БИОХИМИЯ, 2022, том 87, вып. 2, с. 278 - 288
УДК 577.25
ЭКСПРЕССИЯ ГЕНОВ ОПСИНОВ В СЕТЧАТКЕ ГЛАЗА САМОК
И САМЦОВ ТРЁХИГЛОЙ КОЛЮШКИ Gasterosteus aculeatus L.:
ЗАВИСИМОСТЬ ОТ ПРЕСНОВОДНОЙ АДАПТАЦИИ И ПРОЛАКТИНА
© 2022
Н.С. Павлова1*, А.Р. Гизатулина2, Т.В. Неретина3, О.В. Смирнова1
1 Московский государственный университет имени М.В. Ломоносова, биологический факультет,
кафедра физиологии человека и животных, 119991 Москва, Россия; электронная почта: pav.nad.ser@gmail.com
2 Московский государственный университет имени М.В. Ломоносова, факультет фундаментальной медицины,
кафедра физиологии и общей патологии, 119991 Москва, Россия
3 Московский государственный университет имени М.В. Ломоносова, биологический факультет,
Беломорская биологическая станция им. Н.А. Перцова; 186671 пос. Приморский,
Лоухский р4н, Карельская республика, Россия
Поступила в редакцию 01.01.2022
После доработки 24.01.2022
Принята к публикации 24.01.2022
Чувствительность цветового зрения играет важную роль в адаптивных процессах у рыб, в особенности - при
миграциях и в репродуктивном цикле. В обоих этих процессах у рыб важную функцию выполняет гормон
пролактин и его паралог, пролактин!подобный гормон. Для изучения возможного вклада пролактиновой
оси в пресноводную адаптацию цветового зрения рыб проведено исследование влияния пресноводной
адаптации в нерестовый период, а также введения пролактина на экспрессию генов опсинов (SWS1, SWS2,
RH2, LWS) в сетчатке глаза самок и самцов трёхиглой колюшки Gasterosteus aculeatus L. Показано, что у са!
мок, в отличие от самцов, при пресноводной адаптации растёт экспрессия гена пролактина!1 и также, как
у самцов, падает экспрессия гена пролактин!подобного гормона в мозге. При пресноводной адаптации, а
также при введении пролактина в условиях морской воды в сетчатке глаз самок и самцов снижается
экспрессия гена опсина SWS2, чувствительного в синей области спектра. В сетчатке глаза самцов при вве!
дении пролактина в условиях морской воды, но не при пресноводной адаптации снижается также экспрес!
сия гена опсина SWS1, чувствительного в ультрафиолетовой области спектра. Экспрессия других опсинов
при пресноводной адаптации не является пролактинзависимой ни у самок, ни у самцов. Можно заключить,
что экспрессия генов некоторых опсинов в сетчатке глаза трёхиглой колюшки регулируется пролактином и
может пролактинзависимо меняться в условиях пресноводной адаптации, что расширяет понимание адап!
тивной значимости пролактиновой оси для организма рыб при пресноводных миграциях.
КЛЮЧЕВЫЕ СЛОВА: пролактин, опсины, трёхиглая колюшка Gasterosteus aculeatus L., пресноводная адап!
тация, адаптация цветового зрения, зависимость от пола.
DOI: 10.31857/S0320972522020099
ВВЕДЕНИЕ
группой - 11!цис!изомером ретиналя, произ!
водным витамина А. Структура опсина опреде!
Цветовое зрение и его чувствительность в
ляет спектр поглощения колбочки - её спект!
разных областях спектра у позвоночных, в том
ральную чувствительность [1]. Опсины, ассоци!
числе у рыб, опосредовано наличием и соотно!
ированные с цветовым зрением, подразделяют!
шением зрительных пигментов в колбочках сет!
ся на четыре группы: SWS1 чувствителен в ульт!
чатки. В состав молекул!фоторецепторов входит
рафиолетово!синей области спектра, SWS2 - в
опсин, ковалентно связанный с хромофорной
синей, RH2 - в зелёной и LWS - в красной [2].
Изменение соотношения экспрессии генов раз!
личных опсинов сопровождается соответствую!
Принятые сокращения: Prl1 - пролактин!1; Prl2 -
пролактин!2 или пролактин!подобный гормон; oPrl - ове!
щим изменением зрительной чувствительности
чий пролактин; SWS1 - опсин колбочек, чувствительных в
в разных областях спектра [3, 4]. В сетчатке гла!
коротковолновой ультрафиолетово!синей области спект! за рыб описаны опсины всех типов, при этом на
ра; SWS2 - опсин колбочек, чувствительных в коротковол! их соотношение влияет ряд параметров, в осо!
новой синей области спектра; RH2 - опсин колбочек,
бенности экология вида и стадия репродуктив!
чувствительных в средневолновой зелёной области спект!
ра; LWS - опсин колбочек, чувствительных в длинновол!
ного цикла [5]. Исследования цихлид показали,
новой красной области спектра.
что виды, обитающие на большей глубине, бо!
* Адресат для корреспонденции.
лее чувствительны к коротковолновому, нежели
278
ОПСИНЫ СЕТЧАТКИ ГЛАЗА САМОК И САМЦОВ ТРЁХИГЛОЙ КОЛЮШКИ
279
к длинноволновому, излучению видимого
ванных с репродукцией, за исключением про!
спектра, что связано с превалированием соответ!
лактина [8, 10]. В сетчатке трёхиглой колюшки
ствующей части спектра [6]. Интенсивность
присутствуют опсины всех четырёх типов, не
экспрессии опсинов также зависит от прозрач!
имеющие паралогов, в отличие от большинства
ности воды: на флоридской лукании Lucania
других видов рыб, что приближает цветовое зре!
goodei показано, что у особей в прозрачной воде
ние колюшек по функциональным характерис!
экспрессия генов опсинов SWS1 и SWS2 выше, а
тикам к таковому у млекопитающих [14-16].
генов опсинов RH2 и LWS - ниже, чем у особей
В условиях пресноводной миграции в нерес!
в мутной воде. Эти данные воспроизводились в
товый период пролактин участвует в адаптации
экспериментальных условиях [7]. В случае, если
водно!солевого обмена колюшек, как и других
вид эвригалинный, то морская и пресноводная
видов рыб, к пресной воде. При этом у рыб
популяции также различаются по экспрессии
позднее был обнаружен паралог пролактина,
генов опсинов. В работах, посвящённых
которому присвоили название пролактин!по!
чувствительности цветового зрения трёхиглой
добный гормон или пролактин!2 (Prl2), в связи
колюшки Gasterosteus aculeatus L. из морских и
с чем обнаруженному ранее гормону присвоили
пресноводных популяций, показано, что у рези!
название пролактин!1 (Prl1). Ранее нами пока!
дентных морских колюшек более высокие уров!
зано, что при острой (24!часовой) пресновод!
ни экспрессии опсина SWS1 и LWS (последне!
ной адаптации экспрессия генов пролактиновой
го - только в лабораторных условиях), и более
оси в ткани мозга (вместе с гипофизом) самок и
низкие - опсина RH2 по сравнению с резидент!
самцов трёхиглой колюшки меняется неодина!
ными пресноводными колюшками [8]. Однако
ково, что свидетельствует о зависимой от пола
данные по пластичности цветового зрения у
пролактиновой регуляции у колюшек [17]. Опи!
проходных рыб, меняющих среду обитания с
сано влияние пролактина на сетчатку глаза мле!
морской на пресноводную в ходе жизненного
копитающих, однако его эффекты обычно свя!
цикла, отсутствуют.
зывают со стимуляцией васкуляризации, в част!
В условиях пресноводной миграции рыб,
ности во время развития сетчатки, и антиокси!
которая обычно связана с брачным периодом,
дантным эффектом, смягчающим проявления
адаптация цветового зрения может быть важ!
старения организма [18, 19]. В ряде исследова!
ным и даже необходимым условием для распоз!
ний показано наличие элементов пролактино!
навания полового партнёра или конкурента, ес!
вой оси в сетчатке рыб. Так, экспрессия пролак!
ли для вида характерно приобретение брачного
тина!2 была обнаружена в ганглионарном, внут!
окраса, что чаще встречается у самцов. Влияние
реннем и наружном ядерных слоях сетчатки
половых гормонов и репродуктивного периода
Danio rerio, и были доказаны его паракринные
на сетчатку показано для ряда рыб. Так, у самок
эффекты на сетчатку при эмбриональном раз!
цихлид, в частности Astatotilapia burtoni, в брач!
витии. Наружный ядерный слой представлен те!
ный период повышается пластичность и меня!
лами клеток!фоторецепторов: палочек и колбо!
ется чувствительность цветового зрения, опос!
чек. Таким образом, сетчатка рыб, как и прочих
редованная изменением экспрессии генов опси!
позвоночных, чувствительна к пролактину, и в
нов, с превалированием коротковолновых оп!
ней известны паракринные эффекты пролакти!
синов [9]. Для ряда видов рыб (трёхиглая ко!
на!2 [20].
люшка G. aculeatus, золотая рыбка Carassius aura4
Нами была выдвинута гипотеза о возмож!
tus, обыкновенная гамбузия Gambusia affinis) по!
ном и зависимом от пола участии пролактинов в
казано, что андрогены и эстрогены способны
адаптации к пресной воде не только водно!соле!
модулировать зрительную чувствительность к
вого обмена, но и цветового зрения колюшек.
длинноволновому излучению [10-12].
Для её проверки мы проанализировали измене!
Трёхиглая колюшка G. aculeatus L. является
ние экспрессии генов опсинов в сетчатке самок
перспективным объектом для изучения зависи!
и самцов трёхиглой колюшки в условиях острой
мой от пресноводной адаптации и пола особи
и хронической пресноводной адаптации, а так!
пластичности цветового зрения. Это связано, с
же при введении пролактина в условиях морс!
одной стороны, с эвригалинностью вида, с дру!
кой воды.
гой - с наличием сложного репродуктивного
цикла, который сопровождается приобретением
самцами брачного окраса и миграцией части
МАТЕРИАЛЫ И МЕТОДЫ
морской популяции в пресноводные водоё!
мы [13]. Для трёхиглой колюшки было показано
Экспериментальная модель. Работа выполне!
влияние на цветовое зрение как экологических
на на группах половозрелых самок и самцов трёх!
условий, так и некоторых гормонов, ассоцииро!
иглой колюшки G. aculeatus L. морской популя!
БИОХИМИЯ том 87 вып. 2 2022
280
ПАВЛОВА и др.
ции, находящихся в нерестовом состоянии. Жи!
случайные праймеры («Евроген»). Для проведе!
вотных (длина тела самок составила 7 ± 0,32 см,
ния полимеразной цепной реакции (ПЦР) в ре!
самцов - 6,5 ± 0,3 см) отлавливали в Кандалакш!
жиме реального времени использовали ампли!
ском заливе Белого моря в июне 2020 г. Особи
фикатор Bio!Rad CFX96 («Bio!Rad», Канада) и
были разделены по полу, самцы также разделе!
набор реакционных смесей с красителем
ны по интенсивности окраски на альфа! и оме!
SYBRgreen и низкой концентрацией референс!
га!самцов. Внутри этих групп были сформиро!
ного красителя ROX («Евроген») и специфич!
ваны четыре экспериментальные группы: конт!
ные праймеры (таблица). Режим амплифика!
рольная «К» (морская популяция), группа ост!
ции: 95 °С - 5 мин; 95 °С - 15 с, 60 °С - 20 с,
рой пресноводной адаптации «24 часа», группа
72 °С - 20 с, 40 циклов; кривая плавления
хронической пресноводной адаптации «72 ча!
60 - 95 °С, инкремент 0,5 °С - 5 с. Каждую реак!
са», группа с введением морской популяции
цию с использованием кДНК проводили в трёх
овечьего пролактина «oPrl», n = 6 для каждой
независимых экспериментах; для каждого ис!
группы. Группы альфа! и омега!самцов в даль!
следуемого гена также проводили безревертаз!
нейшем были объединены, поскольку ни по од!
ные контроли амплификации фрагмента геном!
ному из изучаемых параметров между ними не
ной ДНК. Если безревертазный контроль был
было обнаружено статистически значимых раз!
отрицательным, значения ПЦР с матрицы
личий, таким образом, для групп самцов n = 12.
кДНК использовали для расчётов. Подробно
Перед началом эксперимента все особи были
методика описана Pierce et al. [24]. При проведе!
адаптированы к условиям содержания (аквариу!
нии ПЦР в реальном времени значения
мы объёмом 20 литров с непрозрачными стенка!
экспрессии генов интереса Prl1 и Prl2 нормиро!
ми, световой и температурный режим соответ!
вали на уровень экспрессии референсных ге!
ствовали естественным условиям) в течение
нов Rpl13α и Ubc [25]. Значения экспрессии ге!
24 ч. Особи группы «К» находились в течение
нов интереса (SWS1, SWS2, RH2 и LWS) норми!
всего эксперимента в условиях морской воды.
ровали на уровень экспрессии референсных ге!
Особи группы «24 часа» были помещены на 24 ч
нов Ubc и GNAT2 [26]. Для расчётов использова!
в пресную воду. Особи группы «72 часа» были
ли формулу, применявшуюся в предыдущих ра!
помещены в пресную воду на 72 ч. Особи груп!
ботах [17].
пы «oPrl» находились в течение 72 ч в условиях
Статистический аналих данных. Статисти!
морской воды, но один раз в сутки им произво!
ческую обработку проводили в програм!
дили внутрибрюшинную инъекцию 50 мкл фи!
ме GraphPad Prism 8 с использованием one!
зиологического раствора с овечьим пролакти!
way ANOVA (однофакторные сравнения) и two!
ном («National hormone and peptide program»,
way ANOVA (двухфакторные сравнения). В обо!
США) в концентрации 14 МЕ/мл. Овечий про!
их случаях использовали тест Даннетта. На гра!
лактин был выбран для применения в экспери!
фиках для каждой группы данные представлены
менте, поскольку широко используется в рабо!
в виде медианы, нижней и верхней квартили, а
тах по изучению влияния пролактина на осмо!
также минимального и максимального значе!
регуляцию и репродукцию рыб и связывается с
ний.
обоими рецепторами пролактина рыб с доста!
точно высоким сродством, хотя и меньшим, чем
гомологичный пролактин [21-23]. Группу поло!
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
жительного контроля, которой вводили физио!
логический раствор в условиях морской воды, в
Влияние адаптации к пресной воде на экс@
эксперименте не использовали, так как наши
прессию генов пролактинов в ткани мозга самок
предыдущие исследования показали, что по
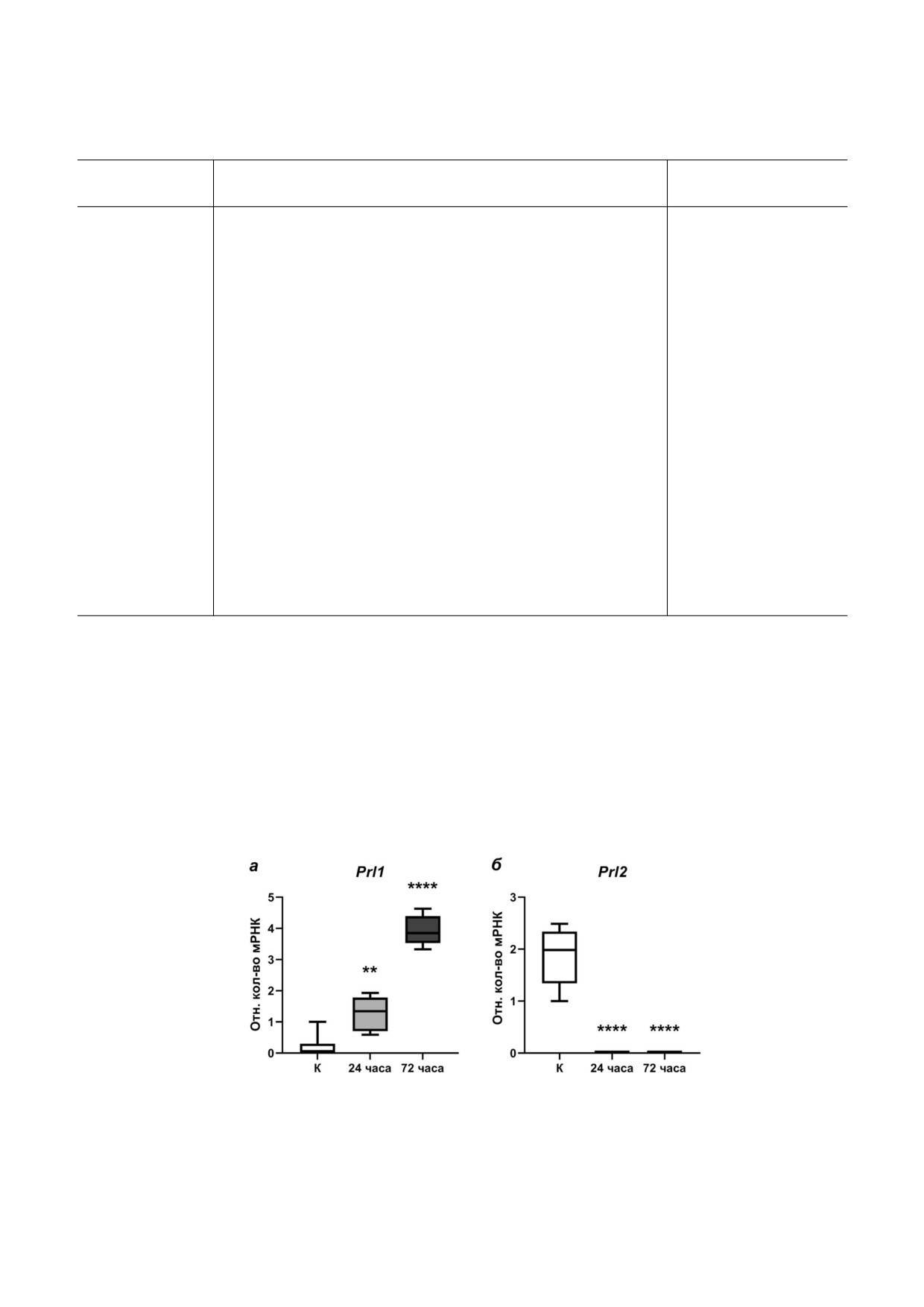
G. aculeatus L. Экспрессия гена пролактина!1 в
изучаемым параметрам особи не отличаются от
ткани мозга самок трёхиглой колюшки повыша!
контрольных. По завершении эксперимента у
лась в ходе адаптации к пресной воде: относи!
особей удаляли мозг вместе с гипофизом (для
тельные количества мРНК гена Prl1 были досто!
контроля экспрессии генов пролактинов), а так!
верно выше в мозге самок как после 24, так и
же ткань сетчатки; образцы фиксировали в
после 72 ч адаптации по сравнению с контроль!
IntactRNA («Евроген», Россия).
ной группой (рис. 1, а). Экспрессия гена про!
Обработка материала. Из ткани мозга (с ги!
лактина!2 в ткани мозга самок, напротив, сни!
пофизом) и сетчатки выделяли тотальную РНК
жалась: мРНК гена Prl2 не была обнаружена в
фенол!хлороформным методом с полиакрил!
мозге самок ни после 24, ни после 72 ч адапта!
амидным осаждением нуклеиновых кислот, ис!
ции к пресной воде, что статистически отлича!
пользуя набор ExtractRNA («Евроген»). Для
лось от данного параметра у контрольной груп!
синтеза кДНК использовали MMLV!ревертазу и
пы (рис. 1, б).
БИОХИМИЯ том 87 вып. 2 2022
ОПСИНЫ СЕТЧАТКИ ГЛАЗА САМОК И САМЦОВ ТРЁХИГЛОЙ КОЛЮШКИ
281
Нуклеотидные последовательности прямого и обратного праймеров генов интереса и референсных генов, а также длина
ПЦР!продукта
Ген
Нуклеотидная последовательность прямого (for) и обратного (rev)
Длина ПЦР!продукта, п.н.
праймеров, 5′-3′
Prl1
for - ACCTGGACTCGCATTTGCCTCTC
125
rev - AAGGTCCGACTCTGGTACTTGAAG
Prl2
for - TCCAATAAAGCCCTAGAGATGAG
113
rev - AGGCTGCTGACGGTGTTGCTTAT
SWS1
for - TGGAGCAGGTACATCCCTGA
184
rev - GAGCTGCTACAGCTCGAAGA
SWS2
for - GCAAGCCGCTTGGTAACTTC
124
rev - TCTGGGATGTACCTGCTCC
RH2
for - GTACCTTCCTGAGGGCATG
135
rev - GGCTGCGGCAGCTTTGACT
LWS
for - GCTGCGGCTAACCCTGGA
126
rev - ACATGAACGGAACTGCCGG
Rpl13α
for - CACCTTGGTCAACTTGAACAGTG
178
rev - TCCCTCCGCCCTACGAC
Ubc
for - AGACGGGCATAGCACTTGC
180
rev - CAGGACAAGGAAGGCATCC
GNAT2
for - GTTACTGCTTGGTGCTGGTG
211
rev - CTTCTGTGCATTCTCCTGTGA
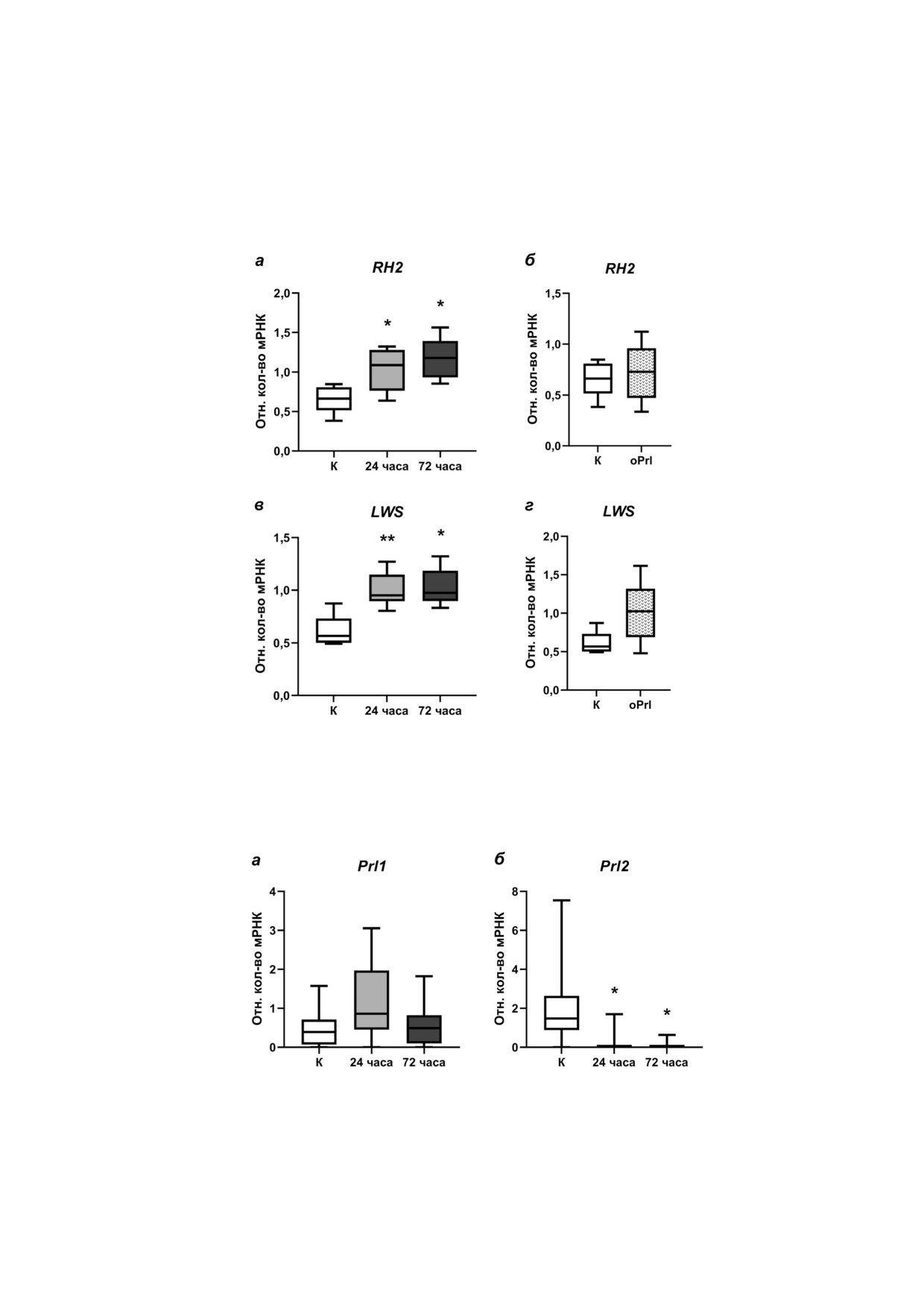
Влияние 24@ и 72@часовой адаптации к прес@
ше по сравнению как с контрольной группой,
ной воде и экзогенного пролактина на экспрессию
так и с группой 72!часовой пресноводной адап!
генов опсинов в ткани сетчатки самок G. acule
тации. Относительные количества мРНК ге!
atus L. Экспрессия гена SWS1, кодирующего оп!
на SWS1 в контрольной группе и группе, пре!
син, чувствительный в ультрафиолетово!синей
терпевшей пресноводную адаптацию в течение
области спектра, в ткани сетчатки глаз самок
72 ч, были на сопоставимом уровне (рис. 2, а).
трёхиглой колюшки в ходе пресноводной адап!
Ежедневное введение овечьего пролактина в те!
тации менялась неодинаково. После 24!часовой
чение 72 ч в условиях морской воды не оказало
адаптации к пресной воде экспрессия ге!
достоверного влияния на экспрессию гена SWS1
на SWS1 в сетчатке самок была достоверно вы!
в ткани сетчатки самок колюшек (рис. 2, б).
Рис. 1. Экспрессия мРНК генов Prl1 (а) и Prl2 (б) в ткани мозга самок трёхиглой колюшки в условиях морской воды (конт!
роль «К» - незакрашенные боксы) и после 24! и 72!часовой адаптации к пресной воде (боксы серого и тёмно!серого цве!
та соответственно), ** p < 0,01; **** p < 0,0001 - статистически значимые различия по сравнению с группой «К» (one!way
ANOVA; n = 6 в каждой из групп)
9 БИОХИМИЯ том 87 вып. 2 2022
282
ПАВЛОВА и др.
Рис. 2. Экспрессия мРНК генов опсинов, чувствительных в коротковолновой области спектра, в сетчатке глаз самок трёх!
иглой колюшки: SWS1 (а, б), SWS2 (в, г). а и в - В условиях морской воды в контроле «К» (незакрашенные боксы) и пос!
ле 24! и 72!часовой адаптации к пресной воде (боксы серого и тёмно!серого цвета соответственно); б и г - в условиях
морской воды в контроле «К» (незакрашенные боксы) и после внутрибрюшинных инъекций овечьего пролактина в тече!
ние 72 ч «oPrl» (пёстрые боксы), * p < 0,05; ** p < 0,01 - статистически значимые различия по сравнению с группой «К»
(one!way ANOVA; n = 6 в каждой из групп)
Экспрессия гена SWS2, кодирующего оп!
ходе пресноводной адаптации повышалась: от!
син, чувствительный в синей области спектра, в
носительные количества мРНК гена LWS были
ткани сетчатки глаз самок трёхиглой колюшки в
достоверно выше в группах 24! и 72!часовой
ходе пресноводной адаптации уменьшалась: от!
пресноводной адаптации по сравнению с конт!
носительные количества мРНК гена SWS2 были
рольной группой (рис. 3, в). В условиях морской
достоверно ниже в группах 24! и 72!часовой
воды после введения экзогенного пролактина
пресноводной адаптации по сравнению с конт!
экспрессия гена LWS в сетчатке росла недосто!
рольной группой (рис. 2, в). В условиях морской
верно (рис. 3, г).
воды на фоне введения пролактина экспрессия
Сравнение экспрессии исследуемых генов у α@
гена SWS2 в сетчатке также была на более низ!
и ω@самцов G. aculeatus L. в контрольной и экспе@
ком уровне по сравнению с контрольной груп!
риментальных группах. По экспрессии всех генов
пой (рис. 2, г).
интереса (Prl1 и Prl2 в ткани мозга с гипофизом,
Экспрессия гена RH2, кодирующего опсин,
SWS1, SWS2, RH2 и LWS в ткани сетчатки) не!
чувствительный в зелёной области спектра, в
парный t!test не выявил достоверных различий
ткани сетчатки глаз самок трёхиглой колюшки в
между α! и ω!самцами ни в группе контроля
ходе пресноводной адаптации повышалась: от!
«К», ни в группах 24!часовой «24 часа» и 72!ча!
носительные количества мРНК гена RH2 были
совой «72 часа» пресноводной адаптации, ни в
достоверно выше в группах 24! и 72!часовой
группах особей, которым в условиях морской
пресноводной адаптации по сравнению с конт!
воды вводили овечий пролактин (oPrl) (p > 0,1).
рольной группой (рис. 3, а). В условиях морской
Поэтому группы α! и ω!самцов объединены, и
воды на фоне введения пролактина уровень
таким образом, в группах самцов размер выбор!
экспрессии гена RH2 в сетчатке был сопоставим
ки составил 12 особей.
с контрольной группой (рис. 3, б).
Влияние адаптации к пресной воде на экс@
Экспрессия гена LWS, кодирующего опсин,
прессию генов пролактинов в ткани мозга самцов
чувствительный в красной области спектра, в
G. aculeatus L. Экспрессия гена пролактина!1 в
ткани сетчатки глаз самок трёхиглой колюшки в
ткани мозга самцов трёхиглой колюшки не ме!
БИОХИМИЯ том 87 вып. 2 2022
ОПСИНЫ СЕТЧАТКИ ГЛАЗА САМОК И САМЦОВ ТРЁХИГЛОЙ КОЛЮШКИ
283
нялась в ходе адаптации к пресной во!
Влияние 24@ и 72@часовой адаптации к прес@
де (рис. 4, а). Экспрессия гена пролактина!2 в
ной воде и экзогенного пролактина на экспрессию
ткани мозга самцов достоверно снижалась как
генов опсинов в ткани сетчатки самцов G. aculea
после 24!, так и после 72!часовой адаптации к
tus L. Экспрессия гена SWS1, кодирующего оп!
пресной воде (рис. 4, б).
син, чувствительный в ультрафиолетово!синей
Рис. 3. Экспрессия мРНК генов опсинов, чувствительных в длинноволновой области спектра, в сетчатке глаз самок трёх!
иглой колюшки: RH2 (а, б), LWS (в, г). а и в - В условиях морской воды в контроле «К» (незакрашенные боксы) и после
24! и 72!часовой адаптации к пресной воде (боксы серого и тёмно!серого цвета соответственно); б и г - в условиях морс!
кой воды в контроле «К» (незакрашенные боксы) и после внутрибрюшинных инъекций овечьего пролактина в течение
72 ч «oPrl» (пёстрые боксы), * p < 0,05; ** p < 0,01 - статистически значимые различия по сравнению с группой «К» (one!
way ANOVA; n = 6 в каждой из групп)
Рис. 4. Экспрессия мРНК генов Prl1 (а) и Prl2 (б) в ткани мозга самцов трёхиглой колюшки в условиях морской во!
ды «К» (незакрашенные боксы) и после 24! и 72!часовой адаптации к пресной воде (боксы серого и тёмно!серого цвета
соответственно), * p < 0,05 - статистически значимые различия по сравнению с группой «К» (one!way ANOVA; n = 12
в каждой из групп)
БИОХИМИЯ том 87 вып. 2 2022
9*
284
ПАВЛОВА и др.
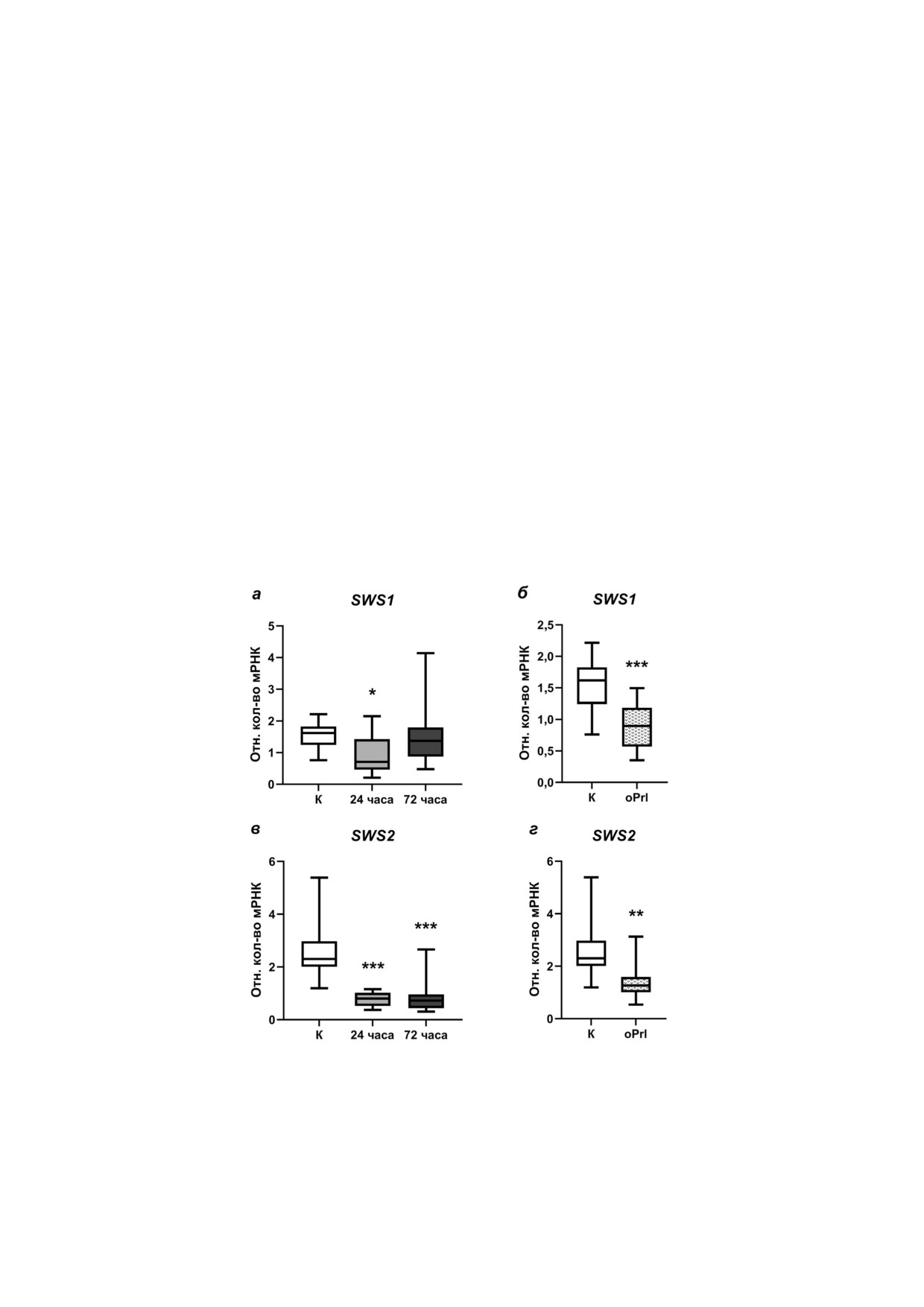
области спектра, в ткани сетчатки глаз самцов
морской воды после введения экзогенного про!
трёхиглой колюшки в ходе пресноводной адап!
лактина экспрессия гена SWS2 в сетчатке сам!
тации менялась неодинаково. После 24!часо!
цов также была на более низком уровне по срав!
вой адаптации к пресной воде экспрессия ге!
нению с контрольной группой (рис. 5, г).
на SWS1 в сетчатке самцов была достоверно ни!
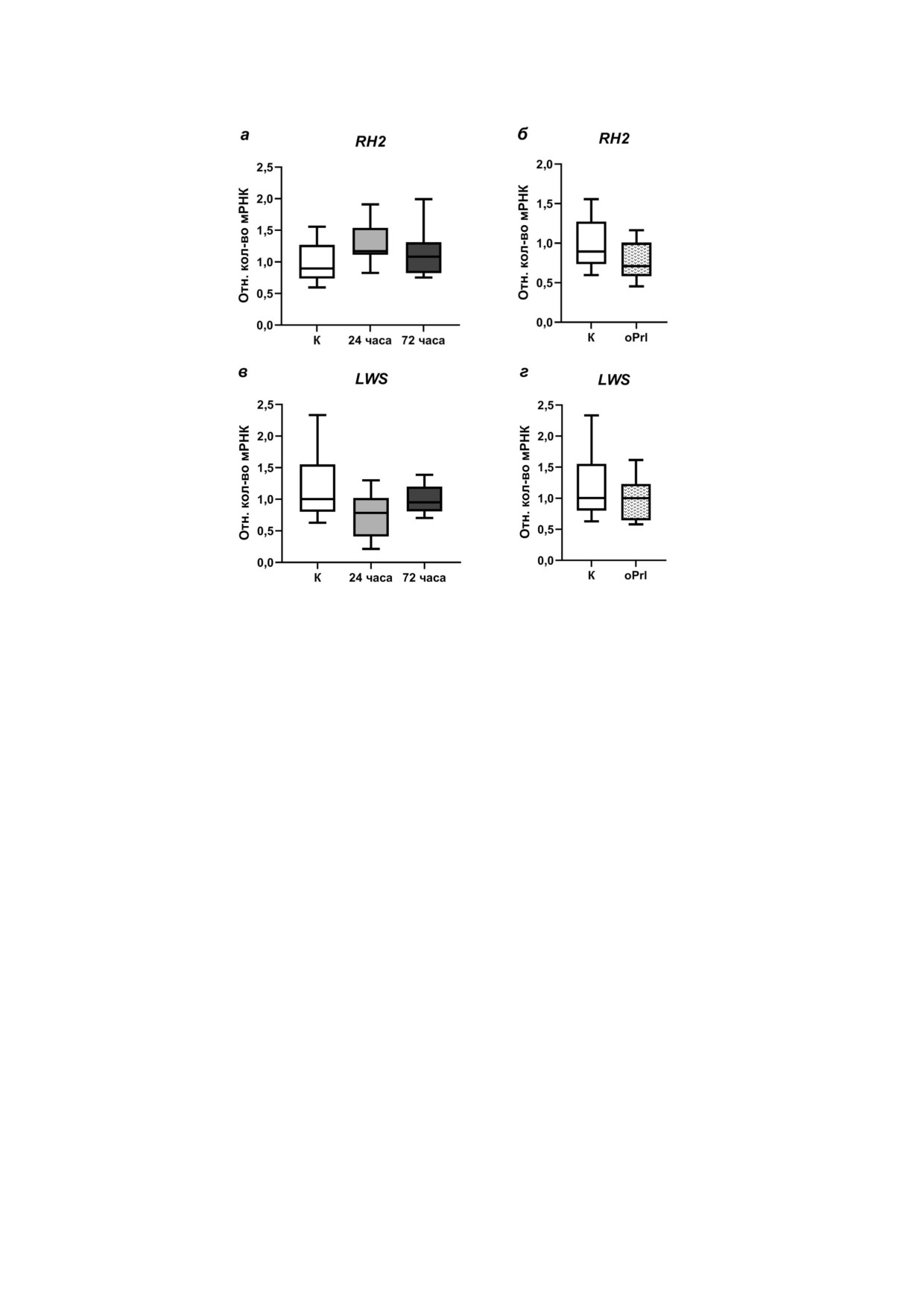
Экспрессия генов RH2 и LWS, кодирующих
же по сравнению как с контрольной группой,
опсины, чувствительные в зелёной и красной
так и с группой 72!часовой пресноводной адап!
области спектра соответственно, в ткани сетчат!
тации. Относительные количества мРНК ге!
ки глаз самцов трёхиглой колюшки в ходе прес!
на SWS1 в контрольной группе и группе, пре!
новодной адаптации не менялась (рис. 6, a и в).
терпевшей пресноводную адаптацию в течение
В условиях морской воды после введения про!
72 ч, были на сопоставимом уровне (рис. 5, а).
лактина экспрессия генов RH2 и LWS в сетчатке
После ежедневного введения пролактина в те!
была сопоставима с их экспрессией в контроль!
чение 72 ч относительные количества мРНК ге!
ной группе (рис. 6, б и г).
на SWS1 в сетчатке самцов были достоверно
ниже по сравнению с контрольной груп!
пой (рис. 5, б).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Экспрессия гена SWS2, кодирующего оп!
син, чувствительный в синей области спектра, в
Изменения уровней экспрессии генов Prl1
ткани сетчатки глаз самцов трёхиглой колюшки
и Prl2 в мозге трёхиглой колюшки в условиях
в ходе пресноводной адаптации уменьшалась:
пресноводной адаптации зависят от пола. Из!
относительные количества мРНК гена SWS2
вестно, что при пресноводной адаптации про!
были достоверно ниже в группах 24! и 72!часо!
лактиновая ось рыб стимулируется, однако эти
вой пресноводной адаптации по сравнению с
эффекты были обнаружены и изучены на сме!
контрольной группой (рис. 5, в). В условиях
шанных выборках, без разделения особей по
Рис. 5. Экспрессия мРНК генов опсинов, чувствительных в коротковолновой области спектра, в сетчатке глаз самцов трёх!
иглой колюшки: SWS1 (а, б), SWS2 (в, г). а и в - В условиях морской воды в контроле «К» (незакрашенные боксы) и пос!
ле 24! и 72!часовой адаптации к пресной воде (боксы серого и тёмно!серого цвета соответственно); б и г - в условиях
морской воды в контроле «К» (незакрашенные боксы) и после внутрибрюшинных инъекций овечьего пролактина в тече!
ние 72 ч «oPrl» (пёстрые боксы), * p < 0,05; ** p < 0,01; *** p < 0,001 - статистически значимые различия по сравнению с
группой «К» (one!way ANOVA; n = 12 в каждой из групп)
БИОХИМИЯ том 87 вып. 2 2022
ОПСИНЫ СЕТЧАТКИ ГЛАЗА САМОК И САМЦОВ ТРЁХИГЛОЙ КОЛЮШКИ
285
Рис. 6. Экспрессия мРНК генов опсинов, чувствительных в длинноволновой области спектра, в сетчатке глаз самцов трёх!
иглой колюшки: RH2 (а, б), LWS (в, г). а и в - В условиях морской воды в контроле «К»(незакрашенные боксы) и после
24! и 72!часовой адаптации к пресной воде (боксы серого и тёмно!серого цвета соответственно); б и г - в условиях морс!
кой воды в контроле «К» (незакрашенные боксы) и после внутрибрюшинных инъекций овечьего пролактина в течение
72 ч «oPrl» (пестрые боксы). n = 12 в каждой из групп
полу [27-29]. Мы обнаружили различия в мо!
наблюдали снижение экспрессии гена Prl2 в
дификации экспрессии генов пролактинов в ус!
мозге (рис. 1, б и 4, б). Следовательно, измене!
ловиях пресноводной адаптации у самок и сам!
ния экспрессии Prl1 (повышение) и Prl2 (сни!
цов. Ранее нами были обнаружены модифика!
жение) в мозге самок трёхиглой колюшки были
ции пролактиновой оси колюшек в условиях
разнонаправленными в условиях как острой, так
пресноводной адаптации, различные у самок и
и хронической пресноводной адаптации, а в
самцов [17]. В этой связи влияние пресновод!
мозге самцов в этих условиях менялась только
ной адаптации, а также эффекты вводимого
экспрессия Prl2 (снижение).
пролактина на сетчатку глаза описаны для са!
Известно, что оценку чувствительности
мок и самцов по отдельности. Поскольку у са!
цветового зрения можно проводить, измеряя со!
мок, в отличие от самцов, экспрессия гена Prl1
отношения экспрессии генов различных опси!
в ткани мозга росла как при острой (24 ч), так и
нов [3, 4]. Для проверки гипотезы о пролактин!
при хронической (72 ч) адаптации к пресной
зависимом изменении чувствительности зрения
воде, мы предполагаем больший вклад пролак!
в условиях пресноводной адаптации мы сравни!
тина!1 в адаптацию организма самок, но не
ли относительные уровни мРНК генов опсинов
самцов к пресноводным условиям (рис. 1, а
у самок и самцов трёхиглой колюшки в услови!
и 4, а). Таким образом, пролактиновая ось са!
ях острой и хронической пресноводной адапта!
мок трёхиглой колюшки стимулировалась при
ции с введением им пролактина в условиях
пресноводной адаптации сходным с другими
морской воды. Использованный овечий про!
видами рыб образом при исследованиях без
лактин имитирует эффекты обоих пролактинов
разделения по полу (зебрафиш Danio rerio, мо!
рыб, что было показано на тилапии O. mossambi4
замбикская тилапия Oreochromis mossambicus и
cus: оба пролактина тилапии, как и овечий про!
серебристый горбыль Argyrosomus regius) [30-32].
лактин, связывались пролактинчувствительны!
Одновременно с этим и у самок, и у самцов мы
ми тканями [33].
БИОХИМИЯ том 87 вып. 2 2022
286
ПАВЛОВА и др.
Экспрессия гена SWS1 у самок трёхиглой ко@
ной адаптации отсутствие эффектов, оказывае!
люшки в условиях пресноводной адаптации не на@
мых на сетчатку пролактином, может быть объ!
ходится под контролем пролактина, в отличие от
яснено отсутствием увеличения пролактиновой
самцов. Различный ответ пролактинов (Prl1
регуляции сетчатки (рис. 4, а и б). Таким обра!
и Prl2) самок и самцов трёхиглой колюшки на
зом, ген SWS1, кодирующий коротковолновый
острую и хроническую пресноводную адапта!
опсин SWS1, в сетчатке глаза регулируется про!
цию частично подтверждается отсутствием у са!
лактином у самцов, но не у самок трёхиглой ко!
мок и наличием у самцов пролактин!индуциро!
люшки.
ванного ингибирования экспрессии гена SWS1,
Снижение экспрессии гена SWS2 у самок и
кодирующего коротковолновый опсин SWS1,
самцов трёхиглой колюшки в условиях пресно@
чувствительный в ультрафиолетово!синей об!
водной адаптации может индуцироваться пролак@
ласти спектра, в ткани сетчатки (рис. 2, б и 5, б).
тином. Экспрессия гена SWS2, кодирующего
Что примечательно, в сетчатке глаза самок ко!
опсин, чувствительный в синей области спект!
люшек после острой адаптации к пресной воде
ра, в сетчатке глаза трёхиглой колюшки меня!
экспрессия гена SWS1 повышалась, но в группе
лась схожим образом у самок и самцов как в мо!
хронической адаптации этот показатель был на
дели пресноводной адаптации, так и после вве!
уровне, сопоставимом с контрольной группой.
дения пролактина (рис. 2, в и г; 5, в и г). Как у
Введение пролактина не оказывало влияния на
самок, так и у самцов при острой и хроничес!
экспрессию гена SWS1 (рис. 2, а и б). Мы свя!
кой пресноводной адаптации экспрессия ге!
зываем подобный эффект с возможной регуля!
на SWS2 в сетчатке глаза снижалась. Аналогич!
цией экспрессии опсина SWS1 в сетчатке глаз
ный эффект оказало введение пролактина в ус!
самок колюшек со стороны кортизола, являю!
ловиях морской воды. Таким образом, можно с
щегося ключевым гормоном острой пресновод!
высокой долей вероятности утверждать, что
ной адаптации у рыб [31]. В ряде работ на мле!
снижение чувствительности цветового зрения
копитающих описано влияние кортизола на
самок и самцов трёхиглой колюшки к синей об!
большинство структур глаза, включая сетчатку
ласти спектра в условиях пресноводной адапта!
и фоторецепторные клетки [34]. Однако подоб!
ции опосредовано пролактином. Также нельзя
ное предположение требует дальнейшего более
исключать возможный эффект кортизола, ко!
детального изучения. В сетчатке глаза самцов
торый также классически ассоциируют с прес!
острая пресноводная адаптация сопровожда!
новодной адаптацией рыб [31]. Однако данное
лась снижением экспрессии гена SWS1, в отли!
предположение требует более подробного изу!
чие от самок, у которых наблюдалось повыше!
чения.
ние его экспрессии (рис. 2, а и 5, а). Изменение
Повышение экспрессии генов RH2 и LWS у
экспрессии гена SWS1 у самцов частично согла!
самок и самцов трёхиглой колюшки в условиях
суется с литературными данными для общей
пресноводной адаптации не опосредовано
популяции: по данным других исследователей
пролактином. Экспрессия генов RH2 и LWS,
экспрессия гена SWS1 в ткани сетчатки была
кодирующих длинноволновые опсины RH2 и
выше у резидентных морских колюшек в срав!
LWS, чувствительные в зелёной и красной об!
нении с резидентными пресноводными [8]. Од!
ластях спектра соответственно, в сетчатке глаз
новременно с этим при хронической пресно!
самок трёхиглой колюшки повышалась при
водной адаптации экспрессия гена SWS1 в сет!
острой и хронической пресноводной адапта!
чатке самцов колюшек не отличалась от тако!
ции (рис. 3, а и в), что говорит в пользу повы!
вой в контрольной группе. Поскольку эффект
шения чувствительности цветового зрения к
пресноводной адаптации на экспрессию данно!
длинноволновому излучению. Однако, по!
го гена наблюдается только в первые 24 ч, мы
скольку этот эффект не воспроизводился при
предполагаем его пролактиннезависимый ха!
введении пролактина в условиях морской во!
рактер в модели пресноводной адаптации и воз!
ды (рис. 3, б и г), мы предполагаем пролактин!
можное влияние кортизола на экспрессию ге!
независимый характер изменения экспрессии
на SWS1 у самцов. Несмотря на отсутствие эф!
данных опсинов. Изменение экспрессии дан!
фекта 72!часовой пресноводной адаптации на
ных опсинов при пресноводной адаптации мо!
экспрессию гена SWS1 в сетчатке глаза самцов
жет быть результатом влияния эстрогенов, для
колюшек, введение экзогенного пролактина в
которых ранее был показан стимулирующий
течение
72
ч приводило к её снижению
эффект на экспрессию RH2 и LWS в сетчатке
(рис. 5, б). Подобный эффект может быть объ!
обыкновенной гамбузии Gambusia affinis [11].
яснён тем, что при переходе в пресную воду
У самцов, в отличие от самок колюшек, экс!
экспрессия генов Prl1 и Prl2 у самцов не повы!
прессия генов RH2 и LWS в сетчатке глаз не ме!
шалась, и, следовательно, в модели пресновод!
нялась при адаптации к пресной воде, и, как и у
БИОХИМИЯ том 87 вып. 2 2022
ОПСИНЫ СЕТЧАТКИ ГЛАЗА САМОК И САМЦОВ ТРЁХИГЛОЙ КОЛЮШКИ
287
самок, не менялась в условиях морской воды
на SWS2, в то время как экспрессия генов ос!
после введения пролактина (рис. 6). С одной
тальных опсинов не меняется. Тем не менее по!
стороны, это доказывает пролактиннезависи!
добное изменение транскрипционной актив!
мый характер регуляции длинноволновых опси!
ности сетчатки самцов также может способство!
нов у самок и самцов трёхиглой колюшки, с дру!
вать итоговому повышению чувствительности к
гой - может подтверждать гипотезу о возмож!
длинноволновой части спектра.
ном положительном влиянии эстрогенов на эти
Можно заключить, что адаптивные эффек!
опсины у самок, но оставляет открытым вопрос
ты пролактина при пресноводных миграциях
об опосредованном половыми гормонами, а не
трёхиглой колюшки проявляются не только в
пролактином, изменении цветового зрения рыб
регуляции водно!солевого обмена, но также
в условиях пресноводной адаптации.
цветового зрения, и они могут различаться у са!
мок и самцов.
Таким образом, цветовое зрение у самок и
самцов трёхиглой колюшки в условиях пресно!
водной адаптации меняется неодинаково. Для
Финансирование. Исследование выполнено
самок при хронической пресноводной адапта!
в рамках научного проекта государственного за!
ции показано пролактинзависимое снижение
дания МГУ № 121032300075!6 и при финансо!
экспрессии гена SWS2 и пролактиннезависимое
вой поддержке Российского фонда фундамен!
повышение экспрессии генов RH2 и LWS. Соче!
тальных исследований (грант № 18!34!00734).
танное повышение чувствительности зрения са!
Благодарности. Авторы выражают благодар!
мок к длинноволновой области спектра и сни!
ность Николаю Сергеевичу Мюге за плодотвор!
жение чувствительности в коротковолновой
ное обсуждение данной работы.
способствуют лучшему распознаванию красно!
Конфликт интересов. Авторы заявляют об
го и зелёного цвета, что может иметь ключевое
отсутствии конфликта интересов.
значение для распознавания полового партнёра
Соблюдение этических норм. Все примени!
во время нереста. Изменение цветового зрения
мые международные, национальные и/или инс!
самцов при пресноводной адаптации сопряже!
титуциональные принципы ухода и использова!
но с уменьшением чувствительности в коротко!
ния животных были соблюдены. Все процедуры
волновой части спектра, опосредованным про!
были одобрены комиссией МГУ по биоэтике
лактинзависимым снижением экспрессии ге!
(№ протокола 98a; № собрания комиссии 108!o).
СПИСОК ЛИТЕРАТУРЫ
1.
Nathans, J. (1989) The genes for color vision, Sci. Am.,
9. Butler, J. M., and Maruska, K. P. (2021) Opsin expression
260, 42!49.
varies with Reproductive state in the cichlid fish
2.
Fernald, R. D. (1993) Vision, in The Physiology of Fishes
Astatotilapia burtoni, Integr. Comp. Biol., 61, 240!248.
(Evans, D. H., ed.) CRC Press Inc, USA, Florida, pp. 161!
10. Shao, Y. T., Wang, F. Y., Fu, W. C., Yan, H. Y., Anraku, K.,
189.
et al. (2014) Androgens increase lws opsin expression and
3.
Carleton, K. L., and Kocher, T. D. (2001) Cone opsin
red sensitivity in male three!spined sticklebacks, PLoS
genes of African cichlid fishes: tuning spectral sensitivity by
One, 9, e100330.
differential gene expression, Mol. Biol. Evol., 18, 1540!
11. Friesen, C. N., Ramsey, M. E., and Cummings, M. E.
1550.
(2017) Differential sensitivity to estrogen!induced opsin
4.
Parry, J. W., Carleton, K. L., Spady, T., Carboo, A., Hunt,
expression in two poeciliid freshwater fish species, Gen.
D. M., et al. (2005) Mix and match color vision: Tuning
Comp. Endocrinol., 246, 200!210.
spectral sensitivity by differential opsin gene expression in
12. Yue, S., Wadia, V., Sekula, N., Dickinson, P. S., and
Lake Malawi cichlids, Curr. Biol., 15, 1734!1739.
Thompson, R. R. (2018) Acute effects of sex steroids
5.
Cortesi, F., Mitchell, L. J., Tettamanti, V., Fogg, L. G., de
on visual processing in male goldfish, J. Comp.
Busserolles, F., et al. (2020) Visual system diversity in coral
Physiol. A Neuroethol. Sens. Neural. Behav. Physiol., 204,
reef fishes, Semin. Cell Dev. Biol., 106, 31!42.
17!29.
6.
Terai, Y., Miyagi, R., Aibara, M., Mizoiri, S., Imai, H.,
13. Bell, M. A., and Foster, S. A. (1994) Introduction to the
et al. (2017) Visual adaptation in Lake Victoria cichlid
evolutionary biology of the threespine stickleback, Evol.
fishes: Depth!related variation of color and scotopic
Biol. Threespine Stickleback, 1, 27.
opsins in species from sand/mud bottoms, BMC Evol.
14. Rennison, D. J., Owens, G. L., and Taylor, J. S. (2012)
Biol., 17, 200.
Opsin gene duplication and divergence in ray!finned fish,
7.
Fuller, R. C., Carleton, K. L., Fadool, J. M., Spady, T. C.,
Mol. Phylogenet. Evol., 62, 986!1008.
and Travis, J. (2005) Genetic and environmental variation
15. Flamarique, I. N., Cheng, C. L., Bergstrom, C., and
in the visual properties of bluefin killifish, Lucania goodei,
Reimchen, T. E. (2013) Pronounced heritable variation
J. Evol. Biol., 18, 516!523.
and limited phenotypic plasticity in visual pigments and
8.
Rennison, D. J., Owens, G. L., Heckman, N., Schluter,
opsin expression of threespine stickleback photoreceptors,
D., and Veen, T. (2016) Rapid adaptive evolution of colour
J. Exp. Biol., 216, 656!667.
vision in the threespine stickleback radiation, Proc. Biol.
16. Cortesi, F., Musilová, Z., Stieb, S. M., Hart, N. S.,
Sci., 283, 20160242.
Siebeck, U. E., et al. (2015) Ancestral duplications and
БИОХИМИЯ том 87 вып. 2 2022
288
ПАВЛОВА и др.
highly dynamic opsin gene evolution in percomorph fishes,
the three!spined stickleback Gasterosteus aculeatus, BMC
Proc. Natl. Acad. Sci. USA, 112, 1493!1498.
Mol. Biol., 9, 18.
17.
Pavlova, N. S., Neretina, T. V., and Smirnova, O. V. (2020)
26. Dalton, B. E., Lu, J., Leips, J., Cronin, T. W., and
Dynamics of prolactin axis genes in the brain of male and
Carleton, K. L. (2015) Variable light environments induce
female three!spined stickleback Gasterosteus aculeatus
plastic spectral tuning by regional opsin coexpression in the
(Gasterostaidae) during short!term freshwater adaptation,
African cichlid fish, Metriaclima zebra, Mol. Ecol., 24,
J. Ichthyol., 60, 299!304.
4193!4204.
18.
Clapp, C., Thebault, S., Macotela, Y., Moreno!
27. Manzon L. A. (2002) The role of prolactin in fish osmo!
Carranza, B., Triebel, J., et al. (2015) Regulation of blood
regulation: A review, Gen. Comp. Endocrinol., 125, 291!
vessels by prolactin and vasoinhibins, Adv. Exp. Med. Biol.,
310.
846, 83!95.
28. Lee, K. M., Kaneko, T., Katoh, F., and Aida, K. (2006)
19.
Thébault, S. (2017) Potential mechanisms behind the
Prolactin gene expression and gill chloride cell activity in
antioxidant actions of prolactin in the retina, Exp. Eye
fugu Takifugu rubripes exposed to a hypoosmotic environ!
Res., 160, 56!61.
ment, Gen. Comp. Endocrinol., 149, 285!293.
20.
Huang, X., Hui, M. N., Liu, Y., Yuen, D. S., Zhang, Y.,
29. Sakamoto, T., and McCormick, S. D. (2006) Prolactin and
et al. (2009) Discovery of a novel prolactin in non!mam!
growth hormone in fish osmoregulation, Gen. Comp.
malian vertebrates: evolutionary perspectives and its involve!
Endocrinol., 147, 24!30.
ment in teleost retina development, PLoS One, 4, e6163.
30. Shu, Y., Lou, Q., Dai, Z., Dai, X., He, J., et al. (2016)
21.
Inokuchi, M., Breves, J. P., Moriyama, S., Watanabe, S.,
The basal function of teleost prolactin as a key regulator on
Kaneko, T., et al. (2015) Prolactin 177, prolactin 188, and
ion uptake identified with zebrafish knockout models, Sci.
extracellular osmolality independently regulate the gene
Rep., 6, 18597.
expression of ion transport effectors in gill of Mozambique
31. Watanabe, S., Itoh, K., and Kaneko, T. (2016) Prolactin
tilapia, Am. J. Physiol. Regul. Integr. Comp. Physiol., 309,
and cortisol mediate the maintenance of hyperosmoregula!
R1251!R1263.
tory ionocytes in gills of Mozambique tilapia: exploring
22.
Bollinger, R. J., Ellis, L. V., Bossus, M. C., and Tipsmark,
with an improved gill incubation system, Gen. Comp.
C. K. (2018) Prolactin controls Na+,Cl- cotransporter via
Endocrinol., 232, 151!159.
Stat5 pathway in the teleost gill, Mol. Cell. Endocrinol.,
32. Mohammed!Geba, K., González, A. A., Suárez, R. A.,
477, 163!171.
Galal!Khallaf, A., Martos!Sitcha, J. A., et al.
(2017)
23.
Franco!Belussi, L., De Oliveira, C., and Sköld, H. N.
Molecular performance of Prl and Gh/Igf1 axis in the Medi!
(2018) Regulation of eye and jaw colouration in three!
terranean meager, Argyrosomus regius, acclimated to dif!
spined stickleback Gasterosteus aculeatus, J. Fish Biol., 92,
ferent rearing salinities, Fish Physiol. Biochem., 43, 203!
1788!1804.
216.
24.
Pierce, A. L., Fox, B. K., Davis, L. K., Visitacion, N.,
33. Dauder, S., Young, G., and Bern, H. A. (1990) Effect of
Kitahashi, T., et al. (2007) Prolactin receptor, growth hor!
hypophysectomy, replacement therapy with ovine pro!
mone receptor, and putative somatolactin receptor in
lactin, and cortisol and triiodothyronine treatment on pro!
Mozambique tilapia: tissue specific expression and differ!
lactin receptors of the tilapia (Oreochromis mossambicus),
ential regulation by salinity and fasting, Gen. Comp.
Gen. Comp. Endocrinol., 77, 378!385.
Endocrinol., 154, 31!40.
34. Sulaiman, R. S., Kadmiel, M., and Cidlowski, J. A. (2018)
25.
Hibbeler, S., Scharsack, J. P., and Becker, S.
(2008)
Glucocorticoid receptor signaling in the eye, Steroids, 133,
Housekeeping genes for quantitative expression studies in
60!66.
OPSINE GENES EXPRESSION IN EYE RETINA OF FEMALE
AND MALE THREESPINED STICKLEBACKS Gasterosteus aculeatus L.
DEPENDS ON FRESHWATER ADAPTATION AND PROLACTIN
N. S. Pavlova1*, A. R. Gizatulina2, T. V. Neretina3, and O. V. Smirnova1
1 Department of Human and Animal Physiology, Faculty of Biology, Lomonosov Moscow State University,
119991 Moscow, Russia; e4mail: pav.nad.ser@gmail.com
2 Department of Physiology and General Pathology, Faculty of Fundamental Medicine,
Lomonosov Moscow State University, 119991 Moscow, Russia
3 Pertsov White Sea Biological Station, Moscow State University, 186671 poselok Seaside,
Loukhsky District, Republic Karelia, Russia
Color vision sensitivity is crucial for fish adaptation during migration and reproduction. Prolactin and prolactinlike
hormone are important hormonal regulators in both these processes. We hypothesized that prolactin might influence
color vision sensitivity during freshwater migrations in fish. We studied the effects of prolactin and freshwater adapta!
tion during spawning period on opsin gene expression (SWS1, SWS2, RH2, LWS) in the retina of female and male
threespined sticklebacks Gasterosteus aculeatus L. Prolactin gene expression elevates in the brain of female, but not
male, and prolactinlike hormone gene expression decreases in the brain of both male and female sticklebacks during
freshwater adaptation. Opsin SWS2 gene expression decreases in female and male retina during freshwater adaptation
and after prolactin administration. In the retina of male sticklebacks opsin SWS1 gene expression decreases after pro!
lactin administration but not freshwater adaptation. Opsins RH2 and LWS gene expression did not depend on pro!
lactin administration in male and female sticklebacks. We conclude that some opsin genes retinal expression is regu!
lated by prolactin in sticklebacks and could depend on sex and freshwater adaptation. This expands the knowledge of
adaptive effects of prolactin on fish during freshwater migrations.
Keywords: prolactin, opsins, threespined stickleback Gasterosteus aculeatus L., freshwater adaptation, color vision
adaptation, sex dependant effects
БИОХИМИЯ том 87 вып. 2 2022