БИОХИМИЯ, 2022, том 87, вып. 6, с. 780 - 793
УДК 62.018.2:017.12
Т-ЛИМФОЦИТЫ КАК МИШЕНИ ДЛЯ SARS-CoV-2
Обзор
© 2022 Е.М. Куклина
ФГБУН Пермский федеральный исследовательский центр УрО РАН,
«Институт экологии и генетики микроорганизмов УрО РАН»,
614081 Пермь, Россия; электронная почта: ibis_07@mail.ru
Поступила в редакцию 19.04.2022
После доработки 17.05.2022
Принята к публикации 17.05.2022
Несмотря на многочисленные данные об отсутствии или слабой экспрессии Т-клетками главного
функционального рецептора SARS-CoV-2, ангиотензин I-превращающего фермента 2 (ACE2), по-
следние данные литературы демонстрируют способность нового коронавируса эффективно инфи-
цировать Т-лимфоциты. Обзор посвящён анализу этих работ: он рассматривает альтернативные
(АСЕ2-независимые) пути инфицирования клеток, определяет Т-клеточные субпопуляции, служа-
щие наиболее вероятными мишенями SARS-CoV-2, обсуждает формат взаимодействия вируса с клет-
кой, включая как инфекционные, так и неинфекционные механизмы регуляции Т-лимфоцитов,
а также оценивает роль вирус-зависимого поражения Т-лимфоцитов в патогенезе COVID-19. Особое
внимание уделено регуляторным Т-клеткам, как потенциальным мишеням SARS-CoV-2, а также воз-
можному участию экзосом в регуляции чувствительности к вирусу Т-лимфоцитов, присутствующих
в периферических тканях.
КЛЮЧЕВЫЕ СЛОВА: SARS-CoV-2, T-лимфоциты, ACE2, CD147, Treg.
DOI: 10.31857/S0320972522060069, EDN: AUEWQV
ВВЕДЕНИЕ
рецепторов к различным поверхностным струк-
турам вируса, в первую очередь к белку шипа
Коронавирус
2019 г., вызывающий тя-
(Spike, S-белок). Главным функциональным
жёлый острый респираторный синдром (Severe
рецептором SARS-CoV-2 является ангиотензин
acute respiratory syndrome corоnavirus 2, SARS-
I-превращающий фермент 2 (angiotensin I-con-
CoV-2), имеет тропность к широкому спектру
verting enzyme 2, ACE2) - он распознает S-белок
клеток и тканей, прежде всего к эпителиальным
вируса, а сериновые протеазы клетки-мишени,
клеткам лёгких и респираторного тракта, к со-
выступая в роли корецепторов, обеспечивают
судистому эндотелию, а также к клеткам ки-
проникновение вируса в клетку [1, 2]. Харак-
шечника, печени, почек, мозга [1], что обуслов-
тер распределения АСЕ2 в тканях организма в
ливает многообразие клинических проявлений
целом совпадает с профилем инфицирования
и пост-ковидных осложнений. Тропность виру-
SARS-CoV-2 [1].
са определяется наличием на клетках-мишенях
Клетки иммунной системы и, в частности
Т-лимфоциты, первоначально не рассматрива-
Принятые сокращения: ACE2 - ангиотензин I-превра-
лись в качестве потенциальных мишеней SARS-
щающий фермент 2; ASGR1 - асиалогликопротеиновый
CoV-2, исходя из данных об отсутствии экспрес-
рецептор 1; AXL - рецепторная тирозинкиназа; CD - мем-
сии рецептора для вируса (АСЕ2) в лимфоидных
бранные маркёры лейкоцитов; COVID - болезнь, вызывае-
органах - селезёнке, тимусе и лимфатических
мая коронавирусом; KREMEN1 - Kringle-домен-содержа-
щий трансмембранный белок 1; MOI - множественность
узлах человека [3], а также в различных субпо-
заражения; N-белок - нуклеокапсидный белок; RBD - ре-
пуляциях циркулирующих Т-лимфоцитов [4].
цептор-связывающий домен; S-белок - белок шипа ви-
Однако интенсивные исследования последних
руса; SARS-CoV - коронавирус, вызывающий тяжёлый
лет позволили идентифицировать альтерна-
острый респираторный синдром; TCR - Т-клеточный ре-
цептор; Treg - регуляторные Т-лимфоциты.
тивные рецепторы SARS-CoV-2, некоторые из
780
Т-ЛИМФОЦИТЫ КАК МИШЕНИ ДЛЯ SARS-CoV-2
781
которых представлены на мембране Т-клеток.
вую очередь TMPRSS2 и фурином [2, 11]. Эндо-
Так, трансмембранный гликопротеин CD147 [5]
сомальные цистеиновые протеазы, катепсины B
конститутивно экспрессируется интактными
и L (CatB/L), также вносят вклад в этот процесс,
CD4+/CD8+ Т-лимфоцитами периферической
хотя делают это только альтернативным путём,
крови человека и существенно усиливает экс-
после эндоцитоза вириона [2]. Молекула ACE2
прессию в ответ на поликлональную актива-
широко представлена в тканях человека с мак-
цию клеток [5], так же, как и другой кандидат на
симальной экспрессией на альвеолярных и
роль рецептора для SARS-CoV-2 - рецепторная
бронхиальных эпителиальных клетках, энтеро-
тирозинкиназа AXL [6, 7]. А экспрессия гена
цитах, сосудистом эндотелии [1, 3]. Такой ха-
Kringle-домен-содержащего трансмембранного
рактер распределения рецепторов совпадает с
белка 1 (Kringle containing transmembrane pro-
профилем инфицирования SARS-CoV-2 [1], но
tein 1, KREMEN1) выявлена в субпопуляции ре-
не объясняет преимущественного повреждения
гуляторных Т-лимфоцитов (Treg) [8, 9]. И хотя
клеток лёгких и дыхательных путей. Фурин так-
в традиционных тканях-мишенях вируса эти
же выявляется повсеместно, тогда как экспрес-
рецепторы играют вспомогательную роль [7, 8],
сия TMPRSS2 носит ограниченный характер с
они могут выходить на первый план в АСЕ2-де-
максимумом как раз на пневмоцитах, что, как
фицитных клетках - например, в иммунных.
полагают, вносит вклад в распространение ви-
Эти данные подняли вопрос о Т-лимфо-
руса и специфику патологии [12, 13].
цитах, как о потенциальных мишенях нового
Однако ACE2 - не единственный рецептор
коронавируса. На сегодняшний день имеется
SARS-CoV-2. Недавние исследования выявили
6 независимых работ, с разной степенью дета-
ещё целый ряд кандидатов на эту роль. Первый
лизации продемонстрировавших способность
и главный из них - CD147, трансмембранный
нового коронавируса SARS-CoV-2 инфициро-
белок семейства иммуноглобулинов (извест-
вать Т-лимфоциты человека. Настоящий об-
ный также как басигин или EMMPRIN), уча-
зор посвящён анализу данных работ, обсужде-
ствующий в инвазии Plasmodium falciparum [14],
нию возможных механизмов инфицирования
некоторых бактериальных и вирусных инфек-
Т-клеток и вклада этих механизмов в патогенез
циях [15, 16], включая и коронавирус 2003 г.,
COVID-19.
SARS-CoV-1 [17]. В 2020 г. Wang et al. [5] сообщи-
ли о прямом взаимодействии CD147 с S-белком
SARS-CoV-2 и его участии в инфицировании
РЕЦЕПТОРЫ SARS-CoV-2
клетки вирусом - на основе связи инфективно-
сти SARS-CoV-2 с уровнем экспрессии CD147 на
Молекула ACE2, ключевой компонент ре-
клетках-мишенях. Правда, две работы 2021 г. не
нин-ангиотензиновой системы, идентифици-
выявили прямого взаимодействия CD147 с ре-
рована как основной входной рецептор для
комбинантным S-белком SARS-CoV-2 [18, 19],
SARS-CoV-2 [1, 2], так же, как и для родствен-
равно как и изменения уровня инфицирования
ного коронавируса 2003 г., SARS-CoV-1 [10].
клеток вирусом в случае подавления экспрессии
ACE2 связывается с S-белком SARS-CoV-2 че-
гена, кодирующего CD147 [18], или функцио-
рез С-терминальный домен S1-субъединицы,
нальной блокады данной молекулы [19]. Тем
так называемый рецептор-связывающий домен
не менее последняя работа по данной пробле-
(receptor-binding domain, RBD) [1, 2]. Следую-
ме [20] подтвердила результаты, полученные
щим шагом в ACE2-зависимом инфицировании
Wang et al. [5]. Таким образом, вопрос о роли
клетки SARS-CoV-2 является протеолитиче-
CD147 в инфицировании клеток SARS-CoV-2
ское расщепление S-белка вируса между S1/S2-
до конца не решён и требует дальнейших ис-
субъединицами и при S2′-сайте, что приводит к
следований - возможно, CD147 участвует в
высвобождению фузогена - фактора, обеспечи-
этом процессе не напрямую. Молекула CD147
вающего слияние оболочки вируса с мембраной
широко представлена в различных тканях: она
клетки-мишени [2]. Слияние может происхо-
экспрессируется эпителиальными и нейрональ-
дить двумя путями - на клеточной мембране по-
ными клетками, а также лейкоцитами - отсюда
сле связывания рецептора или в эндосоме после
её название, CD147, в соответствии с номенкла-
поглощения вириона. В последнем случае обо-
турой лейкоцитарных дифференцировочных
лочка вируса сливается с эндосомальной мем-
антигенов [4].
браной. Расщепление S-белка осуществляется
Другие кандидаты на роль рецепторов,
сериновыми протеазами клетки-хозяина, в пер-
обеспечивающих вход SARS-CoV-2 в клетку, -
БИОХИМИЯ том 87 вып. 6 2022
782
КУКЛИНА
рецепторная тирозинкиназа AXL [7], а также
сульфаты [29] или лектиновые рецепторы [30].
трансмембранный белок KREMEN1 и асиало-
Нейропилин-1 напрямую связывается с по-
гликопротеиновый рецептор 1 (Asialoglycopro-
лиосновным мотивом в расщеплённой фури-
tein receptor 1, ASGR1) [8].
ном S1-субъединице S-белка вируса и заметно
Рецепторная тирозинкиназа AXL широ-
усиливает инфицирование клетки: блокада та-
ко представлена в различных органах и тканях
кого взаимодействия с помощью РНК-интер-
человека и регулирует многие физиологиче-
ференции или селективных ингибиторов сни-
ские процессы, включая клеточное выживание,
жает вход SARS-CoV-2 в клетку [27]. При этом
пролиферацию и дифференцировку [21, 22].
нейропилин-1 не способен вызывать инфици-
Показано, что она специфически связывается
рование клетки вирусом самостоятельно, но
с N-терминальным доменом S1-субъединицы
эффективно делает это в случае коэкспрессии с
S-белка SARS-CoV-2, но не с классическим
ACE2 и TMPRSS2 [27]. Гепарансульфат - дру-
RBD [7], и опосредует вход вируса в клетку:
гой необходимый фактор, усиливающий ин-
блокада экспрессии AXL существенно снижает
фицирование различных клеток-мишеней
инфицирование клеток лёгочной линии SARS-
SARS-CoV-2 [29]. Он взаимодействует с RBD
CoV-2, тогда как сверхэкспрессия этого фактора
S-белка вируса в сайте, не перекрывающимся с
промотирует вирусную инфекцию [7]. Тем не
мотивом, участвующим в контакте с ACE2. Свя-
менее репликация вируса в клетках лёгких лишь
зывание гепарансульфата инициирует транс-
незначительно подавляется на фоне блокады
формацию S-белка из закрытой конформации
AXL [7], указывая на второстепенную роль дан-
в открытую, повышая доступность RBD для
ного фактора в традиционных мишенях SARS-
связывания ACE2 [29]. Некоторые лектиновые
CoV-2.
рецепторы также участвуют в инфицировании
Два дополнительных рецептора для вируса -
клетки SARS-CoV-2 за счёт связывания глика-
трансмембранные белки ASGR1 и KREMEN1:
нов, ассоциированных с S-белком [30]. Боль-
ASGR1 опосредует эндоцитоз и лизосомальную
шинство лектинов С-типа взаимодействуют с
деградацию ряда гликопротеинов, играя крити-
сайтом S-белка за пределами RBD. И хотя такое
ческую роль в их гомеостазе [23], а KREMEN1
связывание не способно напрямую индуциро-
участвует в регуляции WNT-зависимого сигна-
вать инфицирование клетки SARS-CoV-2, лек-
ла [24], и оба фактора служат входными рецепто-
тиновые рецепторы усиливают вход вируса в
рами для многих вирусов [25], включая и новый
ACE2-позитивные клетки, выступая в качестве
коронавирус. Молекулы ASGR1 и KREMEN1
неспецифических «молекул прикрепления» для
напрямую взаимодействуют с S-белком SARS-
SARS-CoV-2 [30]. К тому же N-терминальный
CoV-2 (как с N-терминальным доменом, так
домен S1-субъединицы S-белка вируса содер-
и с RBD), причём аффинность их связывания
жит мотивы, связывающие сиаловые кислоты,
сопоставима с таковой для ACE2 [8, 26]. Экто-
и эти мотивы могут опосредовать взаимодей-
пической экспрессии ASGR1 или KREMEN1
ствие вируса с различными сиалопротеинами,
достаточно для входа SARS-CoV-2 в клетку [8],
гликопротеинами или ганглиозидами на кле-
однако избыточная экспрессия этих рецепто-
точной мембране [28]. Таким образом, хотя пе-
ров в ACE2-негативных клетках лишь частично
речисленные выше неспецифические молекулы
восстанавливает инфективность SARS-CoV-2, с
не являются самодостаточными, они вносят
существенно меньшей эффективностью, чем в
существенный вклад в инфицирование клетки
ACE2-позитивных [8].
SARS-CoV-2 - либо обеспечивая лучшее при-
Каждый из четырёх альтернативных рецеп-
крепление вируса к поверхности клетки-мише-
торов (CD147/AXL/KREMEN1/ASGR1) потен-
ни, либо изменяя конформацию S-белка, что
циально может опосредовать вход SARS-CoV-2
приводит к появлению дополнительных сайтов
в клетку независимо от ACE2, обеспечивая
связывания с рецептором или повышает до-
дополнительные пути инфицирования виру-
ступность RBD для контакта с ACE2.
сом различных тканей, хотя в традиционных
АСЕ2-позитивных мишенях вируса их роль вто-
рична.
ЭКСПРЕССИЯ РЕЦЕПТОРОВ
В отличие от независимых рецепторов, есть
SARS-CoV-2 Т-ЛИМФОЦИТАМИ
ряд мембранных молекул, способных усиливать
инфективность SARS-CoV-2, таких как нейро-
Вопрос о наличии у Т-лимфоцитов главно-
пилин-1 [27], сиаловые кислоты [28], гепаран-
го функционального рецептора SARS-CoV-2 на
БИОХИМИЯ том 87 вып. 6 2022
Т-ЛИМФОЦИТЫ КАК МИШЕНИ ДЛЯ SARS-CoV-2
783
сегдняшний день является спорным. В 2004 г.
терферон-стимулируемым [34], а Т-лимфоциты
Hamming et al. [3] продемонстрировали с по-
активно отвечают на интерфероны.
мощью иммуногистохимического анализа
Кроме того, возможен и ещё один меха-
отсутствие экспрессии ACE2 в лимфоцитах
низм появления АСЕ2 на Т-клеточной мем-
центральных и периферических лимфоидных
бране - он связан с экзосомами: клетки, не
органов человека, а именно: тимуса (число ис-
экспрессирующие или слабо экспрессирующие
следованных образцов, n = 4), селезёнки (n = 4)
рецептор SARS-CoV-2, могут получать этот ре-
и лимфатических узлов (n = 6). Именно на эту
цептор от других клеток с помощью экстракле-
работу до недавнего времени ссылалось боль-
точных микровезикул, которые высвобождают-
шинство авторов, высказывая сомнение в воз-
ся от поверхности любых клеток в норме или
можности инфицирования Т-клеток вирусом
при стимуляции, содержат биомолекулы, такие
SARS-CoV-2 [31]. В подтверждение этих данных
как РНК и белки, и осуществляют межклеточ-
в 2020 г. Radzikowska et al. [4] показали отсут-
ную коммуникацию [35]. Известно, что АСЕ2
ствие экспрессии генов, кодирующих ACE2 и
является традиционным компонентом микро-
TMPRSS2 в различных субпопуляциях Т-лим-
везикул, которые способны переносить его в
фоцитов периферической крови человека,
совместной культуре от клеток со сверхэкспрес-
включая наивные CD4+/CD8+ Т-лимфоциты
сией АСЕ2 клеткам-реципиентам, лишённым
и терминально дифференцированные эффек-
данного фактора [36]. Более того, в недавней
торные CD4+/CD8+ Т-клетки - правда, опять
работе показано присутствие АСЕ2+-микро-
на ограниченной группе (n = 4). В то же время
везикул в плазме больных COVID-19 - их уро-
Bertram et al. [32] продемонстрировали, так-
вень имел высокую вариабельность и в целом
же с помощью иммуногистохимии, высокую
был сопоставим с таковым у здоровых доноров,
стабильную экспрессию ACE2 и TMPRSS2 в
однако в группе больных COVID-19 количество
лимфоидных клетках различных тканей ре-
АСЕ2+-микровезикул коррелировало с тяже-
спираторного и желудочно-кишечного тракта,
стью заболевания [37].
включая респираторный синус, слизистые мин-
В отличие от АСЕ2, недавно идентифици-
далин и ворсинки кишечника. А ещё в одном
рованный новый функциональный рецептор
исследовании с помощью секвенирования РНК
SARS-CoV-2, трансмембранный гликопротеин
на уровне отдельного ядра (single-nuclei RNA
CD147 [5], конститутивно экспрессируется на
sequencing analysis) [33] показана заметная экс-
мембране интактных CD4+/CD8+ Т-лимфоци-
прессия гена, кодирующего TMPRSS2, и в сле-
тов периферической крови человека и суще-
довых количествах гена, кодирующего АСЕ2,
ственно усиливает экспрессию в ответ на поли-
Т-лимфоцитами, присутствующими в здоро-
клональную активацию клеток [5], так же, как и
вой ткани лёгких онкологических пациентов
другой кандидат на роль рецептора для SARS-
(n = 12). Противоречия в представленных выше
CoV-2 - рецепторная тирозинкиназа AXL [6].
данных нет. В первом случае объектом исследо-
А экспрессия гена трансмембранного белка
вания являлись преимущественно Т-клеточные
KREMEN1 выявлена в субпопуляции регуля-
предшественники или наивные Т-лимфоциты,
торных Т-лимфоцитов [9].
тогда как во втором случае речь, по-видимому,
Таким образом, хотя вопрос об экспрессии
идёт либо о лимфоидной ткани, ассоциирован-
Т-лимфоцитами главного функционального ре-
ной со слизистыми, либо об активированных
цептора АСЕ2 не до конца ясен, убедительно
Т-лимфоцитах, инфильтрирующих перифери-
подтверждено присутствие на мембране кле-
ческие ткани. Кроме того, размеры исследован-
ток альтернативных рецепторов SARS-CoV-2.
ных групп невелики, а экспрессия АСЕ2 гене-
И даже если в традиционных мишенях эти ре-
тически детерминирована. В итоге, имеющиеся
цепторы играют второстепенную роль, они мо-
на сегодняшний день данные позволяют гово-
гут выходить на первый план в клетках с низкой
рить об отсутствии конститутивной экспрес-
экспрессией АСЕ2 или её отсутствием - напри-
сии ACE2 и TMPRSS2 основной популяцией
мер, в иммунных клетках.
Т-лимфоцитов, наивными αβТ-клетками, но
Важно также отметить, что недавнее струк-
не исключают присутствия данных молекул на
турное компьютерное исследование иммунных
отдельных Т-клеточных субпопуляциях, а также
рецепторов выявило целый ряд новых потен-
их появления в условиях стимуляции или про-
циальных мишеней SARS-CoV-2: показано,
воспалительного окружения, особенно учиты-
что молекулы CD26, CD2, CD56, CD7, CCR9,
вая, что ген, кодирующий АСЕ2, является ин-
CD150, CD4, CD50, XCR1 и CD106, теоретиче-
БИОХИМИЯ том 87 вып. 6 2022
784
КУКЛИНА
ски, имеют более высокую аффинность связы-
больных COVID-19
[40]. При инфицирова-
вания с RBD S-белка вируса, чем классический
нии вирусом CD4+ Т-клеточных линий Jurkat
рецептор ACE2 [38]. Многие из этих молекул
или MT4 in vitro (множественность заражения
экспрессируются Т-лимфоцитами и даже слу-
(Multiplicity of infection, MOI) равна 0,01) в клет-
жат маркёрами Т-клеточных субпопуляций
ках регистрировали вирусную РНК (район, ко-
(например, CD2, CD7, CD4), но их участие в
дирующий RBD), субгеномную вирусную РНК
инфицировании требует экспериментального
(маркёр репликации вируса в клетке), а также
подтверждения.
вирусный белок (N-белок - вестерн-блоттин-
гом/проточной цитометрией) и вирусные ча-
стицы (электронной микроскопией). Оценка
Т-ЛИМФОЦИТЫ И SARS-CoV-2
инфективности вируса в отношении первичных
Т-лимфоцитов периферической крови здоро-
Инфицирование Т-лимфоцитов SARS-CoV-2.
вых доноров (n = 3) также продемонстрировала
Несмотря на данные об отсутствии экспрессии
присутствие вирусной РНК (RBD), более высо-
Т-лимфоцитами молекулы АСЕ2 [3], для род-
кое в предварительно активированных (CD3/
ственного коронавируса SARS-CoV-1, исполь-
CD28/IL-2) Т-клетках [40]. При этом авторы
зующего тот же функциональный рецептор, по-
выявили массированный апоптоз в Т-лим-
казана способность эффективно инфицировать
фоцитах, инфицированных вирусом in vitro, а
Т-клетки: у 27% пациентов с SARS-CoV-1 выяв-
также повышенный уровень апоптотических
лено присутствие вирусных частиц (электрон-
Т-лимфоцитов в периферической крови боль-
ная микроскопия) и мРНК (гибридизация in situ
ных COVID-19 по сравнению с Т-клетками здо-
и RT-PCR) в циркулирующих Т-лимфоцитах
ровых доноров [40].
(CD3+-клетках), а также во вторичных лимфо-
Ещё две работы, непосредственно отно-
идных органах - селезёнке и лимфатических
сящиеся к обсуждаемой проблеме, находят-
узлах [39]. И лимфопению, сопровождающую
ся в настоящее время на этапе препринта, но
данное заболевание, авторы связывают именно
определённо заслуживают обсуждения, по-
с поражением вирусом иммунной системы [39].
скольку перекликаются с предыдущими. В пер-
Неудивительно, что аналогичные исследо-
вой из них показана способность SARS-CoV-2
вания в отношении нового коронавируса SARS-
(MOI = 0,1) инфицировать in vitro CD4+, но не
CoV-2 начались с момента его идентификации.
CD8+ Т-лимфоциты периферической крови
Первой вышла работа Wang et al. [5]. Ав-
здоровых доноров, что подтверждено на уров-
торы выявили вирионы SARS-CoV-2 в Т-лим-
не вирусной РНК, вирусного белка (S-белка
фоцитах (CD3+-клетках), инфильтрирующих
SARS-CoV-2, иммунофлуоресценцией), а так-
лёгочные ткани больных COVID-19 [5], а в экс-
же вирусных частиц (электронной микроско-
перименте in vitro с помощью люциферазного
пией) [41]. При этом в инфицированных CD4+
репортёрного анализа продемонстрировали
Т-клетках выявлена негативная (антисмысло-
дозозависимое инфицирование CD4+/CD8+
вая) РНК SARS-CoV-2, что указывает на ре-
Т-лимфоцитов периферической крови челове-
пликацию вируса [41]. В подтверждение этих
ка (n = 6) псевдовирусом SARS-CoV-2, суще-
результатов, исследования ex vivo выявили при-
ственно более эффективное в случае предвари-
сутствие вирусной РНК в CD4+, но не в CD8+
тельной поликлональной активации Т-клеток
Т-лимфоцитах периферической крови больных
(анти-CD3/CD28) [5].
COVID-19, причём вирусная нагрузка напря-
В марте 2022 г. была опубликована работа
мую зависела от тяжести заболевания [41].
Shen et al. [40], которая, расширив спектр ис-
Во второй работе, находящейся на этапе
пользуемых методов исследования, в целом
препринта, инфицирование мононуклеарных
подтвердила представленные выше результа-
клеток периферической крови здоровых до-
ты. Так, показано присутствие антигена SARS-
норов (n = 5) вирусом SARS-CoV-2 (MOI = 1)
CoV-2 (нуклеокапсидного белка, N-белка) в
in vitro выявило присутствие вирусных антиге-
Т-лимфоцитах (CD3+-клетках) перифериче-
нов как в CD4+-, так и в CD8+ Т-клетках (~13-
ской крови и постмортальных срезов лёгких
14%, проточной цитометрией с использованием
больных COVID-19 [40], причём для клеток пе-
гипериммунной сыворотки мышей, иммунизи-
риферической крови показан существенно бо-
рованных SARS-CoV-2) [42]. При этом процесс
лее высокий уровень N-белка в CD4+ Т-лимфо-
сопровождался выраженным апоптозом обеих
цитах по сравнению с таковым в CD8+-клетках
Т-клеточных субпопуляций: до 70% SARS-CoV-
БИОХИМИЯ том 87 вып. 6 2022
Т-ЛИМФОЦИТЫ КАК МИШЕНИ ДЛЯ SARS-CoV-2
785
2-инфицированных клеток имели апоптоти-
процесс, указывая на то, что AXL - не главный
ческие изменения [42]. Кроме того, у больных
рецептор входа вируса в Т-клетки данной ли-
COVID-19 (n = 22) показано присутствие ви-
нии, но может вносить вклад в инфицирова-
рус-инфицированных CD4+ Т-лимфоцитов в
ние [40].
периферической крови (иммунофлуоресцент-
Работа Davanzo et al. [41], продемонстри-
ным анализом с использованием для выявления
ровавшая участие в процессе инфицирова-
вирусных антигенов сыворотки выздоровевших
ния Т-лимфоцитов маркёра Т-хелперной
пациентов), и у основной части Т-клеток, несу-
субпопуляции, мембранной молекулы CD4,
щих вирусные антигены, выявлена двунитевая
заслуживает особого внимания: показана ко-
РНК SARS-CoV-2, маркёр репликации [42].
преципитация S-белка SARS-CoV-2 и реком-
А данные посмертного иммуногистохимиче-
бинантной полноразмерной молекулы CD4,
ского исследования показали присутствие ин-
а инфицирование CD4+ Т-лимфоцитов пери-
фицированных вирусом CD4+ Т-лимфоцитов в
ферической крови SARS-CoV-2 in vitro дозоза-
тканях лёгких больных COVID-19 [42].
висимо подавлялось в случае предобработки
Наряду с представленными выше работами,
клеток моноклональными антителами к CD4,
вирусная РНК регистрировалась в Т-лимфоци-
хотя и менее эффективно, чем на фоне блока-
тах (CD3+-клетках), присутствующих в образ-
ды традиционного для вируса рецептора ACE2
цах бронхоальвеолярного лаважа (n = 6) и мо-
и протеазы TMPRSS2. Интересно, что те же
кроты (n = 2) тяжёлых больных COVID-19 [43].
моноклональные антитела против CD4 способ-
А белки SARS-CoV-2 и вирионы выявлялись в
ны блокировать вход в CD4+ Т-клетки вируса
клетках селезёнки и лимфатических узлов при
иммунодефицита человека (Human immunode-
посмертном исследовании тканей больных
ficiency virus, HIV) [45], причём в случае HIV-
COVID-19 [44], хотя в последнем случае тип ин-
инфекции одного CD4 также недостаточно для
фицированных клеток не определялся.
входа вируса в клетку, он действует совместно
Рецепторы, опосредующие инфицирование
с корецепторами, в частности, хемокиновыми
Т-лимфоцитов SARS-CoV-2. Практически во
рецепторами CCR5 или CXCR4 [46].
всех представленных выше работах по инфи-
В итоге, на сегодняшний день имеется
цированию Т-лимфоцитов SARS-CoV-2 основ-
6 независимых работ, с разной степенью дета-
ной вирусный рецептор и корецептор ACE2/
лизации продемонстрировавших способность
TMPRSS2 либо не выявлялись на инфициро-
нового коронавируса SARS-CoV-2 инфициро-
ванных Т-клетках [5, 43], либо не участвовали
вать Т-лимфоциты человека. Как правило, речь
в инфицировании, что подтверждалось инги-
идёт о Т-клетках, инфильтрирующих поражён-
биторным анализом с помощью подавления
ные ткани больных COVID-19 [5, 40, 42, 43] или
экспрессии гена, кодирующего АСЕ2, и/или
о периферической крови пациентов [40-42],
функциональной блокады рецепторов
[40].
однако в четырёх работах инфицирование ви-
Единственное исключение - работа Davan-
русом [40-42] или псевдовирусом [5] in vitro по-
zo et al. [41], в которой инфицирование CD4+
казано и для Т-лимфоцитов периферической
Т-лимфоцитов периферической крови SARS-
крови здоровых доноров. Размер исследуемых
CoV-2 in vitro эффективно снижалось на фоне
групп невелик, но результаты хорошо согласу-
блокады ACE2 и TMPRSS2. В то же время пока-
ются друг с другом (таблица). Так, большин-
зано участие альтернативных рецепторов SARS-
ство авторов отмечает исключительное [41, 42]
CoV-2 в этом процессе. Так, в клетках-мишенях
или преимущественное [40] инфицирование
вируса выявлена экспрессия CD147, существен-
вирусом субпопуляции CD4+ Т-лимфоцитов, и
но возраставшая в ответ на активацию [5, 43],
работа Davanzo et al. [41] даёт объяснение это-
а инфицирование in vitro CD4+/CD8+ Т-лим-
му феномену, демонстрируя участие молекулы
фоцитов человека псевдовирусом SARS-CoV-2
CD4 в инфицировании клетки SARS-CoV-2.
отменялось моноклональными антителами к
Поражение вирусом обеих Т-клеточных субпо-
CD147 [5]. Другой альтернативный рецептор
пуляций (CD4+/CD8+ Т-клеток), показанное
SARS-CoV-2, тирозинкиназа AXL, выявлен
Pontelli et al. [42] в эксперименте in vitro, свя-
на уровне мРНК в вирус-инфицированных
зано, по-видимому, с избыточным количе-
Т-лимфоцитах больных COVID, а в Т-клетках
ством вирионов, вносимых в культуру в расчё-
линии Jurkat сверхэкспрессия AXL усиливала
те на клетку-мишень (MOI = 1,0), поскольку
(в 1,5 раза) инфицирование клеток вирусом,
в других работах этот показатель существен-
хотя нокдаун гена не оказывал влияния на этот
но ниже - MOI = 0,1 [41] или MOI = 0,01 [40].
7
БИОХИМИЯ том 87 вып. 6 2022
786
КУКЛИНА
Инфицирование Т-лимфоцитов* SARS-CoV-2
Рецептор/
Выявляемый
Тип Т-клеток
Локализация Т-клеток
Ссылки
корецептор
вирусный материал
Т-лимфоциты в тканях больных COVID-19
N-белок вируса
[40]
CD3+T-клетки
лёгкие
н/о
антигены вируса**
[42]
вирионы
[5]
бронхоальвеолярный
CD4+/CD8+
н/о
РНК вируса
[43]
лаваж, мокрота
РНК вируса
[41]
T-клетки
периферическая кровь
н/о
двунитевая РНК вируса
[42]
CD4+T-клетки
периферическая кровь
н/о
антигены вируса
[42]
Т-лимфоциты здоровых доноров, инфицированные in vitro
CD147***
РНК вируса
[5]
CD4+/CD8+
периферическая кровь
AXL?
РНК вируса
[40]
T-клетки
н/о
антигены вируса
[42]
АСЕ2/
РНК вируса
CD4+ T-клетки
периферическая кровь
TMPRSS2
антисмысловая РНК вируса
[41]
CD4
S-белок вируса, вирионы
Примечание. N-белок - нуклеокапсидный белок, S-белок - белок шипа вируса; н/о - рецепторы/корецепторы не
определялись.
* В таблице представлены данные только по инфицированию первичных Т-лимфоцитов, не Т-клеточных линий.
Информация о методах выявления вирусного материала и оценки вклада конкретных рецепторов в инфицирование
клетки, а также о размере исследуемых групп - в тексте.
** Вирусные антигены не идентифицировались, они выявлялись с помощью сывороток мышей, иммунизированных
SARS-CoV-2, или выздоровевших пациентов.
*** Представлены только рецепторы/корецепторы, для которых экспериментально подтверждено участие в инфициро-
вании Т-лимфоцитов SARS-CoV-2.
Т-клетки, инфицированные SARS-CoV-2
В инфицированных вирусом первичных
in vitro, подвергались выраженному апопто-
CD4+ Т-лимфоцитах и CD4+ Т-клеточных ли-
зу [40, 42], и в периферической крови больных
ниях Jurkat и MT4 различными способами реги-
COVID-19 уровень апоптотических Т-лим-
стрировали субгеномную вирусную РНК [40],
фоцитов также был повышен по сравнению
негативную (антисмысловую) РНК SARS-
с Т-клетками здоровых доноров [40]. Следует
CoV-2 [41] или двунитевую РНК вируса [42],
отметить, что апоптоз является закономерным
то есть маркёры репликации вируса в клетке,
результатом инфицирования клетки вирусом,
однако данных о сборке вирусных частиц и
он может быть как следствием прямого воз-
их выходе из заражённой клетки пока нет. Ни
действия вируса, так и классическим ответом
одна из имеющихся на сегодняшний день работ
иммунной системы на появление в организме
не позволяет однозначно сказать, является ли
клетки, несущей чужеродные антигены.
инфицирование вирусом Т-лимфоцитов про-
БИОХИМИЯ том 87 вып. 6 2022
Т-ЛИМФОЦИТЫ КАК МИШЕНИ ДЛЯ SARS-CoV-2
787
дуктивным или абортивным. В исследовании
ствующим белком SARS-CoV-1, а также с по-
Pontelli et al. [42] продуктивность инфекции
следовательностью трансмембранного домена
подтверждена для нефракционированного пула
α-цепи TCR, высокая вероятность такого меха-
мононуклеарных клеток периферической кро-
низма предполагается и для нового коронави-
ви, но для Т-лимфоцитов таких данных нет.
руса, хотя это ещё нужно показать [49].
Неинфекционные механизмы вирус-зависи-
мой регуляции Т-лимфоцитов. Говоря об инфи-
цировании Т-лимфоцитов SARS-CoV-2, следует
ИНФИЦИРОВАНИЕ Т-ЛИМФОЦИТОВ
учитывать, что даже в случае непродуктивного
SARS-CoV-2 И ПАТОГЕНЕЗ COVID-19
взаимодействия вируса с клеткой SARS-CoV-
2-зависимый сигнал может регулировать кле-
Новая коронавирусная инфекция сопро-
точную активность - об этом свидетельствуют
вождается существенными изменениями в
данные двух недавних исследований.
работе иммунной системы, и потенциаль-
Так, с помощью структурного компьютер-
ная способность SARS-CoV-2 инфицировать
ного моделирования показано, что S-белок
Т-лимфоциты может иметь прямое отноше-
SARS-CoV-2, но не других коронавирусов, со-
ние к целому ряду феноменов, ассоцииро-
держит структурные мотивы, имеющие вы-
ванных с болезнью. Первый и наиболее оче-
сокое сходство с таковыми у бактериальных
видный - лимфопения, которая выявляется у
суперантигенов (то есть антигенов, вызываю-
большинства больных COVID-19 [50, 51] и ас-
щих массовую неспецифическую активацию
социирована с тяжестью заболевания [50, 51].
Т-лимфоцитов), и способен напрямую связы-
Инфицирование Т-клеток вирусом и индукция
ваться с антигенным рецептором Т-лимфоци-
апоптоза в этих клетках вносит, по-видимому,
та (T cell receptor, TCR) [47]. Следствием этого
существенный вклад в развитие лимфопении
должна быть избыточная активация Т-клеточ-
при COVID-19, а возможно, и является его ос-
ного звена иммунной системы, и данный ме-
новной причиной.
ханизм может вносить вклад в гипервоспали-
Функциональное истощение Т-лимфоци-
тельный синдром, характерный для COVID-19.
тов - также часто регистрируемая ситуация у
В подтверждение этого у больных COVID-19 с
больных COVID-19 [52, 53]. Это вариант Т-кле-
гипервоспалением выявлен нетипичный (ис-
точной дисфункции, при котором кратковре-
кажённый) репертуар TCR, характерный для
менная гиперактивация Т-лимфоцита в ответ
активации суперантигеном, в отличие от па-
на антиген сменяется прогрессивным сниже-
циентов с лёгким или умеренным течением
нием пролиферативной активности, утратой
COVID-19 [47].
эффекторных функций, экспрессией ингиби-
Другой кандидат на роль регулятора Т-кле-
торных рецепторов, таких как PD-1 и CTLA,
точной активации - вирусный фузоген, или
эпигенетическим и транскрипционным репро-
белок, обеспечивающий слияние оболочки
граммированием [52, 53]. Частично за сниже-
вируса с мембраной клетки-мишени - вер-
ние функций Т-клеток при COVID-19 может
нее, не сам белок, а соответствующий домен
быть ответственна их непродуктивная инфек-
в составе S-белка SARS-CoV-2. Известно, что
ция SARS-CoV-2, а также неинфекционные
фузоген родственного коронавируса 2003 г.,
механизмы регуляции вирусом Т-лимфоцитов,
SARS-CoV-1, участвует не только в инфици-
связанные с непосредственным взаимодей-
ровании Т-клеток, но и в прямом ингибиро-
ствием вирусного S-белка с TCR и блокадой
вании TCR-зависимого сигнала: анализ его
или искажением антиген-зависимого сигнала.
первичной последовательности показал, что
Третий феномен, ассоциированный с но-
фузогенный домен S-белка SARS-CoV-1 ими-
вой коронавирусной инфекцией, - это не-
тирует трансмембранный домен α-цепи TCR и
контролируемая избыточная воспалительная
может нарушать взаимодействия между цепями
реакция (цитокиновый шторм), которая явля-
TCR-комплекса, препятствуя антиген-зависи-
ется одной из наиболее частых причин леталь-
мой сигнализации - его специфическая им-
ного исхода при COVID-19 [50, 51]. Как пра-
муносупрессивная активность подтверждена в
вило, гипервоспаление при данной патологии
эксперименте in vivo на модели коллаген-инду-
связывают с неадекватной активацией клеток
цированного артрита у мышей [48]. Учитывая
неспецифической защиты - основных проду-
близкое сходство аминокислотной последо-
центов провоспалительных факторов в орга-
вательности фузогена SARS-CoV-2 с соответ-
низме [50, 51], однако клетки адаптивного им-
БИОХИМИЯ том 87 вып. 6 2022
7*
788
КУКЛИНА
мунитета и, в частности Т-лимфоциты, должны
имеет повышенную экспрессию на мембране
играть непосредственную роль в этом процес-
Treg, причём маркирует активированные клет-
се. В первую очередь речь идёт о субпопуляции
ки (CD45R0+ Treg) с высокой супрессивной
регуляторных Т-клеток, Treg, которые в кон-
активностью [54]. Уровень CD147 на мембра-
тексте данной работы заслуживают отдельного
не Treg коррелирует с экспрессией главного
обсуждения.
транскрипционного фактора и маркёра данной
SARS-CoV-2 и регуляторные Т-лимфоциты.
субпопуляции, FoxP3 [54]. Более того, для под-
Т-хелперная субпопуляция Treg играет ключе-
держания стабильной экспрессии FoxP3 регу-
вую роль в поддержании аутотолерантности и
ляторным клеткам необходим сигнал, который
иммунном гомеостазе: естественные Treg (natu-
они получают как раз через CD147 при связы-
ral Treg, nTreg) созревают в тимусе, и их основ-
вании его c физиологическим лигандом CD98,
ной задачей является подавление иммунного
экспрессируемым неиммунным окружени-
ответа на аутоантигены, а индуцибельные Treg
ем [55]. Показано, что индуцибельные Treg с
(inducible Treg, iTreg) формируются на поздних
высокой экспрессией CD147 (CD147high iTreg)
стадиях любого иммунного ответа и призва-
эффективно подавляют воспалительный от-
ны ограничить избыточное воспаление, пред-
вет в модели экспериментальных колитов у
упреждая повреждение тканей продуктами ак-
гуманизированных мышей, в отличие от ана-
тивированных иммунных клеток.
логичных клеток с низкой экспрессией CD147
Целый ряд данных указывает на Treg, как
(CD147low iTregs) [55].
на вероятную мишень SARS-CoV-2. Так, мо-
Экспрессия двух других рецепторов SARS-
лекула CD147, альтернативный входной ре-
CoV-2, трансмембранного белка KREMEN1 и
цептор для вируса, опосредующий инфи-
рецепторной тирозинкиназы AXL, также выяв-
цирование первичных Т-лимфоцитов
[5], лена в субпопуляции Treg [9, 56], причём AXL
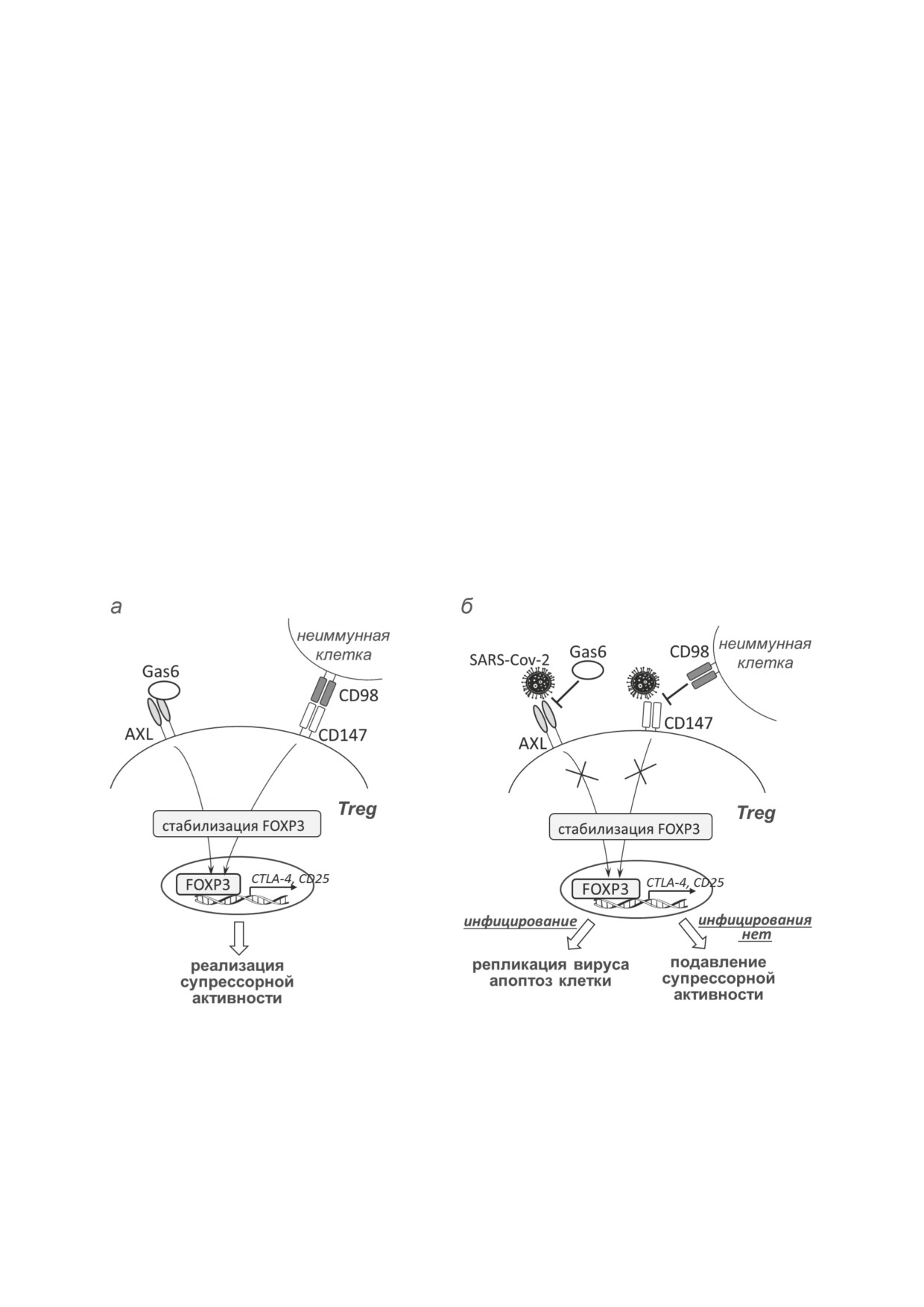
Потенциальные механизмы взаимодействия SARS-CoV-2 с регуляторными Т-лимфоцитами. Молекулы CD147 и AXL
имеют высокую экспрессию на мембране Treg и вовлечены в их функционирование: сигналы, которые клетки получа-
ют при связывании этих молекул с эндогенными лигандами CD98/Gas6, необходимы для поддержания стабильной экс-
прессии ключевого транскрипционного фактора FoxP3 (а). Одновременно CD147 и AXL служат входными рецепторами
SARS-CoV-2, причём встреча Treg с вирусом может приводить как к инфицированию клетки, так и к её неинфекционной
регуляции (б): в первом случае клетка-мишень, как правило, подвергается апоптозу, тогда как при отсутствии инфициро-
вания вирус, связываясь с CD147/AXL, может конкурентно ингибировать взаимодействие этих рецепторов с эндогенны-
ми лигандами (CD98/Gas6), препятствуя таким образом стабилизации клеток и реализации их супрессорной активности.
Подробные пояснения - в тексте статьи
БИОХИМИЯ том 87 вып. 6 2022
Т-ЛИМФОЦИТЫ КАК МИШЕНИ ДЛЯ SARS-CoV-2
789
играет в функционировании этих клеток,
CD147-зависимое инфицирование может регу-
по-видимому, такую же роль, как и CD147: её
лировать Т-клеточное звено иммунитета как на
эндогенный лиганд Gas6 (Growth arrest-specific)
этапе антигеннезависимой дифференцировки,
дозозависмо повышает in vivo и in vitro экс-
так и при ответе Т-лимфоцита на антиген.
прессию транскрипционного фактора FoxP3
в CD4+CD25+ Tregs и усиливает супрессор-
ную активность регуляторных Т-клеток в от-
ЗАКЛЮЧЕНИЕ
ношении эффекторных CD4+ T-лимфоцитов,
причём действие Gas6 in vitro отменяется пол-
Первые исследования взаимодействия
ностью или частично в случае нокдауна гена,
SARS-CoV-2 с Т-лимфоцитами показали, что
кодирующего AXL или функциональной блока-
вирус способен инфицировать Т-клетки и что
ды этого рецептора [56].
АСЕ2, главный функциональный рецептор ви-
Вопрос об инфицировании Treg корона-
руса в неиммунных тканях, либо не участвует в
вирусом пока нигде не поднимался, но этот
этом процессе [40], либо не выявляется на ви-
вариант весьма вероятен и требует проверки.
рус-позитивных Т-клетках [5, 43] - на первый
Кроме того, даже при отсутствии инфицирова-
план в инфицировании Т-лимфоцитов выхо-
ния регуляторных Т-клеток SARS-CoV-2 может
дят альтернативные рецепторы SARS-CoV-2, в
связываться с мембранными рецепторами этих
первую очередь молекулы CD147 и AXL [5, 40].
клеток CD147/AXL и конкурентно ингибиро-
Тем не менее вопрос об экспрессии АСЕ2 не
вать их взаимодействие с эндогенными лиган-
снимается с повестки: во-первых, несмотря
дами (CD98/Gas6), препятствуя таким образом
на убедительное подтверждение присутствия
стабилизации клеток и реализации их супрес-
в инфицированных Т-лимфоцитах вирусной
сорной активности (рисунок). Именно с пря-
РНК, вирусных белков и самих вирионов, уро-
мым действием вируса на регуляторные Т-клет-
вень инфицирования Т-клеток не сопоставим
ки может быть связано существенное снижение
с таковым для традиционных тканей-мишеней
количества Treg в циркуляции у пациентов с тя-
вируса
[40], имеющих стабильную экспрес-
жёлым течением COVID-19 [50, 57]. А посколь-
сию АСЕ2; во-вторых, тот факт, что АСЕ2 от-
ку субпопуляция Treg играет ключевую роль в
сутствует или слабо представлен на общей
ограничении избыточного иммунного ответа,
Т-клеточной популяции, не исключает экс-
снижение численности и/или активности этих
прессии рецептора отдельными субпопуля-
клеток должно вносить вклад в неконтролиру-
циями Т-лимфоцитов. В этом плане в первую
емую воспалительную реакцию (цитокиновый
очередь интерес представляют регуляторные
шторм), ассоциированную с тяжёлым течени-
Т-лимфоциты, провоспалительные Т-хелпер-
ем COVID-19, а возможно, и играть ключевую
ные субпопуляции, а также вирус-специфич-
роль в этом процессе. Кроме того, поражение
ные активированные CD4+/CD8+ Т-клетки,
Treg вирусом может иметь отношение к фе-
инфильтрирующие поражённые вирусом тка-
номену функционального истощения Т-лим-
ни. Не случайно инфицированные SARS-CoV-2
фоцитов, препятствующему эффективному
Т-клетки стабильно выявляются в тканях лёг-
противовирусному ответу [52]: именно с нару-
ких или в бронхоальвеолярном лаваже больных
шением контроля со стороны Treg может быть
COVID-19 - в очаге поражения под действием
связана гиперактивация эффекторных Т-лим-
микроокружения репертуар вирусных рецепто-
фоцитов, предшествующая их функционально-
ров и уровень их экспрессии могут меняться.
му истощению [53].
Кроме того, следует учитывать и возмож-
Наконец, говоря о молекуле CD147, как о
ность получения Т-лимфоцитами рецепто-
функциональном рецепторе для SARS-CoV-2,
ров SARS-CoV-2, в частности АСЕ2, от других
следует отметить, что она актуальна не только
клеток с помощью экстраклеточных микрове-
для Treg - данная молекула тесно связана с раз-
зикул, для которых данная молекула является
витием и функционированием Т-лимфоцитов
традиционным компонентом [36]. Вполне ве-
в целом. Так, CD147 высоко экспрессирован
роятно, что резидентные Т-клетки барьерных
на дубль-негативных тимоцитах и участвует в
органов или вирус-специфичные Т-лимфоци-
их экспансии [58]. На периферии CD147 регу-
ты, инфильтрирующие поражённые ткани, с
лирует миграцию активированных Т-лимфо-
помощью таких АСЕ2+-микровезикул получа-
цитов [59] и пролиферативный ответ на поли-
ют главный функциональный рецептор SARS-
клональную стимуляцию [60]. Как следствие,
CoV-2 от традиционных мишеней вируса с вы-
БИОХИМИЯ том 87 вып. 6 2022
790
КУКЛИНА
сокой экспрессией АСЕ2, таких как лёгочный
тельно, исходя из имеющихся на настоящий
эпителий или эндотелий сосудов, приобретая
момент данных, уровень инфицирования
таким образом чувствительность к вирусу. Это
Т-лимфоцитов новым коронавирусом невы-
в том случае, если микровезикулы секретиру-
сок, продуктивность инфекции пока не под-
ются неинфицированными клетками. Если же
тверждена
- возможно, вирус, проникая в
клетка, секретирующая АСЕ2+-микровезику-
Т-клетки, делает ставку в первую очередь на
лы, вирус-позитивна, что очень вероятно для
регуляцию активности этих клеток и/или на их
очага поражения, к рецепторам для вируса до-
транспортные функции.
бавляется и сам вирусный материал: показано,
что микровезикулы, формируемые SARS-CoV-
Финансирование. Исследование выполнено
2-инфицированными клетками, содержат РНК
в рамках государственного задания (тема госре-
вируса [61] и могут участвовать в распростране-
гистрации № АААА-А19-119112290007-7).
нии инфекции.
Конфликт интересов. Автор заявляет об от-
В этой связи важно заметить, что и сами
сутствии конфликта интересов.
Т-лимфоциты в случае инфицирования мо-
Соблюдение этических норм. Настоящая
гут участвовать в распространении вируса -
статья не содержит описания каких-либо ис-
Pontelli et al. [42] отводит Т-клетке в данной
следований с участием людей или животных
ситуации роль «Троянского коня». Действи-
в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1. Zhou, P., Yang, X. L., Wang, X. G., Hu, B.,
epithelial cells, Cell Res., 31, 126-140, doi: 10.1038/
Zhang, L., et al.
(2020) A pneumonia outbreak
s41422-020-00460-y.
associated with a new coronavirus of probable bat
8. Gu, Y., Cao, J., Zhang, X., Gao, H., Wang, H.,
origin, Nature, 579, 270-273, doi: 10.1038/s41586-020-
et al. (2020) Interaction network of SARS-CoV-2
2012-7.
with host receptome through spike protein, BioRxiv,
2. Hoffmann, M., Kleine-Weber, H., Schroeder, S.,
doi: 10.1101/2020.09.09.287508.
Krüger, N., Herrler, T., et al. (2020) SARS-CoV-2 cell
9. Grigoriou, M., Banos, A., Hatzioannou, A.,
entry depends on ACE2 and TMPRSS2 and is blocked
Kloetgen, A., Kouzis, P., et al. (2021) Regulatory
by a clinically proven protease inhibitor, Cell, 181, 271-
T cell transcriptomic reprogramming characterizes
280 e8, doi: 10.1016/j.cell.2020.02.052.
adverse events by checkpoint inhibitors in solid
3. Hamming, I., Timens, W., Bulthuis, M. L., Lely,
tumors, Cancer Immunol. Res.,
9,
726-734,
A. T., Navis, G., et al. (2004) Tissue distribution
doi: 10.1158/2326-6066.CIR-20-0969.
of ACE2 protein, the functional receptor for SARS
10. Kuba, K., Imai, Y., Rao, S., Gao, H., Guo, F.,
coronavirus. A first step in understanding SARS
et al. (2005) A crucial role of angiotensin converting
pathogenesis, J. Pathol., 203, 631-637, doi: 10.1002/
enzyme 2 (ACE2) in SARS coronavirus-induced lung
path.1570.
injury, Nat. Med., 11, 875-879, doi: 10.1038/nm1267.
4. Radzikowska, U., Ding, M., Tan, G., Zhakparov, D.,
11. Hoffmann, M. A., Kleine-Weber, H., and
Peng, Y., et al. (2020) Distribution of ACE2, CD147,
Pöhlmann, S.
(2020) Multibasic cleavage site in
CD26 and other SARS-CoV-2 associated molecules
the spike protein of SARS-CoV-2 is essential for
in tissues and immune cells in health and in asthma,
infection of human lung cells, Mol. Cell, 78, 779-784,
COPD, obesity, hypertension, and COVID-19 risk
doi: 10.1016/j.molcel.2020.04.022.
factors, Allergy, 75, 2829-2845, doi: 10.1111/all.14429.
12. Shulla, A., Heald-Sargent, T., Subramanya, G.,
5. Wang, K., Chen, W., Zhang, Z., Deng, Y., Lian, J.-Q.,
Zhao, J., Perlman, S., et al. (2011) A transmembrane
et al. (2020) CD147-spike protein is a novel route for
serine protease is linked to the severe acute respiratory
SARS-CoV-2 infection to host cells, Signal. Transduct.
syndrome coronavirus receptor and activates
Target Ther., 5, 283, doi: 10.1038/s41392-020-00426-x.
virus entry, J. Virol.,
85,
873-882, doi:
10.1128/
6. Schmid, E. T., Pang, I. K., Silva, E. A. C., Bosurgi, L.,
JVI.02062-10.
Miner, J. J., et al. (2016) AXL receptor tyrosine kinase
13. Iwata-Yoshikawa, N., Okamura, T., Shimizu, Y.,
is required for T cell priming and antiviral immunity,
Hasegawa, H., Takeda, M., et al. (2019) TMPRSS2
eLife, 5, e12414, doi: 10.7554/eLife.12414.
contributes to virus spread and immunopathology
7. Wang, S., Qiu, Z., Hou, Y., Deng, X., Xu, W., et al.
in the airways of murine models after coronavirus
(2021) AXL is a candidate receptor for SARS-CoV-2
infection, J. Virol.,
93, e01815-18, doi:
10.1128/
that promotes infection of pulmonary and bronchial
JVI.01815-18.
БИОХИМИЯ том 87 вып. 6 2022
Т-ЛИМФОЦИТЫ КАК МИШЕНИ ДЛЯ SARS-CoV-2
791
14.
Zhang, M. Y., Zhang, Y., Wu, X.-D., Zhang, K.,
ASGR1 as alternative functional receptors of SARS-
Lin, P, e al.
(2018) Disrupting CD147-RAP2
CoV-2, Cell Res., 32, 24-37, doi: 10.1038/s41422-021-
interaction abrogates erythrocyte invasion by
00595-6.
Plasmodium falciparum, Blood,
10,
1111-1121,
27.
Cantuti-Castelvetri, L., Ojha, R., Pedro L. D.,
doi: 10.1182/blood-2017-08-802918.
Djannatian, M., Franz, J., et al. (2020) Neuropilin-1
15.
Pushkarsky, T., Zybarth, G., Dubrovsky, L.,
facilitates SARS-CoV-2 cell entry and infectivity,
Yurchenko, V., Tang, H., et al.
(2001) CD147
Science, 370, 856-860, doi: 10.1126/science.abd2985.
facilitates HIV-1 infection by interacting with virus-
28.
Seyran, M., Takayama, K., Uversky, V. N.,
associated cyclophilin A, Proc. Natl. Acad. Sci. USA,
Lundstrom, K., Palù, G., et al. (2020) The structural
11, 6360-6365, doi: 10.1073/pnas.111583198.
basis of accelerated host cell entry by SARS-CoV-2
16.
Bernard, S. C. Simpson, N., Join-Lambert, O.,
dagger, FEBS J., doi: 10.1111/febs.15651.
Federici, C., Laran-Chich, M.-P., et al.
(2014)
29.
Clausen, T. M., Sandoval, D. R., Spliid, C. B., Pihl,
Pathogenic Neisseria meningitidis utilizes CD147
J., Perrett, H. R., et al. (2020) SARS-CoV-2 infection
for vascular colonization, Nat. Med.,
7,
725-731,
depends on cellular heparan sulfate and ACE2, Cell,
doi: 10.1038/nm.3563.
183, 1043-1057, doi: 10.1016/j.cell.2020.09.033.
17.
Chen, Z. Mi, L., Xu, J., Yu, J., Wang, X., et al. (2005)
30.
Thépaut, M., Luczkowiak, J., Vivès, C., Labiod, N.,
Function of HAb18G/CD147 in invasion of host cells
Bally, I., et al. (2021) DC/L-SIGN recognition of
by severe acute respiratory syndrome coronavirus,
spike glycoprotein promotes SARS-CoV-2 trans-
J. Infect. Dis., 5, 755-760, doi: 10.1086/427811.
infection and can be inhibited by a glycomimetic
18.
Shilts, J., Crozier, T. W. M., Greenwood, E. J. D.,
antagonist, PLoS Pathog., 17, e1009576, doi: 10.1371/
Lehner, P. J., and Wright, G. J. (2021) No evidence for
journal.ppat.1009576.
basigin/CD147 as a direct SARS-CoV-2 spike binding
31.
Yan, S., and Wu, G. (2020) Is lymphopenia different
receptor, Sci. Rep., 11, 413, doi: 10.1038/s41598-020-
between SARS and COVID-19 patients? FASEB J., 35,
80464-1.
e21245, doi: 10.1096/fj.202002512.
19.
Ragotte, R. J., Pulido, D., Donnellan, F. R., Hill,
32.
Bertram, S., Lavender, A. H. H., Gierer, S.,
M. L., Gorini, G., et al. (2021) Human basigin
Danisch, S., Perin, P., et al. (2012) Influenza and
(CD147) does not directly interact with SARS-Cov-2
SARS-coronavirus activating proteases TMPRSS2
spike glycoprotein, mSphere, 6, e0064721, doi: 10.1128/
and HAT are expressed at multiple sites in human
mSphere.00647-21.
respiratory and gastrointestinal tracts, PLoS One, 7,
20.
Fenizia, C., Galbiati, S., Vanetti, C., Vago, R.,
e35876, doi: 10.1371/journal.pone.0035876.
Clerici, M., et al. (2021) SARS-CoV-2 entry: at
33.
Lukassen, S., Chua, R., Trefzer, T., Kahn, N. C.,
the crossroads of CD147 and ACE2, Cells,
10,
Schneider, M. A., et al. (2020) SARS-CoV-2 receptor
doi: 10.3390/cells10061434.
ACE2 and TMPRSS2are primarily expressed in
21.
Goruppi, S., Ruaro, E., and Schneider, C. (1996)
bronchial transient secretory cells, EMBO J., 39,
Gas6, the ligand of Axl tyrosine kinase receptor, has
e105114, doi: 10.15252/embj.20105114.
mitogenic and survival activities for serum starved
34.
Ziegler, C. G. K., Allon, S. J., Nyquist, S. K., Mbano,
NIH3T3 fibroblasts, Oncogene, 12, 471-480.
I. M., Miao, V. N., et al. (2020) SARS-CoV-2 receptor
22.
Stitt, T. N., Conn, G., Gore, M., Lai, C., Bruno, J.,
ACE2 is an interferon-stimulated gene in human
et al. (1995) The anticoagulation factor protein S
airway epithelial cells and is enriched in specific
and its relative, Gas6, are ligands for the Tyro 3/Axl
cell subsets across tissues, Cell, 181, 1016-1035.e19,
family of receptor tyrosine kinases, Cell, 80, 661-670,
doi: 10.1016/j.cell.2020.04.035.
doi: 10.1016/0092-8674(95)90520-0.
35.
Gurunathan, S., Kang, M. H., and Kim, J.-H.
23.
Seidah, N. G., Chretien, M., and Mbikay, M. (2018)
(2021) Diverse effects of exosomes on COVID-19:
The ever-expanding saga of the proprotein convertases
a perspective of progress from transmission to
and their roles in body homeostasis: emphasis on
therapeutic developments, Front. Immunol.,
12,
novel proprotein convertase subtilisin kexin number 9
716407, doi: 10.3389/fimmu.2021.716407.
functions and regulation, Curr. Opin. Lipidol., 29, 144-
36.
Wang, J., Chen, S., and Bihl, J. (2020) Exosome-
150, doi: 10.1097/MOL.0000000000000484.
mediated transfer of ACE2 (angiotensin-converting
24.
Mao, B., Wu, W., Davidson, G., Marhold, J., Li, M.,
enzyme 2) from endothelial progenitor cells promotes
et al. (2002) Kremen proteins are Dickkopf receptors
survival and function of endothelial cell, Oxid. Med.
that regulate Wnt/beta-catenin signaling, Nature, 417,
Cell Longev., 4213541, doi: 10.1155/2020/4213541.
664-667, doi: 10.1038/nature756.
37.
El-Shennawy, L., Hoffmann, A. D., Dashzeveg,
25.
Staring, J., van den Hengel, L. G., Raaben, M.,
N. K., Mehl, P. J., Yu, Z., et al. (2020) Circulating
Blomen, V. A., Carette, J. I., et al. (2018) KREMEN1
ACE2-expressing exosomes block SARS-CoV-2
is a host entry receptor for a major group of
infection as an innate antiviral mechanism, bioRxiv,
enteroviruses, Cell Host Microbe, 23, 636-643.e635,
doi: 10.1101/2020.12.03.407031.
doi: 10.1016/j.chom.2018.03.019.
38.
Mobini, S., Chizari, M., Mafakher, L., Rismani, E.,
26.
Gu, Y., Cao, J., Zhang, X., Gao, H., Wang, Y., et al.
and Rismani, E.
(2021) Structure-based study
(2022) Receptome profiling identifies KREMEN1 and
of immune receptors as eligible binding targets
БИОХИМИЯ том 87 вып. 6 2022
792
КУКЛИНА
of
coronavirus SARS-CoV-2 spike protein,
CoV-2 pneumonia in Wuhan, China: a single-
J. Mol. Graph. Model., 108, 107997, doi: 10.1016/
centered, retrospective, observational study, Lancet
j.jmgm.2021.107997.
Respir. Med., doi: 10.1016/S2213-2600(20)30079-5.
39.
Gu, J., Gong, E., Zhang, B., Zheng, J., Gao, Z., et al.
52.
Diao, B., Wang, C., Tan, Y., Chen, X., Liu, Y.,
(2005) Multiple organ infection and the pathogenesis
et al. (2020) Reduction and functional exhaustion
of SARS, J. Exp. Med., 202, 415-424, doi: 10.1084/
of T cells in patients with Coronavirus Disease 2019
jem.20050828.
(COVID-19), Front. Immunol., 11, 1-7, doi: 10.3389/
40.
Shen, X.-R., Geng, R., Li, Q., Chen, Y., Li, S.-
fimmu.2020.00827.
F., et al.
(2022) ACE2-independent infection of
53.
Kim, C. G., Kim, G., Kim, K. H., Park, S.,
T lymphocytes by SARS-CoV-2, Signal. Transduct.
Shin, S., et al. (2021) Distinct exhaustion features
Target Ther., 7, 83, doi: 10.1038/s41392-022-00919-x.
of T lymphocytes shape the tumor-immune
41.
Davanzo, G. G., Codo, A. C., Brunetti, N. S.,
microenvironment with therapeutic implication
Boldrini, V., Knittel, T. L., et al. (2020) SARS-CoV-2
in patients with non-small-cell lung cancer,
uses CD4 to infect T helper lymphocytes, MedRxiv,
J. Immunother. Cancer, 9, e002780, doi: 10.1136/
doi: 10.1101/2020.09.25.20200329.
jitc-2021-002780.
42.
Pontelli, M. C., Castro, I. A., Martins, R. B., Veras,
54.
Solstad, T., Bains, S. J., Landskron, J., Aandahl,
F. P., La Serra, L., et al. (2020) Infection of human
E. M., Thiede, B., et al. (2011) CD147 (Basigin/
lymphomononuclear cells by SARS-CoV-2, bioRxiv,
Emmprin) identifies FoxP3+CD45RO+CTLA4+-
doi: 10.1101/2020.07.28.225912.
activated human regulatory T cells, Blood,
118,
43.
Ren, X., Wen, W., Fan, X., Hou, W., Su, B., et al.
5141-5151.
(2021) COVID-19 immune features revealed by a
55.
Geng, J., Chen, R., Yang, F.-F., Lin, P., Zhu, Y.-M.,
large-scale single-cell transcriptome atlas, Cell, 184,
et al. (2021) CD98-induced CD147 signaling stabilizes
1895-1913.e19, doi: 10.1016/j.cell.2021.01.053.
the Foxp3 protein to maintain tissue homeostasis,
44.
Bian, X.W., COVID-19 Pathology Team
(2020)
Cell. Mol. Immunol., 18, 2618-2631, doi: 10.1038/
Autopsy of COVID-19 victims in China, Natl. Sci.
s41423-021-00785-7.
Rev., 7, 1414-1418, doi: 10.1093/nsr/nwaa123.
56.
Zhao, G.-J., Zheng, J.-Y., Bian, J.-L., Chen, L.-
45.
Shaik, M., Peng, H., Lu, J., Rits-Volloch, S., Xu, C.,
W., Dong, N, et al.
(2017) Growth arrest-
et al. (2019) Structural basis of coreceptor recognition
specific
6 enhances the suppressive function of
by HIV-1 envelope spike, Nature,
565,
318-323,
CD4+CD25+ regulatory T cells mainly through
doi: 10.1038/s41586-018-0804-9.
Axl receptor, Mediators Inflamm.,
6848430,
46.
Iliopoulou, M., Nolan, R., Alvarez, L., Watanabe, Y.,
doi: 10.1155/2017/6848430.
Coomer, C. A., et al. (2018) A dynamic three-step
57.
Qin, C., Zhou, L., Hu, Z., Zhang, S., Yang, S., et al.
mechanism drives the HIV-1 pre-fusion reaction, Nat.
(2020) Dysregulation of immune response in patients
Struct. Mol. Biol., 25, 814-822, doi: 10.1038/s41594-
with COVID-19 in Wuhan, China, Clin. Infect. Dis.,
018-0113-x.
71, 762-768, doi: 10.1093/cid/ciaa248.
47.
Cheng, M. H., Zhang, S., Porritt, R. A., Rivas,
58.
Renno, T., Wilson, A., Dunkel, C., Coste, I.,
M. N., Paschold, L., et al. (2020) Superantigenic
Maisnier-Patin, K., et al. (2002) A role for CD147 in
character of an insert unique to SARS-CoV-2 spike
thymic development, J. Immunol., 168, 4946-4950,
supported by skewed TCR repertoire in patients with
doi: 10.4049/jimmunol.168.10.4946.
hyperinflammation, Proc. Natl Acad. Sci. USA, 117,
59.
Damsker, J. M., Bukrinsky, M. I., and Constant, S. L.
25254-25262, doi: 10.1073/pnas.2010722117.
(2007) Preferential chemotaxis of activated human
48.
Shen, Z. T., and Sigalov, A. B.
(2016) SARS
CD4+ T cells by extracellular cyclophilin A, J. Leukoc.
coronavirus
fusion
peptide-derived
sequence
Biol., 82, 613-618, doi: 10.1189/jlb.0506317.
suppresses collagen-induced arthritis in DBA/1J
60.
Koch, C., Staffler, G., Huttinger, R., Hilgert, I.,
Mice, Sci. Rep., 6, 28672, doi: 10.1038/srep28672.
Prager, E., et al. (1999) T cell activation-associated
49.
Sigalov, A. B.
(2022) SARS-CoV-2 may affect
epitopes of CD147 in regulation of the T cell response,
the immune response via direct inhibition of
and their definition by antibody affinity and antigen
T cell receptor: mechanistic hypothesis and
density, Int. Immunol., 11, 777-786, doi: 10.1093/
rationale, Biochimie,
195,
86-89, doi:
10.1016/
intimm/11.5.777.
j.biochi.2021.11.005.
61.
Kwon, Y., Nukala, S. B., Srivastava, S.,
50.
Wang, F., Hou, H., Luo, Y., Tang, G., Wu, S., et al.
Miyamoto, H., Ismail, N. I., et al.
(2020)
(2020) The laboratory tests and host immunity of
Detection of viral RNA fragments in
human
COVID-19 patients with different severity of illness,
iPSC-cardiomyocytes following treatment with
JCI Insight, 5, doi: 10.1172/jci.insight.137799.
extracellular vesicles from SARS-CoV-2 coding-
51.
Yang, X., Yu, Y., and Xu, J. (2020) Clinical course
sequence-overexpressing lung epithelial cells, bioRxiv,
and outcomes of critically ill patients with SARS-
doi: 10.1101/2020.05.14.093583.
БИОХИМИЯ том 87 вып. 6 2022
Т-ЛИМФОЦИТЫ КАК МИШЕНИ ДЛЯ SARS-CoV-2
793
T LYMPHOCYTES AS TARGETS FOR SARS-CoV-2
Review
E. M. Kuklina
Perm Federal Research Center, Institute of Ecology and Genetics of Microorganisms,
Ural Branch of the Russian Academy of Sciences,
614081 Perm, Russia; e-mail: ibis_07@iegm.ru
Despite numerous data on the absence or weak expression of the main functional receptor of SARS-CoV-2
angiotensin I-converting enzyme 2 (ACE2) by T cells, recent data demonstrate the ability of a new coronavirus
to effectively infect T lymphocytes. The review is devoted to the analysis of these works: it considers alterna-
tive (ACE2-independent) pathways of cell infection, identifies T cell subpopulations that serve as the most
likely targets of SARS-CoV-2, and discusses the format of virus-cell interaction, including both infectious and
non-infectious mechanisms of T lymphocyte regulation, and also evaluates the role of virus-dependent damage
of T lymphocytes in the pathogenesis of COVID-19. Particular attention is paid to regulatory T cells as potential
targets of SARS-CoV-2, as well as the possible involvement of exosomes in the regulation of sensitivity to the
virus of T lymphocytes present in peripheral tissues.
Keywords: SARS-CoV-2, T lymphocytes, ACE2, CD147, Treg
БИОХИМИЯ том 87 вып. 6 2022