БИОХИМИЯ, 2022, том 87, вып. 3, с. 307 - 320
УДК 577.112
ГИБРИДНЫЕ БЕЛКИ С КОРОТКИМИ КОНФОРМАЦИОННЫМИ
ЭПИТОПАМИ РЕЦЕПТОР СВЯЗЫВАЮЩЕГО ДОМЕНА
Spike БЕЛКА ВИРУСА SARS CoV 2 СПОСОБСТВУЮТ ВЫРАБОТКЕ
ВИРУСНЕЙТРАЛИЗУЮЩИХ АНТИТЕЛ ПРИ ИММУНИЗАЦИИ
© 2022
А.С. Карягина1,2,3*, А.В. Громов1, Т.М. Грунина1,2, А.М. Лящук1,
М.С. Попонова1, Д.А. Клейменов1, Н.В. Струкова1, М.С. Генералова1, А.В. Рязанова1,
З.М. Галушкина1, О.Ю. Добрынина1, Т.Н. Большакова1, М.В. Сергеева4,
Е.А. Романовская Романько4, И.В. Красильников5, М.Е. Субботина1,2*, В.Г. Лунин1,2
1 Национальный исследовательский центр эпидемиологии и микробиологии им. Н.Ф. Гамалеи Минздрава России,
123098 Москва, Россия; электронная почта: akaryagina@gmail.com, subbotinam@gmail.com
2 Всероссийский научно*исследовательский институт сельскохозяйственной биотехнологии, 127550 Москва, Россия
3 НИИ физико*химической биологии им. А.Н. Белозерского,
Московский государственный университет имени М.В. Ломоносова, 119991 Москва, Россия
4 НИИ гриппа им. А.А. Смородинцева Минздрава России, 197376 Санкт*Петербург, Россия
5 Санкт*Петербургский институт вакцин и сывороток ФМБА России, 198320 Санкт*Петербург, Россия
Поступила в редакцию 31.01.2022
После доработки 16.02.2022
Принята к публикации 16.02.2022
На основе предложенного ранее подхода синтезом в клетках Escherichia coli получены гибридные рекомби
нантные белки - потенциальные компоненты эпитопных вакцин, содержащие короткие конформацион
ные эпитопы рецептор связывающего домена Spike белка вируса SARS CoV 2 (144-153, 337-346, 414-425,
496-507 а.о.), реализующие большое количество белок белковых взаимодействий в комплексах с нейтрали
зующими антителами и ACE2 (ангиотензин превращающий фермент 2). Полученными белками трёхкрат
но с интервалом в 2 недели иммунизировали мышей и исследовали иммуногенность белковых антигенов и
способность сывороток взаимодействовать с инактивированным вирусом SARS CoV 2 и с рецептор связы
вающим доменом (RBD) Spike белка, полученным в эукариотическом продуценте. Все рекомбинантные
белки демонстрировали высокую иммуногенность. Наиболее высокий титр при связывании с RBD Spike
белка продемонстрировала сыворотка, полученная при иммунизации белком на основе эпитопа 414-425.
При этом титры сывороток при связывании с инактивированным вирусом и RBD в случае остальных бел
ков оказались существенно ниже титров для полученных ранее четырёх белков на основе петлеобразных
эпитопов 452-494 и 470-491, конформация которых дополнительно фиксировалась дисульфидной связью.
Для всех белков - полученных ранее и в данной работе - проведено исследование активации клеточного
иммунитета, детектируемой по уровню синтеза цитокинов спленоцитами иммунизированных мышей. Мак
симально выраженный уровень повышения синтеза цитокинов наблюдался в случае ответа на белки, вклю
чающие эпитопы с дисульфидными связями (452-494, 470-491), а также эпитопы 414-425 и 496-507. Пос
ле оптимизации адъюванта для некоторых рекомбинантных белков с короткими конформационными эпи
топами RBD Spike белка были получены сыворотки мышей, демонстрирующие вируснейтрализующий эф
фект в реакции микронейтрализации с живым вирусом SARS CoV 2 (hCoV 19/Russia/StPetersburg 3524/
2020 EPI_ISL_415710 GISAID). Полученные результаты могут быть использованы для разработки эпитоп
ных вакцин для профилактики COVID 19 и других вирусных инфекций.
КЛЮЧЕВЫЕ СЛОВА: SARS CoV 2, Spike белок, RBD, RBM, эпитопная вакцина, эпитоп, альдолаза.
DOI: 10.31857/S0320972522030010
ВВЕДЕНИЕ
инфекций является чрезвычайно актуальной за
дачей. Цельновирионные вакцины, субъединич
Разработка новых вариантов вакцин для
ные белковые вакцины, включающие полнораз
профилактики COVID 19 и других вирусных мерные белки, векторные вакцины и РНК вак
Принятые сокращения: ДТТ - дитиотреитол; СГТ - средний геометрический титр; ACE2 - ангиотензин превра
щающий фермент 2; ALD - альдолаза; COVID 19 - коронавирусная инфекция 2019 г. (COronaVIrus Disease 2019); HBD -
гепарин связывающий домен; RBD - рецептор связывающий домен; RBM - рецептор связывающий мотив; TLR - Toll
подобные рецепторы; Tri - α спираль, опосредующая тримеризацию Spike белка SARS CoV 2.
* Адресат для корреспонденции.
307
308
КАРЯГИНА и др.
цины, которые способствуют выработке в орга
для очистки. Для всех четырёх вариантов гиб
низме вирусных белков, индуцируют образова
ридных белков показана высокая иммуноген
ние широкого спектра антител, не все из кото
ность и способность антител, образующихся в
рых являются протективными [1]. В ряде случа
ответ на иммунизацию этими белками, в высо
ев могут образовываться антитела, способству
ком титре взаимодействовать как с рецептор
ющие антителозависимому усилению инфекции
связывающим доменом (RBD) Spike белка, так
при заражении после вакцинирования [2-4].
и с инактивированным SARS CoV 2.
Эпитопные вакцины, включающие отдельные
Целью данной работы было получение по
небольшие белковые эпитопы инфекционного
ранее разработанной схеме ещё нескольких ва
агента, позволяют избежать вышеперечислен
риантов гибридных белков, несущих конформа
ных проблем и при этом характеризуются высо
ционные эпитопы Spike белка коронавируса
кой степенью стандартности и слабой реакто
SARS CoV 2, исследование иммуногенности
генностью. Недостатками эпитопных вакцин
полученных белков и способности индуциро
может быть их низкая иммуногенность и неспо
ванных ими антител связываться с RBD Spike
собность вызывать образование вируснейтрали
белка и инактивированным SARS CoV 2, ха
зующих антител, а также возможность быстрого
рактеристика цитокинового ответа спленоцитов
ухода новых штаммов вируса от иммунного от
после иммунизации белками, несущими весь
вета, вызванного такими вакцинами. Повысить
спектр исследуемых эпитопов, исследование
иммуногенность и способствовать выработке
вируснейтрализации наиболее перспективными
вируснейтрализующих антител можно, исполь
гибридными белками на модели микронейтра
зуя такие подходы, как мультимеризация эпито
лизации клинического изолята коронавируса
пов [5] и оптимизация адъюванта [6]. Перспек
SARS CoV 2.
тивным подходом при выборе эпитопов - участ
ков поверхности белков, способных эффектив
но индуцировать образование антител - пред
МАТЕРИАЛЫ И МЕТОДЫ
ставляется выбор не линейных, а непрерывных
конформационных эпитопов, обеспечивающих
Штамм и векторы. В работе использовали
большее соответствие участку структуры вирус
бактериальный штамм Escherichia coli BL21
ного белка. Основываясь на данных о контак
(DE3) (E. coli B F- dcm ompT hsdS(r–m–) gal
тах, реализуемых в комплексах Spike белка с
λ(DE3), «Agilent Technologies», США), модифи
нейтрализующими антителами и ACE 2 (ангио
цированный плазмидный вектор pQE6
тензин превращающий фермент 2), на преды
(«Qiagen», США), содержащий вместо промото
дущих этапах исследования нами были выбраны
ра T5 промотор T7, и плазмиду pRep4 из E. coli
две детерминанты в области рецептор связыва
M15 [pRep4] («Qiagen»).
ющего мотива (RBM) - 452-494 и 470-491 а.о.,
Получение генно инженерных конструкций,
реализующие большое количество контактов
кодирующих белки Rop D2 Rop Tri HBD, Rop
как с антителами, так и с ACE 2 [7]. Обе детер
D3 Rop Tri HBD, Rop D2 Rop ALD HBD и Rop
минанты характеризуются петлеобразной кон
D3 Rop ALD HBD детально описано в работе
формацией и включают два остатка цистеина,
Karyagina et al. [7]. Последовательности этих
образующих дисульфидную связь в структуре
белков содержат участки D2 (470-490 а.о.) и
Spike белка. Для иммунизации были получены
D3 (453-494 а.о.) Spike белка вируса SARS
два варианта гибридных белков, содержащих
CoV 2 (UniProtKB: locus SPIKE_SARS2, acces
антигенные детерминанты, включённые в место
sion P0DTC2), вставленные между аминокис
поворота в мотиве «спираль поворот спираль»
лотными последовательностями двух α спира
Rop подобного белка из Methylococcus capsulatus
лей Rop подобного белка из M. capsulatus (пер
с целью сближения N и C концов и сохранения
вая спираль Rop: 2-34 а.о., вторая спираль Rop:
петлеобразной конформации эпитопов. Rop
34-65 а.о., PDB: 2JS5_A, RefSeq WP_010959602).
подобный белок в данном случае выполняет
Далее присоединена последовательность α спи
функцию так называемого эпитопного каркаса.
рали, опосредующей тримеризацию Spike белка
Показано, что при синтезе такого рода белков в
SARS CoV 2 (Tri) (958-991 а.о., UniProtKB:
Escherichia coli и последующей очистке дисуль
locus SPIKE_SARS2, accession P0DTC2), или
фидная связь образуется [7]. Гибридные белки
последовательность альдолазы из T. mariti*
содержат также либо альдолазу из Thermotoga
ma (2-201 а.о., PDB: 1WA3). На С конце белков
maritima, либо α спиральный участок, соответ
находится короткий участок гепарин связываю
ствующий 958-991 а.о. Spike белка SARS CoV 2,
щего гемагглютинина HBHA из Мycobacterium
обеспечивающие тримеризацию, а также гепа
tuberculosis, соответствующий гепарин связыва
рин связывающий участок, использующийся
ющему домену (HBD) (160-174 а.о., UniProtKB/
БИОХИМИЯ том 87 вып. 3 2022
КОНФОРМАЦИОННЫЕ ЭПИТОПЫ RBD Spike БЕЛКА SARS CoV 2
309
Swiss Prot: A1KFU9.1). В составе конструкций
центрифугированием в течение 30 мин при
также имеются дополнительные остатки, пред
9000 g. Затем проводили аффинную хроматогра
ставляющие собой линкерные последователь
фию на Heparin Sepharose CL 6B
(«GE
ности, и остатки, соответствующие нуклеотид
Healthcare», США), уравновешенной раствором
ным последовательностям с введёнными рест
4 М мочевины в 20 мМ Tris HCl буфере (рН 8,0),
рикционными сайтами.
с элюцией линейным градиентом NaCl с концен
Получение генно инженерных конструкций
трацией 0-1 М в растворе 4 М мочевины в
Rop D4 Rop Tri HBD, Rop D4 Rop ALD HBD,
25 мМ Tris HCl буфере (рН 8,0) и последующим
Rop D6 Rop Tri HBD, Rop D6 Rop ALD HBD,
диализом против буфера
25
мМ Tris НСl
Rop D8 Rop Tri HBD, Rop D8 Rop ALD HBD,
(рН 6,8), постепенно снижая концентрацию мо
Rop D13 Rop Tri HBD, Rop D13 Rop ALD HBD,
чевины в диализном буфере до нуля (ступенями
Rop RBM Rop Tri HBD проводили по схемам,
по 0,5 М) в течение 24 ч при 4 °С.
аналогичным описанным в работе Karyagina
Хроматографию растворимых белков прово
et al. [7]. Получали синтетические последова
дили на сорбенте WorkBeads 40S, уравновешен
тельности, кодирующие участки Spike белка ко
ном 20 мМ Tris НCl (рН 8,0), с элюцией раство
ронавируса SARS CoV 2: D4 (496-507 а.о.),
ром NaCl (линейным градиентом концентрации
D6
(144-153
а.о.), D8
(337-346
а.о.),
0-1 М) в том же буферном растворе, диализова
D13 (414-425 а.о.), RBM (433-511 а.о.), опти
ли против 20 мМ Tris НCl (pH 8,0) в течение 24 ч
мизированные с помощью программы JCat
и центрифугировали в течение 30 мин при
9000 g. Затем проводили аффинную хроматогра
скорректированные для улучшения вторичной
фию на гепарин сефарозе, уравновешенной
структуры транскрибируемой РНК с помощью
20 мМ Tris HCl буфером (рН 8,0), с элюцией
градиентом концентрации NaCl
0-1
М в
edu/?q=DINAMelt/Two state folding) с целью
20 мМ Tris HCl (рН 8,0). Полученные фракции
повышения экспрессии в E. coli. Синтетические
белков диализовали против 20 мМ Tris HCl
последовательности клонировали в плазмиды
(рН 8,0), 50 мМ NaCl в течение 24 ч при темпе
pL1003, кодирующую белок Rop D2 Rop Tri
ратуре 4 °C с трёхкратной сменой буфера.
HBD и pL989, кодирующую белок Rop D2 Rop
Концентрацию и общее количество белков
ALD HBD, получение которых описано ра
определяли по поглощению раствора при 280 нм
нее [7], замещая детерминанту D2. Cинтетичес
с использованием бицинхониновой кислоты.
кие последовательности получали в компании
Полученные препараты белков лиофильно вы
«Евроген» (Россия).
сушивали и использовали для дальнейших ис
Выращивание штаммов продуцентов и разру
следований.
шение клеток E. coli проводили по протоколам,
Молекулярную массу и теоретические значе
описанным в работе Karyagina et al. [7]. При
ния некоторых параметров рекомбинантных
этом белки Rop D4 Rop Tri HBD, Rop D8
белков рассчитывали с использованием интер
Rop Tri HBD, Rop D13 Rop Tri HBD и Rop
RBM Rop Tri HBD синтезировались в нерас
Иммуногенные композиции и эксперименты по
творимой форме в виде телец включений, а бел
иммунизации. На основе полученных рекомби
ки Rop D13 Rop ALD HBD, Rop D4 Rop
нантных белков, содержащих конформацион
ALD HBD и Rop D8 Rop ALD HBD - в рас
ные эпитопы Spike белка SARS CoV 2 (включая
творимой форме. Синтеза белка Rop D6 Rop
четыре белка, полученных ранее), были подго
ALD HBD не наблюдалось.
товлены иммуногенные композиции, содержа
Очистку белков проводили в соответствии с
щие в одной дозе (объёмом 150 мкл) 1 нмоль ре
ранее опубликованными протоколами хрома
комбинантного белка и соответствующий адъю
тографической очистки - для нерастворимых и
вант. В первом эксперименте по иммунизации
для растворимых белков [7]. Хроматографию
мышей использовали семь рекомбинантных
белков, синтезировавшихся в тельцах включе
белков (белки в строках 5-11 в табл. 1), в имму
ний, растворенных в 8 М мочевине и 10 мМ Tris
ногенные композиции вводили адъювант 1: ди
НCl (рН
8,0), проводили на колонке с
этиламиноэтиловый (ДЭАЭ) декстран 500 -
WorkBeads 40S («Bio Works», Швеция), уравно
1 мг, монтанид ISA 201 («Seppic SA», Франция) -
вешенной раствором
8
М мочевины и
75
мкл
(50%),
ретинола пальмитат
10 мМ Tris НCl (рН 8,0), с элюцией линейным
(100 000 МЕ/мл) (АО «Ретиноиды», Россия) -
градиентом NaCl с концентрацией 0-1 М в том
1,5 мкл (1%). Сыворотки, полученные после
же буферном растворе и последующим диали
этой иммунизации, использовали для оценки
зом против раствора 4 М мочевины в 25 мМ Tris
иммуногенности рекомбинантных белков, спо
НCl буфере (рH 8,0) в течение 24 ч при 4 °С и
собности взаимодействовать с RBD и инактиви
БИОХИМИЯ том 87 вып. 3 2022
310
КАРЯГИНА и др.
рованным вирусом SARS CoV 2, постановки
ных опытных и контрольной групп вводили в
первой реакции микронейтрализации, сплено
наркоз ингаляцией изофлурана
(«Piramal
циты - для исследования клеточного иммуни
Enterprises», Индия) и проводили сбор крови из
тета. Во втором эксперименте по иммунизации
сердечной полости (в случае первого экспери
использовали те же рекомбинантные белки, ра
мента по иммунизации также проводили извле
нее полученные рекомбинантные белки [7]
чение селезёнки, см. подраздел про оценку ак
(строки 1-4 в табл. 1), а также белки Rop RBM
тивации клеточного иммунитета). Вакуумные
Rop Tri HBD и RBD («Mount Sinai», США), в
пробирки с кровью выдерживали при комнат
иммуногенные композиции вводили адъю
ной температуре 20 мин и центрифугировали
вант 2: ДЭАЭ декстран 500 - 1 мг, липополиса
30 мин при 1500 g без охлаждения. Надосадоч
харид (LPS) из E. сoli O55:B5 («Sigma», США) -
ную жидкость (сыворотку крови) переносили в
0,1 мкг, олигодезоксинуклеотиды CpG (ODN 1826)
стерильные пробирки объёмом 1,5 мл и замора
(ЗАО «Евроген», Россия) - 30 мкг. Полученные
живали при -80 °С.
сыворотки использовали для постановки второй
Иммуноферментный анализ. В качестве анти
реакции микронейтрализации.
генов на планшеты сорбировали белки, исполь
Детальный протокол иммунизации лабора
зованные для иммунизации, вирус SARS CoV 2,
торных животных описан ранее [7]. Для имму
инактивированный формалином, и RBD Spike
низации использовали самок мышей сингенной
белка вируса SARS CoV 2. Получение инакти
инбредной линии BALB/c в возрасте 5-6 недель
вированного SARS CoV 2 и RBD Spike белка
весом 18-20 г (НИЦЭМ им. Н.Ф. Гамалеи
вируса SARS CoV 2, а также детальный прото
Минздрава России). Мышам опытных групп
кол ИФА описаны ранее [7]. Кратко, в 96 луноч
(n = 7) трёхкратно вводили по 150 мкл иммуно
ных планшетах Maxisorp («Thermo Scientific»,
генных композиций, содержащих рекомбинант
Дания) в течение ночи при 4 °С проводили сорб
ные белки, с интервалом в 2 недели. Животным
цию антигенов, после отмывки планшеты инку
контрольной группы (n = 7) вводили физиоло
бировали в течение 2 ч с блокирующим буфером
гический раствор (150 мкл). Через 2 недели пос
(PBS, содержащий 5% сухого молока; «ТФ Ди
ле последней инъекции лабораторных живот
тол», Россия), повторно отмывали и вносили се
Таблица 1. Характеристика полученных рекомбинантных белков
Поглощение
водного
Поглощение
раствора
водного раствора
Молеку
Локализация белка
Теоретическая
белка
белка
№
Белок
лярная
после разрушения
изоэлектрическая
(1 мг/мл) при
(1 мг/мл)
масса, Да
биомассы
точка (pI)
280 нм для
при 280 нм для
полностью
восстановленной
окисленной
формы
формы
1
Rop D2 Rop Tri HBD
18 714
тельца включения
8,83
0,325
0,318
2
Rop D3 Rop Tri HBD
21 359
тельца включения
9,39
0,355
0,349
3
Rop D2 Rop ALD HBD
36 102
растворимая фракция
8,58
0,486
0,483
4
Rop D3 Rop ALD HBD
38 747
растворимая фракция
9,10
0,491
0,488
5
Rop D4 Rop Tri HBD
17 491
тельца включения
9,45
0,256
6
Rop D8 Rop Tri HBD
17 627
тельца включения
9,49
0,169
7
Rop D4 Rop ALD HBD
34 878
растворимая фракция
9,06
0,457
8
Rop D8 Rop ALD HBD
35 014
растворимая фракция
9,07
0,412
9
Rop D6 Rop Tri HBD
34 966
тельца включения
9,05
0,456
10
Rop D13 Rop ALD HBD
35 215
растворимая фракция
9,13
0,495
11
Rop D13 Rop Tri HBD
17 827
тельца включения
9,29
0,334
12
Rop RBM Rop Tri HBD
245 628
тельца включения
8,43
0,836
0,831
Примечание. Для белков, не образующих дисульфидную связь, поглощение водного раствора белка при 280 нм для вос
становленной формы не исследовали. Данные для белков в строках 1-4 получены ранее [7] и приведены для удобства
сравнения.
БИОХИМИЯ том 87 вып. 3 2022
КОНФОРМАЦИОННЫЕ ЭПИТОПЫ RBD Spike БЕЛКА SARS CoV 2
311
рии десятикратных разведений сывороток мы
pH 7,0-7,6) и инкубировали 40 с в темноте во
шей. В качестве вторичных антител использова
льду, после чего разбавляли ледяной сре
ли меченые пероксидазой хрена Goat anti mouse
дой DMEM и центрифугировали при 200 g
IgG (H+L) HRP («Invitrogen», США) в разведе
15 мин. Супернатант полностью удаляли, а бе
нии 1 : 1200 (вносили по 100 мкл в лунку). В ка
лый клеточный осадок ресуспендировали в пол
честве цветного субстрата использовали рас
ной ростовой культуральной среде RPMI 1640,
твор TMB («Биосервис», Россия), реакцию оста
включающей 10% FBS («HyClone»), 2 мМ L глу
навливали добавлением 1 N H2SO4. Поглощение
тамина
(«ПанЭко»), антибиотик антимико
при длине волны 450 нм измеряли на мульти
тик
(«Sigma»),
0,05
мМ 2 меркаптоэтано
планшетном ридере с LVF монохроматорами
ла («Sigma»), среду RPMI 1640 без глутами
CLARIOstar («BMG Labtech», США).
на («ПанЭко»). Затем проводили подсчёт клеток
Оценку активации клеточного иммунитета
в суспензии с помощью камеры Горяева. Кле
проводили по уровню синтеза цитокинов спле
точную суспензию разводили до концентрации
ноцитами иммунизированных мышей методом
5 × 106-107 кл./мл и вносили по 100 мкл в каж
мультиплексного иммунологического анализа.
дую лунку планшета. Клетки инкубировали в
При этом использовали панель MILLIPLEX
течение 1-1,5 ч в СО2 инкубаторе, после чего
MAP Mouse Cytokine/Chemokine Magnetic Bead
вносили разведения антигенов и контролей.
Panel («Merck Millipore», США), основанную на
Планшет инкубировали в течение 18-24 ч в
измерении уровня 32 регуляторных, про и про
СО2 инкубаторе, после чего переносили культу
тивовоспалительных цитокинов, хемокинов и
ральную среду в микропробирки, замораживали
факторов роста (G CSF, GM CSF, IFN γ, IL 1α,
при -80 °С и хранили до проведения анализа.
IL 1β, IL 2, IL 3, IL 4, IL 5, IL 6, IL 7, IL 9, IL
Анализ спленоцитов, полученных после пер
10, IL 12 (p40), IL 12 (p70), IL 13, IL 15, IL 17,
вого эксперимента по иммунизации (бел
IP 10, KC, LIF, LIX, MCP 1, M CSF, MIG, MIP
ки 5-11 в табл. 1 с адъювантом 1), проводили
1α, MIP 1β, MIP 2, RANTES, TNF α, VEGF,
одновременно с анализом спленоцитов, выде
эотаксин/CCL11). Все анализы проводили в со
ленных ранее после иммунизации белками 1-4
ответствии с протоколом производителя, ис
из табл. 1 с адъювантом 1 [7] (планшеты храни
пользуя рекомендуемые разведения образцов и
ли при -80 °С до проведения анализа).
концентрации стандартной кривой. Измерения
Реакция микронейтрализации. Для обнаруже
проводили на приборе MAGPIX с программным
ния нейтрализующих антител использовали ре
обеспечением xPONENT версии 4.2 («Luminex
акцию микронейтрализации на основе
Corp.», США).
TCID50 (Median Tissue Culture Infectious Dose)
Спленоциты (макрофаги, дендритные клет
[8]. Сыворотки мышей, полученные после трёх
ки, Т и В лимфоциты) выделяли из селезёнки
кратной иммунизации с двухнедельными ин
мышей через 2 недели после первого экспери
тервалами, прогревали 60 мин при 56 °С, чтобы
мента по иммунизации. Эвтаназированных мы
избежать связанного с комплементом снижения
шей обмывали 96% ным спиртом, после чего в
вирусной активности. Затем готовили серию
асептических условиях проводили вскрытие
двукратных разведений сыворотки на среде
брюшной полости, извлекали селезёнки и поме
MEM с добавлением 2% HI FBS в объёме
щали их в стерильную холодную среду DMEM
60 мкл, начиная с 1 : 10 (в пересчёте на цельную
(«ПанЭко», Россия). Для выделения культуры
сыворотку). Каждое разведение сыворотки сме
спленоцитов использовали набор для разруше
шивали в равном объёме (60 мкл) с живым ви
ния тканей Tissue Grinder Homogenizer
русом SARS CoV 2 (hCoV 19/Russia/StPetersburg
Kit («Sigma», США). Стерилизацию инструмен
3524/2020 EPI_ISL_415710 GISAID), содержа
тов проводили путём промывания в 96% ном
щим 25 TCID50/50 мкл, и инкубировали 1 ч при
спирте и прожигания в пламени спиртовки.
37 °С. Затем 100 мкл разведений переносили в
Стерильно собирали инструмент для разруше
96 луночные планшеты с монослоем клеток
ния тканей, использовали сетки 100 mesh и
Vero (ATCC CCL 81). Планшеты инкубировали
200 mesh («Sigma»). Селезёнки нарезали на
при 37 °С в атмосфере 5% CO2 в течение 4 сут.,
фрагменты и протирали через сито в 9 см чашки
после чего проверяли на наличие ЦПД (цитопа
Петри с ледяной средой DMEM, после чего сус
тическое действие). Нейтрализацию регистри
пензию переносили в стерильные центрифуж
ровали, если 100% клеток в лунке сохранялись
ные пробирки объёмом 15 мл. Суспензию цент
без видимых бляшек или цитопатического эф
рифугировали при 200 g 15 мин, после чего су
фекта. Нейтрализующий титр сыворотки выра
пернатант удаляли. Клеточный осадок ресус
жали как значение, обратное самому высокому
пендировали в буфере для лизиса эритроцитов
разведению сыворотки, в котором была показа
(150 мМ NH4Cl, 10 мМ KHCO3, 1 мМ EDTA,
на нейтрализующая активность [9].
БИОХИМИЯ том 87 вып. 3 2022
312
КАРЯГИНА и др.
Эксперименты с использованием реакции
подобного белка введены гибкие глицин содер
микронейтрализации проводили дважды. В пер
жащие спейсеры. В качестве доменов, обеспе
вой реакции микронейтрализации анализирова
чивающих мультимеризацию (в данном слу
ли сыворотки, полученные в данной работе и
чае - тримеризацию) эпитопов, выбраны либо
ранее [7], после иммунизации белками Rop D2
α спираль, участвующая в тримеризации Spike
Rop Tri HBD, Rop D3 Rop Tri HBD, Rop D2
белка SARS CoV 2 (Tri), либо альдолаза из
Rop ALD HBD, Rop D3 Rop ALD HBD, Rop
T. maritima (ALD). Кроме того, в состав гибрид
D4 Rop Tri HBD, Rop D4 Rop ALD HBD и
ных белков включен фрагмент гемагглютини
Rop D13 Rop Tri HBD с адъювантом 1, во вто
на HBHA из М. tuberculosis, соответствующий ге
рой реакции - сыворотки, полученные после
парин связывающему домену (HBD).
иммунизации теми же белками, а также белками
Молекулярные массы мономерных белков
Rop RBM Rop Tri HBD и RBD («Mount Sinai»)
находятся в пределах 17-19 кДа для белков на
с адъювантом 2. Все эксперименты проводили в
основе тримеризатора из коронавируса и
лаборатории BSL3.
35-39 кДа - для белков на основе альдолазы,
Статистический анализ. Статистический ана
что с учётом тримеризации должно быть доста
лиз проводили с использованием программ па
точно для эффективной выработки антител.
кета Statistica 12.0 («Statsoft», США). Нормаль
В качестве одного из контрольных белков
ность распределения проверяли с помощью тес
для исследования вируснейтрализующей актив
та Колмогорова-Смирнова. Данные представ
ности сывороток на основе тех же белковых
ляли как среднее
± стандартное отклоне
компонентов был сконструирован гибридный
ние (SD) или как среднее геометричес
белок Rop RBM Rop Tri HBD, в котором в по
кое ± стандартное геометрическое отклонение
ворот между двумя спиралями Rop подобного
логарифма по основанию 2 титра сывороток ан
белка был вставлен фрагмент Spike белка SARS
тител. Статистическую значимость различий
CoV 2 с 433 по 511 а.о., включающий последова
оценивали с помощью двухстороннего диспер
тельности детерминант D2, D3 и D4.
сионного анализа (one way ANOVA) с апостери
Получение и очистка рекомбинантных бел
орным анализом по Тьюки. Различия считали
ков. Клонирование генов рекомбинантных бел
значимыми при p < 0,05.
ков проводили на основе ранее полученных
конструкций [7], как описано в разделе «Мате
риалы и методы». Все генно инженерные
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
конструкции обеспечивали высокий уровень
экспрессии генов, за исключением гена белка
Дизайн рекомбинантных белков. На основе
Rop D6 Rop ALD HBD, который по этой при
анализа структур Spike белка коронавируса
чине не использовали в работе. Белки на осно
SARS CoV 2 в комплексе с ACE 2 и нейтрали
ве тримеризационного домена из коронавируса
зующими антителами в дополнение к ранее
нарабатывались в виде телец включений, а бел
выбранным двум эпитопам Spike белка вируса
ки на основе альдолазы синтезировались в
SARS CoV 2
- D2
(470-490
а.о.) и D3
растворимой форме. Соответственно, схемы
(453-494 а.о.) [7] - было выбрано ещё четыре
выделения этих белков различались: белки с
поверхностных эпитопа: D4 (496-507 а.о.),
тримеризатором из SARS CoV 2 выделяли в де
D6
(144-153
а.о.), D8
(337-346
а.о.),
натурирующих условиях в присутствии мочеви
D13 (414-425 а.о.) (рис. 1). Все детерминанты,
ны, а белки с альдолазой - в неденатурирую
за исключением D8, имеющей α спиральную
щих условиях. При этом на первой стадии
конформацию, имеют петлеобразную конфор
очистки для всех белков использовали хрома
мацию со сближенными N и С концами. Ре
тографию на сорбенте WorkBeads 40S, на вто
комбинантные белки, несущие эпитопы D4, D6,
рой - на гепарин сефарозе, что обеспечивалось
D8 и D13, организованы аналогично ранее по
присутствием гепарин связывающего домена в
лученным белкам с эпитопами D2 и D3 [7]. Для
белках.
дополнительной фиксации N и C концов эпи
Молекулярные массы, вычисленные на ос
топов и обеспечения их конформационной ста
новании аминокислотных последовательнос
бильности аминокислотная последовательность
тей, и теоретические значения некоторых пара
всех эпитопов фланкирована взаимодействую
метров рекомбинантных белков представлены в
щими между собой α спиралями Rop подобно
табл. 1 (в табл. 1 включены также данные для ра
го белка из M. capsulatus. При этом петлеобраз
нее полученных белков с эпитопами D2 и D3 [7]
ные детерминанты вставлены в поворот между
для удобства сравнения).
спиралями непосредственно, а в случае эпито
Электрофоретическое разделение очищен
па D8 - между эпитопом и α спиралями Rop
ных белков показано на рис. 2. Различная
БИОХИМИЯ том 87 вып. 3 2022
КОНФОРМАЦИОННЫЕ ЭПИТОПЫ RBD Spike БЕЛКА SARS CoV 2
313
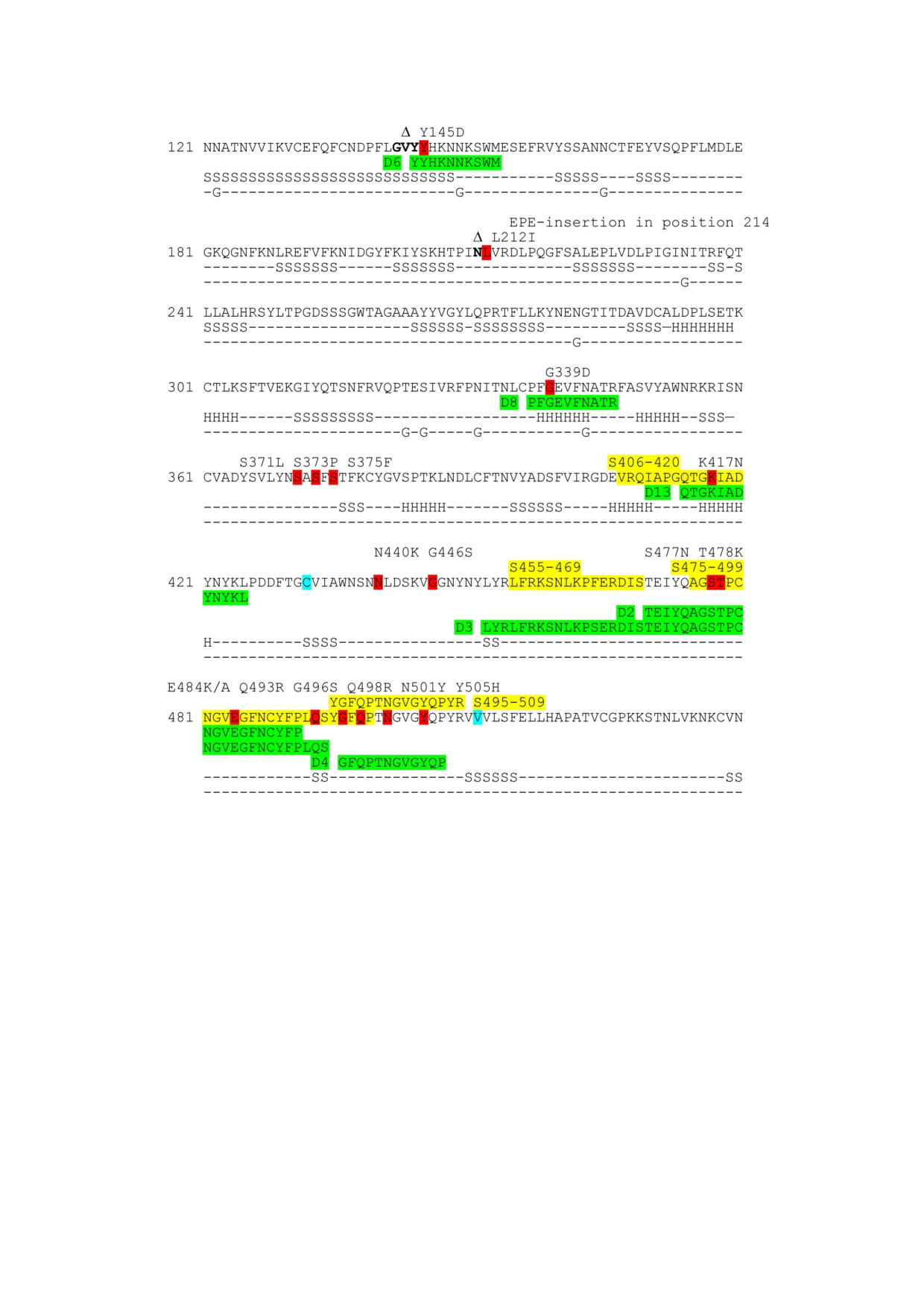
Рис. 1. Участок последовательности Spike белка вируса SARS CoV 2 и последовательности эпитопов, используемые в ра
боте (выделены зелёным цветом). Жёлтым цветом выделены а.о., входящие в линейные эпитопы S406-420, S455-469,
S475-499 и S495-509 (из статьи Lu et al. [5]), последовательности которых перекрываются с последовательностями иссле
дуемых конформационных эпитопов. Красным цветом выделены вариабельные а.о. в различных вариантах SARS CoV 2.
Полужирным шрифтом и значком Δ выделены а.о., делетированные в Spike белке варианта SARS CoV 2 Омик
рон (B.1.1.529). Бирюзовым цветом выделены начальный и концевой а.о. фрагмента, входящего в состав конструкции
Rop RBM Rop Tri HBD. Элементы вторичной структуры (α спиральные участки (H), β тяжи (S)) и сайты гликозилиро
вания (G) отмечены в строках ниже аминокислотной последовательности напротив соответствующих остатков в соответ
ствии с данными из комплекса Spike белка SARS CoV 2 с нейтрализующим антителом (цепь А, PDB: 7C2L)
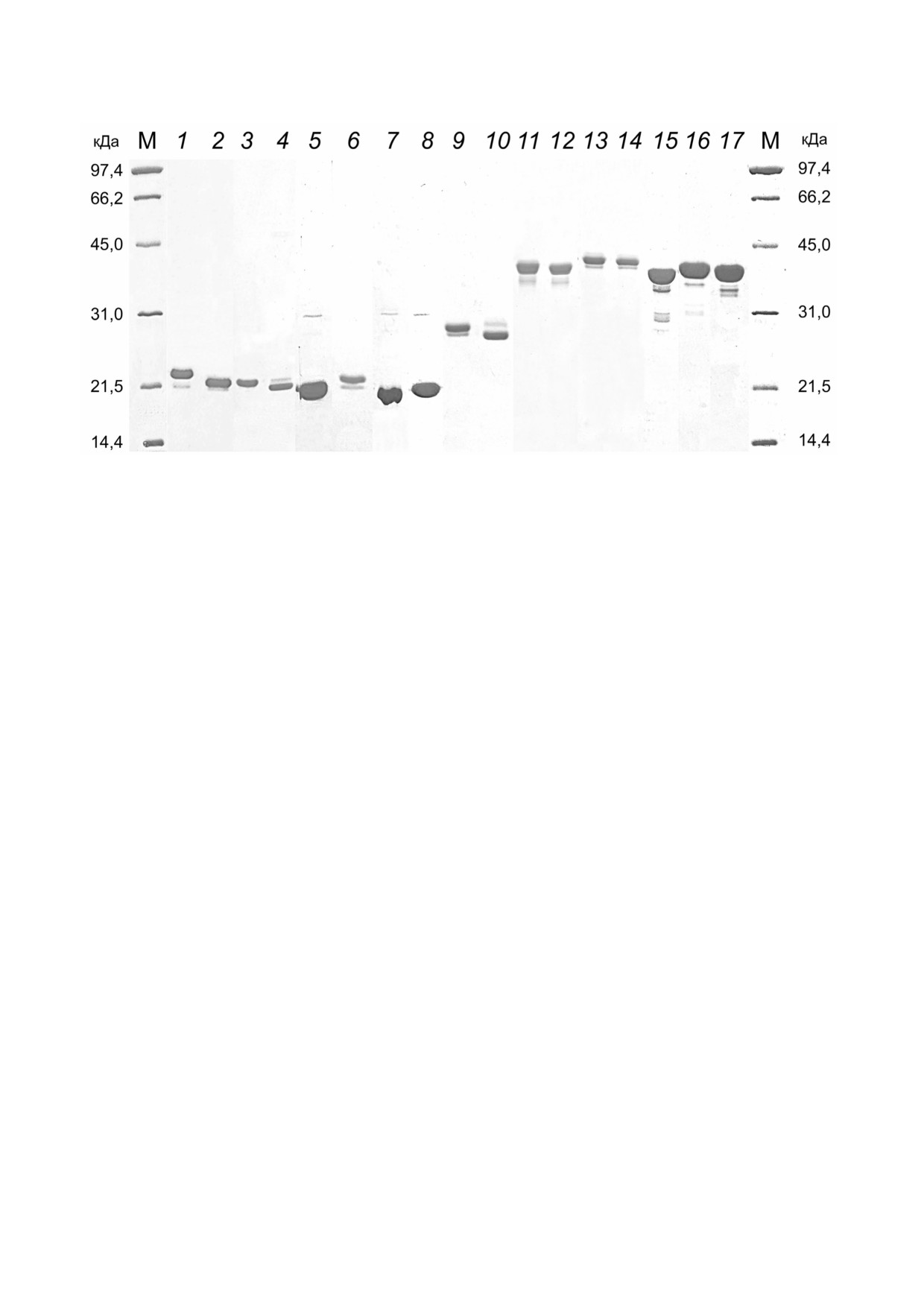
электрофоретическая подвижность белков с
спектрометрическим анализом триптических
эпитопами D2 и D3, содержащими в аминокис
пептидов [7].
лотных последовательностях по два цистеина, в
Иммуногенные композиции, иммунизация мы
присутствии и при отсутствии восстанавливаю
шей. На основе семи рекомбинантных белков с
щего агента дитиотреитола (ДТТ), свидетель
петлеобразными эпитопами (их характеристика
ствует о том, что очищенные белки формируют
приведена в строках 5-11 табл. 1) и адъюванта 1
дисульфидную связь. Наличие дисульфидной
были получены композиции для иммунизации
связи в белке, содержащем последовательность
мышей инбредной линии BALB/C. Наличие
с детерминантами D2/D3, показано ранее масс
специфических иммуноглобулинов класса G
БИОХИМИЯ том 87 вып. 3 2022
314
КАРЯГИНА и др.
Рис. 2. Электрофореграммы разделения препаратов рекомбинантных белков в 12% ном ПААГ. M - маркер молекулярной
массы 14,4-97,4 кДа («Bio Rad», США); 1 - белок Rop D2 Rop Tri HBD в присутствии ДТТ; 2 - белок Rop D2 Rop Tri
HBD без ДТТ; 3 - белок Rop D3 Rop Tri HBD в присутствии ДТТ; 4 - белок Rop D3 Rop Tri HBD без ДТТ; 5 - белок
Rop D4 Rop Tri HBD; 6 - белок Rop D8 Rop Tri HBD; 7 - белок Rop D6 Rop Tri HBD; 8 - белок Rop D13 Rop Tri
HBD; 9 - белок Rop RBM Rop Tri HBD в присутствии ДТТ; 10 - белок Rop RBM Rop Tri HBD без ДТТ; 11 - белок Rop
D2 Rop ALD HBD в присутствии ДТТ; 12 - белок Rop D2 Rop ALD HBD без ДТТ; 13 - белок Rop D3 Rop ALD HBD
в присутствии ДТТ; 14 - белок Rop D3 Rop ALD HBD без ДТТ; 15 - белок Rop D4 Rop ALD HBD; 16 - белок Rop D8
Rop ALD HBD; 17 - белок Rop D13 Rop ALD HBD
(IgG) в образцах сывороток мышей определяли
водили, измеряя уровень синтеза цитокинов
через 2 недели после трёхкратной иммунизации
спленоцитами иммунизированных мышей. При
с интервалом в 2 недели. В качестве антигенов
этом спленоциты (макрофаги, дендритные
использовали те же белки, которые применяли
клетки, Т и В лимфоциты), выделенные из се
для иммунизации, вирус SARS CoV 2, инакти
лезёнок мышей через 2 недели после трёхкрат
вированный формалином, и RBD Spike белка
ной (с двухнедельными перерывами) иммуниза
вируса SARS CoV 2. Результаты анализа, а так
ции с адъювантом 1, подвергали суточному воз
же ранее полученные данные для белков, содер
действию теми же антигенами, после чего изме
жащих эпитопы D2 и D3, представлены в
ряли степень их ответа на стимуляцию путём
табл. 2. Все гибридные белки вызывали у мышей
определения уровней цитокинов, хемокинов и
образование антител класса IgG, в высоком тит
факторов роста в среде, где проходил этот про
ре взаимодействующих с использованным для
цесс. Панель, используемая для оценки иммун
иммунизации антигеном. Белки, содержащие
ного ответа, основана на измерении уровня
детерминанты D2 и D3, как было показано ра
32 регуляторных, про и противовоспалитель
нее [7], с достаточно высоким титром взаимо
ных цитокинов, хемокинов и факторов роста.
действовали также и с инактивированным виру
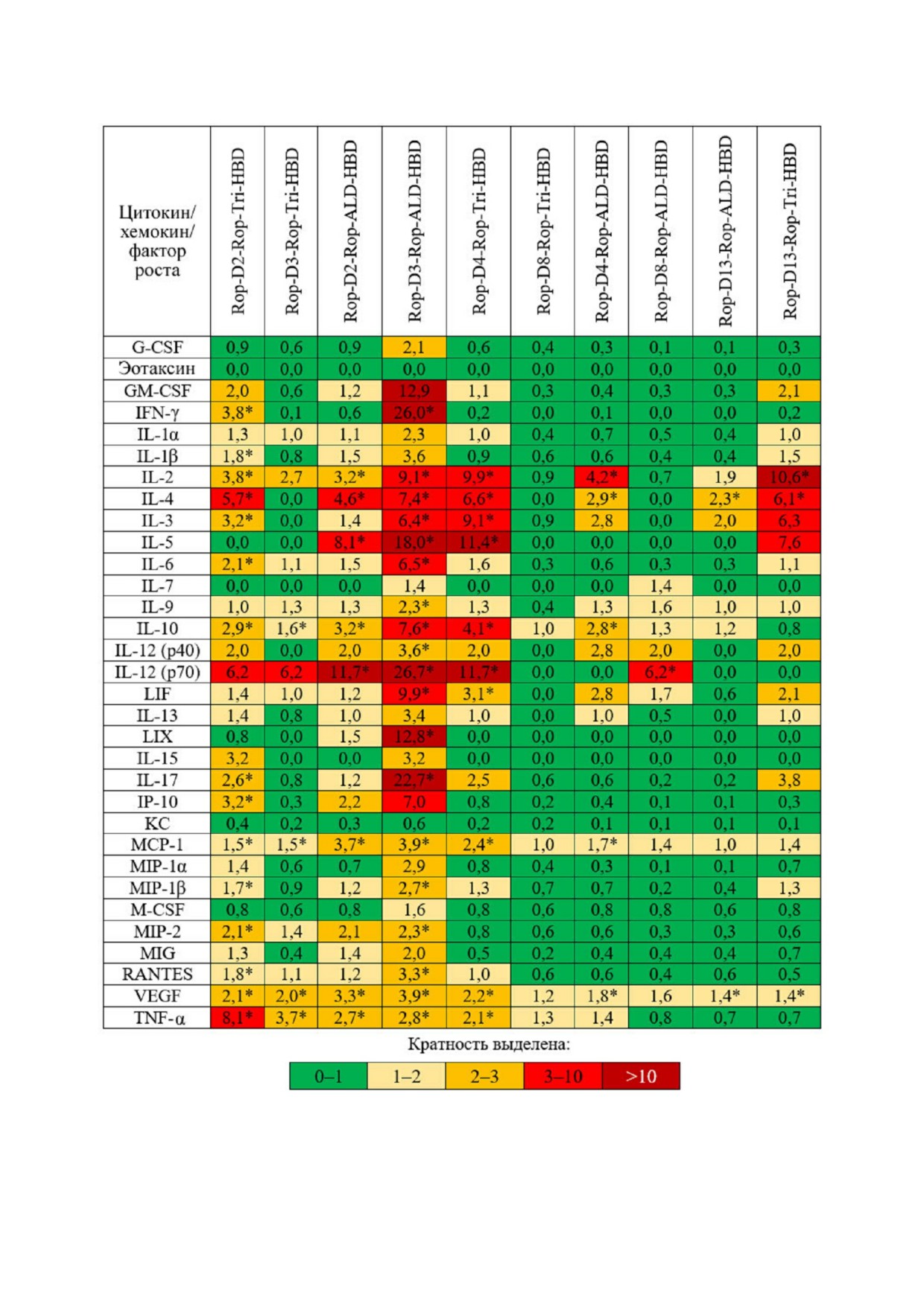
Результаты эксперимента представлены на
сом, и с RBD. Также с RBD взаимодействовал
рис. 3. Максимально выраженный уровень по
белок Rop D13 Rop Tri HBD. Наиболее имму
вышения цитокинов наблюдался в случае ответа
ногенными являются все белки с альдолазным
на белок Rop D3 Rop ALD HBD, несколько
доменом и белки Rop D13 Rop Tri HBD и Rop
менее выраженный - в случае белков Rop D2
D4 Rop Tri HBD. Наиболее выраженный ответ
Rop Tri HBD, Rop D3 Rop Tri HBD, Rop D2
к RBD наблюдался в случае белка Rop D3 Rop
Rop ALD HBD, Rop D4 Rop Tri HBD, Rop
ALD HBD. Наиболее выраженный ответ к
D4 Rop ALD HBD и Rop D13 Rop Tri HBD.
инактивированному вирусу SARS CoV 2 на
Максимальный ответ на белок Rop D3 Rop
блюдался в группе животных, иммунизирован
ALD HBD (рис. 3) совпадает с максимальным
ных белком Rop D2 Rop ALD HBD.
титром антител, специфичных к RBD Spike бел
Исследование клеточного иммунитета. Иссле
ка SARS CoV 2, характерным для данного белка
дование активации клеточного иммунитета про (табл. 2).
БИОХИМИЯ том 87 вып. 3 2022
КОНФОРМАЦИОННЫЕ ЭПИТОПЫ RBD Spike БЕЛКА SARS CoV 2
315
Таким образом, максимально выраженный
второго эксперимента по иммунизации (трёх
клеточный ответ вызывали гибридные белки
кратная иммунизация через 2 недели, но с ис
Rop D2 Rop Tri HBD, Rop D3 Rop Tri HBD,
пользованием адъюванта 2). Помимо отобран
Rop D2 Rop ALD HBD, Rop D3 Rop ALD
ных рекомбинантных белков, содержащих ис
HBD, Rop D4 Rop Tri HBD, Rop D4 Rop
следуемые эпитопы, для иммунизации также
ALD HBD и Rop D13 Rop Tri HBD. Далее ин
использовали ранее выделенные рекомбинант
тересно было выяснить, обладают ли антитела,
ные белки с эпитопами D2 и D3 [7], полученный
образующиеся у мышей после иммунизации
в данной работе белок Rop RBM Rop Tri HBD
этими белками, вируснейтрализующей актив
и коммерческий препарат RBD («Mount Sinai»).
ностью.
Данные по реакции микронейтрализации при
Реакция микронейтрализации. Для обнаруже
ведены в табл. 2. В результате модификации
ния нейтрализующих антител проводили реак
адъюванта в случае ряда разведений сывороток
цию микронейтрализации на основе TCID50 с
было показано отсутствие цитопатического
сыворотками иммунизированных мышей.
действия на клетки живого вируса SARS CoV 2
В первой реакции микронейтрализации ана
(hCoV 19/Russia/StPetersburg 3524/2020
лизировали сыворотки мышей, полученные
EPI_ISL_415710 GISAID), это свидетельствует о
после первого эксперимента по иммунизации
том, что в ответ на иммунизацию гибридными
(трёхкратная иммунизация с двухнедельными
белками, содержащими небольшие антигенные
интервалами с адъювантом 1), а также сыворот
детерминанты Spike белка SARS CoV 2, обра
ки, полученные ранее при иммунизации белка
зуются специфические антитела, способные
ми с эпитопами D2 и D3 и адъювантом 1 [7].
нейтрализовать клинический изолят вируса.
Ни одна из сывороток в условиях реакции мик
Максимальные значения среднего геометричес
ронейтрализации (см. «Материалы и методы»)
кого титра (СГТ) продемонстрировали сыворот
не оказалась способной нейтрализовать вирус
ки, полученные при иммунизации белками
SARS CoV 2.
Rop D13 Rop Tri HBD (6,7), Rop D2 Rop Tri
Во второй реакции микронейтрализации
HBD (7,4), Rop D4 Rop Tri HBD (8,2). Сыво
анализировали сыворотки, полученные после ротки, полученные при иммунизации контроль
Таблица 2. Титры сывороточных антител, определённых методом иммуноферментного анализа к различным антигенам в
сыворотках мышей после трёхкратной иммунизации, а также в экспериментах по вируснейтрализации
Группа
Антиген,
Инактивированный
СГТ в реакции
мышей
Белок
использованный
формалином
RBD
микронейтрализации
при иммунизации
SARS CoV 2
(разброс значений)
1
физиологический раствор
< 7,0 ± 1,00
< 7,0 ± 1,00
< 7,0 ± 1,00
5,0
2
Rop D2 Rop Tri HBD
14,21 ± 1,04
8,64 ± 1,00
10,73 ± 1,12
7,4 (5; 10)
3
Rop D3 Rop Tri HBD
13,48 ± 1,19
9,67 ± 1,18
11,44 ± 1,19
5,0
4
Rop D4 Rop Tri HBD
> 15,64 ± 1,00
< 7,0 ± 1,00
< 7,0 ± 1,00
8,2 (5; 20)
5
Rop D6 Rop Tri HBD
10,57 ± 1,40
< 7,0 ± 1,00
< 7,0 ± 1,00
-
6
Rop D8 Rop Tri HBD
8,64 ± 1,32
< 7,0 ± 1,00
< 7,0 ± 1,00
-
7
Rop D13 Rop Tri HBD
> 15,64 ± 1,00
< 7,0 ± 1,00
10,12 ± 1,16
6,7 (5; 10)
8
Rop D2 Rop ALD HBD
> 15,64 ± 1,00
9,88 ± 1,12
11,36 ± 1,17
5,5 (5; 10)
9
Rop D3 Rop ALD HBD
> 15,64 ± 1,00
9,75 ± 1,10
12,61 ± 1,08
5,5 (5; 10)
10
Rop D4 Rop ALD HBD
> 5,64 ± 1,00
< 7,0 ± 1,00
< 7,0 ± 1,00
5,5 (5; 10)
11
Rop D8 Rop ALD HBD
> 15,64 ± 1,00
< 7,0 ± 1,00
< 7,0 ± 1,00
-
12
Rop D13 Rop ALD HBD
> 15,64 ± 1,00
< 7,0 ± 1,00
< 7,0 ± 1,00
-
13
Rop RBM Rop Tri HBD
-
-
-
11,0 (5; 20)
14
RBD
-
-
-
44,2 (20; 320)
Примечание. Данные по ИФА представлены в виде среднего геометрического титра для группы (log2) ± стандартное гео
метрическое отклонение, данные по вируснейтрализации - в виде среднего геометрического титра и разброса значений
титров сывороток; данные по ИФА для белков Rop D2 Rop Tri HBD, Rop D3 Rop Tri HBD, Rop D2 Rop ALD HBD и
Rop D3 Rop ALD HBD получены ранее [7] и приведены для удобства сравнения. Прочерк означает, что данный белок в
соответствующем эксперименте не использовали. СГТ - средний геометрический титр.
БИОХИМИЯ том 87 вып. 3 2022
316
КАРЯГИНА и др.
Рис. 3. Кратные различия в медианных уровнях цитокинов/хемокинов/факторов роста у мышей, иммунизированных раз
личными антигенами, по сравнению с контрольной группой. * Статистически значимые отличия от контрольной группы
(p < 0,05) по критерию Тьюки
БИОХИМИЯ том 87 вып. 3 2022
КОНФОРМАЦИОННЫЕ ЭПИТОПЫ RBD Spike БЕЛКА SARS CoV 2
317
ным белком Rop RBM Rop Tri HBD и реком
са IgG, в высоком титре взаимодействующих с
бинантным RBD («Mount Sinai»), показали СГТ
использованным для иммунизации антигеном,
на уровне 11,0 и 44,2 (табл. 2).
лишь белки, содержащие эпитопы D2 и D3, в
достаточно высоком титре взаимодействовали
также и с инактивированным вирусом. С RBD
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Spike белка SARS CoV 2, синтезированным в
клетках прокариот, помимо белков с D2 и D3,
В данной работе мы существенно расширили
взаимодействовал также белок Rop D13 Rop
список исследуемых поверхностных эпитопов
Tri HBD. Те же белки, а также белки Rop D4
Spike белка SARS CoV 2: к исследованным ранее
Rop ALD HBD и Rop D4 Rop Tri HBD вызы
эпитопам D2 (470-490 а.о.) и D3 (453-494 а.о.) [7]
вали максимально выраженное повышение
добавилось ещё четыре эпитопа: D4 (496-507 а.о.),
уровня цитокинов в исследовании активации
D6
(144-153
а.о.), D8
(337-346
а.о.) и
клеточного иммунитета. При рассмотрении
D13 (414-425 а.о.) (рис. 1).
профиля повышенных в результате стимуляции
Важным обстоятельством при выборе эпито
спленоцитов антигеном Rop D3 Rop ALD
пов было то, что их последовательности должны
HBD (и в меньшей степени другими белками)
включать а.о. Spike белка, взаимодействующие
уровней цитокинов наблюдается развитие реак
с ACE2 и вируснейтрализующими антителами.
ций врождённого иммунного ответа с потенци
В связывании RBD с рецептором ACE2 участву
альной дифференцировкой B клеток. Также
ют а.о. в следующих положениях: 417, 446, 449,
можно отметить вовлечение самых ранних
453, 455, 456, 475, 486, 487, 489, 493, 496, 498,
признаков развития и других звеньев адаптив
500, 504, 502 и 505 [10]. Большинство этих а.о.
ного иммунного ответа на антиген, причём от
входит в состав выбранных эпитопов D2, D3,
мечается присутствие признаков дифференци
D4, D8 и D13. Ряд описанных в литературе
ровки Т клеток в Th1 (IFN γ), а также присут
нейтрализующих антител связывается с эпито
ствие компонентов Th2 и Th17 (IL 4, IL 5 и IL
пами в RBD, перекрывающимися с эпитопами,
17). Наличие регуляторных цитокинов - стиму
анализируемыми в наших работах: P2B 2F6 [11],
лирующих (IL 1 и IL 12) и супрессирующих
BD23
[12], COVA2 39
[13], REGN10933 и
(IL 10) молекул - характеризует полноценность
REGN10987 [14], LY CoV555 [15].
вызванного антигенами иммунного ответа. Тем
N концевой домен S1 субъединицы Spike
не менее, несмотря на выраженный иммунный
белка (NTD) ограничен 20-286 а.о. Выделено
ответ, исследование вируснейтрализующей ак
несколько нейтрализующих антител, связываю
тивности сывороток, полученных при иммуни
щихся с эпитопами в этом участке: 4A8 [16],
зации мышей с адъювантом 1, в реакции микро
COV2 2676 и COV2 2489 [17], S2L28, S2M28 и
нейтрализации показало отрицательный ре
S2X333 [18]. Так что создание конструкций с
зультат.
эпитопами, соответствующими районам NTD
Известно, что выбор оптимального адъю
(D6), также представляется перспективным.
ванта может иметь решающее значение для фор
Все эпитопы, за исключением D8, представ
мирования иммунного ответа с образованием
ляющего собой α спираль, в структуре Spike
протективных антител при формулировании
белка SARS CoV 2 имеют петлеобразную кон
вакцины для определённого патогена [19]. Име
формацию со сближенными N и C концами.
ются данные о том, что агонисты Toll подобных
Конформацию выбранных эпитопов фиксиро
рецепторов (TLR) могут функционировать в ка
вали с помощью встраивания в место поворота
честве компонентов адъюванта для повышения
между двумя взаимодействующими друг с дру
эффективности инактивированной ультрафио
гом α спиралями Rop подобного белка из
летом вакцины против SARS CoV [20] или вак
M. capsulatus. К такой конструкции на C конец
цинной композиции на основе рекомбинантно
присоединяли тримеризующий модуль, пред
го белка, содержащего эктодомен Spike белка
ставляющий собой либо альдолазу из T. mariti*
SARS CoV [6]. При этом иммунизация мышей
ma, либо тримеризующую α спираль из Spike
эктодоменом Spike белка SARS CoV с наночас
белка SARS CoV 2. На C конец конструкции с
тицами из золота в качестве адъюванта не при
тримеризующим модулем помещали фрагмент
водила к получению протективных антител [6].
гемагглютинина из М. tuberculosis для облегче
Учитывая вышесказанное, мы модифицировали
ния очистки гибридных белков с помощью хро
адъювант для иммунизации - ввели в него вмес
матографии на гепарин сефарозе.
то монтанида ISA 201 и ретинола пальмитата
Интересно отметить, что, несмотря на то что
агонисты TLR - LPS из E. сoli O55:B5 и CpG
абсолютно все полученные гибридные белки
олигодезоксинуклеотиды (ODN 1826). ДЭАЭ
вызывали у мышей образование антител клас
декстран 500, являющийся компонентом обоих
БИОХИМИЯ том 87 вып. 3 2022
318
КАРЯГИНА и др.
адъювантов, в случае второго варианта адъюван
тельно, поскольку все выбранные нами детер
та способствует также удержанию в месте введе
минанты расположены на поверхности Spike
ния (депонированию) CpG олигонуклеотидов.
белка, а тот участок RBD, в пределах которого
При иммунизации мышей гибридными белками
расположены вируснейтрализующие эпитопы,
со вторым вариантом адъюванта удалось полу
непосредственно вовлечён во взаимодействие
чить сыворотки, некоторые из которых проде
с ACE2. Высокая «кучность» вариабельных
монстрировали значимый уровень нейтрализа
участков в области нейтрализующих эпитопов
ции клинического изолята вируса SARS CoV 2
позволит на основе уже имеющихся конструк
в реакции микронейтрализации (табл. 2). При
ций быстро получать измененные варианты
этом наиболее выраженную нейтрализующую
последовательностей и корректировать нейтра
активность продемонстрировали сыворотки,
лизующий ответ, направленный на соответству
полученные при иммунизации гибридными
ющий мутантный вариант коронавируса. Воз
белками Rop D13 Rop Tri HBD, Rop D2 Rop
можно, имеет смысл расширить границы эпито
Tri HBD и Rop D4 Rop Tri HBD. Интересно
пов и использовать для разработки эпитопной
отметить, что все гибридные белки, вызываю
вакцины весь участок RBM, несущий макси
щие образование нейтрализующих антител, по
мальное число вариабельных а.о. Это оправда
лучены с использованием сравнительно неболь
но, поскольку в области RBM отсутствуют сай
шого тримеризационного домена из Spike белка
ты гликозилирования, рекомбинантный белок с
коронавируса, а не достаточно крупного альдо
RBM эффективно нарабатывается в E. coli, де
лазного домена, что позволяет отдать предпоч
монстрирует высокую степень очистки, подвер
тение менее высокомолекулярным белковым
гается правильному рефолдингу с образованием
носителям, смещающим соотношение в сторону
дисульфидной связи и имеет максимальный
преобладания антигенной части белка. Возмож
уровень СГТ в реакции микронейтрализации из
но, положительную роль также играет то, что
всех гибридных белков, полученных микробио
используемый тримеризационный домен про
логическим синтезом.
исходит из Spike белка.
Следует отметить, что, несмотря на положи
Гибридные белки, приводящие к максималь
тельный результат, показанный в реакции мик
но выраженному иммунному ответу с образова
ронейтрализации, демонстрирующий принци
нием нейтрализующих антител, содержат ко
пиальную возможность образования измеримых
роткие петлеобразные эпитопы D2 и D4, распо
количеств нейтрализующих антител при имму
ложенные в пределах RBM, или в области RBD,
низации рекомбинантными белками с неболь
предшествующей RBM (D13). Эти эпитопы в
шими петлеобразными эпитопами RBD Spike
значительной степени перекрываются с линей
белка вируса SARS CoV 2, полученные значе
ными эпитопами S406-420, S475-499 и
ния СГТ сывороток невелики по сравнению с
S495-509 из статьи Lu et al. [5] (рис. 1). По дан
полноразмерным RBD, синтезированным в
ным, представленным в этой статье, все эти ли
клетках эукариот. Это определяет необходи
нейные эпитопы являются вируснейтрализую
мость дальнейшей работы по созданию эпитоп
щими в отношении варианта G614 вируса
ных вакцин для профилактики коронавирусной
SARS CoV 2 [5]. Это хорошо соответствует на
инфекции, которая возможна по следующим
шим результатам по вируснейтрализации кли
направлениям: 1) поиск новых перспективных
нического изолята вируса hCoV 19/Russia/
вируснейтрализующих эпитопов в составе бел
StPetersburg 3524/2020 EPI_ISL_415710 GISAID,
ков SARS CoV 2, например, на основе исполь
также несущего замену D614G. При этом мак
зования экспериментальных данных по характе
симальную способность к образованию нейтра
ристике линейных эпитопов, хорошо коррели
лизующих антител, приближающуюся по значе
рующих с данными по конформационным эпи
нию СГТ к белку с целым RBM, продемонстри
топам; 2) комбинированное использование не
ровал белок, несущий короткий петлеобразный
скольких гибридных белков для иммунизации с
эпитоп D4.
варьированием дозы антигенов; 3) укрупнение
Все выбранные нами эпитопы несут а.о., му
петлеобразных эпитопов, одним из которых мо
тировавшие в Spike белке варианта SARS CoV 2
жет быть RBM; 4) оптимизация платформы для
Омикрон (B.1.1.529), вызвавшего максималь
мультимеризации эпитопов.
ную волну заболеваемости COVID 19 по срав
Поскольку одним из наиболее значимых ре
нению с предыдущими [21]. При этом наиболь
зультатов работы является подбор адъюванта,
шее количество вариабельных а.о. сосредоточе
способствующего формированию вируснейтра
но именно в области эпитопов, демонстрирую
лизующих антител даже при использовании в
щих максимальный уровень вируснейтрализа
качестве антигенов рекомбинантных белков с
ции по нашим данным (рис. 1). Это неудиви
очень короткими участками последовательнос
БИОХИМИЯ том 87 вып. 3 2022
КОНФОРМАЦИОННЫЕ ЭПИТОПЫ RBD Spike БЕЛКА SARS CoV 2
319
ти Spike белка вируса SARS CoV 2, было бы ин
вация клеточного иммунитета при иммуниза
тересно охарактеризовать иммуногенность бел
ции рекомбинантными белками по уровню син
ков, вызывающих образование нейтрализую
теза цитокинов спленоцитами иммунизирован
щих антител, способность получаемых при им
ных мышей. После модификации адъюванта,
мунизации ими сывороток связываться с RBD
заключающейся во введении в него агонис
Spike белка SARS CoV 2 и инактивированным
тов TLR - LPS и CpG олигонуклеотидов - для
вирусом, а также оценить влияние иммуниза
ряда белков показана нейтрализующая актив
ции этими белками на клеточный иммунитет,
ность сывороток в реакции микронейтрализа
чтобы иметь возможность предсказывать ней
ции в отношении живого вируса SARS CoV 2.
трализующую активность белков на основании
более простых экспериментов без использова
Финансирование. Исследование выполнено
ния живого вируса. Это также может быть од
при поддержке Российского фонда фундамен
ним из направлений дальнейших исследований.
тальных исследований (проект № 20 04 60373\
20). Получение белка Rop RBM Rop Tri HBD и
Таким образом, в результате проведённой
исследование его способности вызывать образо
работы получены гибридные рекомбинантные
вание вируснейтрализующих антител было про
белки, включающие небольшие петлеобразные
ведено в рамках государственного задания Ми
конформационные эпитопы из RBD и N конце
нистерства здравоохранения Российской Феде
вого домена Spike белка вируса SARS CoV 2.
рации № 056 00119 21 00 («Разработка препара
Показана высокая иммуногенность полученных
та блокатора рецептора АСЕ2 для профилактики
белков. Для ряда белков, содержащих эпитопы
и терапии коронавирусной инфекции»).
из RBD Spike белка вируса SARS CoV 2, проде
Конфликт интересов. Авторы заявляют об от
монстрирована способность сывороток, полу
сутствии конфликта интересов.
ченных при иммунизации данными белками, в
Соблюдение этических норм. Все примени
высоком титре связываться с RBD, синтезиро
мые международные, национальные и/или ин
ванным в клетках эукариот, и с инактивирован
ституциональные принципы ухода и использо
ным вирусом SARS CoV 2. Исследована акти
вания животных были соблюдены.
СПИСОК ЛИТЕРАТУРЫ
1.
Burton, D. R. (2002) Antibodies, viruses and vaccines,
8. Manenti, A., Maggetti, M., Casa, E., Martinuzzi, D.,
Nat. Rev. Immunol., 9, 706 713, doi: 10.1038/nri891.
Torelli, A., et al. (2020) Evaluation of SARS CoV 2 neu
2.
Neu, K. E., Henry Dunand, C. J., and Wilson, P. C. (2016)
tralizing antibodies using a CPE based colorimetric live
Heads, stalks and everything else: how can antibodies erad
virus micro neutralization assay in human serum sam
icate influenza as a human disease? Curr. Opin. Immunol.,
ples, J. Med. Virol., 92, 2096 2104, doi: 10.1002/jmv.
42, 48 55, doi: 10.1016/j.coi.2016.05.012.
25986.
3.
Sok, D., and Burton, D. R. (2018) Recent progress in
9. Barchuk, A., Shirokov, D., Sergeeva, M., Tursun Zade, R.,
broadly neutralizing antibodies to HIV, Nat. Immunol., 19,
Dudkina, O., et al. (2021) Evaluation of the performance
1179 1188, doi: 10.1038/s41590 018 0235 7.
of SARS CoV 2 antibody assays for a longitudinal popula
4.
Dejnirattisai, W., Supasa, P., Wongwiwat, W.,
tion based study of COVID 19 spread in St. Petersburg,
Rouvinski, A., Barba Spaeth, G., et al. (2016) Dengue
Russia, J. Med. Virol., 93, 5846 5852, doi: 10.1002/jmv.
virus sero cross reactivity drives antibody dependent
27126.
enhancement of infection with zika virus, Nat. Immunol.,
10. Lan, J., Ge, J., Yu, J., Shan, S., Zhou, H., et al. (2020)
17, 1102 1108, doi: 10.1038/ni.3515.
Structure of the SARS CoV 2 spike receptor binding
5.
Lu, S., Xie, X. X., Zhao, L., Wang, B., Zhu, J., et al. (2021)
domain bound to the ACE2 receptor, Nature, 581, 215
The immunodominant and neutralization linear epitopes
220, doi: 10.1038/s41586 020 2180 5.
for SARS CoV 2, Cell Rep., 34, 108666, doi: 10.1016/
11. Ju, B., Zhang, Q., Ge, J., Wang, R., Sun, J., et al. (2020)
j.celrep.2020.108666.
Human neutralizing antibodies elicited by SARS CoV 2
6.
Sekimukai, H., Iwata Yoshikawa, N., Fukushi, S.,
infection, Nature, 584, 115 119, doi: 10.1038/s41586 020
Tani, H., Kataoka, M., et al. (2020) Gold nanoparticle
2380 z.
adjuvanted S protein induces a strong antigen specific IgG
12. Cao, Y., Su, B., Guo, X., Sun, W., Deng, Y., et al. (2020)
response against severe acute respiratory syndrome related
Potent neutralizing antibodies against SARS CoV 2 iden
coronavirus infection, but fails to induce protective anti
tified by high throughput single cell sequencing of conva
bodies and limit eosinophilic infiltration in lungs,
lescent patients’ B cells, Cell,
182,
7384.e16,
Microbiol. Immunol., 64, 3351, doi: 10.1111/1348
doi: 10.1016/j.cell.2020.05.025.
0421.12754.
13. Brouwer, P., Caniels, T. G., van der Straten, K., Snitselaar,
7.
Karyagina, A. S., Gromov, A. V., Grunina, A. M.,
J. L., Aldon, Y., et al. (2020) Potent neutralizing antibodies
Lyaschuk, A. M., Grishin, A. V., et al. (2021) Development
from COVID 19 patients define multiple targets of vulner
of a platform for producing recombinant protein compo
ability, Science, 369, 643650, doi: 10.1126/science.
nents of epitope vaccines for the prevention of COVID 19,
abc5902.
Biochemistry (Moscow), 86, 12751287, doi: 10.1134/
14. Hansen, J., Baum, A., Pascal, K. E., Russo, V.,
S0006297921100096.
Giordano, S., et al. (2020) Studies in humanized mice and
БИОХИМИЯ том 87 вып. 3 2022
320
КАРЯГИНА и др.
convalescent humans yield a SARS CoV 2 antibody cock
18. McCallum, M., De Marco, A., Lempp, F. A., Tortorici,
tail, Science, 369, 10101014, doi: 10.1126/science.
M. A., Pinto, D., et al. (2021) N terminal domain antigenic
abd0827.
mapping reveals a site of vulnerability for SARS CoV 2,
15.
Jones, B. E., Brown Augsburger, P. L., Corbett, K. S.,
Cell, 184, 2332 2347.e16, doi: 10.1016/j.cell.2021.03.028.
Westendorf, K., Davies, J., et al. (2021) The neutralizing
19. Sarkar, I., Garg, R., and van Drunen Littel van den Hurk,
antibody, LY CoV555, protects against SARS CoV 2
S. (2019) Selection of adjuvants for vaccines targeting spe
infection in nonhuman primates, Sci. Transl. Med., 13,
cific pathogens, Expert Rev. Vaccines, 18, 505521,
eabf1906, doi: 10.1126/scitranslmed.abf1906.
doi: 10.1080/14760584.2019.1604231.
16.
Chi, X., Yan, R., Zhang, J., Zhang, G., Zhang, Y., et al.
20. Iwata Yoshikawa, N., Uda, A., Suzuki, T., Tsunetsugu
(2020) A neutralizing human antibody binds to the N ter
Yokota, Y., Sato, Y., et al. (2014) Effects of Toll like recep
minal domain of the Spike protein of SARS CoV 2,
tor stimulation on eosinophilic infiltration in lungs of
Science, 369, 650 655, doi: 10.1126/science.abc6952.
BALB/c mice immunized with UV inactivated severe
17.
Suryadevara, N., Shrihari, S., Gilchuk, P., VanBlargan,
acute respiratory syndrome related coronavirus vaccine,
L. A., Binshtein, E., et al. (2021) Neutralizing and protec
J. Virol., 88, 8597 8614, doi: 10.1128/JVI.00983 14.
tive human monoclonal antibodies recognizing the N ter
21. He, X., Hong, W., Pan, X., Lu, G., and Wei, X. (2021)
minal domain of the SARS CoV 2 spike protein, Cell, 184,
SARS CoV 2 Omicron variant: Characteristics and pre
2316 2331.e15, doi: 10.1016/j.cell.2021.03.029.
vention, MedComm, 2, 838 845, doi: 10.1002/mco2.110.
HYBRID PROTEINS WITH SHORT CONFORMATIONAL EPITOPES
OF THE RECEPTOR BINDING DOMAIN OF THE SARS CoV 2 SPIKE
PROTEIN PROMOTE THE PRODUCTION OF VIRUS NEUTRALIZING
ANTIBODIES DURING IMMUNIZATION
A. S. Karyagina1,2,3*, A. V. Gromov1, T. M. Grunina1,2, A. M. Lyaschuk1, M. S. Poponova1,
D. A. Kleymenov1, N. V. Strukova1, M. S. Generalova1, A. V. Ryazanova1, Z. M. Galushkina1,
O. Yu. Dobrynina1, T. N. Bolshakova1, M. V. Sergeeva4, E. A. Romanovskaya Romanko4,
I. V. Krasilnikov5, M. E. Subbotina1,2*, and V. G. Lunin1,2
1 Gamaleya National Research Center of Epidemiology and Microbiology,
Ministry of Healthcare of the Russian Federation, 123098 Moscow, Russia;
e*mail: akaryagina@gmail.com; e*mail: subbotinam@gmail.com
2 All*Russia Research Institute of Agricultural Biotechnology, 127550 Moscow, Russia
3 Belozersky Institute of Physico*Chemical Biology, Lomonosov Moscow State University, 119992 Moscow, Russia
4 Institute of Influenza, Ministry of Healthcare of the Russian Federation, 197376 St. Petersburg, Russia
5 Saint Petersburg Institute of Vaccines and Sera, FMBA, 198320 St. Petersburg, Russia
Based on the previously proposed approach, hybrid recombinant proteins, potential components of epitope vaccines
containing short conformational epitopes of the receptor binding domain of the Spike protein of the SARS CoV 2
virus, were obtained by synthesis in Escherichia coli cells (144 153, 337 346, 414 425, 496 507 a.a.), which imple
ment a large number of protein protein interactions in complexes with neutralizing antibodies and ACE2
(angiotensin converting enzyme 2). The obtained proteins were used for immunization of mice three times with an
interval of 2 weeks, and the immunogenicity of protein antigens and the ability of sera to interact with the inactivat
ed SARS CoV 2 virus and with the RBD of the Spike protein obtained in a eukaryotic producer were studied. All
recombinant proteins showed high immunogenicity. The highest titer upon binding to the RBD of the Spike protein
was demonstrated by the serum obtained by immunization with a protein based on epitope 414 425. At the same time,
the serum titers upon binding to the inactivated virus and RBD in the case of other proteins turned out to be signifi
cantly lower than the titers for the previously obtained four proteins based on loop like epitopes 452 494 and 470 491,
the conformation of which was additionally fixed by a disulfide bond. For all proteins obtained earlier and in this work,
we studied the activation of cellular immunity, which was detected by the level of cytokine synthesis by splenocytes of
immunized mice. The most pronounced level of increased cytokine synthesis was observed in the case of a response
to proteins, including epitopes with disulfide bonds (452 494, 470 491), as well as epitopes 414 425 and 496 507.
After optimization of the adjuvant for some recombinant proteins with short conformational epitopes of the RBD of
the Spike protein, mouse sera were obtained showing a virus neutralizing effect in the microneutralization reaction
with live virus SARS CoV 2 (hCoV 19/Russia/StPetersburg 3524/2020 EPI_ISL_415710 GISAID). The results
obtained can be used to develop epitope vaccines for the prevention of COVID 19 and other viral infections.
Keywords: SARS CoV 2, Spike protein, RBD, RBM, epitope vaccine, epitope, aldolase
БИОХИМИЯ том 87 вып. 3 2022