БИОХИМИЯ, 2023, том 88, вып. 10, с. 1800 - 1817
УДК 577.352.5
ИЗМЕНЕНИЯ АКТИВНОСТИ Н+-АТРазы
ПЛАЗМАТИЧЕСКОЙ МЕМБРАНЫ КАК СВЯЗУЮЩЕЕ ЗВЕНО
МЕЖДУ ФОРМИРОВАНИЕМ ЭЛЕКТРИЧЕСКИХ СИГНАЛОВ
И РАЗВИТИЕМ ФОТОСИНТЕТИЧЕСКИХ ОТВЕТОВ
У ВЫСШИХ РАСТЕНИЙ
Обзор
© 2023 Е.М. Сухова, Л.М. Юдина, В.С. Сухов*
Нижегородский государственный университет им. Н.И. Лобачевского,
603022 Нижний Новгород, Россия; электронная почта: vssuh@mail.ru
Поступила в редакцию 22.06.2023
После доработки 05.09.2023
Принята к публикации 13.09.2023
Действие многочисленных неблагоприятных факторов окружающей среды на высшие растения
является пространственно-неоднородным; это означает, что для развития системного адаптаци-
онного ответа необходима генерация и распространение стрессовых сигналов. Вызванные локаль-
ным действием стрессоров электрические сигналы (ЭС), которые представлены потенциалом
действия, вариабельным потенциалом и системным потенциалом, играют важную роль в развитии
быстрых физиологических изменений на уровне целого растения, включая фотосинтетические
ответы. Развитие всех отмеченных электрических сигналов сопровождается изменениями актив-
ности Н+-ATPазы, которая является основной системой электрогенного транспорта протонов
на плазматической мембране. На основании литературных данных, представляется вероятным,
что изменения активности Н+-ATPазы и связанные с ними изменения вне- и внутриклеточного рН
играют ключевую роль в формировании вызванной ЭС инактивации фотосинтетических процес-
сов в интактных частях растений. Такая инактивация обусловлена как нарушением притока СО2
в клетки мезофилла листа, вызванным защелачиванием апопласта и, возможно, закислением цито-
плазмы, так и непосредственным влиянием закисления стромы и люмена хлоропластов на про-
цессы световой и, возможно, темновой стадии фотосинтеза. Конечным результатом вызванной ЭС
фотосинтетической инактивации является повышение устойчивости фотосинтетического аппара-
та к действию неблагоприятных факторов и вероятности выживания растения.
КЛЮЧЕВЫЕ СЛОВА: высшие растения, электрические сигналы, потенциал действия, вариабельный
потенциал, системный потенциал, Н+-ATPаза, внутриклеточный рН, внеклеточный рН, фотосинтез.
DOI: 10.31857/S0320972523100068, EDN: OSZEJJ
ВВЕДЕНИЕ
Вследствие этого у растений должны суще-
ствовать системы «дальних» стрессовых сиг-
Растения существуют в нестабильной окру-
налов, которые возникают в зоне локального
жающей среде и могут быть подвержены дей-
действия стрессора и распространяются в
ствию неблагоприятных факторов, многие из
интактные части растительного организма,
которых действуют пространственно-неодно-
вызывая адаптивные физиологические изме-
родно (например, избыточное освещение, вы-
нения. Электрические сигналы (ЭС) являются
сокие и низкие температуры, недостаток воды,
важной группой стрессовых сигналов, оказы-
биотическое поражение и многие другие). вающих влияние на широкий спектр физио-
логических процессов и приводящих к воз-
растанию устойчивости растения к действию
Принятые сокращения: АФК - активные формы кис-
неблагоприятных факторов [1-7].
лорода; ВП - вариабельный потенциал; ПД - потенциал
действия; СП - системный потенциал; ЭС - электриче-
ЭС представляют собой обратимые и спо-
ские сигналы.
собные к распространению по растению изме-
* Адресат для корреспонденции.
нения разности электрических потенциалов
1800
РОЛЬ Н+-АТРазы В ЭС И ОТВЕТАХ ФОТОСИНТЕЗА
1801
на плазматической мембране, основным ме-
растание устойчивости растительного орга-
тодом измерения которых является примене-
низма к действию неблагоприятных факторов
ние неполяризующихся электродов [1, 2]. При
среды [6, 7, 23, 31]. В частности, показано, что
этом в случае внеклеточного отведения изме-
распространение ЭС уменьшает повреждение
рительные электроды контактируют с различ-
клеточных мембран и фотосинтетического
ными участками поверхности растения, что
аппарата при избыточном освещении [32, 33],
позволяет более эффективно исследовать рас-
снижает негативное влияние тепла [33-39] и
пространение сигналов; в случае внутрикле-
холода [34] на фотосинтетические процессы, а
точного отведения - измерительный электрод
также повышает устойчивость на уровне цело-
вводится внутрь растительной клетки, что по-
го растения к низким [40] и высоким [36] тем-
зволяет более детально изучать параметры ЭС
пературам.
и их ионные механизмы.
Таким образом, ЭС играют важную роль в
Известно, что ЭС могут вызывать у выс-
системной адаптации высших растений к не-
ших растений стимуляцию экспрессии защит-
стабильным условиям окружающей среды [6,
ных генов (в частности, pin1, pin2 и vsp2,
7, 31]. Значительный вклад в такую адаптацию
участвующих в защите от насекомых [8, 9]),
вносят фотосинтетические ответы, развитие
усиление продукции стрессовых фитогормо-
которых важно для формирования вызван-
нов (например, абсцизовой и жасмоновой
ного ЭС повышения устойчивости растений
кислот [10-12]), активацию дыхания [13-15],
к действию неблагоприятных факторов [23].
возрастание содержания ATP в листьях [16],
Вследствие этого основной целью нашего об-
остановку потока ассимилятов во флоэме [17-
зора стал анализ путей формирования таких
19], изменения транспирации [20, 21], замед-
ответов при различных типах электрических
ление ростовых процессов [22] и многие дру-
сигналов, опирающийся на их ионные меха-
гие ответы. Роль ЭС в развитии подобных
низмы.
физиологических ответов подтверждается свя-
зью их параметров с характеристиками элек-
трических сигналов [2, 6]. Такая роль допол-
ТИПЫ ЭЛЕКТРИЧЕСКИХ СИГНАЛОВ
нительно показана путем выявления индукции
РАСТЕНИЙ И РОЛЬ H+-ATPазы
физиологических ответов при имитации ге-
ПЛАЗМАТИЧЕСКОЙ МЕМБРАНЫ
нерации ЭС (обработка ингибиторами про-
В ИХ ФОРМИРОВАНИИ
тонного транспорта или протонофорами) и
подавления развития ответов при блокирова-
Потенциал действия (ПД), вариабельный
нии распространения сигнала локальным дей-
потенциал (ВП) и системный потенциал (СП)
ствием низких температур или азида натрия
являются основными типами ЭС у высших ра-
на стебель [2, 6, 13, 23]. В частности, показано,
стений [5-7].
что экспрессия защитных генов наблюдается
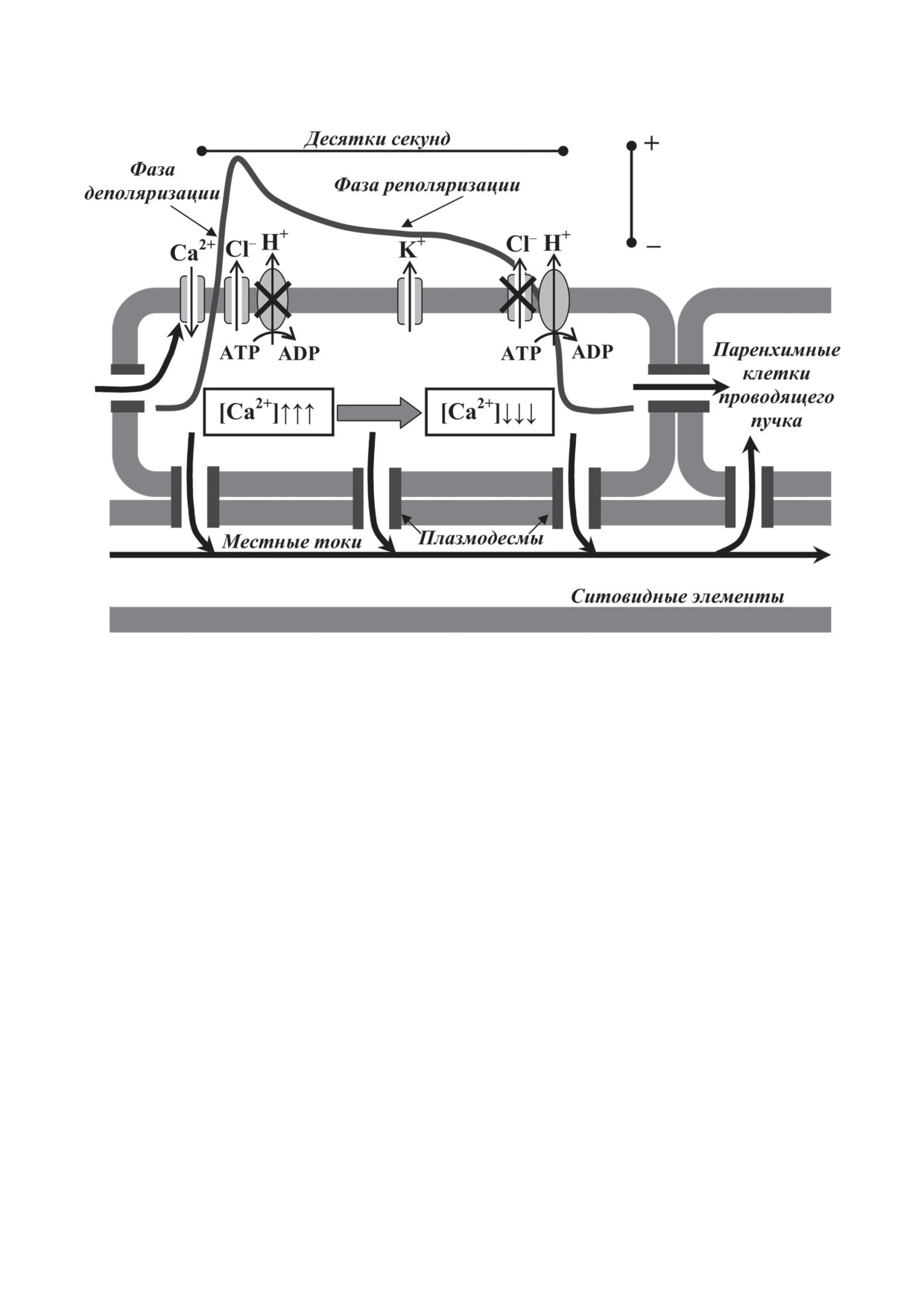
ПД представляет собой относительно
только в тех листьях арабидопсиса, в которые
короткую импульсную реакцию (десятки се-
проходят ЭС с большой длительностью или
кунд), возникающую при локальном действии
которые подвержены прямому действию элек-
неповреждающих стрессоров (например, охла-
трического тока [9].
ждение на несколько градусов) и включающую
В качестве отдельной «мишени» для дей-
в себя две фазы (деполяризация и реполяриза-
ствия ЭС можно отметить фотосинтетические
ция) [1, 6, 7] (рис. 1). У высших растений та-
процессы, так как известно, что электриче-
кой сигнал характеризуется наличием порога
ские сигналы могут вызывать как быструю (5-
генерации, подчинением закону «все или ни-
10 мин), так и длительную (десятки минут и,
чего», длительным периодом рефрактерности
возможно, часы) инактивацию фотосинтеза [6,
(в течение часов) и способностью к активному
7, 23]. Такая инактивация проявляется в сниже-
распространению по ситовидным элементам
нии фотосинтетической ассимиляции CO2 [10,
и/или по симпласту паренхимных клеток про-
11, 16, 20, 24], квантовых выходов фотосистем I
водящих пучков растений за счет идущих по
и II [25-27] и нециклического потока электро-
плазмодесмам местных электрических токов.
нов [28] и в возрастании нефотохимического
Следует отметить, что механизмы генера-
тушения флуоресценции хлорофилла [14, 27-
ции ПД являются сходными у высших расте-
29] (в частности, энергозависимой компонен-
ний и харовых водорослей, вследствие чего
ты [30]), а также циклического потока электро-
ионные механизмы ПД и связь его генерации
нов вокруг фотосистемы I [28].
с изменениями активности протонного транс-
Предполагаемым результатом вызванных ЭС
порта были впервые показаны у харовых водо-
физиологических изменений является воз-
рослей [41-44] и подтверждены в дальнейшем
БИОХИМИЯ том 88 вып. 10 2023
1802
СУХОВА и др.
Рис. 1. Потенциал действия у высших растений и схема механизмов его генерации и распространения по проводящим
пучкам растений
у высших растений. Фактором, инициирую-
мальный нагрев, механическое повреждение)
щим генерацию ПД, является деполяризация
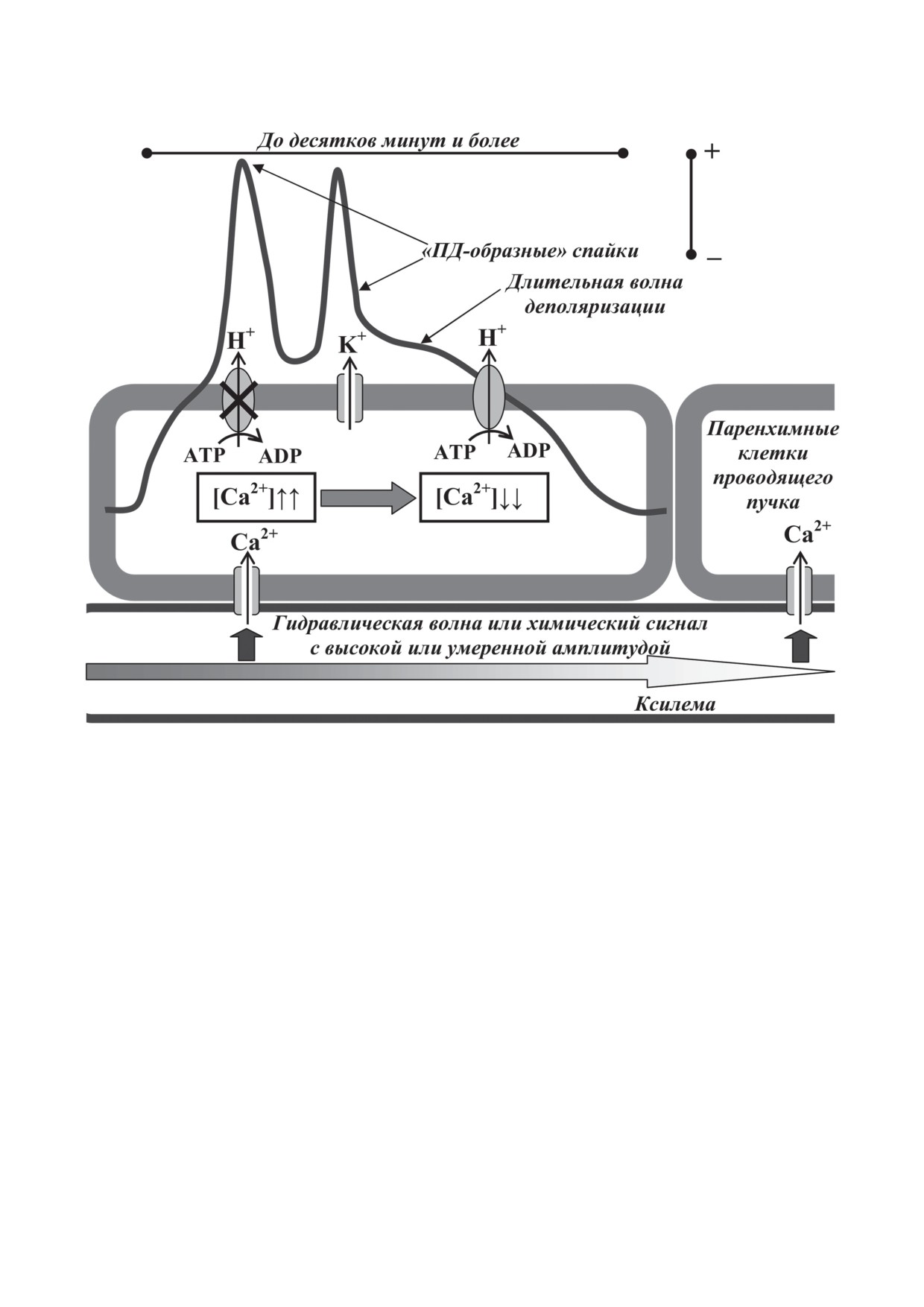
и имеет большую длительность (до десят-
мембранного потенциала до порогового уров-
ков минут и более) и сложную форму [6, 23,
ня, приводящая к активации потенциал-зави-
48], включающую две компоненты: длитель-
симых Ca2+-каналов плазматической мембра-
ную волну деполяризации и «ПД-образные»
ны и к входу ионов кальция в клетку [1, 45-47].
спайки, присутствие которых не является обя-
Значительное повышение концентрации Ca2+
зательным (рис.
2).
«ПД-образные» спайки
в цитоплазме [47] приводит к активации ани-
представляют собой типичные потенциалы
онных каналов и выходу Cl- из клетки [45],
действия, вызванные положительным сдвигом
а также к временной инактивации H+-ATPазы
мембранного потенциала в ходе формирова-
плазматической мембраны [46]; результатом
ния длительной волны деполяризации, кото-
обоих процессов является формирование фазы
рые имеют такие же ионные механизмы, как
деполяризации ПД. Деполяризация приводит
и ПД [48]; в то же время именно длительная
к активации outwardly-rectifying K+-каналов,
волна является основным компонентом ВП.
обеспечивающих выход ионов калия из цито-
Важной особенностью ВП (прежде всего, дли-
плазмы во внеклеточную среду [47]. Значи-
тельной волны деполяризации) является сни-
тельная деполяризация приводит также к
жение скорости его распространения [49, 50]
закрытию Ca2+-каналов, уменьшению кон-
и/или амплитуды [16, 29, 50] с увеличением
центрации ионов кальция в цитоплазме и, как
расстояния от зоны повреждения.
следствие, к снижению активности анионных
ВП является местной электрической ре-
каналов и реактивации H+-ATPазы [47]. Такие
акцией на распространение сигнала неэлек-
процессы приводят к формированию фазы
трической природы [48], который может быть
реполяризации и возвращению мембранного
гидравлическим или химическим. Гидравличе-
потенциала к исходным значениям.
ская гипотеза [22, 48, 51, 52] предполагает, что
ВП возникает при действии повреждаю-
при локальных повреждениях в зоне действия
щих раздражителей (включая ожог, экстре- стрессора формируется область повышенного
БИОХИМИЯ том 88 вып. 10 2023
РОЛЬ Н+-АТРазы В ЭС И ОТВЕТАХ ФОТОСИНТЕЗА
1803
Рис. 2. Вариабельный потенциал у высших растений и схема потенциальных механизмов его генерации и распростра-
нения по проводящим пучкам растений. На рисунке приведена схема формирования длительной волны деполяриза-
ции, связанной с сильным или умеренным повышением концентрации Са2+ в цитоплазме. Механизмы «ПД-образ-
ных» спайков аналогичны механизмам ПД (см. рис. 1) и не показаны на рисунке
давления, индуцирующая распространение по
Ca2+-каналов и развитие ВП [48]; при низкой
растению затухающей гидравлической вол-
величине такого сигнала индукция вариабель-
ны, которая вызывает активацию механо-
ного потенциала отсутствует. В качестве потен-
чувствительных Ca2+-каналов и последующее
циального раневого вещества часто рассматри-
развитие ВП [44]. Экспериментальные [51] и
вается Н2О2 [54-56]; однако нельзя исключать
теоретические [53] работы показывают, что
участие других соединений (например, систе-
уменьшение амплитуды гидравлической вол-
мина [57] или глутамата [58]). Ключевой про-
ны приводит к возрастанию лаг-фазы перед
блемой химической гипотезы является низкая
началом изменений электрического потен-
скорость распространения раневого вещества
циала и тем самым вызывает замедление ка-
посредством молекулярной диффузии
[48].
жущейся скорости распространения ВП с
Существует несколько подходов, позволяющих
увеличением расстояния от зоны действия
преодолеть такое ограничение. Во-первых, в
стрессора. При низкой амплитуде гидравличе-
соответствии с гипотезой Malone [59], которая
ской волны ВП не возникает.
была теоретически проверена на серии мате-
Альтернативная гипотеза предполагает,
матических моделей [60, 61], локальное повы-
что в зоне повреждения происходит выделе-
шение давления в зоне повреждения может
ние специального химического соединения
приводить к возникновению дополнительных
(«раневое вещество»), распространение кото-
водных потоков по ксилеме, которые перено-
рого вызывает активацию лиганд-зависимых
сят раневое вещество в неповрежденные части
БИОХИМИЯ том 88 вып. 10 2023
1804
СУХОВА и др.
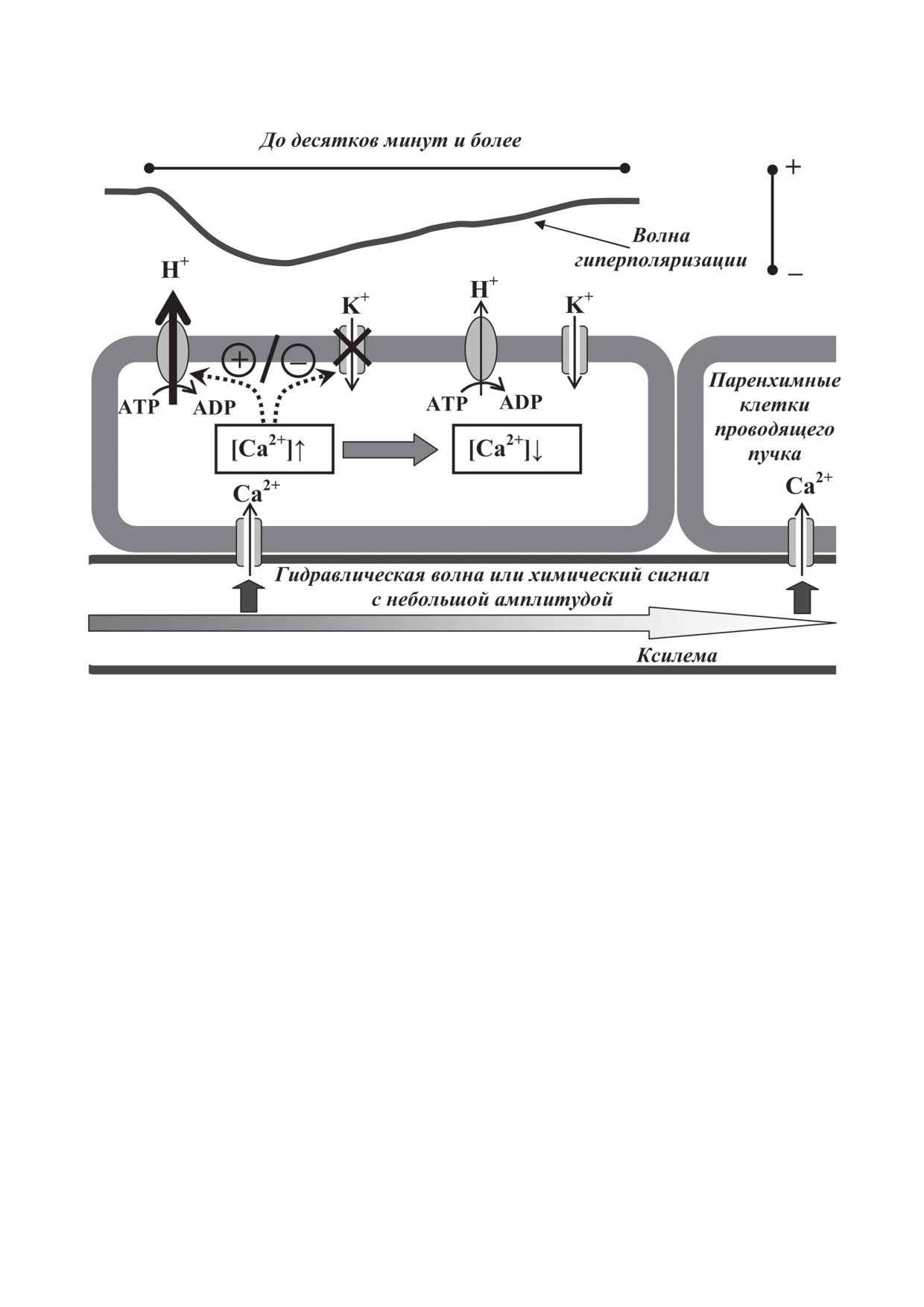
Рис. 3. Системный потенциал у высших растений и схема возможных механизмов его генерации и распространения
по проводящим пучкам растений. Пунктирными линиями показано возможное влияние небольшого повышения кон-
центрации Са2+ в цитоплазме на H+-ATPазу (активация) и на inwardly-rectifying K+-каналы плазматической мембраны
(инактивация)
растения. Во-вторых, нами ранее была пока-
свидетельствуют как экспериментальные дан-
зана возможность ускоренного распростране-
ные [63], показывающие распространение вол-
ния химического агента по растению, которое,
ны АФК (повышение их концентрации) и по-
вероятно, связано с усилением конвективной
давление такой волны при действии ингибито-
(турбулентной) диффузии в потоке жидко-
ров оксидаз RbohD, так и результаты матема-
сти по ксилеме [49, 62]. Следует отметить, что
тического моделирования.
несмотря на относительно низкие скорости
Считается, что распространение хими-
движения жидкости по сосудам ксилемы, тур-
ческого сигнала или гидравлической волны
булентные потоки могут потенциально воз-
приводит к долговременной активации ли-
никать в результате периодических сужений
ганд-зависимых или механочувствительных
их диаметра [62]. В-третьих, согласно гипотезе
Ca2+-каналов плазматической мембраны со-
Mittler et al. [54], развитой в ряде последующих
ответственно и входу ионов кальция в клет-
работ [55, 56, 63], при распространении хими-
ку [48]. Умеренное (или сильное) и долго-
ческого сигнала может происходить вторич-
временное повышение концентрации Ca2+ в
ная генерация Н2О2 (базирующаяся на актива-
цитоплазме (500-600 нМ и более [49]) при-
ции Са2+-каналов и возрастании концентрации
водит к инактивации Н+-ATPазы [64] и фор-
Са2+, что приводит к активации NADPН окси-
мированию длительной волны деполяризации.
даз RbohD в плазматической мембране, кото-
На ее фоне мембранный потенциал может
рые продуцируют активные формы кислоро-
достигать пороговых значений и активировать
да (АФК)); такая вторичная генерация может
потенциал-зависимые Ca2+-каналы, форми-
ускорять распространение химического сиг-
руя «ПД-образные» спайки, механизм которых
нала по растению. В пользу данной гипотезы
аналогичен механизму ПД [48, 49].
БИОХИМИЯ том 88 вып. 10 2023
РОЛЬ Н+-АТРазы В ЭС И ОТВЕТАХ ФОТОСИНТЕЗА
1805
СП, который представляет собой рас-
ранее проведенным теоретическим анализом
пространяющуюся волну гиперполяризации
разработанной нами математической модели
(рис. 3), является наименее изученным элек-
электрогенеза клетки высшего растения [47]),
трическим сигналом у растений [6]. Известно,
снижение проницаемости K+-каналов должно
что СП может возникать при локальном дей-
приводить к гиперполяризации. При этом раз-
ствии широкого спектра стрессоров, включая
витие гиперполяризации будет обусловлено
неорганические соли, умеренный и сильный
уменьшением вклада электрической проводи-
нагрев, ожог, механическое повреждение и
мости inwardly-rectifying K+-каналов в общую
атаки насекомых [9, 65-71]. При этом форма
электрическую проводимость плазматической
и длительность СП может существенно варь-
мембраны. Последнее может быть подтвер-
ировать в зависимости от типа стрессора,
ждено изменениями мембранного потенциа-
вида растения и расстояния от зоны повреж-
ла (EM), упрощенно описанного уравнением:
дения [9, 65-71]. Следует отметить, что СП
EP + gK/gP EK,
может быть тесно связан с ВП, так как (i) ряд
EM =
1 + gK/gP
стрессоров вызывают генерацию ВП вблизи
зоны воздействия на растение, а на удалении от
где EP
- электродвижущая сила (ЭДС)
нее наблюдается появление СП [9, 67, 69-71];
Н+-ATPазы (около -450 мВ) и EK - потенциал
(ii) локальный ожог вызывает ВП у растений с
Нернста для ионов калия (около -100 мВ; на
регулярным поливом, но может вызывать СП
основе [49, 76, 77]) в условиях уменьшения
при сильном водном дефиците [69]; (iii) стрес-
собственной проводимости К+-каналов (gK)
соры могут вызывать ВП при первом дей-
по отношению к собственной проводимо-
ствии, а при повторном - СП [66]; (iv) в зави-
сти Н+-ATPазы (gP). Приведенное уравнение,
симости от участка растения действие одних
опирающееся на параллельное электрическое
и тех же стрессоров может вызывать как ВП,
соединение inwardly-rectifying K+-каналов и
так и СП [65].
Н+-ATPазы, показывает стационарное реше-
Считается, что основным механизмом ге-
ние для мембранного потенциала.
нерации СП является активация H+-ATPазы
Гипотеза об инактивации inwardly-recti-
плазматической мембраны [66], что подтвер-
fying K+-каналов хорошо согласуется с по-
ждается угнетением сигнала при предвари-
давлением генерации СП при действии
тельном ингибировании этого транспортера
специфичного блокатора К+-каналов, тетра-
(ортованадат натрия) и индукцией СП при
этиламмония
[65]. Важным преимуществом
локальном действии активатора H+-ATPазы
предложенной гипотезы является ее способ-
(фузикокцин); однако природа сигнала, вызы-
ность объяснить связь ВП и СП, так как при
вающего такую активацию, остается дискусси-
уменьшении амплитуды гидравлической вол-
онной. Ранее мы предположили [69-71], что
ны (с уменьшением силы стимула при раз-
передача СП связана с распространением гид-
витии засухи или с увеличением расстояния
равлического сигнала с малой амплитудой, ко-
от зоны стимуляции [51, 53, 68-71]) должен
торый активирует механочувствительные Са2+-
уменьшаться вход Са2+ в клетку, что приво-
каналы плазматической мембраны и вызывает
дит к изменению типа ЭС: длительная волна
небольшое возрастание концентрации ионов
деполяризации (при сильном или умеренном
кальция в цитоплазме. Существуют отдельные
повышении концентрации ионов кальция в
электрофизиологические данные [72, 73], по-
цитоплазме) сменяется волной гиперполяри-
казывающие, что небольшое возрастание кон-
зации (при небольшом возрастании концен-
центрации Са2+ может вызывать активацию
трации Са2+). Следует, однако, отметить, что
Н+-ATPазы плазматической мембраны; и, та-
предложенный механизм потенциально совме-
ким образом, происходит гиперполяризация.
стим и с химической гипотезой распростране-
С другой стороны, ряд работ показывает, что
ния сигнала; в этом случае будет происходить
небольшое возрастание концентрации Са2+ в
индукция входа ионов кальция через лиганд-
цитоплазме (200-400 нМ) вызывает инактива-
зависимые Са2+-каналы [48].
цию inwardly-rectifying K+-каналов плазмати-
Таким образом, общей чертой ПД, ВП
ческой мембраны [74, 75], которые обеспечи-
и СП является участие изменений активности
вают вход ионов калия из внеклеточной среды
Н+-ATPазы плазматической мембраны в их
в цитоплазму клетки. Учитывая, что inwardly-
генерации, что хорошо согласуется с ключе-
rectifying K+-каналы и Н+-ATPаза плазматиче-
вой ролью этих ATPаз Р-типа в поддержании
ской мембраны являются основными систе-
ряда физиологических процессов растений
мами электрогенного ионного транспорта в
и рецепции действия факторов окружающей
условиях покоя [47, 76, 77] (в соответствии с
среды [78, 79], включая изменения активности
БИОХИМИЯ том 88 вып. 10 2023
1
806
СУХОВА и др.
Н+-ATPазы при действии абиотических и био-
степени связаны с наличием противополож-
тических стрессоров. При этом ключевым ме-
но направленных протонных потоков. В соот-
ханизмом изменения активности Н+-ATPазы
ветствии с ранее разработанной нами матема-
является фосфорилирование аминокислотных
тической моделью электрогенеза клетки выс-
остатков R-домена на С-конце фермента [78,
шего растения [47, 77], такие потоки могут
79]: фосфорилирование Thr947 обеспечивает
быть обусловлены активностями H+/K+-анти-
присоединение белков 14-3-3 к С-концу и вы-
портера и 2H+/Cl--симпортера и утечкой про-
зывает активацию фермента; напротив, фос-
тонов через плазматическую мембрану. В част-
форилирование ряда других аминокислотных
ности, подобные потоки могут обеспечивать
остатков нарушает связывание белков 14-3-3
быстрый транспорт протонов внутрь клетки
и приводит к инактивации Н+-ATPазы. Следу-
при снижении компенсирующего выхода H+ в
ет отметить, что Са2+-зависимая инактивация
условиях инактивация Н+-ATPазы. Потенци-
Н+-ATPазы, которая предотвращается при об-
ально, отмеченные противоположные потоки
работке ингибитором протеинкиназы H-7 [46],
протонов также могут регулироваться теми или
связана, по-видимому, с фосфорилированием
иными факторами. Однако такая регуляция не
одного из аминоксилотных остатков (Ser944,
является необходимым условием для реали-
Thr942, Tyr946) и нарушением присоединения
стичного описания электрических ответов ра-
белков 14-3-3 к С-концу фермента [79].
стений при моделировании [47, 76, 77], вслед-
По-видимому, непосредственный вклад
ствие чего гипотеза о регуляции направленных
Н+-ATPазы в формирование электрических
внутрь потоков H+ является, скорее, избыточ-
сигналов варьирует. Так, в случае ПД исклю-
ной в случае клеток высших растений.
чение инактивации Н+-ATPазы слабо влияет
В целом участие изменений активности
на форму и амплитуду сигнала, но полностью
Н+-ATPазы в генерации всех основных ти-
подавляет сопровождающее ПД защелачи-
пов ЭС позволяет предположить, что такие
вание апопласта [46, 47]. С другой стороны,
изменения могут играть определенную физио-
инактивация Н+-ATPазы является основным
логическую роль; в частности, значительный
механизмом генерации ВП [48], так как сни-
интерес вызывает их участие в регуляции фо-
жение активности этого транспортера приво-
тосинтетических процессов электрическими
дит к существенному подавлению длительной
сигналами у высших растений [23].
волны деполяризации, составляющей основу
вариабельного потенциала [49, 64, 80].
В случае СП наблюдается более сложная
ИЗМЕНЕНИЯ АКТИВНОСТИ H+-ATPазы
картина. Базовая гипотеза [66, 67, 69-71], опи-
ПЛАЗМАТИЧЕСКОЙ МЕМБРАНЫ
сывающая механизмы СП, предполагает воз-
КАК МЕХАНИЗМ ВЛИЯНИЯ
растание абсолютной величины активности
ЭЛЕКТРИЧЕСКИХ СИГНАЛОВ
Н+-ATPазы; однако такое предположение про-
НА ФОТОСИНТЕЗ
тиворечит данным о защелачивании апоплас-
та во время развития системного потенциа-
Как отмечалось выше, существует ряд аргу-
ла [66]. Аналогичное защелачивание, которое
ментов в пользу сходного снижения абсолют-
наблюдается при инактивации Н+-ATPазы во
ной активности Н+-ATPазы плазматической
время генерации ПД [46, 47] и ВП [25, 27], по-
мембраны при генерации всех трех типов ЭС;
казывает, скорее, снижение активности этого
прежде всего, это наблюдаемое при генера-
транспортера. Предположение о возрастании
ции ПД [46, 47], ВП [25, 27] и СП [66] заще-
относительной активности Н+-ATPазы вслед-
лачивание апопласта. В том случае, если такое
ствие Ca2+-зависимого снижения активности
снижение действительно связано с вызван-
inwardly-rectifying K+-каналов [74, 75] в боль-
ной ЭС инактивацией фотосинтеза, можно
шей степени согласуется с такими данными,
ожидать, что подобная инактивация будет на-
так как развитие гиперполяризации должно
блюдаться при распространении всех трех ти-
снижать активный транспорт протонов из
пов ЭС. Данные литературы показывают, что
клетки и приводить к защелачиванию апо-
ПД [14, 81, 82], ВП [16, 24-30, 83] и СП [66, 71]
пласта вследствие возрастания электрохими-
действительно могут вызывать сходное изме-
ческого градиента протонов, направленного
нение интенсивности фотосинтетических про-
против такого транспорта.
цессов у растений, включая, в частности, сни-
Завершая раздел, следует отдельно отметить,
жение интенсивности ассимиляции СО2, умень-
что изменения pH, сопровождающие измене-
шение квантового выхода фотосистемы II
ния активности Н+-ATPазы плазматической
и увеличение нефотохимического тушения
мембраны при развитии ЭС, в значительной
флуоресценции хлорофилла.
БИОХИМИЯ том 88 вып. 10 2023
РОЛЬ Н+-АТРазы В ЭС И ОТВЕТАХ ФОТОСИНТЕЗА
1807
Вызванные ЭС ответы фотосинтеза вклю-
Прежде всего, следует отметить, что такая ин-
чают в себя быструю (5-10 мин) и длительную
активация может быть следствием всех осталь-
(десятки минут и, возможно, часы) инактива-
ных приведенных механизмов. В частности,
цию [6, 7, 23]. В некоторых случаях у растений
известно, что повышение концентрации ионов
может наблюдаться только длительная фото-
кальция может приводить к подавлению ак-
синтетическая инактивация (см., например,
тивности Н+-ATPазы плазматической мембра-
работы Hlavácková et al. [10], Herde et al. [84]
ны [86], распространение Н2О2 считается свя-
и Sherstneva et al. [85]). В ряде работ пока-
занным с активацией Са2+-каналов, входом
зано [14, 26, 35], что снижение концентрации
ионов кальция и инактивацией Н+-ATPазы [6,
СО2 в среде приводит к инактивации световой
54-56]. Действие же абсцизовой кислоты на
стадии фотосинтеза, сходной с вызванной ЭС,
фотосинтез также может опосредоваться сни-
и уменьшает амплитуду индуцированных элек-
жением активности Н+-ATPазы [87, 88].
трическими сигналами фотосинтетических от-
Результаты наших предыдущих работ [88,
ветов; однако полного подавления быстрых и
89] показывают, что умеренное возрастание
длительных изменений параметров световой
или снижение исходной активности Н+-ATPазы
стадии не происходит. На основании этого
при предварительной обработке растений ак-
было предположено [6, 23], что быстрая и, воз-
тиваторами (фузикокцин) или ингибиторами
можно, длительная инактивация фотосинтеза
(ортованадат натрия) этого ионного транспор-
связаны с двумя различными группами меха-
тера приводит, соответственно, к возрастанию
низмов.
или снижению фотосинтетического ответа, воз-
(i) Со снижением интенсивности темновой
никающего при последующей индукции ВП.
стадии фотосинтеза, приводящим к уменьше-
Такие результаты подтверждают участие Н+-
нию отношений ADP/ATP и NADP+/NADPH
ATPазы в формировании вызванных ЭС изме-
в строме хлоропластов и, соответственно, к
нений фотосинтеза, так как при сохранении
угнетению процессов световой стадии вслед-
постоянной относительной величины измене-
ствие роста электрохимического градиента на
ния ее активности абсолютная величина тако-
тилакоидной мембране (результат снижения
го изменения будет возрастать при предвари-
активности Н+-ATP-синтазы в условиях не-
тельной активации Н+-ATPазы или снижаться
хватки субстрата) и нарушения «стока» элек-
при ее предварительной инактивации, при-
тронов из электрон-транспортной цепи [14, 26].
водя к увеличению или снижению амплитуды
Вероятными механизмами подавления темно-
вызванного ЭС фотосинтетического ответа
вой стадии фотосинтеза являются уменьшение
соответственно.
проводимости мезофилла листа для СО2 [24] и,
Сопровождающие инактивацию Н+-ATPазы
возможно, закрытие устьиц [21].
изменения внутри- и внеклеточного рН рас-
(ii) С подавлением световой стадии фото-
сматриваются как вероятный механизм индук-
синтеза, не вызванным замедлением функ-
ции ЭС фотосинтетических ответов [2, 6, 23].
ционирования цикла Кальвина, которое, по-
Существует ряд аргументов, подтверждающих
тенциально, также может иметь различные
такую гипотезу. Во-первых, значительное чис-
механизмы (в частности, возрастание нефото-
ло работ показывает, что генерация различных
химического тушения [27, 29] и снижение ак-
типов ЭС у высших растений сопровождается
тивности ферредоксин-NADP+-редуктазы [26]).
возрастанием pH в апопласте и снижением pH
Существует несколько гипотез, объяс-
в цитоплазме [25, 27, 45, 46, 66, 85]. Более того,
няющих механизмы инактивации фотосинте-
непрямыми методами выявлено сопровождаю-
за [23]. Среди них можно отметить подавление
щее генерацию ВП закисление стромы и лю-
активности ферментов цикла Кальвина вслед-
мена хлоропластов [90]. При этом некоторые
ствие поступления Са2+ в цитоплазму и далее,
работы [91, 92] показывают наличие сильной
в строму хлоропластов [81] и инактивирующее
связи между изменениями pH и параметров
влияние АФК (вероятно, Н2О2) [83]. В случае
фотосинтеза, подтверждая их роль в индукции
длительной инактивации фотосинтеза также
фотосинтетических ответов. Во-вторых, пока-
рассматривается участие стрессовых фитогор-
зано, что искусственная индукция изменения
монов (абсцизовой и жасмоновой кислоты [10,
pH путем действия протонофоров на участки
11, 50]), которые приводят к закрытию устьиц
листа [27] или путем ингибирования активно-
и подавлению фотосинтеза. В то же время об-
сти Н+-ATPазы у протопластов ортованадатом
ратимая инактивация Н+-ATPазы плазматиче-
натрия [89] приводит к индукции фотосинте-
ской мембраны выглядит наиболее вероятным
тических ответов, сходных с наблюдаемыми
механизмом развития вызванных ЭС фотосин-
при распространении ЭС (снижение кван-
тетических ответов у высших растений [6, 23].
тового выхода фотосистемы II и возрастание
БИОХИМИЯ том 88 вып. 10 2023
1808
СУХОВА и др.
нефотохимического тушения). В-третьих, за-
важную роль в поступлении углекислого газа в
кисление среды инкубации хлоропластов выс-
клетки высших растений [98] и могут регули-
ших растений, имитирующее снижение pH
роваться pH [99-101]. Такая гипотеза хорошо
в цитоплазме при развитии ЭС, также инду-
согласуется с результатами моделирования [97],
цирует фотосинтетические ответы, близкие
показывающими зависимость стационарной
к ответам, вызванным электрическими сиг-
интенсивности фотосинтеза от проводимости
налами [25, 27, 85]; амплитуда таких ответов
плазматической мембраны для СО2; однако
линейно коррелирует с величиной измене-
данные литературы [99-101] показывают, что
ний pH [27, 85]. Аналогичный эффект был вы-
проводимость аквапоринов снижается при за-
явлен при перфузии клеток харовых водорос-
кислении цитоплазмы, а не при защелачивании
лей растворами с пониженным pH [93].
апопласта. Несмотря на то что оба процесса
Наши предыдущие работы показывают
сопровождают генерацию ЭС [25, 27], наши
[91, 92], что сопровождающее ВП возраста-
данные показывают [91, 92] слабую связь закис-
ние внеклеточного pH связано, прежде все-
ления цитоплазмы со снижением интенсивно-
го, со снижением фотосинтетической асси-
сти темновой стадии фотосинтеза; т.е. имеется
миляции СО2, в то время как уменьшение
определенное противоречие между гипотезой
внутриклеточного pH связано с увеличением
об участии аквапоринов в развитии вызван-
нефотохимического тушения флуоресценции.
ных ЭС фотосинтетических ответов и экспери-
На основании этого можно предположить [6,
ментальными особенностями таких ответов.
23], что вызванное ЭС защелачивание апо-
Таким образом, вопрос о конкретных ме-
пласта влияет преимущественно на поступле-
ханизмах влияния изменений pH на поступ-
ние СО2 в клетки, а закисление цитоплазмы,
ление СО2 в клетки мезофилла остается от-
стромы и люмена хлоропластов может непо-
крытым. Потенциально, выявленные противо-
средственно влиять на реакции световой ста-
речия могут быть объяснены неоднородным
дии фотосинтеза. Такое предположение хоро-
пространственным распределением карбоан-
шо согласуется с вызванным ЭС уменьшением
гидраз, которые значительно ускоряют взаим-
проводимости мезофилла листа для СО2 [24] и
ное превращение между СО2 и НСО–, так как
с наличием индуцированных электрическими
такая неоднородность может обеспечить ло-
сигналами изменений световой стадии фото-
кальные pH-зависимые изменения соотноше-
синтеза, которые не были связаны с инактива-
ния нейтральной и заряженной форм углекис-
цией темновой стадии [26, 27, 29].
лого газа, что будет влиять на его прохождение
В качестве простого механизма влияния
через плазматическую мембрану без существен-
защелачивания апопласта на поступление СО2
ного изменения средней концентрации таких
в фотосинтезирующую клетку можно предпо-
форм в апопласте (в предложенной фотосин-
ложить pH-зависимое снижение отношения
тетической модели листа [96, 97] подобная воз-
СО2/НСО– [23], так как заряженная форма зна-
можность не учитывалась). Отдельные работы,
чительно хуже проникает через биологические
показывающие совместное функционирование
мембраны по сравнению с нейтральной [94].
карбоангидраз и аквапоринов в плазматиче-
Такое предположение было предварительно
ской мембране [102] и влияние карбоангидраз
подтверждено анализом упрощенной матема-
на проводимость мезофилла для СО2 [103], яв-
тической модели, описывающей влияние ин-
ляются аргументом в пользу такой гипотезы.
активации Н+-ATPазы на долю нейтральной
Следует отметить, что индуцированное ЭС
формы СО2 [95]; однако дальнейшее иссле-
закрытие устьиц, которое может участвовать в
дование более детализированной фотосинте-
длительной инактивации фотосинтетических
тической модели листа не подтвердило суще-
процессов [21], также связано с подавлением
ственного вклада этого механизма в снижение
активности Н+-ATPазы, так как величина от-
активности фотосинтеза при защелачивании
вета может быть модифицирована изменением
апопласта (по крайней мере для стационарных
начальной активности этого транспортера [36].
потоков СО2 при однородном описании апо-
В то же время непосредственным механиз-
пласта и при использованных значениях пара-
мом закрытия устьиц является, скорее, не ин-
метров модели) [96, 97].
активация Н+-ATPазы, а вызванные такой ин-
Альтернативная гипотеза, описывающая
активацией изменения активности ионных
влияние вызванных ЭС изменений pH на про-
каналов, приводящие к выходу ионов из замы-
водимость мезофилла листа для СО2, была
кающих клеток. В частности, в ряде работ по-
предложена в работе Gallé et al. [24]. В соот-
казан значительный выход ионов хлора, сопро-
ветствии с ней такие изменения влияют на
вождающий развитие электрических сигналов
проводимость аквапоринов, которые играют
различного типа [6, 7, 48, 66].
БИОХИМИЯ том 88 вып. 10 2023
РОЛЬ Н+-АТРазы В ЭС И ОТВЕТАХ ФОТОСИНТЕЗА
1809
Непосредственное влияние закисления ци-
что в условиях отсутствия освещения и, соот-
топлазмы на параметры световой стадии фо-
ветственно, изменения активности темновой
тосинтеза обусловлено, по-видимому, про-
стадии фотосинтеза, ЭС могут приводить к из-
хождением протонов из цитоплазмы в строму
менению распределения поглощенного света
и люмен хлоропластов [23]. На основе изме-
между фотосистемами I и II.
рения электрохромного сдвига поглощения
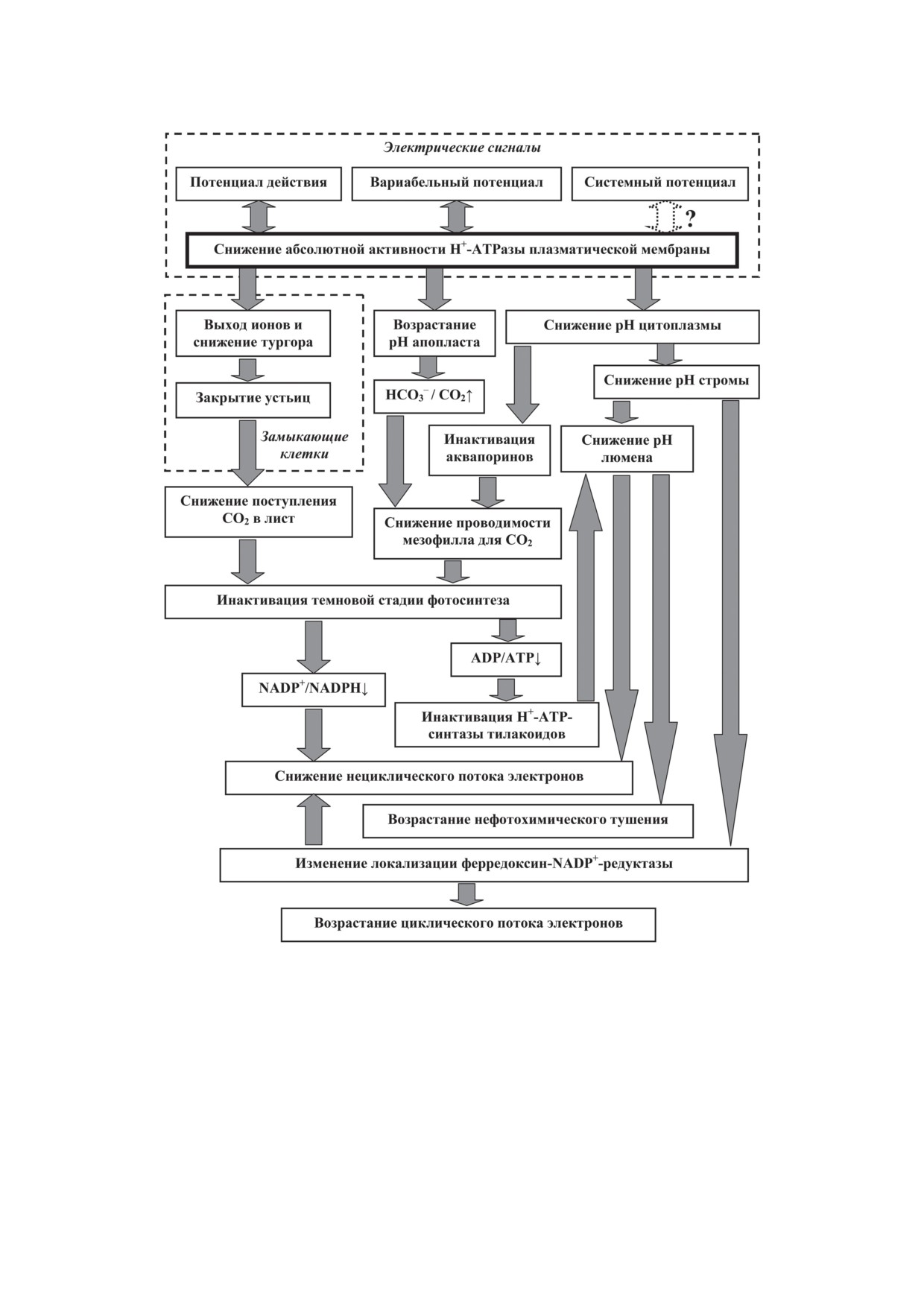
Таким образом, снижение абсолютной
света листом и светорассеяния на длине волны
величины активности Н+-ATPазы плазматиче-
535 нм нами ранее было косвенно показано
ской мембраны, сопровождающее генерацию
сопровождающее развитие ВП снижение pH
ПД, ВП и, возможно, СП, приводит к возра-
в строме и люмене [90], которое подтверждает
станию внеклеточного и уменьшению внутри-
транспорт протонов через оболочки хлоро-
клеточного pH и, как следствие, вызывает фо-
пластов и тилакоидные мембраны.
тосинтетическую инактивацию, включающую
В соответствии с работами Alte et al. [104]
в себя ряд связанных друг с другом механиз-
и Benz et al. [105], снижение pH стромы хло-
мов (рис. 4).
ропластов может приводить к увеличению
Вероятным результатом вызванных ЭС
сродства ферредоксин-NADP+-редуктазы со
изменений фотосинтеза в интактных частях
специальными сайтами связывания (Tic62
растения является повышение устойчиво-
и TROL), что обеспечивает пространственное
сти фотосинтетического аппарата к действию
удаление фермента от фотосистемы I и тем
стрессоров на эти части [6, 7, 23, 31]. Подобное
самым снижение потока электронов на акцеп-
повышение будет иметь высокую значимость
торной стороне этой фотосистемы. Такой ме-
для растения в том случае, если локальное дей-
ханизм хорошо согласуется с вызванным ВП
ствие стрессора, индуцирующее электрические
ростом потерь на акцепторной стороне фото-
сигналы, является предвестником действия
системы I [26], который практически не зави-
неблагоприятных факторов на другие части
сит от интенсивности темновой стадии фото-
растительного организма
[6, 7]. Механизмы
синтеза. По-видимому, этот механизм может
ЭС-индуцированного повышения устойчиво-
также способствовать вызванной ЭС актива-
сти фотосинтетического аппарата могут быть
ции циклического потока электронов вокруг
разделены на несколько групп [6, 7, 23, 31].
фотосистемы I [28], так как уменьшение нецик-
(i) Положительное влияние ЭС на устой-
лического потока, конкурирующего за суб-
чивость фотосистемы II при умеренной интен-
страт (восстановленный ферредоксин), должно
сивности стрессоров (на примере повышенной
приводить к возрастанию других потоков, ис-
и пониженной температуры) [34, 39] может
пользующих этот же субстрат [106].
быть связано с возрастанием энергозависи-
Вызванное ЭС закисление люмена хлоро-
мой компоненты нефотохимического тушения
пластов будет прежде всего индуцировать воз-
флуоресценции [30] и активацией циклическо-
растание энергозависимой компоненты не-
го потока электронов [28], которые являются
фотохимического тушения флуоресценции,
эффективными механизмами защиты фото-
так как электрические сигналы могут вызы-
синтетического аппарата при действии широ-
вать значительное возрастание этой компо-
кого спектра стрессоров [107-100, 113, 114].
ненты [30], для которой показана тесная связь
(ii) В случае действия высокой температуры
с pH люмена
[107-109]. Непосредственное
влияние ЭС на устойчивость фотосинтетиче-
снижение потока электронов через фото-
ского аппарата имеет более сложный харак-
синтетическую электрон-транспортную цепь
тер [35, 36]: снижение повреждения фотосисте-
также может быть результатом закисления
мы I сопровождается усилением повреждения
люмена хлоропластов, так как одной из ли-
фотосистемы II; при этом оба процесса явля-
митирующих стадий процесса является окис-
ются связанными [37]. Такой результат может
ление пластохинолов в комплексе цитохро-
быть объяснен гипотезой [115, 116] о том, что
мов b6 f [110, 111]; считается, что эта стадия
в условиях экстремального действия стрессо-
имеет высокую чувствительность к величи-
ров, повреждение фотосистемы II, способной к
не pH в люмене и замедляется при увеличе-
относительно быстрой репарации, может при-
нии концентрации протонов. Потенциально,
водить к сохранению фотосистемы I в резуль-
еще одной «мишенью» снижения pH в лю-
тате прекращения притока электронов от воды,
мене хлоропластов может быть компонента
уменьшения избыточного восстановления
нефотохимического тушения флуоресценции,
электрон-транспортной цепи и, как следствие,
обусловленная перемещением главного пиг-
снижения образования АФК. Важно отметить,
мент-белкового светособирующего комплекса
что сохранение фотосистемы I также важно для
(«state transition») [112]; так как показано [29],
последующей репарации фотосинтетического
7
БИОХИМИЯ том 88 вып. 10 2023
1810
СУХОВА и др.
Рис. 4. Схема возможных путей участия инактивации Н+-ATPазы плазматической мембраны в регуляции фотосин-
теза электрическими сигналами. Для СП приведен вариант, предполагающий участие инактивации inwardly-rectifying
K+-каналов в гиперполяризации
аппарата [23], так как в этом случае сохраняется
фотосинтетического аппарата после повреж-
возможность функционирования циклического
дающего действия стрессоров. Положительное
потока электронов и синтеза ATP.
влияние ATP на устойчивость фотосинтетиче-
(iii) Так как возрастание содержания ATP
ского аппарата растений [117] и данные о вы-
в листьях является важным следствием вы-
званном ЭС ускорении репарации фотосисте-
званных ЭС фотосинтетических ответов [16],
мы II [34] подтверждают эту гипотезу.
можно предположить, что это возрастание
Итоговым результатом вызванного ЭС
является механизмом ускорения репарации увеличения устойчивости фотосинтетического
БИОХИМИЯ том 88 вып. 10 2023
РОЛЬ Н+-АТРазы В ЭС И ОТВЕТАХ ФОТОСИНТЕЗА
1811
аппарата является общее повышение устойчи-
По-видимому, снижение абсолютной ак-
вости растения, которое проявляется в мень-
тивности Н+-ATPазы плазматической мем-
шем подавлении роста [36] и электрической
браны является основным механизмом инак-
активности [40] в условиях системного дей-
тивирующего влияния потенциалов действия,
ствия стрессоров.
вариабельных потенциалов и, вероятно, си-
стемных потенциалов на фотосинтез. Вызван-
ная электрическими сигналами инактивация
ЗАКЛЮЧЕНИЕ
фотосинтеза проявляется в снижении ассими-
ляции СО2, квантовых выходов фотосистем I
Проведенный обзор литературы показы-
и II и нециклического потока электронов,
вает, что формирование всех типов электри-
а также в возрастании нефотохимического ту-
ческих сигналов высших растений связано с
шения флуоресценции и циклического потока
изменениями активности Н+-ATPазы плазма-
электронов вокруг фотосистемы I. При этом
тической мембраны; при этом инактивация
индуцированное электрическими сигналами
Н+-ATPазы играет важную роль в формиро-
возрастание pH апопласта обеспечивает сни-
вании потенциалов действия и вариабельных
жение поступления СО2 в клетку, инактива-
потенциалов. В случае системного потенциала
цию темновой стадии фотосинтеза и, как след-
вопрос о конкретных изменениях активности
ствие, подавление процессов световой стадии
Н+-ATPазы имеет более сложный характер: с
фотосинтеза; в свою очередь, уменьшение pH
одной стороны, нельзя исключать возраста-
цитоплазмы и, как следствие, стромы и люме-
ние абсолютной активности транспортера,
на хлоропластов приводит к дополнительному
которое может быть обусловлено двухфазной
подавлению световой стадии.
зависимостью такой активности от концен-
Конечным результатом вызванных элек-
трации Са2+ в цитоплазме (активация при
трическими сигналами фотосинтетических
небольшом возрастании концентрации и ин-
ответов является, по-видимому, повышение
активация при умеренном и высоком воз-
устойчивости фотосинтетического аппарата к
растании). С другой стороны, ряд аргументов
действию стрессоров и облегчение его после-
(защелачивание апопласта во время генерации
дующей репарации. Такие изменения, в свою
системного потенциала, роль калиевых кана-
очередь, вносят вклад в общее повышение
лов в такой генерации) показывают большую
устойчивости растения к системному дей-
вероятность участия относительного возраста-
ствию неблагоприятных факторов и способ-
ния активности Н+-ATPазы в формировании
ствуют его выживанию в меняющихся услови-
системного потенциала; такое возрастание
ях окружающей среды.
обусловлено Са2+-зависимой инактивацией
inwardly-rectifying K+-каналов плазматиче-
Вклад авторов. Все авторы участвовали в
ской мембраны при небольшом повышении
формулировке концепции статьи, подготовке
концентрации ионов кальция. Абсолютная
материала и написании черновика обзора.
активность Н+-ATPазы может при этом сни-
В.С. Сухов осуществлял итоговое редактиро-
жаться вследствие возрастания при гипер-
вание текста обзора.
поляризации величины электрохимического
Финансирование. Работа выполнена при
градиента протонов, направленного внутрь
финансовой поддержке Российского научного
клетки. Возрастание электрохимического гра-
фонда (грант № 21-74-10088).
диента протонов может также снижать общую
Конфликт интересов. Авторы заявляют об
эффективность транспорта Н+ из клетки за
отсутствии конфликта интересов.
счет активации систем вторичного активно-
Соблюдение этических норм. Настоящая
го транспорта (например, протон-анионный
статья не содержит каких-либо исследований с
симпортер [118]) и усиления пассивных пото-
участием людей и животных в качестве объек-
ков Н+ в цитоплазму [119].
тов исследования.
СПИСОК ЛИТЕРАТУРЫ
1. Fromm, J., and Lautner, S. (2007) Electrical signals
2. Gallé, A., Lautner, S., Flexas, J., and Fromm, J. (2015)
and their physiological significance in plants, Plant
Environmental stimuli and physiological responses:
Cell Environ., 30, 249-257, doi: 10.1111/j.1365-3040.
the current view on electrical signaling, Environ. Exp.
2006.01614.x.
Bot., 114, 15-21, doi: 10.1016/j.envexpbot.2014.06.013.
БИОХИМИЯ том 88 вып. 10 2023
7*
1812
СУХОВА и др.
3.
Choi, W.G., Hilleary, R., Swanson, S.J., Kim,
photosynthetic limitation upon trigger hair irritation in
S.H., and Gilroy, S. (2016) Rapid, long-distance
the carnivorous plant Venus flytrap (Dionaea muscipula
electrical and calcium signaling in plants, Annu.
Ellis), J. Exp. Bot., 62, 1991-2000, doi: 10.1093/jxb/
Rev. Plant Biol., 67, 287-307, doi: 10.1146/annurev-
erq404.
arplant-043015-112130.
15.
Lautner, S., Stummer, M., Matyssek, R., Fromm, J.,
4.
Hedrich, R., Salvador-Recatalà, V., and Dreyer, I.
and Grams, T. E. E. (2014) Involvement of respiratory
(2016) Electrical wiring and long-distance plant
processes in the transient knockout of net CO2 uptake
communication, Trends Plant Sci.,
21,
376-387,
in Mimosa pudica upon heat stimulation, Plant Cell
doi: 10.1016/j.tplants.2016.01.016.
Environ., 37, 254-260, doi: 10.1111/pce.12150.
5.
Szechyńska-Hebda, M., Lewandowska, M., and
16.
Surova, L., Sherstneva, O., Vodeneev, V., Katiche-
Karpiński, S. (2017) Electrical signaling, photosynthe-
va, L., Semina, M., and Sukhov, V. (2016) Variation
sis and systemic acquired acclimation, Front. Physiol.,
potential-induced photosynthetic and respiratory
8, 684, doi: 10.3389/fphys.2017.00684.
changes increase ATP content in pea leaves, J. Plant
6.
Sukhov, V., Sukhova, E., and Vodeneev, V. (2019)
Physiol., 202, 57-64, doi: 10.1016/j.jplph.2016.05.024.
Long-distance electrical signals as a link between the
17.
Furch, A. C., van Bel, A. J., Fricker, M. D., Felle,
local action of stressors and the systemic physiological
H. H., Fuchs, M., and Hafke, J. B. (2009) Sieve
responses in higher plants, Progr. Biophys. Mol. Biol.,
element Ca2+ channels as relay stations between remote
146, 63-84, doi: 10.1016/j.pbiomolbio.2018.11.009.
stimuli and sieve tube occlusion in Vicia faba, Plant
7.
Sukhova, E., and Sukhov, V. (2021) Electrical sig-
Cell, 21, 2118-2132, doi: 10.1105/tpc.108.063107.
nals, plant tolerance to actions of stressors, and pro-
18.
Furch, A. C., Zimmermann, M. R., Will, T., Hafke,
grammed cell death: is interaction possible? Plants, 10,
J. B., and van Bel, A. J. (2010) Remote-controlled stop
1704, doi: 10.3390/plants10081704.
of phloem mass flow by biphasic occlusion in Cucurbita
8.
Wildon, D. C., Thain, J. F., Minchin, P. E. H.,
maxima, J. Exp. Bot., 61, 3697-3708, doi: 10.1093/
Gubb, I. R., Reilly, A. J., Skipper, Y. D., Doherty,
jxb/erq181.
H. M., O’Donnell, P. J., and Bowles, D. (1992)
19.
Van Bel, A. J., Furch, A. C., Will, T., Buxa, S. V.,
Electrical signalling and systemic proteinase inhibitor
Musetti, R., and Hafke, J. B. (2014) Spread the news:
Induction in the wounded plant, Nature, 360, 62-65,
systemic dissemination and local impact of Ca2+ sig-
doi: 10.1038/360062a0.
nals along the phloem pathway, J. Exp. Bot., 65,
9.
Mousavi, S. A., Chauvin, A., Pascaud, F., Kellen-
1761-1787, doi: 10.1093/jxb/ert425.
berger, S., and Farmer, E. E. (2013) Glutamate recep-
20.
Kaiser, H., and Grams, T. E.
(2006) Rapid
tor-like genes mediate leaf-to-leaf wound signalling,
hydropassive opening and subsequent active stomatal
Nature, 500, 422-426, doi: 10.1038/nature12478.
closure follow heat-induced electrical signals
10.
Hlavácková, V., Krchnák, P., Naus, J., Novák, O.,
in Mimosa pudica, J. Exp. Bot.,
57,
2087-2092,
Spundová, M., and Strnad, M. (2006) Electrical
doi: 10.1093/jxb/erj165.
and chemical signals involved in short-term systemic
21.
Yudina, L. M., Sherstneva, O. N., Mysyagin, S. A.,
photosynthetic responses of tobacco plants to local
Vodeneev, V. A., and Sukhov, V. S. (2019) Impact of
burning, Planta, 225, 235-244, doi: 10.1007/s00425-
local damage on transpiration of pea leaves at various
006-0325-x.
air humidity, Russ. J. Plant Physiol.,
66,
87-94,
11.
Hlavinka, J., Nožková-Hlaváčková, V., Floková, K.,
doi: 10.1134/S1021443719010163.
Novák, O., and Nauš, J.(2012) Jasmonic acid
22.
Stahlberg, R., and Cosgrove, D. J. (1996) Induction
accumulation and systemic pho-to-synthetic and
and ionic basis of slow wave potentials in seedlings of
electrical changes in locally burned wild type tomato,
Pisum sativum L., Planta, 200, 416-425, doi: 10.1007/
ABA-deficient sitiens mutants and sitiens pre-
BF00231397.
treated by ABA, Plant Physiol. Biochem., 54, 89-96,
23.
Sukhov, V. (2016) Electrical signals as mechanism of
doi: 10.1016/j.plaphy.2012.02.014.
photosynthesis regulation in plants, Photosynth. Res.,
12.
Farmer, E. E., Gao, Y. Q., Lenzoni, G., Wolfender,
130, 373-387, doi: 10.1007/s11120-016-0270-x.
J. L., and Wu, Q. (2020) Wound- and mechanostim-
24.
Gallé, A., Lautner, S., Flexas, J., Ribas-Carbo, M.,
ulated electrical signals control hormone responses,
Hanson, D., Roesgen, J., and Fromm, J. (2013) Pho-
New Phytol., 227, 1037-1050, doi: 10.1111/nph.16646.
tosynthetic responses of soybean (Glycine max L.)
13.
Filek, M., and Kościelniak, J. (1997) The effect
to heat-induced electrical signalling are predomi-
of wounding the roots by high temperature on
nantly governed by modifications of mesophyll con-
the respiration rate of the shoot and propagation
ductance for CO2, Plant Cell Environ., 36, 542-552,
of electric signal in horse bean seedlings (Vicia
doi: 10.1111/j.1365-3040.2012.02594.x.
faba L. minor), Plant Sci., 123, 39-46, doi: 10.1016/
25.
Grams, T. E., Lautner, S., Felle, H. H., Matyssek, R.,
S0168-9452(96)04567-0.
and Fromm, J. (2009) Heat-induced electrical signals
14.
Pavlovič, A., Slováková, L., Pandolfi, C., and
affect cytoplasmic and apoplastic pH as well as pho-
Mancuso, S. (2011) On the mechanism underlying
tosynthesis during propagation through the maize leaf,
БИОХИМИЯ том 88 вып. 10 2023
РОЛЬ Н+-АТРазы В ЭС И ОТВЕТАХ ФОТОСИНТЕЗА
1813
Plant Cell Environ., 32, 319-326, doi: 10.1111/j.1365-
37.
Surova, L., Sherstneva, O., Vodeneev, V., and
3040.2008.01922.x.
Sukhov, V. (2016) Variation potential propagation
26.
Sukhov, V., Orlova, L., Mysyagin, S., Sinitsina, J.,
decreases heat-related damage of pea photosystem I by
and Vodeneev, V. (2012) Analysis of the photosyn-
2 different pathways, Plant. Sign. Behav., 11, e1145334,
thetic response induced by variation potential in ge-
doi: 10.1080/15592324.2016.1145334.
ranium, Planta, 235, 703-712, doi: 10.1007/s00425-
38.
Sukhov, V., Gaspirovich, V., Mysyagin, S., and Vode-
011-1529-2.
neev, V. (2017) High-temperature tolerance of pho-
27.
Sukhov, V., Sherstneva, O., Surova, L., Katicheva, L.,
tosynthesis can be linked to local electrical responses
and Vodeneev, V. (2014) Proton cellular influx as a
in leaves of pea, Front. Physiol., 8, 763, doi: 10.3389/
probable mechanism of variation potential influence
fphys.2017.00763.
on photosynthesis in pea, Plant Cell Environ., 37,
39.
Grinberg, M. A., Gudkov, S. V., Balalaeva, I. V.,
2532-2541, doi: 10.1111/pce.12321.
Gromova, E., Sinitsyna, Y., Sukhov, V., and
28.
Sukhov, V., Surova, L., Sherstneva, O., Katicheva, L.,
Vodeneev, V. (2021) Effect of chronic β-radiation on
and Vodeneev, V. (2015) Variation potential influence
long-distance electrical signals in wheat and their role
on photosynthetic cyclic electron flow in pea, Front.
in adaptation to heat stress, Environ. Exp. Bot., 184,
Plant Sci., 5, 766, doi: 10.3389/fpls.2014.00766.
104378, doi: 10.1016/j.envexpbot.2021.104378.
29.
Sukhova, E., Mudrilov, M., Vodeneev, V., and
40.
Ретивин В. Г., Опритов В. А., Федулина С. Б.
Sukhov, V. (2018) Influence of the variation potential
(1997) Предадаптация тканей стебля CUCURBITA
on photosynthetic flows of light energy and electrons
PEPO к повреждающему действию низких темпе-
in pea, Photosynth. Res., 136, 215-228, doi: 10.1007/
ратур, индуцированная потенциалами действия,
s11120-017-0460-1.
Физиология растений, 44, 499-510.
30.
Sukhov, V., Sukhova, E., Gromova, E., Surova, L.,
41.
Williamson, R. E., and Ashley, C. C. (1982) Free Ca2+
Nerush, V., and Vodeneev, V. (2019) The electrical
and cytoplasmic streaming in the alga Chara, Nature,
signal-induced systemic photosynthetic response is
296, 647-650, doi: 10.1038/296647a0.
accompanied by changes in the photochemical re-
42.
Lunevsky, V. Z., Zherelova, O. M., Vostrikov, I. Y.,
flectance index in pea, Func. Plant Biol., 46, 328-338,
and Berestovsky, G. N. (1983) Excitation of Characeae
doi: 10.1071/FP18224.
cell membranes as a result of activation of calcium
31.
Sukhova, E., and Sukhov, V. (2023) Electrical sig-
and chloride channels, J. Membrain Biol., 72, 43-58,
nals in systemic adaptive response of higher plants:
doi: 10.1007/BF01870313.
Integration through separation, Bioelectricity,
5,
43.
Bulychev, A. A., Kamzolkina, N. A., Luengviriya, J.,
126-131, doi: 10.1089/bioe.2022.0042.
Rubin, A. B., and Müller, S. C. (2004) Effect of a
32.
Szechyńska-Hebda, M., Kruk, J., Górecka, M.,
single excitation stimulus on photosynthetic activity
Karpińska, B., and Karpiński, S. (2010) Evidence for
and light-dependent pH banding in Chara cells,
light wavelength-specific photoelectrophysiological
J. Membr. Biol., 202, 11-19, doi: 10.1007/s00232-
signaling and memory of excess light episodes in
004-0716-5.
Arabidopsis, Plant Cell, 22, 2201-2218, doi: 10.1105/
44.
Bulychev, A. A., and Krupenina, N. A. (2009) Tran-
tpc.109.069302.
sient removal of alkaline zones after excitation of
33.
Zandalinas, S. I., Fichman, Y., Devireddy, A. R.,
Chara cells is associated with inactivation of high con-
Sengupta, S., Azad, R. K., and Mittler, R. (2020)
ductance in the plasmalemma, Plant. Signal. Behav.,
Systemic signaling during abiotic stress combination in
4, 727-734, doi: 10.4161/psb.4.8.9306.
plants, Proc. Natl. Acad. Sci. USA, 117, 13810-13820,
45.
Felle, H. H., and Zimmermann, M. R. (2007) Sys-
doi: 10.1073/pnas.2005077117.
temic signalling in barley through action poten-
34.
Ретивин В. Г., Опритов В. А., Лобов С. А., Тара-
tials, Planta,
226,
203-214, doi:
10.1007/s00425-
канов С. А, Худяков В. А (1999). Модификация
006-0458-y.
устойчивости фотосинтезирующих клеток к охла-
46.
Vodeneev, V. A., Opritov, V. A., and Pyatygin, S. S.
ждению и прогреву после раздражения корней
(2006) Reversible changes of extracellular pH during
раствором КСl, Физиология растений, 46, 790-798.
action potential generation in a higher plant Cucurbita
35.
Sukhov, V., Surova, L., Sherstneva, O., and
pepo, Russ. J. Plant Physiol., 53, 481-487, doi: 10.1134/
Vodeneev, V. (2014) Influence of variation potential on
S102144370604008X.
resistance of the photosynthetic machinery to heating
47.
Sukhov, V., and Vodeneev, V. (2009) A mathematical
in pea, Physiol. Plant., 152, 773-783, doi: 10.1111/
model of action potential in cells of vascular plants,
ppl.12208.
J. Membr. Biol., 232, 59-67, doi: 10.1007/s00232-
36.
Sukhov, V., Surova, L., Sherstneva, O., Bushueva, A.,
009-9218-9.
and Vodeneev, V. (2015) Variation potential induces
48.
Vodeneev, V., Akinchits, E., and Sukhov, V. (2015)
decreased PSI damage and increased PSII damage
Variation potential in higher plants: mechanisms of
under high external temperatures in pea, Funct. Plant.
generation and propagation, Plant Sign. Behav., 10,
Biol., 42, 727-736, doi: 10.1071/FP15052.
e1057365, doi: 10.1080/15592324.2015.1057365.
БИОХИМИЯ том 88 вып. 10 2023
1814
СУХОВА и др.
49.
Sukhov, V., Akinchits, E., Katicheva, L., and
sion, Front. Plant Sci., 10, 1393, doi: 10.3389/fpls.
Vodeneev, V. (2013) Simulation of variation potential
2019.01393.
in higher plant cells, J. Membr. Biol., 246, 287-296,
62.
Vodeneev, V., Orlova, A., Morozova, E., Orlova, L.,
doi: 10.1007/s00232-013-9529-8.
Akinchits, E., Orlova, O., and Sukhov, V.
(2012)
50.
Ladeynova, M., Mudrilov, M., Berezina, E.,
The mechanism of propagation of variation potentials
Kior, D., Grinberg, M., Brilkina, A., Sukhov, V., and
in wheat leaves, J. Plant Physiol., 169, 949-954,
Vodeneev, V. (2020) Spatial and temporal dynamics of
doi: 10.1016/j.jplph.2012.02.013.
electrical and photosynthetic activity and the content
63.
Evans, M. J., Choi, W. G., Gilroy, S., and Morris,
of phytohormones induced by local stimulation of pea
R. J. (2016) A ROS-assisted calcium wave dependent
plants, Plants, 9, 1364, doi: 10.3390/plants9101364.
on the AtRBOHD NADPH oxidase and TPC1 cation
51.
Stahlberg, R., and Cosgrove, D. J. (1997) The prop-
channel propagates the systemic response to salt
agation of slow wave potentials in pea epicotyls, Plant
stress, Plant Physiol., 171, 1771-1784, doi: 10.1104/
Physiol., 113, 209-217, doi: 10.1104/pp.113.1.209.
pp.16.00215.
52.
Mancuso, S. (1999) Hydraulic and electrical transmis-
64.
Julien, J. L., Desbiez, M. O., de Jaeger, G.,
sion of wound-induced signals in Vitis vinifera, Aust. J.
Frachisse, J. M. (1991) Characteristics of the wave
Plant Physiol., 26, 55-61, doi: 10.1071/PP98098.
of depolarization induced by wounding in Bidens
53.
Sukhova, E., Akinchits, E., Gudkov, S. V.,
pilosa L., J. Exp. Bot., 42, 131-137, doi: 10.1093/
Pishchalnikov, R. Y., Vodeneev, V., and Sukhov,
jxb/42.1.131.
V. A (2021) Theoretical analysis of relations between
65.
Lautner, S., Grams, T. E. E., Matyssek, R. Fromm, J.
pressure changes along xylem vessels and propagation
(2005) Characteristics of electrical signals in poplar
of variation potential in higher plants, Plants,
and responses in photosynthesis, Plant Physiol.,
10, 372, doi: 10.3390/plants10020372.
138, 2200-2209, doi: 10.1104/pp.105.064196.
54.
Miller, G., Schlauch, K., Tam, R., Cortes, D., Torres,
66.
Zimmermann, M. R., Maischak, H., Mithöfer, A.,
M. A., Shulaev, V., Dangl, J. L., and Mittler, R. (2009)
Boland, W., and Felle, H. H. (2009) System potentials,
The plant NADPH oxidase RBOHD mediates rapid
a novel electrical long-distance apoplastic signal
systemic signaling in response to diverse stimuli,
in plants, induced by wounding, Plant Physiol.,
Sci. Signal., 2, ra45, doi: 10.1126/scisignal.2000448.
149, 1593-1600, doi: 10.1104/pp.108.133884.
55.
Suzuki, N., and Mittler, R. (2012) Reactive oxygen
67.
Zimmermann, M. R., Mithöfer, A., Will, T., Felle,
species-dependent wound responses in animals
H. H., and Furch, A. C. (2016) Herbivore-triggered
and plants, Free Radic. Biol. Med., 53, 2269-2276,
electrophysiological reactions: candidates for systemic
doi: 10.1016/j.freeradbiomed.2012.10.538.
signals in higher plants and the challenge of their iden-
56.
Choi, W. G., Miller, G., Wallace, I., Harper, J.,
tification, Plant Physiol., 170, 2407-2419, doi: 10.1104/
Mittler, R., and Gilroy, S. (2017) Orchestrating rapid
pp.15.01736.
long-distance signaling in plants with Ca2+, ROS and
68.
Vuralhan-Eckert, J., Lautner, S., and Fromm, J.
electrical signals, Plant J., 90, 698-707, doi: 10.1111/
(2018) Effect of simultaneously induced environmental
tpj.13492.
stimuli on electrical signalling and gas exchange
57.
Peña-Cortés, H., Fisahn, J., and Willmitzer, L.
in maize plants, J. Plant Physiol.,
223,
32-36,
(1995) Signals involved in wound-induced proteinase
doi: 10.1016/j.jplph.2018.02.003.
inhibitor II gene expression in tomato and potato
69.
Yudina, L., Gromova, E., Grinberg, M., Popova, A.,
plants, Proc. Natl. Acad. Sci. USA, 92, 4106-4113,
Sukhova, E., and Sukhov, V. (2022) Influence of
doi: 10.1073/pnas.92.10.4106.
burning-induced electrical signals on photosynthesis
58.
Toyota, M., Spencer, D., Sawai-Toyota, S., Jiaqi, W.,
in pea can be modified by soil water shortage, Plants,
Zhang, T., Koo, A. J., Howe, G. A., and Gilroy, S.
11, 534, doi: 10.3390/plants11040534.
(2018) Glutamate triggers long-distance, calcium-
70.
Yudina, L., Sukhova, E., Popova, A., Zolin, Yu.,
based plant defense signaling, Science, 361, 1112-1115,
Abasheva, K., Grebneva, K., and Sukhov V. (2023)
doi: 10.1126/science.aat7744.
Local action of moderate heating and illumination
59.
Malone, M. (1994) Wound-induced hydraulic signals
induces propagation of hyperpolarization electrical
and stimulus transmission in Mimosa pudica L.,
signals in wheat plants, Front. Sustain. Food Syst.,
New Phytol., 128, 49-56, doi: 10.1111/j.1469-8137.
6, 1062449, doi: 10.3389/fsufs.2022.1062449.
1994.tb03985.x.
71.
Yudina, L., Sukhova, E., Popova, A., Zolin, Y.,
60.
Evans, M. J., and Morris, R. J. (2017) Chemical agents
Abasheva, K., Grebneva, K., and Sukhov, V. (2023)
transported by xylem mass flow propagate variation
Hyperpolarization electrical signals induced by local
potentials, Plant J.,
91,
1029-1037, doi:
10.1111/
action of moderate heating influence photosynthetic
tpj.13624.
light reactions in wheat plants, Front. Plant Sci.,
61.
Blyth, M. G., and Morris, R. J. (2019) Shear-en-
14, 1153731, doi: 10.3389/fpls.2023.1153731.
hanced dispersion of a wound substance as a can-
72.
Lew, R. R. (1989) Calcium activates an electro-
didate mechanism for variation potential transmis-
genic proton pump in neurospora plasma mem-
БИОХИМИЯ том 88 вып. 10 2023
РОЛЬ Н+-АТРазы В ЭС И ОТВЕТАХ ФОТОСИНТЕЗА
1815
brane, Plant Physiol.,
91,
213-216, doi:
10.1104/
84.
Herde, O., Peña-Cortés, H., Fuss, H., Willmitzer,
pp.91.1.213.
L., and Fisahn, J. (1999) Effects of mechanical
73.
Grinberg, M., Mudrilov, M., Kozlova, E., Sukhov, V.,
wounding, current application and heat treatment on
Sarafanov, F., Evtushenko, A., Ilin, N., Vodeneev, V.,
chlorophyll fluorescence and pigment composition
Price, C., and Mareev, E. (2022) Effect of extremely
in tomato plants, Physiol. Plant.,
105,
179-184,
low-frequency magnetic fields on light-induced
doi: 10.1034/j.1399-3054.1999.105126.x.
electric reactions in wheat, Plant Signal. Behav., 17,
85.
Sherstneva, O. N., Vodeneev, V. A., Katicheva,
e2021664, doi: 10.1080/15592324.2021.2021664.
L. A., Surova, L. M., and Sukhov, V. S.
(2015)
74.
Grabov, A., and Blatt, M. R.
(1999) A steep
Participation of intracellular and extracellular pH
dependence of inward-rectifying potassium channels
changes in photosynthetic response development
on cytosolic free calcium concentration increase
induced by variation potential in pumpkin seedlings,
evoked by hyperpolarization in guard cells, Plant
Biochemistry (Moscow), 80, 776-784, doi: 10.1134/
Physiol., 119, 277-288, doi: 10.1104/pp.119.1.277.
S0006297915060139.
75.
Gao, Y. Q., Wu, W. H., and Wang, Y.
(2019)
86.
Kinoshita, T., Nishimura, M., and Shimazaki, Ki.
Electrophysiological identification and activity
(1995) Cytosolic concentration of Ca2+ regulates
analyses of plasma membrane K+ channels in
the plasma membrane H+-ATPase in guard cells of
maize guard cells, Plant Cell Physiol., 60, 765-777,
fava bean, Plant Cell, 7, 1333-1342, doi: 10.1105/
doi: 10.1093/pcp/pcy242.
tpc.7.8.1333.
76.
Sukhova, E., Akinchits, E., and Sukhov, V. (2017)
87.
Sukhov, V. S., Gaspirovich, V. V., Gromova, E. N.,
Mathematical models of electrical activity in plants,
Ladeynova, M. M., Sinitsyna, Yu. V., Berezina,
J. Membr. Biol., 250, 407-423, doi: 10.1007/s00232-
E. V., Akinchits, E. K., and Vodeneev, V. A. (2017)
017-9969-7.
Decrease of mesophyll conductance to CO2 is a
77.
Sukhova, E., Ratnitsyna, D., and Sukhov, V. (2021)
possible mechanism of abscisic acid influence on
Stochastic spatial heterogeneity in activities of H+-
photosynthesis in seedlings of pea and wheat, Biochem.
ATP-ases in electrically connected plant cells de-
Moscow Suppl. Ser. A, 11, 237-247, doi: 10.1134/
creases threshold for cooling-induced electrical re-
S1990747817030096.
sponses, Int. J. Mol. Sci., 22, 8254, doi: 10.3390/
88.
Yudina, L., Sukhova, E., Sherstneva, O., Grinberg, M.,
ijms22158254.
Ladeynova, M., Vodeneev, V., and Sukhov, V. (2020)
78.
Falhof, J., Pedersen, J. T., Fuglsang, A. T., and
Exogenous abscisic acid can influence photosyn-
Palmgren, M. (2016) Plasma membrane H+-ATPase
thetic processes in peas through a decrease in activi-
regulation in the center of plant physiology, Mol.
ty of H+-ATP-ase in the plasma membrane, Biology,
Plant., 9, 323-337, doi: 10.1016/j.molp.2015.11.002.
9, 324, doi: 10.3390/biology9100324.
79.
Fuglsang, A. T., and Palmgren, M. (2021) Proton and
89.
Yudina, L., Sherstneva, O., Sukhova, E., Grinberg, M.,
calcium pumping P-type ATPases and their regulation
Mysyagin, S., Vodeneev, V., and Sukhov, V. (2020) In-
of plant responses to the environment, Plant Physiol.,
activation of H+-ATPase participates in the influence
187, 1856-1875, doi: 10.1093/plphys/kiab330.
of variation potential on photosynthesis and respiration
80.
Katicheva, L., Sukhov, V., Akinchits, E., and
in peas, Plants, 9, 1585, doi: 10.3390/plants9111585.
Vodeneev, V. (2014) Ionic nature of burn-induced
90.
Sukhov, V., Surova, L., Morozova, E., Sherstneva, O.,
variation potential in wheat leaves. Plant Cell Physiol.,
and Vodeneev, V. (2016) Changes in H+-ATP synthase
55, 1511-1519, doi: 10.1093/pcp/pcu082.
activity, proton electrochemical gradient, and pH
81.
Krupenina, N. A., and Bulychev, A. A. (2007) Action
in pea chloroplast can be connected with variation
potential in a plant cell lowers the light requirement
potential, Front. Plant Sci., 7, 1092, doi: 10.3389/
for non-photochemical energy-dependent quenching
fpls.2016.01092.
of chlorophyll fluorescence, Biochim. Biophys. Acta,
91.
Sherstneva, O. N., Surova, L. M., Vodeneev, V. A.,
1767, 781-788, doi: 10.1016/j.bbabio.2007.01.004.
Plotnikova, Yu. I., Bushueva, A. V., and Sukhov, V. S.
82.
Krausko, M., Perutka, Z., Šebela, M., Šamajová, O.,
(2016) The role of the intra- and extracellular protons
Šamaj, J., Novák, O., and Pavlovič, A.
(2017)
in the photosynthetic response induced by the variation
The role of electrical and jasmonate signalling in the
potential in pea seedlings, Biochem. Moscow Suppl.
recognition of captured prey in the carnivorous sundew
Ser. A, 10, 60-67, doi: 10.1134/S1990747815050116.
plant Drosera capensis, New Phytol., 213, 1818-1835,
92.
Sherstneva, O. N., Vodeneev, V. A., Surova, L. M.,
doi: 10.1111/nph.14352.
Novikova, E. M., and Sukhov, V. S. (2016) Application
83.
Białasek, M., Górecka, M., Mittler, R., and Kar-
of a mathematical model of variation potential for
piński, S. (2017) Evidence for the Involvement of
analysis of its influence on photosynthesis in higher
electrical, calcium and ROS signaling in the systemic
plants, Biochem. Moscow Suppl. Ser. A, 10, 269-277,
regulation of non-photochemical quenching and
doi: 10.1134/S1990747816030089.
photosynthesis, Plant Cell Physiol.,
58,
207-215,
93.
Bulychev, A. A., Alova, A. V., and Rubin, A. B. (2013)
doi: 10.1093/pcp/pcw232.
Fluorescence transients in chloroplasts of Chara
БИОХИМИЯ том 88 вып. 10 2023
1816
СУХОВА и др.
corallina cells during transmission of photoinduced
105. Benz, J. P., Stengel, A., Lintala, M., Lee, Y. H.,
signal with the streaming cytoplasm, Russ. J. Plant
Weber, A., Philippar, K., Gügel, I. L., Kaieda, S.,
Physiol., 60, 33-40, doi: 10.1134/S1021443712060039.
Ikegami, T., Mulo, P., Soll, J., and Bölter, B.
94. Tholen, D., and Zhu, X.-G. (2011) The mechanistic
(2010) Arabidopsis Tic62 and ferredoxin-NADP(H)
basis of internal conductance: a theoretical analysis of
oxidoreductase form light-regulated complexes that
mesophyll cell photosynthesis and CO2 diffusion, Plant
are integrated into the chloroplast redox poise, Plant
Physiol., 156, 90-105, doi: 10.1104/pp.111.172346.
Cell, 21, 3965-3983, doi: 10.1105/tpc.109.069815.
95. Sukhova, E. M., and Sukhov, V. S. (2018) Depen-
106. Joliot, P., and Alric, J. (2013) Inhibition of CO2
dence of the CO2 uptake in a plant cell on the plasma
fixation by iodoacetamide stimulates cyclic electron
membrane H+-ATPase activity: theoretical analysis,
flow and non-photochemical quenching upon far-
Biochem. Moscow Suppl. Ser. A,
12,
146-159,
red illumination, Photosynth. Res.,
115,
55-63,
doi: 10.1134/S1990747818020149.
doi: 10.1007/s11120-013-9826-1.
96. Sukhova, E., Ratnitsyna, D., Gromova, E., and
107. Müller, P., Li, X. P., and Niyogi, K. K. (2001) Non-
Sukhov, V. (2022) Development of two-dimensional
photochemical quenching. A response to excess light
model of photosynthesis in plant leaves and analysis
energy, Plant Physiol., 125, 1558-1566, doi: 10.1104/
of induction of spatial heterogeneity of CO2 assim-
pp.125.4.1558.
ilation rate under action of excess light and drought,
108. Jajoo, A., Mekala, N. R., Tongra, T., Tiwari, A.,
Plants, 11, 3285, doi: 10.3390/plants11233285.
Grieco, M., Tikkanen, M., and Aro, E. M. (2014) Low
97. Sukhova, E., Ratnitsyna, D., and Sukhov, V. (2022)
pH-induced regulation of excitation energy between
Simulated analysis of influence of changes in
the two photosystems, FEBS Lett., 588, 970-974,
H+-ATPase activity and membrane CO2 conductance
doi: 10.1016/j.febslet.2014.01.056.
on parameters of photosynthetic assimilation in leaves,
109. Ruban, A. V. (2016) Nonphotochemical chlorophyll
Plants, 11, 3435, doi: 10.3390/plants11243435.
fluorescence quenching: mechanism and effectiveness
98. Uehlein. N., Otto, B., Hanson, D. T., Fischer, M.,
in protecting plants from photodamage, Plant Physiol.,
McDowell, N., and Kaldenhoff, R. (2008) Function
170, 1903-1916, doi: 10.1104/pp.15.01935.
of Nicotiana tabacum aquaporins as chloroplast gas
110. Tikhonov, A. N. (2013) pH-dependent regulation of
pores challenges the concept of membrane CO2
electron transport and ATP synthesis in chloroplasts,
permeability, Plant Cell, 20, 648-657, doi: 10.1105/
Photosynth. Res., 116, 511-534, doi: 10.1007/s11120-
tpc.107.054023.
013-9845-y.
99. Luu, D.-Y., and Maurel, C. (2005) Aquaporins in a
111. Tikhonov, A. N. (2014) The cytochrome b6 f complex
challenging environment: molecular gears for adjusting
at the crossroad of photosynthetic electron transport
plant water status, Plant Cell Environ., 28, 85-96,
pathways, Plant Physiol. Biochem.,
81,
163-183,
doi: 10.1111/j.1365-3040.2004.01295.x.
doi: 10.1016/j.plaphy.2013.12.011.
100. Chaumont, F., and Tyerman, S. D. (2014) Aquaporins:
112. Rochaix, J.-D., Lemeille, S., Shapiguzov, A.,
highly regulated channels controlling plant water
Samol, I., Fucile, G., Willig, A., and Goldschmidt-
relations, Plant Physiol., 164, 1600-1618, doi: 10.1104/
Clermont, M. (2012) Protein kinases and phosphatases
pp.113.233791.
involved in the acclimation of the photosynthetic appa-
101. Kapilan, R., Vaziri, M., and Zwiazek, J. J. (2018)
ratus to a changing light environment, Philos. Trans. R.
Regulation of aquaporins in plants under stress, Biol.
Soc. B, 367, 3466-3474, doi: 10.1098/rstb.2012.0064.
Res., 51, 4, doi: 10.1186/s40659-018-0152-0.
113. Joliot, P., and Joliot, A. (2006) Cyclic electron flow
102. Wang, C., Hu, H., Qin, X., Zeise, B., Xu, D.,
in C3 plants, Biochim. Biophys. Acta, 1757, 362-368,
Rappel, W. J., Boron, W. F, and Schroeder, J. I.
doi: 10.1016/j.bbabio.2006.02.018.
(2016) Reconstitution of CO2 regulation of SLAC1
114. Joliot, P., and Johnson, G. N. (2011) Regulation of
anion channel and function of CO2-permeable PIP2;1
cyclic and linear electron flow in higher plants, Proc.
aquaporin as CARBONIC ANHYDRASE4 interactor,
Natl. Acad. Sci. USA, 108, 13317-13322, doi: 10.1073/
Plant Cell, 28, 568-582, doi: 10.1105/tpc.15.00637.
pnas.1110189108.
103. Flexas, J., Ribas-Carbó, M., Diaz-Espejo, A.,
115. Tikkanen, M., and Aro, E. M. (2014) Integrative
Galmés, J., and Medrano, H. (2008) Mesophyll
regulatory network of plant thylakoid energy
conductance to CO2: current knowledge and
transduction, Trends Plant Sci.,
19,
10-17,
future prospects, Plant Cell Environ., 31, 602-621,
doi: 10.1016/j.tplants.2013.09.003.
doi: 10.1111/j.1365-3040.2007.01757.x.
116. ikkanen, M., Mekala, N. R., and Aro, E. M.
104. Alte, F., Stengel, A., Benz, J. P., Petersen, E.,
(2014) Photosystem II photoinhibition-repair cycle
Soll, J., Groll, M., and Bölter, B. (2010) Ferredox-
protects Photosystem I from irreversible damage,
in:NADPH oxidoreductase is recruited to thylakoids
Biochim. Biophys. Acta, 1837, 210-215, doi: 10.1016/
by binding to a polyproline type II helix in a pH-
j.bbabio.2013.10.001.
dependent manner, Proc. Natl. Acad. Sci. USA, 107,
117. Allakhverdiev, S. I., Nishiyama, Y., Takahashi, S.,
19260-19265, doi: 10.1073/pnas.1009124107.
Miyairi, S., Suzuki, I., and Murata, N.
(2005)
БИОХИМИЯ том 88 вып. 10 2023
РОЛЬ Н+-АТРазы В ЭС И ОТВЕТАХ ФОТОСИНТЕЗА
1817
Systematic analysis of the relation of electron transport
119. Tyerman, S. D., Beilby, M., Whittington, J.,
and ATP synthesis to the photodamage and repair
Juswono, U., Neyman, L., and Shabala, S. (2001)
of photosystem II in Synechocystis, Plant Physiol.,
Oscillations in proton transport revealed from simul-
137, 263-273, doi: 10.1104/pp.104.054478.
taneous measurements of net current and net proton
118. Gradmann, D. (2001) Impact of apoplast volume
fluxes from isolated root protopasts: MIFE meets
on ionic relations in plant cells, J. Membr. Biol.,
patch-clamp, Aust. J. Plant Physiol., 28, 591-604,
184, 61-69, doi: 10.1007/s00232-001-0074-5.
doi: 10.1071/PP01030.
CHANGES IN ACTIVITY OF H+-ATPase OF PLASMA MEMBRANE
AS A LINK BETWEEN FORMING ELECTRICAL SIGNALS
AND INDUCTION OF PHOTOSYNTHETIC RESPONSES
IN HIGHER PLANTS
Review
E. M. Sukhova, L. M. Yudina, and V. S. Sukhov*
N. I. Lobachevsky State University of Nizhny Novgorod,
603022 Nizhny Novgorod, Russia; e-mail: vssuh@mail.ru
Action of numerous adverse environmental factors on higher plants is spatially-heterogenous; it means
that induction of a systemic adaptive response requires generation and propagation stress signals. Electrical
signals (ESs) induced by local action of stressors include action potential, variation potential, and system
potential and participate in forming fast physiological changes at the level of the whole plant, including pho-
tosynthetic responses. Generation of these ESs is accompanied by changes in activity of H+-ATPase which
is the main system of electrogenic proton transport across the plasma membrane. Literature data show that
the changes in H+-ATPase activity and related changes in intra- and extracellular pH play a key role in
ESs-induced photosynthetic inactivation in non-irritated parts of plants. This inactivation is caused by both
suppression of CO2 flux into mesophyll cells in leaves, which can be induced by the apoplast alkalization
and, probably, cytoplasm acidification, and direct influence of acidification of stroma and lumen of chlo-
roplasts on photosynthetic light and, probably, dark reactions. Result of the ESs-induced photosynthetic
inactivation is increasing tolerance of photosynthetic machinery to action of adverse factors and probability
of survive of plants.
Keywords: higher plants, electrical signals, action potential, variation potential, system potential, H+-ATPase,
intracellular pH, extracellular pH, photosynthesis
БИОХИМИЯ том 88 вып. 10 2023