БИОХИМИЯ, 2022, том 87, вып. 12, с. 2019 - 2032
ГИПОТЕЗА
УДК 577.24
ПЛАНЕТАРНЫЙ МЕТРОНОМ КАК РЕГУЛЯТОР
ПРОДОЛЖИТЕЛЬНОСТИ ЖИЗНИ И ТЕМПА СТАРЕНИЯ:
МЕТРОНОМНАЯ ГИПОТЕЗА
© 2022
А.М. Оловников
Институт биохимической физики им. Н.М. Эмануэля РАН,
119334 Москва, Россия; электронная почта: olovnikov@gmail.com
Поступила в редакцию 11.11.2022
После доработки 16.11.2022
Принята к публикации 16.11.2022
Предложен метрономный механизм управления длительностью периодов онтогенетического цик-
ла животного организма. Метрономная система состоит из серии компонентов. В их числе: вен-
трикулярная система мозга, планета Земля как участница генерации метрономных сигналов и тем-
поральная ДНК (тДНК) как субстрат, эпигенетически маркируемый для учета истекшего времени
онтогенеза. Метрономная система генерирует повторяющиеся сигналы в виде гидродинамических
возмущений в спинномозговой жидкости (СМЖ). Метрономный эффект возникает благодаря нало-
жению двух процессов - пристеночного однонаправленного потока СМЖ и осцилляций в движени-
ях планеты. Гидродинамические удары метронома трансформируются в нервные импульсы, которые
инициируют акты эпигенетической модификации тДНК в нейронах, меняя содержание факторов,
экспрессируемых этой ДНК для иннервируемых мишеней организма. Продолжительность периодов
онтогенетического цикла, включая продолжительность взрослой жизни, зависит от темпа актов мо-
дификации тДНК. Этот темп зависит главным образом от частоты метрономных сигналов, исполь-
зуемых каждым конкретным биологическим видом. Но на эти эпигенетические акты могут также
влиять факторы, способные модулировать метаболизм и темп модификаций хроматина, например
калорийно ограниченная диета и др.
КЛЮЧЕВЫЕ СЛОВА: онтогенез, старение, теории старения, вентрикулярная система мозга, СМЖ-кон-
тактирующие нейроны, гипоталамус, движения Земли, нутации, полярное движение.
DOI: 10.31857/S0320972522120181, EDN: NIKQYE
ВВЕДЕНИЕ
парадигму, которая бы революционизировала
эту область. Подчеркивая необходимость уни-
Теории процесса старения делятся на два
версальной концепции старения, Gladyshev [11]
основных класса - стохастические и программ-
сравнивает ее потенциальную будущую роль со
ные [1-4]. Накапливается все больше данных в
значением таких идей, как «роль протонного
пользу запрограммированности процесса, вы-
градиента в синтезе ATP, периодическая таб-
зывающего старение. Ведущая к старению пред-
лица химических элементов и эволюция путем
полагаемая программа онтогенеза исторически
естественного отбора».
восходит к Weismann [5]. Она обсуждается уже
Высказывалась, впрочем, уверенность, что
более сотни лет [6], но продолжают появляться ее
нужная парадигма уже существует, и что ею
новые варианты, как и альтернативные подходы
является идея гиперфункции [12, 13]. Однако,
к проблемам эволюционной геронтологии [7-9].
согласно альтернативному мнению, ни идея
Несмотря на несомненные достижения био-
гиперфункции, ни другие теории пока не пред-
геронтологии, некоторые исследователи оце-
ложили той интегративной концепции, которая
нивают ее текущее состояние как кризисное.
может помочь поставить правильные вопросы
В частности, Gems и de Magalhães [10] указывают
для дальнейшего продвижения в изучении ста-
на настоятельную необходимость искать новую рения [11]. К аналогичному заключению прихо-
дят и другие авторы, подчеркивая отсутствие це-
лостного понимания взаимодействий генетики
Принятые сокращения: СМЖ - спинномозговая жид-
кость; ПЖ - продолжительность жизни; тДНК - темпо-
и окружающей среды в объяснении старения и
ральная ДНК.
продолжительности жизни (ПЖ) [14].
2019
17*
2020
ОЛОВНИКОВ
В любом случае успехи, достигнутые в
ция. Организм многоклеточных животных не
идентификации и изучении индикаторов ста-
способен самостоятельно, без использования
рения (нестабильность генома, укорочение
внешнего независимого источника, обеспе-
теломер, эпигенетические изменения, потеря
чить необходимую продолжительность време-
протеостаза, нарушение чувствительности к
ни разных периодов своего онтогенетическо-
питательным веществам, митохондриальная
го цикла. Для этого он нуждается в помощи
дисфункция, клеточное старение, истощение
внешнего источника. В качестве такового, как
стволовых клеток и изменение межклеточной
я полагаю, может служить сама планета Земля
коммуникации) уже позволяют заглянуть за го-
с ее неравномерными движениями в космиче-
ризонт [15]. Тот факт, что появление признаков
ском пространстве.
старения упорядочено во времени, по-видимо-
Однако для детекции темпоральных под-
му, ясно говорит нам, что за этим стоит работа
сказок планеты животные должны были создать
программы. Но какой программы?
особую физиологическую систему, обозначен-
К списку иерархически упорядоченных
ную здесь как метрономная, и расположить ее
показателей старения, иллюстрирующих ги-
в желудочках мозга. Вполне возможно, что и
перфункциональную активность стареющего
создание самой центральной нервной системы
человека, в частности, были отнесены следую-
на основе нервной трубки, и появление желу-
щие: гипертония, гиперлипидемия, гиперин-
дочков мозга в эволюции в значительной мере
сулинемия, гипергликемия, провоспаление,
было подчинено этой центральной задаче -
гиперплазия, измененный протеолиз и др. [13].
созданию аппарата для контроля за течением
Интерпретация длинного и важного списка
времени в ходе выполнения онтогенетического
возраст-ассоциированных патологий как про-
цикла.
явлений гиперфункции создает, однако, некото-
рую иллюзию достигнутого понимания главного
механизма старения. В упомянутой работе Gems
РОЛЬ ПЛАНЕТЫ ЗЕМЛЯ
и de Magalhães, критикующих подобную ил-
В РАБОТЕ МЕТРОНОМА ЖИВОТНЫХ
люзию, метко сказано, что как муха-журчалка,
не имея жала, мимикрирует под осу, так и не-
Известных в молекулярной биологии фак-
которые классификации признаков старения
торов, таких как транскрипция, трансляция
создают видимость найденной парадигмы [10].
и т.д., по-видимому, принципиально недоста-
Ведь то обстоятельство, что «постростовое» на-
точно для измерения хода времени. ДНК са-
рушение внутриклеточных сигнальных путей
мостоятельно, без особого процесса, вероятно,
ведет к возрастным заболеваниям на организ-
не может участвовать в контроле хода времени.
менном уровне, еще не является объяснением.
И сам этот процесс клетки самостоятельно ор-
Требуется понять первичную причину этих воз-
ганизовать тоже не могут. Но если биохимиче-
раст-зависимых внутриклеточных отклонений,
ские и биофизические факторы оказываются
инициирующих старение. Кризис современной
недостаточными, то ничто не мешало изобре-
биогеронтологии объясняется отсутствием по-
тательной Природе обратиться в таком слу-
нимания того, что же именно является движущей
чае к помощи геофизических факторов. К их
силой старения. Хотя из двух противоборствую-
числу относятся особенности неравномерного
щих позиций - стохастика или программа - все
движения Земли в космическом пространстве.
больше доводов в пользу программы, остается
Именно это свойство нашей планеты живот-
по-прежнему загадкой, что является драйвером
ные могли взять за основу при создании метро-
развертывания во времени той онтогенетиче-
номной системы, используемой ими для тем-
ской программы, выполнение которой приводит
поральной разметки онтогенеза.
к старению. Сегодня это неизвестно, и именно
Движения Земля в пространстве включа-
поэтому, по оценкам некоторых авторов, биоге-
ют ряд периодически повторяющихся процес-
ронтология находится на распутье.
сов, в том числе изменения скорости осевого
вращения Земли, а также полярное движение
Земли, т.е. движение оси вращения Земли от-
ОНТОГЕНЕЗ
носительно ее коры, и, кроме того, нутации,
И ХОД БИОЛОГИЧЕСКОГО ВРЕМЕНИ
т.е. покачивания оси вращения Земли. Мульти-
масштабные вариации параметров вращения
Разметка событий онтогенеза во времени,
Земли состоят из высокочастотных вариаций
по-видимому, не может быть организована
полярных движений и более длиннопериодич-
только на основе событий типа стимул → ответ,
ных флуктуаций [16, 17]. Полярные движения
индукция → дифференцировка и пролифера-
Земли, или колебания земного полюса, - это
БИОХИМИЯ том 87 вып. 12 2022
МЕТРОНОМ ДЛЯ ПРОГРАММЫ ПЖ
2021
смещения оси вращения в теле Земли, т.е. из-
осцилляции планеты, относятся: полярное
менения ориентации этой оси в космическом
движение (колебание оси вращения Земли
пространстве. Осцилляции оси вращения Зем-
вокруг оси ее фигуры), нутация (кивающее
ли, называемые нутациями, зависят от притя-
движение оси вращения планеты) и вариации
жения экваториального избытка масс Земли
длины суток. Что касается нутации, то она со-
Луной и Солнцем, от наклона оси вращения
стоит из множества микронутаций, имеющих
Земли к плоскости орбиты, а также от непре-
небольшую амплитуду и высокую частоту. Ам-
рывных изменений позиции небесных тел, гра-
плитуды нутации зависят также от внутренней
витационно взаимодействующих с Землей и
структуры планеты, в том числе от жидкого
между собой. Осцилляции Земли зависят также
ядра внутри постоянно вращающейся мантии
от динамических изменений в структурах Зем-
как системы, подверженной внешнему при-
ли, как в недрах, так и на поверхности [18-22].
ливному воздействию; дополнительный вклад
Гравитационное притяжение, в основном
в картину осцилляций оси вращения Земли
со стороны Солнца и Луны, а также от других
вносят также поверхностные процессы, свя-
небесных тел (приливные силы), вызывает не-
занные с океанами и атмосферой [25-28].
большие периодические изменения в ориента-
Колебательно-вращательные процессы в
ции оси вращения Земли, которые можно изме-
движении Земли, в том числе изменения ори-
рить, используя наблюдения за очень далекими
ентации оси вращения, можно наблюдать как
квазарами; приливное воздействие в суточном
перемещения полюсов в пространстве. В воз-
диапазоне частот вызывает резонансное воз-
мущенных вращательно-колебательных дви-
буждение нутации свободного жидкого ядра и
жениях Земли под действием моментов гра-
свободные нутации внутреннего ядра [23, 24].
витационных сил от Солнца и Луны выявлен
Таким образом, к основным типам дви-
приливной механизм возбуждения колебаний
жения оси вращения Земли, влияющим на
полюса, обусловленный вращательно-посту-
периодические изменения ее ориентации в
пательным движением барицентра «двойной
пространстве и тем самым обусловливающим
планеты» Земля-Луна вокруг Солнца
[29].
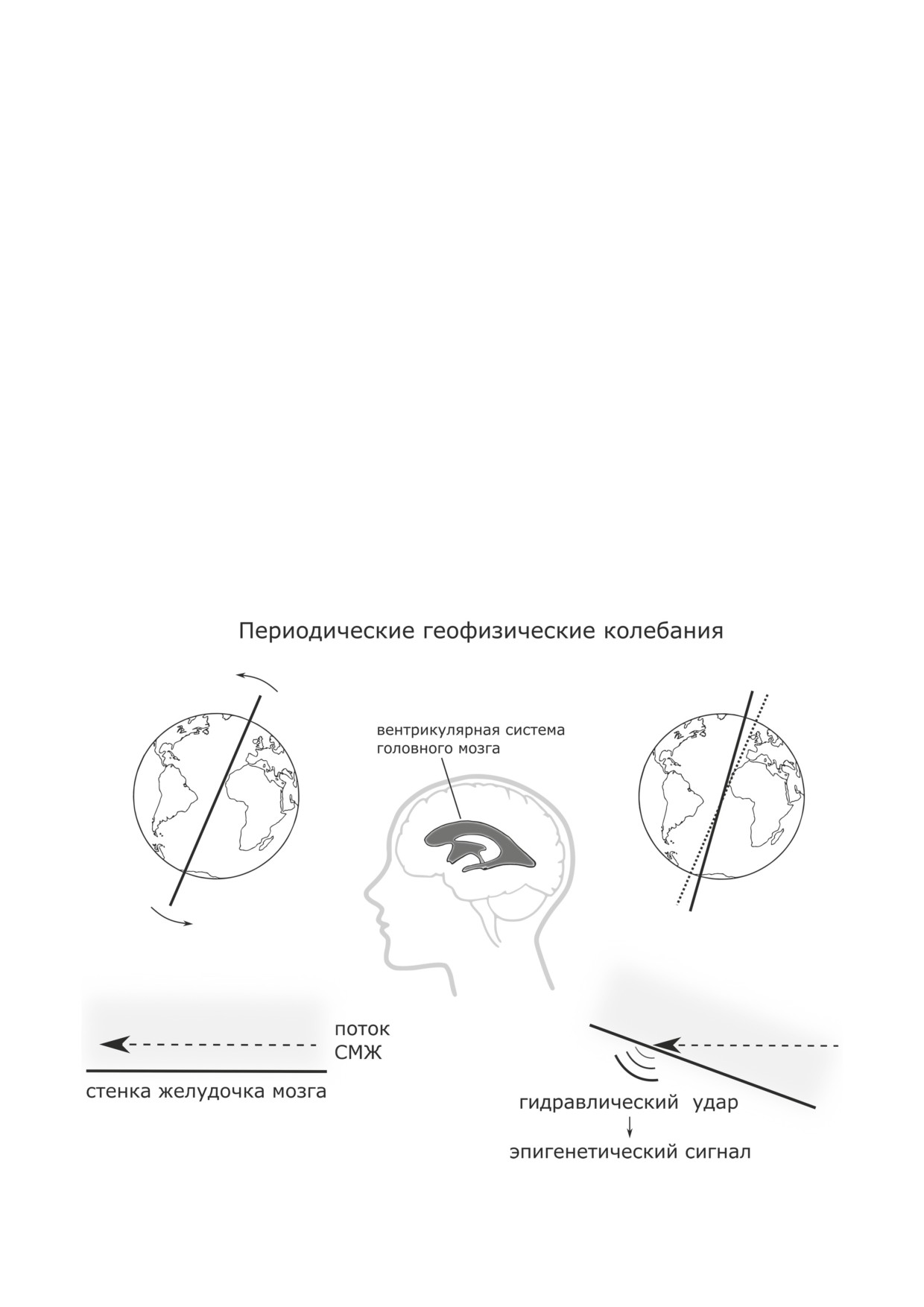
Основные принципы метрономной теории старения (модель)
БИОХИМИЯ том 87 вып. 12 2022
2022
ОЛОВНИКОВ
Основные характеристики этих колебаний
ВЕНТРИКУЛЯРНАЯ СИСТЕМА МОЗГА
остаются стабильными. Анализ данных наблю-
КАК СТРУКТУРА ДЛЯ ГЕНЕРАЦИИ
дений и измерений движений полюса Земли на
МЕТРОНОМНЫХ УДАРОВ
длительном интервале времени показал, что ос-
С УЧАСТИЕМ ПЛАНЕТЫ ЗЕМЛЯ
цилляции полюса Земли синфазны с прецесси-
И ПРИСТЕНОЧНОГО ПОТОКА
онными движениями орбиты Луны, отражаю-
СПИННОМОЗГОВОЙ ЖИДКОСТИ
щими изменения ориентации оси вращения
Луны в пространстве [30]. Продемонстриро-
Можно предположить, что постоянные от-
вано также существование внутрисуточных
клонения в траектории движений Земли ис-
колебаний полюса Земли, вызванных грави-
пользуются ее животными обитателями в рабо-
тационно-приливными моментами Солнца и
те метрономной системы, предлагаемой здесь
Луны [31].
на роль ключевого средства юстировки про-
В процессе орбитального движения пла-
должительности периодов онтогенетического
нета Земля все время испытывает небольшие
цикла. Структурой, которую могли бы избрать
колебания мгновенной оси вращения, кото-
для такой цели обладатели мозга (или его ана-
рые характеризуются различными частотами.
лога), является вентрикулярная система. Она
Серии коротких периодов колебаний вызыва-
представлена соединенными полостями, вдоль
ются большей частью приливными моментами
которых как по каналам течет СМЖ.
Солнца и Луны. Многие колебания Земли име-
Желудочки и центральный канал спинно-
ют субсуточные частоты, хотя вариации пла-
го мозга выстланы эпендимоцитами (клетками
нетарных колебаний меняются во временных
нейроглии). Эпендимальные клетки экспони-
масштабах от субсуточных до многодесятилет-
руют свои подвижные реснички, миниатюр-
них [19, 27, 32]. Вклад в эти гравитационно-
ные хлыстообразные органеллы, в желудочки
опосредованные колебания вносят и геофи-
головного мозга и способствуют оттоку СМЖ.
зические процессы в виде перераспределения
Подвижные реснички эпендимальных клеток
масс внутри Земли и в ее внешней оболоч-
строго координируют свои сокращения, что-
ке, т.е. океанах и атмосфере. В совокупности
бы способствовать мощному и направленному
гравитационные силы Солнца, Луны и гео-
току СМЖ. Этот поток может участвовать в пи-
физические процессы, ассоциированные с их
тании, защите мозга и удалении отходов [38].
воздействиями, участвуют в краткосрочных и
Кроме того, молодые нейробласты, рождаю-
долгосрочных периодических изменениях ори-
щиеся в субвентрикулярной зоне взрослого
ентации оси вращения Земли в космическом
мозга, используют этот поток в своей мигра-
пространстве [17, 33-37].
ции от стенок боковых желудочков до обоня-
Итак, Земля постоянно испытывает такие
тельной луковицы [39].
физические движения, как небольшие коле-
Сложная и внешне даже причудливая ар-
бания и вращение в различных формах. Ори-
хитектоника вентрикулярной системы моз-
ентация оси вращения Земли постоянно ме-
га, однако, не может быть объяснена только
няется во времени. Что касается траектории
перечисленным
[40]. Различные популяции
полета Земли в космическом пространстве,
подвижных реснитчатых клеток (и биение их
разнообразные малые осцилляции являются
ресничек) пространственно организованы так,
проявлением постоянных отклонений от рав-
чтобы направленный поток СМЖ ограничи-
номерного и строго однонаправленного дви-
вался отдельными желудочковыми полостями
жения планеты. Благодаря этому Земля как
с небольшим обменом жидкости между желу-
планета способна играть роль «трансформато-
дочками, несмотря на пульсирующие смеще-
ра», который транслирует свои гравитационно
ния СМЖ, вызванные сердцебиением [41].
обусловленные движения в такой вид гидроди-
Яркой особенностью этой системы явля-
намических сигналов спинномозговой жидко-
ются нейроны мозга, напрямую омываемые
сти (СМЖ), которые могут быть распознаны
потоком СМЖ. Vigh et al. [42, 43] подробно
специализированными структурами живых су-
описали морфологию системы СМЖ-контак-
ществ. У высших животных, включая челове-
тирующих нейронов, подчеркнув, что они,
ка, за это отвечает метрономная система, воз-
очевидно, играют роль в восприятии разных
никшая в эволюции на базе желудочков мозга
сигналов, так или иначе связанных с СМЖ.
и потока СМЖ. Аналог метрономной системы
Эти нейроны контактируют с СМЖ через свои
должен существовать и работать также у беспо-
дендриты, перикарионы или аксоны, и боль-
звоночных. Зависимость работы метронома от
шинство из них посылает дендритные отрост-
периодических геофизических колебаний по-
ки в полость желудочка, где они образуют рес-
казана на рисунке.
нитчатые окончания.
БИОХИМИЯ том 87 вып. 12 2022
МЕТРОНОМ ДЛЯ ПРОГРАММЫ ПЖ
2023
Значительная часть этих сенсорных кле-
ток СМЖ, вместе с планетой и одновременно
ток присутствует в различных областях ги-
с ней меняют свою позицию в пространстве.
поталамуса, таких как паравентрикулярный
Поэтому поток СМЖ, кратковременно со-
орган и сосудистый мешок. Было высказано
храняя прежнее направление движения, при
предположение, что СМЖ-контактирующие
таких условиях должен ударяться в стенку ка-
нервные клетки из специализированных ядер
нала либо отклоняется от нее в зависимости
гипоталамуса, распознавая в СМЖ различные
от направления мгновенного смещения пла-
биоактивные субстанции, участвуют в гипо-
неты. Эти меняющиеся механические взаимо-
таламо-аденогипофизарной регуляции. Дру-
действия потока жидкости со стенками канала
гие СМЖ-контактирующие нейроны, по мне-
служат причиной неизбежного возникновения
нию той же группы исследователей, могли бы
гидродинамических возмущений в пристеноч-
быть чувствительными к давлению или пото-
ном потоке СМЖ.
ку СМЖ, а также к световому воздействию на
Таким образом, потоковые гидродина-
ткани мозга [42].
мические возмущения неизбежно должны
СМЖ-контактируюшие клетки, которые
возникать в непосредственной близости от
присутствуют в стенках желудочков головного
ограничивающей поверхности вентрикуляр-
мозга и центрального канала спинного мозга,
ной системы. Именно там, т.е. на внутренней
найдены у всех исследованных позвоночных.
поверхности полостей мозговых желудочков,
При использовании миноги, как удобной мо-
выставлены, как отмечено выше, механосен-
дели спинного мозга, показана способность
сорные окончания нервов перивентрикуляр-
реснитчатых соматостатин/ГАМК-экспресси-
ной сети нервной системы, которые способны
рующих СМЖ-контактирующих нейронов
улавливать механические возмущения, воз-
работать как pH-сенсорами, так и механоре-
никающие в пристеночном потоке жидкости
цепторами. СМЖ-контактирующие нейроны
вследствие неравномерных движений плане-
локализуются в основном в гипоталамической
ты. Реснички своими биениями поддержива-
области и спинном мозге. Доказано, что гипо-
ют строгую однонаправленность пристеноч-
таламические СМЖ-контактирующие нейроны
ного потока СМЖ, что как раз и позволяет
отвечают на механическую стимуляцию дви-
сенсорным нейронам вентрикулярной систе-
жений жидкости вдоль стенки третьего желу-
мы распознавать отклонения движений Зем-
дочка [44]. В гипоталамусе миноги этот ответ
ли и переводить гидродинамические сигна-
осуществляется через каналы, подобные кислото-
лы потока СМЖ на язык нервной системы,
чувствительным ионным каналам 3 (ASIC3).
т.е. генерировать специфические нервные
Эти механорецепторные СМЖ-контактирую-
импульсы.
щие нейроны гипоталамуса имеют обширные
Из множества гидродинамических возму-
аксональные разветвления. Подобные нейроны
щений, генерируемых движениями планеты в
есть и у грызунов.
пристеночном потоке СМЖ, каждый вид мно-
Что касается самой СМЖ, то эта прозрач-
гоклеточных животных, обладающих вентри-
ная как вода жидкость, омывающая централь-
кулярной системой или, у более примитивных,
ную нервную систему, совершает пульсирую-
ее аналогом, выбирает в ходе эволюции такие
щие движения. Сердечная и артериальная
возмущения, которые повторяются с опти-
пульсации являются основными источниками
мальной для данного вида частотой. Эти перио-
пульсирующего движения СМЖ, за исключе-
дические возмущения транслируются в нерв-
нием глубокого брюшного дыхания, которое
ные импульсы в специализированых нейронах
тоже способно помогать направленному пото-
перивентрикулярной области, которые можно
ку СМЖ [45, 46].
обозначить как «хрононейроны». Периодиче-
Пристеночный поток и поток жидкости
ски возникающие в хрононейронах нервные
в просвете каналов вентрикулярной системы
импульсы служат теми метрономными сиг-
организованы подвижными ресничками, кото-
налами, которые необходимы для упорядочи-
рые действуют как наномашины, которые пе-
вания продолжительности каждого периода
ремещают жидкость в одном направлении по
онтогенетического цикла. Выбранная в эво-
внутренней поверхности каналов [41, 47, 48].
люции частота многократно повторяющихся
Пристеночный поток СМЖ вынужден, благо-
метрономных ударов должна быть оптимальна
даря эффекту инерции, постоянно стремить-
для каждого вида.
ся сохранять свое мгновенное направление в
Сигналы метронома отвечают за темп
пространстве, несмотря на тоже мгновенные
эпигенетических модификаций темпораль-
смещения планеты. Стенки вентрикулярного
ной ДНК (тДНК) в хрононейронах. При про-
канала, по которому течет пристеночный по-
чих равных условиях редкие удары метронома
БИОХИМИЯ том 87 вып. 12 2022
2024
ОЛОВНИКОВ
благоприятны для организации медленного
головного мозга, и такая архитектура, в свою
расходования тДНК и, в частности, для долгой
очередь, способствует организации направ-
репродуктивной жизни. Метрономный ритм
ленных потоков жидкости в вентрикулярных
высокой частоты, напротив, соответствует бо-
полостях мозга, что важно для успешной рабо-
лее быстрому исчерпанию тДНК, контролирую-
ты предложенной метрономной системы [50].
щей продолжительность определенного онто-
Равномерные биения ресничек детерми-
генетического периода. Сказанным, однако, не
нируют направление пристеночного потока
исключается, что один и тот же вид животных
жидкости в каждом желудочке независимо от
в разные периоды своего онтогенетического
размера его полости. Поэтому механизм спо-
цикла может ориентироваться на удары метро-
собен выполнять роль метронома независимо
нома, совершающиеся с разной частотой. Дви-
от размера мозга. Сложность нервной системы
жения Земли генерируют в потоке СМЖ ши-
для его работы не важна, он может работать
рокий спектр возмущений, так что недостатка
как у позвоночных, так и у беспозвоночных.
в эволюционном выборе необходимых вариан-
Достаточно иметь в нервной системе стабиль-
тов не существует.
но работающий пристеночный поток жидко-
Учитывая сложное устройство вентрику-
сти и в нем механосенсорные нейроны.
лярной системы, исследователи неоднократ-
Контактирующие с СМЖ нейроны присут-
но предполагали, что у СМЖ, эволюционно
ствуют во всем исследованном филуме позво-
консервативной транспортной системы мозга,
ночных. Они армированы ресничками, помо-
должна существовать какая-то еще, возможно,
гающими воспринимать гидродинамические
ключевая физиологическая функция, до сих
эффекты в СМЖ. Вентрикулярная система,
пор остающаяся неизвестной [38, 49].
подобно органу слуха, конвертирует энер-
В целом, создается впечатление, что каж-
гию колебаний в процесс нервного возбужде-
дый желудочек выполняет свою часть неко-
ния [44]. Механосенсорные, погруженные в
ей общей задачи, ради выполнения которой
поток жидкости, нейроны являются, по-ви-
вся вентрикулярная система сформировалась.
димому, идеальным кандидатом для улавли-
Сложная и, как выше упомянуто, причудли-
вания сигналов, генерируемых движениями
вая форма системы полостей мозга может быть
планеты.
объяснена тем, что ее определенные компарт-
Каждый специфический для вида ме-
менты имеют нервные связи с разными регио-
трономный сигнал, обработанный в специа-
нами мозга и, соответственно, тела для того,
лизированных нейронах мозга, хрононей-
чтобы посылать в них разные регион-специ-
ронах, инициирует в них акт очередной
фичные метрономные сигналы. Не исключе-
эпигенетической записи путем последова-
но, что в разных желудочках их метрономная
тельной модификации регуляторных сайтов.
система эволюционно настроена на генериру-
Полное заполнение этими записями региона
емые планетой гидродинамические сигналы,
тДНК, отведенного эволюцией на выполнение
имеющие разные характеристики. Чем боль-
очередного периода онтогенетического цикла,
ше период, через который следует очередной
есть необходимое и достаточное условие для
метрономный удар, тем при прочих равных
перехода к следующему периоду онтогенеза.
условиях медленнее меняется биологический
Частоты ударов метронома, и значит, периоды
возраст мишени, темпоральная биологическая
метрономного ритма, очевидно, сильно отли-
судьба которой подконтрольна метрономным
чаются у коротко- и долгоживущих видов, та-
сигналам конкретного вентрикулярного ком-
ких как муха и слон.
партмента. В конечном итоге это могло бы
Следует подчеркнуть, что ритм, исполь-
вести к гетерохронности осуществления он-
зуемый видом в работе своего метронома,
тогенетических процессов в разных органах и
принципиально отличается от биологических
тканях единого организма, т.е. к неидентично-
ритмов, иногда привлекаемых с целью объяс-
му темпу развития, созревания и старения его
нения особенности развития или старения,
разных тканей и органов.
ведь метрономный ритм имеет экзогенное, а не
Следует особо подчеркнуть, что движения
эндогенное происхождение. И потому он ста-
самого организма несопоставимо медленнее,
бильнее, что существенно для разметки боль-
чем мгновенные смещения осциллирующей
ших интервалов времени. Вероятно, именно с
планеты. Поэтому его индивидуальные движе-
целью создания будущего метронома нервная
ния не могут оказывать влияния на характери-
система проходит, как упомянуто выше, через
стики генерируемых метрономных сигналов.
стадию нервной трубки, из которой в дальней-
Механосенсорные гены способствуют по-
шем развивается система полостей, известная
ляризации эпителиальных клеток желудочков
как вентрикулярная система мозга.
БИОХИМИЯ том 87 вып. 12 2022
МЕТРОНОМ ДЛЯ ПРОГРАММЫ ПЖ
2025
ГИДРОМЕХАНИЧЕСКИЕ
система каждого животного вида утратила чув-
УДАРЫ ПОТОКА СМЖ
ствительность в ходе эволюции. Необходимая
ПО МЕХАНОСЕНСОРНЫМ НЕЙРОНАМ
фильтрация достигается либо на уровне нейро-
сенсоров, погруженных в СМЖ, либо на уров-
В чем состоит смысл поддержания строго
не сети хрононейронов.
направленного пристеночного потока СМЖ в
Важно также учитывать, что скорость са-
желудочках мозга? Это необходимо для гене-
мостоятельных движений организма несопо-
рации сигналов метрономной системы. Меха-
ставимо меньше скоростей осциллирующих
носенсорные окончания нейронов находятся в
смещений Земли в пространстве. Поэтому
стенках желудочков мозга и омываются пото-
произвольные изменения позиции организма
ком СМЖ. Пока планета Земля, а вместе с ней и
не могут влиять на необходимые характеристи-
организм с его потоком СМЖ несутся с огром-
ки генерируемых метрономных сигналов.
ной скоростью в космическом пространстве в
В целом, метрономная конструкция, ис-
постоянном направлении, механосенсорные
пользуя столь разные факторы, как механо-
нейроны не получают механических сигналов.
сенсорные нейроны, поток СМЖ и движения
Однако, когда Земля производит очередное не-
планеты, выполняет в организме важную фи-
большое изменение в ориентации своего дви-
зиологическую функцию, а именно упорядо-
жения, происходит следующее событие.
чение онтогенетических периодов во времени.
Организм и стенка его вентрикулярного
Происходит в том числе и упорядочение хода
канала вместе с планетой меняют свою пози-
старения.
цию, тогда как траектория движения присте-
ночного потока СМЖ, в силу инерции движе-
ния потока, сохраняет прежнее направление.
ТЕМПОРАЛЬНАЯ ДНК
То есть поток СМЖ, поддерживая по инерции
КАК СУБСТРАТ ДЛЯ ПЛАНЕТАРНОГО
траекторию своего движения, вынужден стал-
МЕТРОНОМА В ЕГО КОНТРОЛЕ
киваться со стенкой канала, которая мгновен-
ПРОДОЛЖИТЕЛЬНОСТИ РАЗНЫХ
но поменяла свою позицию в пространстве,
ПЕРИОДОВ ОНТОГЕНЕТИЧЕСКОГО
причем одновременно с организмом и всей
ЦИКЛА, ВКЛЮЧАЯ СТАРЕНИЕ И ПЖ
осциллирующей планетой. В итоге механо-
сенсорные нейроны желудочка, погруженные
Основные периоды онтогенетического
в поток СМЖ, подвергаются неизбежному гид-
цикла многоклеточного животного организ-
ромеханическому воздействию, т.е. получают
ма включают развитие, рост, половое созре-
удар. Благодаря этому периодические возму-
вание, продолжительность репродуктивной
щения в потоке жидкости, воспринимаемые
жизни и пострепродуктивного периода жизни.
механонейросенсорами, могут использоваться
Все эти процессы являются примерами время-
организмом как тиканье своеобразных плане-
зависимых изменений, их текущее изучение
тарных часов, которые функционируют благо-
сосредоточено в основном на межорганных
даря гравитационным взаимодействиям Земли
связях и регуляции размеров тела [51, 52]. Все
с другими небесными телами, главным образом
больше данных в этой области указывают на
с Луной и Солнцем. В результате механосен-
ключевую роль эпигенома в регуляции генов в
соры, встроенные в стенку вентрикулярного
гипоталамусе и гипофизе [53].
канала, получают механические сигналы, по-
В настоящей работе предлагается темпо-
тенциально способные выполнять роль внеш-
ральный механизм, объясняющий с единой по-
них сигналов. Каждый сигнал, оцененный
зиции регуляцию продолжительности жизни
как специфичный для данного биологическо-
животных и происхождение их старения. Пред-
го вида, служит триггером для осуществления
полагается, что по крайней мере позвоночные
акта нанесения очередного эпигенетического
животные для регуляции во времени деталей
маркера на темпоральную ДНК в хрононейро-
своего онтогенетического цикла используют
нах, отвечающих за контроль времени.
две важные сущности - метрономную систе-
Существенно отметить, что организму
му и специализированную тДНК. Совместно
совершенно не требуется воспринимать все
они выполняют функцию хронографа, отме-
движения Земли через гидромеханическую си-
чающего пройденную часть онтогенетического
стему потока СМЖ. Более того, это дезоргани-
цикла, т.е. продолжительность уже прожитого
зовало бы всю работу метрономной системы.
организмом времени.
Достаточно воспринимать лишь определен-
Что заставляет организм животного или
ные гидромеханические сигналы, игнорируя
человека, только что завершившего разви-
все прочие сигналы, к которым метрономная
тие и рост, генерировать условия, чреватые
БИОХИМИЯ том 87 вып. 12 2022
2026
ОЛОВНИКОВ
отклонениями, ведущими к старению? Почему
являть свои координирующие свойства в кле-
организм не может остановиться на достигну-
точной культуре и, наоборот, в наибольшей
том? Как именно и каким способом регламен-
степени - в целостном организме.
тируется продолжительность взрослой репро-
Геронтологи неоднократно предлагали
дуктивной жизни, а также и других основных
рассматривать старение как следствие процес-
периодов онтогенетического цикла?
са развития [13, 55-59], в том числе развивая
Все периоды онтогенеза осуществляются,
идеи Дильмана относительно роли гипотала-
как известно, при участии множества генов и
муса в старении.
сигнальных путей, но, как здесь постулирует-
Почему отбор не может устранить старение
ся, они находятся под интеграционным кон-
как неадаптивное явление? Вряд ли из-за «за-
тролем особого механизма, который в своей
боты» о защите экологической ниши от пере-
работе использует несколько совершенно раз-
полнения. Истинная причина существования
нородных компонентов, выполняющих еди-
старения подавляющего большинства видов
ную функцию. К ним относятся: 1) желудочки
животных (и человека), как можно предпо-
мозга (вентрикулярная система), 2) темпораль-
ложить, заключается в совершенно другом:
ная ДНК (тДНК), специализирующаяся на
физически один и тот же субстрат в виде тДНК
учете хода времени в онтогенезе, и 3) планета
отвечает как за продолжительность репродук-
Земля, постоянно совершающая небольшие
тивной производительности организма, так и у
осциллирующие движения в ходе своего ос-
большинства видов за ПЖ. Что касается зна-
новного орбитального движения вокруг Солн-
чительного увеличения пострепродуктивной
ца. Представление об этом механизме, назван-
продолжительности жизни, например, у че-
ном метрономным по его ключевой функции,
ловека и косаток, то это, вероятно, есть про-
предлагается здесь в качестве фундамента не-
сто вторичное приобретение, созданное есте-
достающего пока объяснения происхождения
ственным отбором в качестве ответа на пользу
организменного старения и путей регуляции
длительной заботы родителей о потомстве [60].
видовой ПЖ.
Работа программы продолжительности
Каждому периоду онтогенеза соответствует
репродуктивной жизни как части программы
своя последовательность тДНК. «Расходова-
онтогенетического цикла животных осущест-
ние» тДНК (в виде ее эпигенетического марки-
вляется на уровне нейронов (хрононейронов),
рования) происходит в нейронах, или хроно-
обрабатывающих сигналы метронома, посту-
нейронах, специализирующихся на контроле
пающие из вентрикулярной системы. В хроно-
течения онтогенетического времени. Эпигене-
нейронах с каждым ударом метронома имеет
тическое маркирование сайтов тДНК, отвеча-
место один акт последовательной эпигенетиче-
ющих за контроль продолжительности периода
ской модификации очередного сайта их тДНК.
взрослой репродуктивной жизни, снижает эф-
Это происходит на уровне промотора или дру-
фективность функционирования генов, работа
гой специфической последовательности тДНК.
которых модулируется регуляторными РНК,
В ходе выполнения репродуктивного периода
транскрибируемыми с матриц тДНК. Сниже-
онтогенетического цикла указанный процесс
ние продуктивности маркируемой тДНК (в
эпигенетической модификации тДНК мог бы
отношении, возможно, и состава, и концен-
вести, в частности, к постепенному ослаблению
трации ее транскриптов) могло бы быть связа-
деятельности некоторых ингибиторных путей
но не только с модификацией ее промоторов,
регуляции, вызывая в ответ, например, небла-
но и других последовательностей тДНК. Сум-
гоприятное для организма гиперфункциони-
марно эти процессы ведут к формированию
рование соответствующих процессов. Другие
возраст-зависимых дисфункций в организме,
эпигенетические модификации могли бы вести
т.е. к старению. Старение характеризуется, сре-
к потере эффективного протеостаза и иным
ди прочего, потерей оптимальных взаимодей-
неблагоприятным для выживания событиям.
ствий между клетками в разных тканях и органах
Важно подчеркнуть, что при этом непре-
и ослаблением их координации. Так, например,
рывно работает адаптивная программа регу-
два наиболее распространенных возрастных
ляции продолжительности фертильного перио-
нейродегенеративных заболевания - болезнь
да, поддерживаемая естественным отбором.
Альцгеймера и болезнь Паркинсона - харак-
При таком раскладе вред для организма ока-
теризуются синаптической дисфункцией, а это
зывается прямым побочным следствием ука-
ведет к потере контроля над мишенями [54].
занной пользы. Таким образом, как упомянуто
Следует подчеркнуть, что координирующие
выше, процесс последовательного эпигенети-
функции транскриптов, кодируемых тДНК,
ческого маркирования тДНК, выполняя про-
вероятно, лишь в малой мере имеют шанс про-
грамму продолжительности репродуктивной
БИОХИМИЯ том 87 вып. 12 2022
МЕТРОНОМ ДЛЯ ПРОГРАММЫ ПЖ
2027
жизни, оказывается вынужденным параллель-
нивать ход времени является центральной для
но вызывать старение, а затем и дряхление
понимания проблемы старения и организации
организма. Ярким примером отклонения от
продолжительности жизни организмов.
подобного сценария являются случаи острого
Что закодировано в тДНК? Наиболее веро-
феноптоза, такие как, например, запрограм-
ятно, что ее экспрессия сопряжена с поставкой
мированная гибель тихоокеанских лососей
к мишеням нейронов таких факторов, как ней-
сразу после нереста, осуществляемая ради
ропептиды, нейромедиаторы, регуляторные
успешного выживания их потомства [8].
РНК и др., с участием которых, специализи-
На исполнение функции тДНК могли бы,
рованные на темпоральный контроль нейро-
в частности, претендовать некоторые сегмен-
ны (хрононейроны) обеспечивают регуляцию
ты хромосом, которые у животных ассоции-
подчиненных им мишеней, включая клетки ав-
рованы с их продолжительностью жизни. Так,
тономной нервной системы и другие системы,
например, при поисках мышиных локусов,
которые, в свою очередь, важны для согласо-
ассоциированных с долгожительством, прово-
ванной работы всех тканей и органов.
дившихся в ходе межвидовых сравнений гене-
тически гетерогенных мышей с геномами дру-
гих млекопитающих, были идентифицированы
ОБСУЖДЕНИЕ
несколько частей генома, влияющих на про-
должительность жизни мышей. Эти хромосом-
Цель живой природы
- поддержание
ные участки оказались большими. Одна часть
жизни. Этой цели подчинена и структура он-
хромосомы 12 влияла на продолжительность
тогенетического цикла каждого вида. Соот-
жизни у всех мышей. На ожидаемую продол-
ношение величин отдельных темпоральных
жительность жизни самок также влиял участок
периодов этого цикла согласовано между собой
мышиной хромосомы 3. Некоторые из этих ге-
и в конечном счете подчинено решению глав-
нетических признаков различались у самцов и
ной задачи - обеспечению размножения вида.
самок мышей, а некоторые влияли на продол-
Естественный отбор работает на главную цель
жительность жизни только после определенно-
жизни, на ее вечное продолжение, и только по-
го возраста [14].
этому он пренебрегает параллельным появле-
Как правило, генетическая картография
нием возрастных патологий.
долголетия направлена на поиски генов, соче-
При прочих равных условиях, оперируя
таний генов и генетических путей, отвечающих
именно с тДНК, отбор может создавать ва-
за долголетие [61-64]. Анализ продолжитель-
риации, характерные для видов с большой и
ности жизни дрозофилы показал, что боль-
малой ПЖ, с их часто радикальными разли-
шинство локусов ПЖ пол-специфичны [65].
чиями по численности потомства. Большин-
С одной стороны, бесконечный поиск соответ-
ство видов с короткой продолжительностью
ствующих генов вполне оправдан, поскольку и
жизни имеют много потомков, а виды с боль-
старение, и долгожительство - очень сложные
шой продолжительностью жизни имеют мало
признаки, в реализацию которых вовлечено
потомства, и этих животных, согласно их раз-
множество процессов. Однако можно предпо-
ным типам онтогенетического цикла, относят
ложить, что применительно к контролю величи-
к R- и K-стратегам соответственно [66, 67].
ны ПЖ Природа пошла в эволюции по новому
Отбор на отсроченное воспроизводство обыч-
пути. Сформировалась особая фракция ДНК,
но создает популяции с повышенной продол-
которая, помимо кодирования регуляторных
жительностью жизни, и эти популяции часто
продуктов, выполняет принципиально новую
демонстрируют повышенную плодовитость в
миссию - она играет роль своеобразного «рас-
конце жизни [68]. Сообщалось, что селектив-
ходного» материала. Точнее, роль этой тДНК
ное размножение мышей с повышенным ре-
частично похожа на роль ленты для диктофо-
продуктивным долголетием приводило к кор-
на. По сути, организм использует эту ДНК как
релированному улучшению репродуктивного
ленту своеобразного хронографа. Эпигенети-
долголетия и увеличению ПЖ [69]. Исследо-
чески маркируя тДНК, он учитывает величину
вание исторических данных, касающихся Са-
того интервала времени, которое уже истекло в
амских женщин, обнаружило, что естествен-
ходе выполнения организмом прожитой части
ный отбор благоприятствовал более раннему
его сложного онтогенетического цикла.
началу и более позднему прекращению раз-
Поскольку старение - зависящее от вре-
множения. Хотя общая плодовитость не была
мени функциональное изменение многих
связана с продолжительностью жизни, но
процессов [15], представляется оправданным
женщины, давшие последнее потомство в про-
предположить, что именно способность оце-
двинутом возрасте, также жили дольше всех,
БИОХИМИЯ том 87 вып. 12 2022
2028
ОЛОВНИКОВ
т.е. репродуктивное долголетие и продолжи-
При прочих равных условиях, чем больше
тельность жизни взрослого человека в этих по-
хрононейронов заложено в ходе развития, тем
пуляциях были сопряжены [70].
больше может оказаться ПЖ. Причина некото-
Признаки старения и механизмы, усу-
рого увеличения ПЖ в этом случае заключается в
губляющие состояние стареющего организ-
том, что суммарный уровень нейрональных про-
ма [15, 71], представляют собой, как можно
дуктов (нейропептиды, микроРНК и др.) вблизи
предположить, следствие некоего первичного
их мишеней будет выше при большей численно-
процесса, который ранее не учитывался в тео-
сти хрононейронов, чем при небольшой.
риях старения. В настоящей работе постулиро-
Калорийно ограниченная диета замедляет
ваны два взаимодополняющих и незаменимых
эпигенетическую реализацию сигналов, посту-
фактора онтогенеза, важных для понимания
пающих из метрономной системы, и поэтому
видовых лимитов продолжительности жизни и
увеличивает ПЖ. Вносят вклад и другие факто-
происхождения старения. Предполагается, что
ры, такие как измененная в условиях этого де-
существует темпоральная программа продол-
фицита активность митохондрий и гормонов,
жительности взрослой репродуктивной жизни,
но все эти факторы вторичны по отношению к
и что селекция способствует репродуктивному
роли частоты ударов и запаса тДНК. Хотя из-
долгожительству, коррелирующему с продол-
вестно, например, что мыши с генетическим
жительностью жизни.
дефектом или резистентностью к гормону ро-
Что касается инфрадианных биоритмов как
ста живут дольше, чем их нормальные братья
иногда предполагаемой основы темпоральной
и сестры [72], основной причиной эффекта
разметки онтогенеза, то сама их стабильность,
увеличения ПЖ в этой ситуации является, воз-
вероятно, зависит от экзогенных геофизиче-
можно, замедленный процесс эпигенетическо-
ских синхронизаторов. Это резко ограничивает
го маркирования тДНК.
возможности использования эндогенных био-
В ходе эволюции отбор, вероятно, должен
ритмов как реперных точек онтогенеза. При-
был способствовать снижению чувствительно-
менимость в дикой природе для той же цели
сти тех сенсорных систем, которые позволяют
циркадианных ритмов еще менее вероятна для
мозгу осознавать влияние скачкообразных из-
большинства долгоживущих видов ввиду чрез-
менений в ориентации Земли в пространстве.
мерно короткого периода таких ритмов. Метро-
Слишком высокая чувствительность всех сен-
номная система, предложенная здесь, полно-
соров мозга по отношению к движениям Зем-
стью свободна от подобных ограничений.
ли, отражающимся на гидродинамическом
В рассматриваемом контексте важен еще
поведении спинномозговой жидкости, мешала
один вопрос - за счет чего взрослая жизнь
бы сознанию, доставляя массу неудобств и ра-
может оказываться несколько длиннее репро-
ботая как контрадаптивный механизм, систе-
дуктивной? Ответ состоит в следующем. Для
матически вызывающий беспричинное беспо-
эффективности репродуктивной жизни орга-
койство или нарушения жизнедеятельности.
низм должен иметь достаточно высокую жиз-
Существенно отметить, что движения са-
неспособность. Поэтому к концу завершения
мого организма несопоставимо медленнее, чем
репродуктивной жизни у организма должна еще
мгновенные смещения осциллирующей плане-
оставаться небольшая фракция темпоральной
ты. Поэтому самостоятельные передвижения
ДНК, все еще продолжающая экспрессировать-
организма не способны систематически влиять
ся. Если ПЖ существенно длиннее фертильного
на генерацию метрономных сигналов.
периода, как в вышеупомянутом случае с косат-
ками и людьми, то тут эффект связан, по-види-
мому, с дополнительной работой отбора.
ЗАКЛЮЧЕНИЕ
Только лишаясь в продвинутом возрасте
достаточного уровня координирующей под-
В этой работе рассмотрен механизм, объ-
держки нейрональных факторов (эта поддерж-
ясняющий с единой позиции происхождение
ка со стороны эпигенетически модифициру-
старения и регуляцию продолжительности
емой тДНК с течением времени постепенно
жизни животных и человека. Предполагается,
снижается), некоторые гены, возможно, могли
что они используют для регуляции своего раз-
бы начинать вести себя как вредные, прояв-
вития во времени, т.е. для темпоральной регу-
ляя признаки антагонистической плейотро-
ляции онтогенетического цикла, две важные
пии. В самих генах никакой вредоносности
сущности - метрономную систему и специали-
нет, вредна лишь утрата их согласованных дей-
зированную на учет хода времени темпораль-
ствий на фоне дефицита факторов, контроли-
ную ДНК. На выполнение каждого периода
руемых тДНК.
онтогенеза в геноме каждого вида эволюцией
БИОХИМИЯ том 87 вып. 12 2022
МЕТРОНОМ ДЛЯ ПРОГРАММЫ ПЖ
2029
отведена определенная длина тДНК, хотя и с
существует, а упорядоченность процессов ста-
некоторыми индивидуальными отклонениями
рения во времени есть следствие выполнения
в сторону ее уменьшения или увеличения. Она
организмом программы продолжительности
специфична в отношении отдельных периодов
репродуктивной жизни.
онтогенеза. Видовая тДНК и метроном с его
Система желудочков как полостей вентри-
видоспецифической частотой ударов, на кото-
кулярной системы мозга является органом, ко-
рую эволюционно настроена система их вос-
торый был создан эволюцией, по-видимому,
приятия у каждого вида, в совокупности вы-
в первую очередь ради работы метрономной
полняют функцию хронографа. Он записывает
системы и темпоральной разметки онтогенеза.
выполненную на данный момент часть онто-
Вероятно, именно ради этого нервная система
генетического цикла, фиксируя уже прожитый
как архитектура развивается в эмбриогенезе на
организмом срок его жизни.
основе нервной трубки, на базе которой фор-
В структурах и функциях животного орга-
мируются не только отделы мозга, но система
низма, уже завершившего развитие и рост, на-
циркуляции спинномозговой жидкости, без
блюдаются отклонения, ведущие к старению.
которой метроном работать не может.
Это происходит потому, что процесс старения
При выполнении своей функции метро-
обусловлен теми же эпигенетическими моди-
номная система использует небольшие коле-
фикациями той тДНК, которая используется
бания, совершаемые нашей планетой. Ранее
для адаптивно ценного контроля над продолжи-
я рассмотрел роль неравномерных движений
тельностью репродуктивной жизни. Представ-
Земли, которыми могут пользоваться растения
ление об этом предлагается здесь и как объясне-
в их сложном процессе ориентации относи-
ние первопричины организменного старения,
тельно вектора силы тяжести [73]. Здесь роль
и как путь регуляции видовой продолжитель-
движений планеты предложена в связи с по-
ности жизни. Старение в подавляющем боль-
требностями темпоральной организации онто-
шинстве случаев - процесс явно неадаптив-
генетического цикла животных. Если предла-
ный, но, являясь итогом работы адаптивной
гаемый механизм правильный, то контроль над
программы продолжительности репродуктив-
процессом старения возможен через управле-
ной жизни, закодированной в тДНК, он ста-
ние предложенной здесь новой физиологиче-
новится вынужденным побочным феноменом.
ской системой - метрономной.
Поддерживая метроном-зависимое выполне-
ние полезной программы продолжительности
Благодарности. Выражаю благодарность
репродуктивной жизни, естественный отбор не
А.И. Калмыковой и И.А. Оловникову за помощь
имеет возможности отвергнуть старение имен-
в редактировании, за советы и обсуждение.
но потому, что оба процесса выполнятся с уча-
Конфликт интересов. Автор заявляет об от-
стием общего для них субстрата - одной и той
сутствии конфликта интересов.
же последовательности тДНК. Итак, следует
Соблюдение этических норм. Работа выпол-
подчеркнуть, что самостоятельной программы
нена без привлечения животных и людей в ка-
старения для большинства видов животных не
честве испытуемых.
СПИСОК ЛИТЕРАТУРЫ
1. Medvedev, Z. A. (1990) An attempt at a rational clas-
5. Weismann, A. (1889) Essays upon Heredity and Kindred
sification of theories of ageing, Biol. Rev. Cambr. Phi-
Biological Problems, Clarendon Press, Oxford.
los. Soc., 65, 375-398, doi: 10.1111/j.1469-185x.1990.
6. Kirkwood, T. B., and Cremer, T.
(1982)
tb01428.x.
Cytogerontology since 1881: a reappraisal of August
2. Kirkwood, T. B. (2011) Systems biology of ageing and
Weismann and a review of modern progress, Hum.
longevity, Philos. Trans. R. Soc. London Ser. B Biol.
Genet., 60, 101-121, doi: 10.1007/BF00569695.
Sci., 366, 64-70, doi: 10.1098/rstb.2010.0275.
7. Libertini, G., Corbi, G., Conti, V., Shubernetskaya,
3. Khokhlov, A. N. (2013) Does aging need its own
O., and Ferrara, N. (2021) Evolutionary Gerontology
program, or is the program of development quite
and Geriatrics: Why and How We Age, Springer Nature,
sufficient for it? Stationary cell cultures as a tool to
doi: 10.1007/978-3-030-73774-0_1.
search for anti-aging factors, Curr. Aging Sci., 6, 14-20,
8. Skulachev, V. P. (2012) What is “phenoptosis” and
doi: 10.2174/18746098112059990009.
how to fight it? Biochemistry (Moscow), 77, 689-706,
4. Olovnikov, A. M. (2022) Aging theories, Encyclopedia
doi: 10.1134/S0006297912070012.
of Gerontology and Population Aging, Springer Interna-
9. Skulachev, V. P., Shilovsky, G. A., Putyatina, T.
tional Publishing, doi: 10.1007/978-3-030-22009-9_32.
S., Popov, N. A., Markov, A. V., Skulachev, M. V.,
БИОХИМИЯ том 87 вып. 12 2022
2030
ОЛОВНИКОВ
and Sadovnichii, V. A. (2020) Perspectives of Homo
damping of the Earth’s free core nutation,
sapiens lifespan extension: focus on external or inter-
J. Geophys. Res. Solid Earth, 126, e2020JB021042,
nal resources? Aging (Albany NY), 12, 5566-5584,
doi: 10.1029/2020JB021042.
doi: 10.18632/aging.102981.
25.
Krylov, S. S., Perepelkin, V. V., and Soe, W. Y. (2021)
10.
Gems, D., and de Magalhães, J. P. (2021) The hoverfly
Short-term forecast of the Earth’s pole motion, taking
and the wasp: a critique of the hallmarks of aging as a
into account lunar disturbances, Mech. Solids, 55,
paradigm, Ageing Res. Rev., 70, 101407, doi: 10.1016/
892-897, doi: 10.3103/S002565442006014X.
j.arr.2021.101407.
26.
Rekier, J. (2022) Free core nutation and its relation
11.
Gladyshev, V. N. (2016) Aging: progressive decline
to the spin-over mode, Planet. Sci. J.,
3,
133,
in fitness due to the rising deleteriome adjusted by
doi: 10.3847/PSJ/AC6CE2.
genetic, environmental, and stochastic processes,
27.
Rekier, J., Chao, B. F., Chen, J., Dehant, V., Rosat, S.,
Aging Cell, 15, 594-602, doi: 10.1111/acel.12480.
and Zhu, P. (2021) Earth’s rotation: observations and
12.
Gems, D.
(2022) The hyperfunction theory: an
relation to deep interior, Surveys Geophys., 43, 149-
emerging paradigm for the biology of aging, Ageing
175, doi: 10.1007/S10712-021-09669-X.
Res. Rev., 74, 101557, doi: 10.1016/j.arr.2021.101557.
28.
Zotov, L., Bizouard, C., Shum, C. K., Zhang, C.,
13.
Blagosklonny, M. V. (2022) Hallmarks of cancer and
Sidorenkov, N., and Yushkin, V. (2022) Analysis
hallmarks of aging, Aging (Albany NY), 14, 4176-4187,
of Earth’s polar motion and length of day trends in
doi: 10.18632/aging.204082.
comparison with estimates using second degree stokes
14.
Bou Sleiman, M., Roy, S., Gao, A. W., Sadler, M. C.,
coefficients from satellite gravimetry, Adv. Space Res.,
von Alvensleben, G. V. G., Li, H., Sen, S., Harrison, D. E.,
69, 308-318, doi: 10.1016/J.ASR.2021.09.010.
Nelson, J. F., Strong, R., Miller, R. A., Kutalik, Z.,
29.
Akulenko, L. D., Kumakshev, S. A., Markov, Y. G.,
Williams, R. W., and Auwerx, J. (2022) Sex- and age-
and Rykhlova, L. V.
(2005) A gravitational-tidal
dependent genetics of longevity in a heterogeneous
mechanism for the Earth’s polar oscillations, Astron.
mouse population, Science,
377, eabo3191,
Rep., 49, 847-857, doi: 10.1134/1.2085254.
doi: 10.1126/science.abo3191.
30.
Perepelkin, V. V., Rykhlova, L. V., and Soe, W. Y.
15.
Lopez-Otin, C., Blasco, M. A., Partridge, L., Serrano, M.,
(2022) In-phase variations in the parameters
and Kroemer, G. (2013) The hallmarks of aging, Cell,
of the Earth’s pole motion and the lunar orbit
153, 1194-1217, doi: 10.1016/j.cell.2013.05.039.
precession, Astron. Rep., 66, 80-91, doi: 10.1134/
16.
Sibois, A. E., Desai, S. D., Bertiger, W., and Haines, B. J.
S1063772922020081.
(2017) Analysis of decade-long time series of GPS-
31.
Markov, Y. G., Perepelkin, V. V., Rykhlova, L. V.,
based polar motion estimates at 15-min temporal
Filippova, A. S., and Le Zung, N. (2014) Modeling
resolution, J. Geodesy, 91, 965-983, doi: 10.1007/
intraday oscillations of the Earth’s pole, Astron. Rep.,
S00190-017-1001-6.
58, 194-203, doi: 10.1134/S106377291402005X.
17.
Zajdel, R., Sośnica, K., Bury, G., Dach, R., Prange, L.,
32.
Bizouard, C., Nurul Huda, I., Ziegler, Y., and
and Kazmierski, K. (2021) Sub-daily polar motion
Lambert, S. (2020) Frequency dependence of the
from GPS, GLONASS, and Galileo, J. Geodesy, 95,
polar motion resonance, Geophys. J. Int., 220, 753-
1-27, doi: 10.1007/S00190-020-01453-W.
758, doi: 10.1093/GJI/GGZ463.
18.
Bizouard, C. (2020) Geophysical Modelling of the Polar
33.
Ziegler, Y., Lambert, S. B., Nurul Huda, I., Bizouard, C.,
Motion, de Gruyter GmbH & Co KG.
and Rosat, S. (2020) Contribution of a joint Bayesian
19.
Dehant, V., Mathews, P. M.
(2015) Precession,
inversion of VLBI and gravimetric data to the
Nutation, and Wobble of the Earth, Cambridge
estimation of the free inner core nutation and free core
University Press.
nutation resonance parameters, Geophys. J. Int., 222,
20.
Loper, D. E. (2017) Geophysical Waves and Flows.
845-860, doi: 10.1093/GJI/GGAA181.
Theory and Applications in the Atmosphere, Hydrosphere
34.
Dehant, V., Laguerre, R., Rekier, J., Rivoldini, A.,
and Geosphere, Cambridge University Press.
Triana, S. A., Trinh, A., Van Hoolst, T., and Zhu, P.
21.
Sidorenkov, N. S. (2009) The Interaction Between Earth’s
(2017) Understanding the effects of the core on the
Rotation and Geophysical Processes, John Wiley & Sons.
nutation of the Earth, Geodesy Geodynam., 8, 389-395,
22.
Wei, E., Jin, S., Wan, L., Liu, W., Yang, Y., and
doi: 10.1016/J.GEOG.2017.04.005.
Hu, Z. (2015) High frequency variations of Earth
35.
Kumakshev, S. A. (2018) Gravitational-tidal model
Rotation Parameters from GPS and GLONASS
of oscillations of Earth’s poles, Mech. Solids, 53, 159-
observations, Sensors (Basel, Switzerland), 15, 2944-
163, doi: 10.3103/S0025654418020061.
2963, doi: 10.3390/S150202944.
36.
Chao, B. F. (2017) On rotational normal modes
23.
Amoruso, A., and Crescentini, L. (2020) Parameters of
of the Earth: Resonance, excitation, convolution,
the Earth’s free core nutation from diurnal strain tides,
deconvolution and all that, Geodesy Geodynam., 8,
Sci. Rep., 10, 9756, doi: 10.1038/s41598-020-66426-7.
371-376, doi: 10.1016/J.GEOG.2017.03.014.
24.
Triana, S. A., Trinh, A., Rekier, J., Zhu, P.,
37.
Yatskiv, Y. S. (1980) Nearly Diurnal Free Polar Motion
and Dehant, V.
(2021) The viscous and ohmic
Derived from Astronomical Latitude and Time
БИОХИМИЯ том 87 вып. 12 2022
МЕТРОНОМ ДЛЯ ПРОГРАММЫ ПЖ
2031
Observations, in Nutation and the Earth’s Rotation.
48.
Salman, H. E., Jurisch-Yaksi, N., and Yalcin, H.
International Astronomical Union/Union Astronomique
C. (2022) Computational modeling of motile cilia-
Internationale (Fedorov, E. P., Smith, M. L.,
driven cerebrospinal flow in the brain ventricles of
Bender, P. L., eds) vol. 78, Springer, Dordrecht,
zebrafish embryo, Bioengineering, 9, 421, doi: 10.3390/
doi: 10.1007/978-94-010-9568-6_11.
bioengineering9090421.
38.
Kumar, V., Umair, Z., Kumar, S., Goutam, R. S.,
49.
Faubel, R. J., Santos Canellas, V. S., Gaesser, J.,
Park, S., and Kim, J. (2021) The regulatory roles of
Beluk, N. H., Feinstein, T. N., Wang, Y., Yankova, M.,
motile cilia in CSF circulation and hydrocephalus,
Karunakaran, K. B., King, S. M., Ganapathiraju, M. K.,
Fluids Barr. CNS, 18, 31, doi: 10.1186/s12987-021-
and Lo, C. W. (2022) Flow blockage disrupts cilia-
00265-0.
driven fluid transport in the epileptic brain, Acta Neuro-
39.
Sawamoto, K., Wichterle, H., Gonzalez-Perez, O.,
pathol., 144, 691-706, doi: 10.1007/s00401-022-02463-y.
Cholfin, J. A., Yamada, M., Spassky, N., Murcia, N. S.,
50.
Ohata, S., Herranz-Perez, V., Nakatani, J., Boletta, A.,
Garcia-Verdugo, J. M., Marin, O., Rubenstein, J. L.,
Garcia-Verdugo, J. M., and Alvarez-Buylla, A. (2015)
Tessier-Lavigne, M., Okano, H., and Alvarez-
Mechanosensory genes Pkd1 and Pkd2 contribute
Buylla, A. (2006) New neurons follow the flow of
to the planar polarization of brain ventricular
cerebrospinal fluid in the adult brain, Science, 311,
epithelium, J. Neurosci., 35, 11153-11168, doi: 10.1523/
629-632, doi: 10.1126/science.1119133.
JNEUROSCI.0686-15.2015.
40.
Ringers, C., Olstad, E. W., and Jurisch-Yaksi, N.
51.
Texada, M. J., Koyama, T., and Rewitz, K. (2020)
(2020) The role of motile cilia in the development and
Regulation of body size and growth control, Genetics,
physiology of the nervous system, Philos. Trans. R. Soc.
216, 269-313, doi: 10.1534/genetics.120.303095.
Lond. Ser. B. Biol. Sci., 375, 20190156, doi: 10.1098/
52.
Malita, A., and Rewitz, K.
(2021) Interorgan
rstb.2019.0156.
communication in the control of metamorphosis,
41.
Olstad, E. W., Ringers, C., Hansen, J. N., Wens, A.,
Curr. Opin. Insect Sci.,
43,
54-62, doi:
10.1016/
Brandt, C., Wachten, D., Yaksi, E., and Jurisch-
j.cois.2020.10.005.
Yaksi, N. (2019) Ciliary beating compartmentalizes
53.
Shalev, D., and Melamed, P. (2020) The role of
cerebrospinal fluid flow in the brain and regulates
the hypothalamus and pituitary epigenomes in
ventricular development, Curr. Biol., 29, 229-241.
central activation of the reproductive axis at puberty,
e226, doi: 10.1016/j.cub.2018.11.059.
Mol. Cell Endocrinol.,
518,
111031, doi:
10.1016/
42.
Vigh, B., Manzano e Silva, M. J., Frank, C. L., Vincze, C.,
j.mce.2020.111031.
Czirok, S. J., Szabo, A., Lukats, A., and Szel, A.
54.
Wakhloo, D., Oberhauser, J., Madira, A., and
(2004) The system of cerebrospinal fluid-contacting
Mahajani, S. (2022) From cradle to grave: neurogenesis,
neurons. Its supposed role in the nonsynaptic signal
neuroregeneration
and neurodegeneration in
transmission of the brain, Histol. Histopathol., 19, 607-
Alzheimer’s and Parkinson’s diseases, Neural Regen.
628, doi: 10.14670/HH-19.607.
Res., 17, 2606-2614, doi: 10.4103/1673-5374.336138.
43.
Vigh-Teichmann, I., and Vigh, B. (1983) The system of
55.
Dilman, V. M. (1971) Age-associated elevation of
cerebrospinal fluid-contacting neurons, Arch. Histol.
hypothalamic, threshold to feedback control, and its
Japon., 46, 427-468, doi: 10.1679/AOHC.46.427.
role in development, ageine, and disease, Lancet, 1,
44.
Jalalvand, E., Robertson, B., Tostivint, H., Low, P.,
1211-1219, doi: 10.1016/s0140-6736(71)91721-1.
Wallen, P., and Grillner, S. (2018) Cerebrospinal
56.
Zwaan, B. J.
(2003) Linking development and
fluid-contacting neurons sense pH changes and
aging, Sci. Aging Knowledge Environ., 2003, pe32,
motion in the hypothalamus, J. Neurosci., 38, 7713-
doi: 10.1126/sageke.2003.47.pe32.
7724, doi: 10.1523/JNEUROSCI.3359-17.2018.
57.
Partridge, L., and Gems, D. (2002) A lethal side-
45.
Yildiz, S., Grinstead, J., Hildebrand, A., Oshinski, J.,
effect, Nature, 418, 921, doi: 10.1038/418921a.
Rooney, W. D., Lim, M. M., and Oken, B. (2022)
58.
Dilman, V. M.
(1994) Development, Aging and
Immediate impact of yogic breathing on pulsatile
Disease: A New Rationale for an Intervention Strategy,
cerebrospinal fluid dynamics, Sci. Rep., 12, 10894,
Routledge.
doi: 10.1038/s41598-022-15034-8.
59.
De Magalhães, J. P., and Church, G. M. (2005)
46.
Kedarasetti, R. T., Drew, P. J., and Costanzo, F.
Genomes optimize reproduction: aging as a con-
(2020) Arterial pulsations drive oscillatory flow of
sequence of the developmental program, Phys-
CSF but not directional pumping, Sci. Rep., 10, 10102,
iology (Bethesda),
20,
252-259, doi:
10.1152/
doi: 10.1038/s41598-020-66887-w.
physiol.00010.2005.
47.
Siyahhan, B., Knobloch, V., de Zelicourt, D.,
60.
Khan, N., Kim, S. K., Gagneux, P., Dugan, L. L.,
Asgari, M., Schmid Daners, M., Poulikakos, D., and
and Varki, A. (2020) Maximum reproductive lifespan
Kurtcuoglu, V. (2014) Flow induced by ependymal
correlates with CD33rSIGLEC gene number:
cilia dominates near-wall cerebrospinal fluid dynamics
Implications for NADPH oxidase-derived reactive
in the lateral ventricles, J. R. Soc. Interf., 11, 20131189,
oxygen species in aging, FASEB J., 34, 1928-1938,
doi: 10.1098/rsif.2013.1189.
doi: 10.1096/fj.201902116R.
БИОХИМИЯ том 87 вып. 12 2022
2032
ОЛОВНИКОВ
61. Hook, M., Roy, S., Williams, E. G., Bou Sleiman, M.,
67. Reznick, D., Bryant, M. J., and Bashey, F. (2002)
Mozhui, K., Nelson, J. F., Lu, L., Auwerx, J., and
R- and K-selection revisited: the role of population
Williams, R. W.
(2018) Genetic cartography of
regulation in life-history evolution, Ecology, 83, 1509-
longevity in humans and mice: current landscape and
1520, doi: 10.2307/3071970.
horizons, Biochim. Biophys. Acta. Mol. Basis Dis.,
68. McHugh, K. M., and Burke, M. K. (2022) From
1864, 2718-2732, doi: 10.1016/j.bbadis.2018.01.026.
microbes to mammals: the experimental evolution of
62. Capri, M., Santoro, A., Garagnani, P., Bacalini, M. G.,
aging and longevity across species, Evolution, 76, 692-
Pirazzini, C., Olivieri, F., Procopio, A., Salvioli, S.,
707, doi: 10.1111/evo.14442.
and Franceschi, C. (2014) Genes of human longevity:
69. Nagai, J., Lin, C. Y., and Sabour, M. P. (1995) Lines
an endless quest? Curr Vasc. Pharmacol., 12, 707-717,
of mice selected for reproductive longevity, Growth
doi: 10.2174/1570161111666131219110301.
Dev. Aging, 59, 79-91.
63. Brooks-Wilson, A. R. (2013) Genetics of healthy
70. Helle, S., Lummaa, V., and Jokela, J. (2005) Are
aging and longevity, Hum. Genet., 132, 1323-1338,
reproductive and somatic senescence coupled in
doi: 10.1007/s00439-013-1342-z.
humans? Late, but not early, reproduction correlated
64. Caruso, C., Ligotti, M. E., Accardi, G., Aiello, A.,
with longevity in historical Sami women, Proc. Biol.
Duro, G., Galimberti, D., and Candore, G. (2022) How
Sci., 272, 29-37, doi: 10.1098/rspb.2004.2944.
important are genes to achieve longevity? Int. J. Mol.
71. Campisi, J., Kapahi, P., Lithgow, G. J., Melov, S.,
Sci., 23, 5635, doi: 10.3390/ijms23105635.
Newman, J. C., and Verdin, E. (2019) From discoveries
65. Nuzhdin, S. V., Pasyukova, E. G., Dilda, C. L.,
in ageing research to therapeutics for healthy ageing,
Zeng, Z. B., and Mackay, T. F. (1997) Sex-specific
Nature, 571, 183-192, doi: 10.1038/s41586-019-1365-2.
quantitative trait loci affecting longevity in Drosophila
72. Bartke, A. (2022) Somatotropic axis, pace of life and
melanogaster, Proc. Natl. Acad. Sci. USA, 94, 9734-
aging, Front. Endocrinol. (Lausanne),
13,
916139,
9739, doi: 10.1073/pnas.94.18.9734.
doi: 10.3389/fendo.2022.916139.
66. Van Bodegom, D., May, L., Meij, H. J., and Westen-
73. Olovnikov, A. M. (2021) Role of the Earth’s Motions
dorp, R. G. (2007) Regulation of human life histories:
in plant orientation
- planetary mechanism,
the role of the inflammatory host response, Ann. N.Y.
Biochemistry (Moscow), 86, 1388-1394, doi: 10.1134/
Acad. Sci., 1100, 84-97, doi: 10.1196/annals.1395.007.
S0006297921110031.
PLANETARY METRONOME AS A REGULATOR
OF LIFESPAN AND AGING RATE: METRONOMIC HYPOTHESIS
A. M. Olovnikov
N. M. Emanuel Institute of Biochemical Physics, Russian Academy of Sciences,
119334 Moscow, Russia; E-mail: olovnikov@gmail.com
The metronome mechanism for controlling the duration of periods of the ontogenetic cycle of an animal
organism is proposed. The metronome system consists of a series of components. Among them: the ventricular
system of a brain, the planet Earth as a participant in the generation of metronome signals and the temporal
DNA (tDNA) as a substrate epigenetically labeled to account for the elapsed time of ontogeny. The metronome
system generates repetitive signals in the form of hydrodynamic perturbations in the cerebrospinal fluid (CSF).
The metronome effect arises due to the superposition of two processes - the near-wall unidirectional CSF
f low and oscillations in the motions of the planet. Hydrodynamic beats of the metronome are transformed into
nerve impulses that initiate acts of epigenetic modification of tDNA in neurons, changing the content of factors
expressed by this DNA for innervated targets of the body. Duration of the periods of the ontogenetic cycle,
including the duration of adult life, depends on the rate of tDNA modification acts. This rate depends mainly
on the frequency of the metronomic signals used by each particular biological species. But these epigenetic acts
can also be influenced by factors that can modulate metabolism and the rate of chromatin modifications, such
as a calorie-restricted diet, etc.
Keywords: ontogeny, aging, theories of aging, ventricular system of the brain, CSF-contacting neurons, hypothalamus,
Earth motions, nutations, polar motion
БИОХИМИЯ том 87 вып. 12 2022