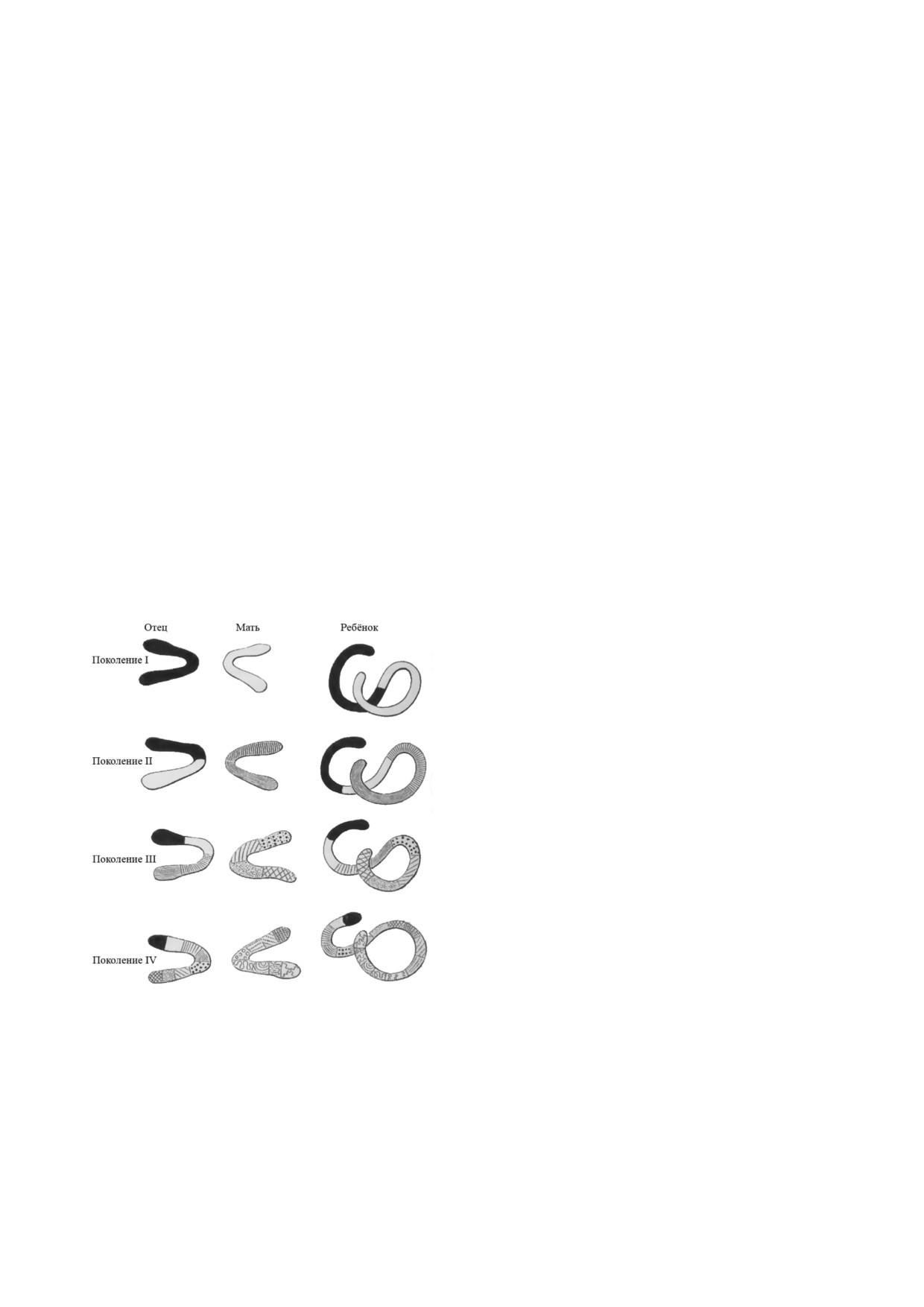БИОХИМИЯ, 2022, том 87, вып. 1, с. 126 - 149
УДК 576.311; 571.27
НАСКОЛЬКО ОБОСНОВАНЫ ПРЕДПОЛОЖЕНИЯ
О ЗАПРОГРАММИРОВАННОМ СТАРЕНИИ В ТЕОРИЯХ ВЕЙСМАНА?
Обзор
© 2022
P. R. Winterhalter*, A. Simm
Martin Luther University of Halle Wittenberg, 06120 Halle (Saale), Germany; e mail: winterhalter.patrick@gmail.com
Поступила в редакцию 02.10.2021
После доработки 30.11.2021
Принята к публикации 15.12.2021
Теории о преимуществах смерти и связанной с этим повышенной вероятности запрограммированного ста
рения противоречивы, и их поддерживает лишь меньшинство. Обоснованность этих предположений необ
ходимо проанализировать. С этой целью в данном обзоре рассмотрены различные подходы к обоснованию
полезности существующих гипотез и источники их возникновения, обращая внимание на возможные пре
имущества быстрой смены поколений, вызванной совместным действием различных эволюционных фак
торов. Были рассмотрены взгляды отца теорий о регулируемом старении - Вейсмана, который в конце сво
ей карьеры придерживался взглядов неадаптивной эволюции. Циркадные ритмы обсуждаются в теорети
ческом эксперименте, как возможный молекулярный источник регуляции старения.
КЛЮЧЕВЫЕ СЛОВА: Август Вейсман, теория старения, запрограммированное старение, семелпарность,
MASON модель, циркадные ритмы, часовые гены.
DOI: 10.31857/S0320972522010092
ВВЕДЕНИЕ. ЗАПРОГРАММИРОВАННОЕ
что нет чёткой тенденции к консенсусу во мне
СТАРЕНИЕ КАК ВЗГЛЯД МЕНЬШИНСТВА
ниях (<65% с одной или другой стороны), явля
ется ли старение генетически запрограммиро
Предполагается, что четыре незапрограмми
ванным, а не просто побочным продуктом несо
рованных эффекта вносят основной вклад в ста
вершенства или эволюционных ограниче
рение. Накопление повреждений, таких как му
ний [1]. Было ясно, что разногласия по вопросу,
тации/разрывы ДНК или агрегация/модифика
является ли старение запрограммированным
ция белков, а также износ биологических струк
или нет, отражают, скорее, расхождения во мне
тур считаются наиболее важными [1]. Более ши
ниях, чем в определениях [1]. Незапрограмми
роко распространены дезадаптации, такие как
рованные эффекты предоставляют множество
антагонистическая плейотропия, адаптация к
разных сценариев старения, что делает выявле
ограничениям, вызывающим решение инвести
ние основного игрока непростой задачей. Гораз
ровать либо в потомство, либо в продление жиз
до более вероятно, что все маркёры вносят опре
ни, а также гемодинамическая дисрегуляция,
деленный вклад, зависящий от генетики и ин
приводящая к разрушению регуляторных се
дивидуального воздействия окружающей среды.
тей [1]. Господствующее мнение о том, что ста
Такой же плавный переход, скорее всего, суще
рение само по себе не приносит пользы и, сле
ствует и для проблемы старения, является ли
довательно, не запрограммировано генетичес
оно запрограммированным или нет; реальная
ки [2], является часто возникающим впечатле
ситуация, скорее всего, зависит от потребностей
нием. Это оправдано идеей, что отбор может
вида. Разнообразие стратегий репродукции
происходить только при наличии различия фе
подсказывает нам это направление.
нотипа, приводящего к различию репродуктив
ного успеха [3]. Удивительно, но опрос в рамках
сообщества исследователей старения показал,
ПОЯВЛЕНИЕ ТЕОРИЙ
ЗАПРОГРАММИРОВАННОГО СТАРЕНИЯ
Принятые сокращения: ЦР - циркадные ритмы;
EGT - эволюционная теория игр; HDAC - гистоацетил
Семелпарность (гибель после репродук
трансфераза; RQH - гипотеза Красной Королевы.
ции) - это явление, известное для нескольких
* Адресат для корреспонденции.
классов растений и животных. Двухлетние рас
126
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
127
тения представляют собой прекрасный пример:
ным и неудивительным, если бы оно было упо
они выживают в условиях холодной зимы в сво
мянуто раньше. Голдсмит отметил далее, что в
ём ареале, но умирают в последующем сезоне,
этом случае больше генетических вариантов вида
скорее всего, в результате запрограммированно
может возникнуть в течение того же самого пе
го жизненного цикла. Существует множество
риода времени [14]. В дополнение к ограничен
теорий, объясняющие возникновение одно
ному числу вариантов, возникающих в результа
кратной репродукции [4]. Обоснования сезонной
те идентичной репродукции, Леопольд отметил
смертности, наблюдаемой для самцов некото
преимущества смерти в способности к распрост
рых видов сумчатых, несколько раз претерпели
ранению. Он написал, что, если благоприятные
изменения. Сначала считалось, что такая сезон
генетические изменения происходят в однолет
ная смертность происходит из за невозможнос
них сорняках, распространение этих изменений
ти накопить достаточные жировые отложения
внутри популяции происходит с максимальной
для сезонов дефицита продуктов питания, затем
скоростью, так как полный оборот размножаю
последовала идея, что такая смертность являет
щейся популяции происходит каждый год [7].
ся результатом рудиментарной неадаптивной
Эта концепция о более быстром распростране
тенденции в этой группе - побочный продукт
нии полезного признака в уже установившемся
преимущественного использования белковых
ареале обитания была далее развита математи
запасов [5]. Эта концепция была опровергнута
чески Либертини, основываясь на теории смер
идеей о том, что смертность самок может быть
ти, ускоренной старением [15]. Затем Скулачёв
причиной семелпарности самцов [6]. Осознание
представил молекулярно биологические пре
того, что быстрая смена поколений предостав
имущества смерти, описывая полезность такого
ляет преимущества, не имеет смысла даже упо
строго регулируемого процесса уничтожения,
минать для такого стиля жизни или можно избе
как апоптоз для предотвращения аутоиммуните
жать упоминания этого целенаправленно, по
та, поддержки морфологических изменений или
скольку это может способствовать концепции
для удаления опасных клеток [16]. В этом случае
запрограммированной смерти.
преимущество смерти на благо вида было назва
Сокращение продолжительности жизни
но феноптозом, и были предложены три мишени
должно привести к уменьшению числа потом
механизмов старения: теломеры и теломераза,
ков, что является сильным аргументом против
индукция белков теплового шока и активные
идеи о индуцировании смерти. Действительно,
формы кислорода (АФК). В дополнение к семел
высокая репродуктивность очень полезна, но не
парности, острый феноптоз включает, например,
всегда является самой успешной стратегией. Од
апоптоз у бактерий, индуцированный фагами,
нако Леопольд упомянул в 1961 г., что у многих
или септический шок у животных. Согласно
растений старение имеет различное и положи
Скулачёву, оба эти процесса являются реакцией
тельное значение с точки зрения экологической
живого организма, направленной на остановку
адаптации, естественного отбора и эффектив
распространяя патогена, которая защищает всю
ности внутренней физиологии [7]. Для случая ес
популяцию [17]. Напротив, предполагаемый по
тественного отбора он также написал, что долгая
жизненный медленный феноптоз встречается у
продолжительность жизни многолетних расте
всех организмов и, предположительно, связан с
ний обеспечивает защиту от быстрых эволюци
изменённой скоростью реакции активных форм
онных изменений. Если бы существовали виды
кислорода в митохондриях [18, 19]. Несколько
растений, которые не старели и были бы почти
позже Либертини тоже предложил возможный
бессмертны, изначальная популяция продолжа
механизм регулируемого старения. В этой связи
лась бы бесконечно, воспроизводя исходный ге
необходимо напомнить, что во всех клетках (за
ном, и поэтому эволюционная способность это
исключением эмбриональных и опухолевых)
го вида к адаптации к изменениям в окружаю
число удвоений напрямую связано с укорочени
щей среде была бы минимальна. Леопольд утвер
ем теломер. Таким образом, была выдвинута ги
ждал, что способность растений стареть является
потеза, согласно которой теломеры защищены
катализатором эволюционной адаптации [7].
кэпом ДНК белкового комплекса и становятся
Голдсмит детально рассмотрел проблему умень
уязвимыми для репликативного старения только
шения геномного разнообразия в свете способ
в некэпированном состоянии. В этом случае пе
ности эволюционного развития [8-12]. Он пред
риод времени, когда теломера остаётся незащи
положил, что более быстрая смена поколений
щённой, кажется пропорциональным укорачи
должна ускорять эволюцию, так как больше по
ванию [20]. Была предложена гипотеза, что регу
ложительных изменений может произойти в
ляция, зависящая от длины теломер, контроли
группе с укороченной продолжительностью жиз
рует способность кэпа взаимодействовать с тело
ни [13, 14]. Это преимущество кажется очевид
мерами. Либертини предположил, что субтело
БИОХИМИЯ том 87 вып. 1 2022
128
WINTERHALTER, SIMM
мерные регуляторные последовательности и их
КОНТРОВЕРСИЯ ГРУППОВОГО ОТБОРА
транскрипция, уменьшающаяся с возрастом, из
меняют покрытие теломер и приводят к клеточ
Возможно, наиболее существенная причина
ным изменениям, имеющим решающее значе
отказа от идеи запрограммированного старения
ние для симптомов старения, с особым акцентом
в настоящее время основана на представлении о
на последовательности TERRA [21, 22].
том, что индивидуальный отбор является гораз
Однако Скулачёв первоначально ссылался
до более эффективным, чем отбор на уровне
на предположения Вейсмана [16], выдвинутые
групп [2]. Вскоре после объявления предпочте
около 140 лет назад, где смерть в условиях пос
ния индивидуального отбора, поддержанного
тоянно ограниченных ресурсов окружающей
Smith [26], была предложена модель с похожими
среды рассматривалась как неизбежность, пото
математическими допущениями, в которой
му что старые и, следовательно, изношенные
альтруистическое поведение имеет преимущест
особи бесполезны и даже опасны, так как зани
во перед личной приспособленностью, не нару
мают место и используют ресурсы более моло
шая концепции индивидуального отбора [27].
дых и лучших особей (стр. 30-31) [23]. Точно не
В модели были использованы гетерогенность
указано, что для Вейсмана означает «лучше»:
популяции, изоляция, отбор и смешение, что
более жизненно или более приспособленно?
привело к постепенному увеличению числа
Однако из за обилия контекста и примеров ка
альтруистов. Такая модель, основанная на изо
жется очевидным, что освободившееся место
ляции и неравномерном распределении субти
выгодно для потомков, и это ускоряет адапта
пов в популяции, по видимому, отражает есте
цию популяции к окружающей среде [23]. В сво
ственную ситуацию гораздо лучше, чем одна
их поздних произведениях Вейсман приводит
большая гомогенная популяция. Далее спорно
конкретный пример конъюгации инфузорий,
обсуждается, основано ли альтруистическое по
указывая, что смешение видов незаменимо для
ведение больше на родственном отборе чем на
развития вида, и что смерть необходима для ис
групповом? Однако групповой отбор может осу
коренения несмешивающихся организмов
ществляться через распространение преиму
(стр. 150) [24]. Вейсман не знал о разнице между
ществ родственного отбора внутри вида. Га
спариванием и конъюгацией и определял все
мильтон предложил первые математические мо
типы взаимодействий, сопровождающиеся со
дели о требуемой степени родства для родствен
единением наследственной информации двух
ного отбора уже в 1964 г. [28]. По видимому, су
организмов, как амфимиксия. Он уточнял, что
ществует также плавный переход между типами
индивидуальная изменчивость составляет осно
отбора (индивидуальный, родственный, груп
ву теории отбора (стр. 21), и что смешение приз
повой). Примером могут служить виды бакте
наков делает возможной эту индивидуальную
рий, потому что большинство их них образуют
изменчивость, которая отвечает за образование
биоплёнки, которые часто бывают либо моно
новых видов путём отбора (стр. 29) [25].
клональной либо близкородственной природы.
Следовательно, он рассматривал смерть, как
Таким образом, родственный отбор может пре
преимущество, позволяющее избежать эволю
вратиться в групповой отбор при определённых
ционного застоя путём остановки или ограниче
условиях, как, например, катастрофическое со
ния вариаций. Предположения, выдвинутые
бытие [21]. Кроме того, точные модели структу
Вейсманом, убедили его в существовании оп
ры популяции с допущением стандартной тео
тимальной продолжительности жизни, и он
рии естественного отбора могут быть более
проповедовал внутреннюю регуляцию жизни,
простым и лучшим подходом, который также
сравнимую с другими приспособлениями, таки
может объяснить групповой отбор [29].
ми как размер тела (стр. 8-9, 41) [23]. Изнаши
Однако предположим, что существует естест
вание биологических структур (тела) не прини
венный отбор на уровне выше индивидуального.
малось в качестве ограничивающего возраст
В этом случае ситуация может заключаться в ус
фактора. Вместо этого он постулировал внут
тановлении равновесия между благоприятными
ренне ограниченный потенциал пролифера
и неблагоприятными факторами в группе.
ции (стр. 27-28, 86-90), основываясь на наблю
Существует несколько примеров, когда при
дениях за эмбриогенезом (стр. 39-40), доказан
рода может поощрять подчинение индивидуаль
ных Хейфликом ~80 лет спустя.
ных интересов групповым. Предупреждающая
Вейсман был первым учёным, который пы
окраска - это наиболее яркий пример, когда ин
тался объяснить полезность смерти в эволюци
дивидуальные интересы подавляются жертвова
онном аспекте и продвигал в начале своей карь
нием на благо вида, включая их собственное по
еры концепт, известный сейчас как запрограм
томство. Неясно, как такое явление могло воз
мированное или регулируемое старение.
никнуть, когда разнообразие окрасок может ле
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
129
жать в основе [30]. Кроме того, в отличие от дру
отцом теории многоуровневого отбора [27].
гих мигрирующих животных, некоторые лососи
В этом случае гены эгоистического поведения
перестают есть, когда попадают в пресную воду
разбавятся со временем, особенно если прини
для спаривания, даже если они теряют массу
мается более распространённая модель полиге
энергии каждый день без пищи [31]. Это пара
нетического промежуточного наследования, а не
доксальное явление может принести пользу для
доминантного. В этом случае, скорее всего, уста
всего вида, потому что в противном случае вся
новится равновесие (Nash), вероятно, зависящее
река будет вычищена массовым наплывом хищ
от степени изоляции, гетерогенности субгруппы
ников. Индивидуум, вероятно, выиграет от еды,
и степени невыгодности эгоизма для группы.
в то время как вид пострадает, так как потомство
Отказ от пищи или от потомства, по види
прошлых лет и их пища будут съедены. По срав
мому, также противоречит личным интересам.
нению с примером подчинения рабочих медо
Нет сомнения в том, что в этом случае часто
носных пчел яйцекладок аргумент родственно
ожидается компенсация, как, например, требо
го отбора кажется неоправданным для лососей,
вание вернуть обратно ресурсы, что наблюдает
так как особи чаще всего не являются близкими
ся среди вампировых летучих мышей [33]. Для
родственниками. Однако пример трутовок
многих организмов, живущих в группах, наблю
представляет собой ещё один пример, когда от
дается такая кооперация, когда особи должны
рицательные последствия для группы будут за
подавить свои эгоистические желания, чтобы
маскированы, так как последующее поколение
поддержать другого члена группы, который мо
будет ослаблено чрезмерным эгоистическим по
жет помочь им в будущем, что делает родствен
ведением и поэтому уменьшит своё социальное
ные/групповые связи сильнее. В этом случае ус
поведение. Обычно проблема решается через
пех поведения одной особи зависит от поведе
принуждение к стерильности и охраной [32].
ния других членов группы. Определение альтру
Это показывает, что социальные группы могут
изма не совсем подходит во многих таких случа
защищать себя от разрушающего эгоистическо
ях. Похоже, он основан на другой форме эгоиз
го поведения, что может быть примером того,
ма, которую можно было бы назвать «соучасти
как могут возникнуть механизмы, предотвраща
ем», и можно рассматривать как дополнитель
ющие поведение индивидуальных особей, не
ный тип преимущества более высокого уровня.
благоприятное для группы, даже если проблема,
В целом соучастие, такое как внутривидовая
провоцирующая это поведение, находится не на
защита (предупреждающий сигнал или окраска,
первом плане эволюционного отбора. Надзира
групповая защита, синхронное спаривание) ка
тельство, такое, как поедание неоплодотворён
жется достаточно сильным фактором, чтобы во
ных яиц, вероятнее всего, предопределяется ге
зобладать, даже в случае, когда такое поведение
нетически, и потеря его приводит к долговре
может негативно отразиться на интересах особи.
менным негативным последствиям. Эти сооб
щества выживут лучше, если им удастся прочно
встроить полицейское поведение в свою генети
РАЗНООБРАЗИЕ РАЗЛИЧИЙ
ку, или, ещё лучше, если такое поведение будет
И ИХ НАСЛЕДОВАНИЕ
связано с жизненно необходимыми процесса
ми. Описанные типы поведения кажутся легко
Необходимо отметить, что Голдсмит разли
разрушаемыми преимуществами для индивиду
чал групповой отбор и концепт эволюционируе
альных особей, но каким то образом они оста
мости, который, согласно его взглядам, создаёт
ются, что может быть объяснено наличием от
условия (вариации), которые должны существо
бора более высокого уровня. Наконец, половое
вать прежде, чем отбор начинается [14]. Он при
размножение само является, скорее всего, наи
водит такой пример. Можно считать, что сред
более распространённой формой групповой се
ний рост индивидуума (здесь можно использо
лекции, так как оно, как кажется, больше связа
вать любую другую характеристику) оптимален
но с опасностями и ущербом для особи, но пре
с точки зрения приспособленности. Следова
имуществами для вида.
тельно, все животные, которые ниже или выше
Признаки, которые выгодны для индивиду
среднего, менее приспособлены. Поэтому особи
ального организма, но не выгодны для группы,
в популяции, состоящей из клонов среднего жи
скорее всего, приведут к уменьшению популяции
вотного, будут более приспособлены, чем боль
в этом конкретном регионе. Этот эгоистический
шинство в популяциях с большими различиями.
признак должен превалировать над неэгоисти
Таким образом, изменчивость способствует раз
ческими признаками в других группах с большей
витию, но снижает приспособленность.
численностью популяции, согласно уже упомя
Для эволюции скорость изменчивости, по
нутой математической модели, предложенной
видимому, очень важный параметр, поэтому
9 БИОХИМИЯ том 87 вып. 1 2022
130
WINTERHALTER, SIMM
классификация необходима. Существует ско
ких вариантов, что было понятно Вейсману
рость внутренней изменчивости, связанная с ви
(рис. 1), и изменениями их регуляции.
дом, которая зависит от таких факторов, как ско
Скорость комбинативной изменчивости за
рость мутаций или внутригеномный обмен. Об
счёт полигенного наследования, обеспечивае
ратная транскриптаза вируса иммунодефицита
мая объёмом и доступностью геномного обмена
человека имеет необычайно высокую частоту
при скрещивании (или через плазмиды), явля
ошибок копирования нуклеотидов (1/1700), что
ется эффективным способом приобретения раз
считается причиной высокой генетической ва
нообразия. Разнообразие обмениваемого ге
риабельности этого вируса [34]. Высокая частота
номного материала зависит от гетерогенности,
мутаций является одной из причин, по которой
количества и присутствующего разнообразия
атака иммунной системы и лечение не дают ре
различий в группе. Многие их этих параметров
зультатов, что является явным преимуществом
зависят от окружающей среды и также могут
выживания вируса. Ещё более высокая частота
косвенно зависеть от вида организма и его ха
мутаций привела бы ко многим ошибкам, кото
рактеристик, таких как повышенная мобиль
рые, вероятно, могли бы привести к нефункцио
ность, групповое спаривание или выбор партнё
нальному организму. Оптимальная скорость
ра. Скорость обмена или рекомбинации, по ви
внутренней изменчивости, по видимому, регу
димому, снова зависит от индивидуального от
лируется классическим индивидуальным отбо
бора. Однако группа должна решить, предпочи
ром идеала. Такие генетические вариации всегда
тает ли она комбинативные вариации однопо
отражаются на всей цепи взаимодействий, в ре
лому размножению.
зультате возникает целая череда последствий,
Внутренние и комбинативные различия при
которая всегда более или менее многофактор
водят к разнообразию в генофонде, что является
ная. Однако единичные гены очень редко ответ
очень полезной предпосылкой для эволюции, в
ственны за существенные изменения и фенотип
то время как тенденция к изменениям, вероятно,
организма. Скорее всего, изменения вызывают
ориентирована в сторону интересов вида. Неяс
ся сложной комбинацией различных генетичес
но, необходимы ли уже существующие различия
для целей селекции без корректировки, учиты
вающей потребности вида. Поэтому вариации,
скорее всего, не могут сами по себе обеспечить
отбор более высокого уровня. Ответ на вопрос о
том, является ли смена поколений, ответствен
ная за постоянное образование вариаций, суще
ственным фактором (прямым или косвенным)
для естественного отбора на уровне выше инди
видуального, может быть ключевым. Поколение
с гетерогенными вариациями и выгоды смены
поколений должны вносить вклад в способность
организма адаптироваться, в то время как умень
шение количества вариаций должно негативно
отражаться на группе.
Существуют ли примеры в природе, когда
ослабленная адаптивность приводит к разруши
тельным последствиям для всего вида? Такие
примеры могут подсказать, что случится, если
изменится регулирование процесса смены поко
лений. С этой точки зрения дальнейшие иссле
дования в области репродукции необходимы.
Рис. 1. Изменения наследуемых характеристик в каждом
ДЕФЕКТЫ ЭВОЛЮЦИИ
поколении, согласно Вейсману (стр. 32) [35]. Вейсман де
лает вывод, что при передаче родительского наследуемого
вещества его количество должно уменьшаться вдвое.
Истинный партеногенез путём однополого
В противном случае количество генетического материала
размножения обнаружен примерно в 90 видах
будет удваиваться с каждым поколением. Это уменьшение
позвоночных [36] и, по видимому, является дос
никогда не происходит одинаково. Поэтому создаются но
тойным упоминания примером ослабленной
вые комбинации наследуемого вещества, что приводит к
разнообразию признаков среди детей и индивидуальным
адаптивной способности. Небольшое количест
различиям (стр. 67-75)
во видов по сравнению со всем семейством поз
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
131
воночных говорит от том, что половое размно
Лайвли экспериментально поддержал идею
жение более предпочтительно эволюционно.
RQH, наблюдая более высокую паразитарную
Это вызывает гендерный парадокс, поскольку
инвазию у однополых рыб (Poeciliopsis) по срав
партеногенез имеет такие преимущества, как
нению с двуполыми аналогами [42]. Эта кло
отсутствие двойных затрат на самцов, не рожаю
нальная гибридогенная рыба существует более
щих детей, и всех трудностей при спаривании
чем 100 000 поколений [43]. Старейшая извест
или поиске партнёра. Smith [37] показал мате
ная однополая линия отделилась от своего дву
матически, что существование пола не является
полого позвоночного родственника (Ambystoma
предпочтительным для эволюции из за сущест
barbouri) примерно 5 млн лет назад [36]. Эта са
венной неэффективности по сравнению с бес
ламандра создаёт много генетических вариаций
полым размножением.
из за сложных межгеномных обменов и через
Почему же тогда спаривание существует?
включение чужого генетического материа
Уильямс первым ответил на этот вопрос в 1975 г.,
ла (спермы) от двуполых особей [36], что могло
сравнив бесполое размножение с копией лоте
быть необходимо, чтобы избежать вымирания.
рейного билета. В отличие от этого, спаривание
Тем не менее остаётся вопрос: является ли это
производит несколько новых комбинаций цифр
все таки истинным партеногенезом, и не явля
в лотерейном билете, что увеличивает вероят
ются ли 5 млн лет просто мигом в истории эво
ность выиграть эволюционный джекпот [38].
люции?
Эволюционный биолог Белл показал в своей ра
Клональное размножение у позвоночных яв
боте в 1982 г., что бесполое размножение наблю
ляется, по видимому, примером того, как нару
дается реже в больших комфортабельных ареалах
шение адаптивной способности может привести
обитания, но в основном встречалось в неболь
к видовому отбору. Он не направлен на выявле
ших, более экстремальных, где экологическое
ние того, оказывает ли и какой эффект (генети
пространство необходимо быстро и чаще засе
ческая изменчивость, смена поколений или
лять [38]. Ранее считалось, что половое размно
распространение выгод) наибольшее влияние.
жение необходимо для адаптации к сложным ус
Тем не менее очевидно, что дефекты эволюции
ловиям обитания. Белл пришёл к выводу, что пол
могут привести к тупиковому концу. В любом
и, следовательно, более высокая степень генети
случае либо истинный партеногенез у позвоноч
ческой изменчивости необходимы для преобла
ных должен найти обратный путь к половому
дания на больших стабильных территориях, ко
размножению, расширить генетическое разно
торые часто уже заселены и довольно сложны.
образие (как в случае Ambystoma barbouri), либо
Он считал, что пол используется во внутривидо
вымирание неизбежно. Любопытно, что Либер
вой конкуренции, но также упомянул сходство с
тини тоже исследовал сходства между полом и
гипотезой Красной Королевы (RQH), предло
старением и отнёс оба эти явления к фенопто
женной в 1973 г. Van Valen [38]. У нескольких ав
зу [44]. Он показал слабость теории RQH мате
торов были параллельные ассоциации пола и
матически и возродил старейшую классическую
RQH [39-41], что в настоящее время является
идею о преимуществах полового размножения -
наиболее общепринятым объяснением развития
гипотезу Фишера-Мюллера. Интересно, что в
полового размножения. RQH описывает ситуа
ограниченной популяции снижение полового
цию, когда продолжающаяся эволюция необхо
преимущества задерживается введением третье
дима, чтобы выжить в постоянной межвидовой
го полезного гена. Многие полезные гены, кото
конкуренции, такой как взаимоотношения меж
рые могут увеличить выгоды этого классическо
ду паразитом и хозяином или хищником и жерт
го предположения, могут быть вовлечены в про
вой. Следует отметить сходство между аргумен
цесс смены поколений в результате плейотроп
тами Вейсмана, Белла и Голдсмита, указываю
ного наследования. Основная идея преимуще
щими, что генетическое разнообразие - это дви
ства может быть частично связана с Вейсманом,
жущая сила эволюции. Также интересен факт,
согласно которому амфимиксис обеспечивает
что Вейсман уже показал преимущества беспо
непрерывную рекомбинацию характеристик,
лого размножения в ареалах обитания с быстро
полезных для групповой эволюции [24, 25].
меняющимися экстремальными условия
ми (стр. 167-168) [24], а далее описал половое
размножение как неоценимое преимущество для
ВОЗМОЖНЫЕ ЭВОЛЮЦИОННЫЕ
приспособляемости видов (стр. 55) [25]. Следо
МЕХАНИЗМЫ СТАРЕНИЯ
вательно, его можно рассматривать как пионера
идеи RQH, поскольку его аргументы основаны
Однополое размножение сужает диапазон
на преимуществах смешения, не ограничивая се
различий в группе, что непросто восстановить,
бя межвидовой конкуренцией.
так как для этого требуются особые условия и,
БИОХИМИЯ том 87 вып. 1 2022
9*
132
WINTERHALTER, SIMM
скорее всего, много времени. Однако старение и
увеличивается значительно для случая полового
его потенциальные преимущества не так просто
размножения, приводя к существенным выго
определить. Признаки старения тоже подверга
дам старения для двуполых организмов по срав
ются нормальному распределению, что затруд
нению с однополыми. Более быстрая смена по
няет объяснение того, почему незначительные
колений создаёт больше вариаций во времени.
изменения средней продолжительности жизни
Следовательно, η надо разделить на (1 - Δg);
могут иметь преимущества для группы. Экспе
fg,Δg = (1 + η/(1 - Δg))g*Δg - 1. При вычислениях
риментально было бы сложно проверить, имеют
выгод смены поколений для популяции необхо
ли субпопуляции с пожилыми людьми повы
димо учитывать её размер и распределение ва
шенную скорость вымирания.
риаций.
Ускоренная смена поколений, связанная с
освобождением места для более быстрого рас
пространения и большим количеством генети
ВОЗМОЖНЫЕ ПУТИ
ческих вариантов, возникающих за определён
ДЛЯ СОКРАЩЕНИЯ ЖИЗНИ
ный промежуток времени, может дать лучшее
объяснение преимуществ старения. Несмотря
В мире, где распределение в каждой группе
на то что трудно определить вклад каждого эф
несовершенно, субпопуляции обитают в более
фекта, можно считать, что, скорее всего, они
или менее изолированных ареалах и имеют не
действуют синергетически. Однако более быст
которые различия, такие как средняя продол
рое распространение полезных признаков бла
жительность жизни. Это происходит потому, что
годаря генетическому обмену, такое как поло
эволюция обычно концентрируется на одних
вое размножение, может происходить только,
признаках, в то время как другие побочные ха
если занятое место освобождается. Либертини
рактеристики остаются несовершенными. Ста
обсуждал распространение преимуществ полез
рение может проявиться в популяции как по
ных признаков, которые могут произвести наи
бочный эффект более выгодной адаптации или
более значимый эффект [15, 44]. Тем не менее
генетического дрейфа. Эффект разделения был
выгоды последовательных улучшений для ко
установлен на высокоинбредных линиях мы
роткоживущих организмов необходимо обсу
шей, где дрейф генов практически исключён.
дить детально, потому что это практически нигде
Тем не менее это не предотвращает генетичес
не обсуждалось в математических терминах.
кие изменения, за которыми следуют сущест
В связи с этим масштаб таких преимуществ мо
венные изменения фенотипа [45]. Если преж
жет быть до сих пор недооценён. Весьма веро
девременная смерть происходит из за адаптив
ятно, что эволюционный процесс не происхо
ных или неадаптивных причин, таких как пред
дит линейно во времени, что было бы выгодно
расположенность к раку или инфаркту, субпопу
эволюции. Приобретённые признаки должны
ляция с более короткой продолжительностью
умножаться синергично, напоминая процесс,
жизни должна будет эволюционировать быстрее
подчиняющийся закону сложных процентов, в
и, следовательно, будет доминировать в группе
котором существенный экспоненциальный
как более адаптированная. Скорее всего, это
рост наблюдается только через несколько поко
может произойти через несколько этапов разде
лений.
ления, смешения и отбора, как описано для раз
Адаптация, зависящая от поколения [fg], долж
вития предупредительного крика [27]. Можно
на определяться средним по популяции значением
ли рассматривать в этом случае предрасполо
предоставленного преимущества на каждое поко
женность к болезням, как предпосылки запрог
ление в процентах [η] и числом поколений [g] как
раммированной смерти? Весьма вероятно, что
показателя степени. Чтобы понять влияние из
такой непреднамеренный отбор привёл к раз
менения на старение, эти вычисления должны
личным причинам смерти, поскольку природе
быть расширены включением изменяющейся
все равно, какой путь был выбран, если резуль
скорости смены поколений в процентах [Δg] до
тат устраивает. Необходимо определить эти не
окончательного зависящего от поколения адап
адаптивные несовершенства, которые могут
тационного изменения [fg,Δg]; fg,Δg = (1+η)g*Δg - 1.
косвенно поддерживать старение/смерть.
Количество возможных полезных относитель
То, что позже можно было бы считать полез
ных различий (относительную изменчивость) [θ]
ной смертностью, могло быть вызвано окружа
включает внутренние и совокупные различия, и
ющей средой по совершенно другой причине.
они должны быть учтены в η вместе с преиму
Примером такой ситуации могут служить бар
ществами распространения (в согласии с идея
хатцы (популярные декоративные растения),
ми Либертини). В результате этого θ сильно за
которые не выживают зимой в Центральной Ев
висит от способа размножения. Эта величина
ропе и могут распространяться только семена
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
133
ми. Такая вынужденная «однолетность» может
программированное старение не может разви
проявиться генетически со временем.
ваться [2]. Следует добавить, что в процитиро
RQH даёт ещё одно объяснение того, почему
ванной статье Кейтли упомянуты два альтерна
старение могло эволюционировать. Далее старе
тивных расчёта с примерно втрое более высоки
ние было предложено как адаптационный меха
ми скоростями мутаций. Но более существен
низм, который является результатом накопле
ным фактором является то, что мутации - это
ния повреждений, а не конкретной програм
только один источник генетической вариабель
мы [46], что было основано на идее RQH с пос
ности, помимо гомологичной рекомбинации,
тоянной конкуренцией коэволюционирующих
иммиграции, эмиграции или транслокации.
паразитов и хищников. Этот подход, по види
И наиболее важным фактором, определяющим
мому, основан на идее выгодного, но косвенно
разнообразие организмов, является комбинация
го отбора. Также упоминалось, что старение
генов (полигены). Принимая это во внимание,
могло возникнуть, чтобы ограничить распрост
можно заключить, что мутации вряд ли являют
ранение болезней [47]. Тем не менее RQH ка
ся основными наследственными двигателями
жется наиболее разумным объяснением того,
эволюции. Эта идея может быть поддержана
почему мог возникнуть адаптивный механизм
долгосрочным эволюционным экспериментом,
старения. Эта гипотеза описывает потребность в
в котором мутации, по видимому, подтвержда
высокой способности к адаптации и, следова
ют эволюционные шаги. Все же выгодное нов
тельно, в большом количестве вариаций. Лайв
шество появилось благодаря модуляции генома,
ли отметил, что в расчётах для отбора на половое
когда дупликация гена транспортёра цитрата
размножение необходимы как распространен
сопровождалась дополнительной транслокаци
ные, так и высоковирулентные инфек
ей правильно расположенного аэробного про
ции [48-50], что ограничивает общую примени
мотора [55]. Редкость таких явлений в большом
мость RQH [51]. Он установил, какая может
геноме должна сопровождаться, помимо смены
быть цена возникновения пола, если использу
поколений, высокой частотой. В противном
ется подход, принимающий во внимание допол
случае такая ситуация была бы маловероятной.
нительные расходы для оценки потенциальных
Благодаря мейозу и всем последующим типам
затрат на сокращение продолжительности жиз
рекомбинаций такие необычные события более
ни [51]. Оказалось, что необходимы нереально
вероятны во время спаривания.
большие изменения/адаптации, чтобы объяс
Кейтли также предполагает, что вредные му
нить возникновение пола в этих моделях. Две из
тации могут удаляться, потому что потомство с
этих моделей пытаются объяснить возникнове
негативным воздействием не будет развивать
ние пола только одной эволюционной потреб
ся [54], что демонстрирует недооценный при
ностью. Однако в дополнение к взаимоотноше
родный фильтр. Почти 43% из более чем 50 000
ниям хозяин/паразит и хищник/жертва сущест
рожавших женщин сообщили, по крайней мере,
вуют и другие движущие силы адаптации много
об одном самопроизвольном аборте в первом
мерного характера, такие как разнообразная
триместре [56], и при этом нет сообщений о
внутри и межвидовая конкуренция за ресурсы
проблемах с оплодотворением. Беременность -
(пища, пространство) и приспособление к окру
это довольно затратный и опасный процесс, и
жающей среде. Только определенный процент
непригодные к жизни организмы, скорее всего,
адаптаций может быть необходим для конкрет
будут удалены на ранних стадиях процесса. Рав
ной цели межвидовой борьбы. Следует отме
ного распределения между вредными и полез
тить, что симуляции, в которых смерть кажется
ными признаками не следует ожидать, так как
полезной на основании сокращения жизни по
явно негативные будут заранее отобраны. Таким
колений [15, 52, 53], были отклонены на основа
образом, частота мутаций не кажется подходя
нии таких же аргументов о нереально быстро из
щим инструментом для предположения о спо
меняющихся полезных мутациях и условиях ок
собности образовывать новые вариации в каж
ружающей среды [2].
дом поколении. Предполагается, что суммарное
Далее было отмечено, что частота спонтан
количество преимуществ может быть больше,
ных мутаций у Drosophila примерно одна на по
особенно в случае полового размножения. Од
коление [54], и из этого был сделан вывод, что
ной из таких выгод может быть то, что спарива
количество полезных мутаций должно быть го
ние помогает удалять вредные мутации путём
раздо меньше. А мутации, вызывающие сущест
рекомбинации [57-59] - ещё одна гипотеза, ко
венный положительный эффект, должны быть
торая может объяснить развитие полового разм
особенно редкими. Таким образом, чтобы дос
ножения (гипотеза храповика Мюллера). Пре
тичь более реалистичных уровней мутаций, доля
дотвращение проблемных генетических накоп
носителей должна быть настолько мала, что за
лений, таких как мутации, также может способ
БИОХИМИЯ том 87 вып. 1 2022
134
WINTERHALTER, SIMM
ствовать использованию преимуществ смены
на то, как определенные навыки отдельных фе
поколений. Однако другой аргумент, отрицаю
нотипов улучшаются или терпят неудачу с тече
щий идею выгодности смены поколений, ука
нием времени. Изменение приспособленности
зывает, что некоторые модели не подходят для
уникальных фенотипов, в свою очередь, влияет
полового размножения, так как половое раз
на частоту их встречаемости в популяции. In sili
множение делает возможным возникновение
co модели могут потребовать большей гибкости в
поколения детей, которые объединяют нестаре
разных конкурирующих стратегиях, чтобы обес
ющий генотип одного родителя с обнаруженной
печить аутентичное поведения эволюции, и кон
полезной мутацией (мутациями) у другого роди
куренция может быть самым критическим ком
теля - потомство «мошенников» [2]. Этот аргу
понентом, увеличивающим давление естествен
мент подходит для симуляции межвидовой кон
ного отбора, которое двигает прогресс. В случае
куренции и гомогенно распределённых агентов.
такого многомерного противостояния в симуля
Предположим, что старение (адаптивное или
ции агентам должно быть разрешено умирать,
косвенное) может иметь большее значение в
если их фенотип неконкурентоспособен по
межвидовой конкуренции, что является наибо
сравнению с резвившимися потомками. Незави
лее разумным объяснением для развития поло
сящей от возраста экологической смертности,
вого размножения. В этом случае уже существу
связанной с окружающей средой [γ], может быть
ющие модели с несмешивающимися компонен
недостаточно для поддержки естественного ко
тами должны подойти для выяснения полезнос
личества новых фенотипов. В этой модели мно
ти старения. Однако до сих пор модели не могут
гие потомки не будут возобладать так же, как их
объяснить само появление старения.
родители, что не является проблемой, так как
некоторые новые агенты будут иметь преимуще
ства, которые подходят лучше для будущих супер
СИМУЛЯЦИЯ СТАРЕНИЯ С ПОМОЩЬЮ
стратегий групп, и такие особи будут размно
МОДЕЛИ MASON
жаться в среднем более эффективно. Согласно
этому предположению, значение имеет не общее
MASON - это регулярно используемая мо
количество, а общее выживание потомства, ко
дель [60] для имитации эффектов изменённой
торое может быть обеспечено несколькими
продолжительности жизни, когда агенты сохра
адаптациями, такими как меньшее по количест
няются в двумерном мире, двигаясь, размножа
ву, но более развитое потомство. Поэтому, если
ясь и умирая [2]. Чтобы проверить предположе
множественная конкуренция отсутствует, как
ния Голдсмита, воспроизводство агентов в
могут развиваться сложные поведенческие стра
MASON зависит от достижения возраста зрелос
тегии? Если такие стратегии не возникают, то не
ти и от набора генов, которые могут иметь значе
будет возможности развития отбора более высо
ния (аллели) в пределах от 0 до 1, которые вносят
кого уровня, что требуется для старения. Макси
аддитивный вклад в вероятность воспроизведе
мальная продолжительность жизни, кажется,
ния (фертильность). Эволюция в этом случае
увеличивается постоянно в модели, адаптиро
направленная, так как фертильность имеет толь
ванной согласно гипотезе Голдсмита. Возможно,
ко положительное значение без всяких недос
это единственный вариант, при котором возмо
татков, таких как повышенное потребление
жен успех группы. Такое отсутствие альтернатив,
энергии. Кроме того, в модель введён переклю
по видимому, является причиной постоянного
чатель, утверждающий, что аллели, мало влияю
увеличения продолжительности жизни.
щие на фертильность, будут постепенно утеря
Наиболее убедительные модели MASON [2]
ны. Чтобы это предотвратить, направление от
основываются на концептах Martins [52, 53].
бора периодически переключается в противопо
В первой модели, предложенной в 2011 г., вновь
ложную сторону (т.е. в этом случае аллели с ма
созданные агенты могут конкурировать с суще
лыми значениями приводят к высокой фертиль
ствующими за место обитания, причём каждая
ности). Необходимость переключателя для про
индивидуальная особь [i] преобладает пропорцио
должения эволюции кажется неестественной
нально fi(t). Агент родитель [i] и потомок [j] кон
для моделирования постоянного развития. Бо
курируют во времени [t], и агент родитель выжи
лее гибкие и требовательные факторы могут по
вает с вероятностью fi(t)/(fi(t) + fj(t)). Родитель
надобиться со своими затратами и преимущест
наследует свою собственную приспособлен
вами. Идея эволюционной теории игр (EGT)
ность, а +/- мутации [M] добавляются в каждом
может больше подходить для таких симуляций.
поколении (fj = fi + m; m = 0, +/- M). Фактор
Агенты должны взаимодействовать друг с другом
скорости изменения окружающей среды [d] ис
на разных уровнях в непрерывных циклах, об
пользуется для уменьшения приспособленности
следовать различные стратегии, которые влияют
на каждом временном этапе fi(t + 1) = fi(t) - d
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
135
(d ≥ 0 для каждой особи (i)). Вторая модель пос
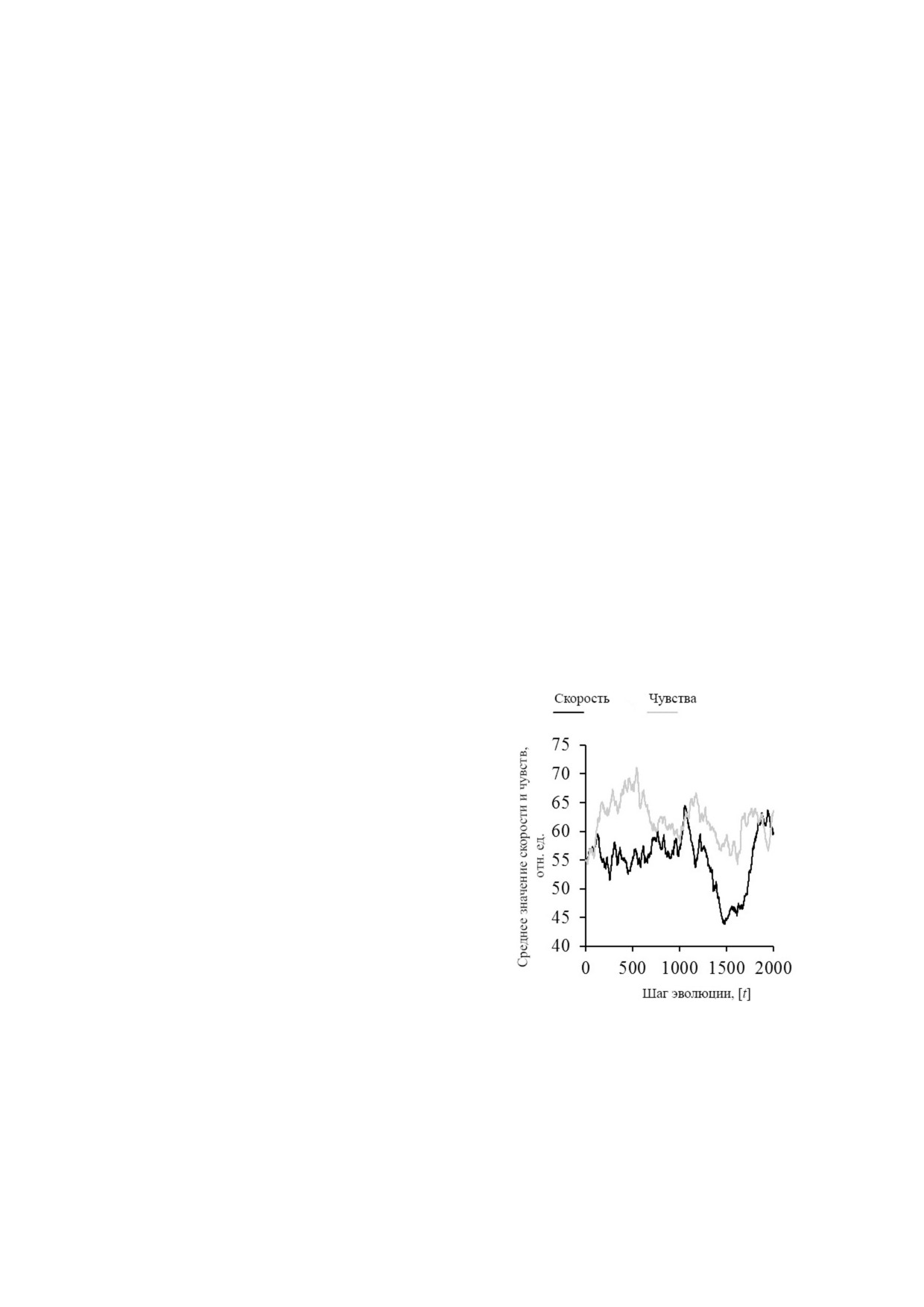
шим выживанием и более успешными потомка
ледовала в 2014 г., в которой параметр d избега
ми, как показано с помощью EGT для развития
ли. Введение случайной смертности видится по
скорости и чувств среднего агента в течение не
лезным, но является ли введение новых правил
скольких поколений (рис. 2). Флуктуация пара
борьбы для новых агентов реалистичной ситуа
метров указывает на постоянное адаптационное
цией, не совсем ясно. Параметр d может быть
давление от окружающих агентов, возникающее
более подходящим для описания экосистемы.
вследствие внутри и межвидовой конкуренции,
В модели отмечено, что угасание приспособлен
которую модель должна учитывать. Поэтому но
ности, вызываемое [d], происходит так быстро,
вые комбинации могут быть полезными для вы
что условия окружающей среды меняются зна
живания в нишах. Такая успешная адаптация
чительно в течение жизни индивидуума [2], но
может установиться, если будет доступно боль
если один гипотетический организм живет при
шее количество пищи, поскольку, по видимому,
мерно в 10 раз дольше, чем другой (как исполь
это способствует установлению субпопуляции
зовано в этих симуляциях; maxLifespan - 5 про
хищников. К сожалению, такие наблюдения
тив 50), соответствующие изменения могут про
трудно подтвердить экспериментально, так как
изойти в течение этого времени. Следует учиты
детальные наблюдения за всеми индивидуаль
вать, что уменьшение приспособленности d
ными агентами в течение нескольких поколе
происходит не так часто в связи с изменениями
ний невозможны. Однако модель Мартин
в ареале обитания во времени. Более вероятная
са 2011 г. с дополнительными подбором значе
ситуация может заключаться в накоплении пос
ний d, вероятным для EGT, оказалась наиболее
ледствий случайных травм у особи (что близко к
ценным предложением по моделированию и ис
идее Вейсмана). Но, скорее всего, основные
пользовалась соответствующим образом.
факторы, влияющие на d, зависят от изменив
Фактор d можно избежать в моделировании,
шихся интересов/стратегий популяции, что яв
основанном на ML&P модели, поскольку дав
ляется следствием EGT. Теория указывает, что
ление адаптации, подобное EGT, уже задано.
если особь приобретает полезный признак, ко
В этом случае средняя жизнь агента составляет
торый позволяет ей использовать новые ресур
примерно два поколения вследствие ограниче
сы или использовать их лучше, чем конкурент,
ний пищи, но агенты могут достичь более дол
такой признак широко распространяется в по
гой продолжительности жизни в модели. Если
пуляции. Это происходит до тех пор, пока борь
агенты с неограниченным и ограниченным мак
ба за ресурсы не становится такой жестокой, что
приобретение нового признака становится бо
лее выгодным. Такой сценарий можно проил
люстрировать на примере золотоискателя, кото
рый нашёл золотую жилу. Другие старатели, уз
нав об этом, тоже собираются в этом месте до
тех пор, пока их не становится слишком много,
и добыча золота в этом месте становится невы
годной, в то время как открытие совершенно
другого бизнеса, например салуна, будет более
выгодно, чем копать золото. Чтобы показать из
менение стратегий в популяции, можно исполь
зовать моделирование эволюции, предложенное
MinuteLabs.io & PRIMER (ML&P). Вопрос со
стоит не в том, подходит ли модель для ответа на
вопрос о развитии старения, и не в том, на
сколько подходящие параметры (такие как вы
сокая скорость изменений в каждом поколении)
и размер модели и количество агентов выбраны.
Важно то, что несколько преимущественных
Рис. 2. Среднее изменение параметров скорости и чувств в
факторов (скорость, размер и чувства) могут из
течение максимального регулируемого жизненного цик
меняться в этой многомерной эволюции в соот
ла [t]. Количество дней - 2000; случайное потомство -
ветствии с изменяющимися затратами. Агенты в
666 666; пища - 50, функция «изменение пищи во време
ни» была деактивирована, использовались только «blue
среднем развивают свои признаки постоянно в
blobs». Следующие параметры были использованы: число
новых комбинациях, поэтому они отличаются
«blobs» вначале - 50; энергия - 500; скорость - 55; раз
от предыдущих признаков старых конкурентов.
мер - 6; чувства - 55; σ2 скорость - 0,5; σ2 размер - 0,5 и
Изменения в большинстве награждаются луч
σ2 чувства - 0,5
БИОХИМИЯ том 87 вып. 1 2022
136
WINTERHALTER, SIMM
симальным числом поколений будут участво
подходящей стратегией. Кроме того, возрастные
вать в межвидовой конкуренции, часто выигры
преимущества в обучении были бы полезны для
вающие стареющие агенты могут указывать на
повышения значимости подхода к моделирова
то, может ли смерть быть выгодной. Тем не менее
нию старения, например, с помощью функции
в этой модели необходимо существенно увели
ln (x), которая вначале является сильной, а со
чить число агентов, чтобы она была подходящей
временем уменьшается.
для исследовательских целей. Кроме того, необ
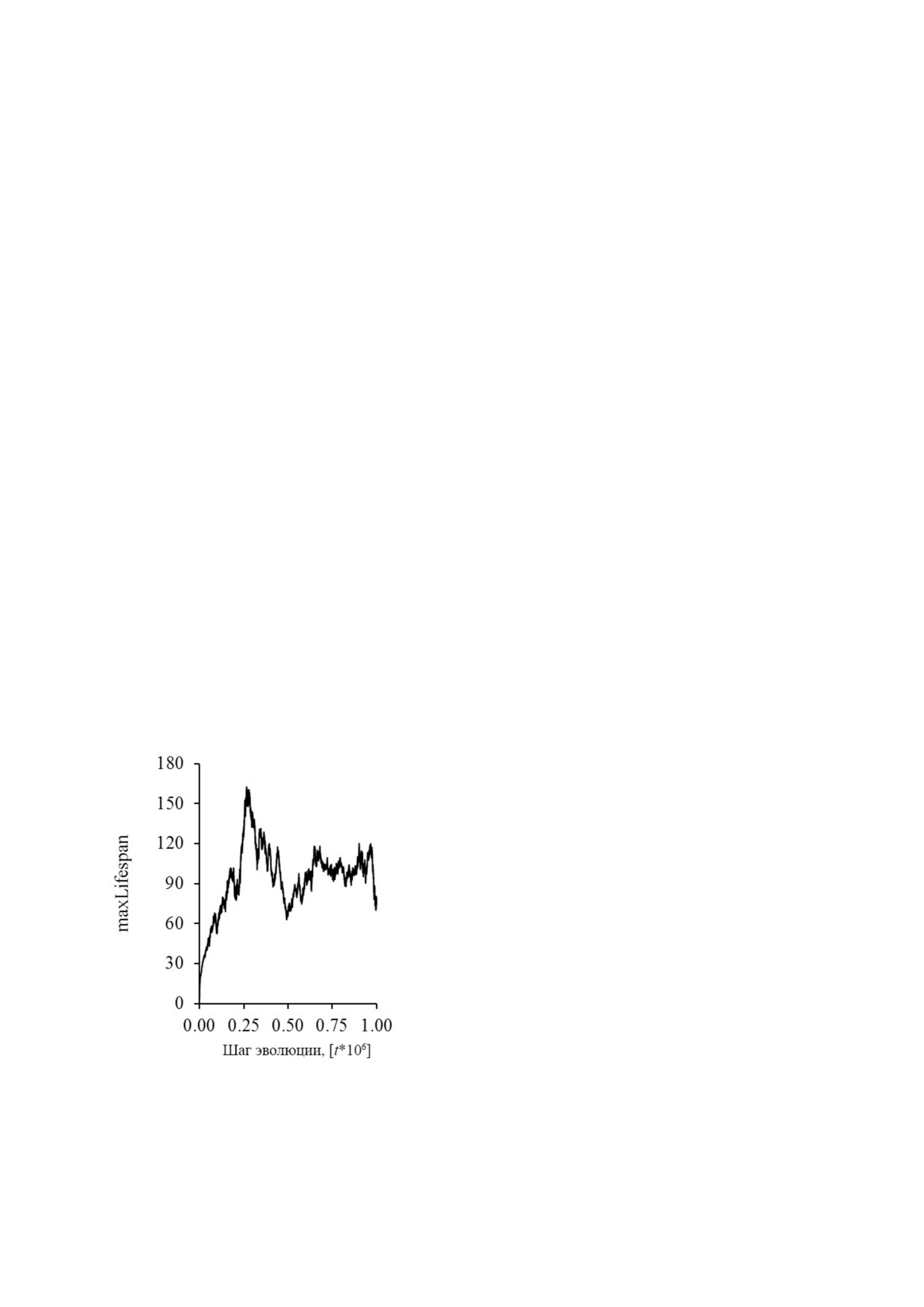
Формирование стабильной максимальной
ходимо добавить более или менее изолирован
продолжительности жизни (maxLifespan) в мо
ные области для изменяющихся временных ин
дели MASON рассматривается, как показатель
тервалов. Дополнительные эволюционные воз
оптимального возраста и, таким образом, явля
можности, такие как изменения окружающей
ется основой возникновения запрограммиро
среды, доступ к пище и её качество или конт
ванного старения. Постоянное увеличение это
роль над паразитами/хищниками, скорее всего,
го параметра указывает на противоположную
не только улучшат предсказательную силу моде
ситуацию [2]. В отличие от оригинальной пуб
ли для старения, но, вероятно, даже определят
ликации, постоянное увеличение maxLifespan
результат моделирования. Кажется, что чем бо
не наблюдалось. Даже для двуполых агентов
лее многомерная симуляция используется, тем
этот параметр осциллировал в широком диапа
более вероятно, что более быстрая эволюция бу
зоне, когда этот параметр отслеживался в тече
дет выгодна.
ние большого числа поколений (рис. 3). Этот
В случае, когда половое размножение доми
факт может противоречить аргументу, что уве
нирует над однополым в модели, уровень разно
личение продолжительности жизни всегда вы
образия, требуемый для такой in silico экосисте
годно, и что это всегда будет происходить, если
мы, может больше соответствовать природному
не появятся некие ограничение, такие, напри
уровню, даже если он невероятно высокий. Па
мер, как повреждения ДНК.
раметры, которые помогают уже взрослым осо
Если агенты выигрывают/проигрывают бо
бям лучше выживать, могут быть более реалис
лее долгоживущим агентам в битвах in silico, это
тичными. Максимальная продолжительность
может предоставить наиболее ценное доказа
жизни в таком случае будет больше зависеть от
тельство преимуществ старения. Это может по
эволюционной приспособленности, чем от та
мочь определить, что является критически важ
ких обстоятельств, как случайное распределе
ным: бóльшая эволюционная приспособлен
ние пищи. Например, введение параметра «вре
ность или большее количество потомков? Если
мя взросления», в течение которого параметры
параметры выбраны подобно тому, как и в ори
молодых агентов заингибированы, может быть
гинальной публикации [2], однополые агенты с
неизменяемой продолжительностью жизни
(epsLifespan - 0) maxLifespan = 5,5 могут выиг
рать (1 из 7 битв; рис. S1, A в Приложении) у
агентов с maxLifespan = 60 в случае, когда нет
изменений в окружающей среде (d = 0), что де
лает развитие старения маловероятным, но не
исключает его. При этом, если двуполые агенты
с неизменяемым maxLifespan = 40 конкурируют
с агентами с maxLifespan = 500 (d = 0), более
быстро стареющая популяция имеет почти та
кие же шансы выиграть (выиграно
11 из
23 битв; рис. S1, B в Приложении). На этом ос
новании можно предположить, что увеличение
maxLifespan выше некого предела не даёт доба
вочных преимуществ в этой модели, что было
бы интересно исследовать, так как ожидается,
что агенты с более длительным сроком жизни
будут постоянно выигрывать, если исключить
биологический износ. Вероятно, относитель
ные эффекты, такие как количество агентов
Рис. 3. Колебания maxLifespan в течение 1 млн поколе
или их распределение, могут иметь больший
ний [t]. Размер мира - 250 × 250; M - 0,03; d - 0,01; началь
эффект, чем продолжительность жизни. Такие
ный maxLifespan - 3,0; epsLifespan - 0,2; b - 1. Такие пара
метры, как потомство 869 413 082 и двуполые агенты, были
случайности могли бы сделать модель не совсем
использованы
подходящей для предсказания преимуществ
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
137
старения, и начальные условия могут значитель
то, как отмечалось ранее, должна присутствовать
но влиять на конечный результат. При этом нет
по крайней мере такая степень адаптации, чтобы
шансов, что двуполые агенты с коротким
имитировать естественную систему in silico. Что
maxLifespan = 5,5 могут выиграть у агентов с
бы определить эти параметры, бесполые агенты
maxLifespan = 60 (d = 0). Короткоживущие аген
должны были конкурировать с половыми в мо
ты с трудом получают возможность достичь
дели Мартинса 2011 г. Бесполые агенты должны
∼250 поколений (рис. S1, C в Приложении) до
были иметь преимущество более быстрого экс
вымирания, в то время как вскоре после начала
поненциального размножения (в 2 раза больше
моделирования почти все ячейки оккупированы
агентов на каждое поколение). Половые агенты
агентами с более высоким maxLifespan, и старею
должны были иметь более высокую эволюцион
щие агенты могут превалировать только с боль
ную приспособленность за счёт обмена наследу
шим трудом. Возможно, быстро стареющие аген
емой информации и повысив изменчивость. Ве
ты не имеют шансов накопить эволюционную
роятность выигрыша будет определяться по
приспособленность. В начале симуляции попу
стандартным правилам, но логично предполо
ляция стареющих агентов уменьшается за счёт
жить, что сражение должно произойти дважды,
более быстрой смерти, которую долгоживущим
если бесполое потомство сталкивается с уже ок
агентам просто нужно пережить. Уже уменьшен
купированной территорией. Параметры, под
ная популяция будет иметь меньше генетических
держивающие двуполое размножение, могут по
вариаций, что уменьшает эволюционную при
мочь определить, какие корректировки модели
способляемость, создавая, таким образом, до
будут более соответствовать реальной ситуации.
полнительное препятствие. Разделение популя
Изменения генома могут произойти за счёт
ции может способствовать разрешению такой не
накопления мутаций или комбинированного
удобной ситуации для группы. В природе геогра
обмена, эффект которых суммируется, но ко
фическая изоляция, впервые описанная
нечное преимущество будет синергично во вре
Эрнстом Майем, является наиболее известным
мени. Чтобы изменения смогли закрепиться,
способом разъединения. Поэтому в модель
они должны в какой то степени быть уместны,
должна быть включена опция для разделения по
что, скорее всего, возможно только в предполо
пуляций поровну в течение регулируемого пери
жении относительного улучшения. Наиболее
ода времени, прежде чем им будет разрешено
существенной предлагаемой корректировкой
сражаться. Преимущество более быстрой адапта
модели in silico является то, как эволюционная
ции должно быть более важно в таких условиях,
приспособленность меняется от поколения к
чем начальные условия. Это тоже может умень
поколению. В модели Мартинса такие парамет
шить гетерогенное распределение агента, но, по
ры, как преимущество распространения гена и
видимому, нет никаких причин, по которым две
большее число агентов, возникающих в связи с
популяции не должны развиваться раздельно до
быстрой сменой поколений в течение опреде
того, как они встретятся на общей границе. Это
лённого промежутка времени, введены автома
кажется более реалистичной ситуацией, напри
тически. Мутации и другие возможные сложные
мер, когда исчезли географические барьеры. Для
комбинации генов способствуют общей измен
генерации старения in silico могут потребоваться
чивости и должны быть адаптированы. Поэтому
несколько изолированных островов.
предлагается заменить M на относительную из
Результаты моделирования, по видимому,
менчивость (θ), которая умножается каждое по
зависят от того, как быстро вид должен приспо
коление (fj = fi × v; v = 1, +/- θ), что является
собиться к таким силам, как RQH или в целом
следствием предполагаемой выгоды от смены
к EGT. Необходимость адаптироваться через
поколений за счёт сложных процентов, уже упо
меж и внутривидовую конкуренцию тоже мо
минавшихся в разделах выше. Подобные отно
жет происходить быстрее, чем ожидается, а
сительные модификации, вероятно, будут пра
сложность модели тоже, по видимому, является
вильными для d и epsLifespan (принятое в моде
фактором, который легко недооценить. Чем бо
ли изменение продолжительности жизни в каж
лее детальная модель используется (количество,
дом поколении), и поэтому тоже могут быть ис
качество, распределение ресурсов, хищников,
пользованы.
и т.д.) и чем масштабнее моделирование, тем
Чтобы продвинуть эволюцию, требуется оп
больше вероятность того, что адаптация повлия
ределенная средняя величина приобретённых
ет на популяцию, и большее давление может
преимуществ на каждую смену поколений. Если
потребоваться для преобладания изменений.
эволюционная приспособленность будет увели
Если половое размножение теоретически
чиваться в модели путём прибавления более или
достаточно выгодно для установления в симуля
менее постоянной величины в каждом поколе
циях с довольно высокой скоростью адаптации,
нии, относительная приспособленность в нача
БИОХИМИЯ том 87 вып. 1 2022
138
WINTERHALTER, SIMM
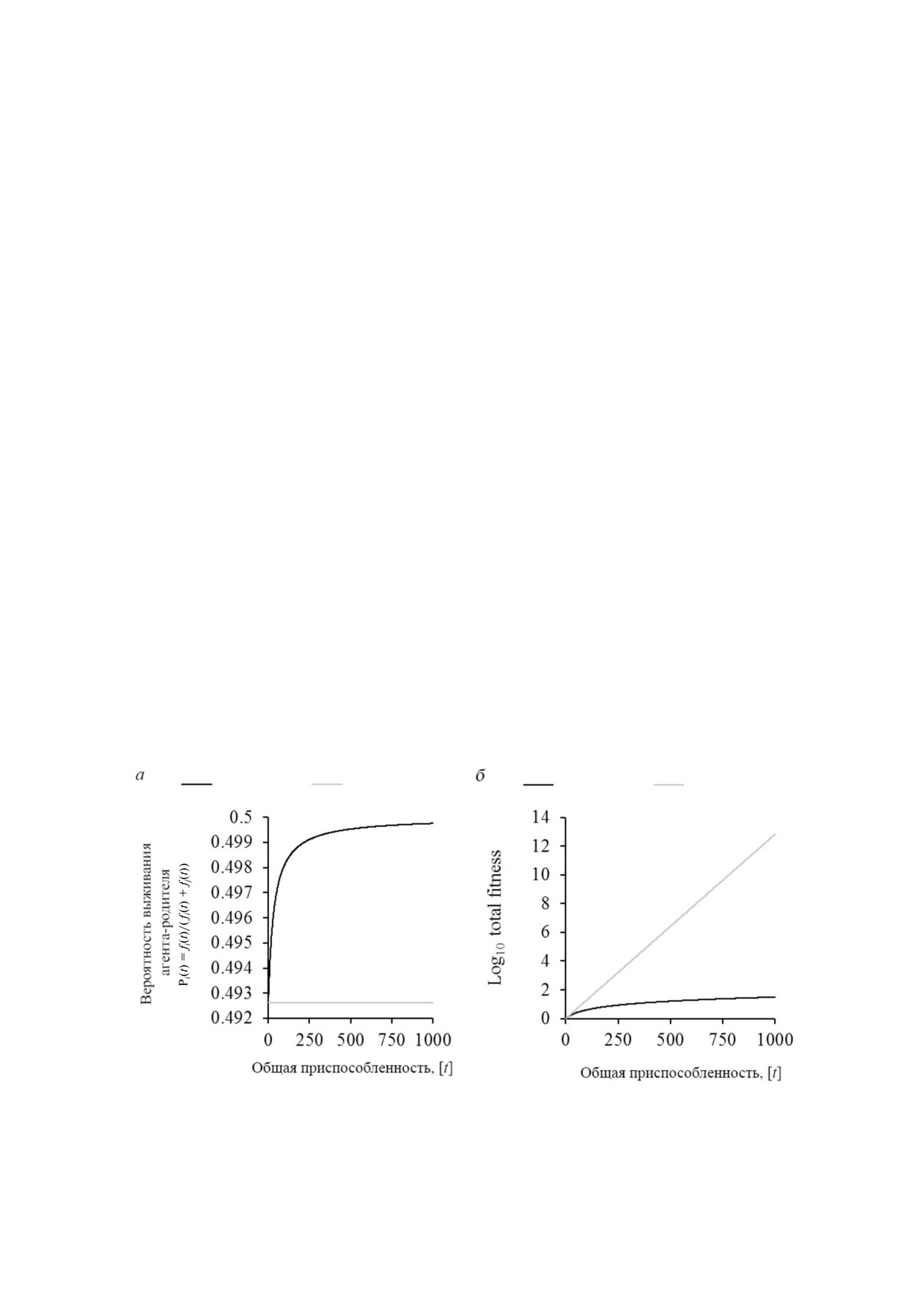
ле была бы высокой. После нескольких поколе
лении вероятность находится на уровне
ний базовая приспособленность группы увели
∼0,492611 (1/(1 + 1,03)), в то время как во втором
чится и достигнет высокого уровня. Дальнейшее
она увеличивается до ∼0,492823 (1,03/(1,03 + 1,06)).
добавление постоянного значения внесёт лишь
Вероятность становится равной (∼0,5), так как
небольшой вклад в общую накопленную спо
преимущество относительной приспособлен
собность. В такой ситуации общая приспособ
ности уменьшается с каждым поколением, в то
ленность потомства не будет сильно отличаться
время как общая приспособленность растёт. Для
от приспособленности родителей. Величина от
сравнения, преимущество остаётся постоянным
носительного преимущества будет несущест
в случае преумножения относительного преиму
венной, так как она приближается все ближе к
щества (рис. 4, а). Приспособленность, следова
нулю с каждым следующим поколением. Эво
тельно, увеличивается экспоненциально благо
люция, основанная только на незначительных
даря предлагаемой адаптации (рис. 4, б). Благо
изменениях, всё же может быть успешна в моде
даря такой впечатляющей экспоненциальной
лях in silico. Вклад случайных эффектов, таких
функции даже малейшие изменения в продол
как количество и распределение агентов, могут
жительности жизни могут привести к сущест
иметь здесь большее значение, чем эволюцион
венному увеличению приспособленности для
ная приспособленность. Почти одинаковый
короткоживущих особей, которое может проя
шанс на успех для агентов с maxLifespan 40 и 500
виться через несколько поколений. Из этого
может быть намёком на такую ситуацию. Если
могла возникнуть высокая выгодность смерти.
больше нет существенных различий в приспо
Было отмечено, что половое размножение
собленности для каждого поколения, случайные
ускоряет достижение наилучших генетических
события, вероятно, решают, кто выживет в при
комбинаций, в то время как старение ускоряет
роде. Отбор будет бессистемным, так как на
распространение выгодных генов и удаление
правление прогресса потеряно; сила, направляю
вредных [44]. К этому утверждению можно до
щая эволюцию, исчезнет, и эволюция остано
бавить, что старение способствует генетичес
вится.
ким выгодам, особенно в случае полового раз
В качестве упрощённого примера без отри
множения.
цательной наследственности используется веро
В случае битвы между агентами, когда
ятность победы родительского агента Pi(t) в мо
maxLifespan зафиксирован, только приспособ
делировании MASON. Эволюция организма на
ленность может меняться. Поэтому в случае по
чинается с уровня приспособленности 1, что яв
лового размножения модель лучше скорректи
ляется обычным начальным условием, который
ровать таким образом, чтобы сделать её более
увеличивается с каждым поколением на поло
естественной, что, вероятно, также усилит пре
жительную величину 0,03 M. В первом поко
имущества спаривания, согласно гипотезе Фи
fj = fi + m
fj = fi *v
fj = fi + m
fj = fi *v
Рис. 4. Сравнение модели MASON для вероятности выживания родительских агентов Pi(t) (а) и общей приспособленнос
ти на поколение [t] (б). Были использованы начальные параметры моделирования Matrins (fj = fi + m; m = 0, +/- M) и пред
ложенные модификации (fj = fi × v; v = 1, +/- θ). Начальная величина была установлена на уровне 1, а коэффициент по
ложительного эффекта - 0,03 для M или θ
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
139
шера-Мюллера. Вероятно, это также уменьшит
(сомой), так как другие функции, такие как дви
требуемую в модели изменчивость до более при
жения и возбудимость, были бы более важными.
емлемого уровня. Вместо одного значения
Действительно, он был первым эволюционис
приспособленности должно быть несколько (по
том, обсуждающим адаптивное и неадаптивное
крайней мере, два). Представляют ли эти значе
старение. Это весьма удивительно, так как изве
ния полезные гены или адаптации (защита, охо
стные отцы неадаптивных теорий Медавара и
та за пищей, умение скрываться, иммунная за
Уильямса несоразмерно критиковали Вейсмана.
щита), является вторичным. Эти значения могут
Для полноты обсуждения необходимо отметить,
случайным образом улучшаться или ухудшаться
что De Grey назвал идею Медавара «предвестни
на некоторую (относительную) величину, при
ком Дж. Б. С. Холдейна» [3]. Медавар и Уильямс
этом общая пригодность определяется средним
просто полагались на ранние идеи Вейсмана об
значением. Таким образом, агент может выиг
адаптивном старении, несмотря на то что Вейс
рать битву, даже если одна из его адапта
ман уже в своих ранних эссе понял, что особь те
ций (умение прятаться) развита не очень хоро
ряет свою ценность для вида после размноже
шо, в то время как другая (защита) была бы до
ния, за некоторыми исключениями, такими как
вольно выгодной. При спаривании, как и при
родительская забота (стр. 9) [23]. Прежде чем
мейозе, каждая величина получена случайным
предложить идею панмиксии, Вейсман уже до
образом от одного из родителей. Таким образом,
пускал эволюционную бесполезность постреп
легко себе представить, что особенно удачные
родуктивной ситуации, когда становится оче
рекомбинации будут активированы, предостав
видным снижение давления естественного от
ляя существенные преимущества агентам, раз
бора на старение. Это может быть критическим
множающимся половым путём. Чем более раз
пунктом в теории старения Медавара, основан
нообразны эти гены или приспособления, тем
ной на накоплении мутаций [61] и родственной
больше будет преимущество полового размно
теории антагонистической плейотропии [62],
жения, и тем более вероятным будут скачки в
предложенной Уильямсом. Вейсман даже пред
эволюционной приспособленности.
полагал, что смертность развилась бы быстрее,
Поправки, предложенные здесь, необходимо
если бы другой выгодный признак (тело) раз
обсудить, и они могут быть очень полезными.
вился бы взамен (стр. 645-646) [63], что связано
Однако считается, что хотя моделирование зас
с основной идеей Уильямса - полезные для эф
тавляет задуматься и обогащает дискуссию об
фективного/раннего воспроизведения гены бу
эволюции старения, эти модели всё ещё слиш
дут отобраны, даже если они могут привести к
ком просты и приблизительны, чтобы ответить
своевременной смерти. В своё время Вейсман
на вопрос об эволюции старения в реальном ми
ничего не знал о генах или мутациях; он утверж
ре. Можно считать, что хотя классические интер
дал, что существует уменьшающееся несовер
претации эволюции старения находятся в рез
шенство в наследовании не очень важных орга
ком контрасте с идеей запрограммированного
нов/функций, и поэтому он практически пред
старения, истина может быть где то посереди
восхитил основную часть теорий Медавара и
не [46], что кажется весьма разумным.
Уильямса, не уточняя геномной причины. Ме
давар тоже упоминает предполагаемую логичес
кую ошибку, которую, по его словам, его студент
МЫСЛИ ВЕЙСМАНА
назвал «порочным кругом». Он писал, что Вейс
О НЕАДАПТИВНОМ СТАРЕНИИ
ман предполагал, что старые особи вида изно
шены и ветхи (в таком же состоянии, происхож
Самый популярный в наше время взгляд на
дение которого он намеревается вывести), а за
неадаптивное старение был описан Вейсманом
тем выдвигал предположение, что из за того,
в его поздних работах. Он считал, что новые
что эти постаревшие особи занимают место здо
формы (как и органы) должны быть не только
ровых, естественный отбор должен привести к
созданы, но и поддержаны отбором. Если нет
истреблению больных и старых. Интересно, что
необходимости поддерживать адаптацию, точ
Медавар предлагал возможный ответ в этой же
ность будет уменьшаться, так как дефекты появ
статье: «Следующая ловушка состоит в том, что
ляются случайным образом и уже не устраняют
популяция предположительно бессмертных жи
ся отбором. Вейсман назвал такое развитие пан
вотных, подверженная опасностям реальной
миксией (стр. 62-63) [25] и предположил, что
смерти, состоит в большинстве своём из очень
функции тоже проходят через этот процесс. По
старых особей и относительно маленького числа
этому, когда природа перестает обращать вни
запуганных молодых, которые путаются между
мание на бессмертие, начинается уничтожение.
ног стариков». Медавар мог сам попасть в такую
Это могло бы случиться с бессмертным телом
логическую ловушку, так как он не верил в то,
БИОХИМИЯ том 87 вып. 1 2022
140
WINTERHALTER, SIMM
что кратковременный процесс отбора достаточ
приспособленным к жизни в течение долгого
но эффективен, чтобы преодолеть преимущест
времени, может быть веками, пока все эти не
ва взрослых или опытных организмов. Это от
большие невосстанавливаемые повреждения не
рицало бы идею порочного круга. Подобно
накопятся и не приведут к смерти. По мнению
Дарвину и Уоллесу, Вейсман был убеждён, что
Вейсмана, если особь занимает свою нишу
селекция происходит очень маленькими шаж
слишком долго, это ускоряет приход смерти.
ками (стр. 47) [64], которые, вероятно, исклю
В среднем смерть предотвращает слишком про
чают предположение, что молодые эволюцио
должительную жизнь. Это важно, потому что
нируют достаточно быстро, чтобы вытеснить
Медавар строит свои аргументы (как в случае с
старых.
порочным кругом) на базе воображаемого орга
Медавар дальше критиковал эту теорию,
низма, который быстро деградирует, а не на ор
приводя статистический пример с пробирками:
ганизме, который ведёт здоровую жизнь долгое
при нормальных условиях объект, который не
время.
уничтожается со временем, будет уничтожен
Тогда зачем Вейсман защищал идею естест
случайно с определённой вероятностью (e-b*x).
венной смерти в своих ранних работах, в то вре
Цель этого была показать, что предположения
мя как он понимал необычность некоторых си
Вейсмана о том, что естественное старение бес
туаций? Для него было настолько само собой
полезно, так как увеличение продолжительнос
разумеющимся, что для процесса отбора важны
ти жизни должно быть непрактичным из за слу
интересы вида, а не индивидуально организма,
чайных стихийных бедствий. Следует отметить,
что он считал это базовой идеей своей лекции, и
что Либертини использовал похожий пример
отказывался говорить об этом, так как считал,
существования определённой вероятности, но
что любой, кто действительно задумывался о
предполагал другое допущение. Он сравнивал
процессе отбора,
должен знать об
гипотетическую смертность, основанную на
этом (стр. 9) [23]. Весьма вероятно, что идеи
случайности, и указал на увеличенные уровня
Вейсмана базировались на отборе более высоко
смертности с возрастом у зебр и буйволов, что
го уровня, что может быть ключевым моментом,
привело его к заключению, что старение может
если в наши дни можно оправдать запрограмми
активно увеличивать уровень смертности [15].
рованное старение.
Тем не менее Вейсман признавал, что смерть -
это действительно случайное событие для каж
дого индивидуума, но поддаётся расчёту для по
ПРОТИВОРЕЧИВОЕ НАСЛЕДИЕ
пуляции, и предполагал, что каждый вид должен
ВЕЙСМАНА
размножаться как можно раньше и как можно
быстрее (стр. 12-13) [23]. Он заявил это ещё до
Теории Вейсмана должны исследоваться в
того, как начал обсуждать свою теорию износа,
широком контексте; в противном случае это, ве
и упомянул бесполезность долгой жизни для не
роятно, приведёт к неполному пониманию его
которых особей, приводя в пример насекомых,
образа мыслей. Вейсман знал, что предложен
так как почти все они довольно рано становятся
ная им теория износа является очень гипотетич
жертвами своих врагов (стр. 24) [23]. Вейсман
ной; он отметил непосредственно в начале свое
показал, что он принял во внимание статисти
го объяснения, что вряд ли можно предполо
ческие доводы Медавара о случайности смерти
жить, что бессмертная особь может избегать все
и никогда не ставил под сомнение тщетность
случайности, разрушающие жизнь, бесконеч
продления жизни во многих случаях. Решающее
но (стр. 30) [23]. В своих работах он обычно при
значение имеют его объяснения возможной це
водит слишком много упрощённых и, соответ
лесообразности сокращения жизни.
ственно, несовершенных примеров, иллюстри
В своей теории износа Вейсман не утвержда
рующих его идеи. В более поздней работе он от
ет, что невозможно идеальное восстановление
ветил на критику своего великого современника
незначительных повреждений; его мысли вра
(Александра Вильгельма Гётте), что он никогда
щались вокруг факта, что полная целостность
не думал о борьбе между бессмертными и смерт
(«in integrum») и идеальное восстановление всех
ными особями, как это иногда предполагается в
потенциальных повреждений вряд ли можно се
наши дни. Вейсман предполагал, что получить
бе представить (стр. 30-31) [23]. Медленное раз
пользу для группы можно уже за счёт сокраще
витие процесса (травма сегодня и через 10 лет)
ния продолжительности жизни (стр. 43) [64].
является ещё одним нюансом и вставками в
В более поздних работах Вейсман отвергал
текст, которые можно воспринять во всей пол
своё прежнее представление о смерти и оправ
ноте только на языке оригинального издания.
дывал её существование необходимым разделе
Это подразумевает, что индивидуум может быть
нием на бессмертный зародыш и высокоспециа
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
141
лизированные клетки тела (сомы) в высших ор
верить в активный отбор, то будет наиболее ра
ганизмах. Он предполагал, что зародышевые
зумно рассматривать этот процесс в контексте
клетки сделали постоянное выживание всего ор
адаптивных теорий, которые подразумевают су
ганизма ненужным и даже непригодным для
ществование позитивного отбора для контроли
сохранения группы (стр. 306-307) [65]. Вейсман
руемого окончания жизни, и это было бы за
уточняет, что тело непригодно для вечной жиз
программированным старением [67]. Аргумен
ни - вероятно, одно из его самым решающим за
ты, свидетельствующие в пользу регулируемого
мечанием. В связи с тем, что он не предоставил
старения и предполагаемые недостатки неадап
никаких объяснений, это утверждение нельзя
тивных теорий, были представлены в других
точно интерпретировать, и контекст не сильно
публикациях [44, 46, 68].
помогает в этом смысле. Бессмертные, но менее
Для организмов, существующих в больших
специализированные клетки могут больше пов
количествах в ограниченных ареалах обитания,
редить эволюционной приспособленности по
таких как лосось или подёнки в реках, смерть
сравнению с высоко оптимизированными, но
после размножения будет полезна для сохране
смертными. В таком случае, почему клетки тка
ния ресурсов или потомства, удобрения среды
ней, в которых метаболизм (функции) наруше
обитания или снижения давления хищник/па
ны, не могут время от времени восстанавливать
разит. Удивительно, что бессмертие или сущест
баланс (регенерироваться)? Этот неразрешён
венное увеличение продолжительности жизни
ный вопрос упомянут в одном из ранних эссе
не развиваются у таких видов, как пчелы и му
Вейсмана (стр. 87) [23]. Однако Вейсман все же
равьи. Увеличение продолжительности жизни
верил в целесообразность смерти, и не было на
наблюдается у репродуктивных организмов, но
мёка в его поздних работах, что он отверг идею о
это увеличение очень далеко от биологического
приоритете интересов видов (групп). В этом слу
бессмертия. Организмы этой высшей касты хо
чае было бы важно определить, как это может
рошо защищены от внешних воздействий, что
мешать его неадаптивным идеям? Тем не менее
существенно уменьшает вероятность смерти,
необходимо отметить, что такое утверждение
вызванной окружающей средой, и у этой груп
могло быть пропущено в работах Вейсмана, ко
пы было достаточно времени (миллионы лет)
торые эклектичны и всеобъемлющи. Кроме того,
для развития. Это может указывать на то, что
он имеет тенденцию вставлять чёткие и ключе
природа не стремится развивать бесконечную
вые утверждения в абсолютно не относящиеся к
продолжительность жизни, так как это до насто
делу скучные разделы, поэтому они иногда оста
ящего времени не было обнаружено, даже если
ются незамеченными.
это кажется целесообразным и осуществимым.
ОБОСНОВАНЫ ЛИ СПОРЫ
ВОЗМОЖНЫЕ МЕХАНИЗМЫ
О НЕАДАПТИВНЫХ ИДЕЯХ?
АКТИВНОГО СТАРЕНИЯ
Почему споры об адаптивной или неадап
Пренебрежение давлением отбора было од
тивной теории старения всё время возвращают
ной из важных причин отказа от идеи запрограм
ся? Может быть, потому, что известны оба об
мированного старения. Новое понимание того,
стоятельства: когда природа стремится отобрать
что старение широко распространено в природе,
полезные для сохранения признаки, а также,
уменьшило значимость одного из традиционных
когда игнорирует несущественные признаки.
аргументов (отсутствие давления отбора) против
Уменьшенная способность к сохранению кле
запрограммированного старения [2], и эту тему,
ток может быть поддержана компонентами за
возможно, следует пересмотреть.
программированного старения, что может ука
Любое сокращение жизни, основанное на
зывать на то, что эти две теории не должны быть
неких дефектах, должен привести к косвенному
взаимоисключающими [66]. Можно себе пред
отбору, если это выгодно. Так как пассивный от
ставить и промежуточную ситуацию, когда не
бор и возможные способы влияния на него огра
адаптивные недостатки приводят к выгодам
ничены, мы сосредоточимся на активном пути
смены поколений, и поэтому отбираются.
старения. В связи с этим необходимо дать точ
Ответы на три фундаментальные вопроса
ное определение регулируемому/запрограмми
определяют наше мнение о смерти. Существуют
рованному старению. Либертини предложил,
ли выгоды от смерти? Если да, то может ли при
что это и есть активная форма старения, харак
рода осуществлять надындивидуальный отбор?
теризующаяся увеличением смертности с увели
Если да, то выбирает ли природа косвенно или
чением хронологического возраста в популяци
посредством контролируемого процесса? Если
ях в дикой природе [15].
БИОХИМИЯ том 87 вып. 1 2022
142
WINTERHALTER, SIMM
Скулачёв считает, что старение способствует
диментарными и активироваться только в опре
прогрессивной эволюции полового размноже
делённых обстоятельствах (у видов, которым
ния [16], предполагая старый эволюционный
присуща семелпария), где это кажется выгод
путь. Вейсман считал, что смерть была изобре
ным. Если бы был возможен активный или кос
тена в многоклеточных организмах (стр. 87)
венный возрастной отбор, они, вероятно, также
[23], тоже склоняясь к раннему эволюционному
взаимодействовали бы друг с другом.
пути развития. Выглядит вполне логично, что,
если запрограммированное старение существу
ет, оно должно было развиться в первичной
ЦИРКАДНЫЕ РИТМЫ
жизни. Если идея смерти способствует эволю
И СВЯЗЬ С ИЗМЕНЕНИЕМ
ционным изменениям за счёт быстрой смены
ПРОДОЛЖИТЕЛЬНОСТИ ЖИЗНИ
поколений, это может быть выгодным и для од
ноклеточных организмов, что указывает на ещё
Периодическая смена дня и ночи существует
более древний механизм.
с тех пор, как возникла Земля, и, вероятно, яв
Вычисления должны быть включены в прог
ляется самым стойким счётчиком времени для
рамму старения, и Голдсмит считал, что, если
биологических организмов. Из этого следует,
старение преднамеренно вызвано биологичес
что циркадные ритмы (ЦР) имели возможность
кой программой, она, вероятно, должна вклю
возникнуть на ранних этапах эволюции. Следо
чать часовой механизм [69]. Оловников предста
вательно, это может быть наиболее подходящим
вил теоретический концепт, в котором незави
фактором старения. Отсутствие таких раздражи
симый астрономический биохронометр регули
телей могло сказаться на продолжительности
рует старение [70]. Точные количества могут то
жизни организмов, живущих в пещерах или на
же определяться с помощью хронологическо
глубоководье. Например, европейский про
го/астрономического времени или по репродук
тей (Proteus anguinus), который весит около
ционному номеру. Оба сигнала могут использо
15-20 г, может дожить примерно до 100 лет [73],
ваться одноклеточными организмами. Счёт вре
возраст которого, вероятно, также достигает ат
мени с определённой гибкостью может быть ос
лантический большеголов (Hoplostethus atlanti
новным путём в сложной жизни, в которой за
cus)
[74], а Гренландская полярная акула
родышевые клетки и клетки тела разделены. Это
(Somniosus microcephalus) в настоящее время счи
не исключает начальную активацию программы
тается самым долгоживущим позвоночным
старения, которая, скорее всего, происходит че
(продолжительность жизни 392 ± 120 лет) [75].
рез половое взросление многоклеточного орга
Голдсмит отмечал, что ЦР и сезоны спарива
низма. Такая ситуации, скорее всего, имеет мес
ния являются самыми полезными биологичес
то у Hydra [71] и может служить начальной точ
кими часами [76], хотя он, скорее всего, не рас
кой для идентификации метаболических путей,
сматривал молекулярные основы этих явлений,
связанных с возрастом.
как драйверов старения, не смотря на то что уже
Воображаемый путь старения, скорее всего,
существовали доказательства ключевой роли ЦР
тесно переплетается с ключевыми процессами
в продолжительности жизни и клеточном старе
развития. В противном случае его можно легко
нии у живых организмаов, а также другие воз
деактивировать. В связи с этим Хмелевски упо
можные варианты продления жизни [77-80].
мянул, что возможность прекращения старения
Была замечена двунаправленная связь между
посредством мутаций, которые ингибируют ге
старением и генами биологических часов: ЦР
нетическую программу, могло бы иметь негатив
гены не только изменялись в процессе старения,
ные, или даже иметь полулетальные, или смер
более того, наблюдалось обратное, когда мута
тельные последствия в дикой природе; он пере
ции модифицировали фенотип старения у пло
числил более слабые модельные организмы, си
довой мушки и у мышей [81]. Также упомина
дящие на диете, а также тяжёлое ингибирование
лась, что существуют связи между ЦР и окисли
передачи сигналов TOR и инсулина/IGF 1 или
тельным стрессом [82]. Но только недавняя
проблемное эпигенетическое репрограммиро
статья осветила ассоциации с запрограммиро
вание [72]. Из этого Хмелевски сделал вывод,
ванным старением [83], были описаны корреля
что такие часто игнорируемые эффекты могут
ции с GSK3B1, влияющими на NRF2. В этой ра
быть причиной ограниченной способности от
боте особое внимание уделялось антиоксидант
бора устранять запрограммированное старение,
ному клеточному ответу, опосредованному
что тоже предполагает связь с летальными пос
NRF2, поскольку исходно повышенные уровни
ледствиями.
NRF2 (и сниженный KEAP1) коррелируют с по
Тем не менее старение или метаболические
вышенными уровнями антиоксидантных белков
пути, инициирующие старение, могут быть ру
(HMOX1, GSTA1 и NQO1) и, по видимому, с
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
143
увеличением продолжительности жизни в пре
ются наиболее точным инструментом для опре
делах семейства грызунов [84]. Этот процесс мо
деления биологического возраста. Таким обра
жет непосредственно регулироваться белка
зом, примечательно, что некоторые признаки
ми ЦР, например, BMAL1 (через NRF2), о чём
эпигенетического старения/долголетия, кажется,
свидетельствует снижение транскрипции
сильно вовлечены в ЦР, например, осциллирую
Hmox1, Gsr и Nqo1 в макрофагах [85]. Однако,
щие модификации цитозина перекрываются с
по видимому, существует петля обратной связи,
возрастными эпигенетическими изменениями
в которой NRF2 регулирует ЦР [86]. В обзорной
ДНК [96]. Кроме того, гиперметилирование и
статье также упомянуты другие механизмы регу
ацетилирование гистонов хорошо коррелируют
ляции изменения NRF2, зависящие от цикли
и часто приводят к эпигенетической транскрип
ческих осцилляций, такие как фосфорилирова
ционной отмене, что оказывает влияние на та
ние, ацетилирование и протеасомная деграда
кие гены, как супрессоры опухолей. Интересно,
ция. Кроме того, изофермент GSK3B1 и его
что изменения в ацетилировании белков, как и в
многочисленные регуляторные возможности,
гистонах, происходят с возрастом и в результате
скорее всего, связанные с NRF2, по видимому,
ограничения калорийности [93], что удивитель
являются центром возрастных изменений. Тем
но, так как ЦР гены (такие как Clock) взаимо
не менее существуют и другие указания на то,
действуют с гистонацетилтрансферазами, при
что ЦР оказывают влияние и могут даже быть
водя к циркадным ритмам ацетилирования [96].
реальной движущей силой старения, и её следу
Само ацетилирование гистонов тоже вовлечено
ет учитывать.
в старение, следовательно, влияние на продол
Мыши с нокаутом генов Bmal1 или Clock де
жительность жизни, вероятно, зависит от ацетил
монстрируют уменьшенную продолжительность
трансферазы [97].
жизни и фенотипы преждевременного старения,
Вещества, продлевающие жизнь, также тес
такие как катаракта, саркопения, уменьшение
но связаны с ЦР зависимыми генами (напри
подкожного жира, усыхание органов или дерма
мер, низкие дозы лития для некоторых организ
тит [77, 87]. Особенно удивительным является
мов, таких как Caenorhabditis elegans или чело
быстрое развитие клеточного старения у мышей с
век) [98]. Вероятно, это работает через ингиби
нокаутом гена Bmal1 [78], что позволяет предпо
рование белка GSK3B [99], которое связано об
ложить существование сигнального пути, кото
ратной связью с ЦР [100]. Изоформа GSK3B
рый изменяет путь старения не только косвенно,
стабилизирует NR1D1 (REV ERBA), в то время
как можно было бы предположить из за повы
как литий вызывает протеасомную деградацию
шенного уровня стресса при нарушенного цикла
и активирует Bmal1 [101]. Литий влияет также на
день/ночь. Например, эффекты изменения
некоторые другие ЦР гены, такие как Per2, Cry1
жизнь у Drosophila хорошо описаны для петли от
и Cry2 [102]. Вальпроевая кислота увеличивает
рицательной обратной связи центральных гете
продолжительность жизни C. elegans [103], изме
родимеров ЦР, когда делеции Per или Tim обеспе
няет экспрессию Per1, Per2, Cry1, Cry2, Bmal1 и
чивали продление жизни, и противоположное
Nr1d1 [104, 105]. Удивительным является тот
происходит, если отсутствовали Clock или
факт, что литий и вальпроевая кислота эффек
Cycle [88]. Кроме того, у мух в диапаузе наблюда
тивно способствуют репрограммированию кле
ется подавление пути ЦР [89]. Все факты, взятые
ток, усиливая экспрессию плюрипотентных ге
вместе, могут свидетельствовать от том, что ЦР
нов [106], что может быть следствием измене
путь сильно влияет, по крайней мере на старение.
ний генов ЦР, также предполагается, что накоп
Другой установленный способ увеличения
ление p53, происходящее с возрастом, может
продолжительности жизни - это ограничение
быть уменьшено литием [107] и что вальпроевая
калорий [90], для которого в 2010 г. были выяв
кислота уменьшает стресс, вызванный старени
лены зависящие от продолжительности жизни
ем во время репрограммирования [108]. Кроме
ассоциации с генами ЦР, такими как «take
того, оба химических вещества также известны
out» [91]. При сравнении кормления ad libitum
как ингибиторы гистоацетилтрансферазы
(питания, ограниченного по времени) и голода
(HDAC), с синергетическими эффектами, а так
ния было показано, что ограничение калорий
же взаимодействуют с несколькими ингибито
регулирует экспрессию таких ЦР генов, как
рами HDAC, такими как трихостатин А (TSA)
Per1, Per2, Cry2, Cry1, Bmal1 и Clock [92]. Ограни
[109, 110]. ТSА, видимо, необходим для эффек
чение калорий также восстанавливает геномные
тивного клонирования мышей (>25 поколе
признаки старения в печёночном транскрипто
ний) [111], что является важной информацией,
ме путём репрограммирования ЦР [93].
так как ингибирование HDAC запускает меха
Эпигенетические часы - ещё один признак
низм, связанный с теоретической возмож
старения [94, 95], в настоящее время они счита
ностью бессмертия соматических клеток. Воз
БИОХИМИЯ том 87 вып. 1 2022
144
WINTERHALTER, SIMM
можность переноса ядер соматических кле
век. Возможно, поэтому Скулачёв различает два
ток (клонирование) доказывает, что каждая
типа старения - острое старение, как у семел
клетка имеет возможность стать бессмертной,
парных видов, и мягкое старение - у других ви
хотя пути репрограммирования всего организма
дов [16]. Противоположное мнение было выска
ещё не изучены. На основании этого можно
зано De Grey, который считает, что фактически
предположить, что такого рода ассоциации со
контрпродуктивно вообще рассматривать се
старением могут быть каким то образом выгод
мелпарность как тип старения [3]. Однако это
ны. В противном случае тело будет способно к
явление наблюдается у широкого круга различ
полной регенерации, если это будет выгодно для
ных существ. Так как регуляция этого процесса
эволюционной приспособленности, как это
не может быть полностью исключена, возника
наблюдается для Hydra [71]. ЦР или затронутые
ет вопрос, присутствуют ли аналогичные про
мишени, такие как HDAC, по видимому, связа
цессы в генетике всех живых существ или не
ны с омоложением.
представляет ли семелпарность совершенно
Другие корреляции тоже стоит отметить. Су
другой тип старения.
ществует наиболее релевантный против старе
ния препарат, рапамицин [112], и детально опи
В заключение можно сказать, что если актив
санные гены долголетия Foxo3 [113] и Ampk [114,
ная регуляция продолжительности жизни суще
115], которые модулируют циркадные ритмы.
ствует, то ЦР могут считаться ключевым регуля
Карликовые мыши известны своим долголети
тором каскада, вызывающего старение, может
ем и экстремальным сокращением клеток пе
быть также регулируемого мозгом (супрахиазма
редней доли гипофиза. При этом сильно снижа
тическое ядро), опосредованно через гормоны.
ется секреция гормона роста, пролактина и ти
Такие заключения слегка отличаются от предло
реотропного гормона [116]. Эти гормоны изве
женного группой Шиловского, которые указы
стны своей зависимостью от ритма день/ночь и
вают, что центральный часовой механизм влияет
вкладом в старение. Другой гормон, изменяю
и на ЦР, и на GSK3B, приводя к увеличению ак
щий дневной цикл - это кортизол, содержание
тивных форм кислорода при старении, опосре
которого можно уменьшить с помощью метфор
дованном NRF2. Можно себе представить ситу
мина - вещества, улучшающего здоровье и про
ацию, когда ЦР сами являются таким централь
должительность жизни [117]. Перепроизводство
ными координаторами, потому что эффекты
кортизола является центральным звеном инду
удаления в рамках механизма обратной связи
цированной смерти у видов, которым присуща
полностью противоположны. Чёткий механизм
семелпария, таких как лососёвые и сумча
обратной связи всегда обеспечивает достижи
тые [118]. Кроме того, хорошо известно, что
мую цель для регулирующего вмешательства.
кортизол снижает активность тимуса и некото
При этом NRF2 и его пути будут одной из мно
рых важных клеток, связанных с иммунной сис
гих мишеней, а поскольку GSK3B колеблется в
темой [119]. У сумчатых основной причиной
течение дня в обратной связи с такими белками,
смертности, по видимому, является недостаточ
как BMAL1, его можно отнести ко всем другим
ность иммунной и противовоспалительной сис
более или менее важным белками ЦР.
тем [5]. У итеропарных позвоночных процесс,
По всей вероятности, окислительный стресс
видимо, очень похож, но происходит медлен
в клетках, опосредованный, скорее всего, мета
нее, причём старение иммунной системы тоже
болическими путями, связанными с глутатио
взаимосвязано с развитием слабого, но увеличи
ном, оказывает большое влияние на старение.
вающегося хронического воспаления (воспали
Однако не трудно себе представить, что ЦР су
тельное старение), в котором такие стрессоры,
щественно влияют по меньшей мере на столь же
как кортизол, играют решающую роль [120].
важные биологические функции, как воспале
Кроме того, другие гормональные измене
ние, контроль качества белка или протеасомную
ния, зависящие от ЦР, могут тоже влиять на из
деградацию, а также на (энергетические) мета
менение продолжительности жизни [121, 122].
болические пути. Кроме того, нельзя исключить
В целом высокая степень пострепродуктивного
существование не обнаруженного или не уста
старения, по видимому, предопределена у се
новленного пути изменения старения, связанно
мелпарных видов [6, 123], и старение, видимо,
го с влиянием ЦР, который может включать ге
происходит схожим образом в некоторых видах.
ны, непосредственно связанные с апоптозом,
Усыхание тимуса с возрастом и старение иммун
контролем повреждений ДНК и активацией ста
ной системы являются примерами, наблюдае
рения. По мнению авторов, старение ещё недос
мыми у сезонных млекопитающих [5, 124] и
таточно изучено, и такие возможности нельзя
рыб [125], а также у организмов с множествен
исключать. Например, недостаточно данных,
ными циклами размножения, таких как чело
объясняющих, как внутриклеточная медь может
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
145
модулировать долголетие [126] - связано ли это
Как бы то ни было, у исследований может
со стрессом, вызванным эффектом ионов тяжё
быть дополнительная многообещающая цель в
лого металла, или вовлечены другие механизмы?
возможных подходах к предотвращению старе
Инициацию программы старения можно се
ния. В результате это может помочь выяснить,
бе представить как механизм изменения ЦР,
является ли старение адаптивным процессом
при которых модификации цитозина во время
или нет.
ежедневных эпигенетических осцилляций не
полностью удаляются и накапливаются после
довательными нарастающими шагами, что при
Финансирование. Работа выполнена при под
водит к изменению транскрипции, вызывая
держке Deutsche Forschungsgemeinschaft (DFG)
медленно нарастающие нарушения клеточных
в учебно исследовательской группе
2155
функций. Другой весьма возможный механизм
(ProMoAge).
может включать изменения в модификации бел
Благодарности. Авторы благодарны Claudia
ков (например, способность к деацетилирова
Rubie (Universitätsklinikum des Saarlandes und
нию) во времени. Например, изменения в аце
Medizinische Fakultät der Universität des
тилировании гистонов могут препятствовать
Saarlandes) за поддержку предположения воз
транскрипции более конденсированными
можности регулирования старения во время
структурами ДНК. Дальнейшие модификации,
персонального разговора на торжественном
которые изменяются в течение жизни, также
обеде во время симпозиума в Heart Centre
возможны и могут взаимодействовать различ
University Hospital of Halle (Saale) в сентябре
ными способами, а механизмы, связанные с
2019 года.
эпигенетическими факторами, могут дополнять
Авторы также благодарны Biodiversity
друг друга. Следовательно, старение может быть
Heritage Library и University of Toronto (Gerstein
вызвано модификациями ДНК и белков, проис
Science Information Centre) и сотрудникам Уни
ходящими в соответствии с циркадными ритма
верситетской библиотеки Freiburg i. Br. за пре
ми. Контролируется ли такая дегенерация или
доставление доступа к работам Вейсмана.
восстановление умышленно предотвращается,
Авторы также благодарят авторов модели
ещё надо выяснить, что, по видимому, не прос
старения MASON за предоставление доступа и
то сделать. Эту идею легко отнести к гипотезам
за удобство пользования [2].
о разрушении сети, поскольку аккультурация
Вклад авторов. PRW был ответственен за
неадаптивных несовершенств, таких как липо
приготовление манускрипта. AS внёс исправле
фусцин, с течением времени также может быть
ния в манускрипт и предоставил доступ к лите
возможным развитием и может привести к дис
ратуре.
регуляции сети ЦР, которая, таким образом, яв
Конфликт интересов. Авторы заявляют об от
ляется лишь побочным продуктом старения.
сутствии конфликта интересов.
В заключение остановимся на вопросе Вейс
Соблюдение этических норм. Настоящая
мана о том, почему соматические клетки не спо
статья не содержит описания каких либо иссле
собны к регенерации время от времени, как это
дований с участием людей или животных в каче
происходит на ранних стадиях развития, соглас
стве объектов.
но современным концепциям. Кроме уже упо
Дополнительные материалы. Приложение к
мянутых возможностей, надо обратить внима
статье на английском языке опубликовано на
ние на ингибирование HDAC, которое, вероят
сайте журнала «Biochemistry» (Moscow) (http://
но, связано с омоложением, что может послу
protein.bio.msu.ru/biokhimiya/) и на сайте изда
жить многообещающим направлением исследо
ваний поведения генов ЦР и модификаций
10541), том 87, вып. 1, 2022.
ДНК/белков во время терапии. Было бы инте
Дополнительная информация. Использован
ресно узнать, являются ли ЦР движущей силой в
ные программы: MinuteLabs.io & PRIMER
накоплении изменений в биологических струк
турах (таких как агрегация ДНК или белков), и
simulator. Код программирования доступен на
какие могут быть последствия этого.
СПИСОК ЛИТЕРАТУРЫ
1.
Cohen, A. A., Kennedy, B. K., Anglas, U., Bronikowski,
2.
Kowald, A., and Kirkwood, T. B. L. (2016) Can aging be
A. M., Deelen, J., et al. (2020) Lack of consensus on an aging
programmed? A critical literature review, Aging Cell, 15,
biology paradigm? A global survey reveals an agreement to dis
986 998, doi: 10.1111/acel.12510.
agree, and the need for an interdisciplinary framework, Mech.
3.
De Grey, A. D. (2007) Calorie restriction, post reproduc
Ageing Dev., 191, 111316, doi: 10.1016/j.mad.2020.111316.
tive life span, and programmed aging: a plea for rigor, Ann.
10 БИОХИМИЯ том 87 вып. 1 2022
146
WINTERHALTER, SIMM
N. Y. Acad. Sci., 1119, 296 305, doi:
10.1196/annals.
23.
Weismann, A. (1882) Ueber die Dauer des Lebens; ein
1404.029.
Vortrag, G. Fischer, Jena.
4.
Hughes, P. W. (2017) Between semelparity and iteroparity:
24.
Weismann, A. (1891) Amphimixis; oder: die Vermischung der
empirical evidence for a continuum of modes of parity,
Individuen, G. Fischer, Jena.
Ecol. Evol., 7, 8232 8261, doi: 10.1002/ece3.3341.
25.
Weismann, A.
(1886) Die Bedeutung der sexuellen
5.
Oakwood, M., Bradley, A. J., and Cockburn, A. (2001)
Fortpflanzung für die Selektions Theorie, G. Fischer, Jena.
Semelparity in a large marsupial, Proc. Biol. Sci., 268, 407
26.
Smith, J. M. (1976) Group selection, Quarterly Rev. Biol.,
411, doi: 10.1098/rspb.2000.1369.
51, 277 283.
6.
Kraaijeveld, K., Kraaijeveld Smit, F. J., and Adcock, G. J.
27.
Wilson, D. S. (1975) A theory of group selection, Proc. Natl.
(2003) Does female mortality drive male semelparity in
Acad. Sci. USA, 72, 143 146, doi: 10.1073/pnas.72.1.143.
dasyurid marsupials? Proc. Biol. Sci., 270 Suppl 2, S251
28.
Hamilton, W. D. (1964) The genetical evolution of social
253, doi: 10.1098/rsbl.2003.0082.
behaviour. I, J. Theor. Biol., 7, 1 16, doi: 10.1016/0022
7.
Leopold, A. C. (1961) Senescence in plant development:
5193(64)90038 4.
the death of plants or plant parts may be of positive ecolog
29.
Nowak, M. A., Tarnita, C. E., and Wilson, E. O. (2010)
ical or physiological value, Science, 134, 17271732,
The evolution of eusociality, Nature, 466, 1057 1062,
doi: 10.1126/science.134.3492.1727.
doi: 10.1038/nature09205.
8.
Dawkins, R. (2003) The Evolution of Evolvability, in On
30.
Briolat, E. S., Burdfield Steel, E. R., Paul, S. C., Ronka,
Growth, Form and Computers (Kumar, S., and Bentley,
K. H., Seymoure, B. M., et al. (2019) Diversity in warning
P. J., eds) Academic Press, pp. 239 255, doi: 10.1016/
coloration: Selective paradox or the norm? Biol. Rev.
B978 012428765 5/50046 3.
Camb. Philos. Soc., 94, 388 414, doi: 10.1111/brv.12460.
9.
Kirschner, M., and Gerhart, J. (1998) Evolvability, Proc.
31.
Kadri, S., Metcalfe, N., Huntingford, F., and Thorpe, J. E.
Natl. Acad. Sci. USA, 95, 8420 8427, doi: 10.1073/pnas.95.
(1995) What controls the onset of anorexia in maturing
15.8420.
adult female atlantic salmon, Funct. Ecol., 9, 790 797.
10.
Wagner, G. P., and Altenberg, L. (1996) Perspective:
32.
Naeger, N. L., Peso, M., Even, N., Barron, A. B., and
Complex adaptations and the evolution of evolvability,
Robinson, G. E. (2013) Altruistic behavior by egg laying
Evolution, 50, 967 976, doi: 10.1111/j.1558 5646.1996.
worker honeybees, Curr. Biol.,
23,
15741578,
tb02339.x.
doi: 10.1016/j.cub.2013.06.045.
11.
Brakefield, P. M. (2006) Evo-devo and constraints on
33.
Carter, G. G., Farine, D. R., Crisp, R. J., Vrtilek, J. K.,
selection, Trends Ecol. Evol., 21, 362 368, doi: 10.1016/
Ripperger, S. P., et al. (2020) Development of new food
j.tree.2006.05.001.
sharing relationships in vampire bats, Curr. Biol., 30, 1275
12.
Landry, C. R., Lemos, B., Rifkin, S. A., Dickinson, W. J.,
1279.e1273, doi: 10.1016/j.cub.2020.01.055.
and Hartl, D. L. (2007) Genetic properties influencing the
34.
Roberts, J. D., Bebenek, K., and Kunkel, T. A. (1988)
evolvability of gene expression, Science, 317, 118 121,
The accuracy of reverse transcriptase from HIV 1, Science,
doi: 10.1126/science.1140247.
242, 1171 1173, doi: 10.1126/science.2460925.
13.
Goldsmith, T. C. (2004) Aging as an evolved character
35.
Weismann, A. (1887) Ueber die Zahl der Richtungskörper
istic - Weismann’s theory reconsidered, Med. Hypotheses,
und über ihre Bedeutung für die Vererbung, G. Fischer, Jena.
62, 304 308, doi: 10.1016/S0306 9877(03)00337 2.
36.
Bi, K., and Bogart, J. P. (2010) Time and time again: uni
14.
Goldsmith, T. C. (2008) Aging, evolvability, and the indi
sexual salamanders (genus Ambystoma) are the oldest uni
vidual benefit requirement; medical implications of aging
sexual vertebrates, BMC Evol. Biol., 10, 238, doi: 10.1186/
theory controversies, J. Theor. Biol., 252, 764768,
1471 2148 10 238.
doi: 10.1016/j.jtbi.2008.02.035.
37.
Smith, J. M. (1971) What use is sex? J. Theor. Biol., 30,
15.
Libertini, G. (1988) An adaptive theory of increasing mor
319 335, doi: 10.1016/0022 5193(71)90058 0.
tality with increasing chronological age in populations in
38.
Trivers, R. (1983) The evolution of sex: the masterpiece of
the wild, J. Theor. Biol., 132, 145 162, doi: 10.1016/s0022
nature: the evolution and genetics of sexuality. Graham
5193(88)80153 x.
Bell, Quarterly Rev. Biol., 58, 62 67, doi: 10.1086/413059.
16.
Skulachev, V. P. (1997) Aging is a specific biological
39.
Hamilton, W. D. (1980) Sex versus non sex versus parasite,
function rather than the result of a disorder in complex
Oikos, 35, 282 290, doi: 10.2307/3544435.
living systems: biochemical evidence in support of
40.
Jaenike, J. (1977) A hypothesis to account for the mainte
Weismann’s hypothesis, Biochemistry (Moscow), 62,
nance of sex in populations, Evol. Theory, 3, 191 194.
1191 1195.
41.
Hartung, J. (1981) Genome Parliaments and Sex with the
17.
Skulachev, V. P. (1999) Phenoptosis: programmed death of
Red Queen, in Natural Selection and Social Behavior
an organism, Biochemistry (Moscow), 64, 1418 1426.
(Alexander, R. D., and Tinkle, D., eds.) Chiron, N.Y.,
18.
Skulachev, M. V., and Skulachev, V. P. (2014) New data on
pp. 382 404.
programmed aging - slow phenoptosis, Biochemistry
42.
Lively, C., Craddock, C., and Vrijenhoek, R. (1990) Red
(Moscow), 79, 977 993, doi: 10.1134/S0006297914100010.
Queen hypothesis supported by parasitism in sexual and
19.
Vyssokikh, M. Y., Holtze, S., Averina, O. A., Lyamzaev, K.
clonal fish, Nature, 344, 864 866, doi: 10.1038/344864a0.
G., Panteleeva, A. A., et al. (2020) Mild depolarization of
43.
Quattro, J. M., Avise, J. C., and Vrijenhoek, R. C. (1992)
the inner mitochondrial membrane is a crucial component
An ancient clonal lineage in the fish genus Poeciliopsis
of an anti aging program, Proc. Natl. Acad. Sci. USA, 117,
(Atheriniformes: Poeciliidae), Proc. Natl. Acad. Sci. USA,
6491 6501, doi: 10.1073/pnas.1916414117.
89, 348 352, doi: 10.1073/pnas.89.1.348.
20.
Blackburn, E. H. (2000) Telomere states and cell fates,
44.
Libertini, G. (2017) Sex and Aging: A Comparison between
Nature, 408, 53 56, doi: 10.1038/35040500.
Two Phenoptotic Phenomena, Biochemistry (Moscow), 82,
21.
Libertini, G. (2015) Phylogeny of aging and related
1435 1455, doi: 10.1134/S0006297917120045.
phenoptotic phenomena, Biochemistry (Moscow), 80,
45.
Dobrowolski, P., Fischer, M., and Naumann, R. (2018)
1529 1546, doi: 10.1134/S0006297915120019.
Novel insights into the genetic background of genetically
22.
Libertini, G., Corbi, G., and Nicola, F. (2020) Importance
modified mice, Transgenic Res.,
27,
265275,
and meaning of TERRA sequences for aging mechanisms,
doi: 10.1007/s11248 018 0073 2.
Biochemistry (Moscow), 85, 15051517, doi: 10.1134/
46.
Lenart, P., and Bienertova Vasku, J. (2017) Keeping up
S0006297920120044.
with the Red Queen: the pace of aging as an adaptation,
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
147
Biogerontology, 18, 693 709, doi: 10.1007/s10522 016
of modern progress, Hum. Genet.,
60,
101121,
9674 4.
doi: 10.1007/BF00569695.
47.
Mitteldorf, J., and Pepper, J. (2009) Senescence as an
68.
Libertini, G. (2015) Non programmed versus programmed
adaptation to limit the spread of disease, J. Theor. Biol.,
aging paradigm, Curr. Aging Sci., 8, 56 68.
260, 186 195, doi: 10.1016/j.jtbi.2009.05.013.
69.
Goldsmith, T. C. (2012) On the programmed/non pro
48.
May, R. M., and Anderson, R. M. (1983) Epidemiology
grammed aging controversy, Biochemistry (Moscow), 77,
and genetics in the coevolution of parasites and hosts, Proc.
729 732, doi: 10.1134/S000629791207005X.
R. Soc. Lond. B Biol. Sci., 219, 281 313, doi: 10.1098/
70.
Olovnikov, A. M. (2003) The redusome hypothesis of aging
rspb.1983.0075.
and the control of biological time during individual devel
49.
Howard, R. S., and Lively, C. M. (1994) Parasitism, muta
opment, Biochemistry (Moscow), 68, 2 33, doi: 10.1023/
tion accumulation and the maintenance of sex, Nature,
a:1022185100035.
367, 554 557, doi: 10.1038/367554a0.
71.
Schaible, R., Sussman, M., and Kramer, B. H. (2014)
50.
Otto, S. P., and Nuismer, S. L. (2004) Species interactions
Aging and potential for self renewal: hydra living in the age
and the evolution of sex, Science, 304, 10181020,
of aging - a mini review, Gerontology, 60, 548556,
doi: 10.1126/science.1094072.
doi: 10.1159/000360397.
51.
Lively, C. M. (2010) Parasite virulence, host life history,
72.
Chmielewski, P. (2017) Rethinking modern theories of
and the costs and benefits of sex, Ecology, 91, 3 6,
ageing and their classification: the proximate mechanisms
doi: 10.1890/09 1158.1.
and the ultimate explanations, Anthropol. Rev., 80, 259
52.
Martins, A. C. (2011) Change and aging senescence as an
272, doi: 10.1515/anre 2017 0021.
adaptation, PLoS One, 6, e24328, doi: 10.1371/journal.
73.
Voituron, Y., de Fraipont, M., Issartel, J., Guillaume, O.,
pone.0024328.
and Clobert, J. (2011) Extreme lifespan of the human fish
53.
Mitteldorf, J., and Martins, A. C. (2014) Programmed life
(Proteus anguinus): A challenge for ageing mechanisms,
span in the context of evolvability, Am. Nat., 184, 289 302,
Biol. Lett., 7, 105 107, doi: 10.1098/rsbl.2010.0539.
doi: 10.1086/677387.
74.
Andrews, A. H., Tracey, D. M., and Dunn, M. R. (2009)
54.
Keightley, P. D., Ness, R. W., Halligan, D. L., and
Lead radium dating of orange roughy (Hoplostethus
Haddrill, P. R. (2014) Estimation of the spontaneous
atlanticus): validation of a centenarian life span, Can. J.
mutation rate per nucleotide site in a Drosophila
Fish. Aquat. Sci., 66, 1130 1140, doi: 10.1139/f09 059.
melanogaster full sib family, Genetics, 196, 313320,
75.
Nielsen, J., Hedeholm, R. B., Heinemeier, J., Bushnell,
doi: 10.1534/genetics.113.158758.
P. G., Christiansen, J. S., et al. (2016) Eye lens radiocarbon
55.
Blount, Z. D., Barrick, J. E., Davidson, C. J., and Lenski,
reveals centuries of longevity in the Greenland shark
R. E. (2012) Genomic analysis of a key innovation in an
(Somniosus microcephalus), Science,
353,
702704,
experimental Escherichia coli population, Nature, 489,
doi: 10.1126/science.aaf1703.
513 518, doi: 10.1038/nature11514.
76.
Goldsmith, T. C. (2016) Emerging programmed aging
56.
Cohain, J. S., Buxbaum, R. E., and Mankuta, D. (2017)
mechanisms and their medical implications, Med.
Spontaneous first trimester miscarriage rates per woman
Hypotheses, 86, 92 96, doi: 10.1016/j.mehy.2015.10.015.
among parous women with 1 or more pregnancies of 24
77.
Kondratov, R. V., Kondratova, A. A., Gorbacheva, V. Y.,
weeks or more, BMC Pregnancy Childbirth, 17, 437,
Vykhovanets, O. V., and Antoch, M. P. (2006) Early aging
doi: 10.1186/s12884 017 1620 1.
and age related pathologies in mice deficient in BMAL1,
57.
Keightley, P. D., and Otto, S. P. (2006) Interference among
the core componentof the circadian clock, Genes Dev., 20,
deleterious mutations favours sex and recombination in
1868 1873, doi: 10.1101/gad.1432206.
finite populations, Nature, 443, 89 92, doi: 10.1038/
78.
Khapre, R. V., Kondratova, A. A., Susova, O., and
nature05049.
Kondratov, R. V. (2011) Circadian clock protein BMAL1
58.
Grieshop, K., Maurizio, P. L., Arnqvist, G., and Berger, D.
regulates cellular senescence in vivo, Cell Cycle, 10, 4162
(2021) Selection in males purges the mutation load on
4169, doi: 10.4161/cc.10.23.18381.
female fitness, Evol. Lett., 5, 328 343, doi: 10.1002/
79.
Nakahata, Y., Sahar, S., Astarita, G., Kaluzova, M., and
evl3.239.
Sassone Corsi, P. (2009) Circadian control of the NAD+
59.
Felsenstein, J. (1974) The evolutionary advantage of
salvage pathway by CLOCK SIRT1, Science, 324, 654
recombination, Genetics, 78, 737 756, doi: 10.1093/genetics/
657, doi: 10.1126/science.1170803.
78.2.737.
80.
Bonaconsa, M., Malpeli, G., Montaruli, A.,
60.
Luke, S., Cioffi Revilla, C., Panait, L., Sullivan, K., and
Carandente, F., Grassi Zucconi, G., et al.
(2014)
Balan, G. (2005) MASON: A multiagent simulation envi
Differential modulation of clock gene expression in the
ronment, Simulation,
81,
517527, doi:
10.1177/
suprachiasmatic nucleus, liver and heart of aged mice, Exp.
0037549705058073.
Gerontol., 55, 70 79, doi: 10.1016/j.exger.2014.03.011.
61.
Medawar, P. B. (1952) An Unsolved Problem of Biology: An
81.
Hood, S., and Amir, S. (2017) The aging clock: Circadian
Inaugural Lecture Delivered at University College, London, 6
rhythms and later life, J. Clin. Invest., 127, 437 446,
December, 1951, H. K. Lewis and Company, London.
doi: 10.1172/JCI90328.
62.
Williams, G. C. (1957) Pleiotropy, Natural Selection, and
82.
Vallee, A., Lecarpentier, Y., and Vallee, J. N.
(2019)
the Evolution of Senescence, Evolution, 11, 398 411,
Curcumin: a therapeutic strategy in cancers by inhibiting
doi: 10.2307/2406060.
the canonical WNT/beta catenin pathway, J. Exp. Clin.
63.
Weismann, A.
(1892) Aufsätze über Vererbung und
Cancer Res., 38, 323, doi: 10.1186/s13046 019 1320 y.
Verwandte Biologische Fragen, G. Fischer, Jena.
83.
Shilovsky, G. A., Putyatina, T. S., Morgunova, G. V.,
64.
Weismann, A.
(1884) Ueber Leben und Tod: Eine
Seliverstov, A. V., Ashapkin, V. V., et al. (2021) A Crosstalk
Biologische Untersuchung, G. Fischer, Jena.
between the biorhythms and gatekeepers of longevity: Dual
65.
Weismann, A. (1904) Vorträge über Deszendenztheorie;
role of glycogen synthase kinase 3, Biochemistry (Moscow),
Band 2, G. Fischer, Jena.
86, 433 448, doi: 10.1134/S0006297921040052.
66.
Sergiev, P. V., Dontsova, O. A., and Berezkin, G. V. (2015)
84.
Lewis, K. N., Wason, E., Edrey, Y. H., Kristan, D. M.,
Theories of aging: an ever evolving field, Acta Naturae, 7, 9 18.
Nevo, E., et al. (2015) Regulation of Nrf2 signaling and
67.
Kirkwood, T. B., and Cremer, T. (1982) Cytogerontology
longevity in naturally long lived rodents, Proc. Natl. Acad.
since 1881: a reappraisal of August Weismann and a review
Sci. USA, 112, 3722 3727, doi: 10.1073/pnas.1417566112.
БИОХИМИЯ том 87 вып. 1 2022
10*
148
WINTERHALTER, SIMM
85. Early, J. O., Menon, D., Wyse, C. A., Cervantes Silva,
tive component of the circadian clock, Science, 311, 1002
M. P., Zaslona, Z., et al. (2018) Circadian clock protein
1005, doi: 10.1126/science.1121613.
BMAL1 regulates IL 1beta in macrophages via NRF2,
102. Noguchi, T., Lo, K., Diemer, T., and Welsh, D. K. (2016)
Proc. Natl. Acad. Sci. USA,
115, E8460 E8468,
Lithium effects on circadian rhythms in fibroblasts and
doi: 10.1073/pnas.1800431115.
suprachiasmatic nucleus slices from Cry knockout mice,
86. Wible, R. S., Ramanathan, C., Sutter, C. H., Olesen,
Neurosci. Lett., 619, 49 53, doi: 10.1016/j.neulet.2016.
K. M., Kensler, T. W., et al. (2018) NRF2 regulates core
02.030.
and stabilizing circadian clock loops, coupling redox and
103. Evason, K., Collins, J. J., Huang, C., Hughes, S., and
timekeeping in Mus musculus, Elife,
7, e31656,
Kornfeld, K. (2008) Valproic acid extends Caenorhabditis
doi: 10.7554/eLife.31656.
elegans lifespan, Aging Cell, 7, 305 317, doi: 10.1111/
87. Dubrovsky, Y. V., Samsa, W. E., and Kondratov, R. V.
j.1474 9726.2008.00375.x.
(2010) Deficiency of circadian protein CLOCK reduces
104. Johansson, A. S., Brask, J., Owe Larsson, B., Hetta, J.,
lifespan and increases age related cataract development in
and Lundkvist, G. B. (2011) Valproic acid phase shifts the
mice, Aging (Albany NY), 2, 936 944, doi: 10.18632/aging.
rhythmic expression of Period2::Luciferase, J. Biol.
100241.
Rhythms, 26, 541 551, doi: 10.1177/0748730411419775.
88. Ulgherait, M., Chen, A., McAllister, S. F., Kim, H. X.,
105. Griggs, C. A., Malm, S. W., Jaime Frias, R., and Smith,
Delventhal, R., et al. (2020) Circadian regulation of mito
C. L. (2018) Valproic acid disrupts the oscillatory expression
chondrial uncoupling and lifespan, Nat. Commun., 11,
of core circadian rhythm transcription factors, Toxicol. Appl.
1927, doi: 10.1038/s41467 020 15617 x.
Pharmacol., 339, 110 120, doi: 10.1016/j.taap.2017.12.005.
89. Kucerova, L., Kubrak, O. I., Bengtsson, J. M., Strnad, H.,
106. Tang, Y., and Cheng, L. (2017) Cocktail of chemical com
Nylin, S., et al. (2016) Slowed aging during reproductive
pounds robustly promoting cell reprogramming protects
dormancy is reflected in genome wide transcriptome
liver against acute injury, Protein Cell, 8, 273283,
changes in Drosophila melanogaster, BMC Genomics, 17,
doi: 10.1007/s13238 017 0373 y.
50, doi: 10.1186/s12864 016 2383 1.
107. Zmijewski, J. W., and Jope, R. S. (2004) Nuclear accumu
90. McCay, C. M., Crowell, M. F., and Maynard, L. A. (1935)
lation of glycogen synthase kinase 3 during replicative
The effect of retarded growth upon the length of life span
senescence of human fibroblasts, Aging Cell, 3, 309 317,
and upon the ultimate body size. 1935, Nutrition, 5, 63 79,
doi: 10.1111/j.1474 9728.2004.00117.x.
doi: 10.1093/jn/10.1.63.
108. Zhai, Y., Chen, X., Yu, D., Li, T., Cui, J., et al. (2015)
91. Bauer, J., Antosh, M., Chang, C., Schorl, C., Kolli, S.,
Histone deacetylase inhibitor valproic acid promotes the
et al. (2010) Comparative transcriptional profiling identi
induction of pluripotency in mouse fibroblasts by suppress
fies takeout as a gene that regulates life span, Aging (Albany
ing reprogramming induced senescence stress, Exp. Cell
NY), 2, 298 310, doi: 10.18632/aging.100146.
Res., 337, 61 67, doi: 10.1016/j.yexcr.2015.06.003.
92. Patel, S. A., Velingkaar, N., Makwana, K., Chaudhari, A.,
109. Leng, Y., Liang, M. H., Ren, M., Marinova, Z., Leeds, P.,
and Kondratov, R. (2016) Calorie restriction regulates cir
et al. (2008) Synergistic neuroprotective effects of lithium
cadian clock gene expression through BMAL1 dependent
and valproic acid or other histone deacetylase inhibitors in
and independent mechanisms, Sci. Rep., 6, 25970,
neurons: roles of glycogen synthase kinase 3 inhibition,
doi: 10.1038/srep25970.
J. Neurosci., 28, 2576 2588, doi: 10.1523/JNEUROSCI.
93. Sato, S., Solanas, G., Peixoto, F. O., Bee, L.,
5467 07.2008.
Symeonidi, A., et al. (2017) Circadian reprogramming in
110. Wu, S., Zheng, S. D., Huang, H. L., Yan, L. C., Yin, X. F.,
the liver identifies metabolic pathways of aging, Cell, 170,
et al. (2013) Lithium down regulates histone deacetylase 1
664 677.e11, doi: 10.1016/j.cell.2017.07.042.
(HDAC1) and induces degradation of mutant huntingtin,
94. Schumacher, A. (2009) An Epigenetic Clock: Anticorrelation
J. Biol. Chem., 288, 35500 35510, doi: 10.1074/jbc.M113.
& DNA Methylation as Biomarker for Aging, doi: 10.13140/
479865.
RG.2.2.12457.83042.
111.
Wakayama, S., Kohda, T., Obokata, H., Tokoro, M.,
95. Bocklandt, S., Lin, W., Sehl, M. E., Sanchez, F. J.,
Li, C., et al. (2013) Successful serial recloning in the mouse
Sinsheimer, J. S., et al. (2011) Epigenetic predictor of age,
over multiple generations, Cell Stem Cell, 12, 293 297,
PLoS One, 6, e14821, doi: 10.1371/journal.pone.0014821.
doi: 10.1016/j.stem.2013.01.005.
96. Oh, G., Ebrahimi, S., Carlucci, M., Zhang, A., Nair, A.,
112. Cao, R., Li, A., Cho, H. Y., Lee, B., and Obrietan, K.
et al. (2018) Cytosine modifications exhibit circadian oscil
(2010) Mammalian target of rapamycin signaling modu
lations that are involved in epigenetic diversity and aging,
lates photic entrainment of the suprachiasmatic circadian
Nat. Commun., 9, 644, doi: 10.1038/s41467 018 03073 7.
clock, J. Neurosci., 30, 6302 6314, doi: 10.1523/JNEUROSCI.
97. Yi, S. J., and Kim, K. (2020) New Insights into the role of
5482 09.2010.
histone changes in aging, Int. J. Mol. Sci., 21, 8241,
113. Chaves, I., van der Horst, G. T., Schellevis, R., Nijman,
doi: 10.3390/ijms21218241.
R. M., Koerkamp, M. G., et al. (2014) Insulin FOXO3 sig
98. Zarse, K., Terao, T., Tian, J., Iwata, N., Ishii, N., et al.
naling modulates circadian rhythms via regulation of clock
(2011) Low dose lithium uptake promotes longevity in
transcription, Curr. Biol., 24, 1248 1255, doi: 10.1016/
humans and metazoans, Eur. J. Nutr., 50, 387389,
j.cub.2014.04.018.
doi: 10.1007/s00394 011 0171 x.
114. Lee, Y., and Kim, E. K. (2013) AMP activated protein
99. Castillo Quan, J. I., Li, L., Kinghorn, K. J., Ivanov, D. K.,
kinase as a key molecular link between metabolism and
Tain, L. S., et al. (2016) Lithium promotes longevity
clockwork, Exp. Mol. Med., 45, e33, doi: 10.1038/emm.
through GSK3/NRF2 dependent hormesis, Cell Rep., 15,
2013.65.
638 650, doi: 10.1016/j.celrep.2016.03.041.
115. Lamia, K. A., Sachdeva, U. M., DiTacchio, L., Williams,
100. Besing, R. C., Paul, J. R., Hablitz, L. M., Rogers, C. O.,
E. C., Alvarez, J. G., et al. (2009) AMPK regulates the cir
Johnson, R. L., et al. (2015) Circadian rhythmicity of
cadian clock by cryptochrome phosphorylation and degra
active GSK3 isoforms modulates molecular clock gene
dation, Science, 326, 437440, doi: 10.1126/science.
rhythms in the suprachiasmatic nucleus, J. Biol. Rhythms,
1172156.
30, 155 160, doi: 10.1177/0748730415573167.
116. Brown Borg, H. M., Borg, K. E., Meliska, C. J., and
101. Yin, L., Wang, J., Klein, P. S., and Lazar, M. A. (2006)
Bartke, A. (1996) Dwarf mice and the ageing process,
Nuclear receptor Rev erbalpha is a critical lithium sensi
Nature, 384, 33, doi: 10.1038/384033a0.
БИОХИМИЯ том 87 вып. 1 2022
О ЦЕЛЕСООБРАЗНОСТИ СМЕРТИ
149
117. Martin Montalvo, A., Mercken, E. M., Mitchell, S. J.,
dyscrasia post reproduction dictate post reproductive
Palacios, H. H., Mote, P. L., et al. (2013) Metformin
lifespan? Lessons from semelparous and iteroparous
improves healthspan and lifespan in mice, Nat. Commun.,
species, Geroscience, 39, 103 116, doi: 10.1007/s11357
4, 2192, doi: 10.1038/ncomms3192.
016 9955 5.
118. Stein Behrens, B. A., and Sapolsky, R. M. (1992) Stress,
123. Jeffries, K. M., Hinch, S. G., Donaldson, M. R., Gale,
glucocorticoids, and aging, Aging (Milano), 4, 197 210,
M. K., Burt, J. M., et al. (2011) Temporal changes in blood
doi: 10.1007/BF03324092.
variables during final maturation and senescence in male
119. Dracott, B. N., and Smith, C. E. (1979) Hydrocortisone
sockeye salmon Oncorhynchus nerka: reduced osmoregu
and the antibody response in mice. I. Correlations between
latory ability can predict mortality, J. Fish Biol., 79, 449
serum cortisol levels and cell numbers in thymus, spleen,
465, doi: 10.1111/j.1095 8649.2011.03042.x.
marrow and lymph nodes, Immunology, 38, 429 435.
124. Poskittt, D. C., Barnett, J., Duffey, K., Kimpton, W. G.,
120. Franceschi, C., Bonafe, M., Valensin, S., Olivieri, F., De
and Muller, H. K. (1984) Involution of the thymus in mar
Luca, M., et al. (2000) Inflamm aging. An evolutionary
supial mice, Dev. Comp. Immunol.,
8,
483488,
perspective on immunosenescence, Ann. N. Y. Acad. Sci.,
doi: 10.1016/0145 305x(84)90056 9.
908, 244 254, doi: 10.1111/j.1749 6632.2000.tb06651.x.
125. McQuillan, H. J., Lokman, P. M., and Young, G. (2003)
121. De Jesus, E. G., and Hirano, T. (1992) Changes in whole
Effects of sex steroids, sex, and sexual maturity on cortisol
body concentrations of cortisol, thyroid hormones, and sex
production: An in vitro comparison of chinook salmon and
steroids during early development of the chum salmon,
rainbow trout interrenals, Gen. Comp. Endocrinol., 133,
Oncorhynchus keta, Gen. Comp. Endocrinol., 85, 55 61,
154 163, doi: 10.1016/s0016 6480(03)00163 1.
doi: 10.1016/0016 6480(92)90171 f.
126. Matos, L., Gouveia, A., and Almeida, H. (2012) Copper
122. Atwood, C. S., Hayashi, K., Meethal, S. V., Gonzales, T.,
ability to induce premature senescence in human fibroblasts,
and Bowen, R. L. (2017) Does the degree of endocrine
Age (Dordr), 34, 783 794, doi: 10.1007/s11357 011 9276 7.
HOW JUSTIFIED IS THE ASSUMPTION OF PROGRAMMED AGING
IN REMINISCENCE OF WEISMANN’S THEORIES?
Review
P. R. Winterhalter* and A. Simm
Martin Luther University of Halle Wittenberg, 06120 Halle (Saale), Germany; E mail: winterhalter.patrick@gmail.com
Theories about the benefits of death and the resulting increased likelihood of programmed aging are controversial,
advocated only by a minority. The extent to which their assumptions might be justified should be investigated. To this
end, various approaches to the possible utility or origin were considered, particularly potential benefits of the faster
generational change caused by possible evolutionary compound interest. Reference was made to the thinking of
Weismann, the father of regulated aging theories, who advocated non adaptive concepts at the end of his career. In a
thought experiment, circadian rhythms are discussed as a possible molecular source of aging regulation.
Keywords: August Weismann, aging theory, programmed aging, semelparity, MASON model, circadian rhythm, clock
genes
БИОХИМИЯ том 87 вып. 1 2022