БИОХИМИЯ, 2023, том 88, вып. 10, с. 1731 - 1741
УДК 577.3
ГЕНЕРАЦИЯ РАЗНОСТИ ЭЛЕКТРИЧЕСКИХ ПОТЕНЦИАЛОВ
ХРОМАТОФОРАМИ ФОТОСИНТЕЗИРУЮЩИХ
БАКТЕРИЙ В ПРИСУТСТВИИ ТРЕГАЛОЗЫ
В ОТВЕТ НА СТАЦИОНАРНОЕ ОСВЕЩЕНИЕ
© 2023 Л.А. Витухновская1,2, А.А. Заспа1, М.Д. Мамедов1*
1 НИИ физико-химической биологии имени А.Н. Белозерского,
Московский государственный университет имени М.В. Ломоносова,
119992 Москва, Россия; электронная почта: mahirmamedov@yandex.ru
2 Федеральный исследовательский центр химической физики им. Н.Н. Семёнова РАН,
119991 Москва, Россия
Поступила в редакцию 27.06.2023
После доработки 20.07.2023
Принята к публикации 26.08.2023
Измерение разности электрических потенциалов (Δψ) в мембранных везикулах (хроматофорах) из
пурпурной бактерии Rhodobacter sphaeroides, ассоциированных с поверхностью нитроцеллюлозного
мембранного фильтра (МФ), пропитанного раствором фосфолипидов в декане, а также погружен-
ных внутрь него, в присутствии экзогенных медиаторов и дисахарида трегалозы продемонстриро-
вало увеличение амплитуды и стабилизацию сигнала при стационарном освещении. В качестве
медиаторов (донора и акцептора электронов) были использованы пара аскорбат/N,N,N′N′-тетра-
метил-п-фенилендиамин и убихинон-0 соответственно. Хотя стабилизация фотоэлектрических
ответов при длительном стационарном освещении наблюдалась в обоих случаях иммобилизации
хроматофоров, только при погружении образцов внутрь МФ сохранялась функциональная ак-
тивность работы реакционного центра в течение месяца при их хранении в темноте при комнат-
ной температуре. Стабильные фотоответы, вероятно, связаны с сохранением интактности бел-
ков хроматофоров внутри пор МФ. Стабилизирующее действие биопротектора трегалозы может
быть обусловлено как его влиянием на белки РЦ, так и на фосфолипидную бислойную мембрану.
Полученные результаты позволят расширить современные представления об использовании полу-
синтетических структур на основе различных интактных фотосинтетических систем, способных
преобразовывать солнечную энергию в электрохимическую форму.
КЛЮЧЕВЫЕ СЛОВА: хроматофоры, реакционный центр, цитохромный bc1-комплекс, мембранный
фильтр, полупроводник, постоянное освещение, электрический потенциал.
DOI: 10.31857/S0320972523100020, EDN: OSMUJE
ВВЕДЕНИЕ
высокая стоимость, применение дорогостоя-
щих компонентов, необходимость их техни-
В течение последних десятилетий уси-
ческого обслуживания и утилизации через
лия многих исследователей были сосредо-
определённое время. В связи с этим одним из
точены на разработке высокоэффективных
перспективных подходов к созданию фото-
искусственных систем на основе различных
электрохимических преобразователей энергии
красителей и полупроводников, способных
является использование природных фотосин-
преобразовывать энергию света в электриче-
тетических систем (см. обзоры Najafpour et al.
ство (см. обзоры Bella et al. и Calió et al. [1, 2]).
и Teodor et al. [3, 4]).
Однако использование этих конструкций свя-
Везикулы, образующиеся при выпячива-
зано с некоторыми недостатками, такими как нии внутрицитоплазматической мембраны
Принятые сокращения: Аск - аскорбат; БХл - бактериохлорофилл; МФ - мембранный фильтр; РЦ - реакционный
центр; ТМФД - N,N,N′N′-тетраметил-п-фенилендиамин; Хром - хроматофоры; цит - цитохром; bc1 - цитохромный
bc1-комплекс; ITO - полупроводник на основе оксида индия-олова; Р870 - димер хлорофилла; QA и QB - первичный
и вторичный хинонные акцепторы; UQ - убихинон; UQ0 -2,3-диметокси-5-метил-1,4-бензохинон; Δψ - трансмем-
бранная разность электрических потенциалов.
* Адресат для корреспонденции.
1731
2*
1
732
ВИТУХНОВСКАЯ и др.
(хроматофоры, Хром) в клетках несерных
ных системах in vitro одним из важных этапов
пурпурных бактерий, содержат фотосинтети-
является их эффективная иммобилизация на
ческий аппарат, идеально подходящий для
носителях (проводники, полупроводники, по-
таких исследований. В отличие от тилакоид-
лимеры, фосфолипидные мембраны) и изме-
ных мембран цианобактерий и хлоропластов,
рение стационарного электрического потен-
Хром фотосинтезирующих бактерий представ-
циала [12-14] и/или тока [12, 15-19]. Однако
ляют собой минимальную структурно-функ-
в приведённых выше работах [12-14] не изуча-
циональную единицу, способную преобразовы-
лась генерация Δψ при длительном стацио-
вать энергию поглощённых фотонов для син-
нарном освещении. Что касается измерений
теза ATP [5]. Светозависимый циклический
стационарного тока в Хром [15-19], то они
перенос электронов в них, например, в Rhodo-
проводились в основном с помощью трёхэлек-
bacter sphaeroides, осуществляется между ре-
тродной системы при подаче внешнего элек-
акционным центром (РЦ) и цитохромным
трического поля, где амплитуда фототока зави-
bc1-комплексом (bc1) с участием мобильного
села от величины этого поля.
мембранного пула убихинонов (UQ) и эндо-
В настоящей работе мы изучали генера-
генного растворимого цитохрома (цит) с2.
цию разности электрических потенциалов
После возбуждения РЦ квантом света
в ответ на стационарное освещение в Хром
фотоактивированный электрон переносится
Rba. sphaeroides, иммобилизованных на по-
от димера бактериохлорофилла Р870 (Р) к пер-
верхности нитроцеллюлозного мембранного
вичному (QA), а затем ко вторичному хинонно-
фильтра (МФ), пропитанного раствором фос-
му акцептору (QB). Восстановление фотоокис-
фолипидов, с помощью прямого электроме-
ленного Р870 происходит в результате переноса
трического метода [13]. Суть этого метода за-
электрона от периферического белка - донора
ключается в регистрации Δψ с помощью пары
электронов для РЦ - цит с2. Согласно моди-
хлорсеребряных (Ag/AgCl) макроэлектродов,
фицированной концепции Q-цикла (см. обзор
погружённых в раствор электролита и соеди-
Crofts et al. [6]), окисление молекулы убихи-
нённых с операционным усилителем. В пред-
нола, образовавшейся в РЦ и перенесённой в
ставленной нами работе полученные резуль-
убихинол-оксидазный центр «Qo» bc1, органи-
таты сопоставлялись с электрометрическими
зовано таким образом, что один электрон воз-
данными для Хром, погружённых внутрь МФ,
вращается через железо-серный белок Риске
зажатого с двух сторон полупроводниковыми
(Fe2S2) и цит c1 и c2 (высокопотенциальная цепь)
стеклянными пластинами на основе оксида
к фотоокисленному P870, а другой передаётся
индия-олова (англ. indium tin oxide или сокра-
через двухгемовый цитохром b (низкопотенци-
щённо ITO) [20]. Полученные данные нагляд-
альная цепь) в убихинон-редуктазный центр
но демонстрируют долговременную и стабиль-
«Qi» для восстановления молекулы UQ из пула
ную фотоэлектрическую активность Хром
убихинонов. Повторный запуск вышеописан-
в этих системах в присутствии дисахарида
ных реакций приводит к образованию «допол-
трегалозы.
нительного» убигидрохинона в сайте «Qi» и к
увеличению выхода протонов в люмен Хром на
каждый поглощённый фотон. Здесь же следует
МАТЕРИАЛЫ И МЕТОДЫ
отметить, что светоиндуцированное векторное
перемещение зарядов в РЦ и bc1 в мембранных
Выращивание клеток и получение Хром.
везикулах сопряжено с генерацией трансмем-
Клетки дикого типа Rba. sphaeroides выращи-
бранной разности электрических потенциалов
вали при 30 °С в анаэробных условиях на сре-
(Δψ) [7-11], являющейся первичной формой
де Ормерода [21] при интенсивности света
запасания энергии.
800 Вт м-2. Для получения мембранных вези-
Таким образом, относительно простое пре-
кул с высоким содержанием эндогенного рас-
паративное получение Хром из бактериальных
творимого цит c2 мы использовали процедуру
клеток, расположение энергопреобразующих
экстракции, описанную в работе Woronowicz
ферментов в интактном липидном окружении,
et al. [22], с небольшими модификациями.
способность РЦ преобразовывать энергию
Клетки, суспендированные в 25 мМ буфере
света с высокой квантовой эффективностью и
HEPES-NaOH (рН 7,5), содержащем несколько
детально исследованный механизм генерации
кристаллов ДНКазы и ингибиторов протеа-
Δψ на отдельных участках электрон-транс-
зы, разрушали с помощью Френч-пресса
портной цепи делают Хром привлекательными
(«Aminco», США). После удаления осадка су-
объектами для фотоэлектрохимических иссле-
пернатант наносили на градиент плотности
дований. При использовании Хром в гибрид-
сахарозы (5-35%, m/m) и центрифугировали
БИОХИМИЯ том 88 вып. 10 2023
ГЕНЕРАЦИЯ ЭЛЕКТРИЧЕСКОГО ОТВЕТА В ХРОМАТОФОРАХ
1733
на вертикальном роторе VTi-50, приобретён-
на поверхность МФ (~1,0 см2) каплями нано-
ном в рамках программы развития МГУ, в те-
сили 30-40 мкл суспензии Хром с концентра-
чение 2 ч при 27 000 об./мин. Нижнюю полосу
цией БХл ~2 мг/мл. После 10-минутной темно-
Хром собирали, дважды диализовали против
вой адсорбции поверхность МФ промывалась
25 мМ буфера HEPES-NaOH (рН 7,5) в тече-
(2 × 250 мкл) 25 мМ буфером HEPES-NaOH
ние 2 ч и концентрировали. Суспензию Хром
(рН 7,5), и после этого второй электрод ITO
(~2 мг бактериохлорофилла (БХл)/мл) замора-
устанавливался сверху.
живали в жидком азоте и хранили при -70 °С.
Следует отметить, что в системе ITO|Хром -
Концентрацию БХл в Хром определяли,
МФ|ITO медиаторы добавлялись в суспензию
как описано ранее [23]. Спектры поглощения
Хром до их иммобилизации внутри МФ.
Хром регистрировали на спектрофотометре
Регистрация электрических потенциалов
Hitachi 3400 («Hitachi», Япония).
в системе ITO|Хром - МФ|ITO проводилась
Измерение разности электрических потен-
с помощью медных проводов, соединённых,
циалов в Хром, иммобилизованных на поверх-
с одной стороны, с предметными стёклами,
ности МФ. Δψ измеряли с помощью прямого
покрытыми ITO, а другой стороны - с опера-
электрометрического метода, разработанного
ционным усилителем («Burr Brown», 3554BM).
в нашей лаборатории [13]. В качестве матрикса
Сигнал от усилителя подавался на АЦП (Gage
для иммобилизации Хром были использова-
CS8012), подключённый к компьютеру. Насы-
ны МФ (Millipore GSTF, «Millipore», США) с
щающее непрерывное освещение обеспечива-
диаметром пор 0,22 мкм и толщиной 150 мкм.
лось лампой накаливания (12 В, 90 Вт).
Круг из МФ диаметром 2,0 см, пропитанный
Каждое фотоэлектрическое измерение по-
декановым раствором азолектина (80 мг/мл)
вторялось не менее 3 раз. Диапазон погрешно-
(L-α-lecithin, type II-S, «Sigma»), зажимали
сти для фотоответов составлял ~5%. Все из-
между двумя частями разборной тефлоновой
мерения проводились при 20 ± 1 °C.
кюветы так, чтобы фильтр перекрывал отвер-
Рисунки подготовлены с помощью пакета
стие диаметром
0,4 см в перегородке, раз-
программ Origin 7.5 («OriginLab Corporation»).
деляющей два отсека. Затем оба отсека запол-
Оптическая спектроскопия. Фотоиндуци-
няли буфером 25 мМ HEPES-NaOH (рН 7,5),
рованные изменения поглощения при 603 нм,
содержащим 20 мМ MgCl2. Хром добавляли
отражающие редокс-свойства первичного до-
только в один отсек ячейки. После перемеши-
нора электронов Р870, регистрировали с помо-
вания в течение 1 ч оба отсека ячейки про-
щью однолучевого дифференциального спек-
мывались десятикратным объёмом буфера без
трофотометра, сконструированного в нашей
MgCl2 с помощью перистальтического насоса
лаборатории. Измеряющий свет от лампы
для удаления магния и не связавшихся с филь-
КГМ-98, питаемой от источника постоянно-
тром Хром.
го тока TEC-15, проходил через монохрома-
Используемые экзогенные медиаторы до-
тор HL-1 («Jobin Ivon», Франция), кювету с об-
бавлялись в отсек ячейки с иммобилизован-
разцом, стеклянный светофильтр, второй мо-
ными на поверхности МФ Хром перед изме-
нохроматор УМ-2, а затем попадал на фото-
рением.
электронный умножитель (ФЭУ). Сигнал с
Фотоэлектрические ответы регистрирова-
ФЭУ через операционный усилитель («Burr
лись при помощи пары хлорсеребряных (Ag/
Brown», 3554BM) подавался на АЦП (Gage
AgCl/3 М KCl) макроэлектродов, погружённых
CS8012), подключённый к компьютеру.
по обе стороны от МФ в раствор электролита.
Электроды были подключены к операционно-
му усилителю («Burr Brown», 3554BM, США),
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
а последний, в свою очередь, к аналого-цифро-
вому преобразователю (АЦП, Gage CS8012)
Целью данной работы было исследова-
и компьютеру. В качестве источника постоян-
ние генерации Δψ в интактных бактериальных
ного света использовалась лампа накаливания
мембранных везикулах при непрерывном осве-
с мощностью 90 Вт (12 В).
щении. Ранее было показано [24], что Хром,
Измерение разности электрических потен-
выделенные из несерной пурпурной бакте-
циалов в Хром, погружённых внутрь МФ. Для
рии Rba. sphaeroides в определённых условиях
этого реконструировалась система ITO|Хром-
(разрушение клеток с помощью Френч-пресса
МФ|ITO. МФ (Millipore GSTF, диаметр пор
при низкой ионной силе), содержат ~70% цит c2
0,22 мкм, толщина 150 мкм) размером 2,5 × 5 см
внутри везикул (см. Материалы и методы).
помещали на поверхность предметного стек-
В таких Хром восстановление фотоокислен-
ла, покрытого полупроводником ITO. Затем
ного Р870 в РЦ может происходить в результате
БИОХИМИЯ том 88 вып. 10 2023
1734
ВИТУХНОВСКАЯ и др.
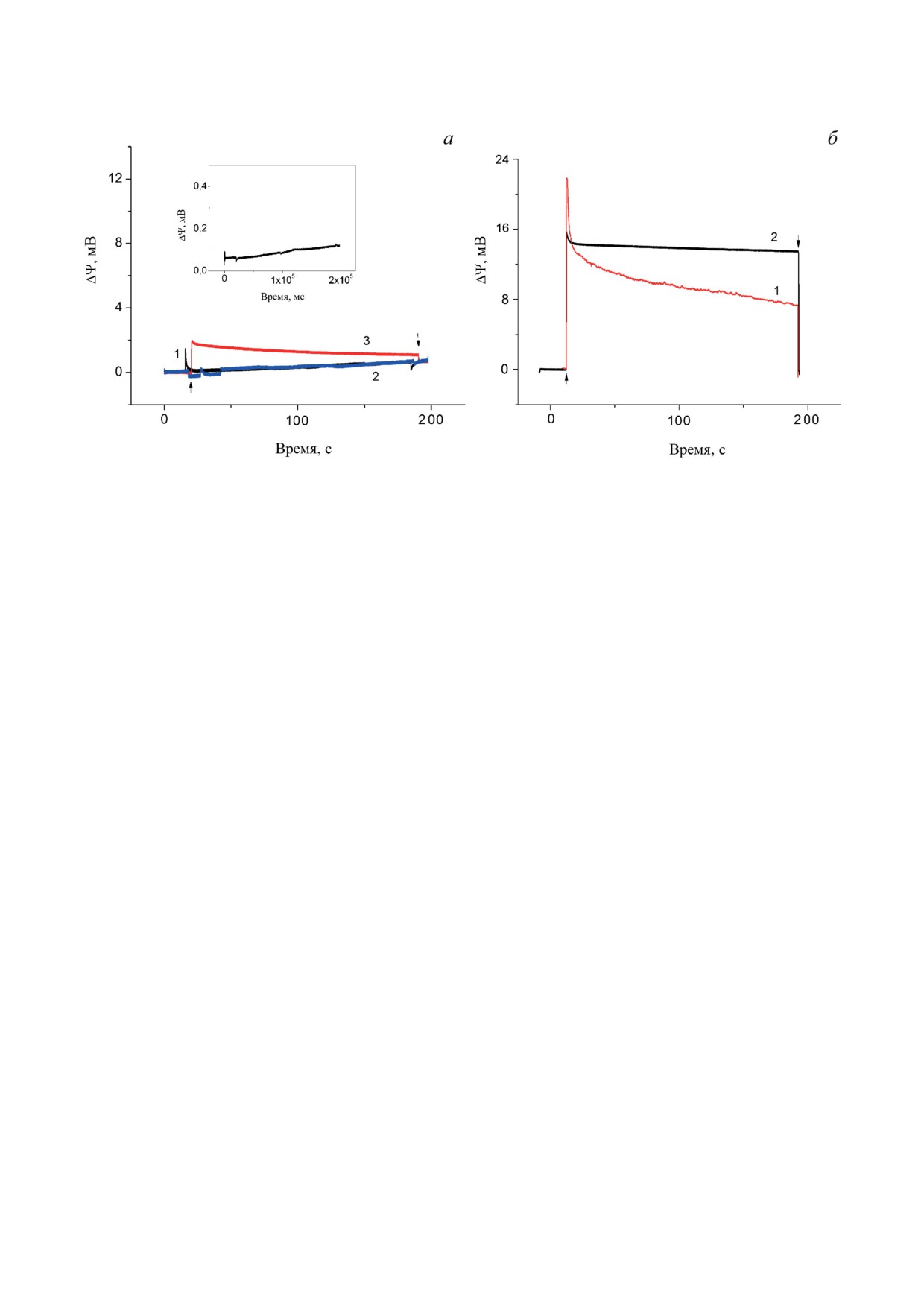
Рис. 1. Генерация электрических потенциалов в Хром. а - Генерация разности электрических потенциалов в Хром,
ассоциированных с поверхностью МФ, пропитанного раствором фосфолипидов при стационарном освещении. Раз-
меры пор 0,22 мкм, толщина фильтра 150 мкм. Фильтр пропитывался декановым раствором азолектина (80 мг/мл).
Хром без добавок (кривая 1), в присутствии пары 2,5 мМ Аск/150 мкМ ТМФД (кривая 2) и в присутствии только
150 мкМ UQ0 (кривая 3). Среда инкубации: 25 мМ HEPES-NaOH (pH 7,5). б - Генерация разности электрических
потенциалов в среде измерения, содержащей 2,5 мМ Аск/150 мкМ ТМФД и 150 мкМ UQ0 в отсутствии (кривая 1)
и в присутствии (кривая 2) 0,75 М трегалозы. Вставка к рис. 1, а - фотоответ в присутствии 5 мМ Аск/150 мкМ ТМФД
и 150 мкМ UQ0, но без Хром. (↑) и (↓) указывают включение и выключение света
прямого переноса электрона от эндогенного
ион-радикальной пары P870Q− внутри белка РЦ.
цит с2 [24, 25]. Другими словами, функциональ-
Однако перенос заряда за пределы РЦ в опи-
но активные Хром должны содержать цит c2,
санной системе может быть реконструирован
который наряду с пулом убихинонов выступает
путём насыщения иммобилизованных Хром
в качестве окислительно-восстановительного
экзогенным хиноном и восстановления эндо-
переносчика между комплексами РЦ и bc1.
генного цит с2 (см. работу Drachev et al. [14]).
Содержание цит с2 было определено нами пу-
В Хром, ассоциированных с поверхно-
тём регистрации абсорбционных изменений
стью МФ, в присутствии только аскорбата
при 603 нм, отражающих редокс-свойства Р870
(Аск) и N,N,N′N′-тетраметил-п-фенилендиами-
в ответ на одиночные вспышки света в сус-
на (ТМФД) в среде измерения фотоэлектри-
пензии Хром в отсутствии и в присутствии 2%
ческий ответ также не наблюдался (рис. 1, а,
детергента Triton Х-100 (не показано) (см. ра-
кривая 2). Использование пары Аск и ТМФД
боту Altamura et al. [26]).
в концентрациях 2,5 мМ и 150 мкМ соответ-
На рис. 1 продемонстрирована генерация
ственно в качестве донора электронов об-
Δψ при постоянном освещении Хром Rba.
условлено необходимостью восстановления
sphaeroides, ассоциированных с поверхностью
растворимого цит с2, поскольку он локализо-
МФ, пропитанного раствором фосфолипида.
ван внутри Хром и не взаимодействует с плохо
Видно, что в отсутствии каких-либо добавок
проникающим через мембрану Аск [9].
(медиаторов) фотоэлектрический ответ не
На рис. 1, а, кривая 3 показана генерация
наблюдается (рис. 1, а, кривая 1). Следует от-
Δψ при стационарном освещении в присут-
метить, что при ассоциации Хром с МФ, про-
ствии только акцептора электронов 2,3-димет-
питанным раствором фосфолипидов в декане,
окси-5-метил-1,4-бензохинона (UQ0) в концен-
как и в случае с коллодиевой плёнкой [9, 10],
трации 150 мкМ. Амплитуда фотоответа в этих
происходит экстракция UQ как из сайта вто-
условиях составляла ~1,8 мВ. UQ0 является эф-
ричного хинонного акцептора QB в белке РЦ,
фективным экзогенным акцептором электрона
так и из мембранного пула. Кроме того, про-
от хинонов в РЦ [27, 28] и широко используется
исходит окисление цит с2 и первичного доно-
в фотоэлектрохимических системах [28].
ра электронов - димера БХл Р870 в РЦ. В этих
В качестве контроля на вставке к рис. 1, а
условиях светоиндуцированный перенос элек-
показана генерация трансмембранной разно-
тронов в Хром ограничивается образованием
сти электрических потенциалов (Δψ) в присут-
БИОХИМИЯ том 88 вып. 10 2023
ГЕНЕРАЦИЯ ЭЛЕКТРИЧЕСКОГО ОТВЕТА В ХРОМАТОФОРАХ
1735
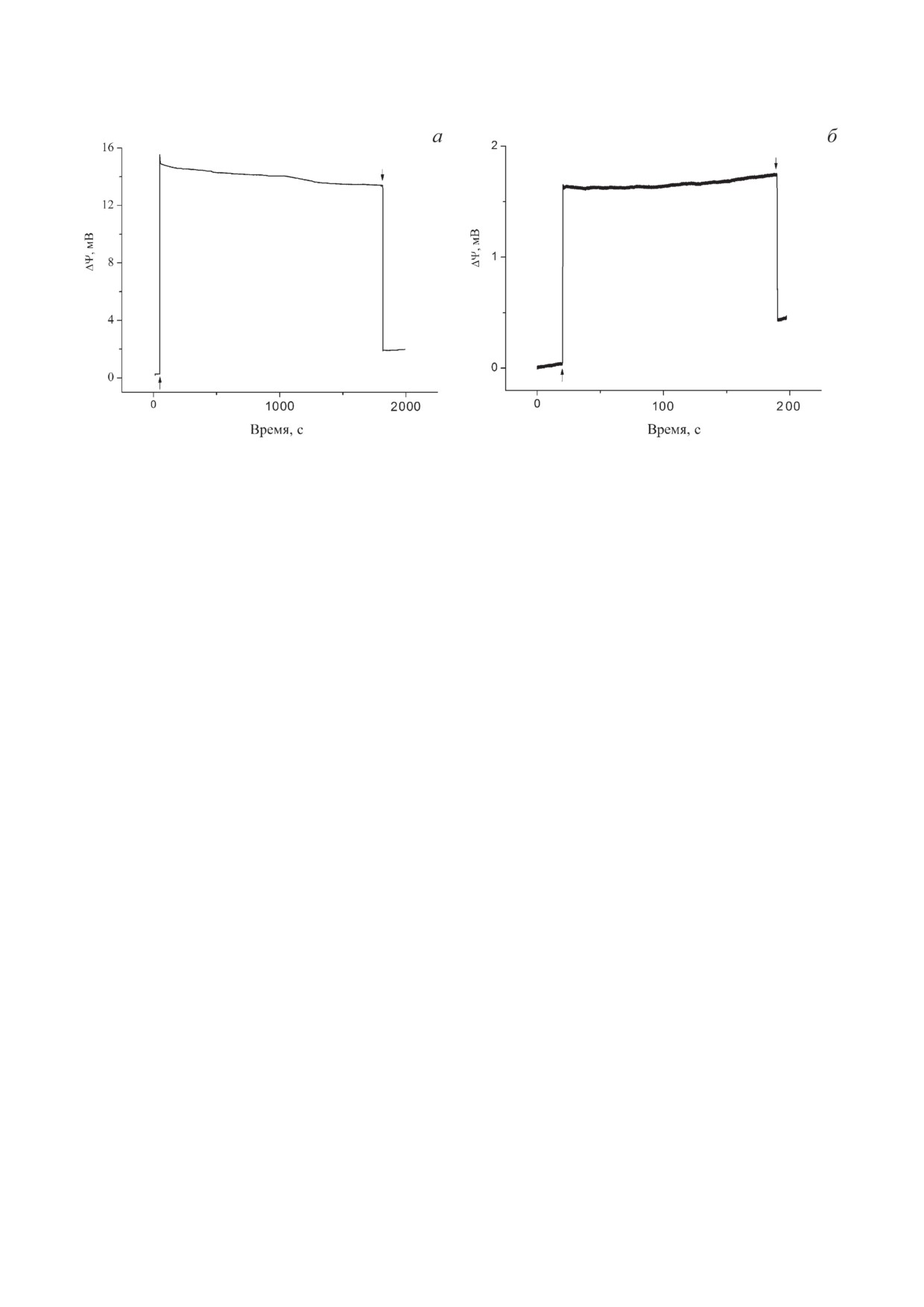
Рис. 2. Генерация электрических потенциалов в Хром в присутствии трегалозы. а - Генерация разности электриче-
ских потенциалов стандартно выделенными Хром, ассоциированными с поверхностью МФ, при стационарном осве-
щении в присутствии 0,75 М трегалозы. б - То же в случае Хром, дефицитных по цит с2. Условия эксперимента как
на рис. 1, б, кривая 2
ствии Аск/ТМФД и UQ0, но без Хром. Видно,
Перенос зарядов внутри белковых ком-
что в этих условиях ответ не наблюдается.
плексов РЦ и bc1 в Хром сопряжён с генерацией
На рис. 1, б, кривая 1 приведён фотоэлек-
Δψ, измеренной как с помощью электрохром-
трический ответ Хром при стационарном осве-
ного сдвига полос поглощения каротиноидов,
щении в присутствии как донора (Аск/ТМФД),
так и прямого электрометрического метода [10].
так и акцептора (UQ0) электронов. Видно, что
На рис. 3, а, кривая 1 продемонстрирована ге-
только в присутствии обоих медиаторов на-
нерация Δψ при добавлении Аск/ТМФД, UQ0
блюдается значительный рост фотоэлектриче-
и трегалозы при стационарном освещении.
ского ответа с последующим двухкомпонент-
Для определения вклада bc1 в общий фото-
ным спадом Δψ - в быстрой (~3 с) и медленной
ответ в среду измерения добавляли антими-
(~180 с) шкалах времени.
цин А, ингибитор убихинон-редуктазного сай-
Следующие эксперименты были прове-
та («Qi») bc1. Видно (рис. 3, а, кривая 2), что до-
дены для изучения влияния биопротектора
бавление антимицина уменьшает амплитуду
дисахарида трегалозы на светозависимое обра-
фотоответа на ~12%. Наблюдаемый эффект, ве-
зование Δψ иммобилизованными на поверхно-
роятно, связан с тем, что используемый нами
сти МФ хроматофорами. В присутствии пары
UQ0 связывается в сайте QB в РЦ [28], получает
Аск/ТМФД и UQ0 добавление 0,75 М трега-
электрон от Q−, а его восстановленная форма
лозы приводило к стабилизации сигнала Δψ
(UQ0H2) выступает в качестве «субстрата» лишь
при стационарном освещении в течение ~190 с
для небольшой фракции bc1.
(рис. 1, б, кривая 2). В этих условиях также
Для доказательства того, что наблюдаемая
удалось наблюдать стабилизацию сигнала Δψ
фотоэлектрическая активность Хром обуслов-
при значительно более длительном освещении
лена функционированием РЦ, эффективный
(~1800 с) (рис. 2, а).
ингибитор переноса электронов между хи-
На рис. 2, б показана генерация Δψ дефи-
нонными акцепторами Q− и Q− атразин был
цитными по цит c2 Хром из Rba. sphaeroides в
добавлен в среду измерения. Действительно,
присутствии пары Аск/ТМФД, UQ0 и трега-
добавление 10 мкМ атразина практически пол-
лозы. Освещение таких Хром приводит к гене-
ностью ингибировало генерацию фотоэлек-
рации разности электрических потенциалов
трического ответа (рис. 3, а, кривая 3). Этот
амплитудой ~1,8 мВ. Этот результат свидетель-
результат показывает, что в исследуемой систе-
ствует о том, что проникающая через мембрану
ме перенос электрона на акцепторном участке
восстановленная форма ТМФД в концентра-
происходит от первичного хинона QA на экзо-
ции 150 мкМ недостаточно эффективно вос-
генный акцептор Q0.
станавливает фотоокисленный Р870 в стацио-
Для изучения стабильности фотоответов
нарных условиях [11].
Хром, ассоциированные с поверхностью МФ,
БИОХИМИЯ том 88 вып. 10 2023
1736
ВИТУХНОВСКАЯ и др.
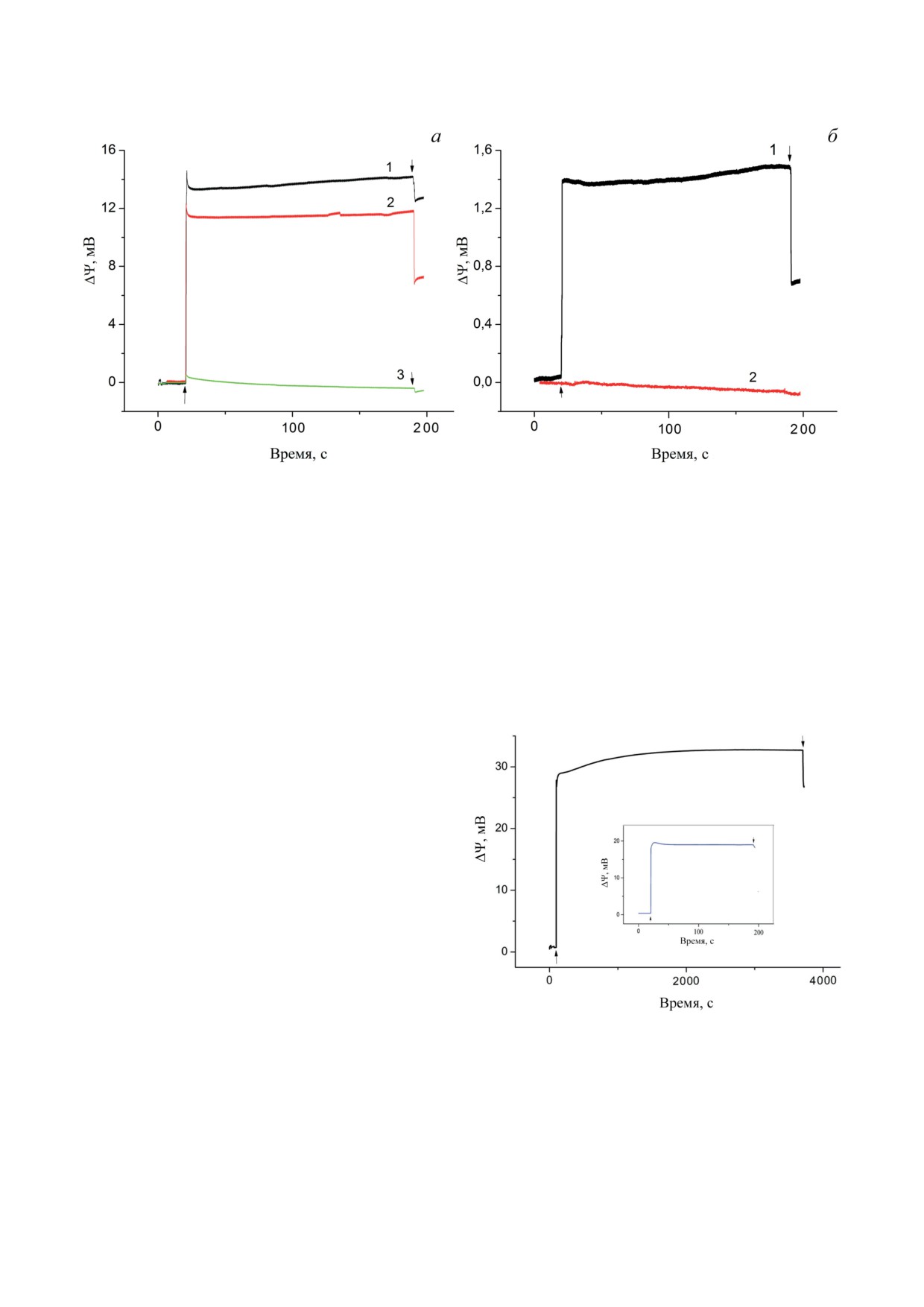
Рис. 3. Генерация электричества в Хром, ассоциированных с поверхностью МФ. а - Генерация разности электриче-
ских потенциалов в Хром, ассоциированных с поверхностью МФ, пропитанного раствором фосфолипидов. Условия
как на рис. 1, б, кривая 2 в отсутствии (кривая 1) и в присутствии 5 мкМ антимицина А (кривая 2); кривая 3 - условия
как на рис. 3, а (кривая 1), но в присутствии 10 мкМ атразина. б - Генерация разности электрических потенциалов
хроматофорами после 18 ч (кривая 1) и 40 ч (кривая 2) хранения при комнатной температуре. Условия эксперимента
как на рис. 3, а, кривая 1
хранились в темноте при комнатной темпера-
условиях амплитуда фотоответа составляла
туре и освещались через определённые про-
~70% от амплитуды ответа контрольного об-
межутки времени. Видно, что амплитуда Δψ,
разца.
изначально составлявшая
~14 мВ (рис. 3, а,
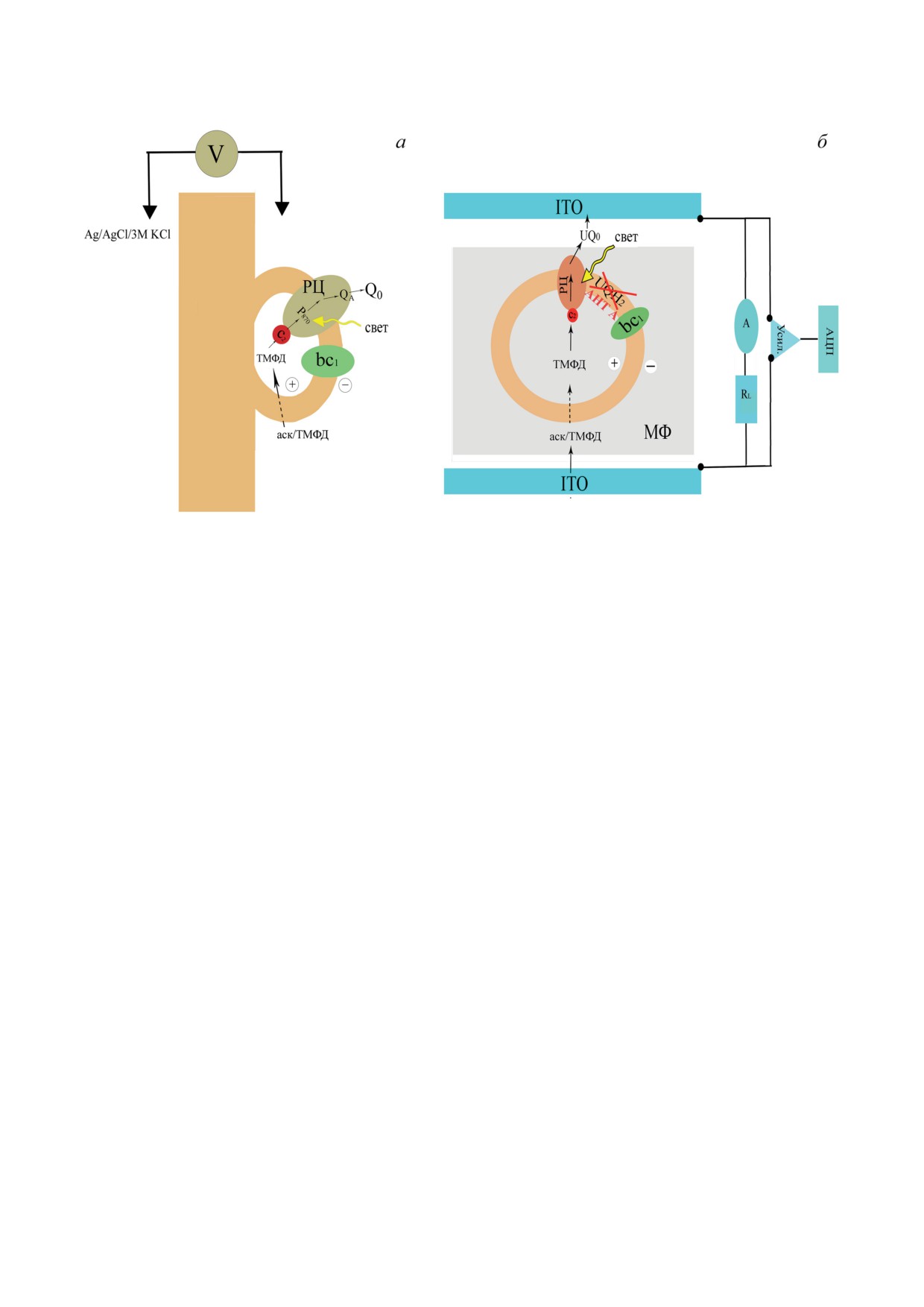
На рис. 5 показаны пути переноса элек-
кривая 1), через 18 ч хранения уменьшилась
тронов в Хром, иммобилизованные двумя раз-
до ~1,4 мВ (рис. 3, б, кривая 1), а через 40 ч
ными способами.
фотоответ не наблюдался (рис. 3, б, кривая 2).
На рис. 4 показана генерация Δψ в Хром,
погруженных внутрь МФ и зажатых с двух
сторон полупроводниковыми стеклянными
пластинами на основе оксида индия-олова
(ITO|Хром-МФ|ITO) с добавленными медиа-
торами (Аск/ТМФД, UQ0), антимицином А,
а также трегалозой. Предполагалось, что ис-
пользуемые редокс-медиаторы свободно пе-
ремещаются внутри МФ и могут взаимодей-
ствовать как с РЦ, так и электродами ITO.
Видно, что при освещении системы в тече-
ние ~4000 с генерируется стабильный неспа-
дающий фотоэлектрический ответ амплиту-
дой ~30 мВ [20].
Следует отметить, что 18-часовая темно-
вая адаптация при комнатной температуре
системы ITO|Хром-МФ|ITO приводила к зна-
Рис. 4. Генерация электричества в Хром, ассоцииро-
ванных с поверхностью МФ, в присутствии трегалозы.
чительному уменьшению сигнала Δψ при осве-
Генерация Δψ в Хром, погруженных внутрь МФ (си-
щении в течение 180 с (не показано). На встав-
стема ITO|Хром-МФ|ITO), в присутствии 2,5 мМ Аск,
ке к рис. 4 показана генерация Δψ при вне-
150 мкМ ТМФД, 150 мкМ UQ0, 0,75 М трегалозы и
сении свежего буфера, 25 мМ HEPES-NaOH
5 мкМ антимицина А при освещении в течение 4000 с
(контроль). Вставка: генерация Δψ после внесения све-
(pH 7,5, ~40 мкл), с медиаторами (Аск/ТМФД
жего буфера 25 мМ HEPES-NaOH (pH 7,5), содержа-
и UQ0) непосредственно в систему ITO|Хром-
щего Аск/ТМФД и UQ0, непосредственно в систему
МФ|ITO после хранения её в течение 30 дней
ITO|Хром-МФ|ITO на 30-й день её хранения при ком-
в темноте при комнатной температуре. В этих
натной температуре
БИОХИМИЯ том 88 вып. 10 2023
ГЕНЕРАЦИЯ ЭЛЕКТРИЧЕСКОГО ОТВЕТА В ХРОМАТОФОРАХ
1737
Рис.
5. Предполагаемые пути переноса электронов, индуцированные стационарным освещением в Хром
Rba. sphaeroides, ассоциированных с поверхностью МФ (а) и погруженных в МФ (б)
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
матрицы для иммобилизации Хром. Ассоциа-
ция образцов с поверхностью МФ достигалась
Исследования механизма генерации Δψ в
путём нейтрализации отрицательных зарядов
Хром, ассоциированных с поверхностью кол-
полярных «головок» фосфолипидов мембран
лодиевой плёнки, пропитанной раствором ли-
Хром и МФ в присутствии катионов Mg2+ или
пидов, в ответ на одиночные вспышки света с
Са2+ [13] (см. «Материалы и методы»).
помощью прямого электрометрического ме-
Для генерации стабильных электрических
тода продемонстрировали наличие следующих
сигналов использовались функционально-ак-
электрогенных реакций: 1) разделение зарядов
тивные Хром, которые характеризуются высо-
между Р870 и QA, 2) ре-восстановление фото-
ким содержанием эндогенного цит с2. Так как
окисленного Р870 эндогенным цит с2, 3) про-
редокс-кофакторы в трансмембранных ком-
тонирование дважды восстановленной формы
плексах РЦ и bc1 погружены глубоко в бел-
вторичного хинонного акцептора QB (см. об-
ковый матрикс, при стационарном освеще-
зор Kaminskaya et al. [11]) и 4) перенос элек-
нии in vitro (в полусинтетической системе) Δψ
тронов и протонов в bc1 [10].
регистрировалась в присутствии экзогенных
Целью данной работы была демонстрация
медиаторов. Использование пары Аск/ТМФД
и изучение механизма генерации Δψ в Хром,
было обусловлено проницаемостью мембра-
иммобилизованных как на поверхности МФ,
ны Хром для восстановленной формы ТМФД
так и погруженных в него при стационарном
и её способностью восстанавливать эндоген-
освещении.
ный цит с2 [9]. Наличие растворимого UQ0,
Следует отметить, что адсорбция биомате-
в свою очередь, поддерживало функциональ-
риалов на твёрдой поверхности (электроды
ную активность РЦ, акцептируя электроны от
Au, TiO2, ITO) может приводить как к струк-
первичного хинона Q− в Хром, ассоцииро-
турным изменениям, так и к возникновению
ванных с поверхностью, и от вторичного хи-
электростатического отталкивания между ис-
нона Q− в случае Хром, погружённых внутрь
следуемыми образцами и поверхностью но-
МФ [20, 28]. UQ0 являлся эффективным ме-
сителя (работа Yaghoubi et al. [29] и ссылки в
диатором между хинон-акцепторным участ-
ней). Принимая во внимание биосовмести-
ком РЦ и электродами [28, 32].
мость, биоразлагаемость, нетоксичность и
В Хром Rba. sphaeroides, ассоциированных
низкую стоимость производного природного
с поверхностью МФ, происходит вымывание
полимера
- пористого нитроцеллюлозного
непрочно связанного QB и пула UQ н-дека-
МФ [30, 31], мы использовали его в качестве
ном, используемым в качестве гидрофобного
БИОХИМИЯ том 88 вып. 10 2023
1
738
ВИТУХНОВСКАЯ и др.
растворителя фосфолипидов при пропиты-
и добавление в среду измерения свежей пор-
вании МФ; следствием этого является блоки-
ции буфера с медиаторами (Аск/ТМФД, UQ0)
рование циклического переноса зарядов [13].
не приводило к ре-восстановлению Δψ (не по-
В этих условиях убигидрохинон (UQ0H2), ге-
казано). Следовательно, уменьшение ампли-
нерируемый при стационарном освещении,
туды Δψ с течением времени, скорее всего,
вероятно, из-за высокой гидрофильности не
связано с конформационно-структурными из-
может выступать в качестве субстрата для bc1
менениями РЦ в случае Хром, ассоциирован-
(рис. 5, а).
ных с поверхностью МФ.
С другой стороны, при погружении Хром
Что касается стабильности фотоэлектри-
внутрь МФ сохраняется функционирование
ческих ответов при погружении Хром внутрь
циклической электрон-транспортной цепи с
МФ, то следует отметить, что после хранения
участием комплексов РЦ и bc1, а также пула
системы ITO|Хром - МФ|ITO при комнатной
UQ и цит с2. Этот факт подтверждается тем, что
температуре в аэробных условиях в течение
добавление ингибитора bc1 - антимицина А -
30 дней при внесении в систему свежего бу-
приводит к значительному (почти в 2 раза)
фера, содержащего донор и акцептор электро-
увеличению амплитуды стационарной Δψ [20].
нов, наблюдалось восстановление амплитуды
Увеличение амплитуды фотоэлектрического
Δψ до ~70% (вставка к рис. 4) относительно
ответа в условиях ингибирования bc1 обуслов-
контрольного образца (рис. 4). Последнее сви-
лено перенаправлением циклического потока
детельствует о том, что большинство Хром,
электронов, не вносящего вклад в регистри-
иммобилизованных на МФ, остаются функ-
руемый в такой системе мембранный потен-
ционально активными, по крайней мере, в те-
циал, в пользу линейного переноса электронов
чение 30 дней.
между электродами ITO и РЦ (рис. 5, б). Такой
Становится очевидным, что стабильные
тип иммобилизации позволяет регистрировать
максимальные фотоответы в обеих системах
стабильный неспадающий сигнал Δψ при дли-
наблюдались только в присутствии уникаль-
тельном освещении (~4000 с) [20].
ного по своим физико-химическим свой-
Таким образом, регистрируемый макси-
ствам дисахарида трегалозы. Ранее такие же
мальный фотоэлектрический ответ, вероятно,
стабильные фотоэлектрические ответы были
обусловлен светоиндуцированным прямым пе-
выявлены в присутствии этого осмолита в изо-
реносом электронов по пути цит с2 → РЦ → QA →
лированных пигмент-белковых комплексах
→ Q0 → O2 в случае Хром, ассоциированных с по-
ФС1 и ФС2, погруженных внутрь МФ [33, 34].
верхностью МФ (рис. 5, а), или ITO → цит c2 →
Отметим, что трегалоза также влияет на харак-
→ РЦ → QA → QB → UQ0 → ITO для Хром, погру-
тер электрического ответа (рис. 1, б, кривая 2),
жённых внутрь МФ (рис. 5, б). Другими сло-
а именно - подавляет быстрый «всплеск» и
вами, в обеих системах генерация стационар-
спад сигнала Δψ в начале освещения (рис. 1, б,
ной Δψ обусловлена работой самого РЦ.
кривая 1). Это может быть связано с умень-
Об этом свидетельствует подавление генера-
шением ионной проницаемости мембраны
ции электрических ответов в присутствии атра-
Хром [35, 36]. Аналогичное влияние трегало-
зина - ингибитора переноса электронов ме-
зы на быстрый «всплеск» и спад фотоответа в
жду хинонными акцепторами в РЦ (рис. 3, а,
отсутствии осмолита также наблюдалось при
кривая 3).
погружении Хром в МФ [20]. Стабилизация
В работах по исследованию преобразова-
фотоответов может быть связана с улучше-
ния энергии света важно не только получение
нием эффективности взаимодействия медиа-
значительной амплитуды тока или электриче-
тора(ов) с белками фотосинтетической элек-
ства при стационарном освещении, но и выяв-
трон-транспортной цепи. Сохранение тонкой
ление условий сохранения функциональной
гидратной оболочки трансмембранных белков
активности образцов при их длительном хра-
в присутствии этого дисахарида может менять
нении при комнатной температуре. В Хром,
их конформацию в пользу более оптимальной
ассоциированных с поверхностью МФ, через
для эффективного функционирования [37-39].
40 ч наблюдалось практически полное паде-
Трегалоза, вероятно, также стабилизирует ли-
ние амплитуды Δψ (рис. 3, б, кривая 2). Паде-
пидные бислои, замещая воду при образова-
ние амплитуды фотоответа может быть свя-
нии водородных связей между её собственны-
зано или с уменьшением сопротивления МФ,
ми ОН-группами и полярными «головками»
пропитанного раствором липидов [13], или
липидов [40]. При этом взаимодействие осмо-
же со временем «деградации» добавленных
лита с трансмембранными белками и с фос-
медиаторов, в частности Аск. Однако, ре-вос-
фолипидами в Хром может осуществляться
становление сопротивления МФ (2 × 108 Oм)
только вблизи внешней стороны мембраны.
БИОХИМИЯ том 88 вып. 10 2023
ГЕНЕРАЦИЯ ЭЛЕКТРИЧЕСКОГО ОТВЕТА В ХРОМАТОФОРАХ
1739
Последнее обстоятельство обусловлено непро-
способных преобразовывать солнечную энер-
ницаемостью мембраны Хром для трегалозы.
гию в электрохимическую форму.
Таким образом, полученные данные позво-
ляют заключить, что трегалоза стабилизиру-
Вклад авторов. М.Д. Мамедов - концеп-
ет фотоэлектрические сигналы, генерируемые
ция и руководство работой, анализ электриче-
Хром, в обоих случаях, но наиболее стабиль-
ских измерений, написание и редактирование
ные фотоответы наблюдаются при погружении
текста статьи; Л.А. Витухновская, А.А. Заспа -
Хром внутрь МФ. Этот крайне простой способ
выделение бактериальных мембранных вези-
иммобилизации образцов, вероятно, позволя-
кул (хроматофоров) и проведение электриче-
ет сохранить интактность фотосинтетических
ских измерений.
белков Хром внутри пор МФ при комнатной
Финансирование. Работа выполнена при
температуре в течение длительного периода
поддержке Российского научного фонда (РНФ),
времени.
грант № 23-74-00025.
Полученные результаты позволят расши-
Конфликт интересов. Авторы заявляют об
рить современные представления об использо-
отсутствии конфликта интересов.
вании полусинтетических структур на основе
Соблюдение этических норм. Настоящая
различных интактных фотосинтетических си-
статья не содержит описания каких-либо ис-
стем (клетки цианобактерий и пурпурных бак-
следований с участием людей или животных
терий [41], тилакоидные мембраны растений),
в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Bella, F., Gerbaldi, C., Barolo, C., and Grätzel, M.
reductase complex of Rhodopseudomonas sphaeroi-
(2015) Aqueous dye-sensitized solar cells, Chem. Soc.
des, Biochim. Biophys. Acta,
766,
322-333, doi:
Rev., 44, 3431-3473, doi: 10.1039/c4cs00456f.
10.1016/0005-2728(84)90248-2.
2.
Calió, L., Kazim, S., Grätzel, M., and Ahmad, S.
9. Drachev, L. A., Kaminskaya, O. P., Konstantinov,
(2016) Hole-transport materials for perovskite solar
A. A., Mamedov, M. D., Samuilov, A. Y., et al. (1986)
cells, Angew. Chem. Int. Ed. Engl., 55, 14522-14545,
Effects of electron donors and acceptors on the kinetics
doi: 10.1002/anie.201601757.
of the photoelectric responses in Rhodospirillum rubrum
3.
Najafpour, M. M., Renger, G., Hołyńska, M.,
and Rhodopseudomonas sphaeroides chromatophores,
Moghaddam, A. N., Aro, E. M., et al.
(2016)
Biochim. Biophys. Acta,
850,
1-9, doi:
10.1016/
Manganese compounds as water-oxidizing catalysts:
0005-2728(86)90002-2.
from the natural water-oxidizing complex to nanosized
10. Drachev, L. A., Kaurov, B. S., Mamedov, M. D.,
manganese oxide structures, Chem. Rev., 116, 2886-
Mulkidjanian, A. Y., Semenov, A. Y., et al. (1989)
2936, doi: 10.1021/acs.chemrev.5b00340.
Flash-induced electrogenic events in the photosyn-
4.
Teodor, A. N., and Bruce, B. D. (2020) Putting photo-
thetic reaction center and bc1 complexes of Rhodobacter
system I to work: truly green energy, Trends Biotechnol.,
sphaeroides chromatophores, Biochim. Biophys. Acta,
38, 1329-1342, doi: 10.1016/j.tibtech.2020.04.004.
973, 189-197, doi: 10.1016/S0005-2728(89)80421-9.
5.
Singharoy, A., Maffeo, C., Delgado-Magnero, K. H.,
11. Kaminskaya, O. P., and Semenov, A. Yu.
(2021)
Swainsbury, D. J. K., Sener, M., et al. (2019) Atoms
The mechanisms of electrogenic reactions in bac-
to phenotypes: molecular design principles of cellular
terial
photosynthetic reaction centers: Stud-
energy metabolism, Cell,
179,
1098-1111, doi:
ies in collaboration with Alexander Konstantin-
10.1016/j.cell.2019.10.021.
ov, Biochemistry (Moscow), 86, 1-7, doi: 10.1134/
6.
Crofts, A. R. (2021) The modified Q-cycle: a look
S0006297921010016.
back at its development and forward to a functional
12. Ajiki, S., Toyotama, H., Hara, M., and Miyake, J.
model, Biochim. Biophys. Acta Bioenerg., 1862, 148417,
(1997) Light-induced electrical response of chro-
doi: 10.1016/j.bbabio.2021.148417.
matophore film in semi-wet photocell with an
7.
Crofts, A. R., Meinhardt, S. W., Jones, K. R., and
agar layer containing an electron mediator, Bio-
Snozzi, M. (1983) The role of the quinone pool in
electrochem. Bioenergetics, 43, 71-75, doi: 10.1016/
the cyclic electron-transfer chain of Rhodopseudo-
S0302-4598(96)05177-X.
monas sphaeroides: a modified Q-cycle mechanism,
13. Drachev, L. A., Kaulen, A. D., Semenov, A. Yu.,
Biochim. Biophys. Acta, 723, 202-218, doi: 10.1016/
Severina, I. I., and Skulachev, V. P. (1979) Lipid-
0005-2728(83)90120-2.
impregnated filters as a tool for studying the electric
8.
Glaser, E. C., and Crofts, A. R. (1984) A new elec-
current-generating proteins, Anal. Biochem.,
96,
trogenic step in the ubiquinol: cytochrome c2 oxido-
250-262, doi: 10.1016/0003-2697(79)90580-3.
БИОХИМИЯ том 88 вып. 10 2023
1
740
ВИТУХНОВСКАЯ и др.
14.
Drachev, L. A., Mamedov, M. D., and Semenov,
(1987) The adaptation of the electron transfer of
A. Y. (1987) The antimycin-sensitive electrogenesis in
Rhodopseudomonas capsulata to different light
Rhodobacter sphaeroides chromatophores, FEBS Lett.,
intensities, Biochim. Biophys. Acta,
890,
335-345,
213, 128-132, doi: 10.1016/0014-5793(87)81477-1.
doi: 10.1016/0005-2728(87)90161-7.
15.
Seibert, M., and Kendall-Tobais, M. W.
(1982)
25.
Dutton, P. L., Petty, K. M., Bonner, H. S., and
Photoelectrochemical properties of electrodes coated
Morse, S. D. (1975) Cytochrome c2 and reaction
with photoactive-membrane vesicles isolated from
center of Rhodopseudomonas sphaeroides Ga. mem-
photosynthetic bacteria, Biochim. Biophys. Acta,
branes. Extinction coefficients, content, half-reduc-
681, 504-511, doi: 10.1016/0005-2728(82)90193-1.
tion potentials, kinetics and electric field alterations,
16.
Hara, M., Ajiki, S., and Miyake, J. (1998) Topo-
Biochim. Biophys. Acta, 387, 536-556, doi: 10.1016/
logical characterization and immobilization of a
0005-2728(75)90092-4.
chromatophore membrane from Rhodopseudomo-
26.
Altamura, E., Albanese, P., Marotta, R., Milano, F.,
nas viridis for application as a photoelectrical de-
Fiore, M., et al. (2021) Chromatophores efficiently
vice, Science, 5, 717-721, doi: 10.1016/S0968-5677
promote light-driven ATP synthesis and DNA tran-
(98)00111-4.
scription inside hybrid multicompartment artificial
17.
Magis, G. J., Hollander, M.-J., Onderwaater, W. G.,
cells, Proc. Natl. Acad. Sci. USA, 118, e2012170118,
Olsen, J. D., Hunter, C. N., et al. (2010) Light har-
doi: 10.1073/pnas.2012170118.
vesting, energy transfer and electron cycling of a na-
27.
Palazzo, G., Mallardi, A., Giustini, M., Monica, M. D.,
tive photosynthetic membrane adsorbed onto a gold
and Venturoli, G. (2000) Interactions of photosynthetic
surface, Biochim. Biophys. Acta,
1798,
637-645,
reaction center with
2,3-dimethoxy-5-methyl-1,4-
doi: 10.1016/j.bbamem.2009.12.018.
benzoquinone in reverse micelles, Phys. Chem. Chem.
18.
Woronowicz, K., Ahmed, S., Biradar, A. A., Biradar,
Phys., 20, 4624-4629, doi: 10.1039/b003905p.
A. V., Birnie, D. P., et al. (2012) Near-IR absorbing
28.
Friebe, V. M., Swainsbury, D. J. K., Fyfe, P. K.,
solar cell sensitized with bacterial photosynthetic
Heijden, W. V. D., Jones, M. R., and Frese, R. N.
membranes, Photochem. Photobiol., 88, 1467-1472,
(2016) On the mechanism of ubiquinone mediated
doi: 10.1111/j.1751-1097.2012.01190.x.
photocurrent generation by a reaction center based
19.
Harrold, J. W., Woronowicz, K., Lamptey, J.,
photocathode, Biochim. Biophys. Acta Bioenerg., 1857,
Baird, J., Moshar, A., et al.
(2013) Functional
1925-1934, doi: 10.1016/j.bbabio.2016.09.011.
interfacing of Rhodospirillum rubrum chromatophores
29.
Yaghoubi, H., Li, Z., Jun, D., Saer, R., and Slota,
to a conducting support for capture and conversion
J. E. (2012) The role of gold-adsorbed photosynthetic
of solar energy, J. Phys. Chem. B, 117, 11249-11259,
reaction centers and redox mediators in the charge
doi: 10.1021/jp402108s.
transfer and photocurrent generation in a bio-
20.
Vitukhnovskaya, L. A., Zaspa, A. A., Semenov, A. Yu.,
photoelectrochemical cell, J. Phys. Chem. C, 116,
and Mamedov, M. D. (2023) Conversion of light
24868-24877, doi: 10.1021/jp306798p.
into electricity in a semi-synthetic system based on
30.
Zdarta, J., Meyer, A. S., Jesionowski, T., and
photosynthetic bacterial chromatophores, Biochim.
Pinelo, M. (2018) A general overview of support
Biophys. Acta Bioenerg., 1864, 148975, doi: 10.1016/
materials for enzyme immobilization. Characteristics,
j.bbabio.2023.148975.
properties, practical utility, Catalysts,
8,
92,
21.
Ormerod, J. G., Ormerod, K. S., and Gest, H. (1961)
doi: 10.3390/catal8020092.
Light-dependent utilization of organic compounds
31.
Imam, H. T., Marr, P. C., and Marr, A. C. (2021)
and photoproduction of molecular hydrogen by
Enzyme entrapment, biocatalyst immobilization
photosynthetic bacteria; Relationships with nitrogen
without covalent attachment, Green Chem.,
23,
metabolism, Arch. Biochem. Biophys., 94, 449-463,
4980-5005, doi: 10.1039/D1GC01852C.
doi: 10.1016/0003-9861(61)90073-x.
32.
Grattieri, M., Rhodes, Z., Hickey, D. P., Beaver, K.,
22.
Woronowicz, K., Sha, D., Frese, R. N., and
and Minteer, S. D.
(2019) Understanding bio-
Niederman, R. A. (2011) The accumulation of the
photocurrent generation in photosynthetic purple bac-
light-harvesting 2 complex during remodeling of the
teria, ACS Catal., 9, 867-873, doi: 10.1021/acscatal.
Rhodobacter sphaeroides intracytoplasmic membrane
8b04464.
results in a slowing of the electron transfer turnover
33.
Zaspa, A. A., Vitukhnovskaya, L. A., Mamedova,
rate of photochemical reaction centers, Biochemistry,
A. M., Semenov, A. Y., and Mamedov, M. D. (2020)
50, 4819-4829, doi: 10.1021/bi101667e.
Photovoltage generation by photosystem II core
23.
Clayton, R. K.
(1966) Spectroscopic analysis of
complexes immobilized onto a Millipore filter on an
bacteriochlorophylls in vitro and in vivo, Photochem.
indium tin oxide electrode, J. Bioenerg. Biomembr.,
Photobiol.,
5,
669-677, doi:
10.1111/j.1751-1097.
52, 495-504, doi: 10.1007/s10863-020-09857-1.
1966.tb05813.x.
34.
Zaspa, A., Vitukhnovskaya, L., Mamedova, A.,
24.
Garcia, A. F., Venturoli, G., Gad’on, N., Fernandez-
Allakhverdiev, S. I., Semenov, A., and Mamedov,
Velasco, J. G., Melandri, B. A., and Drews, G.
M. D. (2022) Voltage generation by photosystem I
БИОХИМИЯ том 88 вып. 10 2023
ГЕНЕРАЦИЯ ЭЛЕКТРИЧЕСКОГО ОТВЕТА В ХРОМАТОФОРАХ
1741
complexes immobilized onto a millipore filter under
38. Mobius, K., Savitsky, A., Malferrari, M., Francia, F.,
continuous illumination, Int. J. Hydrogen Energy, 47,
Mamedov, M., et al. (2020) Soft dynamic confine-
11528-11538, doi: 10.1016/j.ijhydene.2022.01.175.
ment of membrane proteins by dehydrated trehalose
35. Jackson, J. B., and Crofts, A. R. (1971) The kinetics
matrices: high-field EPR and fast-laser studies, Appl.
of light induced carotenoid changes in rhodopseudo-
Magn. Reson.,
50,
773-850, doi:
10.1007/s00723-
monas spheroides and their relation to electrical field
020-01240-y.
generation across the chromatophore membrane,
39. Simongini, M., Puglisi, A., Genovese, F., and
Eur. J. Biochem., 18, 120-130, doi: 10.1111/j.1432-1033.
Hochkoeppler, H. (2023) Trehalose counteracts the
1971.tb01222.x.
dissociation of tetrameric rabbit lactate dehydrogenase
36. Michels, P., Hellingwerf, K. J., Lolkema, J. S.,
induced by acidic pH conditions, Arch. Biochem. Bio-
Friedberg, I., Konings, W. N. (1981) Effects of the
phys., 740, 109584, doi: 10.1016/j.abb.2023.109584.
medium composition on the components of the
40. Tang, M., Waring, A. J., and Hong, M.
(2007)
electrochemical proton gradient in Rhodopseudomo-
Trehalose-protected lipid membranes for determining
nas sphaeroides, Arch. Microbiol., 130, 357-361, doi:
membrane protein structure and insertion, J. Magn.
10.1007/BF00414600.
Reson., 184, 222-227, doi: 10.1016/j.jmr.2006.10.006.
37. Mamedov, M. D., Nosikova, E. S., Vitukhnovskaya,
41. De Moura Torquato, L. D., and Grattieri, M.
L. A., Zaspa, A. A., and Semenov, A. Yu. (2018)
(2022) Photobioelectrochemistry of intact photo-
Influence of the disaccharide trehalose on the
synthetic bacteria: advances and future outlook,
oxidizing side of photosystem II, Photosynthetica, 56,
Curr. Opin. Electrochem., 34, 101018, doi: 10.1016/
236-243, doi: 10.1007/s11099-017-0750-z.
j.coelec.2022.101018.
GENERATION OF ELECTRIC POTENTIAL DIFFERENCE
BY CHROMATOPHORES FROM PHOTOSYNTHETIC
BACTERIA IN THE PRESENCE OF TREHALOSE
UNDER CONTINUOUS ILLUMINATION
L. A. Vitukhnovskaya1,2, A. A. Zaspa1, and M. D. Mamedov1*
1 Belozersky Institute of Physico-Chemical Biology, Moscow State University,
119992 Moscow, Russia; e-mail: mahirmamedov@yandex.ru
2 Semenov Federal Research Center for Chemical Physics, Russian Academy of Sciences,
119991 Moscow, Russia
Measurement of the electrical potential difference (Δψ) in membrane vesicles (chromatophores) from
the purple bacterium Rhodobacter sphaeroides, associated with the surface of a nitrocellulose membrane
filter (MF), impregnated with phospholipid solution in decane, as well as immersed inside it, in the pres-
ence of exogenous mediators and disaccharide trehalose, demonstrated an increase in the amplitude
and stabilization of the signal under continuous illumination. The ascorbate/N,N,N′N′-tetramethyl-p-
phenylenediamine pair and ubiquinone-0 were used as mediators (electron donor and acceptor), respec-
tively. Although the stabilization of photoelectric responses under long-term continuous illumination was
observed in both cases of immobilization of chromatophores, only when the samples were immersed inside
the MF, the functional activity of the reaction centers was maintained for a month when they were stored
in the dark at room temperature. Stable photoresponses are probably associated with the preservation of
the integrity of chromatophore proteins inside MF pores. The stabilizing effect of the bioprotector treha-
lose may be due to both its effect on RC proteins and on the phospholipid bilayer membrane. The results
obtained will expand modern ideas about the use of semi-synthetic structures based on various intact photo-
synthetic systems capable of converting solar energy into an electrochemical form.
Keywords: chromatophores, reaction center, cytochrome bc1 complex, membrane filter, semiconductor, continuous
illumination, electrical potential
БИОХИМИЯ том 88 вып. 10 2023