БИОХИМИЯ, 2022, том 87, вып. 9, с. 1163 - 1181
УДК 577.2
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ
НЕЙРОПРОТЕКТОРНОГО ЭФФЕКТА МЕТИЛЕНОВОГО СИНЕГО
Обзор
© 2022 А.П. Гуреев1,2*, И.С. Садовникова1, В.Н. Попов1,2
1 Воронежский государственный университет,
394018 Воронеж, Россия; электронная почта: gureev@bio.vsu.ru
2 Воронежский государственный университет инженерных технологий, 394036 Воронеж, Россия
Поступила в редакцию 03.05.2022
После доработки 07.07.2022
Принята к публикации 15.07.2022
Метиленовый синий (МС) является первым полностью синтетическим веществом, которое нашло
своё применение в медицине более 120 лет назад для лечения малярии. Сейчас МС одобрен для ле-
чения метгемоглобинемии, но есть предпосылки для его репозиционирования в сторону нейропро-
текции. Перспективность данного соединения подтверждается тем, что на различных моделях была
показана эффективность при терапии болезни Альцгеймера, Паркинсона, Хантингтона, моделях
черепно-мозговых травм, бокового амниотического склероза, депрессивных расстройствах и т.д.
Но задача данного обзора - не столько сконцентрироваться на описании терапевтических эффек-
тов МС при лечении всевозможных нейродегенераций, сколько углубиться в механизмы действия
сигнальных путей, на которые данный препарат оказывает прямое или косвенное влияние. МС мо-
жет функционировать как альтернативный переносчик электронов в дыхательной цепи митохондрий
при нарушении работы цепи переноса электронов. Он может оказывать противовоспалительный
и антиапоптотический эффекты, ингибировать моноаминооксидазы и NO-синтазы, активировать
сигнальные пути, которые обеспечивают обновление митохондриального пула - митохондриаль-
ный биогенез и аутофагию, препятствовать агрегации белков с неправильной укладкой. Понимание
и комплексное изучение всех аспектов прямого и косвенного действия МС, а не только какого-либо
одного эффекта может помочь в его дальнейшем исследовании, в том числе и на клиническом уровне.
КЛЮЧЕВЫЕ СЛОВА: метиленовый синий, нейродегенерация, болезнь Альцгеймера, тау-белок, альтер-
нативный транспорт электронов, Nrf2/ARE сигнальный путь, апоптоз, аутофагия, воспаление, моноамин-
оксидаза, NO-синтаза.
DOI: 10.31857/S0320972522090019, EDN: AZNPIL
ВВЕДЕНИЕ
шо растворим в воде и хлороформе, умеренно
растворим в спирте. Максимумы поглощения
Метиленовый синий (МС; 3,7-бисдиметил-
МС составляют 609 и 668 нм, молекулярная
аминофенотиазин хлорид) представляет собой
масса — 319,852 г/моль [1]. МС не является
темно-зелёные кристаллы или кристалли-
природным веществом и впервые был синте-
ческий порошок с бронзовым блеском, в воде
зирован в 1876 г. химиком Генрихом Каро. Год
образует темно-синий раствор, который обыч-
спустя МС стал первым красителем, запатен-
но используется в качестве красителя. Он хоро-
тованным в Германии [2]. В настоящее время
Принятые сокращения: АФК - активные формы кислорода; БАС - боковой амиотрофический склероз; МАО - мо-
ноаминоксидаза; МС - метиленовый синий; ЭТЦ - электрон-транспортная цепь; AMPK - AMP-активируемая про-
теинкиназа; ARE - антиоксидант-респонсивный элемент; cGMP - циклический гуанозинмонофосфат; CRH - кор-
тикотропин-рилизинг-гормон; GSK3β - киназа гликогенсинтазы 3β; HTT - хантингтин; Keap1 - kelch-подобный
ECH-ассоциированный белок 1; mTORC1 - мишень рапамицина млекопитающих, комплекс 1; NF-κB - ядерный
фактор каппа-B; NOS - NO-синтаза; Nrf2 - ядерный фактор, связанный с эритроидным фактором 2; PGC-1α - гам-
ма-коактиватор 1-альфа-рецептора, активируемого пролифераторами пероксисом; PI3K - фосфоинозитид-3-киназа;
PMCA - Ca2+-ATPаза плазматической мембраны; sGC - растворимая гуанилатциклаза; SOD1 - супероксиддисмутаза 1;
STAT - преобразователь сигнала и активатор транскрипции; TNF-α - фактор некроза опухолей альфа.
* Адресат для корреспонденции.
1163
1164
ГУРЕЕВ и др.
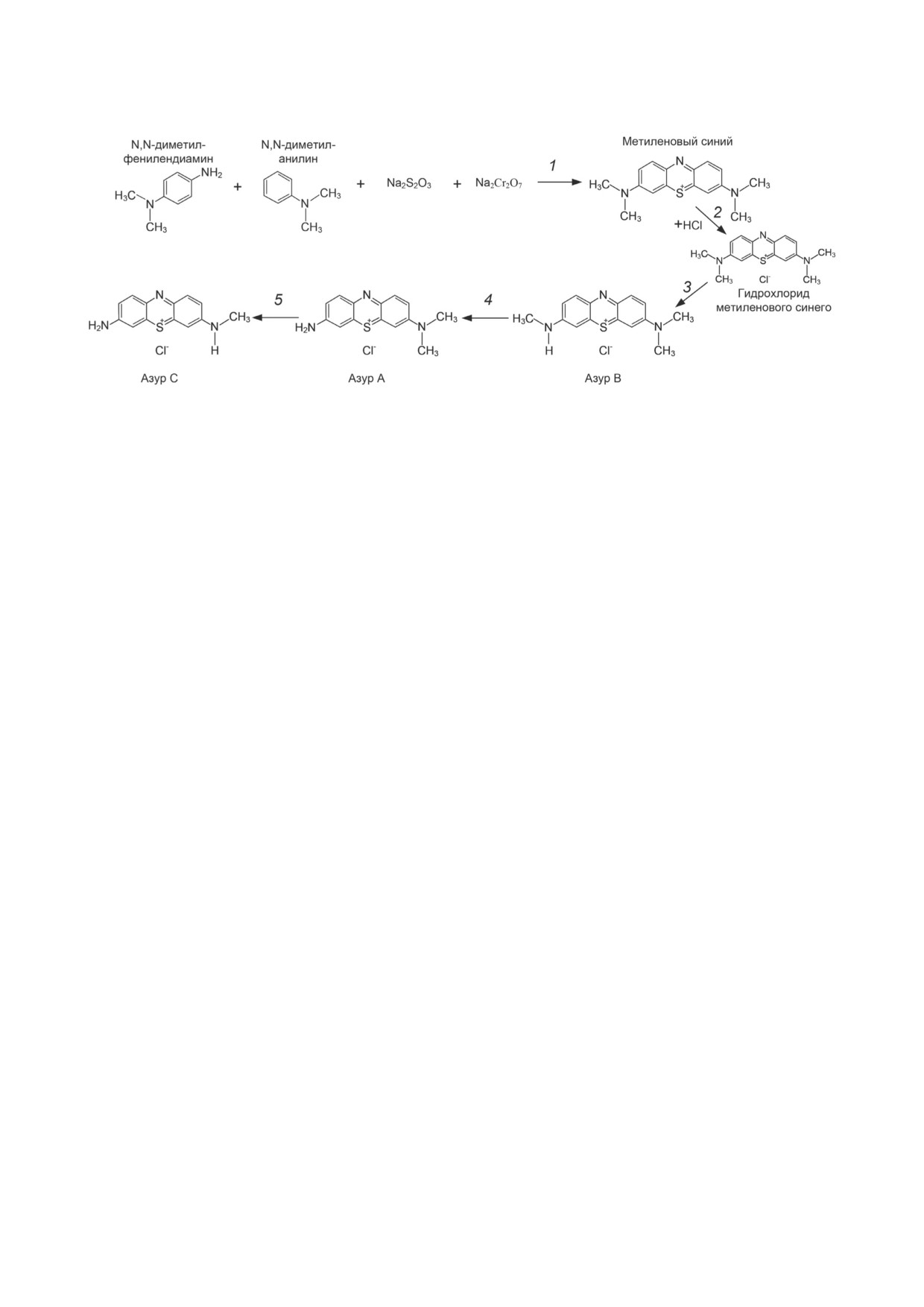
Рис. 1. Синтез и метаболизм MС. 1 - синтез МС. 2 - гидрохлорирование МС. 3-5 - деметилирование MС и последова-
тельное формирование азуров A, B и C
МС синтезируют окислением N,N-диметилфе-
хиатрии, как биологическая психиатрия [9].
нилендиамина дихроматом натрия (Na2Cr2O7)
В это же время МС продолжал использоваться
в присутствии тиосульфата натрия (Na2S2O3)
в медицине, но только в качестве красителя для
и дальнейшим окислением в присутствии
других нейролептиков, что помогало контро-
N,N-диметиланилина [3] (рис. 1, реакция 1).
лировать приём лекарств у пациентов психи-
Гидрохлорид МС получают за счёт добавле-
атрических клиник за счёт окрашивания мочи
ния к раствору красителя 30%-ной соляной
в синий цвет [6]. Единичные исследования про-
кислоты и насыщенного раствора хлорида нат-
водились во второй половине 1930-х гг. Было
рия; после фильтрации продукт промывают
показано, что MС приносит пользу пациентам,
2%-ным раствором хлорида натрия (рис. 1, ре-
страдающим кататонической деменцией [10]
акция 2). Резонансные структуры МС обычно
и снижает смертность при эпилептических
изображают с формальным зарядом, располо-
припадках [11].
женным на атоме серы (рис. 1) или на одном из
Интерес к МС как к нейролептику возро-
атомов азота (не показано) [4]. Кроме того, MС
дился в 70-80-х гг. XX века, когда появились
может быть представлен в виде трёх наиболее
данные, что повышенный уровень ванадия мо-
распространённых N-деметилированных ана-
жет иметь этиологическое значение при маниа-
логов: азура A, азура B и азура C [5] (рис. 1, ре-
кально-депрессивном расстройстве [12]. Было
акции 3-5).
показано, что MС катализирует восстановление
Раствор MС имеет синий цвет в окисли-
ванадата до ванадила, что значительно подав-
тельной среде, но превращается в лейко-MС
ляло депрессивное поведение [13]. Ещё раньше
(бесцветную форму) при воздействии восста-
появились первые данные, демонстрирующие,
новителя [3]. Есть много применений МС в
что МС оказывает влияние на процессы запо-
аналитической химии или в быту, но в большей
минания и сохранения памяти у крыс [14]. В се-
степени этот краситель прославился как пер-
редине 90-х гг. было показано, что МС инги-
вый полностью синтетический препарат, при-
бирует агрегацию тау-белков, а, следовательно,
меняемый в медицине [6].
может иметь клиническое применение для ле-
Наиболее ранние упоминания о влиянии
чения болезни Альцгеймера [15].
МС на психологическое состояние человека от-
Позже на основе МС было разработано
носятся к концу XIX века. Тогда были получены
несколько поколений препаратов, с которыми
данные об обезболивающих эффектах МС [7] и
были проведены клинические исследования,
его седативном эффекте при различных формах
но полученные результаты оказались весьма
психозов [8]. МС, а точнее его фенотиазиновое
противоречивыми. Во II-й фазе клинических
ядро, стал основой для синтеза ряда нейро-
испытаний был показан положительный эф-
лептических препаратов в 1950-х гг., в том числе
фект препарата на основе МС у пациентов с
хлорпромазина и флуфеназина, которые явля-
лёгкой и средней степенью тяжести болезни
ются незаменимыми препаратами для лечения
Альцгеймера (NCT005153330) [16]. Но на этапе
психотических расстройств. Некоторые ис-
фазы III клинических исследований у экспе-
следователи считают, что появление фенотиа-
риментальной группы, получавшей таблетки,
зинов является началом такого раздела в пси-
содержащие 75 и 125 мг МС, не было выявле-
БИОХИМИЯ том 87 вып. 9 2022
НЕЙРОПРОТЕКТОРНЫЙ ЭФФЕКТ МЕТИЛЕНОВОГО СИНЕГО
1165
но улучшений по сравнению с группой пла-
ся основной причиной митохондриальных
цебо (NCT01689246) [17]. Примечательно, что
дефектов [30]. Митохондрии имеют несколь-
группа плацебо получала тот же препарат, но в
ко сайтов, которые ответственны за продук-
концентрации 4 мг, чтобы цвет мочи у группы
цию АФК, и большинство из них расположены
плацебо не отличался от цвета мочи пациентов
на внутренней митохондриальной мембране.
из экспериментальной группы. Позже было
Комплекс I является основным местом про-
показано, что даже в концентрации 4 мг/день
дукции митохондриальных АФК. Кроме того,
МС оказывает тот же эффект, что и в высоких
ингибирование комплекса I приводит к де-
концентрациях. Поэтому некоторые исследо-
фициту ATP и гипервосстановлению многих
ватели причиной неудачи фазы III клиниче-
NAD+-зависимых ферментов [31]. Митохон-
ских испытаний называют некорректный ди-
дрии растений в процессе эволюции приобрели
зайн исследования [18]. В связи с этим в 2018 г.
альтернативные пути дыхания, которые вклю-
было запущено ещё одно клиническое испыта-
чают ротенон-нечувствительные NAD(P)H-де-
ние, где препарат будет использоваться в кон-
гидрогеназы и альтернативную оксидазу. Они
центрации 16 и 8 мг/день, а в качестве плаце-
позволяют за счёт рассеивания мембранного
бо будут использованы другие препараты для
потенциала снизить продукцию АФК и пред-
придания моче характерного цвета. Полное
отвратить перевосстановление компонентов
окончание клинических испытаний ожидается
катаболических путей [32]. У митохондрий жи-
в июне 2023 г. (NCT03446001).
вотных тоже есть механизм рассеивания мем-
В последнее десятилетие проведено много
бранного потенциала - мягкое разобщение,
исследований терапевтического эффекта МС
которое осуществляется белками-разобщите-
при болезни Хантингтона [19, 20], болезни
лями (uncoupling proteins - UCPs), вызываю-
Паркинсона [21, 22], черепно-мозговых трав-
щими утечку протонов, снижение мембранно-
мах [23-25], ишемических повреждений голов-
го потенциала и сокращение уровня продукции
ного мозга [26, 27], травмах спинного мозга [28],
АФК [33]. Имеется целый ряд фармацевтиче-
боковом амиотрофическом склерозе [29] и т.д.
ских разобщителей, среди которых наиболь-
Высока вероятность, что в скором времени
ший интерес представляет 2,4-динитрофенол
начнутся клинические испытания МС для ле-
(2,4-ДНФ), действующий как протонофор.
чения других нейродегенеративных и психи-
Вначале
2,4-ДНФ широко использовался в
ческих заболеваний. Цель данного обзора - не
качестве метаболического препарата для сни-
столько суммировать данные о нейропротек-
жения веса. В дальнейшем была показана его
торных эффектах МС, сколько проанализиро-
значительная острая токсичность, и 2,4-ДНФ
вать основные метаболические и сигнальные
был запрещён для использования в некоторых
пути, на которые оказывает влияние МС. Осо-
странах [34]. Но не только протонофоры могут
бое внимание уделяется способности МС осу-
вызывать рассеивание мембранного потенци-
ществлять альтернативный транспорт электро-
ала. Альтернативный транспорт электронов с
нов в митохондриях, активировать аутофагию
помощью MС является ещё одним способом
и сигнальный путь Nrf2/ARE, ингибировать
модуляции митохондриального метаболиз-
моноаминооксидазы (МАО) и NO-синтазы
ма, который, помимо снижения мембранного
(NOS), препятствовать апоптозу, воспалитель-
потенциала, позволяет обходить нарушенные
ным процессам и агрегации неправильно свёр-
или заингибированные дыхательные комплек-
нутых белков.
сы [35]. MС одобрен управлением по санитар-
ному надзору за качеством пищевых продуктов
и медикаментов (FDA), поэтому повторение
МЕТИЛЕНОВЫЙ СИНИЙ
истории с неожиданным обнаружением ток-
КАК АЛЬТЕРНАТИВНЫЙ
сичности 2,4-ДНФ крайне маловероятно.
ПЕРЕНОСЧИК ЭЛЕКТРОНОВ
Первые сведения о том, что МС может уве-
личивать скорость дыхания, были получены
Этиология большинства нейродегенера-
в конце 1920-х - начале 1930-х гг. Исследова-
тивных заболеваний зачастую комплексная.
ния проводились на безъядерных эритроци-
Наиболее распространёнными факторами,
тах птиц и млекопитающих [36], неоплодот-
которые способствуют нейродегенерации, яв-
ворённых яйцах морских звёзд [37], в которых
ляются митохондриальные дефекты и окисли-
изучались окислительно-восстановительные
тельный стресс. Митохондриальные дефекты
свойства МС. В 1938 г. Elliott и Greig [38] об-
приводят к увеличению скорости образова-
наружили, что сукцинат может окисляться до
ния активных форм кислорода (АФК), а окис-
фумарата в присутствии МС, который «прини-
лительный стресс, в свою очередь, являет-
мает атомы водорода и восстанавливается до
БИОХИМИЯ том 87 вып. 9 2022
1166
ГУРЕЕВ и др.
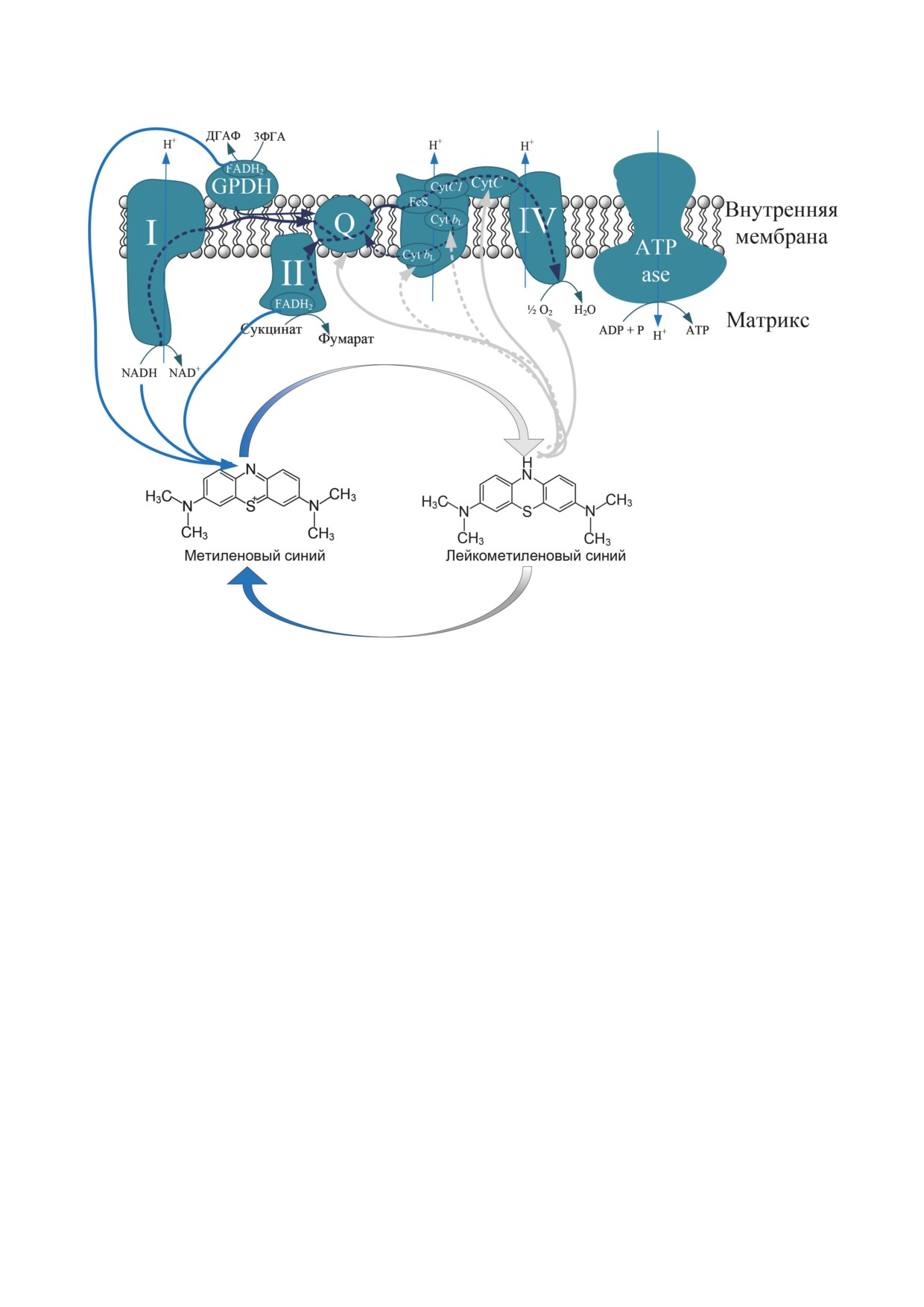
Рис. 2. Функционирование МС в качестве альтернативного переносчика электронов. МС может принимать электроны
от NADH и FADH2, которые в норме окисляются в I- и II-комплексах ЭТЦ, а также α-глицерофосфатдегидрогена-
зы (GPDH), которая катализирует реакцию превращения глицеральдегид-3-фосфата (3-ФГА) в дигидроксиацетонфос-
фат (ДГАФ). При этом сам МС восстанавливается до бесцветной лейкоформы. Лейко-МС может отдавать электроны на
убихинон, комплекс III, цитохром с (Cyt С), а также на молекулярный кислород
лейко-МС». Тогда как в обычных условиях, без
щитель фосфорилированного дыхания [42, 43].
МС, акцептором протонов являлся окислен-
Последующие работы показали, что MС увели-
ный цитохром. В 1949 г. Slater [39] изучал сук-
чивает активность цитохром с-оксидазы [44].
цинатдегидрогеназный комплекс в сердечной
Это подтвердило предположение о том, что
мышце и в почках и показал, что цитохром b
МС функционирует как альтернативный пере-
участвует в катализе восстановления МС сук-
носчик электронов (рис. 2). Донором электро-
цинатом (рис. 2). Далее было обнаружено,
нов для МС является NADH, а цитохром с -
что убихинон и его гомологи могут опосредо-
акцептором [35]. Позже было показано, что
вать реакцию между электрон-транспортной
MС увеличивает скорость дыхания, если в
цепью (ЭТЦ) и MС [40], а также тот факт, что
качестве субстрата для дыхания используется
MС может окислять NADPH [41].
сукцинат и α-глицерофосфат. Таким образом,
Исследования МС на изолированных ми-
сукцинатдегидрогенеза и α-глицерофосфатде-
тохондриях начались ближе к началу XXI века.
гидрогеназа (GPDH) также способны участво-
Было показано, что MС может стимулировать
вать в передаче электронов на МС [45] (рис. 2).
митохондриальное дыхание, которое было ин-
При этом МС может переносить электроны не
гибировано хлорацетальдегидом, модулирую-
только на цитохром с, но и непосредственно
щим митохондриальный окислительный ме-
на О2, что проявлялось в МС-зависимом уве-
таболизм длинноцепочечных жирных кислот
личении скорости продукции Н2О2, которое
в печени. В то же время МС стимулировал пре-
было неоднократно продемонстрировано [43,
имущественно дыхание в состоянии 4 по Чансу
45-47] (рис. 2). Таким образом, механически
(нефосфорилирующее дыхание), а не в состоя-
МС является прооксидантом, который мо-
нии 3 по Чансу (фосфорилирующее дыхание),
жет увеличивать скорость продукции H2O2,
что позволяет характеризовать МС как разоб-
что было показано как при добавлении МС
БИОХИМИЯ том 87 вып. 9 2022
НЕЙРОПРОТЕКТОРНЫЙ ЭФФЕКТ МЕТИЛЕНОВОГО СИНЕГО
1167
к изолированным митохондриям [45, 48], так
повышению уровня АФК и окислительному
и при терапии МС in vivo [46]. Но при этом в
стрессу, вызывают нейровоспаление за счёт
концентрациях до 100 мкМ не было показано
активации микроглии, что приводит к повреж-
генотоксичного эффекта МС в отношении ми-
дению синапсов и гибели нейронов [55].
тохондриального генома [48]. В 2015 г. в работе
В 1996 г. Wischik et al. [15] впервые по-
Atamna et al. [49] окислительный стресс, инду-
казали, что MС в наномолярных концен-
цированный МС, характеризуется термином
трациях ингибирует агрегацию тау-белка и
«мягкий окислительный стресс», который не
нарушает стабильность протеазорезистент-
оказывает деструктивного эффекта на клеточ-
ных парных спиральных филаментов (paired
ные компоненты, но может запускать некото-
helical filaments
- PHF) (рис.
3). Инги-
рые сигнальные механизмы (в том числе и опо-
бирование образования тау-филаментов
средующие антиоксидантный эффект МС),
с помощью MС было впоследствии под-
которые обсуждаются в последующих разделах.
тверждено Taniguchi et al. [56] - в 2005 г., и
Альтернативный транспорт электронов по-
Hattori et al. [57] - в 2008 г. Диета с добавле-
зволяет обходить ингибированный комплекс I.
нием MС приводила к значительному сниже-
Было показано, что МС шунтирует блок, ко-
нию уровня фосфорилированного тау-белка у
торый создавался ротеноном [45, 47, 50, 51] и
мышей линии P301S [58] и линии TauΔK [59].
диметакрилат триэтиленгликолем (Triethylene
MС ингибировал олигомеризацию мономеров
glycol dimethacrylate - TEGDMA) [52]. Од-
амилоида-β
[60]. Также фотовозбуждённые
нако открытым остаётся вопрос о возмож-
молекулы МС блокировали агрегацию амило-
ности обхода комплекса III. Tretter et al. [45]
ида-β за счёт его окисления синглетным кисло-
и Sváb et al. [53] показали, что MС увеличивал
родом, образующимся в ходе фотохимической
скорость дыхания и восстанавливал мембран-
реакции [61]. MС активировал Ca2+-ATPазу
ный потенциал при ингибировании антими-
плазматической мембраны (plasma membrane
цином и миксотиазолом, что согласуется с
Ca2+ATPase - PMCA), играющую важную роль
данными, свидетельствующими о повышении
в поддержании гомеостаза цитозольного каль-
активности цитохром с-оксидазы. В 2019 г.
ция в концентрации, которая необходима для
Gureev et al. [47], напротив, показали, что MС
нормального функционирования нейронов.
не восстанавливает дыхание и мембранный по-
МС препятствует инактивации PMCA, вызван-
тенциал, ингибированные антимицином, но
ной накоплением амилоида-β и тау [62, 63].
при этом восстанавливает дыхание и мембран-
Неправильный фолдинг белков игра-
ный потенциал, ингибируемые ротеноном.
ет ключевую роль в патогенезе болезни Хан-
Это более согласуется с результатами Slater [39],
тингтона (рис. 3). Мутантная форма белка
показавшего участие цитохрома b в катализе
хантингтина (HTT) склонна к расщеплению,
восстановления МС сукцинатом (рис. 2). При-
неправильному фолдингу и агрегации. Накоп-
чина различий нуждается в дальнейшем разъ-
ление агрегатов в конечном итоге приводит к
яснении. Однако эти расхождения в механизме
дисфункции нейронов [64]. MС ингибировал
функционирования не носят принципиальный
агрегацию нерастворимого мутантного бел-
характер, так как нарушения в комплексе I, а не
ка HTT in vitro и снижал нейротоксичность [19].
в комплексе III чаще всего приводят к развитию
Дальнейшие исследования показали, что MС
митохондриально-зависимых патологий [54].
защищает сердце Drosophila melanogaster от
HTT-индуцированной токсичности, но не ока-
зывает эффекта в нейронах из-за плохого про-
МЕТИЛЕНОВЫЙ СИНИЙ
хождения через гематоэнцефалический барьер
КАК ИНГИБИТОР АГРЕГАЦИИ БЕЛКОВ
D. melanogaster [20].
Имеются данные, показывающие, что МС
Неправильный фолдинг, агрегация и ак-
является потенциально эффективным препа-
кумулирование белков в головном мозге зача-
ратом для лечения прионных болезней - се-
стую являются причиной запуска патологи-
мейство редких, но фатальных патологий,
ческих процессов, которые приводят к гибели
поражающих людей и различные виды жи-
нейронов и развитию нейродегенеративных
вотных. Было продемонстрировано, что МС
заболеваний. Накопление белков с неправиль-
влияет на кинетику олигомеризации непра-
ной структурой, таких как амилоид-β и фосфо-
вильно свёрнутого прионного белка (PrP) и
рилированный тау-белок, связано с развитием
уменьшает количество олигомера примерно
болезни Альцгеймера. Неправильно свёрнутые
на 30% [65] (рис. 3).
и агрегированные белки вызывают митохон-
Ряд исследований выявили связь между па-
дриальную дисфункцию, которая приводит к
тогенезом бокового амиотрофического скле-
БИОХИМИЯ том 87 вып. 9 2022
1168
ГУРЕЕВ и др.
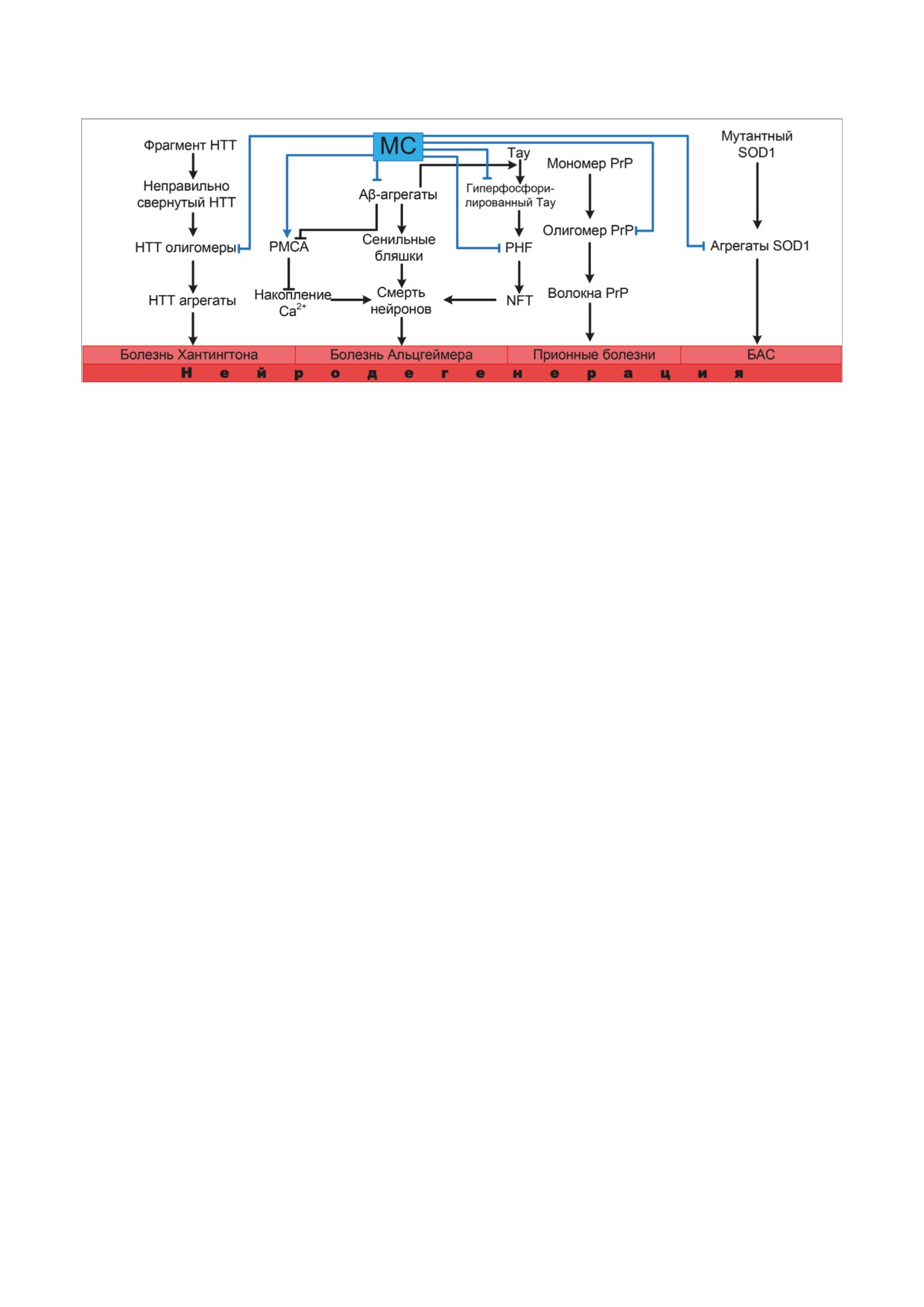
Рис. 3. МС ингибирует олигомеризацию и последующее агрегирование расщеплённого мутантного белка HTT, что мо-
жет замедлять патогенез болезни Хантингтона. МС препятствует накоплению амилоида-β (Aβ), гиперфосфорилиро-
ванию тау-белка и, следовательно, образованию нейрофибриллярных клубков (NFT), способствует активации PMCA,
что в сумме позволяет замедлить патогенез болезни Альцгеймера. МС препятствует олигомеризации неправильно свёр-
нутого прионного белка (PrP) и неправильно свёрнутого SOD1, что замедляет течение прионных заболеваний и БАС
роза (БАС), неправильным фолдингом и агре-
ватором CBP (CREB-binding protein), который
гацией супероксиддисмутазы 1 (SOD1) [66].
обладает гистон-ацетилтрансферазной актив-
Было показано, что MС модулирует спонтан-
ностью, приводящей к изменениям в структу-
ную амилоидную агрегацию SOD1 в клеточных
ре хроматина. Это обеспечивает релаксацию
линиях [67, 68] и животных моделях Danio rerio
ARE-содержащего промотора и связывание с
и Caenorhabditis elegans [69, 70] (рис. 3).
РНК-полимеразой. Подобный механизм регу-
лирует экспрессию большинства генов-мише-
ней Nrf2 [73].
МЕТИЛЕНОВЫЙ СИНИЙ КАК АКТИВАТОР
Было показано, что МС увеличивает экс-
СИГНАЛЬНОГО ПУТИ NRF2/ARE
прессию гена Nfe2l2 (ген, кодирующий Nrf2),
генов антиоксидантов: Ho-1 (кодирующего
Сигнальный путь Nrf2/ARE (Nuclear factor
гем-оксигеназу), Nqo1 (кодирующего NAD(P)H
erythroid 2-related factor 2/antioxidant response
хинондегидрогеназу 1), Gclc и Gclm (кодиру-
element) регулирует экспрессию генов, кодиру-
ющих каталитическую и модифицирующую
ющих белки с цитопротекторными свойствами,
субъединицы гамма-глутамилцистеинсинте-
например, антиоксидантные ферменты, белки
тазы соответственно), Trx1и Trx2 (кодирую-
фазы II детоксикации ксенобиотиков, проти-
щих тиоредоксины 1 и 2), TrxR1 и TrxR2 (ко-
вовоспалительные белки, а также метаболиче-
дирующих тиоредоксинредуктазы 1 и 2), Grx1
ские ферменты и регуляторы, участвующие в
и Grx2 (кодирующих глутаредоксины 1 и 2),
поддержании окислительно-восстановитель-
Prx6 (кодирующего перкосиредоксин
6) и
ного гомеостаза [71] (рис. 4). Ядерный фактор,
препятствует нейротоксичности, ассоцииро-
связанный с эритроидным фактором 2 (Nrf2),
ванной с тау-белком [45]. MС препятствовал
представляет собой короткоживущий белок
возрастному снижению экспрессии антиок-
(время жизни около 15 мин), который при от-
сидантного гена Sod2 (кодирующего суперок-
сутствии активирующих факторов подвергает-
сиддисмутазу) [46] (рис. 4). Участие пути Nrf2/
ся убиквитинированию и протеосомной дегра-
ARE в задержке клеточного старения обсужда-
дации. На сегодняшний день хорошо описаны
лось Atamna et al. [49]. Активация Nrf2, инду-
две системы убиквитинлигаз, которые опо-
цированная MС, оказывала терапевтический
средуют деградацию Nrf2: Kelch-подобный
эффект при колите, индуцированном TNBS
ECH-ассоциированный белок 1 (Keap1) и ки-
(2,4,6-Trinitrobenzene sulfonic acid) [74]. До сих
наза гликогенсинтазы 3β (GSK3β) [72]. Ин-
пор нет прямых доказательств того, что MС
дукторы Nrf2 представляют собой факторы,
непосредственно окисляет Keap1 или GSK3β.
которые способствуют высвобождению Nrf2
Было высказано предположение, что инду-
от Keap1 или GSK3β, что приводит к трансло-
цированная МС продукция H2O2 может сти-
кации Nrf2 в ядро и связыванию с факторами
мулировать окисление Keap1 или GSK3β,
транскрипции основной лейциновой мол-
последующую активацию Nrf2 и усиление анти-
нии (bZip) (чаще всего малыми MAF) и коакти-
оксидантной защиты [45, 46]. Примечательно,
БИОХИМИЯ том 87 вып. 9 2022
НЕЙРОПРОТЕКТОРНЫЙ ЭФФЕКТ МЕТИЛЕНОВОГО СИНЕГО
1169
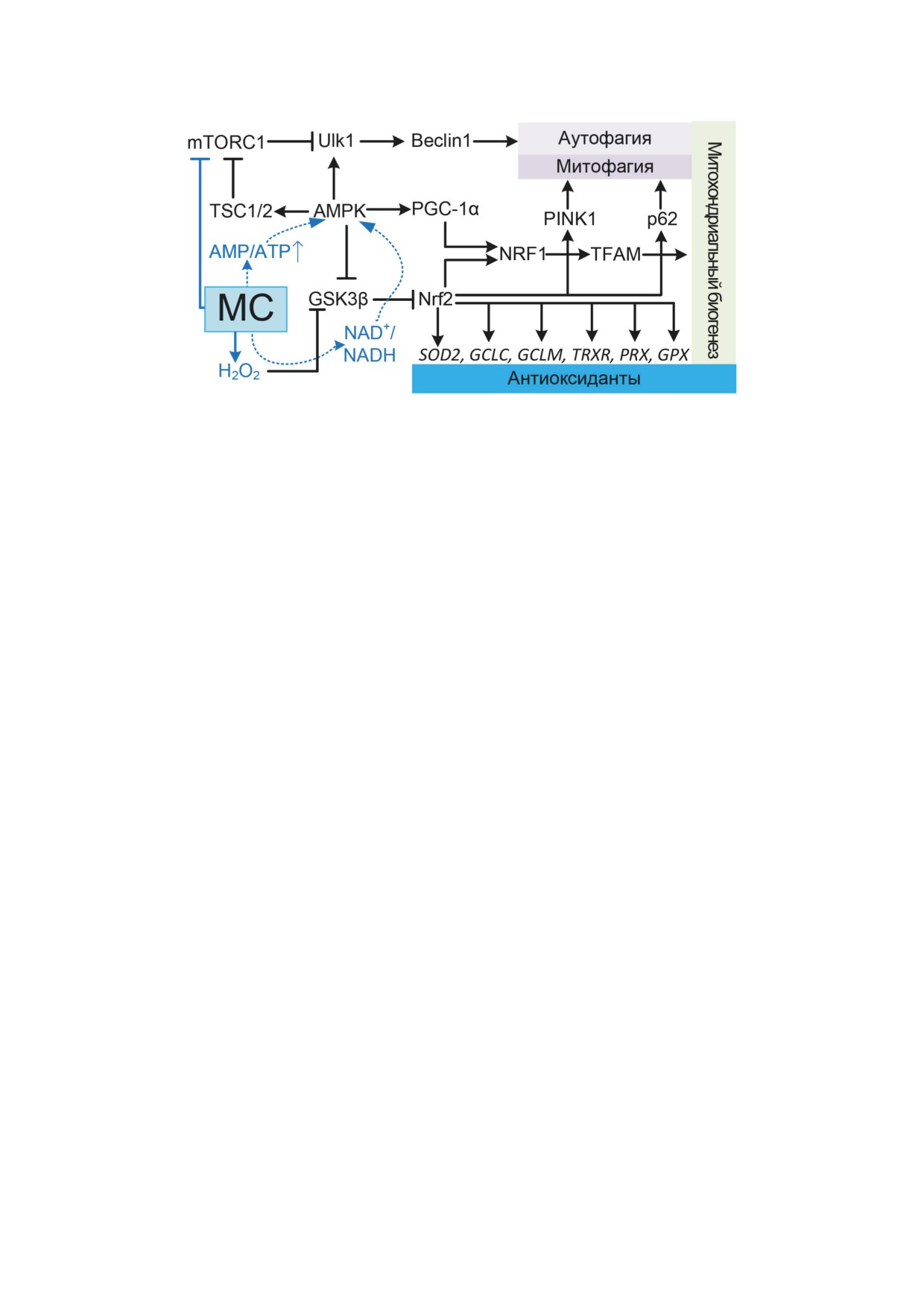
Рис. 4. Влияние МС на сигнальный путь Nrf2/ARE, который регулирует экспрессию генов, кодирующих ключевые ан-
тиоксидантные белки, отвечает за координацию процессов митохондриального биогенеза и митофагии. Влияние МС
на аутофагию путём ингибирования mTORC1, а также по mTORC1-независимому пути
что у Escherichia coli MС также способен ин-
равновесия AMP/ATP в сторону AMP. С другой
дуцировать экспрессию генов, защищающих
стороны, нами было показано, что альтерна-
от окислительного стресса путём активации
тивный транспорт электронов вызывает сни-
регулона soxRS, что натолкнуло авторов ис-
жение трансмембранной разности потенциала,
следования на аналогию с активацией Nrf2/
что, по всей видимости, должно замедлять ско-
ARE-пути [75].
рость синтеза ATP и, следовательно, смещать
Другой важной функцией сигнального
равновесие AMP/ATP в сторону AMP [43].
пути Nrf2/ARE является регуляция митохон-
Atamna et al. [49] предположили, что МС за
дриального биогенеза
[76]. МС увеличива-
счёт альтернативного транспорта электронов
ет количество митохондрий в головном мозге
вызывает смещение равновесия NAD+/NADH
15-месячных мышей, что проявляется в уве-
в сторону NAD+, что приводит к увеличению
личении числа копий мтДНК, увеличении
уровня фосфорилирования AMPK, и подтвер-
экспрессии гена Cox1 (кодируется мтДНК),
дили это экспериментально.
а также Nrf1 и Tfam, которые принимают непо-
средственное участие в процессах транскрип-
ции мтДНК [46] (рис. 4). Влияние МС на ак-
МЕТИЛЕНОВЫЙ СИНИЙ
тивацию митохондриального биогенеза было
КАК АКТИВАТОР АУТОФАГИИ
отмечено Atamna et al. [49], однако указывает-
ся, что это достигается за счёт активации пути
Аутофагия - это процесс самоуничтоже-
AMPK/PGC-1α (AMP activated protein kinase/
ния клеточных компонентов, при котором
Peroxisome proliferator-activated receptor gamma
сначала двухмембранные аутофагосомы изо-
coactivator 1α), который долгое время считал-
лируют органеллы, затем сливаются с лизо-
ся главным регулятором митохондриального
сомами для лизиса гидролазами. Аутофагия
биогенеза [77]. В то же время активация пути
активируется в ответ на внеклеточные или
Nrf2/ARE не указана как путь активации ми-
внутриклеточные стрессовые факторы
[80].
тохондриального биогенеза [49]. В последние
Стресс запускает процессы, которые приво-
годы обсуждаются перекрёстные связи между
дят к активации AMPK или p53, что снижает
путями Nrf2/ARE и AMPK/PGC-1α [76], ко-
активность комплекса 1 мишени рапамицина
торые опровергают упрощённое представле-
млекопитающих (mTORC1, mammalian target of
ние о «главном регуляторе митохондриально-
rapamycin complex 1) (рис. 4). Снижение актив-
го биогенеза» для пути AMPK/PGC-1α [78].
ности mTORC1 приводит к повышению актив-
Не менее интересен и механизм активации
ности комплекса ULK1 (Unc-51 like autophagy
AMPK с помощью МС. С одной стороны, МС
activating kinase), что впоследствии иниции-
восстанавливает митохондриальное дыхание
рует образование фагофора совместно с ком-
и производство АТP в условиях нарушения
плексом фосфоинозитид-3-киназы (PI3K;
функционирования ЭТЦ. По этой причине,
включает белки PI3K-III, Beclin-1, VPS34,
по предположению Xie et al. [79], активация
p150 и ATG14). Рост и созревание фагофора
AMPK не может происходить за счёт смещения
зависит от двух убиквитин-подобных систем
БИОХИМИЯ том 87 вып. 9 2022
1170
ГУРЕЕВ и др.
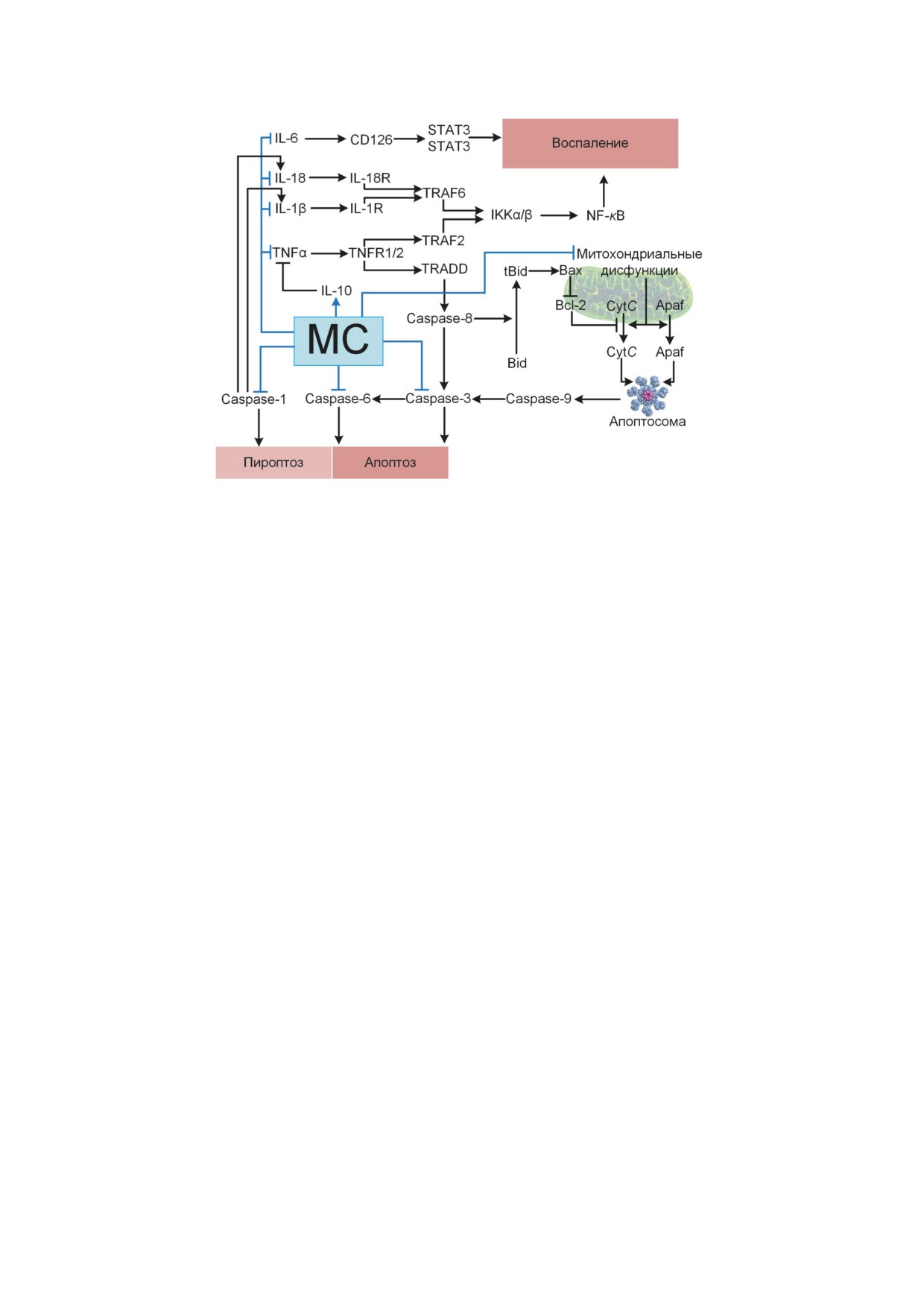
Рис. 5. Влияние МС на апоптоз за счёт ингибирования эффекторных каспаз и устранения митохондриальных дисфунк-
ций, которые являются частой причиной выхода цитохрома с (Cyt C) и Apaf1 в цитоплазму. Влияние МС на пироптоз и
воспалительные процессы за счёт ингибирования каспазы-1, провоспалительных цитокинов и активации противовос-
палительного цитокина
конъюгации, ATG12 и ATG8 [81]. Наруше-
сигнального пути Nrf2/ARE, так как имеются
ния в регуляции аутофагии наблюдаются при
данные, что белок p62 образует петлю обрат-
многих неврологических расстройствах чело-
ной связи с Nrf2 [88]. Кроме того, Nrf2 может
века, таких как болезнь Альцгеймера, болезнь
регулировать экспрессию гена PINK1
[89].
Паркинсона, болезнь Хантингтона и БАС [82].
И p62, и PINK1 играют ключевую роль в регу-
Изначально было показано, что тера-
ляции митофагии (рис. 4).
пия MС не влияет на индукцию аутофагии, по-
скольку не изменялись уровни Beclin-1 (ком-
плекс PI3K), Atg
7 (система ATG12) и
МЕТИЛЕНОВЫЙ СИНИЙ
LC3-II (система ATG8) у мышей 3xTg-AD [83].
КАК СУПРЕССОР АПОПТОЗА
Позже появились данные о том, что MС оказы-
вает влияние на уровень LC3-II. Это позволило
При оценке влияния МС на апоптоз необ-
предположить, что он является мощным индук-
ходимо чётко разграничить условия, при кото-
тором аутофагии у мышей с таупатией. Было
рых происходит терапия. В условиях фотодина-
высказано предположение, что MС функцио-
мической терапии МС будет функционировать
нирует как рапамицин и ингибирует mTOR,
как фотосенсибилизатор и стимулировать
тем самым вызывая активацию аутофагии [84].
апоптоз [90]. Фотодинамическая терапия яв-
Jiang et al. [85] показали, что MС-индуцирован-
ляется достаточно широко используемым ме-
ная аутофагия участвует в инактивации p53,
тодом лечения различных злокачественных и
которая активирует AMPK. AMPK, в свою
доброкачественных опухолей. Фотосенсиби-
очередь, ингибирует mTOR, активируя Tsc2.
лизатор поглощает энергию источника света и
Однако есть данные, что MС также приводит
может передавать её молекулярному кислороду
к аутофагии за счёт активации AMPK, которая
с образованием синглетного кислорода (1O2).
фосфорилирует ULK1, а уровень фосфорили-
1O2 является крайне электрофильной формой
рования mTORC1 не изменяется [79]. ULK1
кислорода и может окислять биологические
фосфорилирует Beclin-1, который необходим
макромолекулы, тем самым повреждая их.
для инициации аутофагии. Уровень Beclin-1
Фотосенсибилизатор должен быть амфифиль-
увеличивается при лечении черепно-мозговой
ным, накапливаться в опухолевой ткани и бы-
травмы с помощью MС [86].
стро выводиться из нормальной ткани, а также
Имеются данные о том, что МС может ин-
иметь низкий уровень токсичности в отсут-
дуцировать митофагию, частный случай ауто-
ствии освещения [91]. Всем этим требованиям
фагии
[87]. Вероятно, МС-индуцированная
отвечает МС [92]. Но в контексте нейропро-
митофагия может быть связана с активацией
текторного эффекта МС нас интересуют его
БИОХИМИЯ том 87 вып. 9 2022
НЕЙРОПРОТЕКТОРНЫЙ ЭФФЕКТ МЕТИЛЕНОВОГО СИНЕГО
1171
антиапоптотические свойства, которые будут
У мышей с ишемией головного мозга, которые
рассмотрены ниже.
подвергались терапии МС, уровни p53 и Bax
Апоптоз можно разделить на три фазы:
были снижены, а количество белка Bcl-2 было
внешний путь, внутренний путь и эффек-
увеличено [85]. MС также снижал уровень Bax
торная фаза. Внешний путь активируется
и увеличивал экспрессию белка Bcl-2 при ко-
лигандами смерти (TNF-α (Tumor necrosis
лите, вызванном TNBS [74], и индуцирован-
factor-alpha), FASL (Fas ligand), TRAIL (TNF-
ном ротеноном повреждении нигростриаль-
related apoptosis-inducing ligand)), которые
ной системы [101]. МС, по всей видимости, не
связываются с соответствующими рецептора-
напрямую взаимодействует с Bcl-2 и Bax, а кос-
ми на клеточной поверхности, запуская про-
венно. Скорее всего, МС оказывает влияние на
цесс их агрегации [93] (рис. 5). Эта агрегация
уровни про- и антиапоптотических факторов,
задействует адапторные белки в цитоплаз-
ингибируя TNF-α [100, 101] или препятствуя
матическом участке рецепторов, формируя
возникновению митохондриальных дисфунк-
сигнальный комплекс, индуцирующий кле-
ций. Например, МС предотвращал открытие
точную гибель (DISC, death-inducing signaling
митохондриальных пор [102].
complex). DISC стимулирует автокаталитиче-
МС снижал количество цитохрома с в ци-
скую активацию каспазы-8 и её высвобожде-
тозоле и активность каспазы-3 [26, 27, 101-
ние в цитоплазму, где она активирует каспаз-
104], каспазы-6 [104, 105], каспазы-8 [26, 103],
ный каскад [94].
каспазы-9 [27, 102] и каспазы-1, отвечающей
Внутренний путь апоптоза ассоциирован
за пироптоз [106]. Для каспазы-1, каспазы-3 и
с митохондриями. Различные митохондри-
каспазы-6 было показано, что МС может непо-
альные дисфункции, например, пермеабили-
средственно окислять их каталитические доме-
зация наружной мембраны, открытие мито-
ны и ингибировать их активность [103] (рис. 5).
хондриальных пор проницаемости, вызывают
Были получены и противоположные данные,
выход цитохрома с и Apaf1 (Apoptotic protease
показывающие, что МС уменьшал острое ише-
activating factor 1) из митохондрий [95] (рис. 5).
мическое повреждение головного мозга, но ко-
Эти два компонента олигомеризуются и обра-
личество апоптотических клеток не уменьша-
зуют сложную структуру - апоптосому, которая
лось, а даже несколько увеличивалось, однако
необходима для расщепления прокаспазы до её
при этом снижался процент некротических
активной формы, каспазы-9, которая, в свою
клеток [87].
очередь, расщепляет и активирует другие про-
каспазы в эффекторные каспазы [96]. Внешний
и внутренний пути апоптоза связываются друг
МЕТИЛЕНОВЫЙ СИНИЙ
с другом с помощью белка Bid (BH3 interacting-
КАК СУПРЕССОР ВОСПАЛЕНИЯ
domain death agonist). Активная каспаза-8 рас-
щепляет белок Bid. Укороченная форма Bid
Воспаление представляет собой сложную
(truncated Bid, tBid) транслоцируется в ми-
реакцию организма на различные внешние
тохондрии и активирует проапоптотические
и внутренние раздражители. Воспаление ха-
белки (Bcl-2-associated X protein, Bax) и деак-
рактеризуется физиологическими процессами
тивирует антиапоптотические белки (B-cell
(увеличением артериального давления, изме-
lymphoma 2, Bcl-2), которые влияют на высво-
нением проницаемости сосудов), клеточными
бождение цитохрома с [97]. Эффекторная фаза
процессами (увеличением количества лейко-
апоптоза осуществляется каспазами 3 и 6. Они
цитов) и молекулярными процессами (увели-
вызывают деградацию ядерного материала, в
чением уровня провоспалительных медиато-
том числе ДНК, а также других клеточных ком-
ров). Неконтролируемое острое воспаление
понентов, в частности цитоскелета [98].
может стать причиной хронического воспале-
Запрограммированная гибель клеток по-
ния, способствуя развитию различных нейро-
средством апоптоза служит естественным ба-
дегенеративных заболеваний [107]. Иммунный
рьером для развития опухолей. С другой сто-
ответ человека регулируется сложной сетью
роны, активация апоптоза играет ключевую
медиаторов, таких как интерлейкины (IL-1β,
роль в прогрессировании неврологических
IL-6, IL-17, IL-18, IL-10), TNF-α и др. (рис. 5).
расстройств, таких как болезнь Альцгеймера,
Воспалительные стимулы связываются со
Паркинсона, Хантингтона, черепно-мозговая
специфическими рецепторами. Активация ре-
травма и БАС, как показано в исследованиях на
цептора запускает важные внутриклеточные
многих моделях [99]. Терапия с MС может по-
сигнальные пути, такие как ядерный фактор
давлять апоптоз через PI3-K/Akt/GSK-3β-за-
каппа-B (NF-κB), митоген-активируемая про-
висимый путь при кровоизлиянии в мозг [100].
теинкиназа (MAPK), янус-киназа (JAK), сиг-
БИОХИМИЯ том 87 вып. 9 2022
1172
ГУРЕЕВ и др.
нальные белки и активаторы транскрипции из
центрации норадреналина, серотонина и до-
семейства белков STAT (Signal transducer and
фамина (рис. 6). МАО имеет две изоформы:
activator of transcription) [108].
МАО типа А и МАО типа В. МАО-А в первую
Уровни провоспалительных цитокинов в
очередь отвечает за метаболизм серотонина.
тканях являются маркерами воспалительных
Обе изоформы ответственны за метаболизм
процессов. TNF-α - один из наиболее мульти-
дофамина [116]. Согласно серотониновой тео-
функциональных и хорошо изученных цито-
рии депрессии, МАО-А катализирует окисли-
кинов. Его основными источниками являются
тельное дезаминирование серотонина (5-НТ),
макрофаги и Т-клетки, но было описано, что
превращая его в 5-гидрокси-3-индолуксусный
многие другие клетки, такие как B-клетки,
альдегид (5-HIAL), который в дальнейшем пе-
нейтрофилы и эндотелиальные клетки, также
рерабатывается в
5-гидрокси-3-индолуксус-
продуцируют TNF-α. Несколько исследований
ную кислоту (5-HIAA) с помощью альдегидде-
показали, что MС вызывает снижение уровня
гидрогеназы [117].
TNF-α в различных тканях и при различных
Астроциты также участвуют в регуляции
физиологических состояниях [99, 100] (рис. 5).
метаболизма серотонина за счёт поглощения
TNF-α связывается с TNFR1/2 и приводит к
серотонина транспортёром обратного захвата
TRAF-2 (TNF receptor-associated factor 2)-зави-
серотонина (SERT) и последующей деграда-
симой активации NF-κB [109]. NF-κB контро-
ции с помощью МАО-А [118]. Кроме того, фер-
лирует экспрессию многих генов, участвующих
мент МАО-А опосредует окисление дофамина
в воспалении, и, по-видимому, NF-κB прояв-
в 3,4-дигидроксифенилацетальдегид (DOPAL)
ляет хроническую активность при многих вос-
наряду с образованием H2O2 в пресинапти-
палительных заболеваниях [110]. MС также
ческих окончаниях. МАО-В метаболизиру-
снижал уровень NF-κB [101].
ет дофамин в 3,4-дигидроксифенилуксусную
IL-1α и IL-1β являются каноническими
кислоту (DOPAC), а катехол-О-метилтрансфе-
активаторами NF-κB. МС значительно снижал
раза (COMT) расщепляет его до гомованили-
экспрессию и уровень белков IL-1α/β [100, 111,
новой кислоты (HVA) в астроцитах [119]. Ан-
112]. IL-6 также является провоспалительным
тидепрессивный эффект ингибирования МАО
цитокином. Передача сигнала IL-6 включа-
заключается в повышении уровня всех биоген-
ет активацию JAK, а затем приводит к акти-
ных аминов [120].
вации транскрипционного фактора STAT3.
Первое упоминание о том, что МС ингиби-
STAT3 фосфорилируется и образует димеры с
рует МАО, было сделано в 1996 г. [121]. MС ин-
другими членами семейства STAT. Активиро-
гибирует обе изоформы МАО, но МАО-А
ванный комплекс STAT3 транслоцируется в
в несколько раз сильнее, чем МАО-В [122]
ядро и регулирует экспрессию широкого спек-
(рис. 6, б). Позже эффективность МС против
тра генов, в том числе генов, кодирующих про-
депрессии была показана на животных мо-
воспалительные цитокины [113]. Неоднократ-
делях [123]. Стоит отметить, что некоторые
но было показано, что МС снижает уровень
производные МС, например азур B, более эф-
IL-6 [29, 74, 100, 112] и других провоспалитель-
фективны при лечении психотических рас-
ных факторов: IL-12 [29], IL-17 [74, 112] и IL-
стройств [4, 60, 124, 125].
18 [100, 112] (рис. 5).
Не все интерлейкины являются провоспа-
лительными. Например, IL-10 играет решаю-
МЕТИЛЕНОВЫЙ СИНИЙ
щую роль в предотвращении воспалительных
КАК ИНГИБИТОР NO-СИНТАЗЫ
и аутоиммунных патологий
[114]. Имеются
данные о том, что IL-10 подавляет работу дру-
Белки семейства NO-синтаз (NOS) явля-
гих провоспалительных цитокинов, таких как
ются важными регуляторами психологическо-
TNF-α [115]. МC повышал уровень IL-10 в тка-
го состояния человека. Существует несколько
нях в соответствии с его противовоспалитель-
форм NOS, выполняющих определённые функ-
ным действием [100, 112] (рис. 5).
ции. Нейрональная NOS (nNOS) локализует-
ся в синаптических шипиках, эндотелиальная
NOS (eNOS) - в эндотелиальных клетках сосу-
МЕТИЛЕНОВЫЙ СИНИЙ
дов головного мозга и двигательных нейронах,
КАК ИНГИБИТОР
индуцируемая NOS (iNOS) - в астроцитах и
МОНОАМИНОКСИДАЗЫ
микроглии и индуцируется при некоторых па-
тологических состояниях [126]. Воспалитель-
Моноаминоксидазы (МАО) регулируют
ные процессы являются важными факторами
синаптические и внутринейрональные кон-
активации iNOS в астроцитах и микроглии гип-
БИОХИМИЯ том 87 вып. 9 2022
НЕЙРОПРОТЕКТОРНЫЙ ЭФФЕКТ МЕТИЛЕНОВОГО СИНЕГО
1173
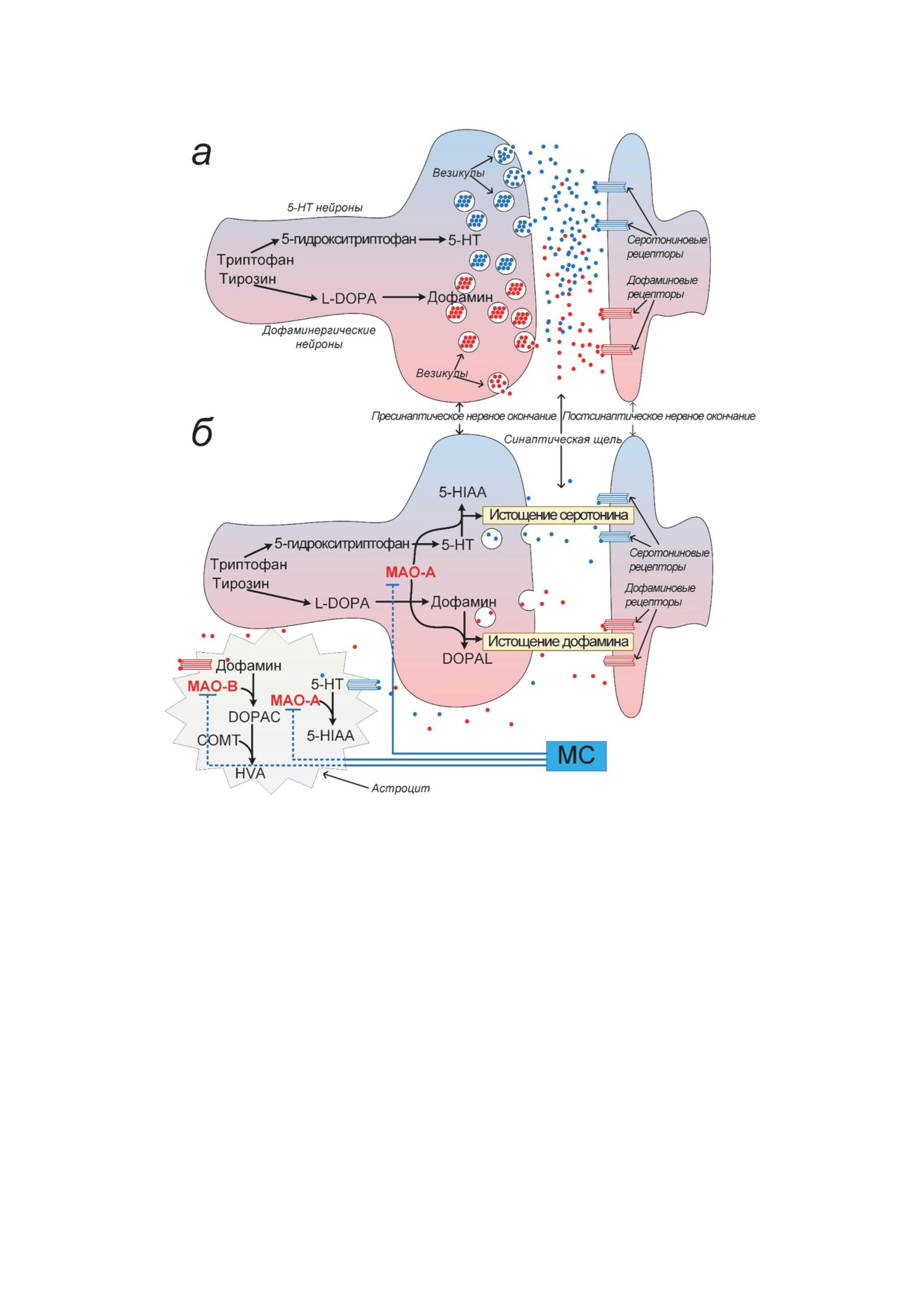
Рис. 6. Влияние МС на метаболизм серотонина и дофамина. а - Синтез и метаболизм серотонина и дофамина в нор-
мальных физиологических условиях. б - Метаболизм серотонина и дофамина в условиях активации MAO-A и МАО-В
в синапсах и астроцитах. МАО-А способствует окислению дофамина в 3,4-дигидроксифенилацетальдегид (DOPAL).
МАО-В метаболизирует дофамин в 3,4-дигидроксифенилуксусную кислоту (DOPAC), а катехол-О-метилтрансфера-
за (COMT) расщепляет его до гомованилиновой кислоты (HVA) в астроцитах. МС препятствует истощению серотонина
и дофамина за счёт ингибирования MAO-A и MAO-B
покампа [127] (рис. 7). Повышение уровня NO
кортикоидный рецептор (MR). В свою очередь,
приводит к активации растворимой гуанилат-
NO подавляет локальную экспрессию глюко-
циклазы (sGC)/циклического гуанозинмоно-
кортикоидных рецепторов (GR) посредством
фосфата (cGMP). Этот сигнальный путь яв-
сигнальных путей sGC/cGMP и пероксинит-
ляется ключевой особенностью когнитивного
рита (ONOO-), регулируемых внеклеточным
дефицита и депрессии [128]. Сигнальный путь
сигналом киназ (ERK). Это приводит к повы-
iNOS/NO/sGC/cGMP в пресинаптических
шению уровня гипоталамического кортикотро-
окончаниях вызывает высвобождение глутама-
пин-рилизинг-гормона (CRH) [130]. Гипотала-
та, который активирует постсинаптические ре-
мическая секреция CRH стимулирует переднюю
цепторы N-метил-D-аспартата (NMDAR) для
долю гипофиза к высвобождению адренокорти-
увеличения концентрации Ca2+ и дальнейшей
котропного гормона (ACTH), который вызыва-
активации NO, полученного от nNOS [129].
ет надпочечниковую секрецию глюкокортикои-
Кроме того, глюкокортикоиды могут активиро-
дов. Повышенный уровень глюкокортикоидов,
вать nNOS через высокоаффинный минерало-
CRH и ACTH, является фактором депрессивно-
БИОХИМИЯ том 87 вып. 9 2022
1174
ГУРЕЕВ и др.
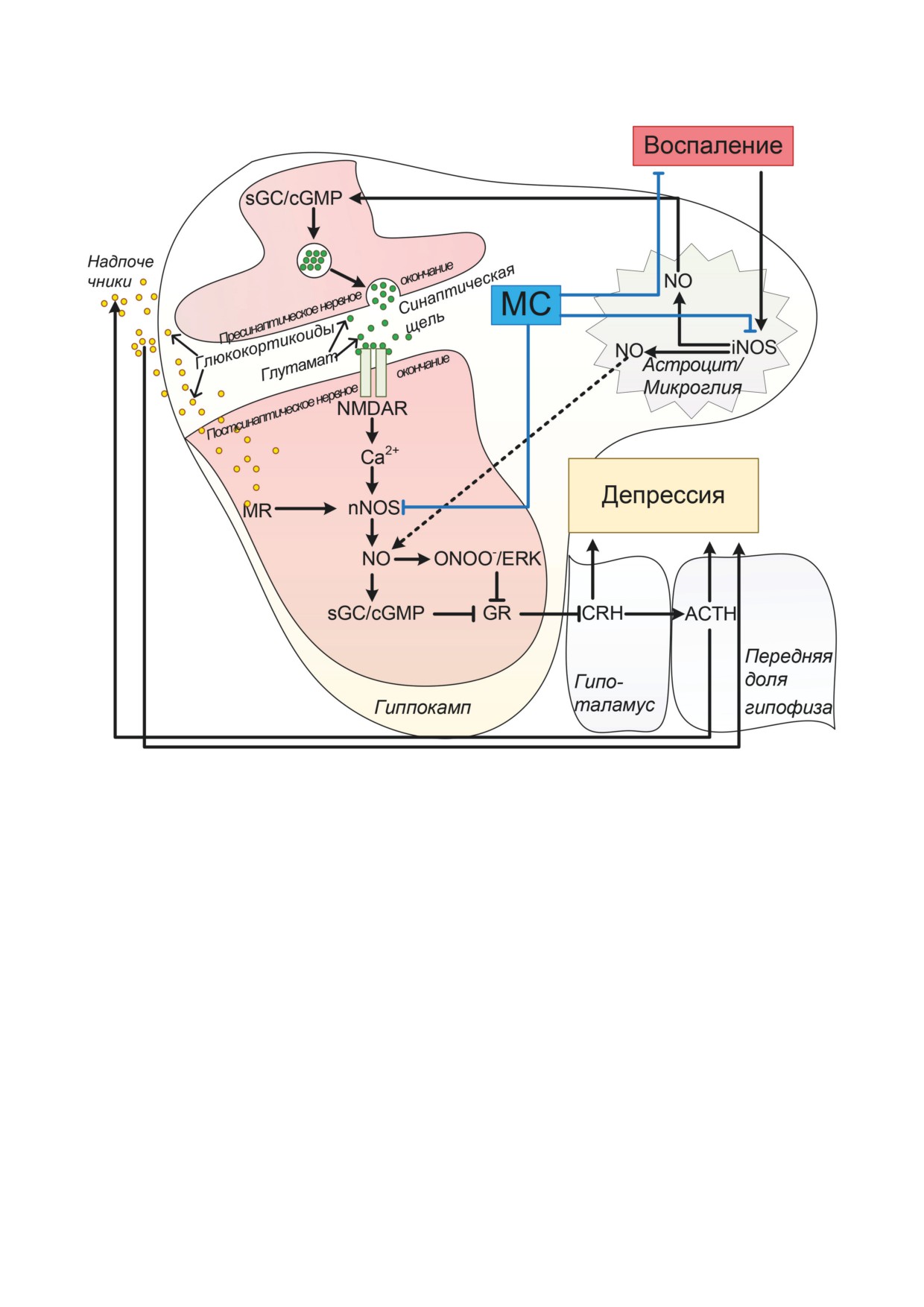
Рис. 7. Влияние МС на iNOS и nNOS способствует блокированию активности sGC/cGMP, снижению гипоталамиче-
ской секреции CRH, секреции глюкокортикоидов и препятствует развитию депрессивного состояния
го поведения, так как в нормальных условиях их
концентрациях: до 1 мM - при введении через
низкий уровень поддерживается отрицательной
микродиализный зонд [135] и 100 мг/кг - при
обратной связью [131].
внутрибрюшинных инъекциях [136]. Хотя по-
Фармакологическое ингибирование NOS
следующие исследования показали, что MС
может снижать показатели депрессивноподоб-
уже в концентрации 3 мг/кг (внутривенная
ного поведения на животных моделях [129]
инъекция) специфически ингибирует iNOS
(рис. 7). Mayer et al. [132] показали, что in vitro
при липополисахарид-индуцированном стрес-
MС полностью ингибирует NOS при кон-
се [137]. В принципе, стоит отметить, что до-
центрации 30 мкМ, а также ингибирует sGC,
зозависимость эффекта МС на ингибирование
но в меньшей степени
(50% ингибирова-
NOS нуждается в дальнейшем изучении, по-
ния - при 60 мкМ и максимальное ингиби-
тому что высокие концентрации МС сами по
рование 72% - при 1 мМ), из-за чего авторы
себе могут быть токсичны.
сделали вывод, что МС нельзя считать мощ-
ным и селективным ингибитор NOS, хотя ра-
нее было показано, что MС ингибирует про-
ЗАКЛЮЧЕНИЕ
дукцию GMP [133]. Позже было показано
ингибирующее действие МС на NOS и sGC не
Итак, МС является плейотропным препа-
только in vitro [134], но и in vivo [135, 136]. Од-
ратом, который может одновременно действо-
нако МС использовался в достаточно высоких
вать на несколько сигнальных путей, которые,
БИОХИМИЯ том 87 вып. 9 2022
НЕЙРОПРОТЕКТОРНЫЙ ЭФФЕКТ МЕТИЛЕНОВОГО СИНЕГО
1175
как правило, между собой крайне тесно пе-
Вклад авторов. А.П. Гуреев
- написа-
реплетаются. Множественные эффекты МС
ние текста, И.С. Садовникова
- поиск и
крайне важно учитывать при проведении кли-
аннотация статей, подготовка рисунков,
нических испытаний. Кроме того, МС обла-
В.Н. Попов - концепция и руководство.
дает крайне интересной фармакодинамикой,
Финансирование. Работа выполнена при
которая зачастую не учитывалась при дизайне
финансовой поддержке Министерства науки
некоторых клинических испытаний и послу-
и Высшего образования Российской Федера-
жила причиной их неудачи [18]. Данный об-
ции в рамках Государственного задания уни-
зор направлен на то, чтобы обобщить и в бу-
верситетам в области научной деятельности
дущих исследованиях учитывать максимально
на 2020-2022 гг. (проект № FZGU-2020-0044),
все возможные эффекты МС при воздействии
стипендии Президента Российской Федера-
на ЦНС, принимать во внимание переплетения
ции молодым учёным и аспирантам (проект
между апоптотическими путями, про/проти-
№ СП-2802.2021.4), гранта Президента Рос-
вовоспалительными процессами, сигнальны-
сийской Федерации для государственной под-
ми каскадами, которые регулируют митохон-
держки молодых российских учёных и по го-
дриальный биогенез, антиоксидатную защиту,
сударственной поддержке ведущих научных
уровни нейромедиаторов и неправильно свёр-
школ (НШ-1375.2022.5).
нутых и агрегированных белков. Глубокое и
Конфликт интересов. Авторы заявляют об
всестороннее понимание всех возможных эпи-
отсутствии конфликта интересов.
генетических, биохимических и физиологиче-
Соблюдение этических норм. Настоящая
ских последствий употребления МС позволит
статья не содержит описания каких-либо ис-
более корректно планировать эксперименты
следований с участием людей или животных в
для оценки его нейропротекторных свойств.
качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1. O’Neil, M. J.
(2013) The Merck Index: An
8. Bodoni, P. (1899) Dell’azione sedativa del bleu di
Encyclopedia of Chemicals, Drugs, and Biologicals,
metilene in varie forme di psicosi [in Italian], Clin.
Cambridge, UK, Royal Society of Chemistry,
Med. Ital., pp. 217-222.
England.
9. Ohlow, M. J., and Moosmann, B.
(2011)
2. Deutsches Reich Patent no. 1886, December 15,
Phenothiazine: the seven lives of pharmacology’s
1877 Badische Anilin- und Sodafabrik
[BASF]
first lead structure, Drug Discov. Today, 16, 119-131,
(Mannheim, Germany), “Verfahren zur Darstellung
doi: 10.1016/j.drudis.2011.01.001.
blauer Farbstoffe aus Dimethylanilin und anderen
10. Allexsaht, W. J. (1938) The use of methylene blue in
tertiaren aromatischen Monaminen” (Method for
the treatment of catatonic dementia praecox patients,
preparation of blue dyes from dimethylaniline and
Psychiatric Quarterly, 12, 245-252.
other tertiary aromatic monoamines), Deutsches
11. Kajdi, L., and Taylor, C. V. (1938) The use of
Reich Patent no. 1886 (December 15, 1877).
intravenous methylene blue in status convulsivus,
3. Berneth, H. (2008) Azine dyes. In: Ullmann’s Encyclo-
Am. J. Insanity,
94,
1369-1376, doi:
10.1176/
pedia of Industrial Chemistry, Weinheim, Germany.
ajp.94.6.1369.
4. Delport, A., Harvey, B. H., Petzer, A., and Petzer,
12. Naylor, G. J., and Smith, A. H. (1981) Vanadium:
J. P.
(2017) The monoamine oxidase inhibition
a possible aetiological factor in manic depressive
properties of selected structural analogues of
illness, Psychol. Med., 11, 249-256, doi: 10.1017/
methylene blue, Toxicol. Appl. Pharmacol., 325, 1-8,
s0033291700052065.
doi: 10.1016/j.taap.2017.03.026.
13. Naylor, G. J., Martin, B., Hopwood, S. E., and
5. Gaudette, N. F., and Lodge, J. W.
(2005)
Watson, Y. (1986) A two-year double-blind crossover
Determination of methylene blue and leucomethylene
trial of the prophylactic effect of methylene blue in
blue in male and female Fischer 344 rat urine and
manic depressive psychosis, Biol. Psychiatry,
21,
B6C3F1 mouse urine, J. Anal. Toxicol., 29, 28-33,
915-920, doi: 10.1016/0006-3223(86)90265-9.
doi: 10.1093/jat/29.1.28.
14. Martinez, J. L., Jensen, R. A., Vasquez, B. J.,
6. Schirmer, R. H., Adler, H., Pickhardt, M., and
McGuinness, T., and McGaugh, J. L.
(1978)
Mandelkow, E. (2011) “Lest we forget you - methylene
Methylene blue alters retention of inhibitory
blue...”, Neurobiol. Aging, 32, 2325.e7-2325.e2.325E16,
avoidance responses, Psychobiology,
6,
387-390,
doi: 10.1016/j.neurobiolaging.2010.12.012.
doi: 10.3758/BF03326744.
7. Ehrlich, P., and Leppmann, A.,
(1890) Ueber
15. Wischik, C. M., Edwards, P. C., Lai, R. Y., Roth, M.,
schmerzstillende Wirkung des Methylenblau, Dtsch.
and Harrington, C. R. (1996) Selective inhibition
Med. Wochenschr., 16, 493-494.
of Alzheimer disease-like tau aggregation by
БИОХИМИЯ том 87 вып. 9 2022
1176
ГУРЕЕВ и др.
phenothiazines, Proc. Natl. Acad. Sci. USA, 93, 11213-
improvement effects of methylene blue in global
11218, doi: 10.1073/pnas.93.20.11213.
cerebral ischemia, Mol. Neurobiol., 53, 5344-5355,
16. Wischik, C. M., Staff, R. T., Wischik, D. J., Bentham,
doi: 10.1007/s12035-015-9455-0.
P., Murray, A. D., et al. (2015) Tau aggregation
27. Li, L., Yang, R., Li, P., Lu, H., Hao, J., et al.
inhibitor therapy: an exploratory phase 2 study in mild
(2018) Combination treatment with methylene
or moderate Alzheimer’s disease, J. Alzheimer’s Dis.,
blue and hypothermia in global cerebral isch-
44, 705-720, doi: 10.3233/JAD-142874.
emia, Mol. Neurobiol., 55, 2042-2055, doi: 10.1007/
17. Gauthier, S., Feldman, H. H., Schneider, L. S.,
s12035-017-0470-1.
Wilcock, G. K., Frisoni, G. B., et al. (2016) Efficacy
28. Lin, Z. H., Wang, S. Y., Chen, L. L., Zhuang, J. Y.,
and safety of tau-aggregation inhibitor therapy in
Ke, Q. F., et al. (2017) Methylene blue mitigates acute
patients with mild or moderate Alzheimer’s disease: a
neuroinflammation after spinal cord injury through
randomised, controlled, double-blind, parallel-arm,
inhibiting NLRP3 inflammasome activation in
phase 3 trial, Lancet, 388, 2873-2884, doi: 10.1016/
microglia, Front. Cell Neurosci., 11, 391, doi: 10.3389/
S0140-6736(16)31275-2.
fncel.2017.00391.
18. Hashweh, N. N., Bartochowski, Z., Khoury, R.,
29. Dibaj, P., Zschüntzsch, J., Steffens, H., Scheffel,
and Grossberg, G. T.
(2020) An evaluation of
J., Göricke, B., et al. (2012) Influence of methy-
hydromethylthionine as a treatment option for
lene blue on microglia-induced inflammation and
Alzheimer’s disease, Expert Opin. Pharmacother., 21,
motor neuron degeneration in the SOD1(G93A)
619-627, doi: 10.1080/14656566.2020.1719066.
model for ALS, PLoS One, 7, e43963, doi: 10.1371/
19. Sontag, E. M., Lotz, G. P., Agrawal, N., Tran, A.,
journal.pone.0043963.
Aron, R., et al.
(2012) Methylene blue mod-
30. Zeevalk, G. D., Bernard, L. P., Song, C., Gluck,
ulates
huntingtin
aggregation
intermediates
M., and Ehrhart, J. (2005) Mitochondrial inhibition
and is protective in Huntington’s disease mod-
and oxidative stress: reciprocating players in
els, J. Neurosci.,
32,
11109-11119, doi:
10.1523/
neurodegeneration, Antioxid. Redox Signal., 7, 1117-
JNEUROSCI.0895-12.2012.
1139, doi: 10.1089/ars.2005.7.1117.
20. Heidari, R., Monnier, V., Martin, E., and Tricoire, H.
31. Andreyev, A. Y., Kushnareva, Y. E., and Starkov,
(2015) Methylene blue partially rescues heart de-
A. A. (2005) Mitochondrial metabolism of reactive
fects in a drosophila model of Huntington’s dis-
oxygen species, Biochemistry (Moscow), 70, 200-214,
ease, J. Huntington’s Dis., 4, 173-186, doi: 10.3233/
doi: 10.1007/s10541-005-0102-7.
JHD-140130.
32. Eubel, H., Heinemeyer, J., Sunderhaus, S.,
21. Bariotto-Dos-Santos, K., Padovan-Neto, F. E.,
and Braun, H. P.
(2004) Respiratory chain su-
Bortolanza, M., Dos-Santos-Pereira, M., Raisman-
percomplexes in
plant mitochondria, Plant
Vozari, R., et al. (2019) Repurposing an established
Physiol. Biochem.,
42,
937-942, doi:
10.1016/
drug: an emerging role for methylene blue in
j.plaphy.2004.09.010.
L-DOPA-induced dyskinesia, Eur. J. Neurosci., 49,
33. Cadenas, S.
(2018) Mitochondrial uncoupling,
869-882, doi: 10.1111/ejn.14079.
ROS generation and cardioprotection, Biochim.
22. Bhurtel, S., Katila, N., Neupane, S., Srivastav, S.,
Biophys. Acta Bioenerg., 1859, 940-950, doi: 10.1016/
Park, P. H., et al. (2018) Methylene blue protects
j.bbabio.2018.05.019.
dopaminergic neurons against MPTP-induced
34. Grundlingh, J., Dargan, P. I., El-Zanfaly, M., and
neurotoxicity
by
upregulating
brain-derived
Wood, D. M.
(2011)
2,4-dinitrophenol (DNP):
neurotrophic factor, Ann. NY Acad. Sci., 1431, 58-71,
a weight loss agent with significant acute toxicity
doi: 10.1111/nyas.13870.
and risk of death, J. Med. Toxicol., 7, 205-212,
23. Stelmashook, E. V., Genrikhs, E. E., Mukhaleva,
doi: 10.1007/s13181-011-0162-6.
E. V., Kapkaeva, M. R., Kondratenko, R. V., et al.
35. Wen, Y., Li, W., Poteet, E. C., Xie, L., Tan, C.,
(2019) Neuroprotective effects of methylene blue
et al.
(2011) Alternative mitochondrial electron
in vivo and in vitro, Bull. Exp. Biol. Med., 167, 455-
transfer as a novel strategy for neuroprotection,
459, doi: 10.1007/s10517-019-04548-3.
J. Biol. Chem.,
286,
16504-16515, doi:
10.1074/
24. Genrikhs, E. E., Stelmashook, E. V., Voronkov,
jbc.M110.208447.
D. N., Novikova, S. V., Alexandrova, O. P., et al.
36. Harrop, G. A., and Barron, E. S. (1928) Stud-
(2020) The delayed neuroprotective effect of
ies on blood cell metabolism: I. The effect of
methylene blue in experimental rat brain trauma,
methylene blue and other dyes upon the oxy-
Antioxidants, 9, 377, doi: 10.3390/antiox9050377.
gen consumption of mammalian and avian eryth-
25. Genrikhs, E. E., Stelmashook, E. V., Voronkov,
rocytes, J. Exp. Med., 48, 207-223, doi: 10.1084/
D. N., Novikova, S. V., Alexandrova, O. P., et al.
jem.48.2.207.
(2020) The single intravenous administration of
37. Barron, E. S., and Hoffman, L. A. (1930) The
methylene blue after traumatic brain injury diminishes
catalytic effect of dyes on the oxygen consumption
neurological deficit, blood-brain barrier disruption
of living cells, J. Gen. Physiol.,
13,
483-494,
and decrease in the expression of S100 protein
doi: 10.1085/jgp.13.4.483.
in rats, Brain Res.,
1740,
146854, doi:
10.1016/
38. Elliott, K. A., and Greig, M. E.
(1938)
j.brainres.2020.146854.
The distribution of the succinic oxidase system
26. Lu, Q., Tucker, D., Dong, Y., Zhao, N., and
in animal tissues, Biochem. J.,
32,
1407-1423,
Zhang, Q. (2016) Neuroprotective and functional
doi: 10.1042/bj0321407.
БИОХИМИЯ том 87 вып. 9 2022
НЕЙРОПРОТЕКТОРНЫЙ ЭФФЕКТ МЕТИЛЕНОВОГО СИНЕГО
1177
39. Slater, E. C. (1949) A respiratory catalyst required
of methylene blue and its derivatives, PLoS One, 7,
for the reduction of cytochrome c by cytochrome b,
e48279, doi: 10.1371/journal.pone.0048279.
Biochem. J., 45, 14-30, doi: 10.1042/bj0450014.
52. Mikulás, K., Komlódi, T., Földes, A., Sváb, G.,
40. Redfearn, E. R.
(1961) The role of ubiquinone
Horváth, G., et al. (2020) Bioenergetic Impairment
(coenzyme Q) and its homologues in mediating the
of triethylene glycol dimethacrylate- (TEGDMA-)
reduction of methylene blue by succinate in heart-
treated dental pulp stem cells (DPSCs) and isolated
muscle preparations, Biochim. Biophys. Acta, 53, 581-
brain mitochondria are amended by redox compound
583, doi: 10.1016/0006-3002(61)90220-7.
methylene blue, Materials, 13, 3472, doi: 10.3390/
41. Tönz, O. (1968) The congenital methemoglobinemias.
ma13163472.
Physiology and pathophysiology of the hemiglobin
53. Sváb, G., Kokas, M., Sipos, I., Ambrus, A., and
metabolism, Bibliotheca Haematologica, 28, 1-146.
Tretter, L.
(2021) Methylene blue bridges the
42. Visarius, T. M., Stucki, J. W., and Lauterburg, B. H.
inhibition and produces unusual respiratory changes
(1997) Stimulation of respiration by methylene blue
in complex III-inhibited mitochondria. Studies on
in rat liver mitochondria, FEBS Lett., 412, 157-160,
rats, mice and guinea pigs, Antioxidants, 10, 305,
doi: 10.1016/s0014-5793(97)00767-9.
doi: 10.3390/antiox10020305.
43. Gureev, A. P., Samoylova, N. A., Potanina, D. V., and
54. Irwin, M. H., Parameshwaran, K., and Pinkert, C. A.
Popov, V. N. (2021) Effect of methylene blue and its
(2013) Mouse models of mitochondrial complex I
metabolite - azure I - on bioenergetic parameters of
dysfunction, Int. J. Biochem. Cell Biol., 45, 34-40,
intact mice brain mitochondria, Biomed. Khim., 67,
doi: 10.1016/j.biocel.2012.08.009.
485-490, doi: 10.18097/PBMC20216706485.
55. Bahn, G., and Jo, D. G.
(2019) Therapeutic
44. Callaway, N. L., Riha, P. D., Wrubel, K. M.,
approaches to Alzheimer’s disease through
McCollum, D., and Gonzalez-Lima, F.
(2002)
modulation of NRF2, Neuromol. Med., 21, 1-11,
Methylene blue restores spatial memory retention
doi: 10.1007/s12017-018-08523-5.
impaired by an inhibitor of cytochrome oxidase
56. Taniguchi, S., Suzuki, N., Masuda, M., Hisanaga,
in rats, Neurosci. Lett., 332, 83-86, doi: 10.1016/
S., Iwatsubo, T., et al. (2005) Inhibition of heparin-
s0304-3940(02)00827-3.
induced tau filament formation by phenothiazines,
45. Tretter, L., Horvath, G., Hölgyesi, A., Essek, F.,
polyphenols, and porphyrins, J. Biol. Chem., 280,
and Adam-Vizi, V.
(2014) Enhanced hydrogen
7614-7623, doi: 10.1074/jbc.M408714200.
peroxide generation
accompanies the beneficial
57. Hattori, M., Sugino, E., Minoura, K., In, Y.,
bioenergetic effects of methylene blue in isolated brain
Sumida, M., et al.
(2008) Different inhibitory
mitochondria, Free Radic. Biol. Med., 77, 317-330,
response of cyanidin and methylene blue for filament
doi: 10.1016/j.freeradbiomed.2014.09.024.
formation of tau microtubule-binding domain,
46. Gureev, A. P., Syromyatnikov, M. Y., Gorbacheva,
Biochem. Biophys. Res. Commun.,
374,
158-163,
T. M., Starkov, A. A., and Popov, V. N.
(2016)
doi: 10.1016/j.bbrc.2008.07.001.
Methylene blue improves sensorimotor pheno-
58. Stack, C., Jainuddin, S., Elipenahli, C., Gerges, M.,
type and decreases anxiety in parallel with acti-
Starkova, N., et al.
(2014) Methylene blue
vating brain mitochondria biogenesis in mid-age
upregulates Nrf2/ARE genes and prevents tau-related
mice, Neurosci. Res.,
113,
19-27, doi:
10.1016/
neurotoxicity, Hum. Mol. Genet.,
23,
3716-3732,
j.neures.2016.07.006.
doi: 10.1093/hmg/ddu080.
47. Gureev, A. P., Shaforostova, E. A., Popov, V. N.,
59. Hochgräfe, K., Sydow, A., Matenia, D., Cadinu, D.,
and Starkov, A. A.
(2019) Methylene blue does
Könen, S., et al.
(2015) Preventive methylene
not bypass Complex III antimycin block in mouse
blue treatment preserves cognition in mice ex-
brain mitochondria, FEBS Lett.,
593,
499-503,
pressing full-length pro-aggregant human Tau,
doi: 10.1002/1873-3468.13332.
Acta Neuropathol. Commun.,
3,
25, doi:
10.1186/
48. Gureev, A. P., Shaforostova, E. A., Laver, D. A.,
s40478-015-0204-4.
Khorolskaya, V. G., Syromyatnikov, M. Y., et al.
60. Necula, M., Breydo, L., Milton, S., Kayed, R.,
(2019)
Methylene blue elicits non-genotoxic
van der Veer, W. E., et al. (2007) Methylene blue
H2O2 production and protects brain mitochondria
inhibits amyloid Abeta oligomerization by promoting
from rotenone toxicity, J. Appl. Biomed., 17, 107-114,
fibrillization,
Biochemistry,
46,
8850-8860,
doi: 10.32725/jab.2019.008.
doi: 10.1021/bi700411k.
49. Atamna, H., Atamna, W., Al-Eyd, G., Shanower,
61. Lee, B. I., Suh, Y. S., Chung, Y. J., Yu, K., and
G., and Dhahbi, J. M. (2015) Combined activation
Park, C. B. (2017) Shedding light on Alzheimer’s
of the energy and cellular-defense pathways may
β-amyloidosis: photosensitized methylene blue
explain the potent anti-senescence activity of
inhibits self-assembly of β-amyloid peptides and
methylene blue, Redox Biol., 6, 426-435, doi: 10.1016/
disintegrates their aggregates, Sci. Rep.,
7,
7523,
j.redox.2015.09.004.
doi: 10.1038/s41598-017-07581-2.
50. Zhang, X., Rojas, J. C., and Gonzalez-Lima, F.
62. Berrocal, M., Corbacho, I., Gutierrez-Merino, C.,
(2006) Methylene blue prevents neurodegeneration
and Mata, A. M. (2018) Methylene blue activates
caused by rotenone in the retina, Neurotox. Res., 9,
the PMCA activity and cross-interacts with amyloid
47-57, doi: 10.1007/BF03033307.
β-peptide, blocking Aβ-mediated PMCA inhibition,
51. Poteet, E., Winters, A., Yan, L. J., Shufelt, K.,
Neuropharmacology,
139,
163-172, doi:
10.1016/j.
Green, K. N., et al. (2012) Neuroprotective actions
neuropharm.2018.07.012.
2
БИОХИМИЯ том 87 вып. 9 2022
1178
ГУРЕЕВ и др.
63. Berrocal, M., Caballero-Bermejo, M., Gutierrez-
Chem. Biol. Interact., 329, 109222, doi: 10.1016/
Merino, C., and Mata, A. M. (2019) Methylene
j.cbi.2020.109222.
blue blocks and reverses the inhibitory effect of
76. Gureev, A. P., Shaforostova, E. A., and Popov, V. N.
Tau on PMCA function, Int. J. Mol. Sci., 20, 3521,
(2019) Regulation of mitochondrial biogenesis as a
doi: 10.3390/ijms20143521.
way for active longevity: interaction between the Nrf2
64. Illarioshkin, S. N., Klyushnikov, S. A., Vigont, V. A.,
and PGC-1α signaling pathways, Front. Genet., 10,
Seliverstov, Y. A., and Kaznacheyeva, E. V. (2018)
435, doi: 10.3389/fgene.2019.00435.
Molecular pathogenesis in Huntington’s disease,
77. Fernandez-Marcos, P. J., and Auwerx, J.
(2011)
Biochemistry (Moscow), 83, 1030-1039, doi: 10.1134/
Regulation of PGC-1α, a nodal regulator of
S0006297918090043.
mitochondrial biogenesis, Am. J. Clin. Nutr., 93,
65. Cavaliere, P., Torrent, J., Prigent, S., Granata,
884S-890S, doi: 10.3945/ajcn.110.001917.
V., Pauwels, K., et al. (2013) Binding of methy-
78. Islam, H., Hood, D. A., and Gurd, B. J. (2020)
lene blue to a surface cleft inhibits the oligom-
Looking beyond PGC-1α: emerging regulators of
erization and fibrillization of prion protein, Bio-
exercise-induced skeletal muscle mitochondrial
chim. Biophys. Acta,
1832,
20-28, doi:
10.1016/
biogenesis and their activation by dietary compounds,
j.bbadis.2012.09.005.
Appl. Physiol. Nutr. Metab., 45, 11-23, doi: 10.1139/
66. Paré, B., Lehmann, M., Beaudin, M., Nordström, U.,
apnm-2019-0069.
Saikali, S., et al. (2018) Misfolded SOD1 pathology
79. Xie, L., Li, W., Winters, A., Yuan, F., Jin, K., et al.
in sporadic amyotrophic lateral sclerosis, Sci. Rep., 8,
(2013) Methylene blue induces macroautophagy
14223, doi: 10.1038/s41598-018-31773-z.
through
5′ adenosine monophosphate-activated
67. Yamashita, M., Nonaka, T., Arai, T., Kametani, F.,
protein kinase pathway to protect neurons from serum
Buchman, V. L., et al. (2009) Methylene blue and
deprivation, Front. Cell Neurosci., 7, 56, doi: 10.3389/
dimebon inhibit aggregation of TDP-43 in cellular
fncel.2013.00056.
models, FEBS Lett., 583, 2419-2424, doi: 10.1016/
80. Kroemer, G., Mariño, G., and Levine, B. (2010)
j.febslet.2009.06.042.
Autophagy and the integrated stress response, Mol.
68. Musteikyte, G., Ziaunys, M., and Smirnovas, V.
Cell, 40, 280-293, doi: 10.1016/j.molcel.2010.09.023.
(2020) Methylene blue inhibits nucleation and
81. Telli, M. L., Nagata, H., Wapnir, I., Acharya,
elongation of SOD1 amyloid fibrils, PeerJ, 8, e9719,
C. R., Zablotsky, K., et al. (2021) Intratumoral
doi: 10.7717/peerj.9719.
plasmid IL12 expands CD8+ T cells and induces
69. Vaccaro, A., Patten, S. A., Ciura, S., Maios, C.,
a CXCR3 gene signature in triple-negative breast
Therrien, M., et al. (2012) Methylene blue protects
tumors that sensitizes patients to anti-PD-1 therapy,
against TDP-43 and FUS neuronal toxicity in
Clin. Cancer Res.,
27,
2481-2493, doi:
10.1158/
C. elegans and D. rerio, PLoS One,
7, e42117,
1078-0432.CCR-20-3944.
doi: 10.1371/journal.pone.0042117.
82. Wong, E., and Cuervo, A. M. (2010) Autophagy gone
70. Vaccaro, A., Patten, S. A., Aggad, D., Julien, C.,
awry in neurodegenerative diseases, Nat. Neurosci.,
Maios, C., et al. (2013) Pharmacological reduction of
13, 805-811, doi: 10.1038/nn.2575.
ER stress protects against TDP-43 neuronal toxicity
83. Medina, D. X., Caccamo, A., and Oddo, S. (2011)
in vivo, Neurobiol. Dis., 55, 64-75, doi: 10.1016/
Methylene blue reduces aβ levels and rescues
j.nbd.2013.03.015.
early cognitive deficit by increasing proteasome
71. Dinkova-Kostova, A. T., and Abramov, A. Y. (2015)
activity, Brain Pathol., 21, 140-149, doi: 10.1111/
The emerging role of Nrf2 in mitochondrial function,
j.1750-3639.2010.00430.x.
Free Radic. Biol. Med., 88, 179-188, doi: 10.1016/
84. Congdon, E. E., Wu, J. W., Myeku, N., Figueroa,
j.freeradbiomed.2015.04.036.
Y. H., Herman, M., et al.
(2012) Methylthi-
72. Hayes, J. D., Chowdhry, S., Dinkova-Kostova,
oninium chloride (methylene blue) induces au-
A. T., and Sutherland, C.
(2015) Dual regula-
tophagy and attenuates tauopathy in vitro and
tion of transcription factor Nrf2 by Keap1 and
in vivo, Autophagy,
8,
609-622, doi:
10.4161/
by the combined actions of β-TrCP and GSK-3,
auto.19048.
Biochem. Soc. Trans., 43, 611-620, doi: 10.1042/
85. Jiang, Z., Watts, L. T., Huang, S., Shen, Q.,
BST20150011.
Rodriguez, P., et al. (2015) The effects of methylene
73. Zenkov, N. K., Kozhin, P. M., Chechushkov,
blue on autophagy and apoptosis in MRI-defined
A. V., Martinovich, G. G., Kandalintseva,
normal tissue, ischemic penumbra and ischemic
N. V., et al.
(2017) Mazes of Nrf2 regulation,
core, PLoS One,
10, e0131929, doi:
10.1371/
Biochemistry (Moscow), 82, 556-564, doi: 10.1134/
journal.pone.0131929.
S0006297917050030.
86. Zhao, M., Liang, F., Xu, H., Yan, W., and Zhang, J.
74. El Sayed, N. S., and Sayed, A. S. (2019) Pro-
(2016) Methylene blue exerts a neuroprotective
tective effect of methylene blue on TNBS-in-
effect against traumatic brain injury by promoting
duced colitis in rats mediated through the mod-
autophagy and inhibiting microglial activation, Mol.
ulation of inflammatory and apoptotic signalling
Med. Rep., 13, 13-20, doi: 10.3892/mmr.2015.4551.
pathways, Arch. Toxicol., 93, 2927-2942, doi: 10.1007/
87. Di, Y., He, Y. L., Zhao, T., Huang, X., Wu, K. W.,
s00204-019-02548-w.
et al. (2015) Methylene blue reduces acute cerebral
75. Kaur, S., and Benov, L. T. (2020) Methylene blue
ischemic injury via the induction of mitophagy, Mol.
induces the soxRS regulon of Escherichia coli,
Med., 21, 420-429, doi: 10.2119/molmed.2015.00038.
БИОХИМИЯ том 87 вып. 9 2022
НЕЙРОПРОТЕКТОРНЫЙ ЭФФЕКТ МЕТИЛЕНОВОГО СИНЕГО
1179
88. Gureev, A. P., Sadovnikova, I. S., Starkov, N. N.,
methylene blue alleviates efavirenz/isoniazid-induced
Starkov, A. A., and Popov, V. N. (2020) p62-Nrf2-p62
oxidant stress and mitochondria-mediated cell death
mitophagy regulatory loop as a target for preventive
in mouse hepatocytes, Redox Biol.,
2,
599-609,
therapy of neurodegenerative diseases, Brain Sci., 10,
doi: 10.1016/j.redox.2014.03.003.
847, doi: 10.3390/brainsci10110847.
103. Abdelkader, N. F., Farid, H. A., Youness, E. R.,
89. Murata, H., Takamatsu, H., Liu, S., Kataoka, K.,
Abdel-Salam, O., and Zaki, H. F. (2020) The role
Huh, N. H., et al. (2015) NRF2 regulates PINK1
of KATP channel blockade and activation in the
expression under oxidative stress conditions, PLoS
protection against neurodegeneration in the rotenone
One, 10, e0142438, doi: 10.1371/journal.pone.0142438.
model of Parkinson’s disease, Life Sci., 257, 118070,
90. Dos Santos, A. F., Terra, L. F., Wailemann, R. A.,
doi: 10.1016/j.lfs.2020.118070.
Oliveira, T. C., Gomes, V. M., et al. (2017) Methylene
104. Pakavathkumar, P., Sharma, G., Kaushal, V.,
blue photodynamic therapy induces selective and
Foveau, B., and LeBlanc, A. C.
(2015) Methy-
massive cell death in human breast cancer cells, BMC
lene blue inhibits caspases by oxidation of the cat-
Cancer, 17, 194, doi: 10.1186/s12885-017-3179-7.
alytic cysteine, Sci. Rep., 5, 13730, doi: 10.1038/
91. Lucky, S. S., Soo, K. C., and Zhang, Y.
(2015)
srep13730.
Nanoparticles in photodynamic therapy, Chem. Rev.,
105. Zhou, L., Flores, J., Noël, A., Beauchet, O.,
115, 1990-2042, doi: 10.1021/cr5004198.
Sjöström, P. J., et al. (2019) Methylene blue inhibits
92. Tardivo, J. P., Del Giglio, A., de Oliveira, C. S.,
Caspase-6 activity, and reverses Caspase-6-induced
Gabrielli, D. S., Junqueira, H. C., et al.
(2005)
cognitive impairment and neuroinflammation in
Methylene blue in photodynamic therapy:
aged mice, Acta Neuropathol. Commun.,
7,
210,
from basic mechanisms to clinical applications,
doi: 10.1186/s40478-019-0856-6.
Photodiagn. Photodyn. Ther., 2, 175-191, doi: 10.1016/
106. Ahn, H., Kang, S. G., Yoon, S. I., Ko, H. J., Kim,
S1572-1000(05)00097-9.
P. H., et al. (2017) Methylene blue inhibits NLRP3,
93. Eimon, P. M., Kratz, E., Varfolomeev, E., Hymowitz,
NLRC4, AIM2, and non-canonical inflammasome
S. G., Stern, H., et al. (2006) Delineation of the
activation, Sci. Rep.,
7,
12409, doi:
10.1038/
cell-extrinsic apoptosis pathway in the zebrafish,
s41598-017-12635-6.
Cell Death Differ.,
13,
1619-1630, doi:
10.1038/
107. Amor, S., Puentes, F., Baker, D., and van der
sj.cdd.4402015.
Valk, P. (2010) Inflammation in neurodegenerative
94. Bellail, A. C., Tse, M. C., Song, J. H., Phuphanich,
diseases, Immunology, 129, 154-169, doi: 10.1111/
S., Olson, J. J., et al. (2010) DR5-mediated DISC
j.1365-2567.2009.03225.x.
controls caspase-8 cleavage and initiation of apoptosis
108. Chen, L., Deng, H., Cui, H., Fang, J., Zuo, Z., et al.
in human glioblastomas, J. Cell Mol. Med., 14, 1303-
(2017) Inflammatory responses and inflammation-
1317, doi: 10.1111/j.1582-4934.2009.00777.x.
associated diseases in organs, Oncotarget, 9, 7204-
95. Garrido, C., Galluzzi, L., Brunet, M., Puig, P. E.,
7218, doi: 10.18632/oncotarget.23208.
Didelot, C., et al. (2006) Mechanisms of cytochrome
109. Zelová, H., and Hošek, J. (2013) TNF-α signalling
c release from mitochondria, Cell Death Differ., 13,
and inflammation: interactions between old
1423-1433, doi: 10.1038/sj.cdd.4401950.
acquaintances, Inflamm. Res.,
62,
641-651,
96. Wu, C. C., and Bratton, S. B. (2013) Regulation of the
doi: 10.1007/s00011-013-0633-0.
intrinsic apoptosis pathway by reactive oxygen species,
110. Monaco, C., Andreakos, E., Kiriakidis, S., Mauri, C.,
Antioxid Redox Signal., 19, 546-558, doi: 10.1089/
Bicknell, C., et al.
(2004) Canonical pathway
ars.2012.4905.
of nuclear factor kappa B activation selectively
97. Roy, S., and Nicholson, D. W. (2000) Cross-talk in
regulates proinflammatory and prothrombotic
cell death signaling, J. Exp. Med., 192, F21-F25.
responses in human atherosclerosis, Proc. Natl.
98. Connolly, P. F., Jäger, R., and Fearnhead, H. O.
Acad. Sci. USA,
101,
5634-5639, doi:
10.1073/
(2014) New roles for old enzymes: killer caspases as
pnas.0401060101.
the engine of cell behavior changes, Front. Physiol., 5,
111. Liu, J. J., Lu, L., Hu, F. Q., Yuan, H., Xu, Q.,
149, doi: 10.3389/fphys.2014.00149.
et al.
(2018) Methylene blue attenuates renal
99. Radi, E., Formichi, P., Battisti, C., and
ischemia-reperfusion injury by negative regula-
Federico, A. (2014) Apoptosis and oxidative stress in
tion of NLRP3 signaling pathway, Eur. Rev. Med.
neurodegenerative diseases, J. Alzheimer’s Dis., 42,
Pharmacol. Sci.,
22,
2847-2853, doi:
10.26355/
S125-S152, doi: 10.3233/JAD-132738.
eurrev_201805_14986.
100. Chen, C., Zhou, F., Zeng, L., Jiang, Z., and Hu, Z.
112. Zheng, J., and Li, Q. (2019) Methylene blue reg-
(2019) Methylene blue offers neuroprotection after
ulates inflammatory response in osteoarthri-
intracerebral hemorrhage in rats through the PI3K/
tis by noncoding long chain RNA CILinc02,
Akt/GSK3β signaling pathway, J. Cell Physiol., 234,
J. Cell Biochem.,
120,
3331-3338, doi:
10.1002/
5304-5318, doi: 10.1002/jcp.27339.
jcb.27602.
101. Abdel-Salam, O., Omara, E., Youness, E., Khadrawy,
113. Wang, S. W., and Sun, Y. M. (2014) The IL-6/JAK/
Y., Mohammed, N., et al. (2014) Rotenone-induced
STAT3 pathway: potential therapeutic strategies in
nigrostriatal toxicity is reduced by methylene blue,
treating colorectal cancer (review), Int. J. Oncol., 44,
J. Neurorestoratol., 2, 65-80, doi: 10.2147/JN.S49207.
1032-1040, doi: 10.3892/ijo.2014.2259.
102. Lee, K. K., and Boelsterli, U. A. (2014) Bypassing the
114. Lobo-Silva, D., Carriche, G. M., Castro, A. G.,
compromised mitochondrial electron transport with
Roque, S., and Saraiva, M.
(2016) Balancing
БИОХИМИЯ том 87 вып. 9 2022
2*
1180
ГУРЕЕВ и др.
the immune response in the brain: IL-10 and its
H. C. (2017) Role of nitric oxide synthase in the
regulation, J. Neuroinflamm., 13, 297, doi: 10.1186/
function of the central nervous system under normal
s12974-016-0763-8.
and infectious conditions, Nitric Oxide Synthase,
115. Riley, J. K., Takeda, K., Akira, S., and Schreiber,
IntechOpen, doi: 10.5772/67816.
R. D. (1999) Interleukin-10 receptor signaling through
127. Czapski, G. A., Cakala, M., Chalimoniuk, M.,
the JAK-STAT pathway. Requirement for two distinct
Gajkowska, B., and Strosznajder, J. B. (2007) Role
receptor-derived signals for anti-inflammatory action,
of nitric oxide in the brain during lipopolysaccharide-
J. Biol. Chem.,
274,
16513-16521, doi:
10.1074/
evoked systemic inflammation, J. Neurosci. Res., 85,
jbc.274.23.16513.
1694-1703, doi: 10.1002/jnr.21294.
116. Shih, J. C., Chen, K., and Ridd, M. J. (1999) Role of
128. Zhou, X. Y., Zhang, F., Ying, C. J., Chen, J.,
MAO A and B in neurotransmitter metabolism and
Chen, L., et al. (2017) Inhibition of iNOS alleviates
behavior, Pol. J. Pharmacol., 51, 25-29.
cognitive deficits and depression in diabetic mice
117. Godar, S. C., Bortolato, M., Frau, R., Dousti,
through downregulating the NO/sGC/cGMP/
M., Chen, K., et al. (2011) Maladaptive defensive
PKG signal pathway, Behav. Brain Res., 322, 70-82,
behaviours in monoamine oxidase A-deficient
doi: 10.1016/j.bbr.2016.12.046.
mice, Int. J. Neuropsychopharmacol., 14, 1195-1207,
129. Chen, H. J., Spiers, J. G., Sernia, C., and Lavidis,
doi: 10.1017/S1461145710001483.
N. A. (2015) Response of the nitrergic system to
118. Watson, C., Bates, J., and Franczak, R. (2009) Sero-
activation of the neuroendocrine stress axis, Front.
tonin regulation by astrocytes, FASEB J., 23, 790.2,
Neurosci., 9, 3, doi: 10.3389/fnins.2015.00003.
doi: 10.1096/fasebj.23.1_supplement.790.2.
130. Zhou, Q. G., Zhu, L. J., Chen, C., Wu, H. Y., Luo,
119. Paudel, P., Park, S. E., Seong, S. H., Jung, H. A.,
C. X., et al. (2011) Hippocampal neuronal nitric
and Choi, J. S. (2019) Novel Diels-Alder type ad-
oxide synthase mediates the stress-related depressive
ducts from morus alba root bark targeting hu-
behaviors of glucocorticoids by downregulating
man monoamine oxidase and dopaminergic re-
glucocorticoid receptor, J. Neurosci., 31, 7579-7590,
ceptors for the management of neurodegenerative
doi: 10.1523/JNEUROSCI.0004-11.2011.
diseases, Int. J. Mol. Sci., 20, 6232, doi: 10.3390/
131. Gragnoli,
C.
(2014)
Hypothesis of
the
ijms20246232.
neuroendocrine cortisol pathway gene role in the
120. Ordway, G. A., Farley, J. T., Dilley, G. E.,
comorbidity of depression, type
2 diabetes, and
Overholser, J. C., Meltzer, H. Y., et al.
(1999)
metabolic syndrome, Appl. Clin. Genet., 7, 43-53,
Quantitative distribution of monoamine oxidase A
doi: 10.2147/TACG.S39993.
in brainstem monoamine nuclei is normal in major
132. Mayer, B., Brunner, F., and Schmidt, K.
(1993)
depression, Brain Res., 847, 71-79, doi: 10.1016/
Inhibition of nitric oxide synthesis by methylene
s0006-8993(99)02043-0.
blue, Biochem. Pharmacol., 45, 367-374, doi: 10.1016/
121. Aeschlimann, C., Cerny, T., and Küpfer, A.
0006-2952(93)90072-5.
(1996) Inhibition of (mono)amine oxidase ac-
133. Ignarro, L. J., Burke, T. M., Wood, K. S., Wolin,
tivity and prevention of ifosfamide encephalopa-
M. S., and Kadowitz, P. J.
(1984) Association
thy by methylene blue, Drug Metabol. Dispos., 24,
between cyclic GMP accumulation and acetylcholine-
1336-1339.
elicited relaxation of bovine intrapulmonary artery,
122. Ramsay, R. R., Dunford, C., and Gillman, P. K.
J. Pharmacol. Exp. Ther., 228, 682-690.
(2007) Methylene blue and serotonin toxicity:
134. Luo, D., Das, S., and Vincent, S. R. (1995) Effects
inhibition of monoamine oxidase A (MAO A)
of methylene blue and LY83583 on neuronal
confirms a theoretical prediction, Br. J. Pharmacol.,
nitric oxide synthase and NADPH-diaphorase,
152, 946-951, doi: 10.1038/sj.bjp.0707430.
Eur. J. Pharmacol.,
290,
247-251, doi:
10.1016/
123. Harvey, B. H., Duvenhage, I., Viljoen, F., Scheepers,
0922-4106(95)00084-4.
N., Malan, S. F., et al. (2010) Role of monoamine
135. Volke, V., Wegener, G., Vasar, E., and Rosenberg, R.
oxidase, nitric oxide synthase and regional brain
(1999) Methylene blue inhibits hippocampal nitric
monoamines in the antidepressant-like effects of
oxide synthase activity in vivo, Brain Res., 826, 303-
methylene blue and selected structural analogues,
305, doi: 10.1016/s0006-8993(99)01253-6.
Biochem. Pharmacol., 80, 1580-1591, doi: 10.1016/
136. Deutsch, S., Rosse, R., Paul, S., Tomasino, V.,
j.bcp.2010.07.037.
Koetzner, L., et al.
(1996)
7-Nitroindazole and
124. Delport, A., Harvey, B. H., Petzer, A., and Petzer,
methylene blue, inhibitors of neuronal nitric ox-
J. P. (2014) Azure B and a synthetic structural
ide synthase and NO-stimulated guanylate cyclase,
analogue of methylene blue, ethylthioninium chloride,
block MK-801-elicited behaviors in mice, Neu-
present with antidepressant-like properties, Life Sci.,
ropsychopharmacology,
15,
37-43, doi:
10.1016/
117, 56-66, doi: 10.1016/j.lfs.2014.10.005.
0893-133X(95)00153-5.
125. Petzer, A., Harvey, B. H., Wegener, G., and Petzer,
137. Lomniczi, A., Cebral, E., Canteros, G., McCann,
J. P. (2012) Azure B, a metabolite of methylene blue,
S. M., and Rettori, V. (2000) Methylene blue in-
is a high-potency, reversible inhibitor of monoamine
hibits the increase of inducible nitric oxide syn-
oxidase, Toxicol. Appl. Pharmacol., 258, 403-409,
thase activity induced by stress and lipopolysac-
doi: 10.1016/j.taap.2011.12.005.
charide in the medial basal hypothalamus of rats,
126. Reis, P. A., Gonçalves de Albuquerque, C. F., Maron-
Neuroimmunomodulation, 8, 122-127, doi: 10.1159/
Gutierrez, T., Silva, A. R, and de Castro Faria Neto,
000054271.
БИОХИМИЯ том 87 вып. 9 2022
НЕЙРОПРОТЕКТОРНЫЙ ЭФФЕКТ МЕТИЛЕНОВОГО СИНЕГО
1181
MOLECULAR MECHANISMS
OF THE NEUROPROTECTIVE EFFECT OF METHYLENE BLUE
Review
A. P. Gureev1,2*, I. S. Sadovnikova1, and V. N. Popov1,2
1 Voronezh State University, 394018 Voronezh, Russia; E-mail: gureev@bio.vsu.ru
2 Voronezh State University of Engineering Technologies, 394036 Voronezh, Russia
Methylene blue (MB) is the first fully synthetic compound that found its way into medicine over 120 years ago
to treat malaria. Today MB is approved for the treatment of methemoglobinemia, but there are premises for its
repositioning towards neuroprotection. The promise of this compound is confirmed by the effectiveness in the
treatment of Alzheimer’s, Parkinson’s, Huntington’s diseases, models of traumatic brain injury, amniotic lateral
sclerosis, depressive disorders, etc. in various models. However, the goal of this review is not so much to focus
on describing the therapeutic effects of MB in the treatment of various neurodegenerations, but to delve into
the mechanisms of direct or indirect effect by this drug on the signaling pathways. MB can act as an alternative
electron carrier in the mitochondrial respiratory chain in condition of disturbed electron transport chain. It can
have anti-inflammatory and anti-apoptotic effects, inhibit monoamine oxidases (MAOs) and NO synthase,
activate signaling pathways that provide renewal of the mitochondrial pool - mitochondrial biogenesis and
autophagy, and prevent the aggregation of misfolded proteins. Understanding and comprehensive study of all
aspects of the direct and indirect action of MB, and not just any one effect, can help in its further research,
including at the clinical level.
Keywords: methylene blue, neurodegeneration, Alzheimer’s disease, tau protein, alternative electron transport, Nrf2/
ARE signaling pathway, apoptosis, autophagy, inflammation, monoamine oxidase, NO synthase
БИОХИМИЯ том 87 вып. 9 2022