БИОХИМИЯ, 2023, том 88, вып. 3, с. 429 - 442
УДК 612.82
ИЗМЕНЕНИЕ СВОЙСТВ ГЛУТАМАТЕРГИЧЕСКОЙ СИСТЕМЫ
ГИППОКАМПА КРЫС В ЛИТИЙ-ПИЛОКАРПИНОВОЙ
МОДЕЛИ ВИСОЧНОЙ ЭПИЛЕПСИИ
© 2023 Г.П. Диеспиров, Т.Ю. Постникова, А.В. Грифлюк,
А.А. Коваленко, А.В. Зайцев*
Институт эволюционной физиологии и биохимии им. И. М. Сеченова РАН,
194223 Санкт-Петербург, Россия; электронная почта: aleksey_zaitsev@mail.ru
Поступила в редакцию 09.11.2022
После доработки 07.01.2023
Принята к публикации 10.01.2023
Эпилептический статус (ЭС) запускает множество патологических изменений в нервной системе,
которые еще не до конца изучены и могут вести к развитию эпилепсии. В этой работе мы изучили
эффекты действия ЭС на характеристики возбуждающей глутаматергической передачи в гиппо-
кампе у крыс в литий-пилокарпиновой модели височной эпилепсии. Исследования проводились
через 1 день (острая фаза модели), 3 и 7 дней (латентная фаза) и 30-80 дней (хроническая фаза)
после ЭС. Используя метод ОТ-ПЦР в реальном времени, мы обнаружили, что в латентную фазу
происходит снижение экспрессии генов субъединиц GluA1 и GluA2 AMPA-рецепторов, что может
также сопровождаться повышением доли кальций-проницаемых AMPA-рецепторов, играющих
существенную роль в патогенезе многих заболеваний ЦНС. В переживающих срезах мозга при
регистрации полевых ответов в области CA1 гиппокампа в ответ на стимуляцию коллатералей
Шаффера электрическим током разной силы мы выявили снижение эффективности возбуждаю-
щей синаптической нейротрансмиссии во всех фазах модели. Однако в хронической фазе обна-
ружили увеличение частоты спонтанных возбуждающих постсинаптических потенциалов, что
свидетельствует о повышенной фоновой активности глутаматергической системы при эпилепсии.
В пользу этого также указывает снижение порога экстензии задних конечностей в тесте макси-
мального электрошока у крыс с височной эпилепсией по сравнению с контрольными животными.
Полученные результаты свидетельствуют о наличии ряда функциональных изменений в глутамат-
ергической системе, связанных с эпилептизацией мозга. Эти данные могут быть использованы для
разработки антиэпилептогенной терапии.
КЛЮЧЕВЫЕ СЛОВА: литий-пилокарпиновая модель эпилепсии, синапс, AMPA-рецептор, гиппокамп,
полевой постсинаптический потенциал, возбуждающий постсинаптический потенциал, тест максималь-
ного электрошока.
DOI: 10.31857/S0320972523030053, EDN: QWLEGQ
ВВЕДЕНИЕ
травма, длительные фебрильные судороги, ин-
фекционное заболевание ЦНС, инсульт [3-5],
Височная эпилепсия (ВЭ) является одним
запускающие эпилептогенез. Эпилептогенез
из распространенных неврологических рас-
представляет собой каскад функциональных
стройств, и почти в 30% от всех случаев ВЭ не
и структурных изменений в ткани мозга, при-
поддается фармакологическому лечению [1],
водящий к развитию спонтанных рецидиви-
что ухудшает качество жизни пациентов [2].
рующих судорог (СРС) [6, 7]. Наиболее часто
ВЭ может развиваться после однократного по-
при ВЭ страдает гиппокамп [8], который иг-
вреждающего воздействия на мозг, такого как
рает важную роль в формировании памяти [9],
эпилептический статус (ЭС), черепномозговая поэтому для ВЭ характерны коморбидные
Принятые сокращения: ВПСП - возбуждающий постсинаптический потенциал; ВЭ - височная эпилепсия;
МЭШ - максимальный электрошок; пПСП - полевой постсинаптический потенциал; ПС - популяционный спайк;
прПС - пресинаптический популяционный спайк; СРС - спонтанные рецидивирующие судороги; ЭС - эпилеп-
тический статус.
* Адресат для корреспонденции.
429
430
ДИЕСПИРОВ и др.
когнитивные нарушения [10, 11]. Несмотря
МАТЕРИАЛЫ И МЕТОДЫ
на то что механизмы эпилептогенеза активно
изучаются, до сих пор полная последователь-
Животные. Исследование проводили на
ность событий остается неясной, а надежных
самцах крыс Вистар. Животных содержали в
терапевтических подходов, предотвращающих
виварии при комнатной температуре, 12-ча-
эпилептогенез, пока не разработано [6, 12].
совом цикле день/ночь и свободном доступе
Для изучения эпилептогенеза и приобре-
к пище и воде.
тенной ВЭ используется моделирование на жи-
Литий-пилокарпиновая модель височной
вотных. Литий-пилокарпиновая модель ВЭ у
эпилепсии у крыс. Крысам в возрасте трех
крыс является общепринятой моделью, т.к. по-
недель внутрибрюшинно (в/б) вводили рас-
вторяет все особенности болезни человека [13].
твор LiCl (127 мг/кг; «Sigma-Aldrich», США),
Основными признаками этой модели явля-
через сутки в/б вводили раствор нитрата ме-
ются: наличие длительного ЭС; острая фаза с
тилскополамина (0,5 мг/кг; «Sigma-Aldrich»),
судорожным состоянием длительностью до су-
еще через 40-50 мин в/б вводили пилокар-
ток; латентная фаза длительностью 3-4 недели
пин (30 мг/кг; «Sigma-Aldrich»), который при-
(фаза эпилептогенеза); хроническая фаза, в
водил к ЭС. Контрольным крысам вводили те
которой у животных развиваются СРС [13, 14].
же препараты, за исключением пилокарпина,
При эпилептизации ткани мозга проис-
который заменяли на 0,9%-ный раствор NaCl.
ходят морфологические и функциональные
Тяжесть судорог оценивали по модифи-
изменения в организации синаптических кон-
цированной шкале лимбических судорог Ра-
тактов
[15]. Например, выявлено разраста-
сина [28]: 1 балл - подергивание челюстей и
ние мшистых волокон и формирование аббе-
лицевых мышц; 2 балла - усиление лицевого
рантных синаптических контактов в зубчатой
клонуса с переходом на мышцы туловища;
извилине и области CA3 гиппокампа [16, 17].
3 балла - клонические сокращения передних
Ранее нами было показано снижение уровня
лап; 4 балла - спячивание (подъем на задние
долговременной синаптической потенциации
лапы с опорой на лапы и хвост); 5 баллов -
в поле CA1 гиппокампа в латентную и хрони-
спячивание и падение на бок с клонически-
ческую фазы литий-пилокарпиновой моде-
ми судорогами конечностей, генерализацией
ли [18-20].
приступа и переходом в тонические судороги
После эпилептических судорог нередко
с полной потерей позы. Судороги с оценкой
нарушается синаптическая передача, вклю-
4 балла и выше, продолжающиеся более 15 мин,
чая изменение вероятности выброса медиато-
оценивали как ЭС. Как правило, через 30 мин
ра [21], числа и плотности глутаматных рецеп-
после введения пилокарпина у большинства
торов на постсинаптической мембране [22, 23],
животных развивались судороги, достигающие
перестройку их субъединичного состава [24-
4 и более баллов. В эксперименты отбирались
27]. Данные, полученные в разных лаборато-
только крысы, у которых судороги от 4 баллов
риях и/или на разных моделях эпилепсии, и
и выше продолжались не менее 90 мин. Тяже-
клинические данные часто являются противо-
лые тонико-клонические судороги, вызванные
речивыми (для обзора см. Zaitsev et al. [15]).
пилокарпином, приводят к высокой смертно-
Это свидетельствует о том, что ЭС может запу-
сти у животных, на уровне ~50%.
скать различные компенсаторные механизмы
Литий-пилокарпиновая модель условно
в нервной системе. В некоторых случаях эти
разделяется на 3 фазы, в каждую из которых
механизмы позволяют предотвратить развитие
были проведены экспериментальные иссле-
эпилепсии, а в других - болезнь прогрессиру-
дования: острая фаза - 1-й день; латентная
ет. Понимание того, какие именно изменения
фаза (эпилептогенез без внешних проявле-
в синапсах происходят после эпилептической
ний) - 3-й и 7-й дни; хроническая фаза (по-
активности, может лечь в основу рационально
явление СРС) наступает через
3-4 недели
обоснованной терапии предотвращения эпи-
после ЭС. В ходе данной работы не проводи-
лептогенеза и развития эпилепсии.
лось отдельных экспериментов по регистра-
В данной работе мы изучили изменения
ции СРС у крыс. Исследование свойств базо-
свойств синаптической передачи нейронов
вой нейротрансмиссии и кратковременной
поля CA1 гиппокампа, уровень экспрессии
пластичности проводили через
30-35 дней
генов субъединиц AMPA-рецепторов в дор-
после ЭС, регистрацию спонтанных синапти-
зальном гиппокампе, а также предрасполо-
ческих событий - через 40-50 дней, биохими-
женность к формированию судорожных со-
ческие исследования - через 60 дней, порог
стояний у крыс в различные периоды времени
развития судорог у крыс - через 70-80 дней.
после пилокарпин-индуцированного ЭС.
Число крыс, использованных в экспериментах,
БИОХИМИЯ том 88 вып. 3 2023
ГЛУТАМАТЕРГИЧЕСКАЯ СИСТЕМА ГИППОКАМПА В МОДЕЛИ ВЭ
431
Таблица 1. Число животных, использованных в экспериментах
Электрофизиологические эксперименты
Фаза
Группа
ОТ-ПЦР
МЭШ
Полевые отведения
Патч-кламп
Контроль
10
8
-
-
Острая фаза
ВЭ
9
14
-
-
Контроль
11
6
-
-
Латентная фаза
ВЭ
6
23
-
-
Контроль
12
10
6
15
Хроническая фаза
ВЭ
7
12
4
21
Примечание. МЭШ - метод максимального электрошока.
указано в табл. 1. Одни и те же животные могли
ли от лучистого слоя поля CA1 гиппокампа
быть использованы в разных экспериментах,
крыс при помощи боросиликатного стеклян-
если это было методически возможно.
ного микроэлектрода (0,2-1,0 Мом; «World
Электрофизиологические эксперименты и
Precision Instruments», США) (рис. 1, а).
анализ результатов. Крысу анестезировали при
Срезы стимулировали парными прямо-
помощи изофлурана («Laboratorios Karizoo»,
угольными импульсами каждые
20 с (дли-
Испания), декапитировали, быстро извлекали
тельность импульса - 0,1 мс; межстимульный
мозг и изготавливали горизонтальные пере-
интервал - 50 мс) с помощью биполярного
живающие срезы мозга, содержащие дорзаль-
нихромового электрода на границе полей
ный гиппокамп, толщиной 350 мкм на вибра-
CA3-CA1 стимулятором А365 («World Precision
томе Microm International HM 650V («Microm
Instruments»). Полевые ПСП регистрировали
International», Германия) в охлажденной до 0 °С
с помощью усилителя Model 1800 («AM Sys-
и аэрируемой карбогеном (95% O2/5% СO2)
tems», США), оцифровывали на ЦАП/АЦП
искусственной спинномозговой жидкости сле-
NI USB-6211 («National Instruments», США) с
дующего состава (в мМ): 126 NaCl, 2,5 KCl,
использованием программного обеспечения
1,25 NaH2PO4, 1 MgSO4, 2 CaCl2, 24 NaHCO3,
WinWCP v5.x («University of Strathclyde», Вели-
10 глюкоза. После изготовления срезы поме-
кобритания).
щали в водяную баню при 35 °С на 60 мин.
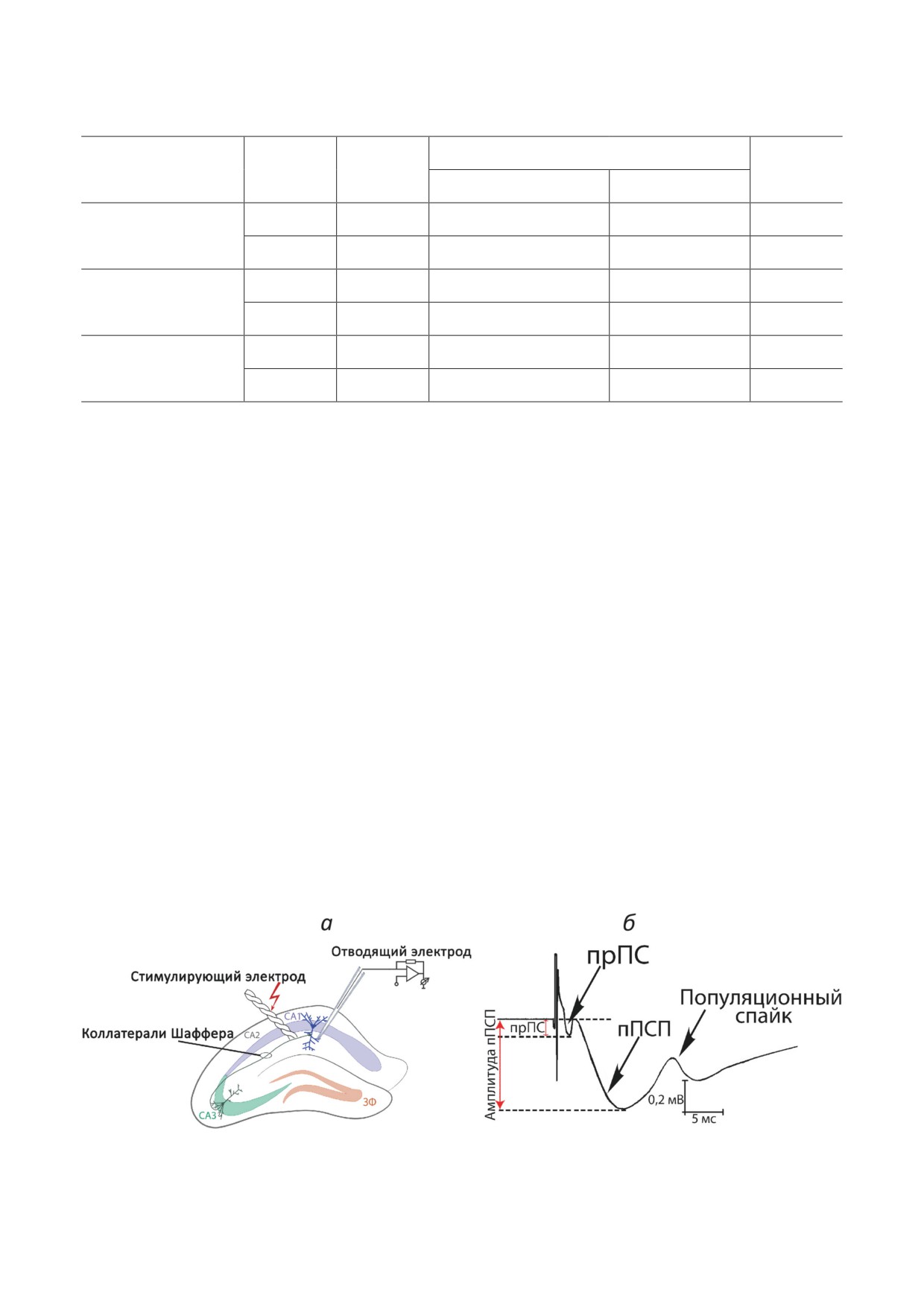
Для каждого среза рассчитывали амплиту-
От одного животного в эксперименте исполь-
ду пресинаптического популяционного спайка
зовали обычно 1-3 среза.
(прПС, fiber volley) и пПСП (рис. 1, б). Изме-
Эксперименты проводили при темпера-
рение соотношения вход/выход (input/output)
туре 30 °С. Регистрацию полевых постсинап-
проводили при помощи стимуляции с возра-
тических потенциалов (пПСП) осуществля-
стающей силой тока в диапазоне 25-200 мкА
Рис. 1. Методика регистрации полевых потенциалов. а - Схема расположения электродов в срезах гиппокампа.
б - Схема регистрируемого полевого ответа с указанием регистрируемых параметров; прПС - пресинаптический по-
пуляционный спайк; пПСП - полевой постсинаптический потенциал
БИОХИМИЯ том 88 вып. 3 2023
432
ДИЕСПИРОВ и др.
с шагом в 25 мкА. Строили зависимость вход/вы-
Полученные образцы кДНК разбавляли в 10 раз
ход, которая представляет собой передаточную
перед проведением ПЦР. ПЦР проводили в
функцию между пресинаптическими потенциа-
общем объеме 6 мкл с 0,8 мкл кДНК. Для ре-
лами действия и ответом на постсинаптических
акции использовали 0,5 ед. TaqM-полимеразы
мембранах. Эффективность синаптической ней-
(«Алкор-Био», Россия). Специфические зонды,
ротрансмиссии определяли по максимальному
прямые и обратные праймеры («ДНК-Син-
наклону кривой вход/выход, который находили
тез») приведены в табл. 2. ПЦР в реальном
путем аппроксимации функцией Гомперца [29].
времени проводили на приборе CFX384 Real-
Оценку величины кратковременной си-
Time System («Bio-Rad», США).
наптической пластичности проводили мето-
ПЦР для генов домашнего хозяйства были
дом парной стимуляции с возрастающими
проведены в мультиплексах (Actb + Gapdh +
межстимульными интервалами 30, 40, 50, 60,
+ B2m, Rpl13a + Ppia + Sdha и Hprt1 + Pgk1 +
70, 80, 90, 100, 150, 200, 300, 400, 500 мс. Вели-
+ Ywhaz) в соответствии с протоколом, описан-
чину пластичности рассчитывали, как отно-
ным ранее [30]. Анализ, выполненный с ис-
шение амплитуды второго пПСП к первому
пользованием онлайн-инструмента RefFinder
для каждого интервала стимулов.
Запись спонтанных возбуждающих пост-
что из перечисленных генов домашнего хозяй-
синаптических потенциалов (ВПСП) на пира-
ства Pgk1, Ywhaz и Gapdh экспрессируются
мидных нейронах гиппокампа производили
наиболее стабильно. Относительная экспрес-
методом патч-клампа в режиме «целая клетка»
сия генов Gria1 и Gria2 была подсчитана с
при потенциале покоя в течение 10 мин. Исполь-
помощью метода 2-ΔΔCt [31]. Нормализацию
зовали стеклянные микроэлектроды с входным
данных проводили с использованием сред-
сопротивлением 3-5 МОм. Электрод заполня-
него геометрического значений Ct генов Pgk1,
ли раствором следующего состава (в мМ): 114 глю-
Ywhaz и Gapdh.
конат калия; 6 KCl; 10 HEPES (4-(2-гидрокси-
Метод максимального электрошока (МЭШ).
этил)-1-пиперазинэтансульфоновая кислота);
Предрасположенность к судорогам у живот-
0,2 EGTA (этиленгликоль-бис(β-аминоэтило-
ных исследовали с помощью МЭШ через
вый эфир)-N,N,N′,N′-тетрауксусная кислота);
2,5 месяца после ЭС. Ток подавался через
4 ATP-Mg; 0,3 GTP. Поиск и анализ ВПСП
ушные электроды с использованием генера-
проводился автоматически при помощи пред-
тора импульсов ECT Unit 57800 («Ugo Basile»,
варительно созданного шаблона в программе
Италия). Использовали силу тока в диапазо-
Clampfit 10.4.2.0 (Molecular Devices), ошибоч-
не 12-100 мА с интервалами 0,1 логарифми-
но отобранные программой ВПСП исключали
ческой шкалы, частота импульсов - 200 Гц,
при визуальной оценке результатов.
длительность одиночного импульса - 0,9 мс,
ОТ-ПЦР в реальном времени. Индукцию
длительность подачи тока - 0,8 с. У каждого
судорог вызывали у крыс в возрасте 6-7 недель.
животного определяли минимальное значение
Для анализа экспрессии генов, кодирующих
тока, при котором наблюдались тонические
GluA1 (Gria1) и GluA2 (Gria2) субъединицы
судороги с экстензией задних конечностей.
AMPA-рецепторов, крыс декапитировали на 3,
В первый день животное получало электро-
7 и 60 день после индукции судорог. Выделение
шок средней силы (40 мА). Если тонические
дорзальной области гиппокампа было выпол-
судороги с экстензией задних конечностей не
нено при помощи замораживающего микрото-
развивались, то силу току увеличивали. Если
ма OTF5000 («Bright Instruments», Великобри-
при 40 мА тонические судороги развивались,
тания). Тотальную РНК из образцов выделяли
то силу тока уменьшали. Повторные экспери-
с помощью реагента ExtractRNA («Евроген»,
менты проводили через 2-3 дня.
Россия). Перед обратной транскрипцией образ-
Статистическая обработка данных. Досто-
цы РНК обрабатывали RQ1 DNAse («Promega»,
верность различий средних в двух выборках
США). Затем РНК осаждали с помощью рас-
оценивали с использованием t-критерия Стью-
твора LiCl. Концентрацию РНК измеряли на
дента. Влияние силы стимулирующего тока на
спектрофотометре NanoDrop Lite
(«Thermo
величину полевых ответов, а также сравнение
Fisher Scientific», США).
пПСП в разных группах проверяли с помощью
кДНК синтезировали из
2 мкг тоталь-
дисперсионного анализа с повторными изме-
ной РНК с использованием олиго-dT (1 мкг),
рениями с последующим использованием апо-
9-мерных случайных
(0,5 мкг) праймеров
стериорного критерия Тьюки. Для сравнения
(«ДНК-Синтез», Россия) и обратной тран-
относительной экспрессии генов использовали
скриптазы M-MLV («Promega»). Реакцию про-
двухфакторный дисперсионный анализ с апо-
водили согласно инструкции производителя.
стериорным тестом Сидака. Данные в тексте
БИОХИМИЯ том 88 вып. 3 2023
ГЛУТАМАТЕРГИЧЕСКАЯ СИСТЕМА ГИППОКАМПА В МОДЕЛИ ВЭ
433
Таблица 2. Последовательности праймеров и зондов для ОТ-ПЦР в реальном времени
Ген
Последовательности праймеров и зондов (5′→3′)
Ссылка
F: TGTCACCAACTGGGACGATA
Actb
[32] (праймеры);
R: GGGGTGTTGAAGGTCTCAAA
NM_031144
[30] (зонд)
Зонд: FAM-CGTGTGGCCCCTGAGGAGCAC-BHQ1
F: TGCACCACCAACTGCTTAG
Gapdh
R: GGATGCAGGGATGATGTTC
[33]
NM_017008
Зонд: R6G-ATCACGCCACAGCTTTCCAGAGGG-BHQ2
F: TGCCATTCAGAAAACTCCCC
B2m
R: GAGGAAGTTGGGCTTCCCATT
[34]
NM_012512
Зонд: ROX-ATTCAAGTGTACTCTCGCCATCCACCG-BHQ1
F: GGATCCCTCCACCCTATGACA
Rpl13a
[35] (праймеры);
R: CTGGTACTTCCACCCGACCTC
NM_173340
[30] (зонд)
Зонд: FAM-CTGCCCTCAAGGTTGTGCGGCT-BHQ1
F: AGACGTTTGACAGGGGAATG
Sdha
[36] (праймеры);
R: TCATCAATCCGCACCTTGTA
NM_130428
[30] (зонд)
Зонд: R6G-ACCTGGTGGAGACGCTGGAGCT-BHQ2
F: AGGATTCATGTGCCAGGGTG
Ppia
R: CTCAGTCTTGGCAGTGCAGA
[26]
NM_017101
Зонд: ROX-CACGCCATAATGGCACTGGTGGCA-BHQ1
F: TCCTCAGACCGCTTTTCCCGC
Hprt1
[37] (праймеры);
R: TCATCATCACTAATCACGACGCTGG
NM_012583
[30] (зонд)
Зонд: FAM-CCGACCGGTTCTGTCATGTCGACCCT-BHQ1
F: ATGCAAAGACTGGCCAAGCTAC
Pgk1
[38] (праймеры);
R: AGCCACAGCCTCAGCATATTTC
NM_053291
[30] (зонд)
Зонд: R6G-TGCTGGCTGGATGGGCTTGGA-BHQ2
F: GATGAAGCCATTGCTGAACTTG
Ywhaz
[38] (праймеры);
R: GTCTCCTTGGGTATCCGATGTC
NM_013011
[30] (зонд)
Зонд: ROX-TGAAGAGTCGTACAAAGACAGCACGC-BHQ1
F: TCAGAACGCCTCAACGCC
Gria1
R: TGTAGTGGTACCCGATGCCA
[39]
NM_031608
Зонд: ROX-TCCTGGGCCAGATCGTGAAGCTAGAAAA-BHQ1
F: CAGTGCATTTCGGGTAGGGA
Gria2
R: TGCGAAACTGTTGGCTACCT
[39]
NM_017261
Зонд: FAM-TCGGAGTTCAGACTGACACCCCA-BHQ1
и на рисунках приведены как среднее со стан-
Относительная экспрессия генов субъеди-
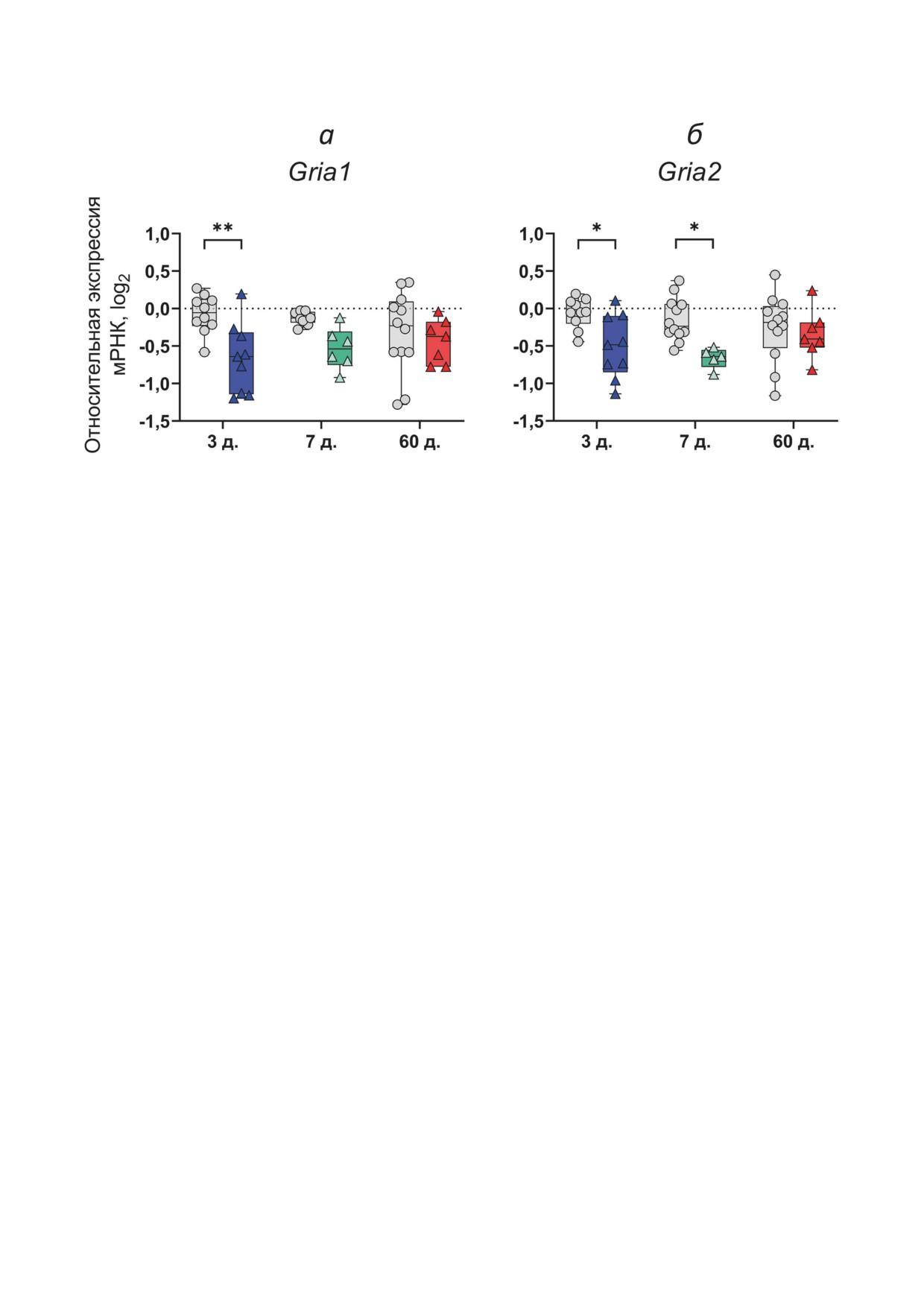
дартной ошибкой среднего, если не указано
ниц AMPA-рецепторов снижена в латентной,
обратного.
но не в хронической фазе. Так как амплитуда
синаптических возбуждающих ответов в ос-
новном определяется AMPA-рецепторами, то
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
мы проанализировали изменяется ли уровень
экспрессии генов, кодирующих GluA1 (Gria1)
Возможные изменения характеристик глу-
и GluA2 (Gria2) субъединицы AMPA-рецепто-
таматергической передачи в гиппокампе крыс
ров (рис. 2).
были проанализированы на нескольких струк-
Мы выявили снижение экспрессии обо-
турных уровнях: молекулярном (экспрессия
их генов на 3-й день после ЭС. Продукция
генов субъединиц AMPA-рецепторов), кле-
мРНК Gria2 остается сниженной до 7-го дня
точном (исследование синаптической нейро-
после пилокарпин-индуцированных судорог,
трансмиссии и вероятности выброса глутамата
но не отличается от контрольных значений
клетками) и на целом животном (МЭШ-тест).
в хроническую фазу (рис. 2, б). Изменений
6
БИОХИМИЯ том 88 вып. 3 2023
434
ДИЕСПИРОВ и др.
Рис. 2. Относительная экспрессия генов Gria1 (а) и Gria2 (б) в дорзальной области гиппокампа крыс. Исследования
проводились на 3-й и 7-й дни после индукции ЭС (латентная фаза модели), а также на 60-й день (хроническая фаза
модели). Контрольная группа показана серым цветом. Данные представлены в виде индивидуальных значений (кружки
и треугольники) с минимальным и максимальным значениями (планки ошибок), медианой выборки (горизонталь-
ная линия), а также первым и третьим квартилями (прямоугольники). Число животных в группах: 1) эпилептическая
группа: 3-й день (n = 9), 7-й день (n = 6), 60-й день (n = 7); 2) контрольная группа: 3-й день (n = 10), 7-й день (n = 11),
60-й день (n = 12); д. - количество дней после ЭС. Двухфакторный дисперсионный анализ: Gria1 - F1,47 (фактор судо-
рог) = 12,4, p = 0,001; Gria2 - F1,49 (фактор судорог) = 13,4, p < 0,001. Значимые различия между группами рассчитаны
с помощью апостериорного теста Сидака - * p < 0,05, ** p < 0,01
экспрессии Gria1 не выявлено ни на 7-й день,
нейротрансмиссии на
1-й день после ЭС
ни в хроническую фазу (рис. 2, а).
(t = 2,77; p < 0,01; рис. 3, г).
Глутаматергическая синаптическая нейро-
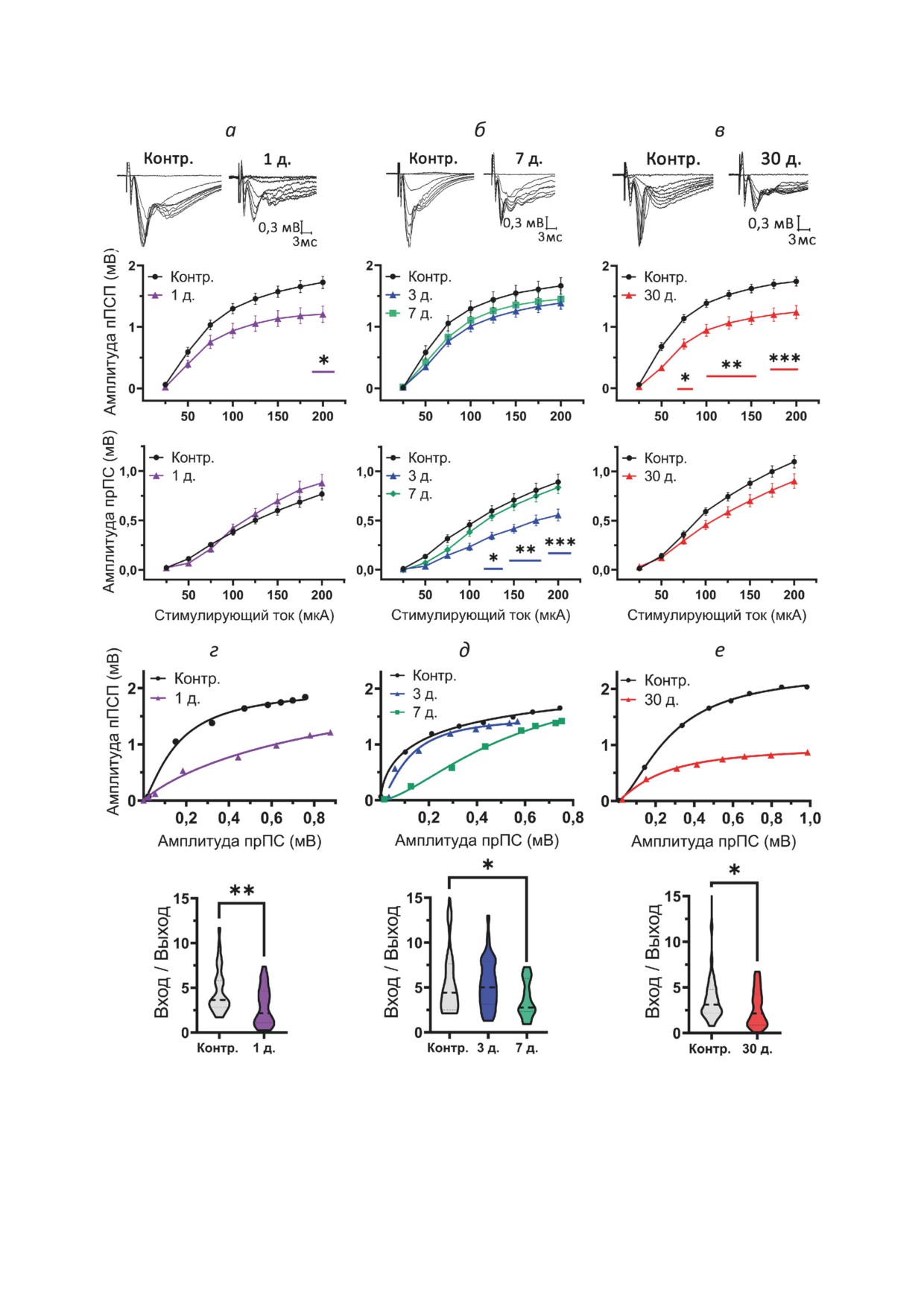
В латентной фазе модели не было выявлено
трансмиссия снижена в разные периоды литий-
отличий амплитуд пПСП ни на 3-й, ни на 7-й
пилокарпиновой модели. Патологические изме-
день (F2,602 = 1,70; p = 0,18; рис. 3, б). Однако
нения в нейронных связях гиппокампа ведут к
было обнаружено снижение прПС на 3-й день
изменениям базовой синаптической трансмис-
в интервале стимуляции 125-200 мкА по срав-
сии. Поэтому мы исследовали, как изменяется
нению с контрольной группой (F2,567 = 8,3;
базовый уровень возбуждающей (глутамат-
p < 0,001; рис. 3. б). Соотношение вход/выход
ергической) синаптической нейротрансмиссии
статистически значимо снижается только к
в синапсах CA3-CA1 пирамидных нейронов
7-му дню после перенесенного ЭС (F2,76 = 3,58;
гиппокампа у крыс в острую, латентную и хро-
p < 0,05; рис. 3, д).
ническую фазы литий-пилокарпиновой моде-
В хронической фазе наблюдается значитель-
ли ВЭ. Для этого мы построили зависимость
ное снижение амплитуды пПСП (F1,707 = 17,0;
амплитуды пПСП и прПС от силы внеклеточ-
p < 0,001; рис. 3, в) в широком диапазоне ин-
ной стимуляции, а также вход/выход-зависи-
тервалов стимуляции (75-200 мкА), однако
мости (рис. 3). Из-за повышенной возбудимо-
изменения амплитуды прПС, выявленные с
сти нейронных сетей у крыс в различные фазы
помощью дисперсионного анализа (F1,637 = 4,2;
после ЭС может изменяться не только ампли-
p < 0,05; рис. 3, в), не достигали уровня значи-
туда, но и форма пПСП, что согласуется с
мости при проверке апостериорными тестами.
результатами, полученными в модели эпилеп-
Также в этот период происходит значительное
тических судорог, вызванных пентилентетразо-
ослабление синаптической нейротрансмиссии
лом [29].
(t = 2,19; p < 0,05; рис. 3, е).
В острой фазе модели амплитуда пПСП
Вероятность выброса глутамата в синапсах
снижена по сравнению с контролем, особен-
области СА1 гиппокампа изменена в хронической
но при большой силе стимуляции (F1,448 = 7,5;
фазе модели эпилепсии. Изменение величины
p < 0,01; рис. 3, а). Изменений амплитуды прПС
синаптической нейротрансмиссии может быть
не было выявлено (F1,441 = 0,67; p = 0,42; рис 3, а).
связано с изменением вероятности выбро-
Анализ вход/выход-зависимости показал зна-
са глутамата в коллатералях Шаффера. Для
чительное снижение уровня синаптической
оценки возможных изменений вероятности
БИОХИМИЯ том 88 вып. 3 2023
ГЛУТАМАТЕРГИЧЕСКАЯ СИСТЕМА ГИППОКАМПА В МОДЕЛИ ВЭ
435
Рис. 3. Репрезентативные примеры записей (сверху), амплитуды пПСП и амплитуды прПС в ответ на стимуляцию (сни-
зу) (а-в). Примеры вход/выход-отношений между амплитудами пПСП и прПС, аппроксимированные сигмоидальной
функцией Гомперца (сверху), и сравнение их максимальных наклонов (снизу) (г-е). На графиках сравнения соот-
ношения вход/выход толстой пунктирной линией указаны медианы, тонкой пунктирной линией - квартили. Число
срезов в группах: 1) эпилептическая группа: 1-й день (n = 39), 3-й день (n = 41), 7-й день (n = 39), 30-й день (n = 41);
2) контрольная группа: 1-й день (n = 35), 7-й день (n = 24), 30-й день (n = 41). прПС - пресинаптический популяци-
онный спайк; Контр. - контроль; д. - количество дней после ЭС. * p < 0,05; ** p < 0,01; *** p < 0,001 (апостериорный
критерий Тьюки - для панелей а, б, в, д; t-тест - для панелей г и е)
БИОХИМИЯ том 88 вып. 3 2023
6*
436
ДИЕСПИРОВ и др.
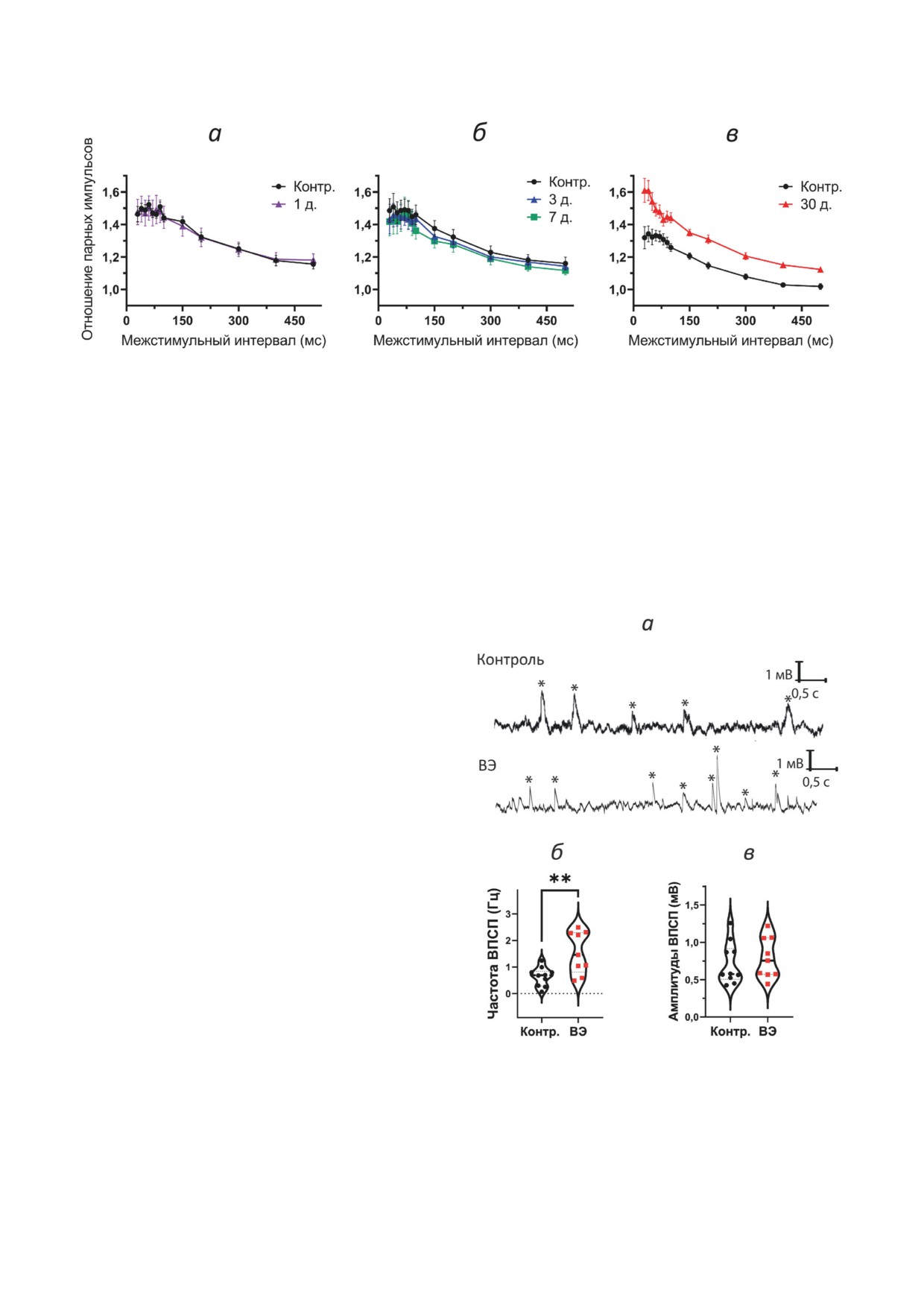
Рис. 4. Кратковременная синаптическая пластичность не изменяется в срезах гиппокампа крыс, перенесших ЭС.
а-в - Графики зависимости отношения парных импульсов от величины межстимульного интервала в острую,
латентную и хроническую фазы соответственно. Число срезов в группах: 1) эпилептическая группа: острая фаза - 1-й
день (n = 10); латентная фаза - 3-й день (n = 6), 7-й день (n = 10); хроническая фаза - 30-й день (n = 20); 2) контрольная
группа: 1-й день (n = 9), 7-й день (n = 7), 30-й день (n = 9); д. - количество дней после ЭС
выброса медиатора часто используют сравне-
Результаты этого эксперимента показыва-
ние отношений амплитуд синаптических отве-
ют, что вероятность выброса медиатора выше
тов при подаче парных стимулов [40]. Поэтому
в эпилептической группе, и это согласуется с
мы измерили отношения амплитуд ответов
данными, полученными другими авторами в
(величину кратковременной фасилитации) при
каинатной модели эпилепсии [21]. Однако в
подаче парных стимулов с разным интервалом
пилокарпиновой модели Owen et al. [41] наблю-
в диапазоне 30-500 мс.
дали увеличение частоты миниатюрных ВПСП
В острой и латентной фазах достоверных
различий выявлено не было (острая фаза -
F1,204 = 0,47; p = 0,93; рис. 4, а; латентная фаза -
F2,240 = 0,18; p = 0,84; рис. 4, б). В хронической
фазе дисперсионный анализ выявляет разли-
чия между контрольной и ВЭ-группами (F1,324 =
= 13,1; p < 0,01; рис. 4, в), однако апостериор-
ный тест Тьюки не подтвердил значимость раз-
личий ни для одного из временных интервалов.
При эпилепсии могут происходить изме-
нения в работе пресинапса [15], включая уве-
личение числа везикул, готовых к выбросу в
пресинаптических терминалях [21], что может
проявляться в увеличении вероятности спонтан-
ного выброса медиатора и увеличении коэффи-
циента фасилитации. Поэтому мы проанализи-
ровали частоту и амплитуду спонтанных ВПСП
в СА1 пирамидных нейронах гиппокампа в
хронической фазе модели. Мы регистрировали
спонтанную активность пирамидных нейронов
поля CA1 при потенциале покоя. На рис. 5, а
представлены репрезентативные примеры реги-
страций, на которых отмечены ВПСП. У ней-
ронов гиппокампа крыс с ВЭ частота ВПСП
была выше на 118%, чем в контрольной груп-
Рис. 5. Частота ВПСП снижена, а амплитуда остается
пе (t = 2,5; p = 0,02; рис. 5, б), однако ампли-
неизменной в пирамидных нейронах гиппокампа крыс
туда ВПСП не различалась (t = 0,60; p = 0,56;
с ВЭ. а - Репрезентативные записи ответов нейронов при
рис. 5, в) и составляла 0,72 ± 0,20 мВ у кон-
потенциале покоя, ВПСП отмечены звездочками. Часто-
трольных животных и 0,79 ± 0,21 мВ - у крыс
та (б) и амплитуда (в) спонтанных ВПСП в двух группах.
Число срезов в группах: 1) эпилептическая группа: 35-й
с ВЭ. Следует отметить, что на рис. 5, б для крыс
день (n = 9); 2) контрольная группа: 35-й день (n = 10);
экспериментальной группы 4 точки на частоте
Контр. - контроль; ВЭ - группа крыс с височной эпилеп-
больше 2 Гц получены от разных животных.
сией; t-тест - ** р < 0,01
БИОХИМИЯ том 88 вып. 3 2023
ГЛУТАМАТЕРГИЧЕСКАЯ СИСТЕМА ГИППОКАМПА В МОДЕЛИ ВЭ
437
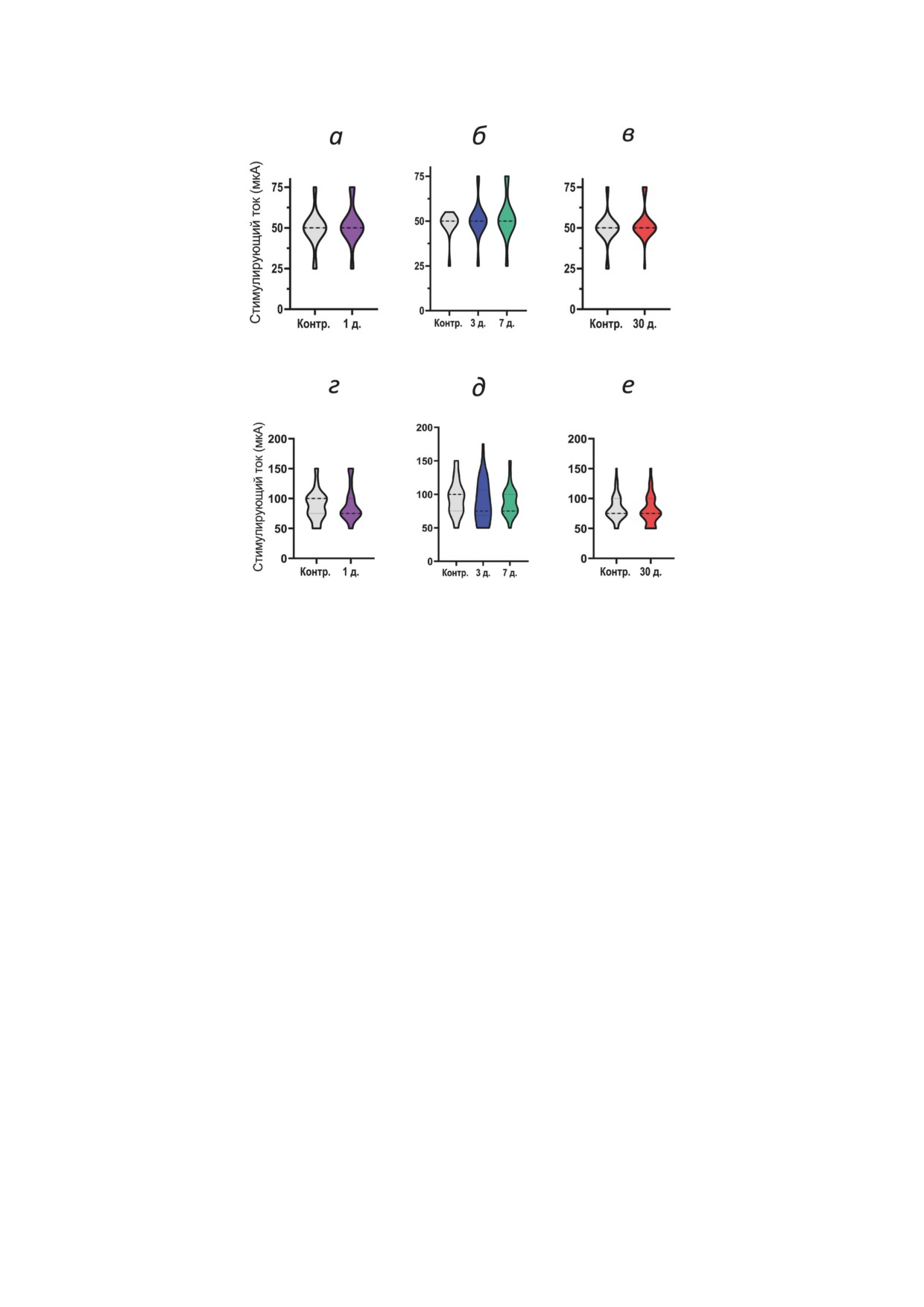
Рис. 6. Пороговые токи, вызывающие пПСП (а-в) и ПС (г-е) не изменяются в острой (а, г), латентной (б, д) и хро-
нической (в, е) фазах литий-пилокарпиновой модели ВЭ. Число срезов в группах: 1) эпилептическая группа: острая
фаза - 1-й день (n = 39); латентная фаза - 3-й день (n = 42), 7-й день (n = 39); хроническая фаза - 30-й день (n = 74);
2) контрольная группа: 1-й день (n = 35), 7-й день (n = 25), 30-й день (n = 59). Контр. - контроль; д. - число дней
после ЭС. Апостериорный критерий Тьюки использован для сравнения данных на панелях б, д; t-тест - для а, в, г и е
в более поздние сроки модели (через
80-
Таким образом, несмотря на снижение эффек-
110 дней), тогда как через 20-60 дней частота
тивности возбуждающей синаптической пере-
событий между контрольной и эпилептической
дачи, сила внеклеточной стимуляции, необ-
группой еще не различалась. Пороги поле-
ходимая для генерации потенциалов действия
вых постсинаптических потенциалов (пПСП)
в пирамидных нейронах области CA1, осталась
и популяционных спайков (ПС) не изменены
прежней.
у крыс с ВЭ.
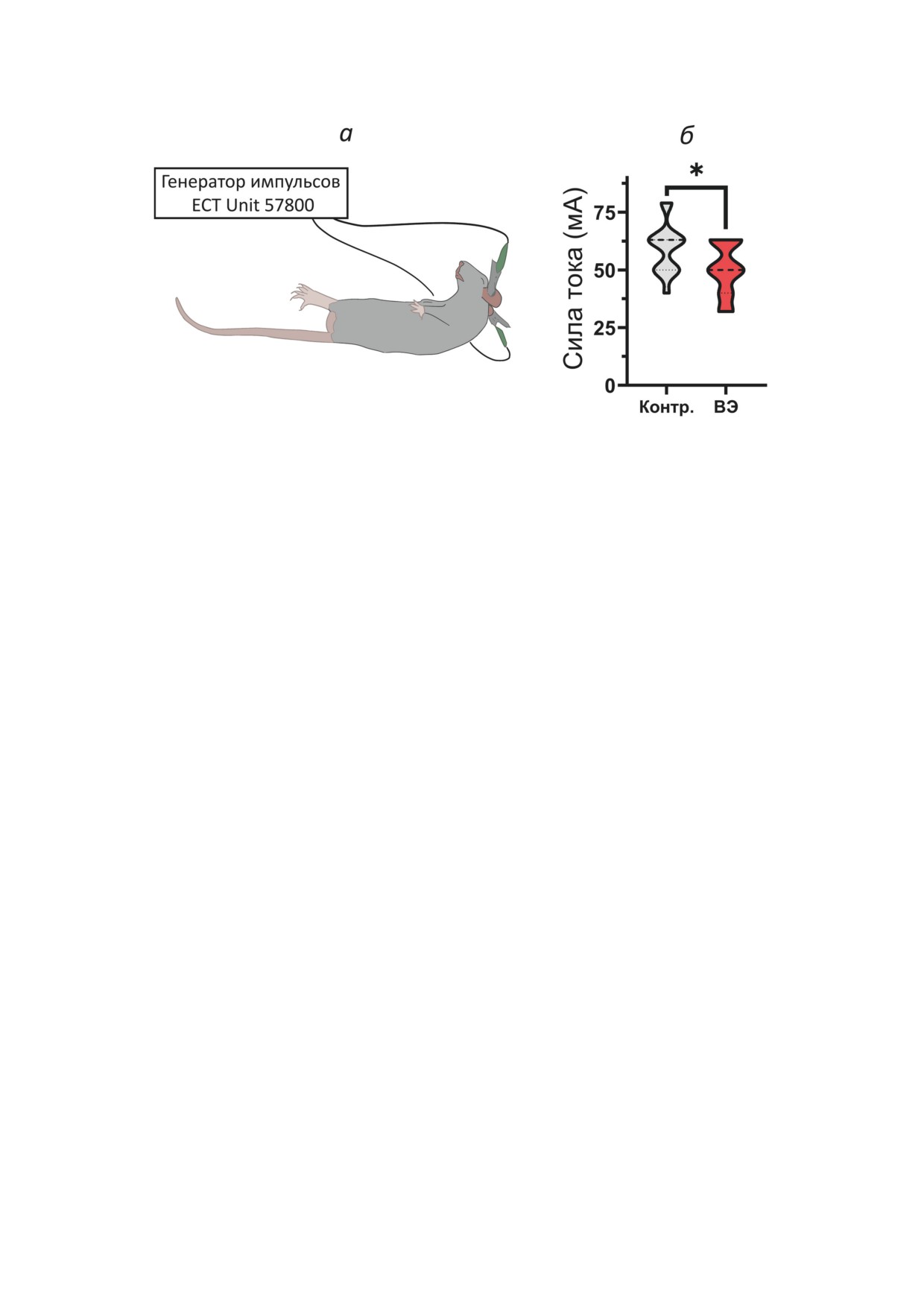
У крыс с височной эпилепсией снижен порог
При эпилепсии вероятность синхрониза-
развития экстензии задних конечностей в МЭШ-
ции и реверберации нервных импульсов в ней-
тесте. Для исследования предрасположенно-
ронных сетях возрастает. Соответственно, бо-
сти к судорогам провели тестирование МЭШ у
лее слабое синаптическое возбуждение может
крыс в хронической фазе модели и у контроль-
приводить к генерации потенциалов действия.
ных животных того же возраста (рис. 7, а).
Так как мы выявили ослабление возбуждаю-
У крыс с ВЭ выявили значительное снижение
щей синаптической передачи в гиппокампе
порога появления экстензии задних конечно-
экспериментальных крыс (рис. 3), то решили
стей (t = 2,4; p < 0,05; рис. 7, б).
проверить изменяются ли пороги возникно-
вения пПСП и ПС в нейронах СА1. Мы не
выявили достоверных различий для порогов
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
возникновения пПСП ни в одном из исследуе-
мых периодов (острая фаза - t = 0,97; p = 0,34;
Результаты наших экспериментов выяви-
рис. 6, а; латентная фаза - F2,102 = 1,11; p = 0,33;
ли ряд особенностей в функционировании
рис. 6, б; хроническая фаза - t = 1,83; p = 0,07;
глутаматергической передачи в гиппокампе
рис. 6, в); ни для порогов генерации ПС (ост-
крыс в разные фазы литий-пилокарпиновой
рая фаза - t = -0,19; p = 0,85; рис. 6, г; латент-
модели ВЭ по сравнению с контролем. Часть
ная фаза - F2,102 = 0,48; p = 0,62; рис. 6, д; хро-
изменений носит компенсаторный характер и
ническая фаза - t = 0,9; p = 0,37; рис. 6, е).
направлена на снижение уровня возбуждения
БИОХИМИЯ том 88 вып. 3 2023
438
ДИЕСПИРОВ и др.
Рис. 7. Метод МЭШ. а - Схематическое изображение крысы с экстензией задних конечностей (тонические судороги),
вызванные подачей электрического тока через ушные электроды. б - Пороговые токи, необходимые для экстензии
задних конечностей у крысы. Число животных в группах: 1) эпилептическая группа: 50-й день (n = 21); 2) контрольная
группа: 50-й день (n = 15). Контр. - контроль; ВЭ - группа крыс с ВЭ. Непарный t-тест - * р < 0,05
нейронных сетей. В частности, мы выявили
увеличению доли AMPA-рецепторов, лишен-
ослабление базового уровня синаптической пе-
ных этой субъединицы и имеющих особые
редачи в срезах гиппокампа крыс. Однако дру-
биофизические свойства и кальциевую про-
гие изменения, такие как повышение частоты
ницаемость [42]. В данной работе мы выявили
спонтанных ВПСП, могут свидетельствовать
снижение продукции мРНК генов субъединиц
о патологически высокой фоновой активации
AMPA-рецепторов Gria1 и Gria2 в дорзальной
глутаматергической системы. На уровне целого
области гиппокампа крыс в латентной фазе
организма это проявляется в снижении порога
литий-пилокарпиновой модели, что согласу-
возникновения судорог, вызванных МЭШ.
ется с ранее полученными нами результатами
В острую фазу модели нарушения синапти-
в коре мозга с помощью электрофизиологи-
ческой трансмиссии могут быть связаны с тем,
ческих методов [26]. Ослабление экспрессии
что после ЭС у части животных наблюдается
GluA2-субъединицы в гиппокампе и височной
почти полная гибель нейронов области СА1
коре ведет к увеличению глутамат-опосредо-
гиппокампа [18], и, скорее всего, именно эти
ванного проникновения ионов Ca2+ в клетку и
животные часто погибают в первые несколько
эксайтотоксичности, что, в свою очередь, мо-
суток после ЭС. С этим может быть связано
жет быть одним из факторов нейродегенера-
почти бимодальное распределение значений
ции и эпилептогенеза [24].
наклона функции вход/выход (рис. 3, г). При
Другой причиной снижения эффективно-
развитии эпилепсии (начиная с 7-го дня) эф-
сти синаптической передачи могут быть изме-
фективность возбуждающей синаптической
нения в пресинапсе. Для оценки вероятности
трансмиссии снижена, что может являться
выброса медиатора изучают кратковременную
компенсаторной реакцией, направленной на
синаптическую пластичность и частоту спон-
ослабление передачи возбуждения в нейрон-
танных ВПСП [40]. В острой и латентной фа-
ных сетях гиппокампа, либо это может быть
зах свойства кратковременной синаптической
проявлением продолжающейся нейродегене-
пластичности экспериментальных животных
рации в гиппокампе [19]. В предыдущих на-
практически не отличались от контроля. В хро-
ших работах мы обнаружили снижение уровня
нической фазе литий-пилокарпиновой модели
долговременной потенциации в поле CA1 в ла-
мы выявили тенденцию к ослаблению фасили-
тентную и хроническую фазы, что также может
тации и увеличение частоты ВПСП, что может
быть вызвано выявленным снижением возбу-
быть связано с увеличением числа везикул,
ждающей нейротрансмиссии [18, 20].
готовых к выбросу в пресинаптических терми-
Другим важным фактором, определяющим
налях, как было показано в каинатной модели
свойства возбуждающей синаптической пере-
эпилепсии [21]. Интересно, что в пентилен-
дачи, является число и субъединичный состав
тетразоловой модели, в которой, в отличии от
AMPA-рецепторов. Известно, что уменьшение
пилокарпиновой и каинатной модели, хрони-
экспрессии GluA2-субъединицы может вести к
ческой эпилепсии не развивается, наблюдается
БИОХИМИЯ том 88 вып. 3 2023
ГЛУТАМАТЕРГИЧЕСКАЯ СИСТЕМА ГИППОКАМПА В МОДЕЛИ ВЭ
439
снижение вероятности выброса глутамата пре-
туры мозга вовлечены в формирование и
синапсами, что может оказывать противоэпи-
поддержание судорог, поэтому нельзя исклю-
лептогенный эффект [29].
чать их вклад в снижение порога генерации
В ходе данной работы при использовании
судорог.
полевых отведений на переживающих срезах
Полученные нами результаты свидетель-
гиппокампа не было выявлено изменений в
ствуют о комплексе изменений в глутамат-
порогах генерации пПСП и ПС у эксперимен-
ергической системе гиппокампа после ЭС,
тальной группы животных. Однако у крыс с ВЭ
которые включают в себя изменения субъ-
в тесте МЭШ порог генерации тонических
единичного состава AMPA-рецепторов в ла-
судорог был значительно снижен по сравне-
тентный период модели, высокую фоновую
нию с контрольными животными. Этот резуль-
активацию глутаматергической системы при
тат противоположен тому, что было получено
снижении эффективности синаптической пе-
нами ранее в петилентетразоловой модели,
редачи в хроническую фазу модели. Эти дан-
где, наоборот, происходило увеличение порога
ные позволяют лучше понять механизмы гене-
тока в тесте МЭШ [29]. Следует подчеркнуть,
рации спонтанных рецидивирующих судорог
что после ЭС, вызванного пентилентетразолом,
и могут помочь в разработке рационально
эпилепсия не развивается, в отличие от литий-
обоснованной терапии предотвращения эпи-
пилокарпиновой модели.
лептогенеза и развития эпилепсии.
Снижение порога генерации судорожной
активности, вероятно, также обусловлено ней-
Вклад авторов. Т.Ю. Постникова, А.В. Зай-
родегенерацией в гиппокампе как в острую,
цев
- концепция и руководство работой;
так и хроническую фазы модели ВЭ [18, 19].
Т.Ю. Постникова, Г.П. Диеспиров, А.А. Ко-
В эти периоды выявлена потеря не только воз-
валенко, А.В. Грифлюк - обработка и про-
буждающих нейронов, но и избирательная
ведение экспериментов; Т.Ю. Постникова,
гибель интернейронов определенных классов
А.А. Коваленко - анализ результатов и стати-
в разных областях гиппокампа [43]. Напри-
стическая обработка данных; Г.П. Диеспиров,
мер, в области СА1 гиппокампа наблюдается
А.В. Грифлюк, А.А. Коваленко - написание
избирательная гибель парвальбуминовых и
текста; Т.Ю. Постникова, А.В. Зайцев - редак-
кальретининовых интернейронов, а в области
тирование текста статьи.
СА3 - глобальное снижение ГАМКергиче-
Финансирование. Работа выполнена при
ской нейротрансмиссии, что ведет к наруше-
финансовой поддержке Российского научного
нию баланса торможения и возбуждения [43].
фонда (грант № 22-75-00131).
В данной работе мы сфокусировались на пато-
Конфликт интересов. Авторы заявляют об
логических изменениях только в гиппокампе,
отсутствии конфликта интересов.
как наиболее уязвимой структуре мозга в дан-
Соблюдение этических норм. Все проводи-
ной модели ВЭ, однако в литий-пилокарпино-
мые исследования выполнялись в соответ-
вой модели различные гистопатологические
ствии с правилами обращения с лаборатор-
изменения выявлены также в ряде областей
ными животными, действующими в ИЭФБ
коры больших полушарий, амигдале, тала-
РАН и утвержденными правовыми актами РФ
мусе, черной субстанции и др. [13]. Эти струк-
и международных организаций.
СПИСОК ЛИТЕРАТУРЫ
1. Fattorusso, A., Matricardi, S., Mencaroni, E.,
4. Engel, J. J. (2001) Mesial temporal lobe epilepsy:
Dell’Isola, G. B., Di Cara, G., Striano, P., and Verrot-
what have we learned? Neuroscientist, 7, 340-352,
ti, A. (2021) The pharmacoresistant epilepsy: an over-
doi: 10.1177/107385840100700410.
view on existant and new emerging therapies, Front.
5. Herman, S. T. (2002) Epilepsy after brain insult:
Neurol., 12, 1030, doi: 10.3389/FNEUR.2021.674483.
targeting epileptogenesis, J. Neurol., 59, S21-S26,
2. Chin, J. H., and Vora, N. (2014) The global burden
doi: 10.1212/wnl.59.9_suppl_5.s21.
of neurologic diseases, J. Neurol.,
83,
349-351,
6. Pitkänen, A., and Lukasiuk, K.
(2011) Mecha-
doi: 10.1212/WNL.0000000000000610.
nisms of epileptogenesis and potential treatment
3. Fordington, S., and Manford, M. (2020) A review of
targets, Lancet Neurol., 10, 173-186, doi: 10.1016/
seizures and epilepsy following traumatic brain injury,
S1474-4422(10)70310-0.
J. Neurol.,
267,
3105-3111, doi:
10.1007/s00415-
7. Goldberg, E. M., and Coulter, D. A. (2013) Mech-
020-09926-w.
anisms of epileptogenesis: a convergence on neural
БИОХИМИЯ том 88 вып. 3 2023
440
ДИЕСПИРОВ и др.
circuit dysfunction, Nat. Rev. Neurosci., 14, 337-349,
19.
Plata, A., Lebedeva, A., Denisov, P., Nosova, O.,
doi: 10.1038/nrn3482.
Postnikova, T. Y., Pimashkin, A., Brazhe, A.,
8.
Thom, M. (2014) Review: hippocampal sclerosis in
Zaitsev, A. V., Rusakov, D. A., and Semyanov, A.
epilepsy: a neuropathology review, Neuropathol. Appl.
(2018) Astrocytic atrophy following status epilepticus
Neurobiol., 40, 520-543, doi: 10.1111/nan.12150.
parallels reduced Ca2+ activity and impaired synaptic
9.
Bliss, T. V. P., and Collingridge, G. L. (1993) A synaptic
plasticity in the rat hippocampus, Front. Mol.
model of memory: long-term potentiation in the hippo-
Neurosci., 11, 215, doi: 10.3389/fnmol.2018.00215.
campus, Nature, 361, 31-39, doi: 10.1038/361031a0.
20.
Kryukov, K. A., Kim, K. K., Magazanik, L. G.,
10.
Titiz, A. S., Mahoney, J. M., Testorf, M. E.,
and Zaitsev, A. V. (2016) Status epilepticus alters
Holmes, G. L., and Scott, R. C. (2014) Cognitive im-
hippocampal long-term synaptic potentiation in a rat
pairment in temporal lobe epilepsy: role of online and
lithium-pilocarpine model, NeuroReport, 27, 1191-
offline processing of single cell information, Hippo-
1195, doi: 10.1097/WNR.0000000000000656.
campus, 24, 1129-1145, doi: 10.1002/hipo.22297.
21.
Clarkson, C., Smeal, R. M., Hasenoehrl, M. G.,
11.
Vlooswijk, M. C. G., Jansen, J. F. A., de Krom,
White, J. A., Rubio, M. E., and Wilcox, K. S. (2020)
M. C. F. T. M., Majoie, H. M., Hofman, P. A.
Ultrastructural and functional changes at the tripartite
M., Backes, W. H., and Aldenkamp, A. P. (2010)
synapse during epileptogenesis in a model of temporal
Functional MRI in chronic epilepsy: associations with
lobe epilepsy, Exp. Neurol., 326, 113196, doi: 10.1016/
cognitive impairment, Lancet Neurol., 9, 1018-1027,
j.expneurol.2020.113196.
doi: 10.1016/S1474-4422(10)70180-0.
22.
Naylor, D. E., Liu, H., Niquet, J., and Waster-
12.
Zavala-Tecuapetla, C., Cuellar-Herrera, M., and
lain, C. G. (2013) Rapid surface accumulation of
Luna-Munguia, H.
(2020) Insights into potential
NMDA receptors increases glutamatergic excitation
targets for therapeutic intervention in epilepsy, Int. J.
during status epilepticus, Neurobiol. Dis., 54, 225-238,
Mol. Sci., 21, 8573, doi: 10.3390/ijms21228573.
doi: 10.1016/j.nbd.2012.12.015.
13.
Curia, G., Longo, D., Biagini, G., Jones, R. S. G.,
23.
Amakhin, D. V., Soboleva, E. B., Ergina, J. L.,
and Avoli, M. (2008) The pilocarpine model of
Malkin, S. L., Chizhov, A. V., and Zaitsev, A. V. (2018)
temporal lobe epilepsy, J. Neurosci. Methods, 172, 143-
Seizure-induced potentiation of AMPA receptor-
157, doi: 10.1016/j.jneumeth.2008.04.019.
mediated synaptic transmission in the entorhinal
14.
Curia, G., Lucchi, C., Vinet, J., Gualtieri, F., Mari-
cortex, Front. Cell. Neurosci., 12, 486, doi: 10.3389/
nelli, C., Torsello, A., Costantino, L., and Biagi-
fncel.2018.00486.
ni, G. (2014) Pathophysiogenesis of mesial temporal
24.
Rajasekaran, K., Todorovic, M., and Kapur, J. (2012)
lobe epilepsy: is prevention of damage antiepilepto-
Calcium-permeable AMPA receptors are expressed in
genic? Curr. Med. Chem., 21, 663-688, doi: 10.2174/
a rodent model of status epilepticus, Ann. Neurol., 72,
0929867320666131119152201.
91-102, doi: 10.1002/ana.23570.
15.
Zaitsev, A. V., Amakhin, D. V., Dyomina, A. V.,
25.
Amakhin, D. V., Malkin, S. L., Ergina, J. L., Kryu-
Zakharova, M. V., Ergina, J. L., Postnikova, T. Y.,
kov, K. A., Veniaminova, E. A., Zubareva, O. E., and
Diespirov, G. P., and Magazanik, L. G. (2021) Synaptic
Zaitsev, A. V. (2017) Alterations in properties of glu-
dysfunction in epilepsy, J. Evol. Biochem. Physiol., 57,
tamatergic transmission in the temporal cortex and
542-563, doi: 10.1134/S002209302103008X.
hippocampus following pilocarpine-induced acute
16.
De Oliveira, D. L., Fischer, A., Jorge, R. S., Da Silva,
seizures in wistar rats, Front. Cell. Neurosci., 11, 264,
M. C., Leite, M., Gonçalves, C. A., Quillfeldt, J. A.,
doi: 10.3389/fncel.2017.00264.
Souza, D. O., E Souza, T. M., and Wofchuk, S.
26.
Malkin, S. L., Amakhin, D. V., Veniaminova, E. A.,
(2008) Effects of early-life LiCl-Pilocarpine-induced
Kim, K. K., Zubareva, O. E., Magazanik, L. G., and
status epilepticus on memory and anxiety in adult
Zaitsev, A. V. (2016) Changes of AMPA receptor prop-
rats are associated with mossy fiber sprouting and
erties in the neocortex and hippocampus following pilo-
elevated CSF S100B protein, Epilepsia, 49, 842-852,
carpine-induced status epilepticus in rats, Neuroscience,
doi: 10.1111/j.1528-1167.2007.01484.x.
327, 146-155, doi: 10.1016/j.neuroscience.2016.04.024.
17.
Morimoto, K., Fahnestock, M., and Racine, R. J.
27.
Zubareva, O. E., Kovalenko, A. A., Kalemenev, S. V.,
(2004) Kindling and status epilepticus models of
Schwarz, A. P., Karyakin, V. B., and Zaitsev, A. V.
epilepsy: rewiring the brain, Prog. Neurobiol., 73, 1-60,
(2018) Alterations in mRNA expression of gluta-
doi: 10.1016/j.pneurobio.2004.03.009.
mate receptor subunits and excitatory amino acid
18.
Postnikova, T. Y., Diespirov, G. P., Amakhin, D. V.,
transporters following pilocarpine-induced seizures
Vylekzhanina, E. N., Soboleva, E. B., and Zaitsev, A. V.
in rats, Neurosci. Lett., 686, 94-100, doi: 10.1016/
(2021) Impairments of long-term synaptic plasticity
j.neulet.2018.08.047.
in the hippocampus of young rats during the latent
28.
Racine, R. J.
(1972) Modification of seizure
phase of the lithium-pilocarpine model of temporal
activity by electrical stimulation. II. Motor seizure,
lobe epilepsy, Int. J. Mol. Sci., 22, 13355, doi: 10.3390/
Electroencephalogr. Clin. Neurophysiol., 32, 281-294,
ijms222413355.
doi: 10.1016/0013-4694(72)90177-0.
БИОХИМИЯ том 88 вып. 3 2023
ГЛУТАМАТЕРГИЧЕСКАЯ СИСТЕМА ГИППОКАМПА В МОДЕЛИ ВЭ
441
29.
Postnikova, T. Y., Amakhin, D. V., Trofimova, A. M.,
various housekeeping genes for their applicability for
Smolensky, I. V., and Zaitsev, A. V. (2019) Chang-
normalization of mRNA expression in dioxin-treated
es in functional properties of rat hippocampal neu-
rats, Chem. Biol. Interact., 160, 134-149, doi: 10.1016/
rons following pentylenetetrazole-induced status ep-
j.cbi.2006.01.001.
ilepticus, Neuroscience, 399, 103-116, doi: 10.1016/
37.
Cook, N. L., Vink, R., Donkin, J. J., and van den
j.neuroscience.2018.12.029.
Heuvel, C. (2009) Validation of reference genes for
30.
Schwarz, A. P., Malygina, D. A., Kovalenko, A. A.,
normalization of real-time quantitative RT-PCR data
Trofimov, A. N., and Zaitsev, A. V. (2020) Multiplex
in traumatic brain injury, J. Neurosci. Res., 87, 34-41,
qPCR assay for assessment of reference gene
doi: 10.1002/jnr.21846.
expression stability in rat tissues/samples, Mol. Cell.
38.
Langnaese, K., John, R., Schweizer, H., Ebmeyer, U.,
Probes, 53, 101611, doi: 10.1016/j.mcp.2020.101611.
and Keilhoff, G. (2008) Selection of reference genes for
31.
Livak, K. J., and Schmittgen, T. D. (2001) Analysis
quantitative real-time PCR in a rat asphyxial cardiac
of relative gene expression data using real-time
arrest model, BMC Mol. Biol., 9, 53, doi: 10.1186/
quantitative PCR and the
2(-Delta Delta C(T))
1471-2199-9-53.
method, Methods,
25,
402-428, doi:
10.1006/
39.
Proudnikov, D., Yuferov, V., Zhou, Y., LaForge, K. S.,
meth.2001.1262.
Ho, A., and Kreek, M. J. (2003) Optimizing primer-
32.
Bonefeld, B. E., Elfving, B., and Wegener, G. (2008)
probe design for fluorescent PCR, J. Neurosci.
Reference genes for normalization: a study of rat brain
Methods, 123, 31-45, doi: 10.1016/S0165-0270(02)
tissue, Synapse, 62, 302-309, doi: 10.1002/syn.20496.
00325-4.
33.
Lin, W., Burks, C. A., Hansen, D. R.,
40.
Zucker, R. S., and Regehr, W. G. (2002) Short-term
Kinnamon, S. C., and Gilbertson, T. A. (2004) Taste
synaptic plasticity, Annu. Rev. Physiol., 64, 355-405,
receptor cells express pH-sensitive leak K+ channels,
doi: 10.1146/annurev.physiol.64.092501.114547.
J. Neurophysiol.,
92,
2909-2919, doi:
10.1152/
41.
Owen, B., Bichler, E., and Benveniste, M. (2021)
jn.01198.2003.
Excitatory synaptic transmission in hippocampal
34.
Yamaguchi, M., Yamauchi, A., Nishimura, M., Ueda,
area CA1 is enhanced then reduced as chronic
N., and Naito, S. (2005) Soybean oil fat emulsion
epilepsy progresses, Neurobiol. Dis., 154, 105343,
prevents cytochrome P450 mRNA down-regulation
doi: 10.1016/j.nbd.2021.105343.
induced by fat-free overdose total parenteral nutrition
42.
Cull-Candy, S. G., and Farrant, M. (2021) Ca2+-
in infant rats, Biol. Pharm. Bull.,
28,
143-147,
permeable AMPA receptors and their auxiliary
doi: 10.1248/bpb.28.143.
subunits in synaptic plasticity and disease, J. Physiol.,
35.
Swijsen, A., Nelissen, K., Janssen, D., Rigo, J. M.,
599, 2655-2671, doi: 10.1113/jp279029.
and Hoogland, G. (2012) Validation of reference genes
43.
Andre, V., Marescaux, C., Nehlig, A., and
for quantitative real-time PCR studies in the dentate
Fritschy, J. M. (2001) Alterations of hippocampal
gyrus after experimental febrile seizures, BMC Res.
GABAergic system contribute to development of
Notes, 5, 685, doi: 10.1186/1756-0500-5-685.
spontaneous recurrent seizures in the at lithium-
36.
Pohjanvirta, R., Niittynen, M., Lindén, J., Boutros, P. C.,
pilocarpine model of temporal lobe epilepsy,
Moffat, I. D., and Okey, A. B. (2006) Evaluation of
Hippocampus, 11, 452-468, doi: 10.1002/hipo.1060.
ALTERATIONS IN THE PROPERTIES
OF THE GLUTAMATERGIC SYSTEM OF THE RAT HIPPOCAMPUS
IN A LITHIUM-PILOCARPINE MODEL OF TEMPORAL LOBE EPILEPSY
G. P. Diespirov, T. Y. Postnikova, A. V. Griflyuk, A. A. Kovalenko, and A. V. Zaitsev*
Sechenov Institute of Evolutionary Physiology and Biochemistry, Russian Academy of Sciences,
194223 St. Petersburg, Russia; e-mail: aleksey_zaitsev@mail.ru
Status epilepticus (SE) triggers many pathological changes in the nervous system that are not yet fully
understood and may lead to the development of epilepsy. In this work, we studied the effects of SE on the
properties of excitatory glutamatergic transmission in the hippocampus in a rat model of lithium-pilocar-
pine temporal lobe epilepsy. Studies were performed 1 day (acute phase of the model), 3 and 7 days (latent
phase), and 30 to 80 days (chronic phase) after SE. Using real-time PCR, we found that in the latent phase
there is a decrease in gene expression of GluA1 and GluA2 AMPA receptor subunits, which may also be
accompanied by an increased proportion of calcium-permeable AMPA receptors, which play an essential
role in the pathogenesis of many CNS diseases. In acute brain slices we found a decrease in the efficiency of
БИОХИМИЯ том 88 вып. 3 2023
442
ДИЕСПИРОВ и др.
excitatory synaptic neurotransmission in all phases of the model when recording field responses in the CA1
region of the hippocampus in response to stimulation of Schaffer collaterals by electric currents of different
intensities. However, in the chronic phase we found an increase in the frequency of spontaneous excitatory
postsynaptic potentials, indicating an increased background activity of the glutamatergic system in epilepsy.
This is also supported by a decrease in the threshold of hind limb extension in the test of maximal electro-
shock seizure in rats with temporal lobe epilepsy compared to control animals. The results obtained indicate
the presence of a number of functional alterations in the glutamatergic system related to epilepsy. These
findings can be used to develop antiepileptogenic therapy.
Keywords: lithium-pilocarpine model of epilepsy, synapse, AMPA receptor, hippocampus, field postsynaptic
potential, excitatory postsynaptic potential, maximal electroshock seizure
БИОХИМИЯ том 88 вып. 3 2023