БИОХИМИЯ, 2023, том 88, вып. 10, с. 1908 - 1925
УДК 577.344;543.421
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ВОЗБУЖДЕННЫХ СОСТОЯНИЙ
ТЕТРАМЕРА ХЛОРОФИЛЛА В ВОДОРАСТВОРИМОМ
ХЛОРОФИЛЛ-СВЯЗЫВАЮЩЕМ БЕЛКЕ BoWSCP
© 2023 Д.А. Черепанов1,2*, К.В. Неверов3,4, Ю.Н. Обухов3, Ю.В. Малеева4,
Ф.Е. Гостев1, И.В. Шелаев1,2, А.В. Айбуш1, М.С. Крицкий3, В.А. Надточенко1,5*
1 ФИЦ химической физики им. Н.Н. Семенова РАН,
119991 Москва, Россия; электронная почта: tscherepanov@gmail.com
2 НИИ физико-химической биологии имени А.Н. Белозерского,
Московский государственный университет имени М.В. Ломоносова, 119992 Москва, Россия
3 ФИЦ «Фундаментальные основы биотехнологии» РАН, Институт биохимии им. А.Н. Баха РАН,
119017 Москва, Россия
4 Московский государственный университет имени М.В. Ломоносова, биологический факультет,
119992 Москва, Россия
5 Московский государственный университет имени М.В. Ломоносова, химический факультет,
119992 Москва, Россия; электронная почта: nadtochenko@gmail.com
Поступила в редакцию 22.06.2023
После доработки 22.09.2023
Принята к публикации 22.09.2023
Методом широкополосной фемтосекундной лазерной спектроскопии «возбуждение-зондирова-
ние» измерена абсорбционная динамика хлорофилла a в симметричном тетрамерном комплексе
водорастворимого хлорофилл-связывающего белка BoWSCP в диапазоне от 400 до 750 нм с вре-
менным разрешением 20 фс-200 пс. При возбуждении BoWSCP в области полосы Соре на длине
волны 430 нм наблюдалась безызлучательная внутримолекулярная конверсия S3→S1 с характерным
временем 83 ± 9 фс. При возбуждении комплекса в области полосы Qy на длине волны 670 нм на-
блюдался релаксационный переход между двумя экситонными состояниями димера хлорофилла
со временем 105 ± 10 фс. Получены спектры поглощения возбужденных синглетных состояний
S1 и S3 хлорофилла a. Продемонстрировано, что делокализация возбужденного состояния между
экситонно-сопряженными молекулами хлорофилла в тетрамере BoWSCP изменяется во времени
и зависит от энергии возбуждения. При возбуждении BoWSCP в области полосы Соре наблюда-
ется сверхбыстрая фотохимическая реакция, обусловленная, по всей видимости, восстановлением
триптофана в ближайшем окружении хлорофилла.
КЛЮЧЕВЫЕ СЛОВА: хлорофилл a, белки семейства WSCP, фемтосекундная лазерная спектроскопия,
спектр возбужденного состояния, экситонная динамика, внутримолекулярная конверсия.
DOI: 10.31857/S0320972523100135, EDN: OTSLKL
ВВЕДЕНИЕ
синтетических комплексов, регистрируемой ме-
тодами фемтосекундной лазерной спектроско-
Анализ фотохимических реакций переноса
пии, представляет значительные методические
энергии и разделения зарядов в фотосинтети-
трудности [1, 2]. Хлорофилл-связывающие бел-
ческих пигмент-белковых комплексах затруд-
ки семейства WSCP (Water-Soluble Chlorophyll
нен их сложной молекулярной организацией
Binding Proteins) из высших растений пред-
и наличием большого количества экситонно-
ставляют собой идеальную модельную систе-
сопряженных молекул хлорофилла (Хл). Ин-
му для изучения механизма фотохимических
терпретация абсорбционной динамики фото- реакций [3-6]. Эти белки водорастворимы,
Принятые сокращения: Хл - хлорофилл; BL - суммарное выцветание в области стимулированного излучения
(GSB + SE); BoWSCP - водорастворимый хлорофилл-связывающий белок семейства WSCP из Brassica oleracea var.
botrytis; GSB - выцветание основного состояния; ESA - поглощение возбужденного состояния; SE - стимулирован-
ное излучение возбужденного состояния; TA - переходное поглощение.
* Адресат для корреспонденции.
1908
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ТЕТРАМЕРА ХЛОРОФИЛЛА
1909
фото- и термостабильны, могут быть эффек-
в низшее возбужденное состояние, а также
тивно модифицированы и продуцированы ме-
электронных переходов между экситонными
тодами генной и белковой инженерии. В клет-
уровнями полосы Qy тетрамера Хл a при воз-
ке растений эти белки локализованы вне
буждении BoWSCP в области полосы Соре и
тилакоидных мембран хлоропластов и не уча-
полосы Qy. Рассматриваются спектральные ха-
ствуют в фотосинтетическом процессе. Пред-
рактеристики, позволяющие определять сте-
полагается, что их физиологические функции
пень делокализации возбужденного состоя-
связаны с активностью антистрессовых систем
ния (экситона) между несколькими молеку-
клетки [4, 7, 8]. По сравнению с фотосинте-
лами Хл в тетрамерном комплексе, а также
тическими пигмент-белковыми комплексами
детектировать реакции переноса электрона с
структура белков WSCP относительно проста.
участием возбужденных молекул Хл a в данном
Холоформы WSCP представляют собой водо-
комплексе.
растворимые гомотетрамеры с молекулярной
массой 69-80 кДа, при этом каждый комплекс
содержит до четырех нековалентно связанных
МАТЕРИАЛЫ И МЕТОДЫ
молекул Хл. В зависимости от степени гомоло-
гии первичной структуры апобелка семейство
Препараты апобелка BoWSCP получали,
WSCP подразделяется на классы I и II, причем
экспрессируя гены этого белка, встроенные
в последнем выделяют подклассы IIа и IIb по
в плазмиды, которыми были трансформиро-
разной аффинности апобелков к Хл a и b [6, 9].
ваны клетки штамма-продуцента Escherichia
Было показано, что при фотовозбуждении Хл
coli BL21(DE3), согласно используемым ра-
в WSCP происходит миграция энергии как
нее протоколам [5, 6, 22]. Благодаря наличию
внутри димера, так и между димерными пара-
на N-концах апобелков шести гистидиновых
ми [10, 11]. Это свойство делает WSCP класса II
остатков (6xHis) очистку их препаратов про-
перспективной моделью для изучения меха-
водили с использованием аффинной хромато-
низмов пигмент-пигментных и пигмент-бел-
графии на колонке с Ni-агарозой с последую-
ковых взаимодействий, с помощью которой воз-
щим диализом препарата. Чистоту препаратов
можно исследовать механизмы переноса энер-
проверяли методом денатурирующего элек-
гии возбуждения между пигментами [12-14] и
трофореза в полиакриламидном геле [5, 6].
взаимодействие возбужденных пигментов с
Самосборку холоформ BoWSCP осуществляли
белковым окружением [15-17].
in vitro путем совместной инкубации апобелков
В данной работе методом широкополос-
и изолированных тилакоидных мембран шпи-
ной фемтосекундной лазерной спектроскопии
ната [4-6, 8, 22]. Учитывая высокую термоста-
«возбуждение-зондирование» изучались фото-
бильность тетрамеров белков WSCP, получен-
химические процессы в симметричном тетра-
ные препараты холоформ прогревали (95 °C,
мерном комплексе белка BoWSCP подклас-
10 мин) с целью удаления посторонних белков,
са IIа из Brassica oleracea var. botrytis, который
после чего проводили окончательную очист-
связывает преимущественно Хл a, то есть их
ку BoWSCP на колонке с Ni-агарозой [1, 23].
пигментный состав можно считать практиче-
Очищенный препарат диализовали против
ски гомогенным [5]. Тетрамерный комплекс Хл
10 мМ Tris-HCl (pH 8,0). Качество препара-
в белке BoWSCP организован в виде двух ди-
тов тетрамеров BoWSCP проверяли методами
меров, причем угол между плоскостями пор-
нативного электрофореза в полиакриламид-
фириновых ядер пигментов в димере состав-
ном геле, абсорбционной, флуоресцентной
ляет около 30° [18]. В совокупности два димера
спектроскопии и спектроскопии кругового
в составе макромолекулы холобелка форми-
дихроизма. Электрофорез белков в денатури-
руют нековалентную тетрамерную структуру, в
рующих и нативных условиях проводили в ка-
которой димерные пары Хл соединены друг с
мере Mini PROTEAN Tetra System («Bio-Rad»,
другом фитольными «хвостами» [19]. Энергия
США). Содержание белка в образцах изме-
экситонного взаимодействия мономеров в ди-
ряли методом Брэдфорда [24]. Спектры по-
мере составляет примерно 100 см-1, взаимодей-
глощения образцов регистрировали на спек-
ствие между димерами в десять раз слабее [13,
трофотометрах Cary-50
(«Agilent», США) и
20]. Недавно было показано, что Хл в составе
Shimadzu-1601PC («Shimadzu», Япония). Спек-
данной структуры обладает фотокаталитиче-
тры кругового дихроизма (КД) регистрирова-
ской активностью в редокс-процессах [21].
ли на спектрометре кругового дихроизма
Настоящая работа посвящена количествен-
Chirascan («Applied Photophysics», США). Пере-
ному анализу процессов безызлучательной
счет амплитуды сигналов (Δε) в единицы эл-
внутримолекулярной конверсии из третьего
липтичности (θ) для спектров кругового ди-
БИОХИМИЯ том 88 вып. 10 2023
1910
ЧЕРЕПАНОВ и др.
хроизма проводили, согласно используемому
и ее производными по времени G′ и G′′, где ши-
ранее протоколу [23].
рина возбуждающего импульса d находилась
Препараты для фемтосекундной спек-
моделированием абсорбционных изменений
троскопии концентрировали путем центри-
чистого растворителя [29]. Сигнал, обуслов-
фугирования в центрифужных концентра-
ленный изменением электронных состояний
торах Amicon Ultra-15 («Merck», Германия) с
пигментов, аппроксимировался сверткой ба-
фильтрами на 30 или 50 кДа. Концентриро-
зовой гауссовой функции G(t) и ступенчатой
вание проводили серией центрифугирова-
функции Хевисайда H(t):
ний препаратов белка BoWSCP в 10 мМ Tris-
HCl (pH 8,0) с добавлением к ним NaCl
χ(t) = G(t)*H(t) = ½Erfc[-t/√2πd2],
(2)
до 150 мМ или 10% (v/v) глицерина для пред-
отвращения агрегации белка в высококонцен-
а также свертками G(t) и экспоненциальных
трированном растворе. Полученный концен-
функций:
трат разбавляли буфером такого же состава
ε(t, τk) = G(t)*exp(-t / τk) =
с NaCl или глицерином до получения 1 мл рас-
(3)
твора с ОП673 = 5.
= ½exp[½(d/τk)2 - t/τk]Erfc[(d/τk - t/d)]/√2,
Сверхбыстрые абсорбционные изменения
ΔA(λi, tm) регистрировались методом широко-
амплитуды Ak(λi) которых находились линей-
полосной фемтосекундной лазерной спек-
ной регрессией, а характерные времена τk
троскопии «возбуждение-зондирование» в оп-
определялись методом нелинейной миними-
тическом диапазоне 400 ≤ λ ≤ 780 нм на вре-
зации.
менных задержках t от 20 фс до 200 пс. Экс-
Математический анализ абсорбционной ди-
периментальная установка и методика изме-
намики включал разложение спектрально-вре-
рений были описаны ранее [25]. Возбуждение
менных матриц ΔA(λl, tm) на линейную ком-
BoWSCP осуществлялось фемтосекундными
бинацию n дискретных экспоненциальных
лазерными импульсами с максимумами на дли-
функций:
нах волн 430 нм (длительность - 40 фс, энер-
n
гия - 20 нДж) и 670 нм (длительность - 16 фс,
(4)
Q[τ](λ, t) = ΣDk(λ)⋅exp(-t/τk)+Dn+1(λ)
k = 1
энергия - 15 нДж). Зондирование обеспечи-
валось широкополосными импульсами супер-
в предположении, что характерные времена
континуума при углах поляризации 0; 54,7
τ = {τk} не зависят от длины волны наблюдения
и 90 градусов относительно поляризации воз-
λ и, следовательно, могут рассматриваться как
буждающего импульса. Разностные спектры
«глобальные» параметры, определяемые нели-
поглощения в диапазоне 400-780 нм снимали
нейной минимизацией нормированной суммы
с помощью ПЗС-камеры SPEC-10 («Roper Sci-
квадратов невязок:
entific», США), соединенной с полихромато-
L M
ром SP-300 («Acton», США). Эксперименты
R[τ] = (L⋅M-n)-1ΣΣ[ΔA(λl, tm)-
l = 1 m = 1
проводились при температуре 6 °С в проточной
(5)
оптической ячейке (толщина - 0,5 мм; диаметр
-Q[τ](λl, tm)]2.
оптических окон - 0,2 мм; оптическая плот-
ность образца - 10 см-1). Скорость циркуляции
Зависящие от длины волны предэкспонен-
в проточной ячейке (9 мл/мин) была доста-
циальные амплитуды Dk(λ), рассчитываемые
точно высокой, чтобы избежать многократного
стандартным методом линейной регрессии для
возбуждения одного и того же объема пробы.
каждой экспоненциальной функции k, опреде-
Спектры корректировались дисперсией груп-
ляют спектры, связанные с распадом (Decay-
повой задержки, как описано ранее [26, 27].
Associated Spectra, DAS), одновременно рас-
Учет когерентного артефакта и деконволю-
считывается финальный спектр Dn+1(λ). После
ции аппаратной функции в области t = 0 про-
того как найдены оптимальные значения {τk}
водился по методике, предложенной в работах
характерных времен, стандартную ошибку их
Dobryakov et al. [26, 28]. Для этого абсорбци-
определения можно оценить с помощью ма-
онные изменения, вызванные нелинейными
трицы Якоби J:
взаимодействиями в области перекрытия воз-
δQj[τ] ,
буждающего и зондирующего импульсов (коге-
Jjk =
(6)
δτk
рентный спайк), аппроксимировались базовой
гауссовой функцией:
где индекс j пробегает L × M эксперименталь-
ных точек спектрально-временной матрицы
G(t) = 1/√2πd2 exp[-½(t/d)2]
(1)
ΔA(λl, tm) [30]. Численную оценку якобиана
БИОХИМИЯ том 88 вып. 10 2023
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ТЕТРАМЕРА ХЛОРОФИЛЛА
1911
можно получить, разложив выражение Qj[τ] в
ны 380 нм соответствует переходу S0 → S4 [32])
ряд Тейлора до первого порядка:
заметно изменены в пользу полосы с максиму-
мом на ~380 нм. Электронная структура спек-
δQj[τ]
Qj[τ + Δτek]-Qj [τ - Δτek],
≈
(7)
тра поглощения Хл a в области полосы Соре
δτk
2Δτk
плохо описывается в рамках четырехорбиталь-
где τ - набор оптимальных значений {τk} ха-
ной модели, предложенной Gouterman [33],
рактерных времен, ek - k-й единичный вектор,
поскольку между возбужденными состояния-
а Δτ - небольшое приращение τ (для число-
ми Bx и By присутствуют состояния не соответ-
вой оценки использовалось значение 0,015τ).
ствующие четырехорбитальной модели, сме-
Стандартные ошибки σk определения харак-
шанные с вибронными переходами полосы Bx,
терных времен можно получить из диагональ-
в результате энергии и дипольные силы этих
ных элементов ковариационной матрицы C =
состояний зависят от диэлектрического окру-
= R[τ] × (JTJ)-1:
жения пигмента [32, 34, 35].
Как видно из рис. 1, а, в белковом окру-
σk = √Ckk.
(8)
жении BoWSCP относительная вероятность
перехода S0 → S3 уменьшена, а вероятность пе-
Значения σk представляют собой стандарт-
рехода S0 → S4 увеличена по сравнению со свой-
ные отклонения, характеризующие неопре-
ствами свободного Хл a в растворе. По этой
деленность оценок {τk}. Согласно t-распреде-
причине моделирование спектра поглощения
лению Стьюдента, 95%-ная достоверность по-
BoWSCP проводилось с помощью эмпири-
лучается при удвоении стандартных отклоне-
ческого выражения, представляющего собой
ний; соответственно, доверительные интер-
сумму модифицированных спектров поглоще-
валы равны τk ± 2σk.
ния Хл a и Хл b и учитывающего перераспре-
деление интенсивности полос Хл a в области
360-500 нм:
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
A(λ) = α⋅AChla(λ - δа) ×
(9)
Следуя методу, предложенному в работе
× [1 - γ⋅tanh((λ - λа / Δ)] + β⋅AChlb(λ - δb),
Hughes et al. [20], пигментный состав полу-
ченных препаратов холоформ BoWSCP был
где α и β - относительные количества Хл a
определен с помощью разложения его спек-
и Хл b в тетрамере BoWSCP; AChla и AChlb - спек-
тра поглощения в области полосы Соре (360-
тры Хл a и Хл b в 90%-ном ацетоне; δa и δb -
500 нм) на относительные вклады Хл a и Хл b
сдвиги спектров Хл a и Хл b в BoWSCP относи-
с использованием спектров поглощения ин-
тельно их спектров в растворе; эмпирическая
дивидуальных пигментов в 90%-ном ацето-
функция γ tanh((λ-λa)/Δ) описывает измене-
не [31]. На рис. 1, а проведено сопоставление
ние соотношения интенсивности полос Bx и By
спектра поглощения BoWSCP со спектрами
Хл a в спектральном интервале шириной Δ с
Хл a и b. В спектре поглощения свободного
центром на длине волны λa. Значения указан-
Хл a полосы поглощения Соре и Qy с макси-
ных параметров, определенные с помощью не-
мумами около 430 и 663 нм имеют близкую
линейной минимизации, составили α = 89,2%;
амплитуду; в спектре Хл b эти полосы сбли-
β = 10,8%; δa = 9,2 нм; δb = 1,4 нм; γ = 0,67;
жены и располагаются соответственно при 458
λa = 413 нм; Δ = 50 нм; модельный спектр по-
и 645 нм, при этом амплитуда полосы Соре
казан на рис. 1, б тонкой сплошной линией.
увеличена, а полосы Qy - уменьшена относи-
Относительные вклады Хл a и Хл b получились
тельно полос Хл a. Присутствие Хл b в спектре
равными 89% и 11% соответственно. Соот-
BoWSCP выявляется по небольшому плечу в
ношение пигментов Хл a/b в пропорции 8 : 1
области 470 нм, тогда как поглощение в ин-
соответствует диапазону значений (6-10) : 1,
тервале 380-430 нм обусловлено преимуще-
полученных в разных работах для нативного
ственно Хл a. Сравнение спектров BoWSCP
белка BoWSCP [3].
и свободного Хл a демонстрирует существен-
Поглощение BoWSCP в области полосы Qy
ный эффект белкового окружения в области
(660-700 нм) обусловлено преимущественно
360-450 нм: во-первых, полосы Хл a сдвинуты
поглощением Хл a, вклад Хл b в этом спек-
в красную область на ~8 нм, во-вторых, ин-
тральном интервале незначителен (рис. 1, б).
тенсивности двух основных переходов Хл a в
Спектральные свойства BoWSCP определя-
области полосы Соре (полоса Bx с максимумом
ются молекулярной структурой пигмент-бел-
на длине волны 430 нм соответствует переходу
кового комплекса: четыре идентичных субъ-
S0 → S3, полоса By с максимумом на длине вол-
единицы образуют тетрамер, в котором три
БИОХИМИЯ том 88 вып. 10 2023
1912
ЧЕРЕПАНОВ и др.
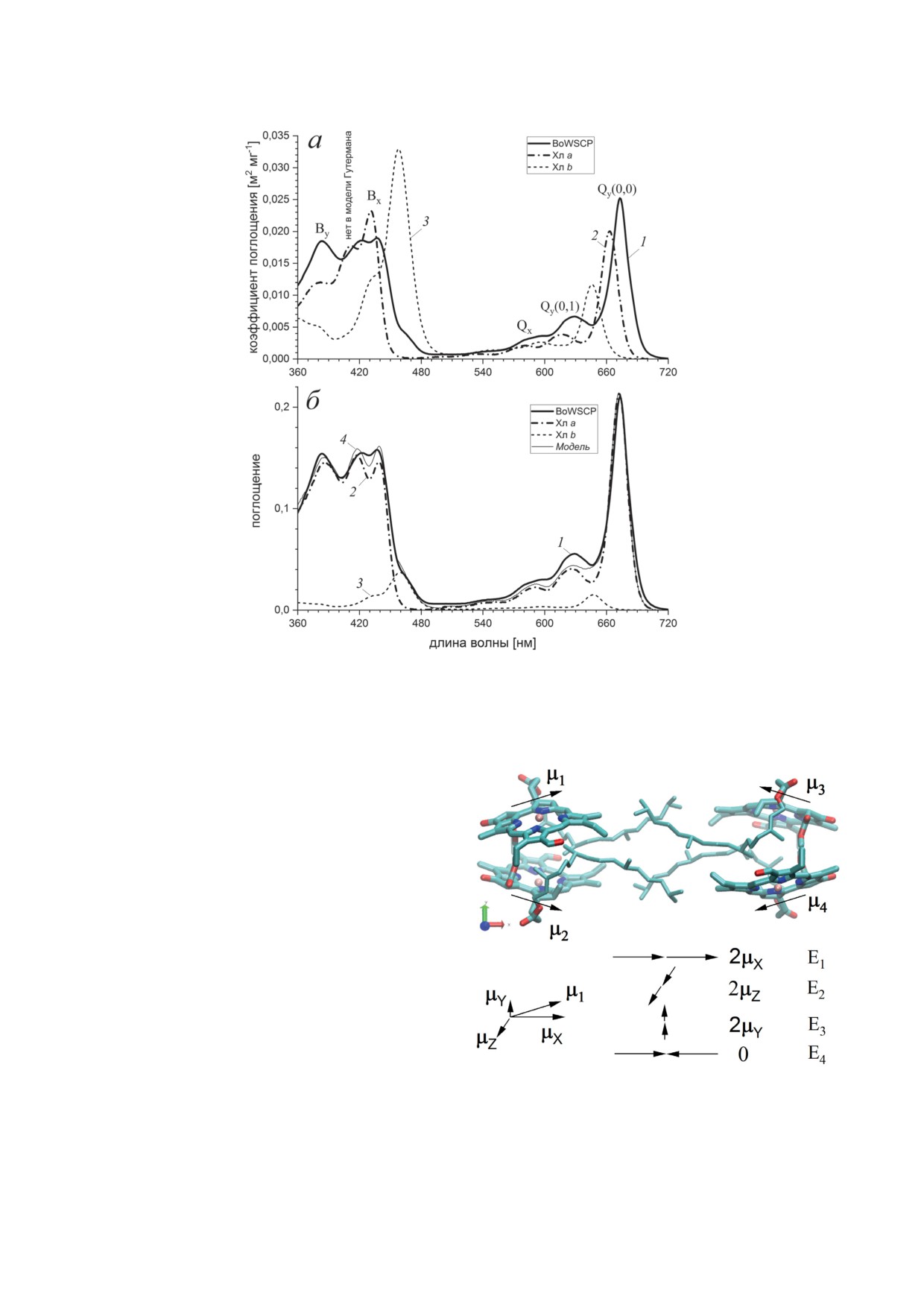
Рис. 1. Определение вкладов Хл a и Хл b в спектр поглощения BoWSCP. а - Сравнение спектра поглощения холо-
формы BoWSCP в буферном растворе (1) со спектрами Хл a (2) и Хл b (3) в 90%-ном ацетоне (взяты из открытой базы
спектральных данных [31]). б - Моделирование спектра BoWSCP (1) с помощью модифицированного, в соответствии
с уравнением (9), спектра Хл a (2) и Хл b (3). В области 500-720 нм амплитуда спектра Хл a увеличена на 15%. Относи-
тельные вклады Хл a и Хл b в модельный спектр (4) составляют 89% и 11% соответственно
взаимно ортогональных оси симметрии вра-
щения C2(X) C2(Y) C2(Z) задают систему коор-
динат XYZ [18]. Четыре молекулы хлорофилла,
находящиеся в гидрофобном локусе, объеди-
нены в двойной димер: взаимодействие между
димерами существенно слабее взаимодействия
мономеров внутри димера (рис. 2).
Преобладающим эффектом в области 660-
700 нм является экситонное взаимодействие
молекул Хл a внутри димера, тогда как взаи-
модействие димеров друг с другом выражено
значительно слабее. Форма спектра поглоще-
ния BoWSCP в области полосы Qy (рис. 3, а)
свидетельствует о наличии двух экситонных
полос, максимумы которых при 672 и 684 нм
соответствуют минимумам второй производ-
ной (рис. 3, б) и максимумам четвертой про-
изводной (рис. 3, в). Следуя методу, предло-Рис. 2. Расположение молекул хлорофилла в тетрамерном
женному для анализа спектров WSCP [20], комплексе BoWSCP по данным рентгеновской кристал-
лографической структуры PDB ID: 6s2z [18], в которой
параметры экситонного взаимодействия Хл a
Хл b заменен на Хл a. Проиллюстрировано направление
были найдены разложением спектра погло-
векторов μk (k 1-4) дипольных моментов S0 → S1 перехо-
да мономеров Хл, а также дипольных моментов четырех
щения BoWSCP в области полосы Qy(0,0)
на гауссовы компоненты. Для корректной экситонных состояний En тетрамерного комплекса
БИОХИМИЯ том 88 вып. 10 2023
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ТЕТРАМЕРА ХЛОРОФИЛЛА
1913
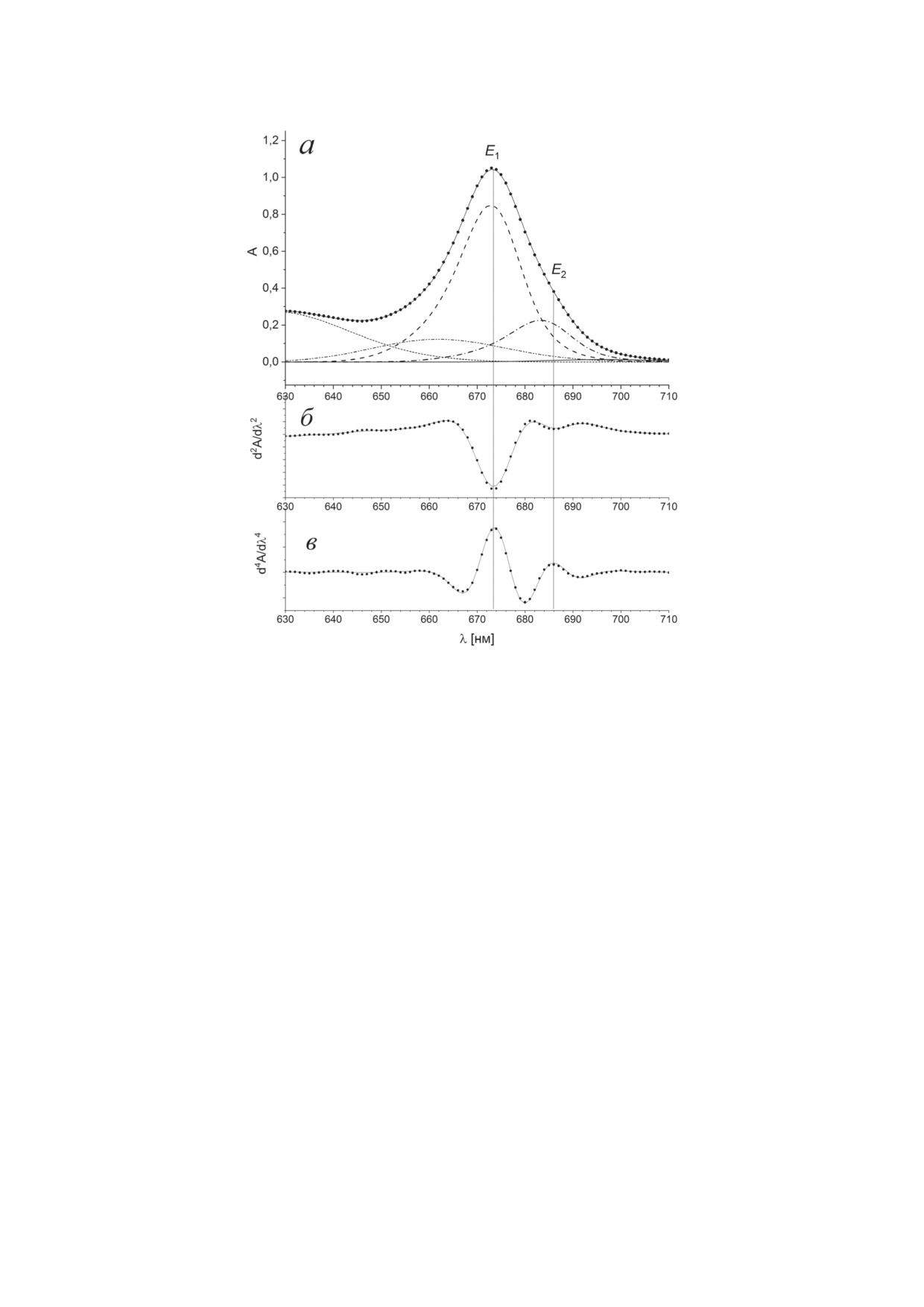
Рис. 3. Спектр поглощения (а), его вторая (б) и четвертая (в) производные холоформы BoWSCP (точки). Сплошными
линиями представлен модельный спектр, его основные гауссовские компоненты показаны пунктирными линиями.
Вертикальным пунктиром отмечены положения максимумов четвертой производной спектра поглощения
аппроксимации полосы Qy(0,0) с учетом внут-
670 нм (б), индуцированные импульсом 670 нм
римолекулярных колебательных мод тетра-
длительностью 16 фс (толстые сплошные ли-
пирольного макроцикла Хл использовались
нии). Когерентный артефакт был аппроксими-
дополнительные гауссовы компоненты в высо-
рован суперпозицией гауссовой функции G(t)
коэнергетической области полосы Qy(0,0) [20,
(уравнение (1)) и ее производных G′ и G′′ с
36, 37]. Относительная интенсивность основ-
параметром d, равным 24 фс (пунктирные ли-
ной полосы E1 с максимумом на длине волны
нии), а абсорбционные изменения BoWSCP -
672 нм составила 79%, вклад второй полосы E2
комбинацией функций χ(t) и ε(t, τk), задавае-
с максимумом на длине волны 684 нм - ~21%,
мых уравнениями (2) и (3) (штрихпунктирные
спектральные характеристики состояния E3
линии). Абсорбционные изменения BoWSCP,
определить не удалось, состояние E4 в прибли-
начиная с нулевой временной задержки (t = 0),
жении точечных диполей имеет нулевую ин-
были получены вычитанием когерентного арте-
тенсивность.
факта и деконволюции результирующего сиг-
Методом широкополосной фемтосекунд-
нала (тонкие сплошные линии).
ной спектроскопии «возбуждение-зондирова-
На рис. 5, а приведены переходные спек-
ние» была измерена абсорбционная динамика
тры при временной задержке 50 фс, индуци-
холоформы BoWSCP, индуцированная возбуж-
рованные импульсами 430 нм (1, сплошная
дением в области длин волн 430 и 670 нм.
линия) и 670 нм (2, штрихпунктирная линия).
Возбуждение в этих спектральных областях
Возбуждение на длине волны 430 нм связано с
связано преимущественно с Хл a (кванто-
переходом Хл преимущественно в третье син-
вый выход >98%) и лишь в малой степени -
глетное возбужденное состояние S0 → S3 (по-
с Хл b (<2%). На рис. 4 приведены абсорбци-
лоса Bx), тогда как при возбуждении на длине
онные изменения в фемтосекундном времен-
волны 670 нм происходило образование со-
ном диапазоне на длинах волн 450 нм (а) и
стояния S1 (полоса Qy). Данную интерпретацию
БИОХИМИЯ том 88 вып. 10 2023
1914
ЧЕРЕПАНОВ и др.
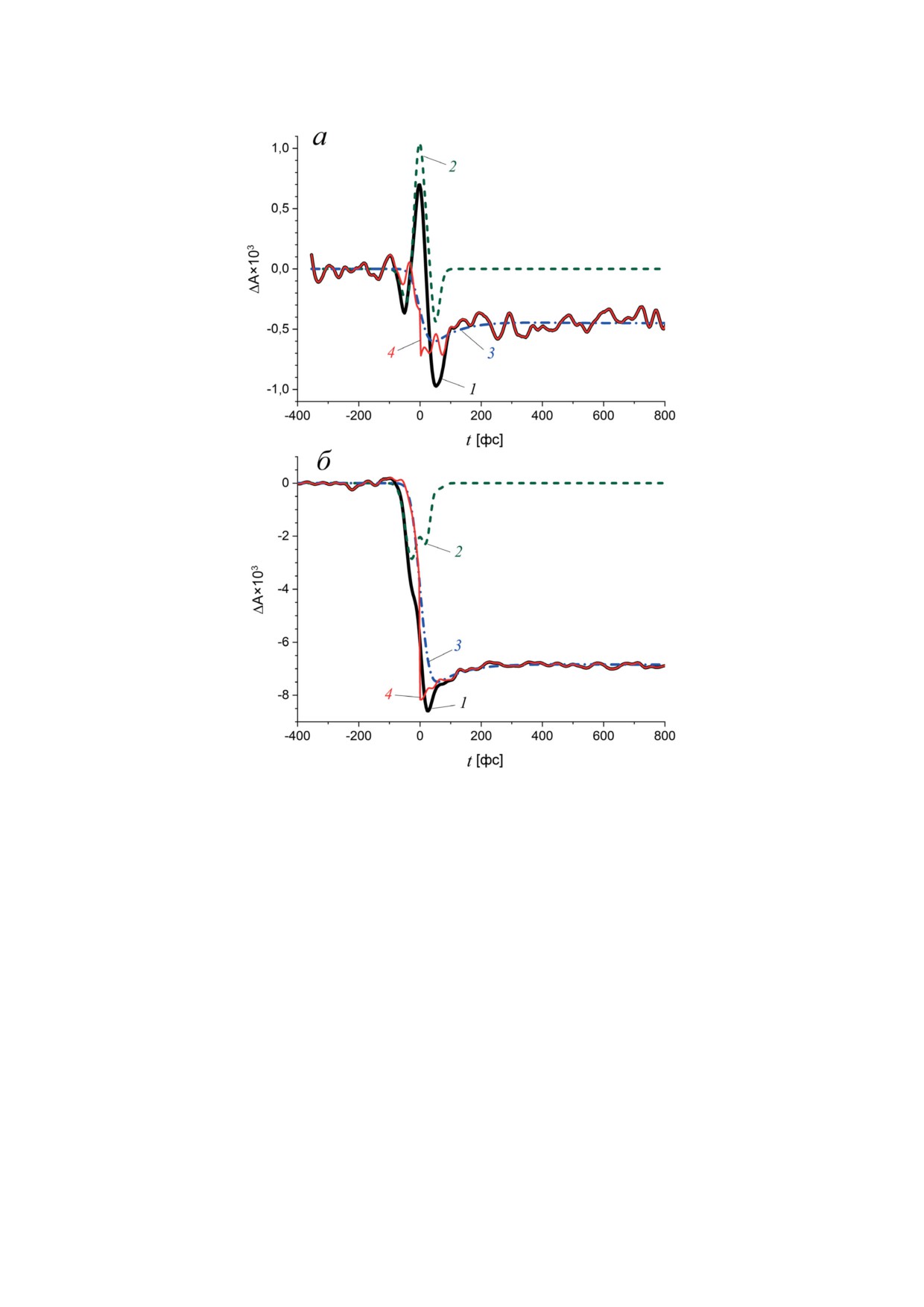
Рис. 4. Абсорбционные изменения BoWSCP на длине волны 450 нм (а) и 670 нм (б), индуцированные импульсом
670 нм длительностью 16 фс (линия 1). Когерентный артефакт был аппроксимирован суперпозицией гауссовой
функции G(t) и ее производных G′ и G′′, рассчитанных для значения параметра d = 24 фс (линия 2), а абсорбцион-
ные изменения BoWSCP - комбинацией функций χ(t) и ε(t, τk) (линия 3). Абсорбционный отклик BoWSCP с нуле-
вой временной задержкой (t = 0) был получен вычитанием когерентного артефакта и деконволюцией результирую-
щего сигнала (линия 4)
подтверждает отрицательный коэффициент
новного состояния S0, его форма соответствует
анизотропии в области полосы Qy при воз-
линейному спектру поглощения с отрицатель-
буждении на длине волны 430 нм, величина
ным знаком. Во-вторых, это спектр поглощения
которого близка к теоретическому пределу -0,1
возникающего возбужденного состояния S*,
(вставка на рис. 5, а).
его вклад всегда положительный. В-третьих,
Амплитуда выцветания полосы Qy в пере-
это спектр стимулированного излучением
ходных спектрах BoWSCP на рис. 5 в 3-5 раз
обратного перехода S* → S0, для нижнего воз-
больше амплитуды выцветания полосы Соре,
бужденного состояния его форма близка спек-
тогда как в спектре поглощения BoWSCP
тру спонтанной флуоресценции, а вклад -
на рис. 1, а полоса Qy сравнима по амплитуде
отрицательный. Основное различие между
с полосой Соре. Переходный спектр, наблю-
спектрами (1) и (2) на рис. 5, а наблюдается
даемый в фемтосекундных измерениях, пред-
в области полосы Qy, где имеет место стимули-
ставляет собой сумму спектров трех состав-
рованное излучение перехода S1 → S0, которое
ляющих. Во-первых, это спектр выцветания,
отсутствует на временах жизни состояния S3
появляющийся в результате исчезновения ос-
при возбуждении на длине волны
430 нм.
БИОХИМИЯ том 88 вып. 10 2023
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ТЕТРАМЕРА ХЛОРОФИЛЛА
1915
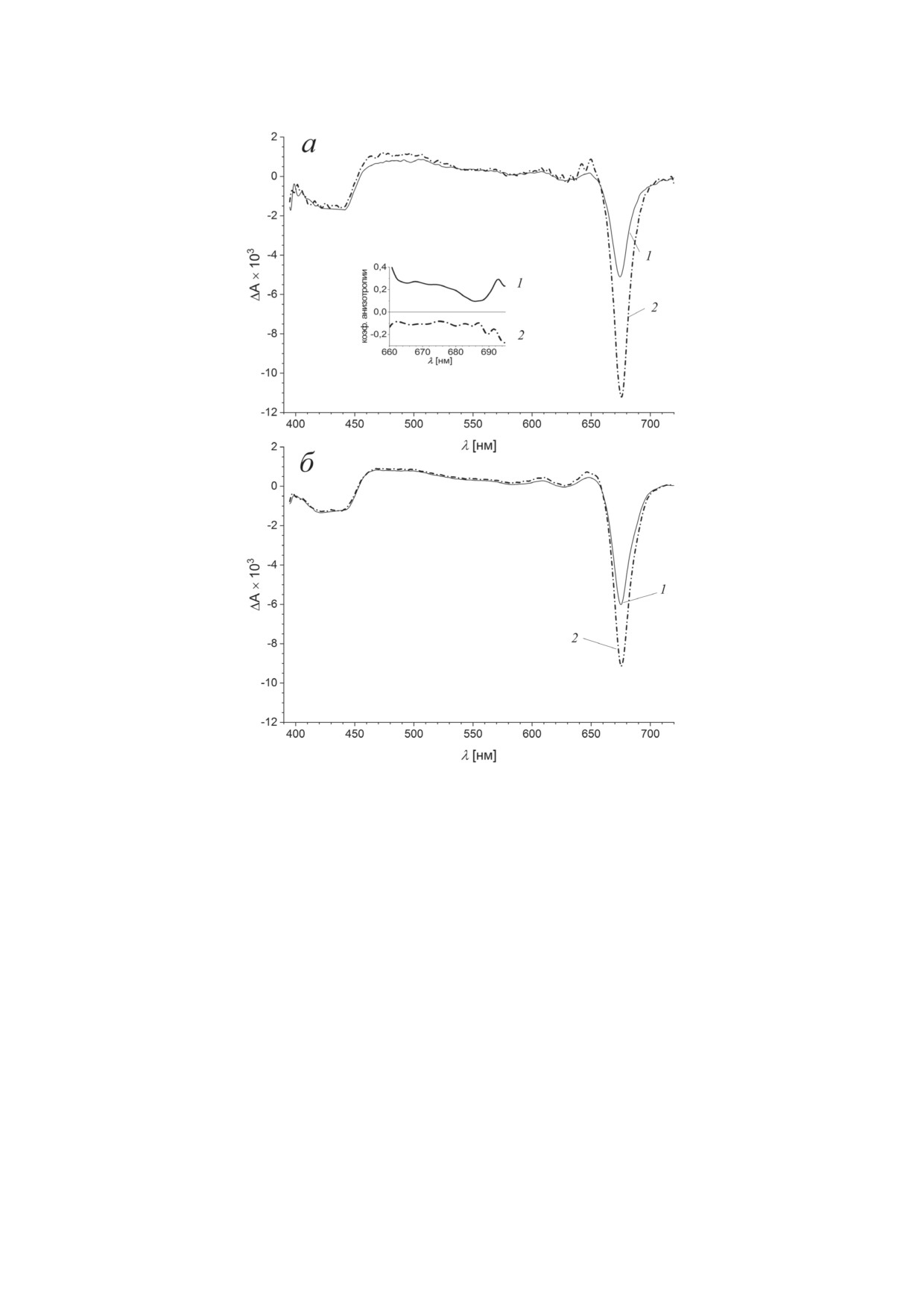
Рис. 5. Переходные спектры BoWSCP на задержках 50 фс (а) и 200 пс (б), инициированные возбуждением на длинах
волн 430 нм (1) и 670 нм (2). Спектры масштабированы по области полосы Соре 400-500 нм при задержке 200 пс;
угол поляризации - 54,7°. На вставке показан коэффициент анизотропии в области полосы Qy
Менее значительное различие между спектра-
Наиболее значимые абсорбционные из-
ми (1) и (2) в области 420-500 нм может быть
менения BoWSCP связаны с увеличением
объяснено стимулированным излучением пе-
амплитуды выцветания полосы Qy после воз-
рехода S3 → S0.
буждения на длине волны 430 нм. Его количе-
На рис. 5, б приведены переходные спек-
ственные характеристики были определены с
тры при временной задержке 200 пс в тех же
помощью разложения абсорбционной динами-
обозначениях: (1) - возбуждение 430 нм, (2) -
ки на экспоненциальные компоненты, связан-
670 нм. В этом временном диапазоне должны
ные с распадом (DAS, см. раздел «Материалы
завершиться процессы внутримолекулярной
и методы»); разложение выявило спектральные
конверсии S3 → S1 и установления теплового
переходы в диапазоне 100 фс и 10 пс (рис. 6).
равновесия в распределении четырех экситон-
На рис. 6, а представлены дифференци-
ных состояний En тетрамерного комплекса.
альные спектры распада для фемтосекундного
Наблюдаемые финальные спектры близки во
диапазона. После возбуждения на длине вол-
всем диапазоне за исключением полосы Qy,
ны 430 нм основные спектральные изменения
где амплитуда выцветания после возбуждения
происходят с характерным временем 83 ± 9 фс,
на длине волны 670 нм значительно превос-
они могут быть приписаны внутримолеку-
ходит выцветание, индуцированное возбужде-
лярной безызлучательной конверсии S3 → S1, в
нием на длине волны 430 нм.
результате которой исчезает стимулированное
БИОХИМИЯ том 88 вып. 10 2023
1916
ЧЕРЕПАНОВ и др.
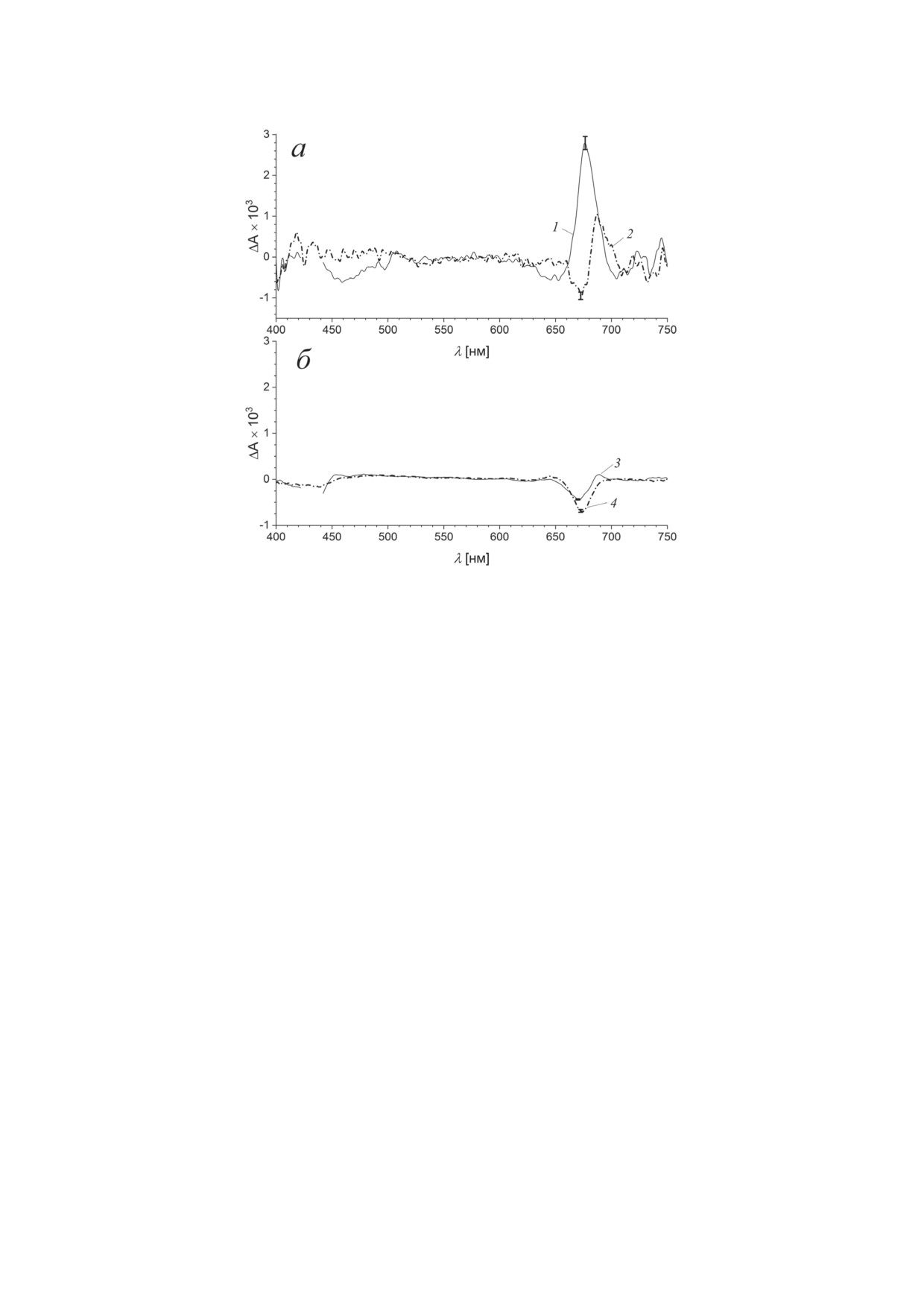
Рис. 6. Ассоциированные с экспоненциальным распадом спектры BoWSCP в диапазоне 100 фс (а) и 10 пс (б) после
возбуждения на длинах волн 430 нм (1, 3) и 670 нм (2, 4). Характерные времена: 1 - 83 фс; 2 - 105 фс; 3 - 19 пс; 4 -
14 пс. Отметки на графиках в области полосы Qy показывают доверительные интервалы, рассчитанные с помощью
уравнения (8)
излучение в области полосы Соре
(450-
выцветание основного состояния (Ground State
480 нм) и появляется стимулированное излу-
Bleach, GSB), поглощение возбужденного со-
чение в области полосы Qy (670-690 нм). При
стояния (Excited State Absorption, ESA) и сти-
возбуждении BoWSCP в области полосы Qy
мулированное излучение (Stimulated Emission,
на длине волны 670 нм небольшие спектраль-
SE). Вероятность поглощения отдельной мо-
ные изменения в субпикосекундном диапа-
лекулой пигмента кванта света энергии ħω0
зоне происходят со временем 105 ± 10 фс, они
пропорциональна квадрату дипольного мо-
могут быть отнесены к релаксационному пере-
мента |μ|2 ее перехода между основным и воз-
ходу между двумя экситонными уровнями E1
бужденным состояниями (дипольной силе
и E2 полосы Qy димера Хл a.
перехода). Для мономера Хл a в белке диполь-
В пикосекундном диапазоне значимые из-
ный момент перехода S0 → S1 равен 5,5 Д [38].
менения поглощения происходят с характерным
Импульс накачки переводит в образце ΔN мо-
временем ~16 пс. Дифференциальные спектры
лекул из основного состояния в возбужденное.
распада, рассчитанные для возбуждения на дли-
Пробный импульс разделяется на два пучка:
нах волн 430 нм (характерное время 19 ± 3 пс)
один проходит через невозбужденный образец,
и 670 нм (характерное время 14 ± 2 пс), близки
другой - через область возбуждения, разница в
во всем спектральном диапазоне (рис. 6, б)
интенсивности прошедших пучков определяет
и связаны с незначительным по амплитуде
дифференциальное поглощение. В области воз-
восстановлением поглощения в области по-
буждения количество молекул в основном со-
лосы Qy с максимумом около 675 нм. Возмож-
стоянии уменьшено на ΔN, поэтому поглоще-
ная интерпретация этих изменений будет рас-
ние на частоте ω0 в этой области уменьшено
смотрена далее.
на величину, пропорциональную дипольной
Для количественной характеристики фото-
силе перехода: ΔAGSB ∝ |μ|2. Одновременно с этим
химических процессов, наблюдаемых с помо-
имеет место стимулированное световой вол-
щью фемтосекундных измерений в экситон-
ной излучение света при обратном переходе
но-сопряженной системе пигментов, было
S1→ S0, вероятность которого также пропорцио-
проведено разложение переходных спектров
нальна квадрату дипольного момента, поэто-
(Transient Absorption, TA) на три составляющих:
му поглощение на частоте ω0 дополнительно
БИОХИМИЯ том 88 вып. 10 2023
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ТЕТРАМЕРА ХЛОРОФИЛЛА
1917
уменьшается на ту же величину ΔASE ∝ |μ|2. Для
В остальной части спектрального диапазона
идеальной двухуровневой системы отношение
выцветание основного состояния пропорцио-
β = ΔASE/ΔAGSB равно единице [39]; для Хл a в
нально дипольной силе свободного мономера:
различных растворителях была получена оцен-
ка β ≈ 0,9 [40, 41].
ΔAGSB(ω) ∝ μ2.
(12)
Для экситонно-сопряженных пигментов
ситуация более сложная. Дипольный момент
Для симметричного тетрамера дипольные
перехода экситонного комплекса можно пред-
моменты перехода в четыре возможных экси-
ставить линейной комбинацией дипольных
тонных состояния находятся унитарным пре-
моментов его мономеров [42]. В кристалло-
образованием [42], момент с наибольшей ам-
графической структуре BoWSCP [18] диполь-
плитудой равен:
ные моменты мономеров μk ориентированы
практически параллельно кристаллографиче-
M1 = ½(μ1 + μ2 - μ3 - μ4).
(13)
ской оси X, а проекции моментов на оси Y и Z
много меньше по абсолютной величине [13],
В этом случае суммарная амплитуда вы-
и в первом приближении их можно положить
цветания переходного спектра вблизи полосы
равными нулю. Проекции дипольного момен-
экситонного расщепления:
та мономера на кристаллографические оси
схематически изображены на рис. 2.
ΔABL ∝ 8μ2.
(14)
При описании взаимодействия световой
волны с молекулами Хл в BoWSCP возникает
В целом количество экситонно-сопряжен-
фундаментальный вопрос: является ли возни-
ных молекул nexcited, между которыми делока-
кающее возбужденное состояние локализован-
лизовано возбужденное состояние, возможно
ным в пределах димера (энергия сопряжения
определить по отношению суммарной ампли-
~100 см-1) или оно делокализовано по всему
туды выцветания переходного спектра на ча-
тетрамерному комплексу (энергия сопряжения
стоте экситонного перехода ω0 и амплитуды
~10 см-1). Данные фемтосекундных измерений
линейного спектра, который не учитывает се-
позволяют определить степень делокализации
лективный характер возбуждения экситонного
возбужденного состояния, то есть количество
комплекса:
молекул Хл, эффективно участвующих в экси-
тонном сопряжении.
nexcited = ½ΔABL/ΔAGSB.
(15)
Для симметричного димера дипольные мо-
менты перехода в два возможных возбужден-
При возбуждении BoWSCP в области 670 нм
ных состояния E+ и E- равны [43]:
амплитуду выцветания основного состоя-
ния ΔAGSB можно определить по полосе Соре.
1
M± =
(μ1 ± μ2).
(10)
На рис. 7 точками показаны переходные спек-
√2
тры BoWSCP при временной задержке 50 фс
Следует отметить, что экситонное взаимо-
после возбуждения на длинах волн 430 нм (а)
действие мономеров не меняет суммарную
и 670 нм (б).
дипольную силу комплекса:
|M+|2 + |M-|2 =
В области 400-500 нм переходные спек-
= μ2 + μ2, поэтому интегральная амплитуда ли-
тры (1) близки по форме к спектру погло-
нейного спектра поглощения остается посто-
щения BoWSCP (2) за исключением области
янной. В силу геометрии BoWSCP дипольная
450-480 нм, где преобладает поглощение Хл b
сила перехода в состояние E+ равна |M+|2 ≅
и стимулированное излучение Хл a из состоя-
≅ μ2 + μ2 = 2μ2, тогда как дипольная сила пере-
ния S3. Предполагая, что спектры поглощения
хода в состояние E- близка к нулю, поэтому
возбужденных состояний S1 и S3 являются глад-
при возбуждении BoWSCP широкополосным
кими функциями в указанном интервале длин
импульсом с центром на длине волны 670 нм
волн и могут быть корректно описаны полино-
(полуширина - 50 нм, время τ = 16 фс) в пер-
мами P3(λ) третьей степени, переходные спек-
воначальном распределении возникающих эк-
тры были аппроксимированы выражением:
ситонных состояний существенно преобладает
состояние E+. В этом случае суммарная ампли-
ΔATA(λ) = ξ⋅ΔAGSB(λ) + P3(λ),
(16)
туда выцветания ΔABL начального переходного
спектра вблизи полосы экситонного расщеп-
в котором масштабирующий множитель ξ
ления на частоте ω0:
и коэффициенты кубического полинома P3
находились стандартным решением задачи
ΔABL = ΔAGSB + ΔASE ∝ (1 + β)2μ2 ≈ 4μ2.
(11)
линейной регрессии. Спектры поглощения
БИОХИМИЯ том 88 вып. 10 2023
1918
ЧЕРЕПАНОВ и др.
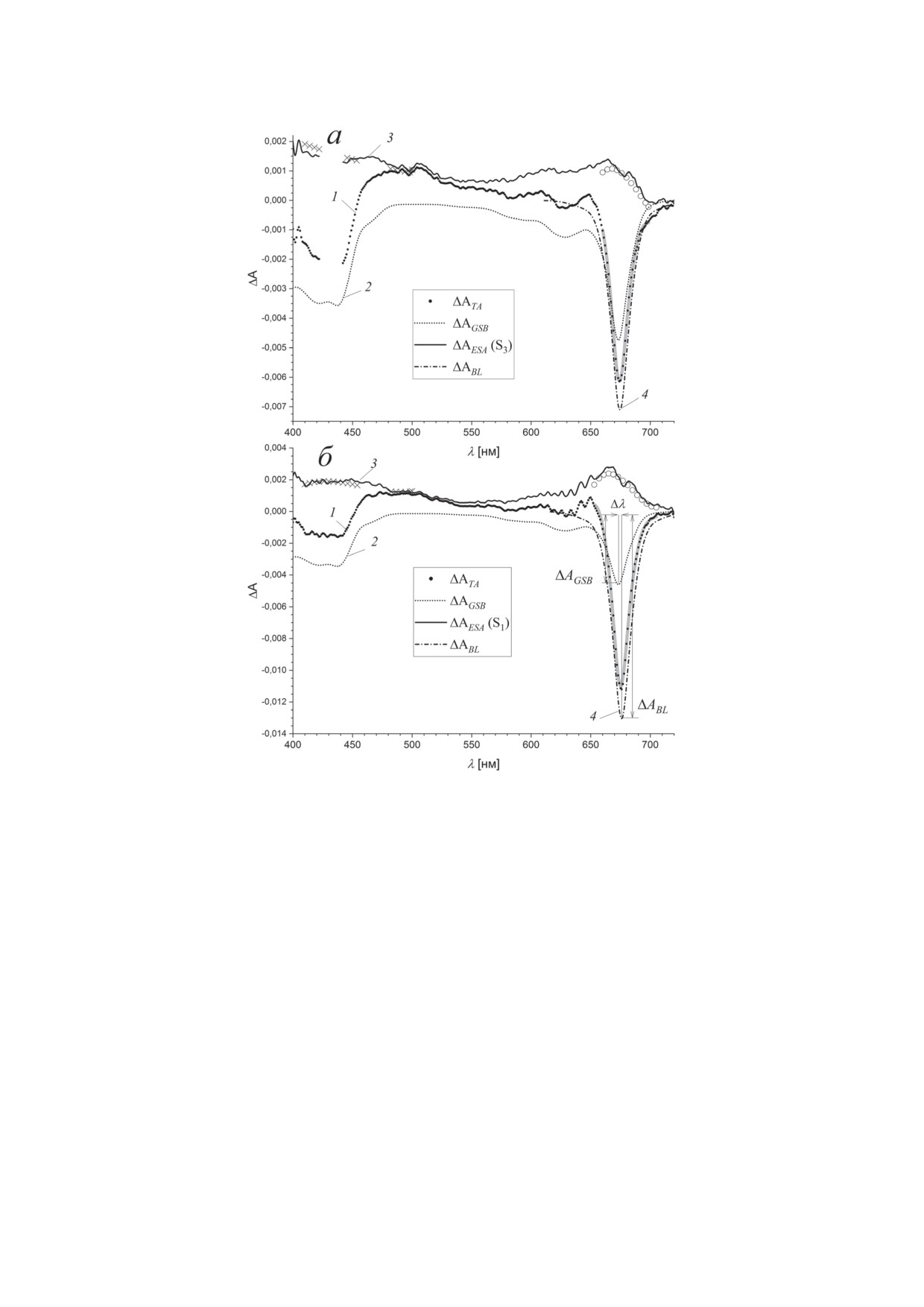
Рис. 7. Разложение переходных спектров поглощения BoWSCP (1 - Transient Absorption, TA) на компоненты выцвета-
ния основного состояния (2 - Ground State Bleaching, GSB), поглощения возбужденного состояния (3 - Excited State
Absorption, ESA) и выцветания в области стимулированного излучения (4 - Bleach, BL). Переходные спектры получе-
ны при задержке 50 фс после возбуждения на длинах волн 430 нм (а) и 670 нм (б). Нормировка спектра выцветания (2)
проводилась по переходному спектру (1) в области 400-500 нм с учетом аппроксимации спектра возбужденного со-
стояния кубической полиномиальной функцией (×). Модельный спектр выцветания в области стимулированного
излучения (4) в диапазоне 650-700 нм был представлен суммой ΔABL = nexcited·(ΔAGSB+ΔASE) линейного спектра поглоще-
ния и его зеркального отражения относительно максимума полосы поглощения Qy и сдвига Стокса Δλ. Относительная
амплитуда модельного спектра nexcited находилась линейной регрессией с помощью полиномиальной аппроксимации
спектра возбужденного состояния (○). Утолщенная серая линия представляет результат данного разложения
возбужденных состояний в области, где отсут-
Поскольку положение
0-0-перехода остава-
ствует стимулированное излучение, находятся
лось неизвестным, величину сдвига Стокса Δλ
как разность ΔAESA(λ) = ΔATA(λ) - ξ⋅ΔAGSB(λ).
спектра стимулированного излучения нахо-
На рис. 7 показаны полученные таким образом
дили нелинейной минимизацией суммарного
спектры выцветания основного состояния (2),
модельного спектра:
полиномиальные функции P3 (×) и спектры
ΔATA(λ) = nexcited ⋅ξ⋅[ΔAGSB(λ) +
поглощения возбужденных состояний (3).
(17)
Модельный спектр выцветания (4) вблизи
+ ΔAGSB(λ0 + Δλ - λ)] + P3(λ).
полосы Qy был представлен суммой линей-
ного спектра поглощения, взятого с обратным
Эффективное число экситонно-сопряжен-
знаком ΔAGSB(λ),и его зеркального отражения
ных молекул nexcited и коэффициенты полино-
относительно максимума полосы Qy ΔASE(λ).
миальной функции P3 находились линейной
БИОХИМИЯ том 88 вып. 10 2023
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ТЕТРАМЕРА ХЛОРОФИЛЛА
1919
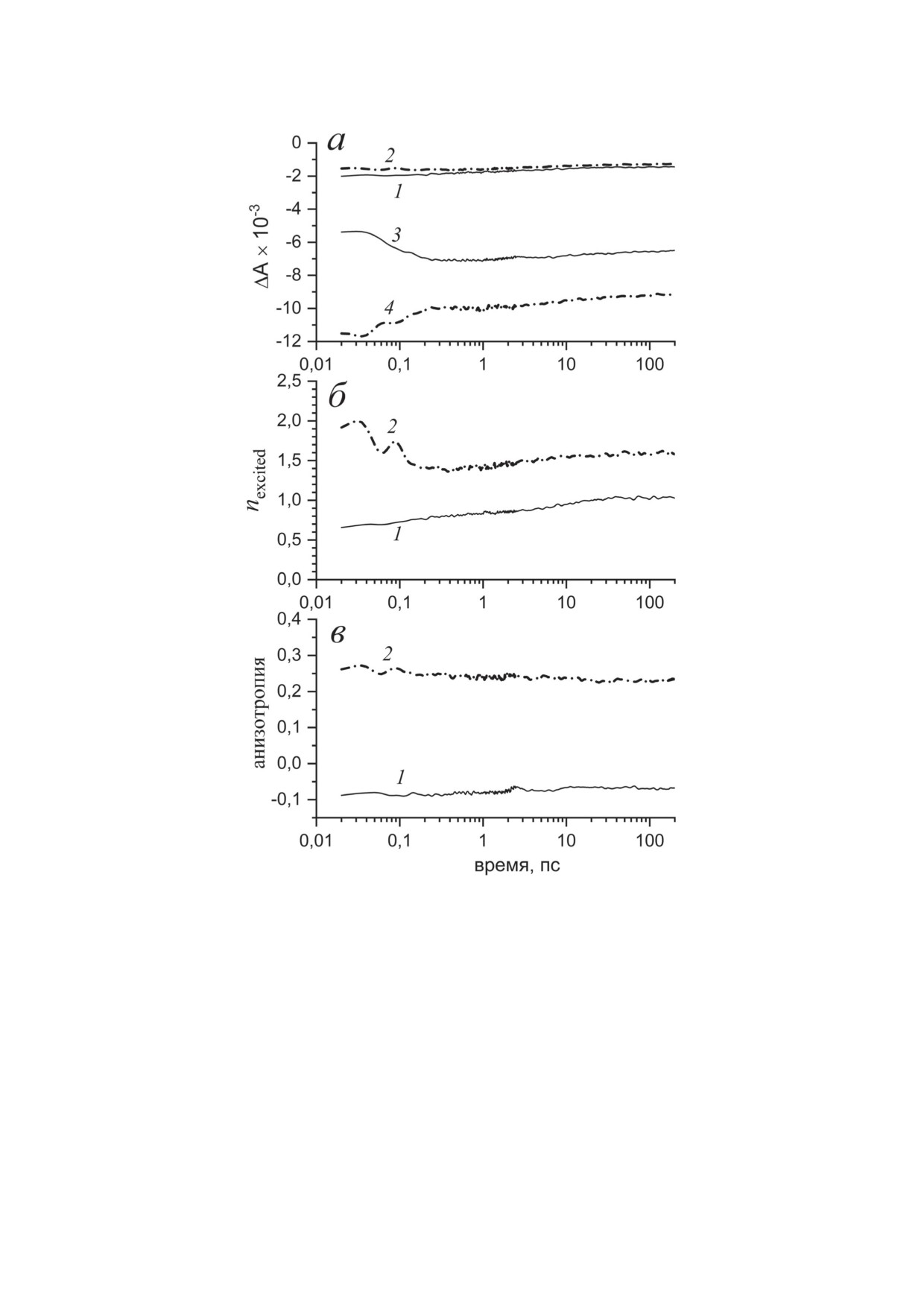
Рис. 8. Изменение во времени основных параметров, характеризующих возбужденное состояние тетрамерного ком-
плекса BoWSCP. а - Амплитуда выцветания полосы Соре (1, 2) и полосы Qy (3, 4) после возбуждения на 430 нм (1, 3)
и 670 нм (2, 4). б - Эффективное число экситонно-сопряженных молекул Хл, рассчитанное по уравнению (15) при
возбуждении на 430 нм (1) и 670 нм (2). в - Коэффициент анизотропии, рассчитанный в области полосы Qy при воз-
буждении на 430 нм (1) и 670 нм (2)
регрессией. Серая утолщенная линия на рис. 7
и полосы Qy (3, 4), инициированная возбужде-
представляет результат данного разложения.
нием BoWSCP на длинах волн 430 нм (сплош-
При возбуждении на длине волны 430 нм на-
ные кривые) и 670 нм (штрихпунктирные кри-
блюдаемая амплитуда выцветания ΔABL поло-
вые). Изменение поглощения во временном
сы Qy лишь незначительно превышает ампли-
диапазоне 100 фс при возбуждении на длине
туду выцветания основного состояния ΔAGSB,
волны 430 нм (3) отражает появление стиму-
тогда как при возбуждении на длине волны
лированного излучения в результате электрон-
670 нм амплитуда ΔABL втрое превышает ам-
ного перехода S3 → S1, его характерное время
плитуду ΔAGSB.
составляет 80 фс. Как было отмечено выше,
На рис. 8, а показана временная зависи-
амплитуда выцветания полосы Qy после воз-
мость амплитуды выцветания полосы Соре (1, 2)
буждения в области 430 нм остается во всем
БИОХИМИЯ том 88 вып. 10 2023
1920
ЧЕРЕПАНОВ и др.
временном интервале существенно меньше
дихроизма, составляет ~100 см-1 [20], что по-
амплитуды выцветания этой полосы после
зволяет рассматривать делокализованные воз-
возбуждения в области 670 нм. Другой особен-
бужденные состояния пигментов в терминах
ностью является высокая амплитуда выцве-
теории экситонов [42]. Величина экситонного
тания полосы Qy после возбуждения на длине
взаимодействия димеров в тетрамере WSCP,
волны 670 нм во всем временном интервале:
рассчитанная на основе рентгеновской струк-
его амплитуда втрое превышает амплитуду вы-
туры комплекса, составляет всего 10 см-1 [13],
цветания полосы Соре.
что позволяет описывать перенос энергии ме-
На рис. 8, б показано изменение во вре-
жду димерами в рамках теории Förster [52].
мени эффективного числа экситонно-сопря-
Измерение абсорбционной динамики бел-
женных молекул Хл, между которыми делока-
ков WSCP подклассов IIa (BoWSCP, из Bras-
лизовано возбужденное состояние тетрамера,
sica oleracea) и IIb (LvWSCP, из Lepidium vir-
рассчитанного по уравнению (15), для двух
ginicum) в фемтосекундном диапазоне про-
вариантов возбуждении BoWSCP. На коротких
водилось ранее в области полосы Qy мето-
задержках при возбуждении на длине волны
дом двухимпульсной спектроскопии «возбужде-
670 нм коэффициент nexcited приближается к 2,
ние-зондирование» [14] и методом четырехим-
то есть комплекс BoWSCP ведет себя как экси-
пульсной 2D-спектроскопии [12, 13]. В работе
тонно-сопряженный димер. При возбуждении
Theiss et al. [14] измерялись абсорбционные
на длине волны 430 нм на коротких задерж-
изменения комплекса BoWSCP, содержащего
ках коэффициент nexcited приближается к 0,5.
20-30% Хл b, в этой системе был зарегистри-
Как было отмечено ранее, это свидетельствует
рован перенос возбуждения от Хл b к Хл a с
о преобладании возбужденного состояния S3
характерным временем 400 фс. Данный про-
в данных условиях.
цесс был отнесен авторами к экситонной ре-
Наконец, на рис. 8, в приведено изменение
лаксации гетеродимера Chl a-Chl b, но позд-
коэффициента анизотропии, r = (ΔA|| - ΔA⊥) /
нее в работе Alster et al. [12] было показано,
(ΔA|| + 2ΔA⊥), рассчитанного в области поло-
что экситонная релаксация гетеродимера про-
сы Qy для двух вариантов возбуждения. Из-
исходит быстрее 100 фс, а процессы со време-
менение амплитуды выцветания полосы Qy
нем 250-400 фс относятся к переносу энергии
на временах ~100 фс практически не связано
между димерами. В работе Fresch et al. [13]
с изменением анизотропии.
методом 2D-спектроскопии изучалась абсорб-
ционная динамика гомотетрамеров LvWSCP и
BoWSCP, содержащих 100% Хл a. В обеих си-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
стемах в субпикосекундном диапазоне наблю-
далась моноэкспоненциальная абсорбционная
Белки класса WSCP представляют собой
динамика со временем 70 и 100 фс соответ-
уникальную по организации модельную си-
ственно, которая была интерпретирована как
стему экситонно-сопряженных молекул хло-
релаксационный переход E1 → E2 в нижнее эк-
рофилла [11], с помощью которой возможно
ситонное состояние [13]. Расчеты в рамках мо-
исследовать механизмы переноса энергии воз-
дифицированной теории Редфилда предска-
буждения между пигментами [12-14] и взаимо-
зывают, что установление равновесия между
действие возбужденных пигментов с белковым
экситонными состояниями должно происхо-
окружением [15-17]. Обмен энергией между
дить в этом временном диапазоне [15]. Спек-
электронной подсистемой и колебательными
тральный переход со временем 110 фс, наблю-
движениями ядер приводит к эволюции элек-
даемый при возбуждении BoWSCP импульсом
тронных состояний [44, 45]. Эффективность
670 нм (рис. 6, а; спектр 2), хорошо соответ-
энергетического обмена экситона с фононны-
ствует полученным ранее данным.
ми модами зависит от спектральной плотности
В данной работе впервые проведены из-
колебательных состояний ядер, для экспери-
мерения абсорбционной динамики тетрамер-
ментального определения которой использу-
ного комплекса Хл a в широком спектральном
ют методы выжигания спектрального прова-
диапазоне
400-750 нм с временным разре-
ла [46-48] и нестационарной флуоресцентной
шением 20 фс, что позволило в одной экспе-
спектроскопии [10]. Вибронные полосы Хл а
риментальной системе зарегистрировать про-
включают более 40 мод в диапазоне от 1500
цессы внутримолекулярной конверсии Хл a
до 260 см-1 [49-51]. Величина электронного
из возбужденного состояния S3 в состояние S1,
взаимодействия ближайших молекул Хл a в
проанализировать релаксационную динамику
тетрамере BoWSCP, полученная совместным
экситонно-сопряженных молекул Хл a и ко-
разложением спектров поглощения и кругового
личественно характеризовать распределение
БИОХИМИЯ том 88 вып. 10 2023
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ТЕТРАМЕРА ХЛОРОФИЛЛА
1921
экситонных состояний E1 и E2, относящихся к
взаимодействие мономеров в BoWSCP, воз-
возбужденному состоянию S1 димера Хл a.
никающее при существенном перекрывании
Количественное сопоставление абсорбци-
волновых функций возбужденных состояний.
онной динамики в областях Соре и полосы Qy
В подобных димерах возникают резонансные
требуется для интерпретации данных, полу-
состояния с переносом заряда, поэтому для
ченных при изучении больших фотосинте-
них характерна высокая энергия электрон-
тических комплексов. Например, амплитуда
фононного сопряжения [61, 62].
выцветания Хл a в области полосы Qy в началь-
2. При возбуждении BoWSCP в области
ных переходных спектрах фотосистемы I циа-
полосы Соре образуется преимущественно
нобактерии Thermosynechococcus elongatus при
электронное состояние S3 (обозначаемое так
возбуждении в дальней красной области [53],
же, как Bx), которое переходит в состояние S1
а также Хл a и Хл f в аналогичных спектрах
в процессе внутримолекулярной конверсии
фотосистемы I цианобактерии Fischerella ther-
с характерным временем 83 ± 9 фс. Однако
malis PCC 7521 [54] в 3-6 раз превышает ам-
стимулированное излучение в этом состоя-
плитуду выцветания полосы Соре, тогда как в
нии характеризуется примерно вдвое мень-
аналогичных спектрах фотосистемы I циано-
шей амплитудой в сравнении с равновесным
бактерии Synechocystis sp. PCC 6803 амплитуда
состоянием, образующимся после возбужде-
выцветания полосы Qy лишь незначитель-
ния на 670 нм. Уменьшение амплитуды сти-
но превышает амплитуду полосы Соре [55].
мулированного излучения может возникать в
Van Stokkum et al. [56] предположили, что за-
результате сверхбыстрой фотохимической ре-
вышенная амплитуда полосы Qy в началь-
акции окисления Хл и восстановления одного
ных спектрах фотосистемы I свидетельствует
из остатков триптофана, которая конкурирует
о тушении большей части возбужденных со-
с реакцией внутримолекулярной конверсии.
стояний окисленным хлорофиллом специаль-
В этом случае обратная реакция рекомбина-
ной пары. Однако фотособирающие антен-
ции ион-радикальной пары Chl+Trp- может
ные комплексы фотосистемы I цианобактерий
происходить медленнее 200 пс. Реакция фото-
T. elongatus и F. thermalis PCC 7521, в отличие от
восстановления триптофана Chl* → Chl+Trp-
антенны Synechocystis sp. PCC 6803, содержат
была зарегистрирована при исследовании
экситонно-сопряженные димеры и тримеры
LvWSCP методами спектроскопии выжига-
Хл с максимумом поглощения в дальней крас-
ния провалов при температуре 4 К в часовом
ной области [57-60]. Результаты, полученные
временном интервале с малым квантовым
в данной работе о влиянии экситонного со-
выходом [47]. Можно предположить, что спек-
пряжения на амплитуду выцветания полосы Qy
тральные изменения с характерным временем
(рис. 8), позволяют объяснить наблюдаемые
τ2 = 16 ± 3 пс (рис. 6, б) могут возникать в ре-
эффекты структурными особенностями фото-
зультате ускорения данной фотохимической
собирающих антенных комплексов.
реакции при температуре 279 К.
Из представленных на рис. 8 зависимо-
3. Увеличение амплитуды выцветания по-
стей амплитуды выцветания полосы Соре и
лосы Qy относительно полосы Соре наблю-
полосы Qy, а также фактора nexcited, представ-
далось в переходных спектрах фотосистемы I
ляющего собой оценку количества экситонно-
различных цианобактерий при специфическом
сопряженных молекул Хл, следует несколько
возбуждении длинноволновых форм хлоро-
выводов о физико-химических свойствах воз-
филла [53, 54, 63]. Возникновение длинновол-
бужденных состояний BoWSCP.
новых форм хлорофилла в светособирающей
1. При возбуждении BoWSCP в области Qy
фотосинтетической антенне и реакционном
образуется преимущественно верхнее экситон-
центре фотосистемы I связано с образованием
ное состояние E1 с максимумом около 672 нм.
димерных и тримерных экситонно-сопряжен-
Тепловое равновесие между состоянием E1 и
ных комплексов [60, 64, 65], что имеет очевид-
более низким по энергии возбуждения состоя-
ную структурную аналогию с организацией
нием E2 (максимум около 684 нм) устанавлива-
молекул хлорофилла в комплексе WSCP.
ется с характерным временем τ1 = 105 ± 10 фс;
однако состояние E1 остается преобладающим
Вклад авторов. Д.А. Черепанов - анализ
на временах сотен пикосекунд (его относи-
результатов спектральных измерений; К.В. Не-
тельный вклад уменьшается до ~80%). Ин-
веров, Ю.Н. Обухов, Ю.В. Малеева - произ-
версию заселенности можно объяснить су-
водство, выделение и биохимическая харак-
щественным вкладом энтропии в разницу
теристика препаратов BoWSCP; Ф.Е. Гостев,
свободных энергий состояний E1 и E2. Это, в
И.В. Шелаев, А.В. Айбуш - разработка мате-
свою очередь, означает сильное электронное
матического аппарата, методики и проведение
14
БИОХИМИЯ том 88 вып. 10 2023
1922
ЧЕРЕПАНОВ и др.
фемтосекундных измерений; М.С. Крицкий,
природных материалов: масс-спектральная
В.А. Надточенко - концепция работы. Все ав-
микроскопия и фемтосекундная лазерная
торы внесли значительный вклад в написание
микроскопия-спектроскопия» (рег. номер:
рукописи.
506694). Экспрессионный вектор pET-24b со
Финансирование. Работа выполнена при
встроенным геном белка BoWSCP был лю-
поддержке Российского научного фонда (грант
безно предоставлен проф. Харальдом Пауль-
№ 21-74-20155).
сеном (Институт молекулярной физиологии
Благодарности. Авторы благодарят А.Л. До-
Университета Иоганна Гутенберга, Майнц,
брякова за помощь в разработке метода аппро-
Германия).
ксимации когерентного артефакта и установ-
Конфликт интересов. Авторы заявляют об
ления аппаратной функции фемтосекундной
отсутствии конфликта интересов.
установки. В работе использовано оборудова-
Соблюдение этических норм. Настоящая
ние (фемтосекундная установка) Центра кол-
статья не содержит описания каких-либо ис-
лективного пользования ФИЦ ХФ РАН «Ана-
следований с участием людей или животных
лиз химических, биологических систем и
в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Charlton, A., and Zachariou, M. (2007) Immobilized
7.
Satoh, H., Uchida, A., Nakayama, K., and Okada, M.
metal ion affinity chromatography of histidine-tagged
(2001) Water-soluble chlorophyll protein in Brassi-
fusion proteins, Methods Mol. Biol., 421, 137-149,
caceae plants is a stress-induced chlorophyll-binding
doi: 10.1007/978-1-59745-582-4_10.
protein, Plant Cell Physiol., 42, 906-911, doi: 10.1093/
2.
Cherepanov, D. A., Semenov, A. Y., Mamedov,
pcp/pce117.
M. D., Aybush, A. V., Gostev, F. E., Shelaev, I. V.,
8.
Horigome, D., Satoh, H., Itoh, N., Mitsunaga, K.,
Shuvalov, V. A., and Nadtochenko, V. A.
(2022)
Oonishi, I., Nakagawa, A., and Uchida, A. (2007)
Current state of the primary charge separation
Structural mechanism and photoprotective function
mechanism in photosystem I of cyanobacteria,
of water-soluble chlorophyll-binding protein, J. Biol.
Biophys. Rev.,
14,
805-820, doi:
10.1007/s12551-
Chem., 282, 6525-6531, doi: 10.1074/jbc.M609458200.
022-00983-1.
9.
Малеева Ю. В., Неверов К. В., Обухов Ю. Н.,
3.
Murata, T., Toda, F., Uchino, K., and Yakushiji, E.
Крицкий М. С. (2019) Водорастворимые хлоро-
(1971) Water-soluble chlorophyll protein of Bras-
филлсвязывающие белки растений: структура,
sica oleracea var. botrys (cauliflower), Biochim. Bio-
свойства и функции, Мол. Биол., 53, 998-1011,
phys. Acta Bioenerg., 245, 208-215, doi: 10.1016/
doi: 10.1134/S0026898419060120.
0005-2728(71)90023-5.
10.
Schmitt, F. J., Trostmann, I., Theíss, C., Pieper, J.,
4.
Satoh, H., Nakayama, K., and Okada, M. (1998)
Renger, T., Fuesers, J., Hubrich, E. H., Paulsen, H.,
Molecular cloning and functional expression of a
Eichler, H. J., and Renger, G. (2008) Excited state
water-soluble chlorophyll protein, a putative carrier of
dynamics in recombinant water-soluble chlorophyll
chlorophyll molecules in cauliflower, J. Biol. Chem.,
proteins (WSCP) from cauliflower investigated by
273, 30568-30575, doi: 10.1074/jbc.273.46.30568.
transient fluorescence spectroscopy, J. Phys. Chem. B,
5.
Takahashi, S., Yanai, H., Nakamaru, Y., Uchi-
112, 13951-13961, doi: 10.1021/jp8024057.
da, A., Nakayama, K., and Satoh, H. (2012) Mo-
11.
Renger, G., Pieper, J., Theiss, C., Trostmann, I.,
lecular cloning, characterization and analysis of the
Paulsen, H., Renger, T., Eichler, H. J., and Schmitt, F. J.
intracellular localization of a water-soluble chl-bind-
(2011) Water soluble chlorophyll binding protein of high-
ing protein from brussels sprouts (Brassica oleracea
er plants: A most suitable model system for basic anal-
var. gemmifera), Plant Cell Physiol., 53, 879-891,
yses of pigment-pigment and pigment-protein interac-
doi: 10.1093/pcp/pcs031.
tions in chlorophyll protein complexes, J. Plant Physiol.,
6.
Takahashi, S., Yanai, H., Oka-Takayama, Y., Zanma-
168, 1462-1472, doi: 10.1016/j.jplph.2010.12.005.
Sohtome, A., Fujiyama, K., Uchida, A., Nakayama, K.,
12.
Alster, J., Lokstein, H., Dostál, J., Uchida, A., and
and Satoh, H. (2013) Molecular cloning, characteri-
Zigmantas, D. (2014) 2D spectroscopy study of water-
zation and analysis of the intracellular localization of
soluble chlorophyll-binding protein from lepidium
a water-soluble chlorophyll-binding protein (WSCP)
virginicum, J. Phys. Chem. B,
118,
3524-3531,
from Virginia pepperweed (Lepidium virginicum),
doi: 10.1021/jp411174t.
a unique WSCP that preferentially binds chloro-
13.
Fresch, E., Meneghin, E., Agostini, A., Paulsen, H.,
phyll b in vitro, Planta, 238, 1065-1080, doi: 10.1007/
Carbonera, D., and Collini, E. (2020) How the protein
s00425-013-1952-7.
environment can tune the energy, the coupling, and
БИОХИМИЯ том 88 вып. 10 2023
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ТЕТРАМЕРА ХЛОРОФИЛЛА
1923
the ultrafast dynamics of interacting chlorophylls:
24.
Bradford, M. M. (1976) A rapid and sensitive meth-
the example of the water-soluble chlorophyll protein,
od for the quantitation of microgram quantities of
J. Phys. Chem. Lett., 11, 1059-1067, doi: 10.1021/
protein utilizing the principle of protein-dye bind-
acs.jpclett.9b03628.
ing, Anal. Biochem.,
72,
248-254, doi:
10.1016/
14.
Theiss, C., Trostmann, I., Andree, S., Schmitt, F. J.,
0003-2697(76)90527-3.
Renger, T., Eichler, H. J., Paulsen, H., and Renger, G.
25.
Cherepanov, D. A., Shelaev, I. V., Gostev, F. E.,
(2007) Pigment-pigment and pigment-protein inter-
Mamedov, M. D., Petrova, A. A., Aybush, A. V.,
actions in recombinant water-soluble chlorophyll pro-
Shuvalov, V. A., Semenov, A. Y., and Nadtochenko,
teins (WSCP) from cauliflower, J. Phys. Chem. B, 111,
V. A. (2017) Mechanism of adiabatic primary electron
13325-13335, doi: 10.1021/jp0723968.
transfer in photosystem I: Femtosecond spectroscopy
15.
Renger, T., Trostmann, I., Theiss, C., Madjet, M. E.,
upon excitation of reaction center in the far-red edge
Richter, M., Paulsen, H., Eichler, H. J., Knorr, A.,
of the Q Y band, Biochim. Biophys. Acta Bioenerg.,
and Renger, G. (2007) Refinement of a structur-
1858, 895-905, doi: 10.1016/j.bbabio.2017.08.008.
al model of a pigment-protein complex by accurate
26.
Dobryakov, A. L., Pérez Lustres, J. L., Kovalenko, S. A.,
optical line shape theory and experiments, J. Phys.
and Ernsting, N. P. (2008) Femtosecond transient ab-
Chem. B, 111, 10487-10501, doi: 10.1021/jp0717241.
sorption with chirped pump and supercontinuum probe:
16.
Friedl, C., Fedorov, D. G., and Renger, T. (2022)
Perturbative calculation of transient spectra with general
Towards a quantitative description of excitonic cou-
lineshape functions, and simplifications, Chem. Phys.,
plings in photosynthetic pigment-protein complexes:
347, 127-138, doi: 10.1016/j.chemphys.2007.11.003.
Quantum chemistry driven multiscale approaches,
27.
Golubeva, E. N., Zubanova, E. M., Melnikov, M. Y.,
Phys. Chem. Chem. Phys., 24, 5014-5038, doi: 10.1039/
Gostev, F. E., Shelaev, I. V., and Nadtochenko, V. A.
d1cp03566e.
(2014) Femtosecond spectroscopy and TD-DFT
17.
Lahav, Y., Noy, D., and Schapiro, I. (2021) Spectral
calculations of CuCl42- excited states, Dalt. Trans.,
tuning of chlorophylls in proteins - electrostatics
43, 17820-17827, doi: 10.1039/C4DT01409J.
vs. ring deformation, Phys. Chem. Chem. Phys., 23,
28.
Dobryakov, A. L., Kovalenko, S. A., Weigel, A., Ṕrez-
6544-6551, doi: 10.1039/d0cp06582j.
Lustres, J. L., Lange, J., Müller, A., and Ernsting,
18.
Agostini, A., Meneghin, E., Gewehr, L., Pedron, D.,
N. P. (2010) Femtosecond pump/supercontinuum-
Palm, D. M., Carbonera, D., Paulsen, H.,
probe spectroscopy: Optimized setup and signal
Jaenicke, E., and Collini, E. (2019) How water-
analysis for single-shot spectral referencing, Rev. Sci.
mediated hydrogen bonds affect chlorophyll a/b
Instrum., 81, 113106, doi: 10.1063/1.3492897.
selectivity in water-soluble chlorophyll protein, Sci.
29.
Kovalenko, S. A., Dobryakov, A. L., Ruthmann, J.,
Rep., 9, 1-10, doi: 10.1038/s41598-019-54520-54524.
and Ernsting, N. P. (1999) Femtosecond spectroscopy
19.
Bednarczyk, D., Dym, O., Prabahar, V., Peleg, Y.,
of condensed phases with chirped supercontinuum
Pike, D. H., and Noy, D. (2016) Fine tuning of chlo-
probing, Phys. Rev. A At. Mol. Opt. Phys., 59, 2369-
rophyll spectra by protein-induced ring deformation,
2384, doi: 10.1103/PhysRevA.59.2369.
Angew. Chemie Int. Ed., 55, 6901-6905, doi: 10.1002/
30.
Šimůnek, J., and Hopmans, J. W. (2002) 1.7 parameter
anie.201512001.
optimization and nonlinear fitting, Methods Soil
20.
Hughes, J. L., Razeghifard, R., Logue, M., Oakley, A.,
Anal. Part 4 Phys. Methods, 5, 139-157, doi: 10.2136/
Wydrzynski, T., and Krausz, E. (2006) Magneto-
sssabookser5.4.c7.
optic spectroscopy of a protein tetramer binding two
31.
Clementson, L. A., and Wojtasiewicz, B.
(2019)
exciton-coupled chlorophylls, J. Am. Chem. Soc., 128,
Dataset on the absorption characteristics of extract-
3649-3658, doi: 10.1021/ja056576b.
ed phytoplankton pigments, Data Br., 24, 103875,
21.
Обухов Ю. Н., Неверов К. В., Малеева Ю. В.,
doi: 10.1016/j.dib.2019.103875.
Крицкий М. С. (2023) Димеры хлорофилла a в
32.
Sirohiwal, A., Berraud-Pache, R., Neese, F., Izsák, R.,
составе водорастворимого белка BoWSCP фото-
and Pantazis, D. A. (2020) Accurate computation of the
сенсибилизируют восстановление цитохрома c,
absorption spectrum of chlorophyll a with pair natural
Докл. Акад. Наук,
509,
191-195, doi:
10.31857/
orbital coupled cluster methods, J. Phys. Chem. B,
S2686738922600790.
124, 8761-8771, doi: 10.1021/acs.jpcb.0c05761.
22.
Takahashi, S., Uchida, A., Nakayama, K., and
33.
Gouterman, M.
(1961) Spectra of porphyrins,
Satoh, H. (2014) Three-step photoconversion of only
J. Mol. Spectrosc., 6, 138-163, doi: 10.1016/0022-2852
three subunits of the water-soluble chlorophyll-binding
(61)90236-3.
protein tetramer from Chenopodium album, Protein J.,
34.
Bricker, W. P., Shenai, P. M., Ghosh, A., Liu, Z.,
33, 337-343, doi: 10.1007/s10930-014-9565-y.
Enriquez, M. G. M., Lambrev, P. H., Tan, H. S., Lo,
23.
Kelly, S. M., Jess, T. J., and Price, N. C. (2005) How to
C. S., Tretiak, S., Fernandez-Alberti, S., and Zhao, Y.
study proteins by circular dichroism, Biochim. Biophys.
(2015) Non-radiative relaxation of photoexcited
Acta Proteins Proteomics, 1751, 119-139, doi: 10.1016/
chlorophylls: theoretical and experimental study, Sci.
j.bbapap.2005.06.005.
Rep., 5, 1-16, doi: 10.1038/srep13625.
БИОХИМИЯ том 88 вып. 10 2023
14*
1924
ЧЕРЕПАНОВ и др.
35.
Götze, J. P., Anders, F., Petry, S., Witte, J. F., and
ture and pigment-protein interactions in the recom-
Lokstein, H. (2022) Spectral characterization of the
binant water-soluble chlorophyll protein. II. Spectral
main pigments in the plant photosynthetic apparatus
hole-burning experiments, J. Phys. Chem. B, 115,
by theory and experiment, Chem. Phys., 559, 111517,
4053-4065, doi: 10.1021/jp111457t.
doi: 10.1016/j.chemphys.2022.111517.
47.
Kell, A., Bednarczyk, D., Acharya, K., Chen, J.,
36.
Umetsu, M., Wang, Z. Y., Kobayashi, M., and
Noy, D., and Jankowiak, R. (2016) New insight into
Nozawa, T. (1999) Interaction of photosynthetic pig-
the water-soluble chlorophyll-binding protein from
ments with various organic solvents. Magnetic cir-
Lepidium virginicum, Photochem. Photobiol.,
92,
cular dichroism approach and application to chloro-
428-435, doi: 10.1111/php.12581.
somes, Biochim. Biophys. Acta Bioenerg., 1410, 19-31,
48.
Adolphs, J., Berrer, M., and Renger, T.
(2016)
doi: 10.1016/S0005-2728(98)00170-4.
Hole-burning spectroscopy on excitonically cou-
37.
Shipman, L. L., Cotton, T. M., Norris, J. R., and
pled pigments in proteins: theory meets experiment,
Katz, J. J. (1976) An analysis of the visible absorption
J. Am. Chem. Soc., 138, 2993-3001, doi: 10.1021/
spectrum of chlorophyll a monomer, dimer,and
jacs.5b08246.
oligomers in solution, J. Am. Chem. Soc., 98, 8222-
49.
Gillie, J. K., Lyle, P. A., Small, G. J., and Golbeck,
8230, doi: 10.1021/ja00441a056.
J. H. (1989) Spectral hole burning of the primary
38.
Черепанов Д. А., Милановский Г. Е., Айбуш А. В.,
electron donor state of Photosystem I, Photosynth.
Надточенко В. А. (2023) Дипольный момент пере-
Res., 22, 233-246, doi: 10.1007/BF00048302.
хода S0→S1 хлорофилла a в растворителях с раз-
50.
Pieper, J., Voigt, J., and Small, G. J. (1999) Chloro-
личным индексом рефракции, Хим. Физ.,
42,
phyll a Franck-Condon factors and excitation en-
77-78, doi: 10.31857/S0207401X23060031.
ergy transfer, J. Phys. Chem. B, 103, 2321-2322,
39.
Berera, R., Van Grondelle, R., and Kennis, J. T. M.
doi: 10.1021/jp984460e.
(2009) Ultrafast transient absorption spectroscopy:
51.
Peternian, E. J. G., Pullerits, T., van Grondelle, R.,
principles and application to photosynthetic systems,
and van Amerongen, H. (1997) Electron-phonon cou-
Photosynth. Res., 101, 105-118, doi: 10.1007/s11120-
pling and vibronic fine structure of light-harvesting
009-9454-y.
complex II of green plants: temperature dependent
40.
Черепанов Д. А., Гостев Ф. Е., Шелаев И. В.,
absorption and high-resolution fluorescence spectros-
Айбуш А. В., Семенов А. Ю., Мамедов М. Д.,
copy, J. Phys. Chem. B, 101, 4448-4457, doi: 10.1021/
Шувалов В. А., Надточенко В. А. (2020) Абсорб-
jp962338e.
ционный спектр возбужденного синглетного
52.
Förster, T. (1948) Intermolecular energy migration
состояния хлорофилла a в видимом и ближнем
and fluorescence [in German], Ann. Phys., 437, 55-75,
инфракрасном диапазонах, Хим. Выс. Энерг., 54,
doi: 10.1002/andp.19484370105.
158-160, doi: 10.31857/S0023119320020059.
53.
Cherepanov, D. A., Shelaev, I. V., Gostev, F. E.,
41.
Cherepanov, D. A., Petrova, A. A., Mamedov,
Aybush, A. V., Mamedov, M. D., Shuvalov, V. A.,
M. D., Vishnevskaya, A. I., Gostev, F. E., Shelaev,
Semenov, A. Y., and Nadtochenko, V. A.
(2020)
I. V., Aybush, A. V., and Nadtochenko, V. A. (2022)
Generation of ion-radical chlorophyll states in the
Comparative absorption dynamics of the singlet
light-harvesting antenna and the reaction center of
excited states of chlorophylls a and d, Biochem.,
cyanobacterial photosystem I, Photosynth. Res., 146,
87, 1179-1186, doi: 10.1134/S000629792210011X.
55-73, doi: 10.1007/s11120-020-00731-0.
42.
Davydov, A. S. (1962) The Theory Of Molecular
54.
Cherepanov, D. A., Shelaev, I. V., Gostev, F. E., Aybush,
Excitons, McGraw-Hill, New York, NY.
A. V., Mamedov, M. D., Shen, G., Nadtochenko, V. A.,
43.
Kasha, M., Rawls, H. R., and El-Bayoumi, M. A. (1965)
Bryant, D. A., Semenov, A. Y., and Golbeck, J. H.
The exciton model in molecular spectroscopy, Pure Appl.
(2020) Evidence that chlorophyll f functions solely as
Chem., 11, 371-392, doi: 10.1351/pac196511030371.
an antenna pigment in far-red-light photosystem I from
44.
Scholes, G. D., and Ghiggino, K. P. (1994) Rate
Fischerella thermalis PCC 7521, Biochim. Biophys. Acta
expressions for excitation transfer I. Radiationless
Bioenerg., 1861 doi: 10.1016/j.bbabio.2020.148184.
transition theory perspective, J. Chem. Phys., 101,
55.
Shelaev, I. V., Gostev, F. E., Mamedov, M. D.,
1251-1261, doi: 10.1063/1.467817.
Sarkisov, O. M., Nadtochenko, V. A., Shuvalov, V. A.,
45.
Renger, T., and Marcus, R. A. (2002) On the rela-
and Semenov, A. Y. (2010) Femtosecond primary
tion of protein dynamics and exciton relaxation in
charge separation in Synechocystis sp. PCC
6803
pigment-protein complexes: an estimation of the
photosystem I, Biochim. Biophys. Acta Bioenerg., 1797,
spectral density and a theory for the calculation of
1410-1420, doi: 10.1016/j.bbabio.2010.02.026.
optical spectra, J. Chem. Phys., 116, 9997-10019,
56.
Van Stokkum, I. H. M., Müller, M. G., Weißenborn, J.,
doi: 10.1063/1.1470200.
Weigand, S., Snellenburg, J. J., and Holzwarth, A. R.
46.
Pieper, J., Rätsep, M., Trostmann, I., Schmitt, F. J.,
(2023) Energy transfer and trapping in photosystem I
Theiss, C., Paulsen, H., Eichler, H. J., Freiberg, A.,
with and without chlorophyll-f, IScience, 26, 107650,
and Renger, G. (2011) Excitonic energy level struc-
doi: 10.1016/J.ISCI.2023.107650.
БИОХИМИЯ том 88 вып. 10 2023
ФЕМТОСЕКУНДНАЯ ДИНАМИКА ТЕТРАМЕРА ХЛОРОФИЛЛА
1925
57. Pålsson, L.-O. O., Flemming, C., Gobets, B., van
61. Young, R. M., and Wasielewski, M. R. (2020) Mixed
Grondelle, R., Dekker, J. P., and Schlodder, E. (1998)
electronic states in molecular dimers: Connecting
Energy transfer and charge separation in photosys-
singlet fission, excimer formation, and symmetry-
tem I: P700 oxidation upon selective excitation of the
breaking charge transfer, Acc. Chem. Res.,
53,
long-wavelength antenna chlorophylls of Synechococ-
1957-1968, doi: 10.1021/acs.accounts.0c00397.
cus elongatus, Biophys. J., 74, 2611-2622, doi: 10.1016/
62. Casanova, D. (2018) Theoretical modeling of singlet
S0006-3495(98)77967-6.
fission, Chem. Rev., 118, 7164-7207, doi: 10.1021/
58. Gobets,B.,Kennis, J.T.M., Ihalainen, J.A.,Brazzoli,M.,
acs.chemrev.7b00601.
Croce, R., Stokkum, I. H. M. van, Bassi, R., Dekker,
63. Petrova, A. A., Casazza, A. P., Shelaev, I. V., Gostev,
J. P., van Amerongen, H., Fleming, G. R., and van
F. E., Aybush, A. V., Nadtochenko, V. A., Semenov,
Grondelle, R. (2001) Excitation energy transfer in
A. Y., Santabarbara, S., and Cherepanov, D. A. (2023)
dimeric light harvesting complex I: a combined streak-
Role of pheophytin a in the primary charge separation
camera/fluorescence upconversion study, J. Phys.
of photosystem I from Acaryochloris marina: femtosec-
Chem. B, 105, 10132-10139, doi: 10.1021/jp011901c.
ond optical studies of excitation energy and electron
59. Riley, K. J., Reinot, T., Jankowiak, R., Fromme, P.,
transfer reactions, Biochim. Biophys. Acta Bioenerg.,
and Zazubovich, V. (2007) Red antenna states of pho-
1864, 148984, doi: 10.1016/j.bbabio.2023.148984.
tosystem I from cyanobacteria Synechocystis PCC 6803
64. Akhtar, P., and Lambrev, P. H. (2020) On the spectral
and Thermosynechococcus elongatus: single-complex
properties and excitation dynamics of long-wavelength
spectroscopy and spectral hole-burning study, J. Phys.
chlorophylls in higher-plant photosystem I, Biochim.
Chem. B, 111, 286-292, doi: 10.1021/jp062664m.
Biophys. Acta Bioenerg., 1861, 148274, doi: 10.1016/
60. Gisriel, C., Shen, G., Kurashov, V., Ho, M. Y.,
J.BBABIO.2020.148274.
Zhang, S., Williams, D., Golbeck, J. H., Fromme, P.,
65. Karapetyan, N. V., Bolychevtseva, Y. V., Yurina, N. P.,
and Bryant, D. A. (2020) The structure of Photosys-
Terekhova, I. V., Shubin, V. V., and Brecht, M. (2014)
tem I acclimated to far-red light illuminates an eco-
Long-wavelength chlorophylls in photosystem I of
logically important acclimation process in photo-
cyanobacteria: origin, localization, and functions,
synthesis, Sci. Adv., 6, eaay6415, doi: 10.1126/sciadv.
Biochemistry (Moscow), 79, 213-220, doi: 10.1134/
aay6415.
S0006297914030067.
PHOTOCHEMICAL REACTIONS OF CHLOROPHYLL TETRAMER
IN WATER-SOLUBLE CHLOROPHYLL-BINDING PROTEIN BoWSCP
D. A. Cherepanov1,2*, K. V. Neverov3,4, Yu. N. Obukhov3, Yu. V. Maleeva4, F. E. Gostev1,
I. V. Shelaev1,2, A. V. Aybush1, M. S. Kritsky3, and V. A. Nadtochenko1,5*
1 Semenov Federal Research Center for Chemical Physics, Russian Academy of Sciences,
119991 Moscow, Russia; e-mail: tscherepanov@gmail.com
2 Belozersky Research Institute of Physical and Chemical Biology, Lomonosov Moscow State University,
119992 Moscow, Russia
3 Bach Institute of Biochemistry, Federal Research Centre
“Fundamentals of Biotechnology” of the Russian Academy of Sciences, 119071 Moscow, Russia
4 Faculty of Biology, Lomonosov Moscow State University, 119991 Moscow, Russia
5 Faculty of Chemistry, Lomonosov Moscow State University,
119991 Moscow, Russia; e-mail: nadtochenko@gmail.com
The absorption dynamics of chlorophyll a in a symmetric tetrameric complex of the water-soluble chloro-
phyll-binding protein BoWSCP was measured by the broadband femtosecond laser pump-probe spectros-
copy within the range from 400 to 780 nm with a time resolution of 20 f-200 ps. When BoWSCP was excited
in the region of the Soret band at a wavelength of 430 nm, a nonradiative intramolecular conversion S3 → S1
was observed with a characteristic time of 83 ± 9 fs. When the complex was excited in the region of the Qy
band at a wavelength of 670 nm, a relaxation transition between two excitonic states of the chlorophyll dimer
was observed with a time of 105 ± 10 fs. Absorption spectra of excited singlet states S1 and S3 of chlorophyll a
were obtained. It has been demonstrated that the delocalization of the excited state between exciton-coupled
Chl molecules in the BoWSCP tetramer changes in time and depends on the excitation energy. Upon excita-
tion of BoWSCP, an ultrafast photochemical reaction is observed in the Soret region, apparently due to the
reduction of tryptophan in the vicinity of chlorophyll.
Keywords: chlorophyll a, proteins from the WSCP family, femtosecond laser spectroscopy, excited state spectrum,
exciton dynamics, intramolecular conversion
БИОХИМИЯ том 88 вып. 10 2023