БИОХИМИЯ, 2022, том 87, вып. 12, с. 1928 - 1946
УДК 577.24
МНОГОГРАННЫЙ НОТОБРАНХИУС
Обзор
© 2022 Е.В. Булавкина1,2, А.А. Кудрявцев2, М.А. Гончарова2, М.С. Ланцова2,
А.И. Шувалова2, М.А. Ковалев2, А.В. Кудрявцева1,2*
1 Институт молекулярной биологии имени В.А. Энгельгардта РАН,
Центр высокоточного редактирования и генетических технологий для биомедицины,
119991 Москва, Россия; электронная почта: rhizamoeba@mail.ru
2 Институт молекулярной биологии имени В.А. Энгельгардта РАН,
лаборатория постгеномных исследований, 119991 Москва, Россия
Поступила в редакцию 03.11.2022
После доработки 11.11.2022
Принята к публикации 11.11.2022
Научный интерес к короткоцикловым рыбам рода Nothobranchius с каждым годом стремительно
возрастает. Всесторонне исследуются самые разные аспекты жизнедеятельности этих икромечущих
карпозубых и особенно их старение. Обитание в пересыхающих водоёмах Африки редко позволяет
нотобранхиусам пережить более одного сезона дождей, поэтому естественный отбор не был направ-
лен на длительное выживание взрослых особей, что привело к снижению эффективности системы
репарации ДНК. Старение нотобранхиусов изучается как в норме, так и при воздействии потен-
циальных геропротекторов, а также генетических модификаций. Большая часть биогеронтологиче-
ских исследований проводится на Nothobranchius furzeri (изолят GRZ) с продолжительностью жиз-
ни 3-7 месяцев. Однако перечень модельных видов нотобранхиусов существенно шире, а спектр
передовых научных направлений с их участием выходит далеко за пределы геронтологии. В обзоре
рассматриваются наиболее интересные и перспективные направления, развивающиеся благодаря
рыбам рода Nothobranchius. Затрагиваются как классические исследования, связанные с контролем
продолжительности жизни, так и относительно новые, включающие специфику протекания диапа-
узы, сложности систематики и филогении, становление механизмов определения пола, изменение
количества хромосом, появление в геноме множества повторяющихся последовательностей ДНК,
когнитивно-поведенческие особенности и социальную стратификацию, а также методические
сложности при работе с нотобранхиусами.
КЛЮЧЕВЫЕ СЛОВА: старение, животные модели старения, Nothobranchius, возраст-зависимые забо-
левания, диапауза, нейродегенерация, продолжительность жизни, долголетие, видообразование, карио-
тип, стресс.
DOI: 10.31857/S0320972522120120, EDN: NHENPO
ВВЕДЕНИЕ
методической платформы для тестирования
потенциальных геропротекторов и анализа
Современная геронтология, как фунда-
механизма их действия на позвоночных жи-
ментальная, так и прикладная, немыслима
вотных [6-8]. Ранее исследования были сфо-
без исследований, проводимых на рыбах рода
кусированы на таких моделях, как Caenorabditis
Nothobranchius [1-5]. Самый короткоживущий
elegans и Drosophila melanogaster [9-13], однако
представитель рода, Nothobranchius furzeri Jubb,
при поиске геропротекторных интервенций
1971 (изолят GRZ) успевает состариться и уме-
для человека лучше использовать более эволю-
реть естественным образом всего за 3-7 меся-
ционно близкие модельные организмы. При
цев (не дольше 12 месяцев для вида в целом),
использовании эволюционно далёких моделей
что открывает огромные перспективы в иссле-
намного выше риск с самого начала отказаться
довании генетических и биохимических ос-
от интервенций, способных хорошо работать
нов процесса старения, а также в разработке
на человеке. Помимо этого, тестирование на
* Адресат для корреспонденции.
1928
МНОГОГРАННЫЙ НОТОБРАНХИУС
1929
позвоночных животных является обязательным
относящиеся к семейству Нотобранхиевых
этапом исследований после проведения экспе-
(Nothobranchiidae), отряду Карпозубообраз-
риментов на беспозвоночных. Поэтому исполь-
ных (Cyprinodontiformes). Род Nothobranchius
зование нотобранхиусов позволяет сократить
довольно большой: описано несколько десят-
срок разработки лекарственных препаратов.
ков валидных видов, каждый год к ним добав-
Самые популярные модельные животные сре-
ляют новые. Территория распространения рода
ди позвоночных живут долго: грызуны живут
Nothobranchius сравнительно небольшая, усло-
2-3 года, а Danio rerio - до 4-5 лет. Таким об-
вия существования сильно ограничены фактора-
разом, нотобранхиусы стали для геронтологов
ми внешней среды, однако экологические ниши
ценной находкой [5, 14], тем более что один из
разных видов практически не различаются.
важнейших принципов биоэтики относитель-
Исследователей интересуют прежде всего
но выбора модельного животного гласит, что по
причины и механизмы формирования такого
возможности следует проводить эксперимент
«высокого» видового разнообразия. Темати-
на организме, находящемся на как можно более
чески связанным оказывается вопрос выбо-
низком уровне эволюционного развития.
ра оптимальных систематических признаков
Тестирование экспрессии генов в различ-
для разграничения видов в пределах рода
ных тканях, анализ микробиома кишечника,
Nothobranchius.
а также оценка когнитивно-поведенческих
Каковы причины и механизм бурного видо-
детерминант при исследовании геропротек-
образования в пределах рода Nothobranchius?
торов позволяют комплексно изучать влияние
По этому вопросу уже сформировалась опре-
на организм различных интервенций [15, 16] и
делённая гипотеза. Поскольку рыбы рода
работать в области продления активной фазы
Nothobranchius населяют сезонно пересыхаю-
жизни, т.е. обеспечивать активное долголе-
щие водоёмы в Центральной и Восточной Аф-
тие [17, 18]. В условиях демографического пе-
рике, их местообитание ограничено неболь-
рехода в Российской Федерации и многих ев-
шими, малоподвижными водно-болотными
ропейских странах такие вопросы становятся
угодьями, включающими поймы в верховьях
архиважными.
рек, сезонно затопляемые долины, маршевые
Преимущества нотобранхиусов не огра-
луга и русла медленно текущих мелких сезон-
ничиваются исследованием механизмов ста-
ных рек. Территория распространения ното-
рения. Существует множество удивительных
бранхиусов также ограничивается областями
особенностей, свойственных представителям
с определённым составом грунта, так как для
этого рода, которые приковывают к себе вни-
длительного сохранения икры необходимо,
мание учёных. Например, тонкая регуляция
чтобы влага задерживалась в достаточном ко-
процессов входа эмбриона в диапаузу и выхо-
личестве. Нотобранхиусы были описаны в
да из неё, что необходимо для коррекции пе-
семи районах Африки: Нило-Судан, Викто-
риода нахождения в стадии икры до момента
рия, Танганьика, Малави, Восточное побере-
наступления погодных условий, подходящих
жье, Замбези и Конго [22]. Представители рода
для выклева малька (сезона дождей) [19-21].
Nothobranchius являются эталонным приме-
Изучение способности к почти полному вы-
ром аллопатрического видообразования. Оно
ключению метаболизма крайне важно для раз-
происходит под влиянием изменений релье-
работки подходов к освоению человеком кос-
фа и обусловливает наличие весьма ограни-
мического пространства.
ченного ареала каждого вида при их общем
В обзоре рассмотрены наиболее перспек-
большом количестве. Распространение рода
тивные направления изучения нотобранхиу-
в значительной степени совпадает с Восточ-
сов, интересующие исследовательские группы
но-Африканской рифтовой долиной, а геобио-
по всему миру - от полового поведения и соци-
логический анализ подтверждает тектониче-
альной иерархической системы до особенно-
скую гипотезу эволюции нотобранхиусов.
стей структуры и функционирования генома.
Видообразование прежде всего связано с
накоплением изменений в геноме. Большое
количество видов в пределах одного рода гово-
ПРОБЛЕМЫ СИСТЕМАТИКИ
рит о том, что для таксона в целом характерна
И ЭВОЛЮЦИИ РОДА Nothobranchius
высокая скорость накопления мутаций. Мож-
но предположить, что у нотобранхиусов такая
Представители рода Nothobranchius Peters,
предрасположенность связана с ускоренным
1868 - стенотопные икромечущие костистые
жизненным циклом, а точнее - с рано при-
рыбы, обитающие в пересыхающих пресно-
ходящей старостью, неизменно связанной с
водных водоёмах на территории Африки и
накоплением генетических ошибок в каждой
БИОХИМИЯ том 87 вып. 12 2022
1930
БУЛАВКИНА и др.
клетке организма [22]. Получается, что систе-
В природных популяциях изредка встречается
ма репарации повреждений ДНК нотобран-
ситуация, когда на одной территории одновре-
хиусов может не быть очень совершенной,
менно обитают разные виды нотобранхиусов.
так как организму не требуется, чтобы она в
В литературе их называют симпатрическими
течение длительного периода его жизни ис-
видами. Однако, во-первых, обычно времени
правляла ошибки. Естественный отбор не был
проведения экспедиции недостаточно для де-
направлен на решение такой задачи. Сочета-
тального анализа видового состава водоёма и
ние высокой частоты мутаций с генетическим
популяционных особенностей, а, во-вторых,
дрейфом при коротком жизненном цикле, за-
идентификация потенциальных гибридов да-
висящем от сезонов дождей, в совокупности
леко не всегда возможна по причине отсутствия
задали быстрые темпы видообразования [23].
разработанной системы маркеров.
Перечисленные признаки вместе с минималь-
Большие надежды возлагались на моле-
ным расхождением по экологическим нишам
кулярную систематику, но и в этой области
говорят о неадаптивном процессе видообразо-
исследователи столкнулись со сложностями.
вания в пределах рода Nothobranchius [24].
Оказалось, что очень ограниченные ареалы рас-
Какие таксономические признаки наиболее
пространения не дают возможности сформиро-
адекватны для разграничения видов нотобран-
вать представление о внутривидовой межпопу-
хиусов? Систематика нотобранхиусов ослож-
ляционной вариабельности, т.е. молекулярные
нена несколькими обстоятельствами. Прежде
методы не позволяют дать точный ответ на
всего, не до конца очевидны видовые критерии,
вопрос, являются ли похожие изоляты одним
т.е. регулярно возникает вопрос, является ли
видом или разными. Однако можно с уверен-
новый изолят представителем уже известного
ностью заключить, что род Nothobranchius - это
вида, или он должен быть описан в качестве но-
уникальный объект для исследования микро-
вого [25]. Эта проблема стоит довольно остро,
эволюции, и учёным ещё предстоит сделать в
так как почти каждая научная экспедиция воз-
этой области много открытий [22].
вращается с ранее не описанными видами ното-
В заключение следует отметить, что из-
бранхиусов. Важным диагностическим призна-
учение флоры и фауны Африки осложняет-
ком является специфический паттерн окраски
ся стремительной урбанизацией территорий,
самцов [26]. Ситуация осложняется, с одной
приводящей к изменению условий среды и,
стороны, наличием различных цветовых морф
следовательно, к сокращению биоразнообра-
одного и того же вида (например, у N. furzeri,
зия. Поэтому уже в ближайшем будущем пла-
Nothobranchius korthause, Nothobranchius hassoni),
нета может лишиться многих известных науке
с другой стороны - существованием изоля-
нотобранхиусов и потерять ещё даже не от-
тов-двойников, фенотипически очень похожих,
крытые виды этих удивительных организмов.
но имеющих различное количество хромосом.
Интересна существующая практика при-
своения видовых эпитетов новым видам но-
РАЗНООБРАЗИЕ КАРИОТИПОВ,
тобранхиусов. Довольно часто новый вид на-
ОСОБЕННОСТИ ОРГАНИЗАЦИИ ГЕНОМА
зывают в честь специалиста, который выделил
И ЭВОЛЮЦИОННОЕ СТАНОВЛЕНИЕ
изолят в природе и привёз его из экспедиции.
СИСТЕМЫ ДЕТЕРМИНАЦИИ ПОЛА
При этом описанием вида занимаются со-
всем другие исследователи, как правило, даже
В течение длительного времени, в доге-
не в соавторстве с первооткрывателем [27, 28].
номную эру, кариология была важным ин-
Встречаются трогательные случаи, когда ис-
струментом систематики и позволяла найти
следователи называют новые виды в честь
дополнительные признаки там, где не хватало
близких, например, Nothobranchius ditte был
морфологических. Кариотипирование помо-
описан известным исследователем короткоцик-
гало также объяснить причины стерильно-
ловых рыб Béla Nagy и получил свой видовой
сти гибридов, обнаружить полиплоидизацию,
эпитет в честь его жены Эдит [29].
определить половую принадлежность в раннем
В систематике одним из главных видовых
возрасте до появления фенотипических при-
критериев считается способность к свободному
знаков. Однако специалисты по кариологии
скрещиванию организмов и появлению пло-
нотобранхиусов до сих пор не могут прийти к
довитого потомства, сохраняющего признаки
единому выводу даже по самым базовым вопро-
вида в ряду поколений. Но при работе с ното-
сам. Дело в том, что кариотип различных пред-
бранхиусами проведение таких тестов в лабора-
ставителей рода Nothobranchius характеризуется
торных условиях сильно ограничено отсутстви-
необыкновенно высокой вариабельностью [30].
ем представительной биоресурсной коллекции.
Количество хромосом варьирует от 16 до 50.
БИОХИМИЯ том 87 вып. 12 2022
МНОГОГРАННЫЙ НОТОБРАНХИУС
1931
Причины и механизмы возникновения такого
Учитывая колоссальный разброс в количе-
разнообразия, а также направление эволюци-
стве хромосом у разных видов, сразу возникает
онного процесса вызывают огромный интерес
предположение, что на каком-то этапе эволю-
исследователей. В настоящее время некото-
ции возникла полиплоидизация, после кото-
рая ясность в эти вопросы уже внесена, одна-
рой произошло расхождение дублированных
ко работ по исследованию кариотипа крайне
хромосом по структуре и функциям. В этом
немного, в связи с популярностью геномного
случае виды с наименьшим количеством хро-
секвенирования, вытесняющего кариотипиро-
мосом являлись бы наиболее эволюционно
вание.
древними. Второй идеей было принятие «мно-
Также представляют научный интерес ме-
гохромосомных» видов (имеющих диплоидный
ханизмы определения пола у нотобранхиусов.
набор более 38 хромосом, в том числе N. ditte,
Многие аквариумисты, которые содержали
Nothobranchius malaissei, Nothobranchius brieni) в
различные виды рода Nothobranchius, замети-
качестве предковых форм. Однако существу-
ли, что при получении мальков и их последу-
ющие филогенетические деревья, а также де-
ющем подращивании соотношение полов 1 : 1
тальное изучение кариотипа различных видов
не соблюдается почти никогда. Например, у
свидетельствуют о том, что, скорее всего, пред-
Nothobranchius guentheri на 1 самку приходится
ковой формой был вид с 38 хромосомами.
около 5 самцов. Распространено также мнение,
У подрода Nothobranchius
[37]
(2n = 38)
что в пределах рода Nothobranchius сосуществу-
изменения кариотипа происходили в основ-
ют как механизмы определения пола фактора-
ном путём перицентрических инверсий. Че-
ми внешней среды, так и генетическая обуслов-
тыре вида - N. furzeri, Nothobranchius kadleci,
ленность полового статуса.
Nothobranchius orthonotus и Nothobranchius
Самое передовое и популярное направле-
kuhntae - отличались друг от друга соотноше-
ние - это, несомненно, проведение омиксных
нием одноплечих и двуплечих хромосом [35].
исследований, прежде всего секвенирование
Для видов с числом хромосом меньше 38 ха-
генома и транскриптома [18]. Знание последо-
рактерна их редукция путём слияний. При этом
вательности ДНК в геноме вида открывает ши-
в кариотипах видов с наименьшим числом хро-
рокие возможности молекулярно-генетических
мосом (16 и 18 соответственно) преобладали
исследований, а также позволяет уточнить его
двуплечие хромосомы (Nothobranchius rachovii -
положение на филогенетическом древе. Одна-
16, Nothobranchius krysanovi - 18).
ко у нотобранхиусов сборка генома сильно ос-
Для рода Nothobranchius характерно обилие
ложняется наличием огромного количества по-
не только внутрихромосомных, но и межхро-
второв, происхождение и роль которых также
мосомных перестроек. В хромосомной эволю-
вызывают жаркие дискуссии.
ции рода выявлены две основные тенденции:
Как эволюционировал кариотип нотобран-
слияние хромосом (или, реже, расщепле-
хиусов, и сколько хромосом было у предко-
ние на две хромосомы) и перицентрические
вого вида? Говоря о причинах и механизмах
инверсии [35].
эволюции кариотипа нотобранхиусов, важно
Подводя итог, можно сказать, что, согласно
отметить, что хромосомные перестройки
-
современным взглядам, эволюционно первич-
естественное и частое явление при высокой
ным является набор хромосом 2n = 38. Далее
скорости видообразования. Наиболее частые
параллельно в нескольких группах рыб шли
изменения связаны со смещением центромер
процессы уменьшения и увеличения количе-
на хромосомах и инверсиями, которые могут
ства хромосом. Все случаи с увеличенным ко-
способствовать локальной адаптации за счёт
личеством хромосом произошли независимо
подавления рекомбинации и, таким образом,
друг от друга. Механизмы увеличения числа
накопления связанных адаптивных генов. Важ-
хромосом связаны не с полиплоидизацией, а с
но, что такие события часто приводят к генети-
расщеплением существующих хромосом на бо-
ческой несовместимости, снижая фертильность
лее мелкие. Процессы уменьшения количества
гибридов, способствуя репродуктивной изоля-
хромосом эволюционно связаны с их слиянием.
ции и видообразованию [31-34].
Каким образом у разных видов нотобранхиу-
Описаны кариотипы более чем 65 видов [35].
сов определяется пол? Среди описанных ка-
Все известные нотобранхиусы диплоидны (2n),
риотипов представителей рода Nothobranchius
однако количество хромосом у разных видов
у пяти видов были описаны морфологически
варьирует от 16 до 50 (чаще всего 36, реже -
различимые половые хромосомы, а именно у
38) [36]. Эта особенность делает нотобранхиусов
N. guentheri [38], N. brieni [36], Nothobranchius
довольно привлекательной моделью для специа-
lourensi, Nothobranchius janpapi и N. ditte [35]. Од-
листов в области эволюции и экологии.
нако у всех изученных видов различается даже
БИОХИМИЯ том 87 вып. 12 2022
1932
БУЛАВКИНА и др.
хромосомный механизм определения пола.
mir-430 - важного регулятора экспрессии ге-
Рассмотрим, например, генотип N. brieni [36].
нов в половой линии у рыб [41]. Конкретный
Количество хромосом N. brieni у самцов со-
механизм действия gdf6 и распространённость
ставило 2n = 49, у самок - 2n = 50. Женский
системы определения пола с его участием у
кариотип состоял из 25 пар акроцентрических
других видов нотобранхиусов требуют дальней-
хромосом, постепенно уменьшающихся в раз-
шего изучения. Наличие таких разнообразных
мерах. Мужской кариотип состоял из 23 пар
систем половых хромосом делает род Notho-
акроцентрических хромосом, одной двупле-
branchius перспективной моделью для исследо-
чей пары и двух непарных акроцентрических
вания путей становления генетической детер-
хромосом. В хромосомах первого мейоза при
минации пола.
сперматогенезе наблюдалось 23 бивалента и
Почему же все-таки у N. guentheri соотно-
тривалента при диакинезе. Таким образом,
шение самцов и самок всегда настолько дале-
вид N. brieni - обладатель многополой хромо-
ко от ожидаемого 1 : 1? Скорость роста и, как
сомной системы типа X1X2Y/X1X1X2X2. Одна
следствие, размеры тела самцов превышают
двуплечая нео-Y-хромосома, скорее всего,
аналогичные показатели у самок, поэтому при
возникла в результате робертсоновского сли-
совместном выращивании мальков самки не
яния между Y-хромосомой и аутосомой, как
выдерживают конкуренции и либо становятся
это было описано для других видов рыб [39].
«ползунами», либо поедаются более крупными
У N. brieni и Nothobranchius sp. «Kasenga» Y-хро-
особями. Как только в нашей лаборатории на-
мосома является крупной метацентрической,
чали сортировать мальков согласно их размеру
а X1- и Х2-хромосомы - акроцентрическими
в пределах одного контейнера, соотношение
разного размера [39]. У четырёх неблизкород-
полов сразу приблизилось к эталонному и со-
ственных видов в подродах Zononothobranchius
ставило 1 : 1.
(N. brieni) и Adiniops (N. guentheri, N. lourensi
Чем необычен геном нотобранхиусов? Разви-
и N. janpapi) [40] множественные половые хро-
тие методов высокопроизводительного секве-
мосомы, предположительно, возникли незави-
нирования позволило не только секвенировать
симо друг от друга [35].
геномы нескольких видов рода Nothobranchius,
У некоторых видов не удаётся выделить
но и перейти к работам в области функцио-
морфологически различимые половые хромо-
нальной геномики. Всё шире применяется
сомы, однако идентифицированы SDR-локу-
анализ транскриптома, исследуются и неко-
сы - последовательности ДНК, конкретные
дирующие РНК, метилом и состав сообществ
различия в которых определяют пол особи.
микроорганизмов, населяющих главным об-
В качестве примера рассмотрим вид N. furzeri,
разом желудочно-кишечный тракт [16]. Ана-
который имеет XY-систему определения пола,
лиз бактериального состава кишечника может
причём его половые хромосомы практически
выполняться как культуральными методами,
неразличимы морфологически. Самцы явля-
так и при помощи высокопроизводительного
ются гетерогаметным полом, несущим уни-
секвенирования [16, 42], позволяя анализиро-
кальную версию гена gdf6, являющегося пред-
вать метаболический потенциал микроорга-
ставителем семейства факторов роста TGF-β и
низмов и их влияние на метаболизм хозяина.
запускающего развитие по мужскому типу [41].
Гаплоидный геном N. furzeri состоит из
Y-вариант gdf6 отличается от X-варианта на-
19 хромосом (2n = 38). Размер генома состав-
личием 15 аминокислотных замен и 3 деле-
ляет примерно ~1,5 млрд п.н. [41], при этом
ций. Важно отметить, что замены произошли
аннотировано более 22 000 белок-кодирующих
в высококонсервативной для позвоночных об-
генов [43]. Секвенирование генома значитель-
ласти, это свидетельствует о сильной положи-
но упростило применение молекулярно-био-
тельной селекции в сторону накопления мута-
логических методов и подходов. Для наиболее
ций в этом гене.
короткоживущего и востребованного изолята
Сразу после выклева оба аллеля gdf6 экс-
N. furzeri GRZ на основе анализа генома был
прессируются в сопоставимой степени, од-
сделан вывод о высоком уровне инбредности.
нако спустя уже 3 дня экспрессия у самцов
Было показано, что геном N. furzeri содержит
значительно превышает экспрессию у самок.
высокую долю повторов, достигающую 45%,
У взрослых рыб проходит зависимая от пола
что очень велико для костистых рыб [41, 43].
экспрессия gdf6: тогда как в яичниках у са-
Эта особенность усложняет сборку генома но-
мок его экспрессия очень низка, в семенни-
тобранхиусов, однако эволюционное проис-
ках у самцов он детектируется, что может быть
хождение такой специфической черты и её
связано с ещё одной делецией, произошед-
роль в жизнедеятельности этих рыб крайне ин-
шей в 3′-UTR и вырезавшей сайт связывания
тересны. Наша гипотеза сводится к тому, что
БИОХИМИЯ том 87 вып. 12 2022
МНОГОГРАННЫЙ НОТОБРАНХИУС
1933
происхождение большого количества повторов
окружении агрессивных факторов внешней
ассоциировано с повышенной активностью
среды. В процессе эволюции они смогли выра-
элементов генома, способных к транспозиции.
ботать ряд адаптационных механизмов, кото-
Повышенную активность транспозонов мы
рые помогают им успешно выживать и активно
наблюдали, в частности, при сопоставлении
эволюционировать. Детальная расшифровка
транскриптомов самцов N. guentheri разного ие-
этих механизмов интересует огромное коли-
рархического статуса. Транспозоны содержат в
чество исследователей, поскольку важна не
своём составе регуляторные последовательно-
только в фундаментальном аспекте, но имеет и
сти, оказывающие цис- и транс-воздействие на
важный прикладной потенциал.
экспрессию специфических белок-кодирующих
Какие основные направления адаптаций от-
генов. Таким образом, они способны изменять
личают нотобранхиусов? Условия существова-
механизмы морфогенеза, играя роль в адапта-
ния нотобранхиусов в сезонно засушливых ме-
ции организма к факторам окружающей среды.
стообитаниях африканских саванн отличаются
В геноме N. furzeri представлены тандемные
чрезвычайной изменчивостью. Многие фак-
повторы, подвижные элементы (транспозоны и
торы изменяются очень быстро и в очень ши-
ретротранспозоны), псевдогены и сегментные
роком диапазоне значений (например, суточ-
дупликации. Среди них тандемные повторы
ные колебания температуры от 14 до 37 °С) [1].
составляют рекордные 20% генома, что силь-
Для того чтобы выживать в таких условиях,
но выделяет N. furzeri среди других рыб. Среди
животные должны сформировать особые меха-
тандемных повторов особенно широко пред-
низмы адаптации. По-видимому, нотобранхиу-
ставлены два GC-богатых минисателлита, ло-
сы имели некую преадаптацию к условиям оби-
кализующиеся в перицентромерных регионах
тания в составе эфемерной фауны, поскольку
и имеющие длину 77 и 49 нуклеотидов. Возле
их геном содержит довольно много мобильных
центромер также локализуется имеющая ши-
элементов, способных к транспозиции и опо-
рокое распространение сателлитная последова-
средующих, таким образом, инсерционный
тельность, состоящая из 348 нуклеотидов с низ-
мутагенез. Этот процесс лежит в основе изме-
ким содержанием GC.
нений геномов при естественном отборе [46].
Подвижные элементы занимают ~ 25% ге-
В эволюции свойство транспозонов переме-
нома и представляют собой важную особен-
щаться в специфические сайты генома, регули-
ность генома N. furzeri [41].
ровать экспрессию генов и взаимодействовать
Как ядерный, так и митохондриальный ге-
с транскрипционными факторами, наряду со
ном нотобранхиусов имеют большие размеры
способностью реагировать на экологические
относительно родственных видов рыб, не явля-
стрессоры, является основой для быстрой из-
ющихся короткоцикловыми. Увеличение разме-
менчивости и видообразования за счёт моду-
ра ядерного генома в значительной степени об-
лирования управления онтогенезом. Таким
условлено повышенным количеством элементов
образом, нотобранхиусы приспособились к
генома, способных к транспозиции, причиной
постоянно меняющимся условиям, в том чис-
которого может быть генетический дрейф. Ми-
ле за счёт увеличения количества транспозонов
тохондриальный геном у N. furzeri, также увели-
в геноме.
ченный (>19,5 кб) по сравнению с другими поз-
Помимо этого, представители рода
воночными (~16 кб), характеризуется большей
Nothobranchius обладают уникальным набором
суммарной длиной некодирующих областей [43].
репродуктивных адаптаций, одна из которых -
Вариабельность кариотипа в пределах рода
наличие диапаузы, т.е. способности эмбрионов
Nothobranchius очень велика по сравнению с
в определённые периоды существенно затор-
другими родами костистых рыб [44, 45], что
маживать обмен веществ. Одним из основных
предполагает возникновение в процессе эво-
преимуществ является возможность регуляции
люции событий, связанных с частой крупно-
общего времени нахождения эмбриона в за-
масштабной реструктуризацией геномов, что
щитной оболочке икры, что критически важно
также соотносится с бурным видообразовани-
при наличии флуктуаций в погодных условиях
ем в пределах рода Nothobranchius [43].
из года в год [44, 45].
Помимо регуляции временного перио-
да, важным адаптационным механизмом ста-
АДАПТАЦИОННЫЙ ПОТЕНЦИАЛ
ло выживание эмбрионов в условиях засухи
НОТОБРАНХИУСОВ
благодаря возникновению толстой оболочки
икры, строение и состав которой эволюци-
В естественных местообитаниях нотобран-
онно сформировались в условиях окружения
хиусы практически постоянно находятся в
смектитовыми глинами. Икринки способны
БИОХИМИЯ том 87 вып. 12 2022
1934
БУЛАВКИНА и др.
сохранять влагу и развиваться в таком грунте в
перед гаструляцией), на этом этапе пул эмбрио-
течение длительного периода времени. Имен-
нов развивается синхронно. Синхронно он
но гидроморфные свойства растрескивающих-
входит и в диапаузу II, которая начинается в
ся глинистых субстратов определяют ключевую
середине сомитогенеза, в период формирова-
роль в неравномерном распространении этих
ния большинства органов. Это случается уже
видов в тропических ландшафтах. Рыбы рода
во время засушливого периода. Диапауза II ха-
Nothobranchius мечут икру в мягкий ил, кото-
рактеризуется снижением синтеза белка, оста-
рый имеет типичные характеристики вертисо-
новкой клеточного цикла, ремоделированием
ля. Критически важно, что входящие в состав
энергетического обмена и контролируется сиг-
минералы группы смектитов (главным обра-
нальной системой инсулиноподобного факто-
зом монтмориллонит) обладают способностью
ра роста. Митохондрии, выделенные из эмбрио-
поглощать молекулы воды в кристаллических
нов, остановленных в диапаузе II, не готовы
слоях. В период засухи вода испаряется, обра-
производить АТР, а, скорее, перемещают угле-
зуя в почве глубокие трещины. Таким образом,
род и электроны через цикл Кребса, миними-
в течение сухого сезона глубоко внутри этих
зируя генерацию протон-движущей силы [48].
растрескивающихся глин, которые поддер-
Диапауза III происходит на заключительной
живают оптимально влажный микроклимат,
стадии развития, непосредственно перед вы-
икринки способны войти в состояние диапа-
клевом [47]. Асинхронность приходилась на
узы и остаться жизнеспособными до следую-
начало и конец засушливого сезона.
щего сезона дождей [26, 44]. Со временем, в
Все три стадии диапаузы являются обя-
процессе эволюции, диапауза стала не просто
зательными, но могут длиться разное время
жизненно необходимой адаптацией для пере-
в зависимости от внешних раздражителей.
жидания засухи, но и приобрела дополнитель-
У Austrofundulus limnaeus процесс входа в диа-
ную роль: прохождение стадий диапаузы необ-
паузу и выхода из неё связан с температурой
ходимо для формирования здоровой особи.
окружающей среды, а также с определёнными
Феномен диапаузы у позвоночных, и механиз-
сигналами, передаваемыми от материнско-
мы её контроля. Диапауза встречается преиму-
го организма [20, 49]. Например, вероятность
щественно у беспозвоночных животных, поэто-
того, что эмбрион войдёт в длительную диа-
му её изучение на модели рыб является особенно
паузу, зависит от возраста самки и количества
ценным в аспекте потенциального практиче-
яиц, которые она уже произвела [49]. При раз-
ского применения результатов. Основная часть
ведении N. guentheri и N. rachovii в лаборатор-
научных исследований диапаузы выполнена
ных условиях мы выявили сходный паттерн.
на родственниках нотобранхиусов из Южной
Обнаружено, что процесс эмбрионального
Америки, срок развития их икры существенно
развития, в частности прохождение диапаузы,
меньше, однако принято считать, что механиз-
различается у рыб, живущих в дикой природе
мы должны быть во многом схожими.
и рыб, содержащихся в условиях лаборатории.
Выделяют три стадии диапаузы, которые
Основным отличительным признаком являет-
в целом рассматриваются как этапы одного
ся сокращение срока диапаузы, например, у
процесса, однако их продолжительность мо-
N. furzeri диапауза III в стандартных условиях
жет существенно варьировать. Такая система
инкубации практически не наблюдается. Ско-
подразумевает различные регуляторные меха-
рее всего, она становится крайне непродолжи-
низмы для каждой стадии диапаузы. Важную
тельной. Это может отражаться на результатах
роль в регуляции прохождения этих стадий
научных экспериментов и вызывает необходи-
играют условия окружающей среды [47]. Были
мость детального изучения различий.
изучены и молекулярные механизмы, опреде-
ляющие возможность протекания диапаузы,
например, её зависимость от сигнального пути
КОГНИТИВНО-ПОВЕДЕНЧЕСКАЯ СФЕРА
витамина D [21]. Сделано предположение, что
И ОСОБЕННОСТИ СОЦИАЛЬНОГО
существует специальный путь для интеграции
ПОВЕДЕНИЯ
информации об окружающей среде и её преоб-
разования в программы развития, связанные с
В отличие от D. rerio, традиционного мо-
эволюционными преобразованиями у живот-
дельного организма, представители рода Notho-
ных. В роли сигнальных молекул рассматрива-
branchius не ведут стайный образ жизни и явля-
ют 7-дигидрохолестерин-производные гормо-
ются хищниками. Всемирная организация здраво-
нов и связанные с ними ядерные рецепторы.
охранения даже рассматривала нотобранхиусов
В природе диапауза I происходит ещё в
в качестве биологического средства борьбы с ма-
сезон дождей (во время дисперсионной фазы
лярией в Африке, предполагая, что они способ-
БИОХИМИЯ том 87 вып. 12 2022
МНОГОГРАННЫЙ НОТОБРАНХИУС
1935
ны массово истреблять личинки комаров [50].
нию рыб, на оценку смелости и склонности к
По-видимому, именно хищничество позволи-
риску. Результаты данного исследования под-
ло нотобранхиусам развить в процессе эволю-
твердили различия в последовательных ин-
ции когнитивные способности и опосредовало
дивидуальных поведенческих реакциях, а все
появление иных поведенческих паттернов по
поведенческие показатели, отражающие дви-
сравнению с другими, нехищными, видами.
гательную активность и склонность к риску,
Можно ли говорить о том, что рыба облада-
оказались повторяемыми [54].
ет характером? Существует крайне интересная
Индивидуальные различия поведения меж-
ниша исследований, направленная на изуче-
ду особями наблюдались и в наших исследова-
ние стабильных индивидуальных различий в
ниях. Для тестирования экспериментального
поведении. Был введён термин «характер жи-
препарата рыбы вида Nothobranchius foershi по-
вотного» (animal personality) [51], и вариации
мещались в пробирки объёмом 50 мл на 1 час
поведения были подтверждены для некоторых
4 раза в день. После нахождения в пробирке
видов рыб [52, 53].
рыбы демонстрировали два разных паттерна
Экспериментально доказано, что особи
поведения. Одни после освобождения стреми-
вида N. furzeri обладают индивидуальными по-
тельно уплывали и прятались у дальней стенки
веденческими вариациями. Для того чтобы до-
аквариума, в то время как другие подплывали
казать эту особенность, были проведены тесты
к передней стенке и демонстрировали поведен-
на склонность к исследовательскому поведе-
ческие признаки агрессии.
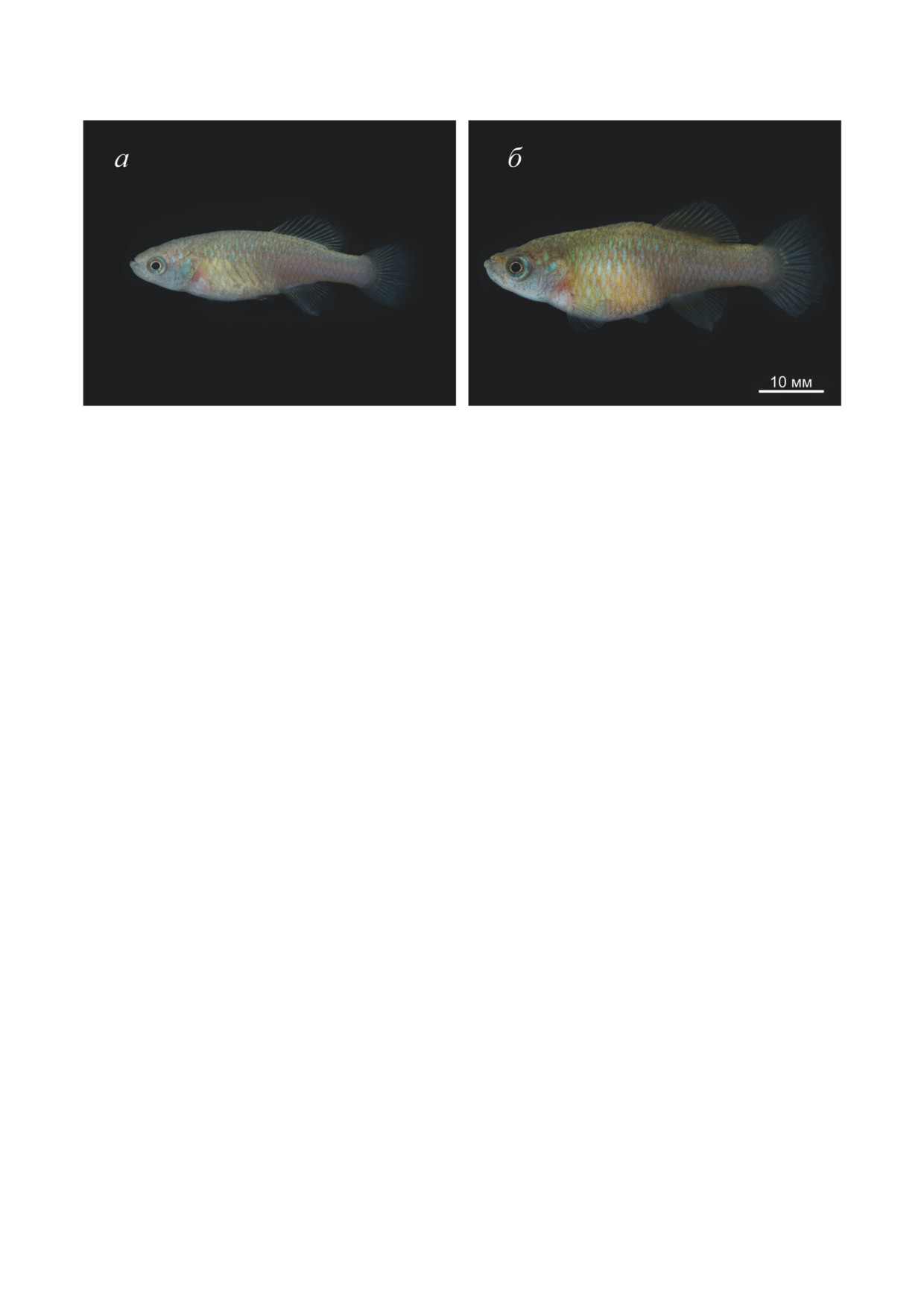
Рис. 1. Социальная стратификация в искусственно сформированных группах самцов Nothobranchius guentheri (а и б)
и Nothobranchius rachovii (в и г) при избыточном кормлении личинками Chironomus. Уровень в иерархической лестнице
находит отражение в размере особи, особенностях её окраски, а также поведенческом паттерне. Доминантные самцы
крупнее и ярче (б и г), в то время как низкоранговые особи имеют меньшие размеры, бледную окраску и менее выра-
женный рисунок на поверхности тела (а и в)
БИОХИМИЯ том 87 вып. 12 2022
1936
БУЛАВКИНА и др.
Рис. 2. В условиях аквакультуры среди нормальных самок N. guentheri средних размеров (а) отдельные особи маскули-
низируются (б). Они могут значительно отличаться по размеру, иметь более тёмную окраску и проявлять высокий уро-
вень агрессии по отношению к самкам и самцам своего вида
Как устроена иерархическая система у но-
морфологическим признакам, так и по пове-
тобранхиусов, и как это учитывать при плани-
денческим паттернам (рис. 1).
ровании экспериментов? Широко известно, что
При лабораторном разведении самок
животные одного вида в большинстве случаев
N. guentheri как в изоляции от самцов, так и при
формируют иерархическую социальную струк-
совместном содержании также наблюдается ди-
туру при попадании в условия с ограниченны-
версификация особей по морфологическим
ми ресурсами [55]. Выявляются доминантные,
признакам. Однако если у самцов формируется
иногда субдоминантные, и низкоранговые осо-
плавный переход между самыми яркими круп-
би [56].
ными и бледными мелкими особями, то для
На начальных этапах работы с нотобранхи-
самок наблюдается немного другая ситуация.
усами (N. guentheri, N. rachovii и Nothobranchius
Самым ярким событием становится появление
flammicomantis) нами было обнаружено, что
крупных агрессивных самок с более тёмной
при групповом содержании в лабораторных ус-
окраской, однако они никогда не приобретают
ловиях рыбы активно взаимодействуют друг с
фенотип и функционал самца (рис. 2).
другом. Характер взаимодействия однозначно
Причины, которые определяют роль ка-
говорил о невозможности проведения любых
кой-то конкретной особи в социальной струк-
длительных экспериментов даже в том случае,
туре, на данный момент неизвестны. Иерархи-
если группа особей формировалась ещё в ран-
ческая структура распространена среди рыб [57],
нем возрасте и в аквариум не добавлялись но-
однако она ещё недостаточно изучена у ното-
вые особи.
бранхиусов и представляет не только фун-
При недостаточно обильном кормлении
даментальный научный интерес, но и важна
(3-4 раза в день до насыщения науплиусами
для корректного составления выборки при
Artemia salina) нотобранхиусы становятся бо-
проведении различных видов эксперимен-
лее агрессивными, взаимодействие особей в
тов. Например, это может быть важным при
популяции часто приводит к увечьям, а иногда
тестировании потенциальных геропротекто-
даже к летальным исходам. Интересно, что та-
ров и проведении сопутствующих поведенче-
кие особенности социального взаимодействия
ских тестов. Также можно рассмотреть воз-
наблюдаются даже у одного из самых неагрес-
можность использования представителей рода
сивных видов, N. rachovii. При избыточном
Nothobranchius в качестве моделей для изучения
кормлении (4-5 раз в день до насыщения ли-
молекулярно-генетических основ различных
чинками комаров семейства Chironomidae)
поведенческих паттернов.
рыбы реже проявляют агрессию, направлен-
В наших экспериментах были обнаружены
ную на смерть противника, однако самцы де-
некоторые молекулярные механизмы, потен-
монстрируют по отношению к представителям
циально связанные с различиями в поведении
своего пола поведенческий паттерн, характер-
нотобранхиусов и их иерархическим поло-
ный для процесса спаривания. Помимо это-
жением. Был проведён анализ транскрипто-
го, можно наблюдать диверсификацию как по
ма самцов N. guentheri разного социального
БИОХИМИЯ том 87 вып. 12 2022
МНОГОГРАННЫЙ НОТОБРАНХИУС
1937
ранга - доминантных и наиболее низкоран-
ности можно считать стремительную потерю
говых. У доминантных самцов обнаружено
способности регулировать положение своего
значительное повышение уровня экспрессии
тела в толще воды. Чаще всего у самцов это
транспозонов, ассоциированное преимуще-
происходит при буллинге со стороны других
ственно с экспрессией белок-кодирующих ге-
самцов в аквариуме, у самок - при форми-
нов, вовлечённых в активность клеток нервной
ровании нерестовой группы, когда одна из
системы. Вероятно, высокая социальная пози-
них не получает внимание самца. Такие осо-
ция служит стрессовым стимулом, из-за кото-
би преимущественно передвигаются по дну,
рого и активируется экспрессия транспозонов.
из-за чего таких рыб называют «ползунами»
Учитывая обнаруженные изменения на
(в англоязычной литературе «belly-sliders»), и
уровне транскриптома мозга в зависимости
социальный стресс - только одна из причин
от иерархического статуса, можно предпола-
возникновения подобной патологии [58, 59].
гать наличие искажений при тестировании
Рис. 3 иллюстрирует значение социального
геропротекторов или при проведении любых
стресса, его механизм и психогенный потен-
других исследований. Полученные результаты
циал влияния на физиологическое состояние
необходимо учитывать при планировании экс-
организма.
периментов и составлении групп сравнения.
Полученные результаты могут частично
Как социальный стресс может влиять на
объяснять широко известную проблему, свя-
жизнедеятельность организма и результаты
занную со сложностями в трансляции резуль-
трансляционных исследований? Нотобранхиу-
татов экспериментов с животными на людей:
сы отличаются высокой чувствительностью к
модельные организмы при проведении экс-
социальному стрессу. В наших исследованиях
перимента содержатся в практически идеаль-
наиболее яркой демонстрацией этой особен-
ных условиях, в то время как люди постоянно
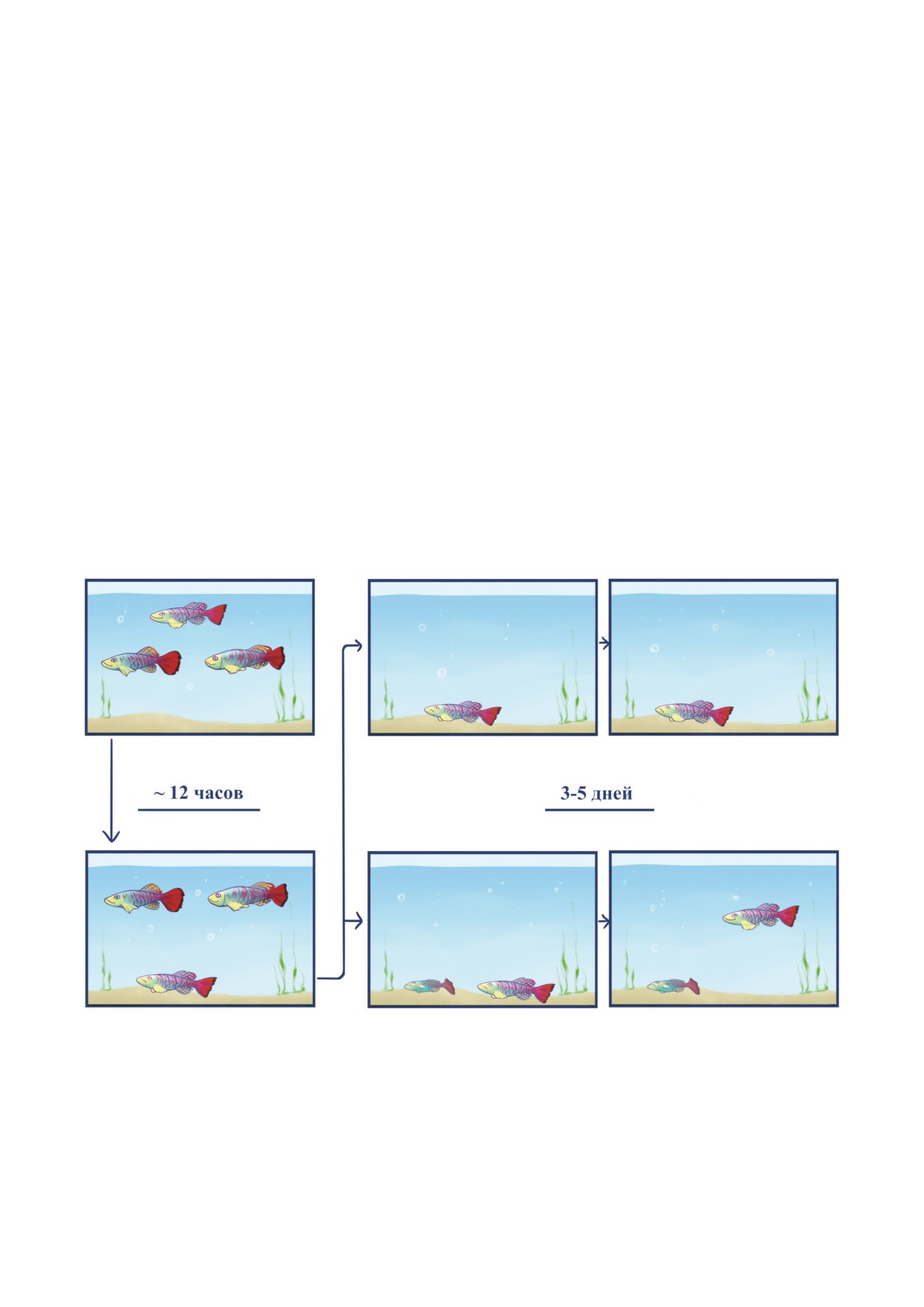
Рис. 3. Социальный стресс у самцов N. guentheri. Потенциал влияния буллинга на обоих участников взаимодействия.
Если поместить в аквариум на ночь несколько самцов, например, троих, и предоставить им корм в избытке, то с боль-
шой долей вероятности один из них к утру окажется ползуном. Начало эксперимента в дневное время, скорее всего,
приведёт к гибели одного из самцов вследствие агрессивного поведения и атак со стороны остальных. Став ползуном,
особь старается найти безопасное место в аквариуме, где её не видно. Если наутро отсадить такую особь в отдельный
аквариум, остановив стрессовое воздействие, идущее от соседей, то есть вероятность, что в течение 3-5 дней рыба вер-
нётся к исходному состоянию и перестанет быть ползуном. Если к такому ползуну подсадить более мелкого ползуна
(самца или самку в ещё более худшем состоянии), то формируется новая иерархия. Подопытный самец сам занимает
главенствующее положение и нападает на маленького. В результате эксперимента оказалось, что если имеется более
низкоранговый самец, которого можно подвергать буллингу, существенно возрастает вероятность возвращения к нор-
мальному состоянию контроля положения тела в толще воды
12
БИОХИМИЯ том 87 вып. 12 2022
1938
БУЛАВКИНА и др.
подвергаются социальному стрессу во время
Изучать механизмы старения полностью на
приёма препаратов. Стресс связан как с осоз-
человеке невозможно, особенно если речь идёт
нанием своей болезни и сложностями в ком-
о тестировании потенциальных лекарственных
муникации, которые происходят на этом фоне,
препаратов. Необходимы модельные животные.
так и с обычными рутинными ситуациями,
При их выборе приоритетными являются такие
которые почти каждый день случаются с чело-
факторы, как эволюционная близость, схожесть
веком. Наша гипотеза заключается в том, что
процессов старения с человеком и наличие ха-
для получения более релевантных результатов
рактерных признаков, которые могут быть ис-
клинических испытаний необходимо допол-
пользованы в качестве биомаркеров процесса
нять их теми факторами, которые окружают
старения. Также желательна небольшая масса
человека в реальной жизни. Подверженность
тела животного и минимальная продолжитель-
нотобранхиусов социальному стрессу предо-
ность жизни. Последние два пункта позволяют
ставляет исследователям такую возможность.
не только быстрее получить результаты рабо-
ты, но и существенно сократить временные и
финансовые затраты на тестируемый препарат.
СТАРЕНИЕ
Прежде чем принять решение о корректности
использования нотобранхиусов в качестве мо-
Подавляющее большинство животных име-
дельного организма, важно подтвердить, что
ют сходные паттерны старения, хотя известны
выбранная модель является адекватной для экс-
и исключения. Различия обычно затрагивают
траполяции результатов исследования.
среднюю продолжительность жизни и динами-
При относительно коротком жизненном
ку проявления возраст-ассоциированных при-
цикле представители рода Nothobranchius про-
знаков/заболеваний - скорость старения.
ходят типичные для млекопитающих, включая
Рис. 4. Самцы N. guentheri разных возрастов: 7 дней (а); 2 недели (б); 3 месяца (в) и 18 месяцев (г)
БИОХИМИЯ том 87 вып. 12 2022
МНОГОГРАННЫЙ НОТОБРАНХИУС
1939
Рис. 5. Самки N. guentheri разных возрастов: 7 дней (а); 2 недели (б); 3 месяца (в) и 18 месяцев (г)
человека, этапы старения. Это выражается в
ных патологий и оценивать биологический воз-
нескольких проявлениях, в том числе посте-
раст при тестировании различных интервенций.
пенности развития признаков: нотобранхиу-
Также внешние признаки старения необходимо
сы теряют насыщенность и чёткость окраски,
учитывать при методологических подходах, на-
теряют мышечную массу и вес тела, накапли-
пример, при составлении выборки для получе-
вают мутации, демонстрируют снижение ло-
ния статистически значимых результатов, осо-
комоторной активности. С возрастом у ното-
бенно если тест проводится на старых рыбах, так
бранхиусов, как и у человека, синхронность
как к этому времени особи в группе могут значи-
календарного и биологического возраста всё
тельно разойтись по биологическому возрасту.
больше нарушается. То есть в одном календар-
Самцы теряют насыщенность окраски тела,
ном возрасте особи могут существенно отли-
узор на анальном и спинном плавнике бледнеет
чаться друг от друга по количеству и специфи-
и теряет чёткость. Некоторые исследователи ви-
ческому набору старческих признаков.
дят в этих процессах у нотобранхиусов парал-
Какие признаки стоит рассматривать при
лель с млекопитающими, которые с возрастом
разработке методов коррекции проявлений ста-
постепенно теряют пигмент волос и кожи [60].
рения на нотобранхиусах? Являясь модельными
На рис. 4 представлен самец N. guentheri на че-
организмами в геронтологии, нотобранхиусы
тырёх стадиях развития - до начала окрашива-
имеют ряд морфологических признаков, кото-
ния, в процессе приобретения окраски, в зре-
рые удобно использовать в качестве маркеров
лости и в старости.
старения. Статус этих маркеров и его измене-
У старых нотобранхиусов заметно наруша-
ния необходимо отслеживать в течение экспе-
ется целостность плавников, причём даже в ус-
римента. Оценка маркеров старения позволяет
ловиях индивидуального содержания, предот-
фиксировать динамику возраст-ассоциирован-
вращающего риск повреждения особями друг
БИОХИМИЯ том 87 вып. 12 2022
12*
1940
БУЛАВКИНА и др.
друга. Значительные нарушения претерпевает
и старением (mtor, 4ebp, s6k, insr, foxo3a/b, ampk
опорно-двигательная система - искривлением
и polg), аналогичная ассоциация была про-
позвоночника страдают как самцы, так и сам-
демонстрирована и для нескольких генов, ко-
ки. У некоторых особей образуется «горб», так-
дируемых митохондриальным геномом
[43].
же наблюдается истощение мышц, потеря веса.
Генетические локусы, ассоциированные с
Старым особям свойственны нарушения зри-
продолжительностью жизни, расположены
тельной системы. Довольно часто, особенно у
у N. furzeri на половой хромосоме и включают:
самок, образуются мутно-белые пятна, которые
31 белок-кодирующий ген, 6 генов длинных не-
могут расти и полностью «застилать» роговицу
кодирующих РНК и 2 гена малых ядерных РНК.
глаза подобно катаракте (рис. 5). У самцов чаще
Можно отметить ген GRN, кодирующий белок
отмечается необратимое постепенное уменьше-
програнулин, который вовлечён в нейроде-
ние зрачка до состояния едва заметной точки.
генеративные заболевания и регуляцию про-
С возрастом нотобранхиусы теряют спо-
должительности жизни у мышей, GSTT1A, ген
собность к регенерации тканей, например,
глутатион-S-трансфераз (окислительно-вос-
8-недельные N. furzeri (штамм MZM-0703) спо-
становительных ферментов гомеостаза, ко-
собны почти полностью регенерировать хво-
торые регулируют продолжительность жиз-
стовой плавник в течение 4 недель, а в возрасте
ни у червей и мышей), а также гены STAT3
54 недель - уже только 46% от первоначаль-
и STAT5.1
(2of6), кодирующие два фактора
ного размера плавника [61]. Подобно боль-
транскрипции, участвующие в регуляции вос-
шинству животных, рыбы рода Nothobranchius
палительных процессов [43, 67].
постепенно утрачивают репродуктивную спо-
Истощение теломер - типичный процесс
собность, которая у N. furzeri начинает сни-
старения клеток людей и многих других орга-
жаться уже с 8-10 недель жизни [58].
низмов, в том числе модельных [68-71], но он
Как и у человека, характерной чертой ста-
не является однозначным биомаркером старе-
рения нотобранхиусов является развитие опу-
ния нотобранхиусов. Было продемонстриро-
холевых новообразований. Опухоли, изученные
вано, что у долгоживущего штамма N. furzeri
в разных штаммах N. furzeri, имели нарушения
теломеры истощаются с возрастом, а у штам-
в таких белках, как Bcl-2, цитокератин-8, кар-
ма GRZ они почти не укорачиваются, возмож-
циноэмбриональный антиген и p53. Наиболее
но, из-за недолгой продолжительности жизни
подверженным новообразованиям органом у
этого штамма [72]. Однако линия N. furzeri с
нотобранхиусов является печень [62]. Вероят-
мутацией теломеразы продемонстрировала
ность возникновения неоплазий увеличивается
преждевременное бесплодие, резкое снижение
по мере старения рыбы [63], что наблюдается и у
количества красных и белых кровяных клеток,
людей [64]. Примечательно, что заболеваемость
аномалии в эпителиальных клетках кишеч-
раком печени у самцов N. furzeri выше, чем у са-
ника, включая снижение полярности и уве-
мок [62], это также повторяет статистику забо-
личение соотношения размеров ядер и цито-
леваемости раком печени у людей [41, 65].
плазмы [73]. По-видимому, теломераза играет
Перечисленные признаки старения ното-
ключевую роль в поддержании гомеостаза ор-
бранхиусов соответствуют некоторым из воз-
ганизма у N. furzeri, но не влияет на продолжи-
растных изменений, происходящих у челове-
тельность жизни штамма GRZ.
ка, мышей и других позвоночных [66]. Таким
У многих видов, включая человека, со ста-
образом, в арсенале исследователей имеется
рением коррелирует нестабильность митохон-
набор биомаркеров старения представителей
дриальной ДНК (мтДНК) [70, 74-77]. Причина
рода Nothobranchius, позволяющий изучать ме-
в том, что мтДНК не имеет гистонов, поэто-
ханизмы возраст-зависимых патологий раз-
му она в большей степени, чем ядерная ДНК,
личных систем органов, оценивать геропро-
подвержена мутагенезу [78]. Также наблюдает-
текторные эффекты тестируемых интервенций
ся снижение копийности мтДНК [74, 79], что
и отслеживать динамику возрастных измене-
приводит к раннему проявлению возраст-ассо-
ний неинвазивным способом, получая реле-
циированных заболеваний [80]. Данный про-
вантные результаты экспериментов на позво-
цесс коррелирует с возрастом во многих тканях
ночном животном за короткий срок.
N. furzeri, включая мозг, печень и мышцы [81].
Какие молекулярно-генетические нарушения
Возрастное понижение активности про-
нотобранхиусов наиболее перспективно изучать в
теасом, отвечающих за контроль качества бел-
аспекте старения? В 2015 году был отсеквениро-
ка, начинающееся после полового созревания,
ван и собран de novo геном N. furzeri, проведена
в итоге приводит к нарушению стехиометрии
аннотация [67]. Оказалось, что некоторые гены
белков при старении. Прогрессирующая утра-
были связаны с продолжительностью жизни
та нормального протеостаза считается одним
БИОХИМИЯ том 87 вып. 12 2022
МНОГОГРАННЫЙ НОТОБРАНХИУС
1941
из основных факторов нейродегенерации как
ЗАКЛЮЧЕНИЕ
у N. furzeri, так и у человека [82].
Для каких нозологических категорий ис-
Нотобранхиусы таят в себе ещё немало за-
пользование нотобранхиусов как модели возраст-
гадок, и в обзоре мы рассмотрели лишь наибо-
зависимых заболеваний наиболее оправдано,
лее актуальные вопросы, которые можно сфор-
обосновано и перспективно? При использова-
мулировать на существующем уровне развития
нии рыб рода Nothobranchius в качестве моде-
техники. С усовершенствованием методических
ли старения человека наибольшее внимание
подходов возникнут и новые, не менее интерес-
уделяется процессам нейродегенерации. На-
ные вопросы. Что же мешает двигаться вперед,
пример, было показано [4], что в глиальных
и какие методические сложности испытывают
клетках головного мозга N. furzeri с возрастом
исследователи при работе с нотобранхиусами?
повышается регуляция глиального фибрил-
Первый блок методических проблем за-
лярного кислого белка GFAP, как это проис-
трагивает особенности содержания животных
ходит при старении у млекопитающих. Из-за
и стандартизацию всех манипуляций с ними.
этого с возрастом накапливаются богатые ли-
Специалистам хорошо известно, что далеко не
пидами пигментные гранулы - липофусцин,
всегда даже опубликованные научные резуль-
что снижает поддержку и защиту нейронов,
таты уважаемых коллективов удаётся воспро-
обеспечиваемых глией. Вместе с этим дегене-
извести в других лабораториях. Важно макси-
рируют сами нейроны - в них накапливаются
мально внимательно относиться к унификации
молекулы бета-амилоида и агрегируют, обра-
процессов, причём в тесном сотрудничестве
зуя бляшки, наблюдаемые у людей с болезнью
с коллегами. Известно, например, что повы-
Альцгеймера, самой значимой и известной
шение температуры воды изменяет среднюю
нейродегенерацией у человека.
продолжительность жизни. Однако такие по-
Также у состарившихся N. furzeri наблюда-
пытки могут иметь и негативные последствия.
ется возраст-ассоциированная нейродегенера-
Создание протоколов, позволяющих избежать
ция норадренергических и дофаминергических
использование не стандартизируемых матери-
нейронов, что напоминает предсимптомную
алов, например торфа, привело к тому, что у
стадию болезни Паркинсона [83]. Данные пат-
лабораторных животных N. furzeri изменилось
терны старения нервной системы свидетель-
соотношение диапауз в процессе эмбриогене-
ствуют о том, что естественные генетические
за - они практически не входили в диапаузу III.
вариации способны влиять на восприимчи-
Вторая группа вопросов - создание воз-
вость дофаминергических нейронов N. furzeri
можности проводить малоинвазивные тесты.
и могут быть использованы для выявления мо-
Это могло бы обеспечить сбор биологического
дифицирующих факторов возрастной нейро-
материала для исследований, не только пост-
дегенерации.
мортально или подвергая организм сильному
Нарушение функций зрительной системы
стрессовому воздействию, и, таким образом,
является значимым признаком старения и про-
получать данные от одного и того же живот-
явлением многих нейродегенеративных забо-
ного в динамике. В настоящее время попыт-
леваний. Более того, были идентифицированы
ки использовать, например, фрагменты плав-
специфические маркеры нейродегенератив-
ников, приводят к тому, что часть животных
ных процессов в сетчатке старых N. furzeri [84].
необратимо становятся ползунами. Причём с
Рыбы рода Nothobranchius используются
возрастом доля таких случаев увеличивается, а
как модель остеопороза, ассоциированного со
плавники регенерируют всё хуже.
старением. Остеопороз - частое возрастное
И, наконец, одна из самых неразработан-
заболевание опорно-двигательной системы
ных в методическом плане областей - это ис-
человека. С возрастом в цитологическом про-
следования поведения нотобранхиусов. Ког-
филе и минеральном составе костей N. furzeri
нитивно-поведенческие тесты требуют от
наблюдаются не только возрастные, но и поло-
экспериментатора одновременно творческого
вые различия, которые могут быть приняты во
подхода, наблюдательности, педантичности и
внимание при изучении гендерных аспектов
огромного временного ресурса. Однако эта об-
возрастных заболеваний опорно-двигательно-
ласть является крайне привлекательной, потому
го аппарата [4].
что в ней есть ещё масса вопросов, на которые
Таким образом, представители рода
можно ответить, не дожидаясь появления но-
Nothobranchius - превосходные модели для те-
вых методов и приборов. Вот лишь некоторые
стирования не только потенциальных геро-
из них. В каких направлениях необходимо опти-
протекторов [17, 85], но возраст-ассоцииро-
мизировать когнитивно-поведенческие тесты, в
ванных патологий человека.
том числе для разных видов и в разном возрасте?
БИОХИМИЯ том 87 вып. 12 2022
1942
БУЛАВКИНА и др.
Что может быть положительным подкреплением
Е.В. Булавкина., М.А. Гончарова., А.И. Шувало-
при проведении когнитивно-поведенческих те-
ва - подготовка иллюстраций. Все авторы уча-
стов? То есть как построить эксперимент, не ис-
ствовали в анализе литературы и написании
пользуя лишь отрицательное подкрепление, что,
обзора.
несомненно, вызывает у особи стресс? Как при
Финансирование. Работа выполнена при
проведении экспериментов решить проблему
поддержке Министерства науки и высшего обра-
влияния разного иерархического статуса тести-
зования РФ, Соглашение № 075-15-2019-1660.
руемых особей? Или же, наоборот, как постро-
Конфликт интересов. Авторы заявляют об
ить эксперимент, чтобы учитывать этот статус?
отсутствии конфликта интересов.
Данный обзор, подчёркивающий потенци-
Соблюдение этических норм. Все проце-
ал модельных рыб рода Nothobranchius, призван
дуры, выполненные в исследованиях с уча-
стимулировать применение данной модели в
стием животных, соответствовали этиче-
борьбе за активное долголетие.
ским стандартам учреждения, в котором
проводились исследования, и утверждён-
Вклад авторов. А.В. Кудрявцева, Е.В. Бу-
ным правовым актам РФ и международных
лавкина - концепция и руководство работой; организаций.
СПИСОК ЛИТЕРАТУРЫ
1.
Cellerino, A., Valenzano, D. R., and Reichard, M.
tseva, A., Shaposhnikov, M., and Moskalev, A. (2015)
(2016) From the bush to the bench: the annual
Effect of low doses (5-40 cGy) of gamma-irradiation
Nothobranchius fishes as a new model system in
on lifespan and stress-related genes expression profile
biology, Biol. Rev., 91, 511-533, doi: 10.1111/brv.12183.
in Drosophila melanogaster, PLoS One, 10, e0133840,
2.
Hu, C.-K., and Brunet, A.
(2018) The African
doi: 10.1371/journal.pone.0133840.
turquoise killifish: a research organism to study
10.
Proshkina, E. N., Shaposhnikov, M. V., Sadritdi-
vertebrate aging and diapause, Aging Cell, 17, e12757,
nova, A. F., Kudryavtseva, A. V., and Moskalev, A. A.
doi: 10.1111/acel.12757.
(2015) Basic mechanisms of longevity: A case study of
3.
Poeschla, M., and Valenzano, D. R.
(2020)
Drosophila pro-longevity genes, Ageing Res. Rev., 24,
The turquoise killifish: a genetically tractable model
218-231, doi: 10.1016/j.arr.2015.08.005.
for the study of aging, J. Exp. Biol., 223, jeb209296,
11.
Proshkina, E., Lashmanova, E., Dobrovolskaya, E.,
doi: 10.1242/jeb.209296.
Zemskaya, N., Kudryavtseva, A., Shaposhnikov, M.,
4.
Butylina, M., Föger-Samwald, U., Gamsjaeger, S.,
and Moskalev, A.
(2016) Geroprotective and
Wahl-Figlash, K., Kothmayer, M., Paschalis, E. P.,
radioprotective activity of quercetin, (-)-epicatechin,
Pusch, O., and Pietschmann, P.
(2022) Notho-
and ibuprofen in Drosophila melanogaster, Front.
branchius furzeri, the turquoise killifish: a model
Pharmacol., 7, doi: 10.3389/fphar.2016.00505.
of age-related osteoporosis? Gerontology,
1-13,
12.
Moskalev, A., Shaposhnikov, M., Proshkina, E.,
doi: 10.1159/000524300.
Belyi, A., Fedintsev, A., Zhikrivetskaya, S., Guvatova, Z.,
5.
Dance, A. (2016) Live fast, die young, Nature, 535,
Sadritdinova, A., Snezhkina, A., Krasnov, G., and
453-455, doi: 10.1038/535453a.
Kudryavtseva, A.
(2016) The influence of pro-
6.
Valdesalici, S., and Cellerino, A. (2003) Extremely
longevity gene Gclc overexpression on the age-
short lifespan in the annual fish Nothobranchius furzeri,
dependent changes in Drosophila transcriptome
Proc. Biol. Sci., 270, doi: 10.1098/rsbl.2003.0048.
and biological functions, BMC Genomics, 17, 1046,
7.
Dong, Y., Cui, P., Li, Z., and Zhang, S. (2017) Aging
doi: 10.1186/s12864-016-3356-0.
asymmetry: systematic survey of changes in age-
13.
Lashmanova, E., Zemskaya, N., Proshkina, E.,
related biomarkers in the annual fish Nothobranchius
Kudryavtseva, A., Volosnikova, M., Marusich, E.,
guentheri, Fish Physiol. Biochem.,
43,
309-319,
Leonov, S., Zhavoronkov, A., and Moskalev, A.
doi: 10.1007/s10695-016-0288-1.
(2017) The evaluation of geroprotective effects of
8.
Li, C., Song, L., Zhou, Y., Yuan, J., and Zhang, S.
selected flavonoids in Drosophila melanogaster and
(2022) Identification of Isthmin1 in the small annual
Caenorhabditis elegans, Front. Pharmacol., 8, 884,
fish, Nothobranchius guentheri, as a novel biomarker
doi: 10.3389/fphar.2017.00884.
of aging and its potential rejuvenation activity,
14.
Yuan, R., Tsaih, S. W., Petkova, S. B., Marin de
Biogerontology, 23, 99-114, doi: 10.1007/s10522-021-
Evsikova, C., Xing, S., Marion, M. A., Bogue, M. A.,
09948-5.
Mills, K. D., Peters, L. L., Bult, C. J., Rosen, C. J.,
9.
Zhikrevetskaya, S., Peregudova, D., Danilov, A.,
Sundberg, J. P., Harrison, D. E., Churchill, G. A.,
Plyusnina, E., Krasnov, G., Dmitriev, A., Kudryav-
and Paigen, B. (2009) Aging in inbred strains of mice:
БИОХИМИЯ том 87 вып. 12 2022
МНОГОГРАННЫЙ НОТОБРАНХИУС
1943
study design and interim report on median lifespans
24.
Lambert, J. W., Reichard, M., and Pincheira-
and circulating IGF1 levels: median lifespans and
Donoso, D. (2019) Live fast, diversify non-adaptively:
IGF1 levels of 31 inbred strains, Aging Cell, 8, 277-287,
evolutionary diversification of exceptionally short-
doi: 10.1111/j.1474-9726.2009.00478.x.
lived annual killifishes, BMC Evol. Biol., 19, 10,
15.
Seidel, J., and Valenzano, D. R. (2018) The role of the
doi: 10.1186/s12862-019-1344-0.
gut microbiome during host ageing, F1000Research, 7,
25.
Costa, W. J. E. M.
(2017) Taxonomic revision
1086, doi: 10.12688/f1000research.15121.1.
of the seasonal killifish genus Nothobranchius
16.
Smith, P., Willemsen, D., Popkes, M., Metge, F.,
from Zanzibar, East Africa (Cyprinodontoidei:
Gandiwa, E., Reichard, M., and Valenzano, D. R.
Aplocheilidae), J. Nat. Hist.,
51,
1609-1624,
(2017) Regulation of life span by the gut microbiota
doi: 10.1080/00222933.2017.1330976.
in the short-lived African turquoise killifish, eLife, 6,
26.
Watters, B. R., Nagy, B., van der Merwe, P. D. W.,
e27014, doi: 10.7554/eLife.27014.
Cotterill, F. P. D., and Bellstedt, D. U. (2022) Review
17.
Bakhtogarimov, I. R., Kudryavtseva, A. V., Krasnov, G. S.,
of the Nothobranchius taeniopygus species group from
Gladysh, N. S., Volodin, V. V., Kudryavtsev, A. A.,
central and western Tanzania with descriptions of
Bulavkina, E. V., Goncharova, M. A., Ledyaeva, V. S.,
five new species and redescription of Nothobranchius
Pastukhov, I. S., Vershinina, Y. S., Starkova, A. M.,
taeniopygus (Teleostei: Nothobranchiidae), DE: Verlag
Snezhkina, A. V., Shuvalova, A. I., Pavlov, V. S.,
Dr. Friedrich Pfeil, 2019. Accessed: Oct. 27, 2022,
Nikiforov-Nikishin, D. L., Moskalev, A. A., and
doi: 10.23788/IEF-1110.
Guvatova, Z. G. (2022) The effect of meclofenoxate
27.
Nagy, B., Watters, B. R., and Raspopova, A. A.
on the transcriptome of aging brain of Nothobranchius
(2021) Nothobranchius nikiforovi, a new species of
guentheri annual killifish, Int. J. Mol. Sci., 23, 2491,
seasonal killifish from the lower Matandu drainage in
doi: 10.3390/ijms23052491.
south-eastern coastal Tanzania (Cyprinodontiformes:
18.
Guvatova, Z. G., Fedorova, M. S., Vershinina, Y. S., Pu-
Nothobranchiidae), Zootaxa,
4950,
103-122,
dova, E. A., Lipatova, A. V., Volodin, V. V., Gladysh, N. S.,
doi: 10.11646/zootaxa.4950.1.5.
Tokarev, A. T., Kornev, A. B., Pavlov, V. S., Bakhtoga-
28.
Shidlovskiy, K. M., Watters, B. R., and Wildekamp, R. H.
rimov, I. R., Krysanov, E. Y., Moskalev, A. A., Kras-
(2019) Notes on the annual killifish species Notho-
nov, G. S., and Kudryavtseva, A. V. (2021) De novo
branchius rachovii (Cyprinodontiformes; Notho-
transcriptome profiling of brain tissue from the annual
branchiidae) with the description of two new species,
killifish Nothobranchius guentheri, Life,
11,
137,
Zootaxa, 2724, 37, doi: 10.11646/zootaxa.2724.1.3.
doi: 10.3390/life11020137.
29.
Nagy, B. (2018) Nothobranchius ditte, a new species
19.
Gao, X., Cai, T., Lin, Y., Zhu, R., Hao, W., Guo, S.,
of annual killifish from the Lake Mweru basin in the
and Hu, G. (2022) The function of glucose metabolism
Democratic Republic of the Congo (Teleostei: Notho-
in embryonic diapause of annual killifish, Comp.
branchiidae), Ichthyol. Explor. Freshw., 28, 115-134.
Biochem. Physiol. Part D Genomics Proteomics, 42,
30.
Arai, R. (2011) Fish Karyotypes, Tokyo, Springer
100965, doi: 10.1016/j.cbd.2022.100965.
Japan, doi: 10.1007/978-4-431-53877-6.
20.
Romney, A. L., and Podrabsky, J. E. (2017) Tran-
31.
Navarro, A., and Barton, N. H. (2003) Chromosomal
scriptomic analysis of maternally provisioned cues
speciation and molecular divergence - accelerated
for phenotypic plasticity in the annual killifish,
evolution in rearranged chromosomes, Science, 300,
Austrofundulus limnaeus, EvoDevo, 8, 6, doi: 10.1186/
321-324, doi: 10.1126/science.1080600.
s13227-017-0069-7.
32.
Kirkpatrick, M., and Barton, N. (2006) Chromosome
21.
Romney, A. L. T., Davis, E. M., Corona, M. M.,
inversions, local adaptation and speciation, Genetics,
Wagner, J. T., and Podrabsky, J. E. (2018) Temperature-
173, 419-434, doi: 10.1534/genetics.105.047985.
dependent vitamin D signaling regulates developmen-
33.
Noor, M. A. F., Grams, K. L., Bertucci, L. A., and
tal trajectory associated with diapause in an annual
Reiland, J.
(2001) Chromosomal inversions and
killifish, Proc. Natl. Acad. Sci. USA, 115, 12763-12768,
the reproductive isolation of species, Proc. Natl.
doi: 10.1073/pnas.1804590115.
Acad. Sci. USA,
98,
12084-12088, doi:
10.1073/
22.
Van der Merwe, P.W., Cotterill, F. P. D., Kandziora, M.,
pnas.221274498.
Watters, B. R., Nagy, B., Genade, T., Flügel, T. J.,
34.
Rieseberg, L. H. (2001) Chromosomal rearrangements
Svendsen, D. S., and Bellstedt, D. U. (2021) Genomic
and speciation, Trends Ecol. Evol.,
16,
351-358,
fingerprints of palaeogeographic history: The tempo and
doi: 10.1016/S0169-5347(01)02187-5.
mode of rift tectonics across tropical Africa has shaped
35.
Krysanov, E., and Demidova, T. (2018) Extensive
the diversification of the killifish genus Nothobranchius
karyotype variability of African fish genus Notho-
(Teleostei: Cyprinodontiformes), Mol. Phylogenet.
branchius (Cyprinodontiformes), Comp. Cytogenet.,
Evol., 158, 106988, doi: 10.1016/j.ympev.2020.106988.
12, 387-402, doi: 10.3897/CompCytogen.v12i3.25092.
23.
Whitlock, M. C., and Phillips, P. C. (2014) Drift:
36.
Krysanov, E., Demidova, T., and Nagy, B. (2016)
Introduction, in eLS, 1st ed., John Wiley & Sons, Ltd,
Divergent karyotypes of the annual killifish genus Notho-
doi: 10.1002/9780470015902.a0001698.pub2.
branchius (Cyprinodontiformes, Nothobranchiidae),
БИОХИМИЯ том 87 вып. 12 2022
1
944
БУЛАВКИНА и др.
Comp. Cytogenet.,
10,
439-445, doi:
10.3897/
populations, Ecol. Evol., 11, 4945-4956, doi: 10.1002/
CompCytogen.v10i3.9863.
ece3.7402.
37.
Dorn, A., Musilová, Z., Platzer, M., Reichwald, K.,
48.
Duerr, J. M., and Podrabsky, J. E. (2010) Mitochon-
and Cellerino, A. (2014) The strange case of East
drial physiology of diapausing and developing embryos
African annual fishes: aridification correlates with
of the annual killifish Austrofundulus limnaeus: impli-
diversification for a savannah aquatic group? BMC
cations for extreme anoxia tolerance, J. Comp. Physi-
Evol. Biol., 14, 210, doi: 10.1186/s12862-014-0210-3.
ol. B, 180, 991-1003, doi: 10.1007/s00360-010-0478-6.
38.
Ewulonu, U. K., Haas, R., and Turner, B. J. (1985)
49.
Podrabsky, J. E., Garrett, I. D. F., and Kohl, Z. F.
A multiple sex chromosome system in the annual
(2010) Alternative developmental pathways associated
killifish, Nothobranchius guentheri, Copeia, 1985, 503,
with diapause regulated by temperature and maternal
doi: 10.2307/1444868.
influences in embryos of the annual killifish
39.
Kitano, J., and Peichel, C. L. (2012) Turnover of sex
Austrofundulus limnaeus, J. Exp. Biol., 213, 3280-3288,
chromosomes and speciation in fishes, Environ. Biol.
doi: 10.1242/jeb.045906.
Fishes, 94, 549-558, doi: 10.1007/s10641-011-9853-8.
50.
Matias, J. R., and Adrias, A. Q. (2010) The use of
40.
Costa, W. J. E. M. (2018) Comparative morphology,
annual killifish in the biocontrol of the aquatic stages
phylogeny
and
classification
of
African
of mosquitoes in temporary bodies of fresh water; a
seasonal killifishes of the tribe Nothobranchiini
potential new tool in vector control, Parasit. Vectors, 3,
(Cyprinodontiformes: Aplocheilidae), Zool. J. Linn.
46, doi: 10.1186/1756-3305-3-46.
Soc., 184, 115-135, doi: 10.1093/zoolinnean/zlx102.
51.
Briffa, M., and Weiss, A. (2010) Animal personality,
41.
Reichwald, K., Petzold, A., Koch, P., Downie, B. R.,
Curr. Biol.,
20, R912-R914, doi:
10.1016/
Hartmann,N., Pietsch, S., Baumgart,M., Chalopin,D.,
j.cub.2010.09.019.
Felder, M., Bens, M., Sahm, A., Szafranski, K.,
52.
Biro, P. A., Adriaenssens, B., and Sampson, P.
Taudien, S., Groth, M., Arisi, I., Weise, A., Bhatt, S. S.,
(2014) Individual and sex-specific differences in
Sharma, V., Kraus, J. M., Schmid, F., Priebe, S.,
intrinsic growth rate covary with consistent individual
Liehr, T., Görlach, M., Than, M. E., Hiller, M.,
differences in behaviour, J. Anim. Ecol., 83, 1186-1195,
Kestler, H. A., Volff, J. N., Schartl, M., Cellerino, A.,
doi: 10.1111/1365-2656.12210.
Englert, C., and Platzer, M. (2015) Insights into sex
53.
Budaev, S., and Brown, C. (2011) Personality Traits
chromosome evolution and aging from the genome of
and Behaviour, in Fish Cognition and Behavior (Brown,
a short-lived fish, Cell, 163, 1527-1538, doi: 10.1016/
C., Laland, K., and Krause, J., eds) 1st Edn., Wiley,
j.cell.2015.10.071.
pp. 135-165, doi: 10.1002/9781444342536.ch7.
42.
Nikiforov-Nikishin, A., Smorodinskaya, S., Kochet-
54.
Thoré, E. S. J., Steenaerts, L., Philippe, C., Grégoir, A.,
kov, N., Nikiforov-Nikishin, D., Danilenko, V.,
Brendonck, L., and Pinceel, T. (2018) Individual
Bugaev, O., Vatlin, A., Abrosimova, N., Antipov, S.,
behavioral variation reflects personality divergence in
Kudryavtsev, A., and Klimov, V. (2022) Effects of
the upcoming model organism Nothobranchius furzeri,
three feed additives on the culturable microbiota
Ecol. Evol., 8, 8448-8457, doi: 10.1002/ece3.4356.
composition and histology of the anterior and posterior
55.
Jolles, J. W., King, A. J., and Killen, S. S. (2020)
intestines of Zebrafish (Danio rerio), Animals, 12,
The role of individual heterogeneity in collective
2424, doi: 10.3390/ani12182424.
animal behaviour, Trends Ecol. Evol., 35, 278-291,
43.
Cui, R., Willemsen, D., and Valenzano, D. R.
doi: 10.1016/j.tree.2019.11.001.
(2020) Nothobranchius furzeri (African Turquoise
56.
Nikiforov-Nikishin, D. L., Kochetkov, N. I.,
Killifish), Trends Genet., 36, 540-541, doi: 10.1016/
Mikodina, E. V., Nikiforov-Nikishin, A. L., Si-
j.tig.2020.01.012.
makov, Y. G., Golovacheva, N. A., Gorbunov, A. V.,
44.
Terzibasi Tozzini, E., and Cellerino, A.
(2020)
Chebotarev, S. N., Kirichenko, E. Y., Zabiyaka, I. Y.,
Nothobranchius annual killifishes, EvoDevo, 11, 25,
Pastukhov, I. S., and Bren, A. B. (2022) Evaluation
doi: 10.1186/s13227-020-00170-x.
of age-dependent changes in the coloration of
45.
Sahm, A., Platzer, M., and Cellerino, A.
(2016)
male killifish Nothobranchius guentheri using
Outgroups and positive selection: the Nothobranchius
new photoprocessing methods, Biology,
11,
205,
furzeri case, Trends Genet., 32, 523-525, doi: 10.1016/
doi: 10.3390/biology11020205.
j.tig.2016.06.002.
57.
McMahon, S. J., Munday, P. L., Wong, M. Y. L.,
46.
Mustafin, R. N., and Khusnutdinova, E. K. (2019)
and Donelson, J. M. (2019) Elevated CO2 and food
The role of transposable elements in the ecological
ration affect growth but not the size-based hierarchy
morphogenesis under the influence of stress, Vavilov
of a reef fish, Sci. Rep., 9, 19706, doi: 10.1038/
J. Genet. Breed, 23, 380-389, doi: 10.18699/VJ19.506.
s41598-019-56002-z.
47.
Polačik, M., Vrtílek, M., Reichard, M., Žák, J.,
58.
Blažek, R., Polačik, M., and Reichard, M. (2013)
Blažek, R., and Podrabsky, J.
(2021) Embryo
Rapid growth, early maturation and short generation
ecology: Developmental synchrony and asynchrony
time in African annual fishes, EvoDevo,
4,
24,
in the embryonic development of wild annual fish
doi: 10.1186/2041-9139-4-24.
БИОХИМИЯ том 87 вып. 12 2022
МНОГОГРАННЫЙ НОТОБРАНХИУС
1945
59.
Fonseca, A. P. da, Volcan, M. V., Romano, L. A., and
locus, Proc. Natl. Acad. Sci. USA, 95, 8648-8653,
Robaldo, R. B. (2018) Metaplasia in swim bladder ep-
doi: 10.1073/pnas.95.15.8648.
ithelium of the endangered annual fish Austrolebias
72.
Hartmann, N., Reichwald, K., Lechel, A., Graf, M.,
nigrofasciatus (Cyprinodontiformes: Rivulidae) results
Kirschner, J., Dorn, A., Terzibasi, E., Wellner, J.,
in inadequate swimming and delayed growth, Neotrop-
Platzer, M., Rudolph, K. L., Cellerino, A., and
ical Ichthyol., 16, doi: 10.1590/1982-0224-20170038.
Englert, C.
(2009) Telomeres shorten while Tert
60.
Geyfman, M., and Andersen, B.
(2010) Clock
expression increases during ageing of the short-lived
genes, hair growth and aging, Aging, 2, 122-128,
fish Nothobranchius furzeri, Mech. Ageing Dev., 130,
doi: 10.18632/aging.100130.
290-296, doi: 10.1016/j.mad.2009.01.003.
61.
Wendler, S., Hartmann, N., Hoppe, B., and Englert, C.
73.
Harel, I., Benayoun, B. A., Machado, B., Singh, P. P.,
(2015) Age-dependent decline in fin regenerative
Hu, C. K., Pech, M. F., Valenzano, D. R., Zhang, E.,
capacity in the short-lived fish Nothobranchius furzeri,
Sharp, S. C., Artandi, S. E., and Brunet, A. (2015)
Aging Cell, 14, 857-866, doi: 10.1111/acel.12367.
A platform for rapid exploration of aging and diseases
62.
Di Cicco, E., Tozzini, E. T., Rossi, G., and
in a naturally short-lived vertebrate, Cell, 160, 1013-
Cellerino, A.
(2011) The short-lived annual fish
1026, doi: 10.1016/j.cell.2015.01.038.
Nothobranchius furzeri shows a typical teleost aging
74.
Barazzoni, R., Short, K. R., and Nair, K. S. (2000)
process reinforced by high incidence of age-dependent
Effects of aging on mitochondrial DNA copy number
neoplasias, Exp. Gerontol., 46, 249-256, doi: 10.1016/
and cytochromec oxidase gene expression in rat
j.exger.2010.10.011.
skeletal muscle, liver, and heart, J. Biol. Chem., 275,
63.
Pompei, F., Polkanov, M., and Wilson, R. (2001)
3343-3347, doi: 10.1074/jbc.275.5.3343.
Age distribution of cancer in mice: the incidence
75.
Tauchi, H., and Sato, T. (1968) Age changes in size
turnover at old age, Toxicol. Ind. Health, 17, 7-16,
and number of mitochondria of human hepatic cells,
doi: 10.1191/0748233701th091oa.
J. Gerontol., 23, 454-461, doi: 10.1093/geronj/23.4.454.
64.
De Magalhães, J. P. (2013) How ageing processes
76.
Yen, T.-C., Chen, Y.-S., King, K.-L., Yeh, S.-H.,
influence cancer, Nat. Rev. Cancer, 13, 357-365,
and Wei, Y.-H. (1989) Liver mitochondrial respiratory
doi: 10.1038/nrc3497.
functions decline with age, Biochem. Biophys.
65.
Nordenstedt, H., White, D. L., and El-Serag, H. B.
Res. Commun., 165, 994-1003, doi: 10.1016/0006-
(2010) The changing pattern of epidemiology in
291X(89)92701-0.
hepatocellular carcinoma, Dig. Liver Dis.,
42,
77.
Yui, R., Ohno, Y., and Matsuura, E. T.
(2003)
S206-S214, doi: 10.1016/S1590-8658(10)60507-5.
Accumulation of deleted mitochondrial DNA in aging
66.
Vanhooren, V., and Libert, C. (2013) The mouse
Drosophila melanogaster, Genes Genet. Syst., 78, 245-
as a model organism in aging research: Usefulness,
251, doi: 10.1266/ggs.78.245.
pitfalls and possibilities, Ageing Res. Rev., 12, 8-21,
78.
Tatarenkov, A., and Avise, J. C. (2007) Rapid concerted
doi: 10.1016/j.arr.2012.03.010.
evolution in animal mitochondrial DNA, Proc. Biol.
67.
Valenzano, D. R., Benayoun, B. A., Singh, P. P.,
Sci., 274, 1795-1798, doi: 10.1098/rspb.2007.0169.
Zhang, E., Etter, P. D., Hu, C. K., Clément-Ziza, M.,
79.
Bratic, A., and Larsson, N.-G. (2013) The role of
Willemsen, D., Cui, R., Harel, I., Machado, B. E.,
mitochondria in aging, J. Clin. Invest., 123, 951-957,
Yee, M. C., Sharp, S. C., Bustamante, C. D., Beyer, A.,
doi: 10.1172/JCI64125.
Johnson, E. A., and Brunet, A. (2015) The African
80.
Vermulst, M., Wanagat, J., Kujoth, G. C., Bielas, J. H.,
Turquoise killifish genome provides insights into
Rabinovitch, P. S., Prolla, T. A., and Loeb, L. A.
evolution and genetic architecture of lifespan, Cell,
(2008) DNA deletions and clonal mutations drive
163, 1539-1554, doi: 10.1016/j.cell.2015.11.008.
premature aging in mitochondrial mutator mice, Nat.
68.
Benetos, A., Okuda, K., Lajemi, M., Kimura, M.,
Genet., 40, 392-394, doi: 10.1038/ng.95.
Thomas, F., Skurnick, J., Labat, C., Bean, K., and
81.
Hartmann, N., Reichwald, K., Wittig, I., Dröse, S.,
Aviv, A. (2001) Telomere length as an indicator of
Schmeisser, S., Lück, C., Hahn, C., Graf, M.,
biological aging: the gender effect and relation with
Gausmann, U., Terzibasi, E., Cellerino, A., Ristow, M.,
pulse pressure and pulse wave velocity, Hypertension,
Brandt, U., Platzer, M., and Englert, C.
(2011)
37, 381-385, doi: 10.1161/01.hyp.37.2.381.
Mitochondrial DNA copy number and function
69.
Harley, C. B., Futcher, A. B., and Greider, C. W. (1990)
decrease with age in the short-lived fish Nothobranchius
Telomeres shorten during ageing of human fibroblasts,
furzeri: decline of mitochondrial function in
Nature, 345, 458-460, doi: 10.1038/345458a0.
aging fish, Aging Cell, 10, 824-831, doi: 10.1111/
70.
López-Otín, C., Blasco,M. A., Partridge, L., Serrano,M.,
j.1474-9726.2011.00723.x.
and Kroemer, G. (2013) The hallmarks of aging, Cell,
82.
Kelmer Sacramento, E., Kirkpatrick, J. M., Mazzetto, M.,
153, 1194-1217, doi: 10.1016/j.cell.2013.05.039.
Baumgart, M., Bartolome, A., Di Sanzo, S., Caterino, C.,
71.
Zhu, L., Hathcock, K. S., Hande, P., Lansdorp, P. M.,
Sanguanini, M., Papaevgeniou, N., Lefaki, M.,
Seldin, M. F., and Hodes, R. J. (1998) Telomere length
Childs, D., Bagnoli, S., Terzibasi Tozzini, E., Di
regulation in mice is linked to a novel chromosome
Fraia, D., Romanov, N., Sudmant, P. H., Huber, W.,
БИОХИМИЯ том 87 вып. 12 2022
1946
БУЛАВКИНА и др.
Chondrogianni, N., Vendruscolo, M., Cellerino, A.,
84. Vanhunsel, S., Bergmans, S., Beckers, A., Etienne, I.,
and Ori, A. (2020) Reduced proteasome activity in
Van Houcke, J., Seuntjens, E., Arckens, L., De Groef, L.,
the aging brain results in ribosome stoichiometry loss
and Moons, L. (2021) The killifish visual system as an
and aggregation, Mol. Syst. Biol., 16, doi: 10.15252/
in vivo model to study brain aging and rejuvenation, Npj
msb.20209596.
Aging Mech. Dis., 7, 22, doi: 10.1038/s41514-021-00077-4.
83. Bagnoli, S., Fronte, B., Bibbiani, C., Terzibasi
85. Valenzano, D. R., Terzibasi, E., Genade, T., Cattaneo, A.,
Tozzini, E., and Cellerino, A. (2022) Quantification
Domenici, L., and Cellerino, A. (2006) Resveratrol
of noradrenergic-, dopaminergic-, and tectal-neurons
prolongs lifespan and retards the onset of age-related
during aging in the short-lived killifish Nothobranchius
markers in a short-lived vertebrate, Curr. Biol., 16,
furzeri, Aging Cell, 21, doi: 10.1111/acel.13689.
296-300, doi: 10.1016/j.cub.2005.12.038.
MULTIFACETED NOTHOBRANCHIUS
Review
E. V. Bulavkina1,2, A. A. Kudryavtsev2, M. A. Goncharova2, M. S. Lantsova2,
A. I. Shuvalova2, M. A. Kovalev2, and A. V. Kudryavtseva1,2*
1 Center for Precision Genome Editing and Genetic Technologies for Biomedicine,
Engelhardt Institute of Molecular Biology, Russian Academy of Sciences,
119991 Moscow, Russia; E-mail: rhizamoeba@mail.ru
2 Postgenomic research laboratory, Engelhardt Institute of Molecular Biology,
Russian Academy of Sciences, 119991 Moscow, Russia
Annual killifish of the genus Nothobranchius are seeing a rapid increase in scientific interest over the years.
A variety of aspects surrounding egg-laying Cyprinodontiformes is being extensively studied, including their
aging. Inhabiting the drying water bodies of Africa rarely allows surviving through more than one rainy season
for Nothobranchius populations. Therefore, there was no lifespan-related bias in natural selection, which has
ultimately led to the reparation system decreasing in efficiency. The aging of Nothobranchius is studied both
in normal conditions and under the influence of potential geroprotectors, as well as genetic modifications.
Most biogerontological studies are conducted using the species Nothobranchius furzeri (GRZ isolate), which
has a lifespan of 3 to 7 months. However, the list of model species of Nothobranchius is considerably wider,
and the range of advanced research areas with their participation extends far beyond gerontology. Thus,
the review paper summarizes the most interesting and promising topics developing due to fish of the genus
Nothobranchius. Both classical studies related to lifespan control and rather new ones are touched upon,
including mechanisms of diapause, challenges of systematics and phylogeny, evolution of sex determination
mechanisms, changes in chromosome count, occurrence of multiple repeated DNA sequences in genome,
cognitive and behavioral features and social stratification, as well as methodological difficulties in working
with Nothobranchius.
Keywords: aging, animal models of aging, Nothobranchius, age related diseases, diapause, neurodegeneration, lifespan,
longevity, speciation, karyotype, stress
БИОХИМИЯ том 87 вып. 12 2022