, входит в состав цикла), частичный заряд и
длительности моделирования для маннозного
принадлежность атома рецептору (-1) или ли
рецептора обусловлено неточностью начальных
ганду (1).
координат, в некоторых случаях снижение шага
Математическая обработка данных. Значения
до 0,5-1,0 фс предпринято во избежание техни
свободных энергий рассчитаны на основе трёх
ческих ошибок. Все связи, содержащие атомы
независимых циклов моделирования путём ус
водорода, были ограничены алгоритмом
реднения, для ConA тетрамера расчёты прове
SHAKE [49].
дены по четырём субъединицам и затем усред
Анализ траекторий и топологических файлов
нены. Нормирование полученных данных на
комплексов. Анализ траекторий выполнен с по
экспериментальные проводили по уравнению:
мощью MMPBSA.py [50] и cpptraj [51]. Расчёт
ΔGbind = ΔGbind (исх) k + b (ккал/моль). В случае
энергий комплексообразования проведён на ос
ConA: k = 0,1125 и b = 2,07 (R2 = 0,71) по методу
нове равновесной (заключительной) части тра
Пуассона-Больцмана для МД; k = 2,1961 и
БИОХИМИЯ том 87 вып. 1 2022
72
ЗЛОТНИКОВ, КУДРЯШОВА
b = -9,05 (R2 = 0,67) - оптимизация нейросети.
для ConA, полученными методом изотермичес
Для CRD4 CD206 использовались коэффициен
кой калориметрии [21], и для домена МР CD206
ты: k = 0,0137 и b = 2,99 (R2 = 0,47) по методу
по сдвигам в спектрах ЯМР [52-53] и конкурент
Пуассона-Больцмана МД; k = 0,3855 и b = 1,33
ному связыванию (рис. 1) [13]. Абсолютные зна
(R2 = 0,78) - НС. Более высокая корреляция
чения свободных энергий ожидаемо не совпада
данных НС по рецептору ConA достигнута при
ют, однако прослеживается достаточная высо
нормировании не энергий, а констант:
кая для методов in silico степень корреляции, что
Kd = 8,24⋅Kd(исх) + 10-5 (М).
визуально можно определить из сходства зави
симостей свободных энергий (рис. 1). Коэффи
циент корреляции Пирсона с литературными
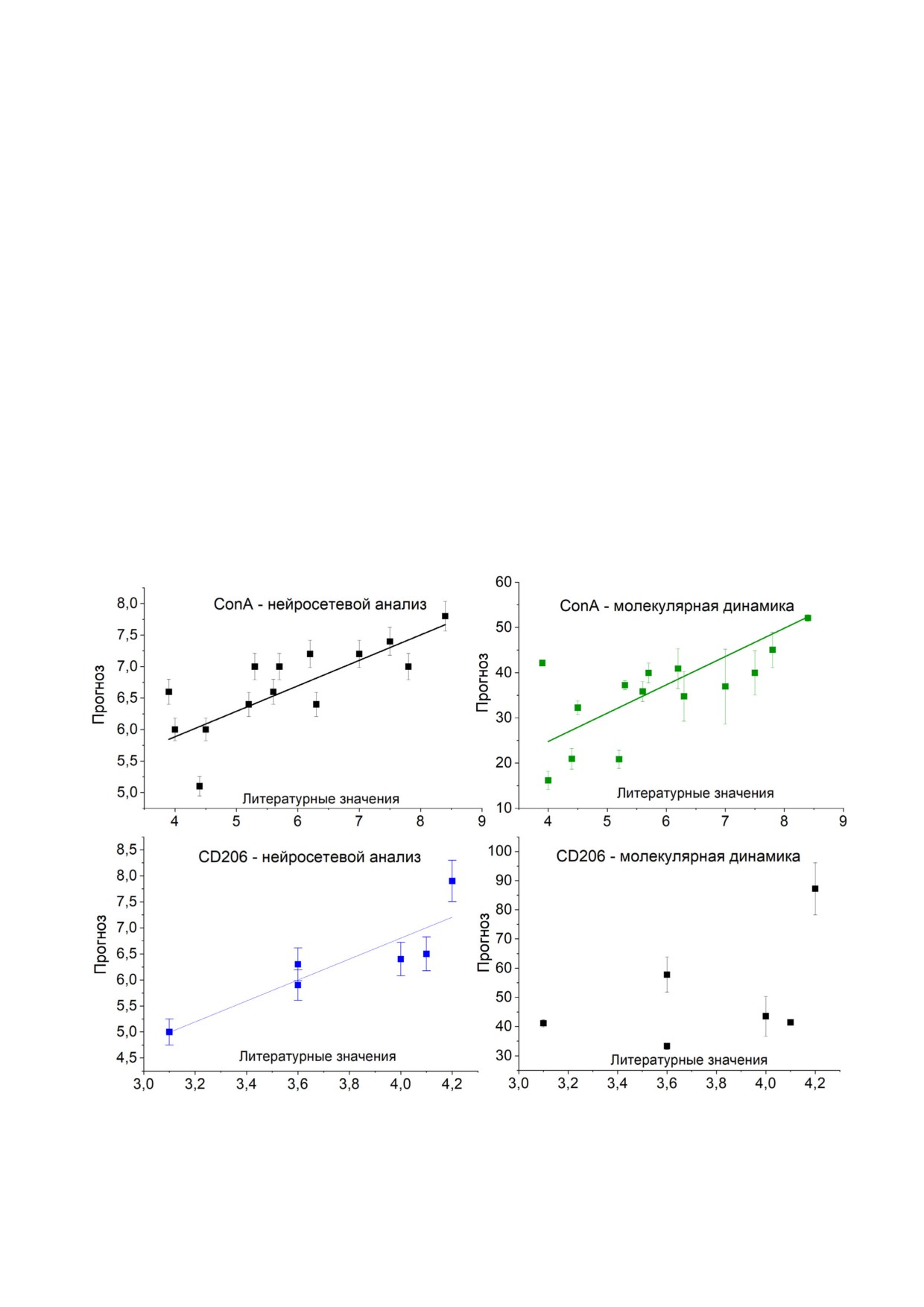
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
данными r = 0,82 для ConA и r = 0,58 для CD206,
что достаточно с учётом малой изученности
Энергетические характеристики комплексооб
маннозного рецептора. Причём наиболее приб
разования ConA и домена CD206 с углеводными
лиженными к экспериментальным оказались
лигандами. Для определения параметров лиган
значения, полученные в ходе нейросетевого
да, увеличивающих сродство к маннозным ре
анализа комплексов рецептор-лиганд пос
цепторам, проведено моделирование взаимо
ле МД. CD206 практически невозможно полу
действия углеводов, состоящих из Man , Gal ,
чить в виде, пригодном для скрининга in vitro, а
Glc , Fuc или GlcNAc остатков (рис. 1, табли
следовательно, компьютерные методы приме
ца, табл. S2 и S3 в Приложении). Релевантность
нительно к данным системам обладают значи
определённых в ходе моделирования значений
тельным потенциалом.
констант и свободных энергий подтверждена
Отметим, что экспериментальные литера
путём сравнения с литературными данными турные данные по комплексообразованию МР
Рис. 1. Энергии комплексообразования ConA и домена маннозного рецептора CRD4 CD206 с углеводными лигандами,
рассчитанные с помощью компьютерного моделирования, в сравнении с литературными данными [13, 20, 21, 26, 52, 53].
Для ConA представлены усреднённые по субъединицам значения. T = 300 К. pH = 7. Концентрация NaCl - 1M
БИОХИМИЯ том 87 вып. 1 2022
МОДЕЛИРОВАНИЕ ConA, CD206 - МАННОЗНЫЕ ЛИГАНДЫ
73
Константы диссоциации комплексов ConA и домена маннозного рецептора CRD4 CD206 с углеводными лигандами
Kd (ConA-лиганд), мкM
*Kd (CD206 CRD4-лиганд), мкM
Номер
Название лиганда, код PDB комплекса
с ConA и номер иллюстрации
Молекулярная
Лит.
Нейросеть
Молекулярная
Лит.
Нейросеть
динамика
данные
динамика
данные
1
α маннопираноза [5CNA]; рис. 5
1450
1100
360
3450
-
3700
и рис. S5 в Приложении
2
Me маннопиранозид [5CNA]; рис. 5, 7,
600
160
190
2900
2400
2360
и рис. S3, S5 в Приложении
3
Me α(1→2) диманнопиранозид
30
7
60
1700
-
20
[1BXH]; рис. 4
4
α(1→2) диманнопиранозид [1BXH];
43
24
190
2290
1280
1700
рис. S4 в Приложении
5
Me α(1→3) диманнопиранозид
14
30
56
1800
-
2860
[1QDO/C]; рис. 4
6
α(1→3) диманнопиранозид; рис. S4
16
71
78
1600
2290
1800
в Приложении
7
α(1→6) диманнопиранозид; рис. S4, S7
35
75
145
2400
1010
1600
в Приложении
8
Me α(1→6) диманнопиранозид; рис. 4,
27
120
80
1500
-
1500
рис. S2 в Приложении
9
Me 3,6 ди O (маннопиранозил) α
6
2
70
1080
-
2860
маннопиранозид [1ONA]
10
3,6 ди O (маннопиранозил) α манно
16
3
40
780
-
650
пираноза; рис. 2
11
3,6 ди O (β(1→2) N ацетилглюкозами
2
0,7
30
480
-
1000
но маннопиранозил) α маннопираноза
[1TEI]; рис. S6 в Приложении
12
Me α глюкопиранозид [1GIC]; рис. S2
590
560
200
>6000*
>6000*
>6000*
в Приложении
13
Me α галактопиранозид [1GIC]; рис. 3,
70
500
370
>6000*
>6000*
>6000*
рис. S2 в Приложении
14
Me GlcNAc
11
1350
150
2400
5300
4200
15
Fuc α(1→3) GlcNAc
3
-
170
2300
–
1400
Примечание. Значения рассчитаны с помощью компьютерного моделирования, в сравнении с литературными данными
по методам изотермической калориметрии [21] и ингибированию Ландштейнера [26], а также рассчитанные по сдвигам в
спектрах ЯМР [52-53] и конкурентному связыванию [13]. T = 300 К. pH = 7. Концентрация NaCl - 1M.
* Неспецифическое взаимодействие.
описаны лишь для ограниченного круга лиган
(CRD4) нами рассчитаны 9 значений констант,
дов: по ЯМР сдвигам и конкурентному связы
ранее не опубликованных в статьях.
ванию [13, 52, 53] - 4 из 15 рассмотренных в
Роль МД в оптимизации структуры комплек
данной работе. Компьютерное моделирова
сов. Несмотря на то что для 9 комплексов ConA
ние CD206 или его углеводсвязывающего доме
и 4 комплексов CD206 с лигандами в качестве
на ранее не проводилось, кроме работы
исходных координат использовались структуры
Asciutto et al. [54], в которой изучалась часть
из PDB, компьютерное моделирование даже та
МР CysRD CTLD2/3 (домены, которые не иг
ких систем позволяет значительно оптимизиро
рают важной роли в связывании углеводов [55]).
вать пространственное расположение рецептора
Для домена маннозного рецептора CD206
и лиганда. Экспериментальное определение
БИОХИМИЯ том 87 вып. 1 2022
74
ЗЛОТНИКОВ, КУДРЯШОВА
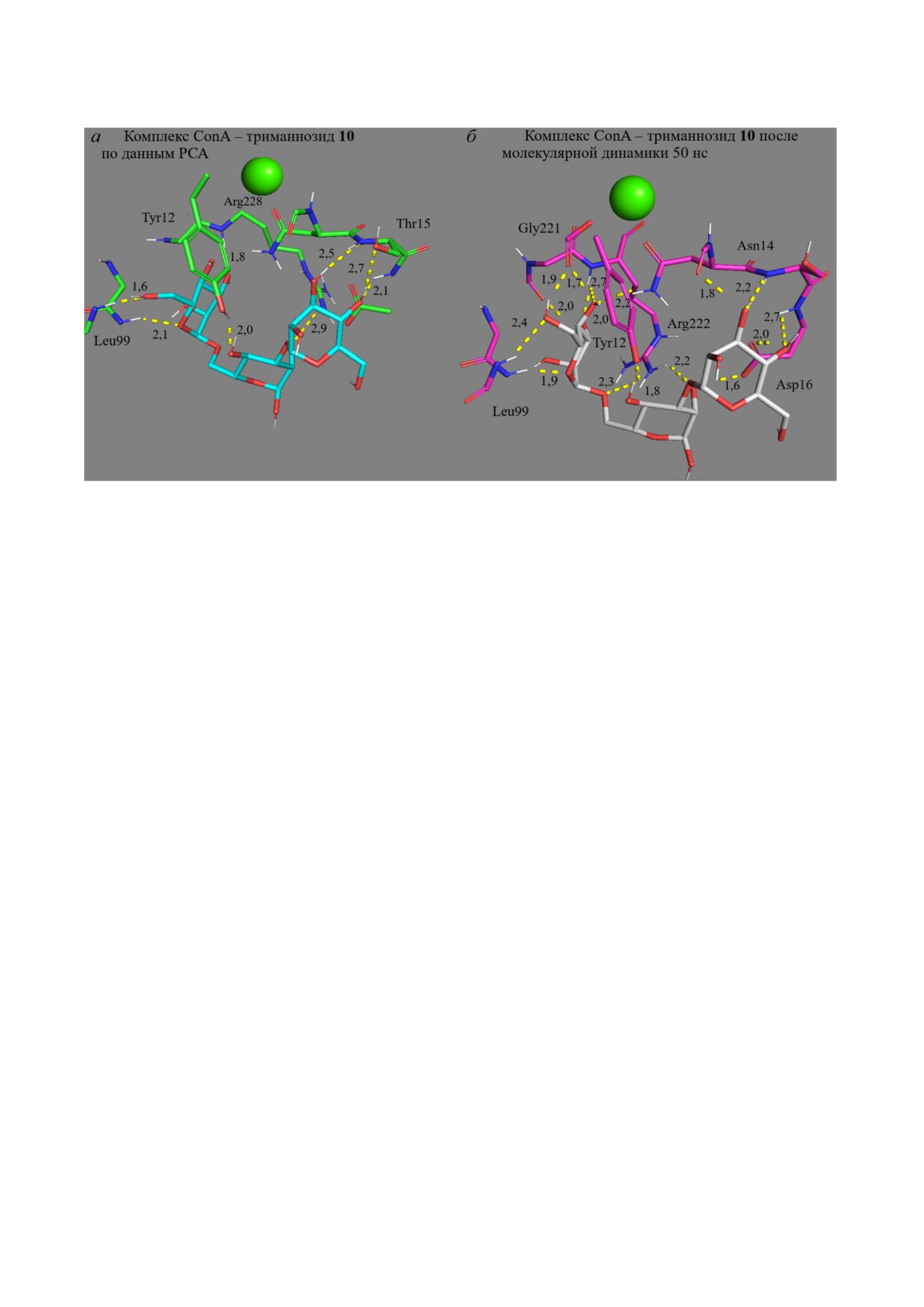
Рис. 2. Поиск оптимального положения лиганда с помощью молекулярной динамики. Показаны комплексы ConA - три
маннозид 10 по данным РСА (а) и после молекулярной динамики 50 нс (б). Зелёный шар - Ca2+
структур комплексов проводится методом рент
лигандов четвёртым доменом МР CD206 пока
геноструктурного анализа (РСА) кристалла [56].
жем на примере взаимодействия диманнозы 8 с
Однако данный метод имеет определенные ог
рецептором (рис. S2 в Приложении). В исход
раничения, связанные с несоответствием усло
ном положении лиганд не образует водородных
вий кристаллизации белка с лигандом и прове
связей с аминокислотными остатками. В ходе
дением экспериментов по комплексообразова
моделирования наблюдается оптимизация
нию, неопределённостью положений атомов во
пространственного расположения диманнозы:
дорода [57] и различием структур рецептора в
маннозный остаток α1 сближается с рецепто
растворе и в кристалле. Метод МД позволяет
ром, происходит связывание с Glu107, Asn121,
устранить эти недостатки и выявить значимые
Hie127 (рис. S2 в Приложении). В то же время
взаимодействия лиганд-рецептор, которые от
остаток α6 проворачивается вокруг гликозид
сутствуют в кристалле [34]. Роль МД в оптими
ной связи, чтобы образовать дополнительные
зации структуры комплекса белок-лиганд про
взаимодействия с МР. Отметим, что при исполь
демонстрирована на рис. 2, где сопоставлены
зовании в качестве исходного положения рецеп
положения триманнозида
10
в комплексе
тора с вручную подведённым лигандом требует
с ConA по данным РСА и после моделирова
ся увеличение времени моделирования в срав
ния 50 нс. При связывании с рецептором три
нении со случаем исходной PDB структуры:
манноза 10 (рис. 2) взаимодействует централь
от 10-30 нс до 50-250 нс. Из рис. S2 в Приложе
ным Man остатком с Arg222 (3 значимые водо
нии следует, что 30 нс в первом случае недоста
родные связи 1,8-2,3 Å). Левый (α1→6) Man
точно для оптимизации структуры комплекса.
остаток координируется бóльшим числом ами
Необходимо не менее 100 нс МД.
нокислот, в том числе Asn14, Gly221 за счёт об
Основные закономерности связывания угле
разования высокоэнергетических водородных
водсодержащих лигандов CRD4 CD206 и модель
связей атомов O4,5,6
(1,7-2,7
Å). Правый
ным лектином ConA. На основании рассчитан
(α1→3) Man остаток сближается с Thr15 на
ных с помощью МД и НС констант диссоциа
0,3 Å и Asp. С помощью моделирования показа
ции комплексов лиганд-рецептор (таблица,
но более энергетически выгодное положение
табл. S2 и S3 в Приложении) [58] и механизмов
лиганда в сайте связывания относительно комп
комплексообразования (рис. 2-5 и рис. S2-S8
лекса PDB (1ONA).
в Приложении) можно выявить следующие за
ConA имеет сходные с CD206 мотивы струк
кономерности. По данным МД, наиболее низ
туры углеводсвязывающего домена. Насколько
кие значения констант диссоциации характер
структурное сходство транслируется в функцио
ны для комплексов рецепторов с биантенными
нальное, показано далее. Механизм связывания
лигандами 9-11 и 15: Kd (ConA - (GlcNAc)2
БИОХИМИЯ том 87 вып. 1 2022
МОДЕЛИРОВАНИЕ ConA, CD206 - МАННОЗНЫЕ ЛИГАНДЫ
75
triMan) = 2 мкМ, Kd (ConA - Fuc α (1→3)
ходима кластеризация бóльшего количества
GlcNAc) = 3 мкМ. Аналогично домен манноз
концевых Man/GlcNAc остатков. Однако, по
ного рецептора образует наиболее прочные
данным калориметрического титрования [21],
комплексы с лигандами 9-11 с Kd 1080, 780 и
гликопептиды Man7-9, которые моделируют
480 мкМ соответственно.
олигосахариды клеточной стенки микроорга
Специфичность рецепторов к типу углеводно
низмов, практически не превосходят в сродстве
го остатка. По данным нейросетевого анализа
к ConA лиганд 11 (производное triMan - три
комплексов, сродство углеводных остатков
маннозида), т.е. существует оптимальное коли
к ConA возрастает в ряду (Gal≈Fuc)
<
чество углеводных остатков, задействованных в
< (Glc≈Man) < GlcNAc, что коррелирует с лите
связывании. Иными словами, модельный лек
ратурными данными [20-21]. Несколько отлич
тин ConA имеет наибольшее сродство к консер
ная закономерность характерна для четвёртого
вативному ядру триманнозида, следовательно,
домена МР CD206: аффинность к лигандам уве
достигнут предел специфичности лиганд-ре
личивается в ряду (Glc≈Gal)
< GlcNAc <
цептор. В случае CRD4 с увеличением количест
< (Fuc≈Man) (таблица). Отметим, что метилиро
ва доступных Man остатков, по данным МД
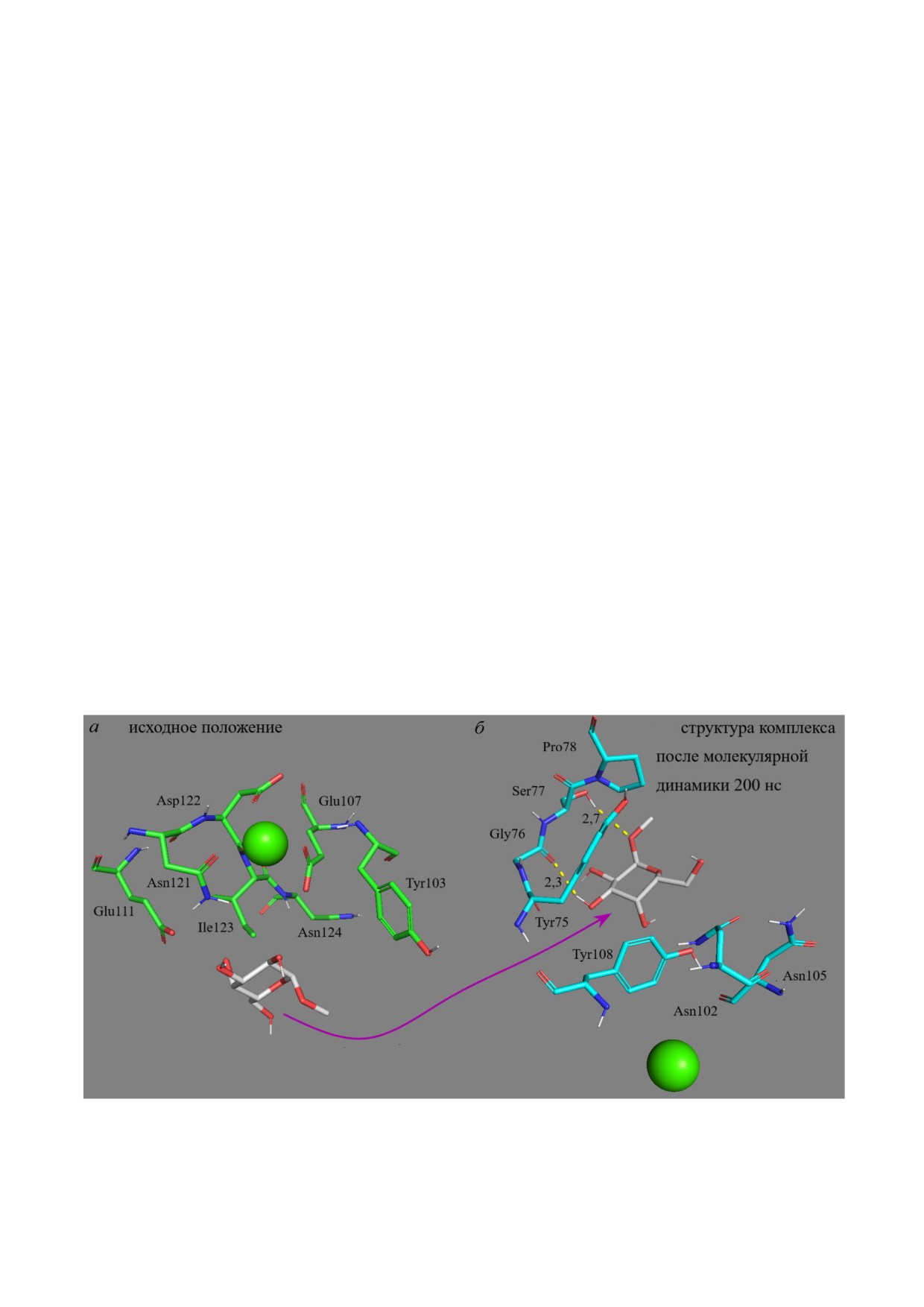
ванные производные α D глюкозы 12 и галак
и НС, сродство лигандов к МР растёт, однако
тозы 13 (рис. 3) крайне слабо взаимодействуют с
менее выражено. Что также следует из данных,
МР, но более аффинно связываются с ConA
полученных методом конкурентного связыва
(рис. S3 в Приложении). В ходе моделирова
ния в работе Feinberg et al. [13]: Man9 специфич
ния CD206 происходит выход лиганда (12, 13) из
нее MeMan более чем на порядок, а триманно
сайта связывания и дальнейшее неспецифичес
зид - всего лишь в 3 раза специфичнее MeMan.
кое взаимодействие с боковыми аминокислот
Аффинность МР к маннозосодержащим лиган
ными остатками (рис. 3). Для остальных иссле
дам возрастает, начиная от 1-2 остатков и
дуемых лигандов наблюдается специфичное
вплоть до крупных олигосахаридов и полимеров
связывание в сайте распознавания углеводов.
(например, Man23, маннан) [15].
Аффинность рецепторов к лигандам в зависи
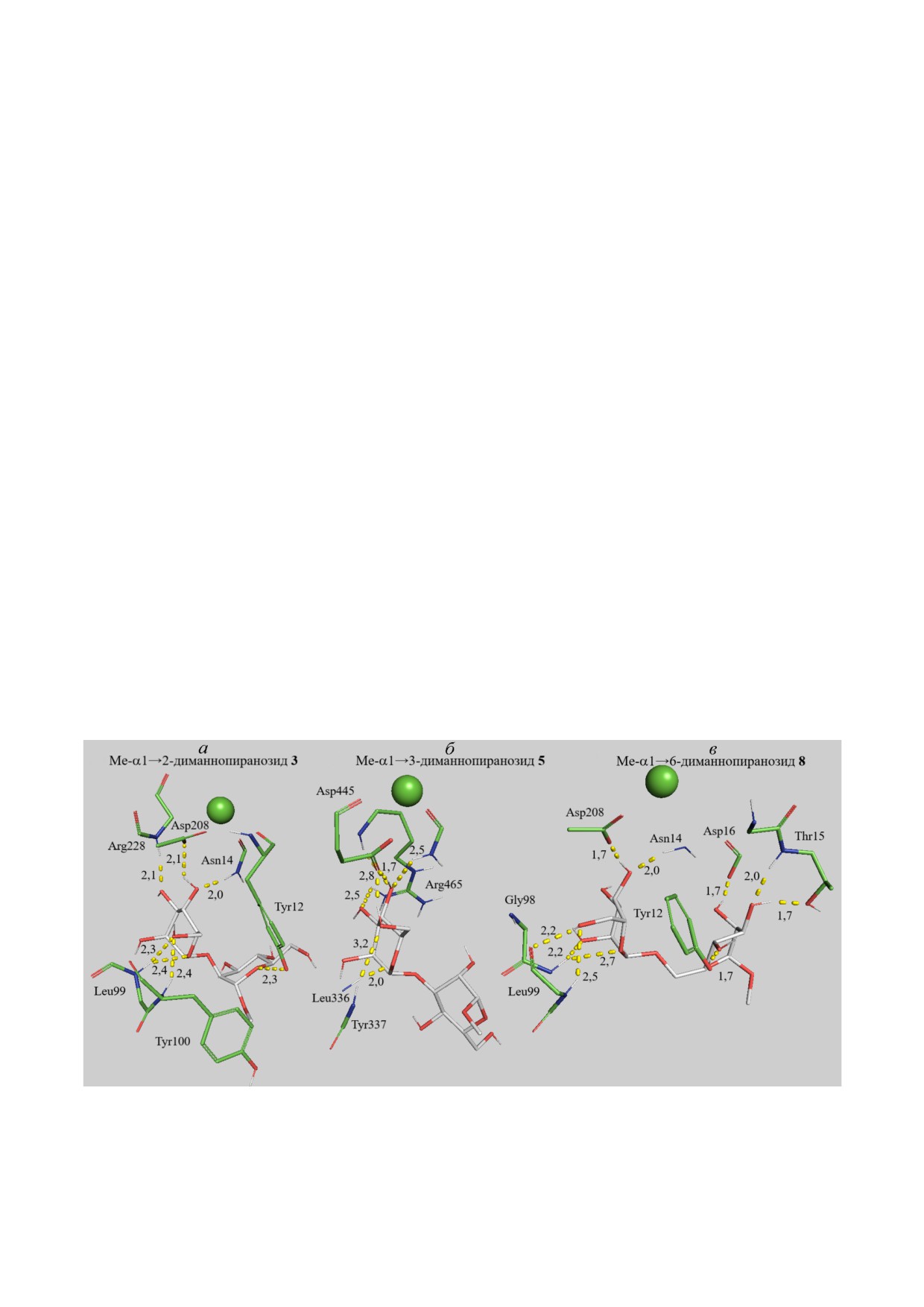
Влияние типа гликозидной связи в диманнози
мости от количества Man остатков. Углеводы,
дах на сродство к рецептору. Рецептор ConA рас
содержащие только одно маннопиранозильное
познаёт диманнозиды 3-8 (рис. 4 и рис. S2
кольцо (рис. S4 в Приложении) - Man 1 и
в Приложении) с аффинностью, возрастающей
MeMan 2 (метил α D маннопиранозид), не об
в ряду: α1→2 < α1→6 < Me α1→6 < α1→3 <<
ладают высокой аффинностью к ConA - необ
<< Me α1→3 ≈ Me α1→2, в котором приведены
Рис. 3. Неспецифическое взаимодействие метил α D галактопиранозида 13 (белый) с доменом маннозного рецепто
ра CD206. а - Исходное положение. б - Структура комплекса после молекулярной динамики 200 нс. Зелёный шар - Ca2+
БИОХИМИЯ том 87 вып. 1 2022
76
ЗЛОТНИКОВ, КУДРЯШОВА
условные обозначения диманнозидов и их мети
тупными для рецептора и повышает аффин
лированных производных (нейросеть, таблица).
ность.
Таким образом, связь Man остатков через атомы
Напротив, биантенным становится α1→2
кислорода у С1 и С2/3 оптимальная. Однако,
диманнопиранозид при взаимодействии с
исходя из структур комплексов, можно сделать
CRD4 CD206 (рис. S5 в Приложении). При этом
вывод, что в случае α1→2 и α1→6 гликозидной
рецептор связывает второй Man остаток за счёт
связи в α D маннопиранозил α D манно
дополнительных взаимодействий Lys113
зе (diMan) достигается эффект вилки или биан
(3,3
ккал/моль; vdW, E, NP) и Glu111
тенны, связывание бóльшим количеством ато
(4,5 ккал/моль; E). По данным НС (таблица),
мов (рис. 4) и задействованы 2 Man остатка. Об
аффинность МР к диманнозе 3 с α1→2 глико
щие мотивы в комплексах diMan-ConA: остат
зидной связью практически на 2 порядка выше,
ки 0MA (терминальные маннопиранозиды) об
чем к диманнозам 6-7 с α1→3 или α1→6.
разуют водородные связи с Leu99, Tyr100, Asn14,
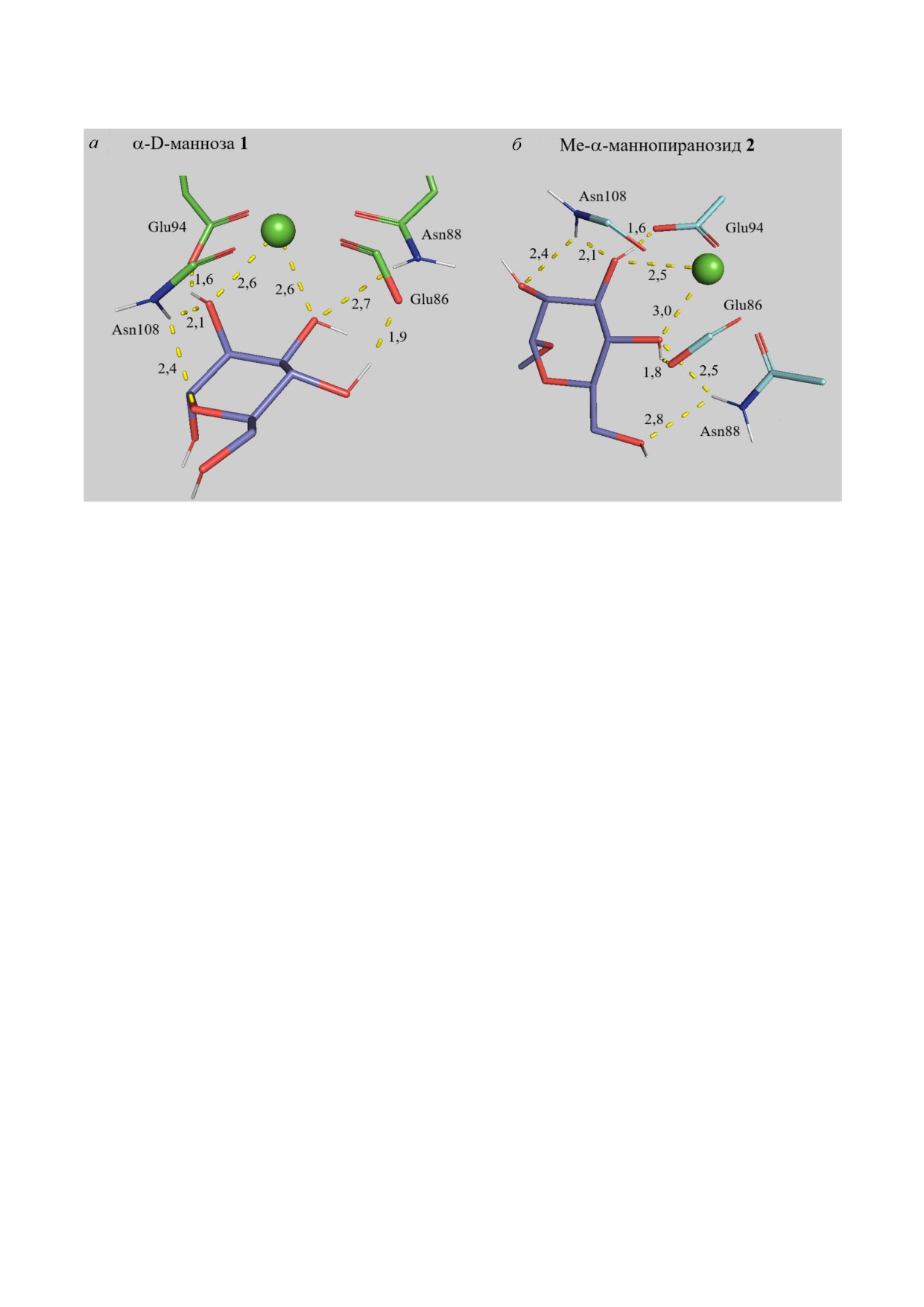
Роль CH3 группы в O1 положении. Метилиро
Asp208, Arg228 - консервативные для большин
вание OH групп у O1 атомов углеводов, по дан
ства комплексов. Вклад в свободную энергию
ным МД, увеличивает сродство к лекти
данных взаимодействий таков: Leu и Tyr - 3,95 и
ну ConA (рис. S6 в Приложении): в случае ман
2,45 ккал/моль (vdW, E, NP); Asn и Asp - 3,0 и
нозы 1-2 - в 2,4 раза; диманнозы 3-4 - в 1,4 ра
4,62 (E); Arg - 3,79 (P); где vdW - ван дер вааль
за; диманнозы 5-6 - в 1,14 раза; диманно
совы взаимодействия, E - электростатические,
зы 7-8 - в 1,3 раза; триманнозидов 9-10 - в 2,7
P - полярная сольватация и NP - неполярная
раза соответственно. Для домена CD206 наблю
сольватация. Однако для остатка 2MA (Man с
дается похожая картина (рис. 5) для маннозы и
позицией для связи α2) наблюдаются различия в
α1→2 и α1→6 изомеров диманнозы, однако ме
комплексах изомеров диманнозы с ConA. Так,
тилирование α1→3 изомера и триманнозы при
в α1→3 изомере (рис. 4) 2MA практически не за
водит к снижению аффинности. Таким образом,
действован в связывании. В других изомерах ре
метильная группа в олигосахаридах мешает свя
ализуются водородные связи с Tyr12 (3,5 ккал/моль;
зыванию с CD206, что хорошо согласуется с от
E, NP), Asn14 и Thr15 (5,6 и 3,8 ккал/моль;
сутствием таковой в гликанах микроорганиз
vdW, E, NP), Asp16 (7,0 ккал/моль; E). Наиболее
мов, которые распознаёт МР.
энергетически выгодные взаимодействия обус
Интересен механизм увеличения аффиннос
ловлены электростатическим притяжением к
ти моносахаридов к МР за счёт введения гидро
остаткам Asp и Asn. Таким образом, α1→6 гли
фобной группы и блокировки O1 атома от по
козидная связь в diMan делает Man остатки дос
тенциального образования водородных связей.
Рис. 4. Влияние типа гликозидной связи на связывание диманнозы (белый) ConA (зелёный). а - Me α1→2 диманнопи
ранозид 3 - субъединица A. б - Me α1→3 диманнопиранозид 5 - субъединица B. в - Me α1→6 диманнопиранозид 8 -
субъединица A. Зелёный шар - Ca2+. Молекулярная динамика 10 нс
БИОХИМИЯ том 87 вып. 1 2022
МОДЕЛИРОВАНИЕ ConA, CD206 - МАННОЗНЫЕ ЛИГАНДЫ
77
Рис. 5. Влияние метилирования атома O1 на связывание маннозы четвёртым доменом маннозного рецептора CD206.
а - α D манноза 1. б - Me α маннопиранозид 2. Молекулярная динамика 200 нс
CH3 группа непосредственно не участвует во
Лиганд 11, в котором к консервативному ядру
взаимодействии с рецептором (рис. 5 и рис. S6
триманнозида прикреплены через гликозидную
в Приложении), однако уменьшает возмож
связь β(1→2) два терминальных остат
ность образования взаимодействий O1H груп
ка GlcNAc, оказался самым специфичным
пы с рецептором, что приводит к изменению
к ConA и CD206 среди представленных:
пространственной ориентации Man остатка
Kd (ConA - лиганд 11) = 2 мкМ; Kd (CRD4 - ли
примерно на 90° и, соответственно, к оптимиза
ганд 11) = 480 мкМ.
ции связывания маннозы аминокислотными
Термодинамические параметры комплексооб
остатками CD206, включая Asn108, Asn88 и
разования рецептор-лиганд. Декомпозиция энер*
Glu94, а также изменяется координация катио
гии. Как обсуждалось выше, с увеличением ко
ном кальция.
личества Man или GlcNAc остатков сродство
Мультивалентное связывание триманнози
лигандов к ConA и CD206 увеличивается. Рас
дов. Триманнозид является природным лиган
смотрены водородные связи, образующиеся в
дом к ConA, консервативное ядро triMan в оли
комплексе между аминокислотными остатками
госахаридах высокоаффинно к рецептору [20,
белка и лигандом. Их количество коррелирует с
21]. Рассмотрим связывание триманнози
аффинностью взаимодействия, однако данный
дов 9-11 (рис. S2 в Приложении) ConA: две
параметр не исчерпывающий. Необходимо до
плоскости маннопиранозильных остатков пер
полнительно учитывать ван дер ваальсо
пендикулярны, а третья наклонена примерно
вы (vdW) и электростатические (E) взаимодей
под углом 45° к центральной. В данном случае
ствия, полярную (P) и неполярную (NP) сольва
связывание происходит по «улучшенному меха
тацию. Большое количество ароматических ос
низму»: образуется в среднем 15 водородных
татков (особенно Tyr) в сайтах связывания угле
связей и 5-9 дополнительно опосредованы во
водов, по видимому, облегчает связывание за
дой. Сайт лектина расширяется: дополнительно
счёт CH π взаимодействий [59]: например, цел
к упомянутым выше задействованы следующие
люлозо синтаза, Escherichia coli β galactosi
аминокислотные остатки: Asp16, Thr15, Pro13,
dase (PDB 1JZ8) [60].
Tyr12, Asp208. Это обусловливает прочность
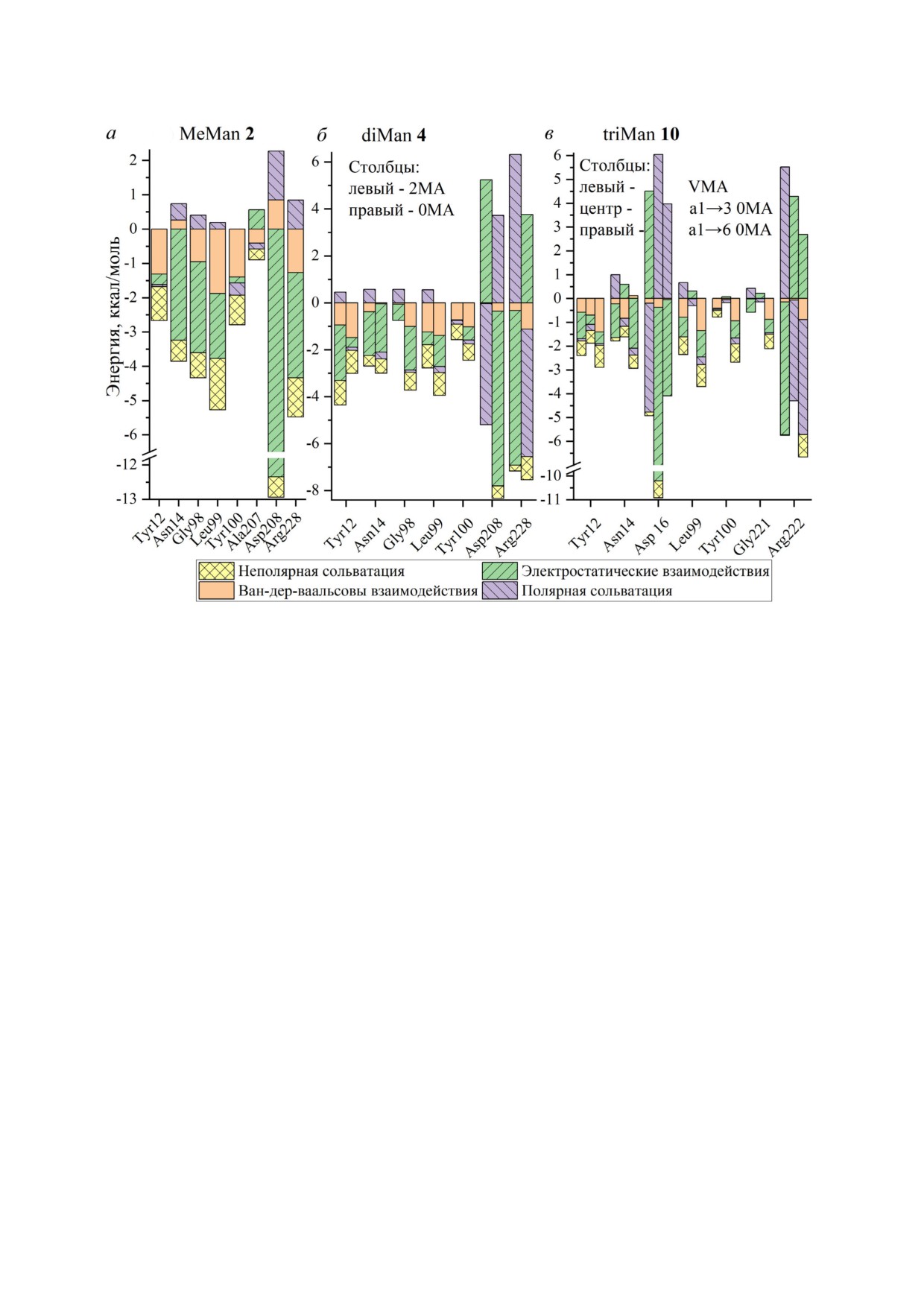
Нами проведено разложение свободной
комплексов с ConA.
энергии комплексообразования на 4 составляю
Введение терминальных GlcNAc остатков
щие, указанные выше, для аминокислотных ос
к triMan приводит к взаимодействию с Thr226,
татков, которые задействованы в связыва
Glu98, Arg228 и Gly224 (рис. S7 в Приложении).
нии (рис. 6).
БИОХИМИЯ том 87 вып. 1 2022
78
ЗЛОТНИКОВ, КУДРЯШОВА
Рис. 6. Компоненты свободной энергии взаимодействия аминокислотных остатков ConA с лигандами MeMan 2 (а),
diMan 4 (б) и MetriMan 10 (в). Значения приведены в ккал/моль. Для лигандов 4 и 10 показано разложение по составляю
щим Man остаткам: 0MA - концевой остаток, 2MA - Man с α2 позицией для связи, VMA - центральный Man остаток в
триманнозиде
При связывании моносахаридов (рис. 6, а)
отталкивания второго остатка (2MA или 0MA).
с ConA наибольший вклад в свободную энергию
Большее значение приобретают гидрофобные
вносят электростатические взаимодействия
взаимодействия с Tyr12, Tyr100 (включая CH π
(70%) атомов H, O пептидных связей и заряжён
стекинг с плоскостями Man остатков), Leu99.
ных групп Asp, Asn, Arg с атомами лиганда. До
Комплексообразование биантенного три
полнительная стабилизация связывания моно
маннозида (рис. 6, в) по компонентам энергии
сахарида достигается за счёт vdW взаимодей
аналогично диманнозиду. Таким образом, свя
ствий (18%) и неполярной сольватации (20%).
зывание углеводных лигандов в значительной
Мешающим фактором является полярная соль
мере обусловлено электростатическими взаимо
ватация - 8% затраты энергии.
действиями [35], что согласуется с фактом поте
При комплексообразовании действительно
ри углеводсвязывающей способности ConA при
могут осуществляться стекинг взаимодей
pH < 4. Однако велика роль гидрофобных остат
ствия (CH π) между Tyr и Man остатком с вкла
ков, в том числе ароматических, для увеличения
дом в свободную энергию 1-2 ккал/моль. Так,
аффинности за счёт CH π взаимодействий.
при переходе от Man к diMan и triMan каждый
Вытеснение молекул воды. Кроме того, с по
новый углеводный остаток взаимодействует
мощью GIST [61-62] показано, что вблизи сай
с Tyr, хотя и с меньшей термодинамической вы
та связывания ConA и CRD4 энергии взаимо
годой, однако общее значение энергетического
действия молекул воды с аминокислотными ос
вклада увеличивается от 2,3 до 4,4 ккал/моль.
татками рецептора неблагоприятны, Esw
=
Связывание диманнозы (рис. 6, б), как и в
= -2 ккал/(моль Å3), в то время как энергии
случае MeMan, обеспечивается электростати
растворитель-растворитель энергетически вы
ческими взаимодействиями заряженных групп
годнее, Eww = -18 ккал/(моль Å3). При комплек
Asp, Asn, Arg, но с меньшей долей (47%), так как
сообразовании триманнозида
9
с доменом
проявляется сглаживание общей энергии за счёт
CD206 происходит снижение ориентационной
БИОХИМИЯ том 87 вып. 1 2022
МОДЕЛИРОВАНИЕ ConA, CD206 - МАННОЗНЫЕ ЛИГАНДЫ
79
энтропии воды с 4,4 до 0,7 ккал/(моль Å3) и уве
вижность лигандов (особенно моносахаридов)
личение позиционной компоненты с
-139
заметна при моделировании в пс-нс временных
до -102 ккал/(моль Å3) за счёт перераспределе
интервалах, однако в экспериментах in vitro наб
ния молекул растворителя. Значит, вытеснение
людается усреднённая картина.
молекул воды лигандом при комплексообразо
Для объяснения различий в субъединицах
вании обеспечивает дополнительную стабили
следует рассмотреть детали механизма комплек
зирующую термодинамическую составляю
сообразования. Заметно общее сходство в свя
щую [63-64].
зывании Me Man (рис. S4 в Приложении) в че
Сравнение домена CD206 и ConA. Одна из за
тырёх субъединицах, однако в A и B только по
дач работы состоит в определении границ при
одной водородной связи, опосредованной во
менимости ConA в качестве модельного ман
дой, в субъединице C нет связи O3 c Arg228, но
нозного рецептора. Знание аспектов сходства и
есть O2-Leu99. Кроме того, расстояния в субъ
отличия рецепторов позволит апробировать
единице D в основном больше, чем в других -
системы доставки лекарств на доступном лекти
сродство ниже.
не и лишь после этого этапа на самих МР, что
Отличающиеся механизмы комплексообра
позволит значительно оптимизировать экспери
зования свидетельствуют о нескольких режимах
менты in vitro и in vivo [3, 20]. Нами рассмотрены
связывания, реализующихся в субъедини
режимы комплексообразования, структуры сай
цах A-D ConA. Возможность вариативного свя
тов связывания, механизмы задействования ка
зывания лигандов CRD4 CD206 не описана в ли
тионов металлов в связывании углеводов, значе
тературе, но, вероятно, проявляется в аспектах
ния энергий комплексообразования и констан
задействования катиона кальция и конкретных
ты диссоциации комплексов лиганд-рецептор,
а.о. с лигандом. Предполагается, что такая вари
а также литературные примеры.
ативность особенно заметна при участии как ми
Наиболее веским подтверждением сходства
нимум нескольких углеводсвязывающих доме
закономерностей связывания углеводов ConA
нов (CRD4-8). В литературе описано подобное
и CD206 является близость относительных изме*
явление на примере маннозо специфичного лек
нений констант диссоциаций комплексов (табли
тина ERGIC 53 [66], а также ConA [67]. Хотя все
ца), что обусловливает обоснованность исполь
4 субъединицы в ConA идентичны по составляю
зования модельного лектина для оптимизации
щим аминокислотам, существуют небольшие от
экспериментов in vitro и in vivo в пользу изучения
личия в их структурной организации (относи
большего количества лигандов и апробации по
тельное расположение цепей) в результате фор
тенциальных носителей лекарств. Так, коэффи
мирования четвертичной структуры белка, что
циент корреляции между свободными энергия
обусловливает обсуждаемые выше явления. По
ми комплексообразования для шести описан
этому наиболее корректные параметры можно
ных в литературе комплексов лигандов (№ 2, 4,
получить, усредняя значения энергий связыва
6-7, 10, 14) с CD206 и ConA составляет 0,90.
ния для всех четырёх субъединиц, поскольку на
В ходе моделирования получены достаточно ре
практике реализуются все варианты связывания.
левантные значения энергий с высокой взаи
Сравнение структур домена CD206 и ConA
мосвязью между комплексами обоих рецепто
(рис. S8 в Приложении). Ключевые сходства
ров: данные МД по комплексообразова
структурной организации ConA и CRD4 CD206
нию CRD4 коррелируют с таковыми для нейро
следующие: примерно одинаковое простран
сетевого анализа ConA с коэффициентом
ственное расположение аминокислот, близкий
r = 0,91. Таким образом, ConA в закономернос
аминокислотный состав сайтов связывания уг
тях связывания моно и олигосахаридов являет
леводов (2 остатка Tyr, Asn, Asp, Ala, Leu, Pro),
ся хорошей моделью маннозного рецептора.
что обеспечивает реализацию связывания за
Режимы комплексообразования маннозосодер
счёт высокоэнергетических электростатических
жащих лигандов с рецептором. Главное отличие
взаимодействий заряженных остатков амино
ConA от домена CD206 - тетрамерная структу
кислот и дополнительной стабилизации гидро
ра. Считается, что все субъединицы ConA взаи
фобными и CH π стекинг взаимодействиями,
модействуют с лигандами одинаково [65], что
обязательно присутствие Ca2+ (зелёный шар)
было поставлено под сомнение в ходе модели
для формирования сайта связывания.
рования (табл. S2 в Приложении). Можно пред
Роль катионов Ca2+ и Mn2+ в связывающем
положить, что это связано с возможностью взаи
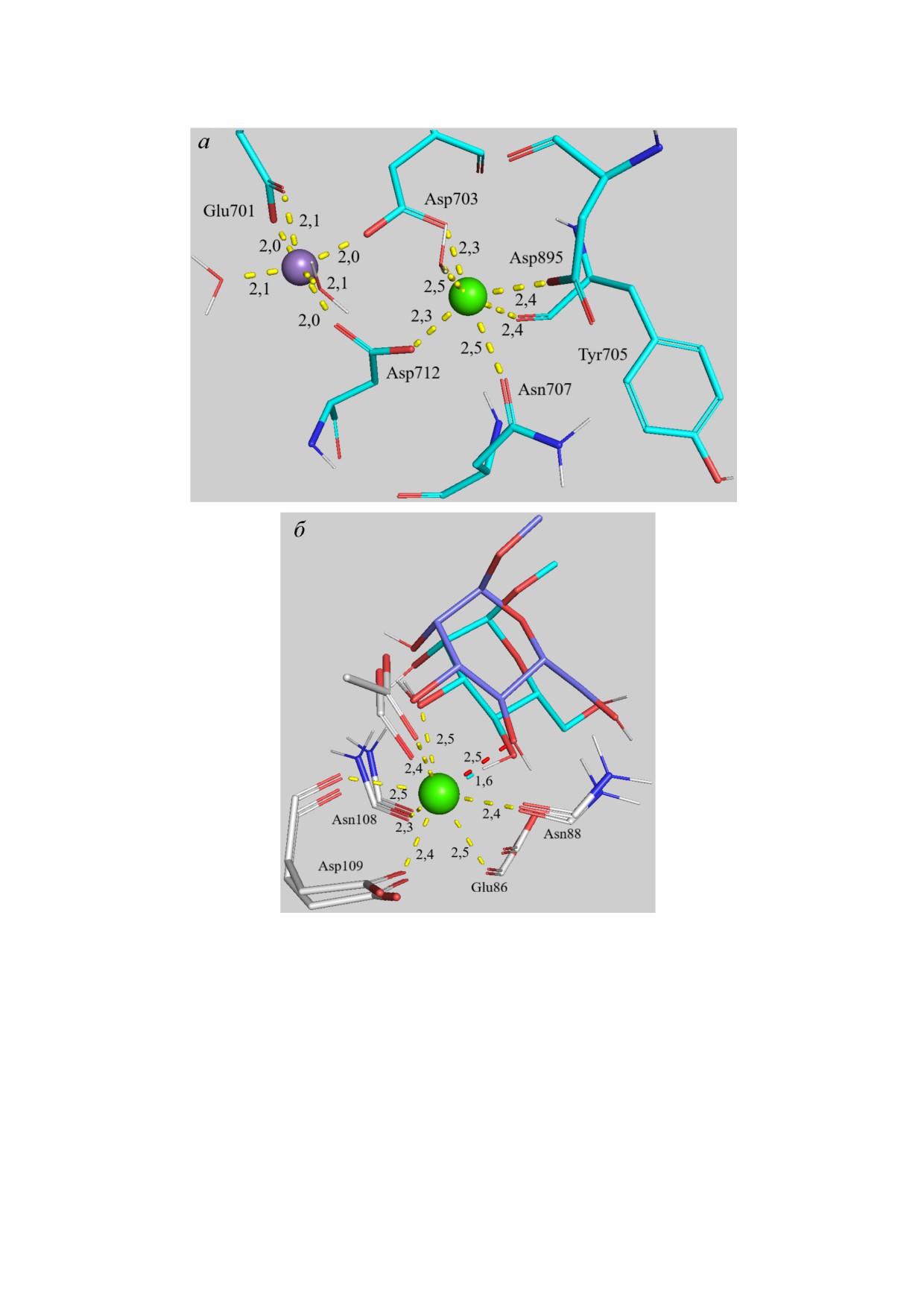
центре рецепторов ConA и CD206 (рис. 7 в При
модействия лигандов с различными аминокис
ложении). Как обсуждалось выше, наличие ка
лотными остатками, а не только определённы
тиона металла в сайте связывания необходимо
ми (например, Gly98 и Leu99, расположенными
для поддержания пространственной ориента
в сайте связывания, вместо Arg228). Такая под
ции аминокислотных остатков рецептора.
БИОХИМИЯ том 87 вып. 1 2022
80
ЗЛОТНИКОВ, КУДРЯШОВА
Рис. 7. а - Координационное окружение Mn2+, Ca2+ в сайте ConA (субъединица C) при связывании триманнози
да 9 (PDB Acc. No. 1ONA) по данным молекулярной динамики 10 нс. б - Координационное окружение Ca2+ в сайте до
мена CD206 при связывании метилманнозида 2 (PDB Acc. No. 7JUB). Сравнение исходных координат PDB (голубой) и
структуры после молекулярной динамики 100 нс (фиолетовый)
В ConA роль опоры играет Ca2+, который фик
ных остатков и кальция достигается за счёт рас
сирует остатки Asp703, Asp712 и Asp895, Asn707,
положенного на расстоянии 4,2-4,4 Å от Ca2+
Tyr705 и водой (рис. 7, а). Координационный
катиона Mn2+, координирующего остатки
полиэдр для катиона кальция - искажённый ок
Glu701, Asp703 и Asp712, а также две молекулы
таэдр (координационное число (КЧ) равно 6,
воды (рис. 7, а). Благодаря этому формируется
ионный радиус 100 пм). Более того, дополни
сайт связывания высокоаффинный к консерва
тельная стабилизация положений аминокислот
тивному ядру триманнозида [68-70].
БИОХИМИЯ том 87 вып. 1 2022
МОДЕЛИРОВАНИЕ ConA, CD206 - МАННОЗНЫЕ ЛИГАНДЫ
81
Специфичность маннозного рецептора
тальными методами. Однако с развитием техно
CD206 к Man , Fuc и GlcNAc остаткам в гли
логий эта проблема постепенно решается. Ней
канах обеспечивается механизмом, аналогич
росети активно совершенствуются, что следует
ным рассмотренному выше для ConA. Ca2+ ко
из результатов
«соревнований CASF 2013»
ординирует Asn88, Asn108, Asp109 и Glu86, но в
[75, 76]. Нами выбрана НС Pafnucy вследствие её
отличие от сайта в ConA с большими координа
лидирующей позиции среди алгоритмов по
ционными числами, равным 8, и ионным ради
предсказанию аффинность лиганд-рецептор.
усом 112 пм. Кроме того, катион Ca2+ в CRD4
Однако исходный алгоритм, обученный разра
играет важную роль в непосредственном связы
ботчиками на основе PDBbind2016, показал
вании лиганда. На рис. 7, б показано взаимодей
низкую точность применительно к рассматри
ствие метилманнозида 2 с Ca2+. При моделиро
ваемым системам. Коэффициенты корреляции
вании связывания на основе исходных коорди
свободных энергий для ConA и CRD4 CD206
нат из PDB продемонстрировано изменение ко
(нейросетевой анализ № 1) оказались не превос
ординационного окружения кальция атомами
ходящими 0,50, что, вероятно, связано с непод
O2,3 MeMan до среднего расстояния 2,5 Å с об
ходящими весами в слоях НС для рассматривае
разованием устойчивой квадратной антиприз
мых систем. Поэтому авторы настоящей работы
мы (рис. 7, б).
подготовили обучающий набор на основе
Литературные данные о сходстве CRD4 с
PDBbind2020, увеличив количество циклов обу
ConA. Согласно литературным данным,
чения до 25 и выбрав подходящие коэффициен
CRD4 демонстрирует высокое сходство с ман
ты (раздел Методы, рис. S1 и табл. S1 в Прило
нан связывающим лектином А, который, в
жении). Нейросеть с новыми параметрами по
свою очередь, похож на ConA в закономернос
казала более высокую точность предсказаний,
тях связывания углеводов [71]. В экспериментах
превосходящую метод МД: коэффициенты кор
in vivo и in vitro при использовании маннозосо
реляции энергий для ConA и CRD4 CD206 (ней
держащих лигандов, высокоспецифичных
росетевой анализ № 2) соответственно рав
к ConA, для модификации средств доставки ле
ны 0,82 и 0,8. Достигнутая точность в среднем
карственных препаратов к макрофагам путём
совпадает с таковой для алгоритмов, описанных
нацеливания на МР наблюдаются положитель
в литературе [77], хотя может быть увеличена.
ные эффекты [20]: поглощение маннозилиро
Перспективы методов in silico широки. Усо
ванных частиц клетками [72, 73], увеличение
вершенствование алгоритмов НС и подготовка
времени циркуляции лекарства в кровотоке и
обучающих наборов с бóльшим количеством ре
повышение концентрации в макрофагах [3], био
левантных данных о комплексах позволят уве
распределение маннозилированных липосом,
личить точность предсказаний, что было пока
преимущественно в лёгкие [74]. Все эти данные
зано выше. Применение молекулярной динами
обусловливают релевантность применения
ки с уточнёнными силовыми полями (amber) и
ConA в качестве модели МР.
увеличенным временем моделирования позво
Настоящая ситуация и перспективы методов
ляет точнее определять термодинамические ве
молекулярной динамики и нейросетевого анализа.
личины и структуры молекул, в том числе комп
Значения энергий комплексообразования и со
лексов белок-лиганд, в условиях, приближен
ответственно констант диссоциации комплек
ным к реальным (раствор, концентрация соли,
сов рецептор-лиганд были рассчитаны для
температура).
15
лигандов с двумя рецепторами ConA и
Таким образом, с помощью молекулярной
CRD4 CD206. Из них ранее в литературе не
динамики и искусственных нейросетей опреде
представлены 1 для конканавалина А и 9 - для
лены параметры лектин лигандных взаимодей
маннозного рецептора. Достоверность получен
ствий с учётом модели оптимизации. Компью
ных данных показана относительно экспери
терные методы позволили получить простран
ментальных данных из литературы. Коэффици
ственные структуры комплексов ConA и
енты корреляции свободных энергий для ConA
CRD4 CD206 с углеводными лигандами, боль
и CRD4 CD206 по методу МД (анализ
шая часть которых ранее не описана. Кроме то
mmpbsa.py) соответственно равны 0,68 и 0,52,
го, изучено комплексообразование маннозного
что можно считать хорошим результатом, учи
рецептора, что крайне затруднительно in vitro.
тывая количество изученных лигандов и новиз
ну исследований МР.
Нужно отметить, что существующие алго
ЗАКЛЮЧЕНИЕ
ритмы далеко не совершенны, и компьютерное
моделирование редко коррелирует с абсолют
Нацеливание средств доставки терапевти
ными значениями, полученными эксперимен
ческих агентов на маннозные рецепторы макро
6 БИОХИМИЯ том 87 вып. 1 2022
82
ЗЛОТНИКОВ, КУДРЯШОВА
фагов представляется перспективным методом
и его производным, а МР наиболее прочно свя
повышения эффективности лечения целого ря
зывает разветвлённые олигосахариды, состоя
да опасных заболеваний, включая аутоиммун
щие из Man остатков, подобные поверхност
ные, онкологические и инфекционные. Поэто
ным углеводам клеточной стенки микроорга
му требуется создание высокоселективных ман
низмов. Открывается возможность моделиро
нозилированных носителей лекарственных
вать комплексообразование маннозного рецеп
средств, оптимальная конфигурация которых
тора CD206 с олиго и полимерными молекула
может быть найдена с помощью компьютерного
ми, модифицированными наиболее аффинны
моделирования. В данной работе с помощью
ми лигандами среди рассмотренных в данной
молекулярной динамики и искусственной ней
работе, например, высокоманнозилированны
росети Pafnucy проведено моделирование комп
ми циклодекстринами и хитозанами - перспек
лексообразования 15 углеводных лигандов, ко
тивными носителями лекарств с функцией ад
торые могут выступать в качестве адресной мет
ресного нацеливания на маннозные рецепторы
ки на молекуле для таргетирования макрофагов,
макрофагов.
с модельным тетрамером ConA и четвёртым до
меном (вносящим наибольший вклад в углевод
Благодарности. Авторы выражают искренние
связывающую способность) маннозного ре
благодарности Дроботу Виктору Валерьевичу
цептора CD206. Искусственные нейросети ак
(МГУ имени М.В. Ломоносова, НИИ им. Бело
тивно разрабатываются и развиваются, однако
зерского, химический факультет, НИВЦ) за
их применение в биологических системах пока
консультации и советы по проведению молеку
не так широко. В данной работе продемонстри
лярной динамики; к.х.н. Виноградову Алексею
ровано, что обученная на основе PDBbind2020
Александровичу за концептуальные, методичес
нейросеть, наряду с молекулярной динамикой,
кие и стилистические предложения по улучше
является эффективным инструментом для изу
нию представленного материала; Злотникову
чения механизмов взаимодействия лиганд-ре
Дмитрию Николаевичу за реализацию техни
цептор. В ходе компьютерного моделирования
ческой платформы для проведения молекуляр
нами рассчитаны константы диссоциации
ных симуляций и нейросетевого анализа с ис
комплексов лиганд-ConA, коррелирующие с
пользованием технологии докеризации.
литературными данными. С использованием от
Конфликт интересов. Авторы заявляют об от
работанной модели впервые изучалось комп
сутствии конфликта интересов.
лексообразование домена маннозного рецепто
Соблюдение этических норм. Настоящая
ра CD206 с лигандами, более того, нами рассчи
статья не содержит описания каких либо иссле
таны 9 констант диссоциации комплексов, ра
дований с участием людей или животных в каче
нее не представленных в статьях. Показано, что
стве объектов.
ConA демонстрирует сходство с четвёртым до
Дополнительные материалы. Приложение к
меном CD206 в закономерностях связывания
статье опубликовано на сайте журнала «Биохи
углеводов, однако выявлены и отличия. Лектин
ConA высокоспецифичен к ядру триманнозида
вып. 1, 2022.
СПИСОК ЛИТЕРАТУРЫ
1.
Stepniewska Dziubinska, M. M., Zielenkiewicz, P., and
5.
Nikitina, E., Larionova, I., Choinzonov, E., and
Siedlecki, P. (2018) Development and evaluation of a
Kzhyshkowska, J. (2018) Monocytes and macrophages as
deep learning model for protein-ligand binding affinity
viral targets and reservoirs, Int. J. Mol. Sci., 19, 2821 2845,
prediction, Bioinformatics, 34, 3666 3674, doi: 10.1093/
doi: 10.3390/ijms19092821.
bioinformatics/bty374.
6.
Tormoen, G. W., Crittenden, M. R., and Gough, M. J.
2.
Banerjee, S., and Roy, S. (2019) Polysaccharide installed
(2018) Role of the immunosuppressive microenvironment
lipid nanoparticles in targeted antituberculosis drug deliv
in immunotherapy, Adv. Rad. Oncol.,
3,
520526,
ery applications, in Polysaccharide Carriers for Drug
doi: 10.1016/j.adro.2018.08.018.
Delivery, pp. 397 411, doi: 10.1016/b978 0 08 102553 6.
7.
Shan, H., Dou, W., Zhang, Y., and Qi, M. (2020) Targeted
00014 3.
ferritin nanoparticle encapsulating CpG oligodeoxynu
3.
Filatova, L. Y., Klyachko, N. L., and Kudryashova, E. V.
cleotides induces tumor associated macrophage M2 phe
(2018) Targeted delivery of anti tuberculosis drugs to
notype polarization into M1 phenotype and inhibits tumor
macrophages: targeting mannose receptors, Russ. Chem.
growth, Nanoscale, 12, 2226822280, doi: 10.1039/
Rev., 87, 374 391, doi: 10.1070/rcr4740.
d0nr04520a.
4.
Wong, M. E., Jaworowski, A., and Hearps, A. C. (2019)
8.
Le Menn, G., and Neels, J. (2018) Regulation of immune
The HIV reservoir in monocytes and macrophages, Front.
cell function by PPARs and the connection with metabolic
Immunol., 10, 14351450, doi: 10.3389/fimmu.2019.
and neurodegenerative diseases, Int. J. Mol. Sci., 19, 1575,
01435.
doi: 10.3390/ijms19061575.
БИОХИМИЯ том 87 вып. 1 2022
МОДЕЛИРОВАНИЕ ConA, CD206 - МАННОЗНЫЕ ЛИГАНДЫ
83
9.
Arora, S., Dev, K., Agarwal, B., Das, P., and Syed, M. A.
носителей для доставки лекарственных средств к аль
(2018) Macrophages: Their role, activation and polariza
веолярным макрофагам, Биотехнология, 37, 28 40,
tion in pulmonary diseases, Immunobiology, 223, 383 396,
doi: 10.21519/0234 2758 2021 37 5 28 40.
doi: 10.1016/j.imbio.2017.11.001.
25.
Le Deygen, I. M., Mamaeva, P. V., Skuredina, A. A., and
10.
Lyadova, I. V., and Panteleev, A. V. (2015) Th1 and Th17
Kudryashova, E. V. (2020) A spectral approach to study
cells in tuberculosis: Protection, pathology, and biomark
interaction between chitosan modified with mannose and
ers, Med. Inflamm., 2015, 854507, doi: 10.1155/2015/
concanavalin A for the creation of address delivery systems
854507.
of antituberculosis drugs, Moscow Univ. Chem. Bull., 75,
11.
Zhang, H. L., Zheng, X. Y., and Zhu, J. (2013) Th1/
213 217, doi: 10.3103/S0027131420040045.
Th2/Th17/Treg cytokines in Guillain-Barré syndrome
26.
Goldstein, I. J., and Poretz, R. D., (1986) The Lectins
and experimental autoimmune neuritis, Cyt. Grow. Fact.
Properties, Functions and Applications in Biology and
Rev., 24, 443 453, doi: 10.1016/j.cytogfr.2013.05.005.
Medicine, Academic Press.
12.
Stahl, P. D. (1990) The macrophage mannose receptor:
27.
Gordon, S. (2003) Alternative activation of macrophages,
Current status, Am. J. Resp. Cell Mol. Biol., 2, 317 318,
Nat. Rev. Immunol., 3, 23 35, doi: 10.1038/nri978.
doi: 10.1165/ajrcmb/2.4.317.
28.
Sharma, A., Porterfield, J. E., Smith, E., Sharma, R.,
13.
Feinberg, H., Jegouzo, S. A. F., Lasanajak, Y., Smith,
Kannan, S., et al. (2018) Effect of mannose targeting of
D. F., Drickamer, K., et al. (2021) Structural analysis of
hydroxyl PAMAM dendrimers on cellular and organ
carbohydrate binding by the macrophage mannose recep
biodistribution in a neonatal brain injury model, J. Control.
tor CD206, J. Biol. Chem.,
296,
100368100385,
Rel., 283, 175 189, doi: 10.1016/j.jconrel.2018.06.003.
doi: 10.1016/j.jbc.2021.100368.
29.
Tran, D. P., and Kitao, A. (2019) Dissociation process of
14.
Feinberg, H., Park Snyder, S., Kolatkar, A. R., Heise,
MDM2/p53 complex investigated by parallel cascade
C. T., Taylor, M. E., et al. (2000) Structure of a C type car
selection molecular dynamics and Markov state model,
bohydrate recognition domain from the macrophage man
J. Phys. Chem. B., 123, 2469 2478, doi: 10.1021/acs.jpcb.
nose receptor, J. Biol. Chem.,
275,
2153921548,
8b10309.
doi: 10.1074/jbc.m002366200.
30.
Hollingsworth, S. A., Nguyen, B. D., Chreifi, G., Arce,
15.
Taylor, M. E., and Drickamer, K. (1993) Structural
A. P., and Poulos, T. L. (2017) Insights into the dynamics
requirements for high affinity binding of complex ligands
and dissociation mechanism of a protein redox complex
by the macrophage mannose receptor, J. Biol. Chem., 268,
using molecular dynamics, J. Chem. Inf. Model., 57, 2344
399 404, doi: 10.1016/S0021 9258(18)54164 8.
2350, doi: 10.1021/acs.jcim.7b00421.
16.
Naismith, J. H., Emmerich, C., Habash, J., Harrop, S. J.,
31.
Tran, D. P., Takemura, K., Kuwata, K., and Kitao, A.
Helliwell, J. R., et al. (1994) Refined structure of con
(2017) Protein-Ligand dissociation simulated by parallel
canavalin A complexed with methyl α D mannopyra
cascade selection molecular dynamics, J. Chem. Theory
noside at 2.0 Å resolution and comparison with the saccha
Comput., 14, 404 417, doi: 10.1021/acs.jctc.7b00504.
ride free structure, Acta Crystallogr. Sec. D Biol. Crystallogr.,
32.
Mollica, L., Decherchi, S., Zia, S. R., Gaspari, R.,
50, 847 858, doi: 10.1107/s0907444994005287.
Cavalli, A., et al. (2015) Kinetics of protein ligand unbind
17.
Derewenda, Z., Yariv, J., Helliwell, J. R., Kalb, A. J.,
ing via smoothed potential molecular dynamics simula
Dodson, E. J., et al. (1989) The structure of the saccha
tions, Sci. Rep., 5, doi: 10.1038/srep11539.
ride binding site of concanavalin A, EMBO J., 8, 2189
33.
Fu, Y., Zhao, J., and Chen, Z. (2018) Insights into the
2193, doi: 10.1002/j.1460 2075.1989.tb08341.x.
molecular mechanisms of protein-ligand interactions by
18.
Brewer, C. F., Brown, R. D., and Koenig, S. H. (1983)
molecular docking and molecular dynamics simulation: a
Metal ion binding and conformational transitions in con
case of oligopeptide binding protein, Comput. Math. Meth.
canavalin-A: A structure-function study, J. Biomol. Struct.
Med., 2018, 1 12, doi: 10.1155/2018/3502514.
Dyn., 1, 961 997, doi: 10.1080/07391102.1983.10507497.
34.
Bradbrook, G. M., Gleichmann, T., Harrop, S. J.,
19.
Kaushik, S., Mohanty, D., and Surolia, A. (2009) The role
Habash, J., Raftery, J., et al. (1998) X Ray and molecular
of metal ions in substrate recognition and stability of con
dynamics studies of concanavalin A glucoside and manno
canavalin A: A molecular dynamics study, Biophys. J., 96,
side complexes relating structure to thermodynamics of
21 34, doi: 10.1529/biophysj.108.134601.
binding, J. Chem. Soc. Faraday Trans., 94, 1603 1611,
20.
Zlotnikov, I. D., and Kudryashova, E. V. (2022) Mannose
doi: 10.1039/a800429c.
receptors of alveolar macrophages as a target for delivery of
35.
Bryce, R. A., Hillier, I. H., and Naismith, J. H. (2001)
medicines to the lungs, Russ. J. Bioorg. Chem., 48,
Carbohydrate-protein recognition: Molecular dynamics
doi: 10.1134/S1068162022010150.
simulations and free energy analysis of oligosaccharide
21.
Mandal, D. K., Kishore, N., and Brewer, C. F. (1994)
binding to concanavalin A, Biophys. J., 81, 1373 1388,
Thermodynamics of lectin-carbohydrate interactions.
doi: 10.1016/s0006 3495(01)75793 1.
Titration microcalorimetry measurements of the binding of
36.
Arcon, J. P., Defelipe, L. A., Modenutti, C. P., Lopez, E. D.,
N linked carbohydrates and ovalbumin to concanavalin A,
Alvarez Garcia, D., et al. (2017) Molecular dynamics in
Biochemistry, 33, 1149 1156, doi: 10.1021/bi00171a014.
mixed solvents reveals protein-ligand interactions, improves
22.
Dam, T. K., Roy, R., Das, S. K., Oscarson, S., and Brewer,
docking, and allows accurate binding free energy predic
C. F. (2000) Binding of multivalent carbohydrates to con
tions, J. Chem. Inf. Model., 57, 846 863, doi: 10.1021/
canavalin A and Dioclea grandiflora lectin, J. Biol. Chem.,
acs.jcim.6b00678.
275, 14223 14230, doi: 10.1074/jbc.275.19.14223.
37.
Zacharias, M. (2004) Rapid protein-ligand docking using
23.
Landschoot, A., Loontiens, F. G., and Bruyne, C. K.
soft modes from molecular dynamics simulations to
(1980) Binding of manno oligosaccharides to con
account for protein deformability: binding of FK506 to
canavalin A. Substitution titration with a fluorescent indi
FKBP, Prot. Str. Fun. Bioinf., 54, 759 767, doi: 10.1002/
cator ligand, Eur. J. Biochem., 103, 307 312, doi: 10.1111/
prot.10637.
j.1432 1033.1980.tb04316.x.
38.
Mollica, L., Theret, I., Antoine, M., Perron Sierra, F.,
24.
Злотников И. Д., Ваничкин Д. А., Кудряшова Е. В.
Charton, Y., et al. (2016) Molecular dynamics simulations
(2021) Методы определения параметров рецептор ли
and kinetic measurements to estimate and predict pro
гандных взаимодействий на модели конканавалина А
tein-ligand residence times, J. Med. Chem., 59, 7167
и маннозилированных хитозанов - перспективных
7176, doi: 10.1021/acs.jmedchem.6b00632.
БИОХИМИЯ том 87 вып. 1 2022
6*
84
ЗЛОТНИКОВ, КУДРЯШОВА
39.
Wang, S., Liu, D., Ding, M., Du, Z., Zhong, Y., et al.
55.
Taylor, M. E., Bezouska, K., and Drickamer, K. (1992)
(2021) SE OnionNet: a convolution neural network for
Contribution to ligand binding by multiple carbohydrate
protein-ligand binding affinity prediction, Front. Gen., 11,
recognition domains in the macrophage mannose receptor,
607824, doi: 10.3389/fgene.2020.607824.
J. Biol. Chem., 267, 17191726, doi: 10.1016/S0021
40.
Durrant, J. D., and McCammon, J. A.
(2011)
9258(18)46005 X.
NNScore 2.0: A neural network receptor-ligand scoring
56.
Berman, H. M., Battistuz, T., Bhat, T. N., Bluhm, W. F.,
function, J. Chem. Inf. Model.,
51,
28972903,
Bourne, P. E., et al. (2002) The protein data bank, Acta
doi: 10.1021/ci2003889.
Cryst. Sect. D Biol. Cryst., 58, 899 907, doi: 10.1107/
41.
Lahey, S. L. J., and Rowley, C. N. (2020) Simulating pro
s0907444902003451.
tein-ligand binding with neural network potentials, Chem.
57.
Böhm, H. J., and Schneider, G. (2003) Protein-Ligand
Sci., 11, 2362 2368, doi: 10.1039/c9sc06017k.
Interactions, in Methods and Principles in Medicinal
42.
Zhou, Y., Wang, F., Tang, J., Nussinov, R., and Cheng, F.
Chemistry, pp. 3 20, doi: 10.1002/3527601813.
(2020) Artificial intelligence in COVID 19 drug repurpos
58.
Huang, K., Luo, S., Cong, Y., Zhong, S., Zhang, J. Z. H.,
ing, Lanc. Dig. Heal., 2, 667 676, doi: 10.1016/s2589
et al. (2020) Accurate free energy estimator: Based on
7500(20)30192 8.
MM/PBSA combined with interaction entropy for pro
43.
Pham, T. H., Qiu, Y., Zeng, J., Xie, L., and Zhang, P.
tein-ligand binding affinity, Nanoscale, 12, 10737 10750,
(2021) A deep learning framework for high throughput
doi: 10.1039/c9nr10638c.
mechanism driven phenotype compound screening and its
59.
Spiwok, V. (2017) CH/π interactions in carbohydrate recog
application to COVID 19 drug repurposing, Nat. Mach.
nition, Molecules, 22, 1038, doi: 10.3390/molecules22071038.
Intel., 3, 247 257, doi: 10.1038/s42256 020 00285 9.
60.
Spiwok, V., Lipovova, P., Skalova, T., Buchtelova, E.,
44.
Книга А. Е., Поляков И. В., Немухин А. В. (2021)
Hasek, J., et al. (2004) Role of CH/π interactions in sub
In silico определение специфичности неоантиген ре
strate binding by Escherichia coli β galactosidase, Carb.
активных T лимфоцитов, Биомедицинская химия, 67,
Res., 339, 2275 2280, doi: 10.1016/j.carres.2004.06.016.
251 258, doi: 10.18097/pbmc20216703251.
61.
Lazaridis, T. (1998) Inhomogeneous fluid approach to sol
45.
Manasaryan, G., Suplatov, D., Pushkarev, S., Drobot, V.,
vation thermodynamics. 1. Theory, J. Phys. Chem. B, 102,
Kuimov, A., et al. (2021) Bioinformatic analysis of the
3531 3541, doi: 10.1021/jp9723574.
nicotinamide binding site in Poly(ADP ribose) polymerase
62.
Nguyen, C. N., Young, T. K., and Gilson, M. K. (2012)
family proteins, Cancers,
13,
1201, doi:
10.3390/
Grid inhomogeneous solvation theory: hydration structure
cancers13061201.
and thermodynamics of the miniature receptor cucur
46.
Timonina, D., Sharapova, Y., Švedas, V., and Suplatov, D.
bit[7]uril, J. Chem. Phys., 137, 044101, doi: 10.1063/
(2021) Bioinformatic analysis of subfamily specific regions
1.4733951.
in 3D structures of homologs to study functional diversity
63.
Hüfner Wulsdorf, T., and Klebe, G. (2020) Protein-ligand
and conformational plasticity in protein superfamilies,
complex solvation thermodynamics: Development, para
Comput. Struct. Biotech. J., 19, 1302 1311, doi: 10.1016/
meterization, and testing of GIST based solvent function
j.csbj.2021.02.005.
als, J. Chem. Inf. Model., 60, 1409 1423, doi: 10.1021/
47.
Case, D. A., Aktulga, H. M., Belfon, K., Ben Shalom,
acs.jcim.9b01109.
I. Y., Brozell, S. R., et al. (2021) Amber 2021: Reference
64.
Ramsey, S., Nguyen, C., Salomon Ferrer, R., Walker, R. C.,
Manual, University of California, San Francisco.
Gilson, M. K., et al. (2016) Solvation thermodynamic map
48.
Jorgensen, W. L., Chandrasekhar, J., Madura, J. D.,
ping of molecular surfaces in AmberTools: GIST, J. Comp.
Impey, R. W., and Klein, M. L. (1983) Comparison of sim
Chem., 37, 2029 2037, doi: 10.1002/jcc.24417.
ple potential functions for simulating liquid water, J. Chem.
65.
Dimick, S. M., Powell, S. C., McMahon, S. A., Moothoo,
Phys., 79, 926 935, doi: 10.1063/1.445869.
D. N., Naismith, J. H., et al. (1999) On the meaning of
49.
Ryckaert, J. P., Ciccotti, G., and Berendsen, H. J. C.
affinity: Cluster glycoside effects and concanavalin A,
(1977) Numerical integration of the Cartesian equations of
J. Am. Chem. Soc., 121, 1028610296, doi: 10.1021/
motion of a system with constraints: molecular dynamics
ja991729e.
of n alkanes, J. Comput. Phys., 23, 327 334, doi: 10.1016/
66.
Satoh, T., Suzuki, K., Yamaguchi, T., and Kato, K. (2014)
0021 9991(77)90098 5.
Structural basis for disparate sugar binding specificities in
50.
Miller, B. R., McGee, T. D., Swails, J. M., Homeyer, N.,
the homologous cargo receptors ERGIC 53 and VIP36,
Gohlke, H., et al. (2012) MMPBSA.py: An efficient pro
PLoS One, 9, 87963, doi: 10.1371/journal.pone.0087963.
gram for end state free energy calculations, J. Chem.
67.
Moothoo, D. N., Canan, B., Field, R. A., and Naismith,
Theory Comput., 8, 3314 3321, doi: 10.1021/ct300418h.
J. H. (1999) Man α1 2 Man α OMe concanavalin A com
51.
Roe, D. R., and Cheatham, T. E. (2013) PTRAJ and CPP
plex reveals a balance of forces involved in carbohydrate
TRAJ: Software for processing and analysis of molecular
recognition, Glycobiology, 9, 539 545, doi: 10.1093/glycob/
dynamics trajectory data, J. Chem. Theory Comput., 9,
9.6.539.
3084 3095, doi: 10.1021/ct400341p.
68.
Naismith, J. H., and Field, R. A. (1996) Structural basis of
52.
Hitchen, P. G., Mullin, N. P., and Taylor, M. E. (1998) Ori
trimannoside recognition by concanavalin A, J. Biol.
entation of sugars bound to the principal C type carbohy
Chem., 271, 972 976, doi: 10.1074/jbc.271.2.972.
drate recognition domain of the macrophage mannose
69.
Gupta, D., Dam, T. K., Oscarson, S., and Brewer, C. F.
receptor, Biochem. J., 333, 601 608, doi: 10.1042/bj3330601.
(1997) Thermodynamics of lectin-carbohydrate interac
53.
Mullin, N. P., Hitchen, P. G., and Taylor, M. E. (1997)
tions, J. Biol. Chem., 272, 6388 6392, doi: 10.1074/jbc.272.
Mechanism of Ca2+ and monosaccharide binding to a C
10.6388.
type carbohydrate recognition domain of the macrophage
70.
Mandal, D. K., Bhattacharyya, L., Koenig, S. H., Brown,
mannose receptor, J. Biol. Chem., 272, 56685681,
R. D., Oscarson, S., et al. (1994) Studies of the binding
doi: 10.1074/jbc.272.9.5668.
specificity of concanavalin A. Nature of the extended bind
54.
Asciutto, E. K., Kopanchuk, S., Lepland, A., Simón
ing site for asparagine linked carbohydrates, Biochemistry,
Gracia, L., Aleman, C., et al. (2019) A phage display
33, 1157 1162, doi: 10.1021/bi00171a015.
derived peptide binds to human CD206 and modeling
71.
Weis, W. I., and Drickamer, K. (1996) Structural basis of
reveals a new binding site in the receptor, J. Phys. Chem. B.,
lectin carbohydrate recognition, Annu. Rev. Biochem., 65,
123, 1973 1982, doi: 10.1021/acs.jpcb.8b11876.
441 473, doi: 10.1146/annurev.bi.65.070196.002301.
БИОХИМИЯ том 87 вып. 1 2022
МОДЕЛИРОВАНИЕ ConA, CD206 - МАННОЗНЫЕ ЛИГАНДЫ
85
72.
Ghotbi, Z., Haddadi, A., Hamdy, S., Hung, R. W.,
75. Li, Y., Liu, Z., Li, J., and Wang, R. (2014) Comparative
Samuel, J., et al. (2010) Active targeting of dendritic cells
assessment of scoring functions on an updated bench
with mannan decorated PLGA nanoparticles, J. Drug
mark: 1. Compilation of the test set, J. Chem. Inf. Model.,
Target, 19, 281 292, doi: 10.3109/1061186X.2010.499463.
54, 1700 1716, doi: 10.1021/ci500080q.
73.
Chen, P., Zhang, X., Jia, L., Prud’homme, R. K.,
76. Li, Y., Han, L., Liu, Z., and Wang, R. (2014) Comparative
Szekely, Z., et al. (2014) Optimal structural design of man
assessment of scoring functions on an updated bench
nosylated nanocarriers for macrophage targeting, J. Control.
mark: 2. Evaluation methods and general results, J. Chem.
Rel., 194, 341 349, doi: 10.1016/j.jconrel.2014.09.006.
Inf. Model., 54, 1717 1736, doi: 10.1021/ci500081m.
74.
Nimje, N., Agarwal, A., Saraogi, G. K., Lariya, N.,
77. Nguyen, D. D., Xiao, T., Wang, M., and Wei, G. W. (2017)
Rai, G., et al. (2009) Mannosylated nanoparticulate carri
Rigidity strengthening: A mechanism for protein-ligand
ers of rifabutin for alveolar targeting, J. Drug Target, 17,
binding, J. Chem. Inf. Model., 57, 1715 1721, doi: 10.1021/
777 787, doi: 10.3109/10611860903115308.
acs.jcim.7b00226.
COMPUTER SIMULATION OF RECEPTOR LIGAND INTERACTIONS
MANNOSE RECEPTOR CD206 IN COMPARISON
WITH THE MODEL LECTIN CONCANAVALIN A
I. D. Zlotnikov and E. V. Kudryashova*
Faculty of Chemistry, Lomonosov Moscow State University,
119991 Moscow, Russia; E*mail: helenakoudriachova@yandex.ru
With using the methods of molecular dynamics and neural network analysis, computer modeling of the complexation
of mono and oligosaccharide ligands with the main (fourth) carbohydrate binding domain of the mannose receptor
CD206 (CRD4), as well as with the model receptor concanavalin A (ConA), was carried out for the first time. It is
shown that Con is a relevant CD206 (CRD4) model due to the similarity of the structural organization of the binding
sites and the high correlation of the values of the free energies of complexation r > 0.9 according to literature data and
computer modeling. The role of the main factors affecting the affinity of ligand receptor interactions is discussed: the
number and nature of carbohydrate residues, the presence of the Me group in the O1 position, the type of glycoside
bond in the dimannose. It is shown that the complexation of ConA and CD206 with ligands is energetically caused by
the electrostatic interactions of charged residues (Asn, Asp, Arg) with oxygen and hydrogen atoms in carbohydrates;
the hydrophobic and van der Waals components contribute less. A possible variant of additional stabilization of com
plexes due to CH π stacking interactions of Tyr with the Man plane is discussed. The role of calcium and manganese
ions in ligand binding processes has been studied. The free energies of complexation calculated in the course of mol
ecular dynamics correlate with experimental data (published for the model ConA): correlation coefficient r = 0.68.
The Pafnucy neural network was trained based on a set of PDBbind2020 ligand-receptor complexes, which signifi
cantly increased the accuracy of energy predictions to r = 0.8 and 0.82, respectively, for CD206 and ConA receptors.
A model of normalization of the complexation energy values for calculating the relevant values of ΔGbind, Kd is pro
posed. Based on the developed technique, the values of the dissociation constants of a series of CD206 complexes with
9 carbohydrate ligands of different structures not previously characterized were determined. The obtained data open
up prospects for using of computer modeling for the development of optimal drug carriers with the function of active
targeting of macrophages, and also determine the limits of the applicability of using ConA as a relevant model for
studying the parameters of CD206 binding to various carbohydrate ligands.
Keywords: molecular dynamics, artificial neural network, CD206, concanavalin A, macrophages
БИОХИМИЯ том 87 вып. 1 2022