БИОХИМИЯ, 2023, том 88, вып. 10, с. 1926 - 1939
УДК 577.352.53;581.17;577.359;576.32/.36;612.014
МИТОХОНДРИАЛЬНАЯ СЕТЬ:
ЭЛЕКТРИЧЕСКИЙ КАБЕЛЬ И МНОГОЕ ДРУГОЕ
Обзор
© 2023 П.А. Абрамичева1, Н.В. Андрианова1, В.А. Бабенко1,2, Л.Д. Зорова1,2,
С.Д. Зоров1,3, И.Б. Певзнер1,2, В.А. Попков1,2, Д.С. Семенович1, Э.И. Якупова1,
Д.Н. Силачев1,2, Е.Ю. Плотников1,2, Г.Т. Сухих2, Д.Б. Зоров1,2*
1 Московский государственный университет имени М.В. Ломоносова,
НИИ физико-химической биологии имени А.Н. Белозерского,
119991 Москва, Россия; электронная почта: zorov@belozersky.msu.ru
2 Национальный медицинский исследовательский центр акушерства, гинекологии
и перинатологии им. академика В.И. Кулакова Минздрава России, 117997 Москва, Россия
3 Московский государственный университет имени М.В. Ломоносова,
факультет биоинженерии и биоинформатики, 119991 Москва, Россия
Поступила в редакцию 12.07.2023
После доработки 20.09.2023
Принята к публикации 22.09.2023
Митохондрии в клетке могут объединяться и организовывать сложные, протяженные структуры,
которые пронизывают весь объем клетки, обеспечивая равномерное снабжение энергией в виде
синтезируемого в митохондриях АТР. В соответствии с хемиосмотической концепцией, энергия
окисления дыхательных субстратов в значительной степени запасается в виде разности электриче-
ских потенциалов на внутренней мембране митохондрий. Теория функционирования протяжен-
ных митохондриальных структур как внутриклеточных электрических проводов предполагает, что
митохондрии обеспечивают максимально быструю доставку электрической энергии по клеточному
объему с последующим использованием этой энергии для синтеза АТР, тем самым ускоряя про-
цесс доставки АТР по сравнению с достаточно медленной диффузией АТР в клетке. В аналитиче-
ском обзоре дана история кабельной теории, перечислены нерешенные критические проблемы,
описаны перестройки митохондриальной сети и роль окислительного стресса в этом процессе.
Кроме уже доказанного функционирования протяженных митохондриальных структур как элек-
трических кабелей, предлагается и ряд дополнительных функций, в частности выдвигается гипо-
теза поддержания митохондриальными сетями редокс-потенциала в клеточном объеме, который
может варьировать в зависимости от физиологического состояния в результате изменения трех-
мерной организации митохондриальной сети (фрагментация/расщепление - слияние). Рассмо-
трен ряд патологий, сопровождаемых нарушением редокс-статуса, и участие в них митохондрий.
КЛЮЧЕВЫЕ СЛОВА: митохондрии, ретикулум, сеть, электричество, мембранный потенциал, фрагмен-
тация, расщепление, кардиомиоциты, сперматозоиды, окислительный стресс, редокс, преэклампсия,
задержка роста плода.
DOI: 10.31857/S0320972523100147, EDN: OVONXX
ВВЕДЕНИЕ. ХЕМИОСМОТИЧЕСКАЯ
создает протондвижущую силу) и реализа-
КОНЦЕПЦИЯ МИТЧЕЛЛА.
цию этой силы для фосфорилирования ADP,
РЕШЕННЫЕ И НЕРЕШЕННЫЕ ВОПРОСЫ
и эта концепция стала доминирующей в био-
энергетике [1-3]. Правда, надо отметить, что
Исторически в шестидесятые годы ХХ века
все еще идут споры о точности терминологии
произошел переход от чисто химической кон-
и о принципах сопряжения. В первом слу-
цепции окислительного фосфорилирования к
чае не очень ясно, насколько термин «осмо-
хемиосмотической, предусматривающей про-
тика» имеет отношение к этой концепции,
странственное разделение и функциональное
ибо во всем протонном цикле осмотических
сопряжение процессов окисления (которое
перестроек не предполагалось. Сам автор этой
концепции (П. Митчелл) трактовал «осмоти-
* Адресат для корреспонденции.
ку» в терминах существования топологически
1926
МИТОХОНДРИАЛЬНАЯ СЕТЬ
1927
замкнутых мембран, названных сопрягающи-
Митчелла хеми-электрической, и только с
ми мембранами, которые представляют собой
учетом недавнего открытия калиевой энерге-
осмотический барьер для веществ вообще и
тики
- электро-хемиосмотической. Однако
для протонов в частности и в пределах кото-
великая ценность открытия Митчелла, как ос-
рых существуют системы транспорта протона,
новополагающей теории, позволяющей при-
осуществляющие осмотическую стабилизацию
писать митохондриям функцию клеточных
и обеспечивающие транспорт метаболитов [4].
электростанций, не подлежит сомнению.
Недавнее обнаружение цикла транспорта иона
К+ в митохондриальной мембране, также обес-
печивающего синтез АТР [5-9], в большей
ВОЗМОЖНОСТЬ ТРАНСПОРТА
мере соответствует названию хемиосмотиче-
ЭЛЕКТРИЧЕСТВА ПО БИОЛОГИЧЕСКИМ
ской концепции по причине того, что транс-
МЕМБРАНАМ. СТРУКТУРНЫЕ ОСНОВЫ
порт ионов калия ввиду его высоких сольвати-
рующих свойств (в отличии от протона) может
В 1971 году В.П. Скулачев в одной из своих
существенным образом менять осмотические
фундаментальных работ [17] постулировал воз-
свойства сред по обе стороны сопрягающей
можность внутриклеточного транспорта энер-
мембраны, что неминуемо будет сопрово-
гии в виде мембранного потенциала. Он писал:
ждаться не всегда скомпенсированным транс-
«Система внутриклеточных (и митохондриаль-
портом воды из одного в другой объем.
ных) мембран и крист может препятствовать
Второй, не всеми признанной, частью
передаче энергии в виде таких гидрофильных
концепции Митчелла был постулат, что со-
энергоносителей, как ATP. Проблема была бы
пряжение окисления и фосфорилирования
упрощена, если бы энергию можно было пере-
происходит за счет первичного переноса
давать вдоль мембраны в форме электрическо-
протона из одной водной фазы, омывающей
го поля. Таким образом, было бы возможно,
сопрягающую мембрану, в водную фазу по
например, объединить в общую систему ты-
другую сторону мембраны с последующим
сячи отдельных ферментативных комплексов,
переносом протона из второй фазы в исход-
вырабатывающих энергию, которые располо-
ную, осуществляя при этом химический син-
жены в пространственно удаленных участках
тез АТР в АТР-синтазе, локализованной в
митохондриальной мембраны».
мембране [10-13]. Эта часть включала концеп-
Одним из самых сильных подтверждений
цию делокализованного (обводненного) про-
догадки Скулачева было обнаружение митохон-
тона в отличие от концепции сопряжения за
дриальной сети в поперечнополосатой мышце
счет локализованного протона, то есть так же
диафрагмы [18] и результаты исследований ее
транспортируемого через мембрану без выхода
развития в онтогенезе [19]. Это была фундамен-
его в водную фазу [14, 15]. При наличии этих
тальная работа, которая сломала имеющуюся
двух элементов хемиосмотической концепции,
на то время парадигму традиционного дву-
которые могут быть подвергнуты некоторой
мерного мышления, основанную на анализе
критике, есть один момент, который остается
одиночных электронно-микроскопических сре-
незыблемым и основополагающим в этой кон-
зов. В работе использовался метод реконструк-
цепции - это постулирование и доказатель-
ции внутриклеточного объема из серии после-
ство генерации электрического потенциала в
довательных тканевых срезов, позволяющий
сопрягающих мембранах [16]. Из известного
установить трехмерную организацию митохон-
набора сопрягающих мембран, включающего
дрий. Обнаруженная сложная структура, обра-
в себя клеточные мембраны некоторых аэроб-
зованная ими, была названа митохондриаль-
ных бактерий, мембраны хроматофоров фото-
ным ретикулумом. Таким образом, эти работы
синтезирующих бактерий, тилакоидные мем-
явились в некоторой степени структурной ос-
браны хлоропластов и внутренние мембраны
новой для подтверждения гипотезы функцио-
митохондрий, в этом обзоре будут обсуждаться
нирования митохондрий как электрического
лишь сопрягающие мембраны митохондрий.
кабеля.
В последних в результате работы протонных
помп комплексов I, III и IV достигается асим-
метричное распределение протонов по обе
ВИЗУАЛИЗАЦИЯ МИТОХОНДРИЙ
стороны внутренней мембраны с результиру-
ПРОНИКАЮЩИМИ
ющим повышением их концентрации в меж-
ФЛУОРЕСЦЕНТНЫМИ АГЕНТАМИ
мембранном пространстве, что соответствует
цитозольной стороне. В итоге более правиль-
Немногим позже был внедрен в практику
ным было бы считать первичную концепцию
метод визуализации митохондрий в клетке при
БИОХИМИЯ том 88 вып. 10 2023
1928
АБРАМИЧЕВА и др.
помощи избирательного накопления положи-
тие флуоресцентной микроскопии позволили
тельно заряженного флуоресцентного зонда
быстро распространить этот подход для опреде-
родамина 123, который представляет собой ме-
ления структуры хондриома в разных клетках.
тиловый эфир незамещенного родамина [20].
Это открытие также надо признать как рево-
люционное, потому что до этого митохондрии
ЛАБИЛЬНОСТЬ
в клетке визуализировали почти исключи-
МИТОХОНДРИАЛЬНОГО РЕТИКУЛУМА
тельно при помощи просвечивающей свето-
вой микроскопии [21]. Самым ранним, хотя и
Через пять лет после введения в практику
не очень чувствительным, способом визуали-
родаминовых красителей для визуализации
зации митохондрий в клетке был очень редко
митохондрий впервые была продемонстриро-
используемый метод окрашивания Янусом Зе-
вана лабильность организации митохондриаль-
леным B [22], основанный на восстановлении
ного ретикулума. Первым агентом, вызываю-
красителя за счет процессов деятельности мито-
щим быстрое превращение митохондриально-
хондрий. Для визуализации митохондрий необ-
го филамента в серию из округлых фрагментов
ходимо было использовать краситель в высоких
(явление было тогда названо митохондриаль-
концентрациях, поскольку детекция основы-
ной фрагментацией, которое соответствует бо-
валась на изменении оптического поглоще-
лее позднему названию митохондриального
ния красителя. Использование флуоресцент-
расщепления) был бензодиазепин диазепам
ных красителей, несущих в молекуле делока-
(который был митохондриально токсичен),
лизованный положительный заряд, позволяло
потенциально вызывающий окислительный
накопить краситель в матриксе митохондрий
стресс [24]. Позднее в разряд инициаторов ми-
в многотысячном превышении его концен-
тохондриальной фрагментации было помеще-
трации по сравнению с цитоплазмой клетки.
но множество классических митохондриальных
В результате митохондрии могли быть визуали-
ядов (ротенон, антимицин А, цианид, олиго-
зированы в клетке при использовании низ-
мицин, дициклогексилкарбоксидиамид и пр.
ких концентраций красителя, при этом изби-
[25, 26]). Стала ясна природа высокой ла-
рательность окрашивания была очень высокой.
бильности митохондриальных филаментов,
Почти одновременно с родамином 123 для окрас-
находящейся в прямой зависимости от окис-
ки структур, несущих мембранный потенциал,
лительного стресса [27-30]. Пример митохон-
являющийся движущей силой накопления флуо-
дриальной фрагментации, вызванной ротено-
ресцентного красителя в митохондриях, был
ном, ингибитором комплекса I митохондрий,
использован этиловый эфир незамещенного
приведен на рис. 1.
родамина [23]. Простота процедуры избира-
Явление фрагментации митохондрий (их
тельной окраски митохондрий в клетке и разви-
расщепление) стало предметом исследования
Рис. 1. Фрагментация митохондрий в клеточной культуре почек эмбриона свиньи. а - Контрольные клетки; б - после
обработки ротеноном (1 мкМ, 24 ч). Масштабная линейка - 5 мкм
БИОХИМИЯ том 88 вып. 10 2023
МИТОХОНДРИАЛЬНАЯ СЕТЬ
1929
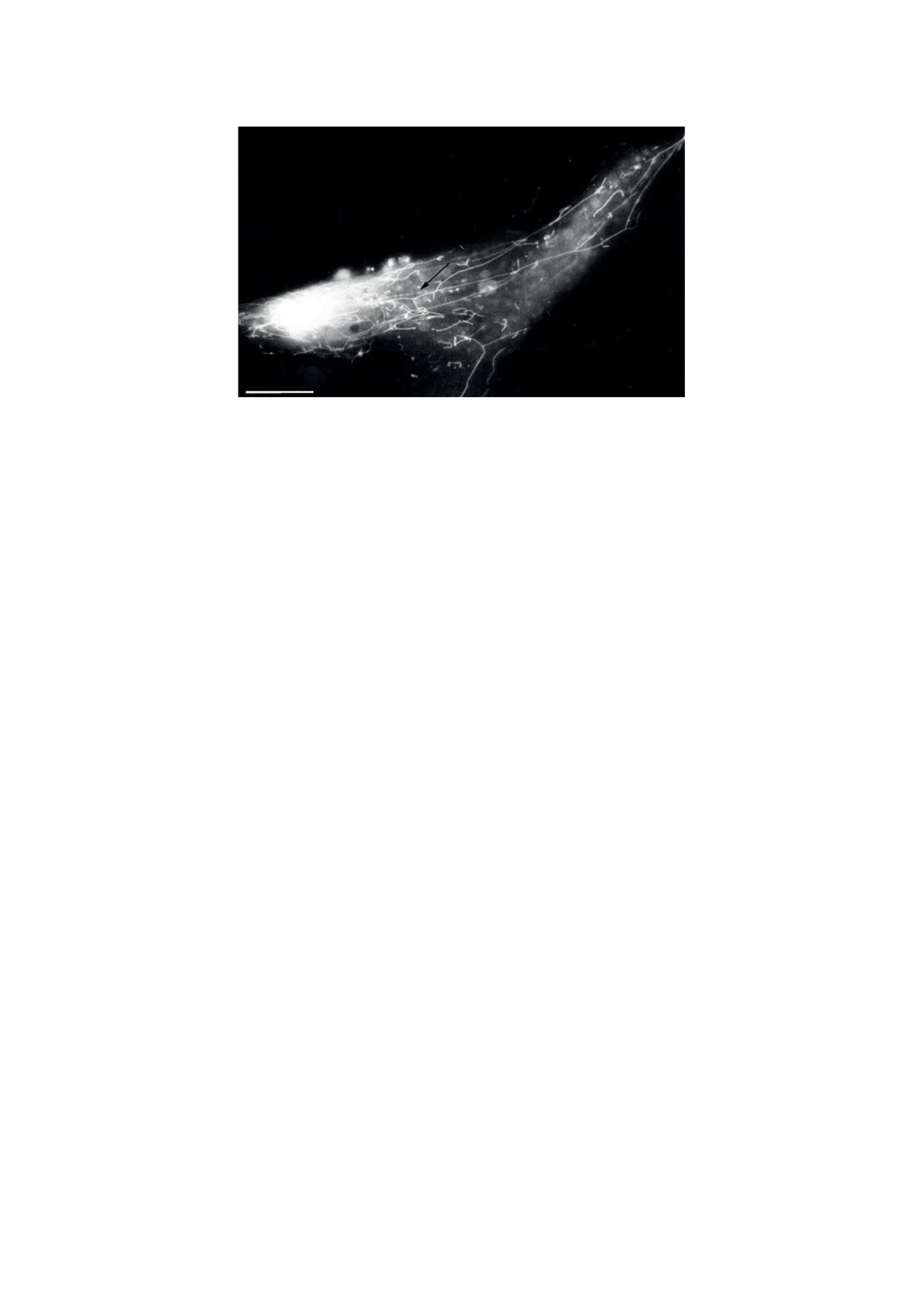
Рис. 2. Митохондриальная сеть в подкожном фибробласте человека (окраска этилродамином). Стрелка указывает
на фрагментированный митохондриальный филамент. Масштабная линейка - 10 мкм
для многих научных групп, и эти работы
астроглиальных клеток млекопитающих, в
продолжаются, в результате чего определен
клетках Saccharomyces cerevisiae [35], в яйце-
целый набор белков, участвующих в этом
клетке Xenopus [36] и сперматидах насекомых,
процессе (например, см. обзор Giacomel-
образующих так называемый небенкерн [37].
lo et al. [31]). Однако пока никто не проводил
По второму сценарию митохондриальный ре-
анализ минимального размера митохондри-
тикулум организован отдельными малыми ми-
ального фрагмента и его состава. Этот кажу-
тохондриальными единицами, каждая со своим
щийся простым вопрос не слишком тривиален
изолированным друг от друга матриксом, объ-
и связан с проблемой гетерогенности популя-
единенными за счет плотных контактов друг с
ции митохондрий и их асимметрического
другом. Самый старый, но не очень известный
деления [32-34], в результате которого обра-
пример - митохондрии в ряде сперматозои-
зуемые продукты фрагментации могут сильно
дов [38-41], в частности млекопитающих, где
отличаться по составу и последующей судьбе.
отдельные митохондрии, соединяясь друг с дру-
Остается при этом неясным и вопрос: все ли
гом при помощи определенного «цемента», об-
митохондриальные фрагменты имеют в своем
разуют средний отдел сперматозоида, органи-
составе митохондриальную ДНК, чтобы хоть в
зованный этими митохондриями по винтовой
малой степени обеспечить себе относительную
линии с разным числом витков в зависимости
автономность? Правда, последний вопрос мо-
от вида животного. Среди разнообразной тер-
жет быть не слишком принципиальным, если
минологии, описывающей межмитохондриаль-
учесть, что при устранении окислительного
ные образования в сперматидах, часто встреча-
стресса наблюдается процесс, обратный фраг-
ется термин «nuage», имеющий более широкое
ментации, когда митохондриальные фрагмен-
применение для описания внутриклеточных
ты могут начинать сливаться. Однако вопрос,
структур [40] (также см. обзор Motta et al. [41]).
насколько разными или одинаковыми должны
В скелетной мышце почти 50 лет назад были
быть фрагменты по составу, чтобы слиться во-
подробно описаны
«межмембранные кон-
едино, остается открытым.
такты» в виде электронноплотного материала,
соединяющего соседние митохондрии в пре-
делах одной клетки (так называемые четырех-
ОРГАНИЗАЦИЯ
мембранные контакты, то есть покрывающие
МИТОХОНДРИАЛЬНОГО РЕТИКУЛУМА
пространство между внутренними мембранами
двух митохондрий, включающее и обе внешние
Митохондриальный ретикулум может быть
мембраны). Одновременно были описаны ше-
организован двумя путями. Одна система обра-
стимембранные контакты такого же типа и для
зована таким образом, что митохондриальный
зоны так называемого нексуса, где между мито-
матрикс един и образует континуум на всем
хондриями образуется контакт, включающий
протяжении митохондрии даже в условиях ее
также две плазматические мембраны соседних
разветвления. Примером такой организации
клеток [18, 19, 42] и была подробно рассмо-
является гигантский митохондриальный рети-
трена организация митохондриального ретику-
кулум у нормальных фибробластов (рис. 2),
лума в онтогенезе [19].
БИОХИМИЯ том 88 вып. 10 2023
1930
АБРАМИЧЕВА и др.
Почти через полвека такого же типа иссле-
Так как флуоресцентные проникающие кра-
дования были продублированы [43, 44], но но-
сители типа родамина 123 или этилродамина
визна этих более поздних работ ограничивалась
позволяют выявить лишь энергизованные
лишь более изощренной методической базой,
митохондрии (у которых есть мембранный
использованием новых терминов «коннектом»
потенциал, позволяющий накопить краси-
и
«интерактом» и соотнесением структур-
тель в матриксе митохондрий теоретически
ной организации митохондриального дерева с
до 10 000-кратных концентраций по сравне-
мышечной активностью. Последнее является
нию с внеклеточным содержанием), то было
важным моментом, ибо предполагается соот-
отмечено изменение мембранного потенциала
ветствующая организация митохондриального
по всей длине протяженной митохондрии по-
ретикулума в скелетной мускулатуре [45], кото-
сле точечного поражения митохондриального
рая наилучшим образом соответствует энер-
филамента. Последнее было осуществлено при
гетическим нуждам мышечной клетки. Надо
помощи сфокусированного лазерного луча,
отметить, что проблема соответствия энерге-
при этом размер его пятна был сопоставим с
тических потребностей и доставки компонен-
толщиной митохондриального тяжа и состав-
тов и производства энергии в клетке (проблема
лял доли микрона. Результат опытов под-
энергетического поступления и расходования)
твердил гипотезу об эквипотенциальности
является одним из ключевых элементов воз-
митохондриального филамента, поскольку ло-
никновения сердечных, мозговых и других кле-
кальный пробой митохондриальной мембраны
точных и органных патологий при нарушении
вызвал полную деэнергизацию всей протяжен-
этого соответствия, и митохондриальная орга-
ной митохондрии, проявившуюся в потере
низация и коммуникация играет принципи-
флуоресценции митохондриального красителя
альную роль в этих процессах [45].
по всей ее длине [50]. Позже были поставлены
Несомненно, что структурное взаимодей-
аналогичные опыты с неонатальными кардио-
ствие митохондрий между собой и с другими
миоцитами, митохондрии которых также орга-
клеточными элементами является лишь од-
низованы в виде разветвленного дерева. Но в
ной из частей внутриклеточной коммуника-
отличие от фибробластов, где матрикс мито-
ции. В основном эта коммуникация реализу-
хондрии непрерывен и распространяется на
ется через обмен химическими сигнальными
всю длину без разрывов, весь митохондриаль-
молекулами, которые могут быть спрятаны
ный ретикулум в данном случае организован
в мембранные везикулы, генерируемые раз-
в виде отдельных митохондриальных единиц,
ными элементами клетки, включая митохон-
объединенных при помощи межмитохондри-
дрии [46-49].
альных контактов. В результате этого митохон-
Подводя итог, гипотеза функционирова-
дриальная сеть неонатальных кардиомиоцитов
ния протяженных митохондриальных структур
имеет множество не соединенных между собой
как электрических энергопередающих кабелей
митохондриальных матриксов. В этом случае
состояла в адекватном и равномерном снабже-
при локальном точечном поражении одного
нии всех компонентов клетки электрической
элемента митохондриальной сети деэнерги-
энергией, которую можно трансформировать
зовалась часть общей сети. Это означало, что
в химическую энергию АТР.
в структурно-функциональном плане в нео-
натальном кардиомиоците митохондрии в
одиночной клетке организованы в виде от-
ДОКАЗАТЕЛЬСТВО
дельных изопотенциальных кластеров, также
ФУНКЦИОНИРОВАНИЯ ПРОТЯЖЕННЫХ
состоящих из отдельных митохондрий, объ-
МИТОХОНДРИАЛЬНЫХ СИСТЕМ
единенных при помощи межмембранных кон-
КАК ЭЛЕКТРИЧЕСКИХ КАБЕЛЕЙ
тактов [51]. Полный электрический пробой,
наблюдаемый при деэнергизации отдельного
Прошло всего три года после открытия
кластера, означал, что межмитохондриальные
фрагментации митохондриального ретику-
контакты являются электропроницаемыми,
лума, когда было проведено эксперимен-
обеспечивая коммуникацию соседних матрик-
тальное доказательство функционирования
сов митохондрий предположительно за счет на-
митохондрии как электрического кабеля [50].
личия ионных каналов во внутренних и внеш-
В качестве объекта использовались подкож-
них мембранах контактирующих митохондрий.
ные фибробласты человека, у которых в норме
Более того, интересной была архитектура крист
митохондрии образуют единую сеть из очень
в межмитохондриальных контактах
- они
протяженных одиночных или объединенных
располагались четко перпендикулярно плану
в сеть филаментов длиной в сотни микрон.
контактной зоны, что, вероятно, облегчало
БИОХИМИЯ том 88 вып. 10 2023
МИТОХОНДРИАЛЬНАЯ СЕТЬ
1931
распространение мембранного потенциала по
приводили как старые, так и современные дан-
митохондриальной сети [52].
ные о структуре и организации крист [60, 61].
Позже было обнаружено, что в межмито-
Эти данные условно можно представить как
хондриальных контактах кардиомиоцитов, ко-
доказательство наличия внутри митохондрий
торые на электронномикроскопических изо-
субмитохондриальных частиц, имеющих или
бражениях представлены осмиофильной суб-
не имеющих электрический контакт с внутрен-
станцией, обильно присутствует поровый белок
ней митохондриальной мембраной, что под-
внешних мембран - вольт-зависимый анион-
рывает старые представления о том, что мито-
ный канал [53], что могло быть рассмотрено
хондриальные кристы образуют континуум с
как структурное подтверждение ионоканаль-
внутренней мембраной. На основании такого
ного сообщения между митохондриями в зоне
представления о структуре митохондрий кри-
контакта.
сты могут представлять собой отдельные орга-
Впоследствии с использованием тех же
низации сопрягающих мембран, не всегда име-
подходов была продемонстрирована электри-
ющие электрическое соединение с внутренней
ческая связь между митохондриями в спермато-
мембраной, и они могут иметь разный транс-
зоиде [54] и в клетках трихомы [55]. Это также
мембранный потенциал. При этом внутренняя
предполагало наличие ионных каналов в зоне
мембрана митохондрий представляет собой
контактов митохондрий сперматозоидов (ко-
непрерывную и эквипотенциальную структуру
торые, как мы уже указывали выше, визуали-
и является материальной основой теории ми-
зировались как электронно-плотные образо-
тохондриального электрического кабеля.
вания, что послужило основой для их названия
Однако нельзя исключить еще один сцена-
«митохондриальным цементом» [40, 41]).
рий, который без критической оценки методи-
Более чем через 25 лет после формули-
ческих особенностей полученных данных [62]
ровки и доказательства существования мито-
может быть интерпретирован как отсутствие
хондрий как электрического кабеля в жур-
кабельных свойств протяженных митохондрий.
нале Nature была опубликована работа из
Это касается опытов по оценке энергизации
лаборатории Роберта Балабана, в точности
митохондрий при использовании красителя
воспроизводящая уже описанную идею, прин-
JC-1, флуоресценция которого зависит от кон-
цип и методический характер организации и
центрации зонда. При высоких концентраци-
функционирования митохондриального рети-
ях JC-1, соответствующих его концентрации
кулума в кардиомиоцитах [56] с той лишь раз-
в матриксе высокоэнергизованных митохон-
ницей, что использовалась более современ-
дрий, образуются так называемые J-агрегаты,
ная методическая база. Позже в ряде других
при возбуждении испускающие свет в красной
публикаций (например, Glancy et al. [57]) из
области, в то время как в низкопотенциаль-
этой же лаборатории по непонятным причи-
ных митохондриях, где концентрация зонда
нам однозначно интерпретируемые опыты,
не столь высокая, J-агрегаты не образуются, и
поставленные в Москве, были поставлены
при возбуждении испускается свет в зеленой
под сомнение («However, there are contradictory
области спектра. Однако при использовании
reports of the functional connectivity of the car-
обычной флуоресцентной микроскопии было
diac mitochondrial reticulum» [57]), правда, с
замечено, что на протяжении одного митохон-
последующим подтверждением правоты ран-
дриального филамента можно было наблюдать
них идей и доказательств.
чередование областей с красной и зеленой
Кроме примеров игнорирования первоис-
флуоресценцией, что могло трактоваться как
точника кабельной теории протяженных сопря-
отсутствие эквипотенциальности по длине
гающих мембран, включая внутренние мембра-
филамента и определенное чередование низ-
ны митохондрий, была осуществлена в какой-то
ко- и высокопотенциальных отсеков по длине
мере критика основ кабельной теории функцио-
одного митохондриального филамента [62].
нирования митохондриального ретикулума, что
Есть два момента, которые надо учитывать
требует особого рассмотрения [58]. На очень хо-
при использовании и интерпретации флуо-
рошей методической основе было показано, что
ресцентных сигналов от JC-1. Во-первых, этот
кристы в пределах одной митохондрии могут
агент достаточно липофилен и будет распреде-
имеет разный трансмембранный потенциал.
ляться не только в соответствии со значения-
Сами авторы данного исследования отчетливо
ми мембранного потенциала на внутренней
понимали некоторое расхождение их данных с
митохондриальной мембране, но и в соответ-
кабельной теорией и предлагали разумное объ-
ствии с липофильным окружением в мито-
яснение, основы которого мы обсуждали ранее
хондрии, которое может быть неодинаково
(см. рис. 3 в статье Зоров и соавт. [59]), когда
по всему объему митохондрий. Но главное -
БИОХИМИЯ том 88 вып. 10 2023
1932
АБРАМИЧЕВА и др.
Рис. 3. Фрагментация митохондрий в клетках. Окраска этилродамином. а - Нормальный фибробласт человека.
б - Та же клетка после 10-секундного воздействия возбуждающим светом. Масштабная линейка - 5 мкм. в - Элек-
тронная микроскопия митохондрии в клетке культуры почек эмбриона свиньи, подвергнутой воздействию ротено-
на (1 мкМ, 24 ч). Видно наличие отдельных по морфологии компартментов, разделенных септой (указана стрелкой).
Масштабная линейка - 0,3 мкм
это то, что эта «многоцветность» одиночной
К такому же выводу о непрерывности ма-
митохондрии в клетке появляется не сразу, а в
трикса в пределах одной несептированной ми-
процессе наблюдения, что может быть резуль-
тохондрии (одиночного электрического кабеля)
татом фотодинамического эффекта, приводя-
и о невозможности функциональной комму-
щего сначала к образованию септ внутри ми-
никации матриксов соседних митохондрий
тохондрий, разделяющих одну протяженную
пришли и другие исследователи. Полученные
митохондрию на ряд электрически изолиро-
данные послужили структурно-функциональ-
ванных отсеков (рис. 3), с последующей фраг-
ной основой понимания морфологической и
ментацией митохондриального филамента.
функциональной гетерогенности митохондрий
Этот процесс образования септ может про-
в клетке [64, 65].
исходить в секундной шкале при облучении
Завершая этот раздел, можно с достаточ-
достаточно сильным светом, возбуждающим
ной уверенностью сказать, что функциониро-
краситель. Электронно-микроскопические дан-
вание митохондрии как электрического кабеля
ные подтверждают, что в процессе инициации
является доказанным. Однако вполне веро-
фрагментации митохондриального ретикулума
ятно, что митохондриальные сети могут нести
септа может разделять митохондрию на отсеки
и другие гипотетические функции, которые
разной конфигурации, соответствующие раз-
мы обсудим.
ной степени энергизации (рис 3, в, а также
рис. 6 в статье Zorov et al. [29]).
Критиков электрической кабельной тео-
ДРУГИЕ ГИПОТЕТИЧЕСКИЕ
рии митохондрий было существенно меньше,
ФУНКЦИИ ПРОТЯЖЕННЫХ
чем тех, которые поддерживали эту концеп-
МИТОХОНДРИАЛЬНЫХ СТРУКТУР
цию. Очень важной работой, подкрепляющей
базовые принципы кабельной теории, было
Обеспечение однородности распределения
решение вопроса о том, насколько непреры-
редокс-потенциала по всему объему клетки.
вен матрикс по всему пространству митохон-
Рассматривая одиночную, но единую, непре-
дриального дерева. В одном из исследований
рывную по матриксу митохондриальную сеть,
использовали фотоактивируемый зеленый
на всем протяжении которой ΔΨ одинаков (вся
флуоресцентный белок, находящийся исклю-
сеть эквипотенциальна), а сам мембранный
чительно в матриксе митохондрий, для мече-
потенциал отражает работу протонных помп,
ния отдельных митохондриальных сетей в клет-
приводимых в действие за счет окисления
ке в сочетании с мониторингом мембранного
восстановленных эквивалентов, прежде всего
потенциала митохондрий в режиме реального
NAD(Р)H, надо учесть, что мембранный потен-
времени [63]. При этом была обнаружена не-
циал (ΔΨ), вернее, электрохимический потен-
прерывность матрикса в пределах одного изо-
циал ионов водорода (ΔμН+) в стационарных
потенциального митохондриального кластера.
условиях должен находиться в термодинамиче-
Неизменная эквипотенциальность отдельных
ском равновесии с редокс-потенциалом, созда-
митохондриальных сетей позволила предполо-
ваемым парой NAD(Р)H/NAD(Р). В свою оче-
жить, что гетерогенный характер мембранного
редь, эта пара находится в равновесии с парой
потенциала в митохондриях клетки отражает
восстановленный глутатион/окисленный глу-
различия между отдельными сетями.
татион (GSH/GSSG), при этом оба являются
БИОХИМИЯ том 88 вып. 10 2023
МИТОХОНДРИАЛЬНАЯ СЕТЬ
1933
основными буферными редокс-системами в
низмом De Grotthuss [68] по упорядоченным
клетке [66].
молекулам воды в примембранных слоях [69].
Таким образом, за счет эквипотенциаль-
Таким образом, как и в случае с предполагае-
ности митохондриальной сети, в идеальном
мым «перемешиванием» редокс-потенциала,
случае покрывающей всю толщу клетки, эта
гигантская митохондрия будет
«перемеши-
сеть обеспечивает равномерное распределе-
вать» рН по всей длине своего окружения.
ние редокс-потенциала по всему объему клет-
Уравновешивание внутриклеточных концен-
ки, являясь в некотором роде «мешалкой», не
траций ионов калия. Учитывая недавно откры-
допуская создания больших локальных обла-
тую возможность митохондриальной калие-
стей с разным редокс-потенциалом. Ситуация
вой энергетики [6-8], приводимой в действие
может резко измениться в условиях вынужден-
трансмембранным потенциалом, мы вправе
ной фрагментации/расщепления митохондри-
ожидать равномерное распределение ионов
альной сети, когда каждый митохондриальный
калия по внутриклеточному объему в окруже-
фрагмент будет создавать вокруг себя редокс-
нии гигантской митохондрии.
окружение в соответствии с величиной мем-
Таким образом, митохондриальная сеть
бранного потенциала на внутренней мембране
по многим параметрам может служить струк-
этого фрагмента. Учитывая, что фрагмента-
турой, обеспечивающей равномерное рас-
ция ассоциируется с наличием окислительного
пределение по клетке разных компонентов, и
стресса, приводящего к увеличению гетеро-
служить своеобразной внутриклеточной «ме-
генности митохондрий по мембранному по-
шалкой».
тенциалу [29, 34], вполне ожидаемой является
гетерогенность распределения редокс-потен-
циала в клетке после фрагментации мито-
МЕДИЦИНСКИЕ АСПЕКТЫ
хондрий.
Проводник кислорода. Известно, что рас-
В приведенном материале мы делали ак-
творимость кислорода в липидных мембранах
цент на проблемах адекватного снабжения
и их гидрофобных компонентах выше, чем в
живой системы необходимым материалом для
водной фазе, в результате чего в митохондри-
нормального течения метаболизма, в част-
альных мембранах концентрация кислорода
ности энергетического метаболизма, будь то
выше, чем в цитозоле. Несмотря на то что
клетка, орган или организм. Полное соответ-
именно во внутренней мембране митохондрий
ствие доставки насущным нуждам живой клет-
локализован основной потребитель кислоро-
ки определяет понятие гомеостаза, и откло-
да (цитохромоксидаза), все равно мембраны
нения в любую сторону могут быть чреваты
можно рассматривать как буферы кислорода,
возникновением патологического фенотипа.
помогающие облегчать диффузию О2 по мито-
Эти отклонения могут быть вызваны физико-
хондрии. Это в какой-то степени обеспечива-
химическим воздействием на систему, и од-
ет уравновешивание распределения кислоро-
ним из таких факторов является воздействие,
да по объему клетки и не допускает создания
сопровождающееся возникновением окисли-
локальных гипоксических областей. Конечно
тельного стресса. Патогенез окислительного
же это будет сильно зависеть от активности ми-
стресса слишком очевиден, и приведение при-
тохондриального дыхания, что определяет диа-
меров такого рода занимает большую часть
метр так называемого цилиндра Крога (объем-
научной литературы. Мы можем лишь кратко
ное распределение кислорода в ткани вокруг
коснуться тех медицинских проблем, кото-
капилляров, несущих кислород, зависящее от
рые составляют малую долю от громадного
скорости доставки кислорода и его потреб-
числа широко обсуждаемых нейрологических
ления [67]).
или кардиологических аспектов, оставляя на
Проводник протонов. В случае равнознач-
заднем плане проблемы акушерства и нео-
ности не только мембранного потенциала по
натологии, хотя задача поддержания редокс-
всей длине митохондрии, но и всего ΔμН+, в
гомеостаза в клетках там имеет не меньшее
состав которого, кроме значения мембранного
значение.
потенциала, входит и градиент ионов водо-
Из нашего логического предположения,
рода (ΔpH), величина ΔpH также будет одина-
что митохондриальная сеть может служить для
кова по всей ее длине. Это приведет к тому,
обеспечения равновесия редокс-потенциала
что окружение гигантской митохондрии будет
в клетке, следует, что окислительный стресс,
иметь одинаковые значения рН, даже без учета
как первопричина или следствие внутренних
возможности ускоренного проведения про-
нарушений и следующие за этим измене-
тонов по мембранам в соответствии с меха-
ния структуры сети, уводит внутриклеточное
БИОХИМИЯ том 88 вып. 10 2023
1934
АБРАМИЧЕВА и др.
содержимое от состояния равновесия, что
предшествует внутренняя перестройка мито-
неминуемо приводит к изменению внутри-
хондрий с последующим отделением фраг-
клеточного метаболического гомеостаза. Это
мента митохондрий с неправильно функцио-
касается громадного ряда патологий, и мы
нирующим содержимым от митохондриальной
лишь в качестве примера возьмем генераль-
сети (которая может сохранять свою трехмер-
ные проблемы акушерства, гинекологии и
ную структуру), и в этом состоит принцип ми-
неонатологии, которые составляют ведущую
тофагии (митоптоза) [29]. Однако и в том, и в
часть заболеваемости и детской и взрослой
другом случае фрагментация (расщепление)
смертности. Особо надлежит обратить вни-
идет с участием белков расщепления (напри-
мание на окислительный стресс, сопрово-
мер, Drp1 и Fis1), уровень которых в плаценте
ждающий разные патологии, являющиеся
коррелирует с тяжестью заболевания матери и
предметом этих отраслей медицины [70, 71],
плода, включая размер последнего [81, 82], что
в частности такие как преэклампсия, опреде-
свидетельствует о прямой связи патологии с
ляемая как гипертензия и протеинурия, воз-
митохондриальной структурой, которая может
никшие после 20 недель гестации, и задержка
служить показателем патологического фено-
роста плода [72, 73]. Одной из причин пре-
типа клетки.
эклампсии является имплантация плаценты,
Очевидность вовлеченности окислитель-
приводящая к ремоделированию артерий ма-
ного стресса в патогенез и необходимость
тери и снижению кровотока к плоду. Это
поддержания редокс-состояния клетки и ее
может вызывать резкую восприимчивость к
компонентов, прежде всего митохондрий,
колебаниям в кровотоке и, как следствие, при-
привели к пониманию, что одной из возмож-
водить к изменениям редокс-статуса клеток
ностей терапевтического вмешательства явля-
как плаценты, так и плода, характерным для
ется нормализация редокс-статуса в клетках
ишемии-реперфузии. Реперфузионное пов-
органа. В недавней работе при использовании
реждение плаценты сопровождается генера-
эндотелиальных клеток пупочной вены чело-
цией активных форм кислорода и азота с обра-
века было показано, что после воздействия
зованием окисленных продуктов, обладающих
на них плазмы крови беременных женщин с
сигнальными и патогенными свойствами при
преэклампсией наблюдается снижение функ-
значительном повышении их концентраций,
ций митохондрий этих клеток, ассоцииро-
что, прежде всего, приводит к дисфункции
ванное с повышенной генерацией активных
клеток эндотелия [74, 75]. Таким образом, эти
форм кислорода [83]. Одновременно в клет-
продукты являются факторами риска сердеч-
ках наблюдалась повышенная экспрессия мар-
но-сосудистых заболеваний, что позволяет
керов воспаления TNF-α, TLR-9 и ICAM-1.
предположить о возможной функциональной
Митохондриально-направленный антиокси-
связи между плацентой и сердечно-сосуди-
дант MitoTempo снижал выработку супер-
стой системой, причем в основе такой сигна-
оксида митохондриями в клетках, подверг-
лизации лежат компоненты, поддерживаю-
шихся воздействию плазмы крови беременных
щие или нарушающие редокс-статус матери
с преэклампсией, нормализовал митохондри-
и плода [76].
альный метаболизм и значительно восстанав-
Пришло понимание того, что именно ми-
ливал воспалительный профиль клеток. Эти
тохондрии в должной мере определяют редокс-
данные подтверждают функциональную роль
статус биологической системы и плаценты, в
митохондриальной редокс-сигнализации в па-
частности [77-80]. Как мы указывали раньше,
тогенезе преэклампсии и предлагают терапев-
именно лабильность митохондриальных струк-
тические пути, направленные на сохранение
тур и функций в значительной степени явля-
митохондриальной структуры и функций.
ется показателем редокс-статуса клетки и не
только. Процесс фрагментации митохондрий,
Вклад авторов. П.А. Абрамичева, Н.В. Ан-
как почти обязательный ответ на возникший
дрианова, В.А. Бабенко, Л.Д. Зорова, С.Д. Зо-
окислительный стресс, является также и обяза-
ров, И.Б. Певзнер, В.А. Попков, Д.С. Семе-
тельным в механизме контроля качества мито-
нович, Э.И. Якупова, Д.Н. Силачев, Е.Ю. Плот-
хондрий [29, 33]. Разница между первой и вто-
ников, Г.Т. Сухих, Д.Б. Зоров - общее обсу-
рой опцией состоит в том, что окислительный
ждение идеологии и планов работы; Д.Б. Зо-
стресс приводит к глобальной фрагментации
ров
- написание рукописи; Д.Н. Силачев,
митохондриальной популяции в одиночной
Е.Ю. Плотников, Г.Т. Сухих, Д.Б. Зоров - руко-
клетке, в то время как в процессе програм-
водство и контроль исследования; П.А. Абрами-
мируемого уничтожения нефункциональных
чева, Н.В. Андрианова, В.А. Бабенко, Л.Д. Зо-
митохондрий фрагментация локальна, и ей
рова, С.Д. Зоров, И.Б. Певзнер, В.А. Попков,
БИОХИМИЯ том 88 вып. 10 2023
МИТОХОНДРИАЛЬНАЯ СЕТЬ
1935
Д.С. Семенович, Э.И. Якупова, Д.Н. Силачев,
Конфликт интересов. Авторы заявляют об
Е.Ю. Плотников, Г.Т. Сухих - редактирование
отсутствии конфликта интересов.
рукописи.
Соблюдение этических норм. Настоящая
Финансирование. Работа выполнена при
статья не содержит каких-либо исследований с
поддержке Российского научного фонда (грант
участием людей и животных в качестве объек-
№ 19-14-00173-П).
тов исследования.
СПИСОК ЛИТЕРАТУРЫ
1.
Mitchell, P.
(1961) Coupling of phosphorylation
10.
Williams, R. J. P. (1961) Possible functions of chains
to electron and hydrogen transfer by a chemi-
of catalysts, J. Theor. Biol., 1, 1-13, doi: 10.1016/
osmotic type of mechanism, Nature, 191, 144-148,
0022-5193(61)90023-6.
doi: 10.1038/191144a0.
11.
Williams, R. J. P. (1962) Possible functions of chains
2.
Mitchell, P. (1966) Chemiosmotic coupling in oxidative
of catalysts II, J. Theor. Biol., 3, 209-220, doi: 10.1016/
and photosynthetic phosphorylation, Biol. Rev. Camb.
S0022-5193(62)80015-0.
Philos. Soc., 41, 445-502, doi: 10.1111/j.1469-185X.
12.
Williams, R. J. P. (1978) The multifarious couplings
1966.tb01501.x.
of energy transduction, Biochim. Biophys. Acta,
3.
Mitchell, P.
(2011) Chemiosmotic coupling in
505, 1-44, doi: 10.1016/0304-4173(78)90007-1.
oxidative and photosynthetic phosphorylation. 1966,
13.
Williams, R. J. P. (2001) The structures of organ-
Biochim Biophys Acta, 1807, 1507-1538, doi: 10.1016/
elles and reticula: localised bioenergetics and me-
j.bbabio.2011.09.018.
tabolism, Chembiochem, 2, 637-641, doi: 10.1002/
4.
Mitchell, P. (1979) David Keilin’s respiratory chain
1439-7633(20010903)2:9<637::AID-CBIC637>
concept and its chemiosmotic consequences, Science,
3.0.CO;2-7.
206, 1148-1159, doi: 10.1126/science.388618.
14.
Kell, D. B. (1979) On the functional proton current
5.
Juhaszova, M., Kobrinsky, E., Zorov, D. B., Aon,
pathway of electron transport phosphorylation.
M. A., Cortassa, S., and Sollott, S. J. (2022) Setting
An electrodic view, Biochim. Biophys. Acta, 549, 55-99,
the record straight: a new twist on the chemiosmotic
doi: 10.1016/0304-4173(79)90018-1.
mechanism of oxidative Phosphorylation, Function
15.
Mulkidjanian, A. Y., Heberle, J., and Cherepanov,
(Oxf), 3, zqac018, doi: 10.1093/function/zqac018.
D. A. (2006) Protons and interfaces: implications for
6.
Juhaszova, M., Kobrinsky, E., Zorov, D. B., Nuss,
biological energy conversion, Biochim. Biophys. Acta,
H. B., Yaniv, Y., Fishbein, K. W., de Cabo, R.,
1757, 913-930, doi: 10.1016/j.bbabio.2006.02.015.
Montoliu, L., Gabelli, S. B., Aon, M.A., Cortassa, S.,
16.
Zorova, L. D., Popkov, V. A., Plotnikov, E. Y.,
and Sollott, S. J. (2021) ATP synthase K+- and H+-
Silachev, D. N., Pevzner, I. B., Jankauskas, S. S.,
fluxes drive ATP synthesis and enable mitochondrial
Babenko, V. A., Zorov, S. D., Balakireva, A. V.,
K+-“Uniporter” function: I. Characterization of ion
Juhaszova, M, Sollott, S. J., and Zorov, D. B. (2018)
fluxes, Function (Oxf), 3, zqab065, doi: 10.1093/
Mitochondrial membrane potential, Anal. Biochem.,
function/zqab065.
552, 50-59, doi: 10.1016/j.ab.2017.07.009.
7.
Juhaszova, M., Kobrinsky, E., Zorov, D. B., Nuss,
17.
Skulachev, V. P. (1971) Energy transformations in the
H. B., Yaniv, Y., Fishbein, K. W., de Cabo, R.,
respiratory chain, Curr. Top. Bioenergetics, 4, 127-190,
Montoliu, L., Gabelli, S. B., Aon, M. A., Cortassa, S.,
doi: 10.1016/B978-0-12-152504-0.50010-1.
and Sollott, S. J. (2022) ATP synthase K+- and
18.
Bakeeva, L. E., Chentsov, Y. S., and Skulachev,
H+-fluxes drive ATP synthesis and enable mitochon-
V. P.
(1978) Mitochondrial framework (reticulum
drial K+-“Uniporter” function: II. Ion and ATP syn-
mitochondriale) in rat diaphragm muscle, Bio-
thase flux regulation, Function (Oxf), 3, zqac001,
chim. Biophys. Acta,
501,
349-369, doi:
10.1016/
doi: 10.1093/function/zqac001.
0005-2728(78)90104-4.
8.
Зоров Д. Б. (2022) Окно в калиевый мир. Откры-
19.
Bakeeva, L. E., Chentsov, Y. S., and Skulachev, V. P.
тие калиевой энергетики в митохондриях и иден-
(1981) Ontogenesis of mitochondrial reticulum in rat
тичности митохондриального ATP-зависимого
diaphragm muscle, Eur. J. Cell Biol., 25, 175-181.
K+-канала, Биохимия, 87, 1007-1013, doi: 10.31857/
20.
Johnson, L. V., Walsh, M. L., and Chen, L. B.
S0320972522080024.
(1980) Localization of mitochondria in living cells
9.
Cortassa, S., Aon, M. A., Juhaszova, M., Kobrinsky, E.,
with rhodamine 123, Proc. Natl. Acad. Sci. USA, 77,
Zorov, D. B., and Sollott, S. J. (2022) Computa-
990-994, doi: 10.1073/pnas.77.2.990.
tional modeling of mitochondrial K+- and H+-driv-
21.
Bereiter-Hahn, J., and Vöth, M. (1994) Dynamics
en ATP synthesis, J. Mol. Cell Cardiol., 165, 9-18,
of mitochondria in living cells: shape changes,
doi: 10.1016/j.yjmcc.2021.12.005.
dislocations, fusion, and fission of mitochondria,
БИОХИМИЯ том 88 вып. 10 2023
1936
АБРАМИЧЕВА и др.
Microsc. Res. Tech.,
27,
198-219, doi:
10.1002/
(2008) Fission and selective fusion govern mito-
jemt.1070270303.
chondrial segregation and elimination by autoph-
22.
Bensley, R. R. (1911) Studies on the pancreas of the
agy, EMBO J., 27, 433-446, doi: 10.1038/sj.emboj.
guinea pig, Am. J. Anat., 12, 297-388, doi: 10.1002/
7601963.
aja.1000120304.
33.
Zorov, D. B., Popkov, V. A., Zorova, L. D., Vorobjev,
23.
Мурванидзе Г. В., Северина И. И., Скулачев В. П.
I. A., Pevzner, I. B., Silachev, D. N., Zorov, S. D.,
(1981) Этилродамин - проникающий катион и
Jankauskas, S. S., Babenko, V. A., and Plotnikov, E. Y.
флуоресцентный индикатор мембранного потен-
(2017) Mitochondrial aging: is there a mitochondrial
циала цианобактерий in vivo, Докл. Акад. Наук
clock? J. Gerontol. A Biol. Sci. Med. Sci., 72, 1171-1179,
СССР, 261, 1252-1254.
doi: 10.1093/gerona/glw184.
24.
Vorobjev, I. A., and Zorov, D. B. (1983) Diazepam
34.
Попков В. А., Плотников Е. Ю., Лямзаев К. Г.,
inhibits cell respiration and induces fragmentation of
Силачев Д. Н., Зорова Л. Д., Певзнер И. Б., Ян-
mitochondrial reticulum, FEBS Lett., 163, 311-314,
каускас С. С., Зоров С. Д., Бабенко В. А., Зоров Д. Б.
doi: 10.1016/0014-5793(83)80842-4.
(2015) Миторазнообразие, Биохимия, 80, 631-641.
25.
Avad, A. S., Vorobjev, I. A., and Zorov, D. B. (1984)
35.
Hoffman, H. P., and Avers, C. H. (1973) Mitochon-
Fragmentation of mitochondrial reticulum, Proceed-
drion of yeast: ultrastructural evidence for one giant,
ings of the XVI Congress of FEBS, abst. XI-80.
branched organelle per cell, Science, 181, 749-751,
26.
Полякова И. А., Зоров Д. Б., Лейкина М. И. (1995)
doi: 10.1126/science.181.4101.749.
Структурно-функциональные изменения хон-
36.
Heasman, J., Quarmby, J., and Wylie, C. C. (1984)
дриома культивированных клеток при нарушении
The mitochondrial cloud of Xenopus oocytes: the
энергетического метаболизма, Докл. Акад. Наук
source of germinal granule material, Dev. Biol., 105,
СССР, 342, 553-555.
458-469, doi: 10.1016/0012-1606(84)90303-8.
27.
Skulachev, V. P., Bakeeva, L. E., Chernyak, B. V.,
37.
Pratt, S. A. (1968) An electron microscope study of
Domnina, L. V., Minin, A. A., Pletjushkina, O. Y.,
nebenkern formation and differentiation in spermatids
Saprunova, V. B., Skulachev, I. V., Tsyplenkova, V. G.,
of Murgantia histrionica (Hemiptera, Pentatomidae),
Vasiliev, J. M., Yaguzhinsky, L. S., and Zorov, D. B.
J. Morphol., 126, 31-65, doi: 10.1002/jmor.1051260104.
(2004) Thread-grain transition of mitochondrial re-
38.
Andre, J. (1962) Contribution to the knowledge of the
ticulum as a step of mitoptosis and apoptosis, Mol.
chondrioma study of its ultrastructural modifications
Cell. Biochem.,
256-257,
341-358, doi:
10.1023/
during spermatogenesis [in French], J. Ultrastruct.
B:MCBI.0000009880.94044.49.
Res., Suppl. 3, 1-185.
28.
Plotnikov, E. Y., Vasileva, A. K., Arkhangelskaya,
39.
Fawcett, D. W. (1958) The structure of the mam-
A. A., Pevzner, I. B., Skulachev, V. P., and Zorov, D. B.
malian spermatozoon, Int. Rev. Cytol., 7, 195-234,
(2008) Interrelations of mitochondrial fragmentation
doi: 10.1016/S0074-7696(08)62688-1.
and cell death under ischemia/reoxygenation and UV-
40.
Eddy, E. M. (1974) Fine structural observations on the
irradiation: protective effects of SkQ1, lithium ions
form and distribution of nuage in germ cells of the rat,
and insulin, FEBS Lett., 582, 3117-3124, doi: 10.1016/
Anat. Rec., 178, 731-757, doi: 10.1002/ar.1091780406.
j.febslet.2008.08.002.
41.
Motta, P. M., Nottola, S. A., Makabe, S., and Heyn, R.
29.
Zorov, D. B., Vorobjev, I. A., Popkov, V. A., Babenko,
(2000) Mitochondrial morphology in human fetal and
V. A., Zorova, L. D., Pevzner, I. B., Silachev, D. N.,
adult female germ cells, Hum. Reprod., 15 Suppl 2,
Zorov, S. D., Andrianova, N. V., and Plotnikov, E. Y.
129-147, doi: 10.1093/humrep/15.suppl_2.129.
(2019) Lessons from the discovery of mitochondrial
42.
Bakeeva, L.E., Chentsov, Y.S., and Skulachev, V. P.
fragmentation (fission): a review and update, Cells,
(1983) Intermitochondrial contacts in myocardiocytes,
8, 175, doi: 10.3390/cells8020175.
J. Mol. Cell Cardiol., 15, 413-420, doi: 10.1016/
30.
Pletjushkina, O. Y., Lyamzaev, K. G., Popova, E. N.,
0022-2828(83)90261-4.
Nepryakhina, O. K., Ivanova, O. Y., Domnina, L. V.,
43.
Bleck, C. K. E., Kim, Y., Willingham, T. B., and
Chernyak, B. V., and Skulachev, V. P. (2006) Effect
Glancy, B. (2018) Subcellular connectomic analyses of
of oxidative stress on dynamics of mitochondrial
energy networks in striated muscle, Nat. Commun., 9,
reticulum, Biochim. Biophys. Acta, 1757, 518-524,
5111, doi: 10.1038/s41467-018-07676-y.
doi: 10.1016/j.bbabio.2006.03.018.
44.
Kim, Y., Ajayi, P. T., Bleck, C. K. E., and Glancy, B.
31.
Giacomello, M., Pyakurel, A., Glytsou, C., and
(2022) Three-dimensional remodelling of the cellular
Scorrano, L. (2020) The cell biology of mitochondrial
energy distribution system during postnatal heart
membrane dynamics, Nat. Rev. Mol. Cell Biol., 21,
development, Philos. Trans. R. Soc. Lond. B Biol. Sci.,
204-224, doi: 10.1038/s41580-020-0210-7.
377, 20210322, doi: 10.1098/rstb.2021.0322.
32.
Twig, G., Elorza, A., Molina, A. J. A., Mohamed, H.,
45.
Yaniv, Y., Juhaszova, M., Nuss, H. B., Wang, S.,
Wikstrom, J. D., Walzer, G., Stiles, L., Haigh, S. E.,
Zorov, D. B., Lakatta, E. G., and Sollott, S. J. (2010)
Katz, S., Las, G., Alroy, J., Wu, M., Py, B. F., Yuan, J.,
Matching ATP supply and demand in mammalian
Deeney, J. T., Corkey, B. E., and Shirihai, O. S.
heart: in vivo, in vitro, and in silico perspectives,
БИОХИМИЯ том 88 вып. 10 2023
МИТОХОНДРИАЛЬНАЯ СЕТЬ
1937
Ann. N Y Acad. Sci., 1188, 133-142, doi: 10.1111/
55.
Severina, I. I., Skulachev, V. P., and Zorov, D. B.
j.1749-6632.2009.05093.x.
(1988) Coupling membranes as energy-transmitting
46.
Boardman N. T., Trani, G., Scalabrin, M., Romanel-
cables. II. Cyanobacterial trichomes, J. Cell Biol., 107,
lo, V., and Wüst, R. C. I. (2023) Intra-cellular to
497-501, doi: 10.1083/jcb.107.2.497.
inter-organ mitochondrial communication in striated
56.
Glancy, B., Hartnell, L. M., Malide, D., Yu, Z. X.,
muscle in health and disease, Endocr. Rev.,
44,
Combs, C. A., Connelly, P. S., Subramaniam, S., and
668-692, doi: 10.1210/endrev/bnad004, doi: 10.1210/
Balaban, R. S. (2015) Mitochondrial reticulum for
endrev/bnad004.
cellular energy distribution in muscle, Nature, 523,
47.
Plotnikov, E. Y., Silachev, D. N., Popkov, V. A.,
617-620, doi: 10.1038/nature14614.
Zorova, L. D., Pevzner, I. B., Zorov, S. D.,
57.
Glancy, B., Hartnell, L. M., Combs, C. A., Femnou, A.,
Jankauskas, S. S, Babenko, V. A., Sukhikh, G. T., and
Sun, J., Murphy, E., Subramaniam, S., and Balaban,
Zorov, D. B. (2017) Intercellular signalling cross-talk:
R. S. (2017) Power grid protection of the muscle
to kill, to heal and to rejuvenate, Heart Lung Circ.,
mitochondrial reticulum, Cell Rep.,
19,
487-496,
26, 648-659, doi: 10.1016/j.hlc.2016.12.002.
doi: 10.1016/j.celrep.2017.03.063.
48.
Turovsky, E. A., Golovicheva, V. V., Varlamova, E. G.,
58.
Wolf, D. M., Segawa, M., Kondadi, A. K., Anand, R.,
Danilina, T. I., Goryunov, K. V., Shevtsova, Y. A.,
Bailey, S. T., Reichert, A. S., van der Bliek, A. M.,
Pevzner, I. B., Zorova, L. D., Babenko, V. A.,
Shackelford, D. B., Liesa, M., and Shirihai, O. S.
Evtushenko, E. A., Zharikova, A. A., Khutornenko,
(2019) Individual cristae within the same mitochon-
A. A., Kovalchuk, S. I., Plotnikov, E. Y., Zorov, D. B.,
drion display different membrane potentials and are
Sukhikh, G. T., and Silachev, D. N. (2022) Mes-
functionally independent, EMBO J., 38, e101056,
enchymal stromal cell-derived extracellular vesicles
doi: 10.15252/embj.2018101056.
afford neuroprotection by modulating PI3K/AKT
59.
Зоров Д. Б., Плотников Е. Ю., Силачев Д. Н.,
pathway and calcium oscillations, Int. J. Biol. Sci., 18,
Зорова Л. Д., Певзнер И. Б., Зоров С. Д., Бабен-
5345-5368, doi: 10.7150/ijbs.73747.
ко В. А., Янкаускас С. С., Попков В. А., Савина
49.
Zorova, L. D., Kovalchuk, S. I., Popkov, V. A.,
П. С. (2014) Микробиота и митобиота. Поставив
Chernikov, V. P., Zharikova, A. A., Khutornenko,
знак равенства между митохондрией и бактерией,
A. A., Zorov, S. D., Plokhikh, K. S., Zinovkin, R. A.,
Биохимия, 79, 1252-1268.
Evtushenko, E. A., Babenko, V. A., Pevzner, I. B.,
60.
Daems, W. T., and Wisse, E. (1966) Shape and
Shevtsova, Y. A., Goryunov, K. V., Plotnikov, E. Y.,
attachment of the cristae mitochondriales in mouse
Silachev, D. N., Sukhikh, G. T., and Zorov, D. B.
hepatic cell mitochondria, J. Ultrastruct. Res., 16,
(2022) Do Extracellular vesicles derived from mes-
123-140, doi: 10.1016/S0022-5320(66)80027-8.
enchymal stem cells contain functional mitochon-
61.
Mannella, C. A., Marko, M., and Buttle, K. (1997)
dria? Int. J. Mol. Sci.,
23,
7408, doi:
10.3390/
Reconsidering mitochondrial structure: new views
ijms23137408.
of an old organelle, Trends Biochem. Sci., 22, 37-38,
50.
Драчев В. А., Зоров Д. Б. (1986) Митохондрия
doi: 10.1016/S0968-0004(96)30050-9.
как электрический кабель. Экспериментальная
62.
Smiley, S. T., Reers, M., Mottola-Hartshorn, C.,
проверка гипотезы, Докл. Акад. Наук СССР, 287,
Lin, M., Chen, A., Smith, T. W., Steele, G. D., Jr.,
1237-1238.
and Chen, L. B. (1991) Intracellular heterogeneity
51.
Amchenkova, A. A., Bakeeva, L. E., Chentsov, Y. S.,
in mitochondrial membrane potentials revealed by
Skulachev, V. P., and Zorov, D. B. (1988) Coupling
a J-aggregate-forming lipophilic cation JC-1, Proc.
membranes as energy-transmitting cables. I. Filamen-
Natl. Acad. Sci. USA, 88, 3671-3675, doi: 10.1073/
tous mitochondria in fibroblasts and mitochondrial
pnas.88.9.3671.
clusters in cardiomyocytes, J. Cell. Biol., 107, 481-495,
63.
Twig, G., Graf, S. A., Wikstrom, J. D., Mohamed, H.,
doi: 10.1083/jcb.107.2.481.
Haigh, S. E., Elorza, A., Deutsch, M., Zurgil, N.,
52.
Picard, M., McManus, M. J., Csordás, G., Várnai, P.,
Reynolds, N., and Shirihai, O. S. (2006) Tagging
Dorn, G. W., 2nd, Williams, D., Hajnóczky, G., and
and tracking individual networks within a complex
Wallace, D. C. (2015) Trans-mitochondrial coordina-
mitochondrial web with photoactivatable GFP, Am. J.
tion of cristae at regulated membrane junctions, Nat.
Physiol. Cell Physiol., 291, C176-C184, doi: 10.1152/
Commun., 6, 6259, doi: 10.1038/ncomms7259.
ajpcell.00348.2005.
53.
Konstantinova, S. A., Mannella, C. A., Skulachev,
64.
Collins, T. J., and Bootman, M. D. (2003) Mitochon-
V. P., and Zorov, D. B. (1995) Immunoelectron
dria are morphologically heterogeneous within cells,
microscopic study of the distribution of porin on outer
J. Exp. Biol., 206 (Pt 12), 1993-2000, doi: 10.1242/
membranes of rat heart mitochondria, J. Bioenerg.
jeb.00244.
Biomembr., 27, 93-99, doi: 10.1007/BF02110336.
65.
Collins, T. J., Berridge, M. J., Lipp, P., and Bootman,
54.
Zorov, D. B., Skulachev, V. P., and Halangk, V. (1990)
M. D. (2002) Mitochondria are morphologically and
Membranous electric cable. 4. Mitochondrial helix of
functionally heterogeneous within cells, EMBO J.,
the rat spermatozoa, Biol. Membr., 7, 243-249.
21, 1616-1627, doi: 10.1093/emboj/21.7.1616.
15
БИОХИМИЯ том 88 вып. 10 2023
1938
АБРАМИЧЕВА и др.
66.
Ghosh, D., Levault, K. R., and Brewer, G. J. (2014)
75.
Ness, R. B., and Sibai, B. M. (2006) Shared and
Relative importance of redox buffers GSH and
disparate components of the pathophysiologies of fetal
NAD(P)H in age-related neurodegeneration and
growth restriction and preeclampsia, Am. J. Obstet.
Alzheimer disease-like mouse neurons, Aging Cell,
Gynecol., 195, 40-49, doi: 10.1016/j.ajog.2005.07.049.
13, 631-640, doi: 10.1111/acel.12216.
76.
Manna, S., Ruano, C. S. M., Hegenbarth, J. C.,
67.
McGuire, B. J., and Secomb, T. W. (2001) A the-
Vaiman, D., Gupta, S., McCarthy, F. P., Méhats, C.,
oretical model for oxygen transport in skeletal
McCarthy, C., Apicella, C., and Scheel, J. (2022)
muscle under conditions of high oxygen demand,
Computational models on pathological redox sig-
J. Appl. Physiol. (1985), 91, 2255-2265, doi: 10.1152/
nalling driven by pregnancy: a review, Antioxidants
jappl.2001.91.5.2255.
(Basel), 11, 585, doi: 10.3390/antiox11030585.
68.
De Grotthuss, C.J.T. (1806) On the decomposition of
77.
Holland, O., Dekker Nitert, M., Gallo, L. A.,
water and the bodies it holds in solution using galvanic
Vejzovic, M., Fisher, J. J., and Perkins, A. V. (2017)
electricity [in French], Ann. Chim. (Paris), 58, 54-73.
Placental mitochondrial function and structure in
69.
Morelli, A. M., Ravera, S., Calzia, D., and Panfoli, I.
gestational disorders, Placenta, 54, 2-9, doi: 10.1016/
(2019) An update of the chemiosmotic theory as sug-
j.placenta.2016.12.012.
gested by possible proton currents inside the cou-
78.
Salazar-Petres, E., Pereira-Carvalho, D., Lopez-
pling membrane, Open Biol., 9, 180221, doi: 10.1098/
Tello, J., and Sferruzzi-Perri, A. N. (2022) Placental
rsob.180221.
structure, function, and mitochondrial phenotype
70.
Mannaerts, D., Faes, E., Cos, P., Briedé, J. J.,
relate to fetal size in each fetal sex in mice, Biol.
Gyselaers, W., Cornette, J., Gorbanev, Y., Bogaerts, A.,
Reprod., 106, 1292-1311, doi: 10.1093/biolre/ioac056.
Spaanderman, M., Van Craenenbroeck, E., and
79.
Vangrieken, P., Al-Nasiry, S., Bast, A., Leermakers,
Jacquemyn, Y. (2018) Oxidative stress in healthy preg-
P. A., Tulen, C. B. M., Schiffers, P. M. H., van
nancy and preeclampsia is linked to chronic inflam-
Schooten, F. J., and Remels, A. H. V. (2021) Placental
mation, iron status and vascular function, PLoS One,
mitochondrial abnormalities in preeclampsia, Reprod.
13, e0202919, doi: 10.1371/journal.pone.0202919.
Sci., 28, 2186-2199, doi: 10.1007/s43032-021-00464-y.
71.
Toboła-Wróbel, K., Pietryga, M., Dydowicz, P., Na-
80.
Vangrieken, P., Al-Nasiry, S., Bast, A., Leermakers,
pierała, M., Brązert, J., and Florek, E. (2020) Asso-
P. A., Tulen, C. B. M., Janssen, G. M. J., Kaminski, I.,
ciation of oxidative stress on pregnancy, Oxid. Med.
Geomini, I., Lemmens, T., Schiffers, P. M. H., van
Cell Longev.,
2020,
6398520, doi:
10.1155/2020/
Schooten, F. J., and Remels, A. H. V. (2012) Hypoxia-
6398520.
induced mitochondrial abnormalities in cells of the
72.
Mert, I., Oruc, A. S., Yuksel, S., Cakar, E. S.,
placenta, PLoS One, 16, e0245155, doi: 10.1371/
Buyukkagnıcı, U., Karaer, A., and Danısman, N.
journal.pone.0245155.
(2012) Role of oxidative stress in preeclampsia and
81.
Bartho, L. A., Fisher, J. J., Cuffe, J. S. M., and
intrauterine growth restriction, J. Obstet. Gynae-
Perkins, A. V. (2020) Mitochondrial transformations
col. Res.,
38,
658-664, doi:
10.1111/j.1447-0756.
in the aging human placenta, Am. J. Physiol. Endocri-
2011.01771.x.
nol. Metab., 319, E981-E994, doi: 10.1152/ajpendo.
73.
Rashid, C., Bansal, A., and Simmons, R. A. (2018)
00354.2020.
Oxidative stress, intrauterine growth restriction,
82.
Kolac, U. K., Kurek Eken, M., Ünübol, M., Donmez
and developmental programming of type 2 diabe-
Yalcin, G., and Yalcin, A. (2021) The effect of ges-
tes, Physiology, 33, 348-359, doi: 10.1152/physiol.
tational diabetes on the expression of mitochondri-
00023.2018.
al fusion proteins in placental tissue, Placenta, 115,
74.
Lyall, F., Robson, S. C., and Bulmer, J. N. (2013)
106-114, doi: 10.1016/j.placenta.2021.09.015.
Spiral artery remodeling and trophoblast invasion in
83.
McCarthy, C., and Kenny, L. C. (2016) Therapeutically
preeclampsia and fetal growth restriction: relationship
targeting mitochondrial redox signalling alleviates
to clinical outcome, Hypertension, 62, 1046-1054,
endothelial dysfunction in preeclampsia, Sci. Rep., 6,
doi: 10.1161/HYPERTENSIONAHA.113.01892.
32683, doi: 10.1038/srep32683.
MITOCHONDRIAL NETWORK: ELECTRIC CABLE AND MORE
Review
P. A. Abramicheva1, N. V. Andrianova1, V. A. Babenko1,2, L. D. Zorova1,2, S. D. Zorov1,3,
I. B. Pevzner1,2, V. A. Popkov1,2, D. S. Semenovich1, E. I. Yakupova1, D. N. Silachev1,2,
E. Y. Plotnikov1,2, G. T. Sukhikh2, and D. B. Zorov1,2*
БИОХИМИЯ том 88 вып. 10 2023
МИТОХОНДРИАЛЬНАЯ СЕТЬ
1939
1 Belozersky Institute of Physico-Chemical Biology, Lomonosov Moscow State University,
119991 Moscow, Russia; e-mail: zorov@belozersky.msu.ru
2 Kulakov National Medical Research Center of Obstetrics, Gynecology and Perinatology,
117997 Moscow, Russia
3 Faculty of Bioengineering and Bioinformatics, Lomonosov Moscow State University,
119991 Moscow, Russia
Mitochondria in a cell can unite and organize complex extended structures that extend over the entire
cellular volume, ensuring a uniform supply of energy to cellular compartments with ATP synthesized in
the mitochondria. In accordance with the chemiosmotic concept, the energy of oxidation of respiratory
substrates is largely stored in the form of an electrical potential difference on the inner membrane of
mitochondria. The theory of the functioning of extended mitochondrial structures as intracellular elec-
trical wires suggests that mitochondria provide the fast delivery of electrical energy through the cellular
volume, with following use of this energy for ATP synthesis, thereby speeding up the process of ATP
delivery compared to the rather slow diffusion of ATP in the cell. The analytical review gives the histo-
ry of the cable theory, solved and unsolved critical problems, the lability of the mitochondrial network
and the role of oxidative stress in this process. In addition to the already proven functioning of extended
mitochondrial structures as electrical cables, a few of additional functions are proposed, in particular,
the hypothesis is put forward that mitochondrial networks provide a uniform distribution of redox poten-
tial throughout the cell, which can change in pathological conditions during fragmentation of the mito-
chondrial reticulum. A few of pathologies accompanied by a violation of the redox status and the partici-
pation of mitochondria in them are considered.
Keywords: mitochondria, reticulum, network, electricity, membrane potential, fragmentation, fission, cardiomyocytes,
spermatozoa, oxidative stress, redox, preeclampsia, fetal growth retardation
БИОХИМИЯ том 88 вып. 10 2023
15*