БИОХИМИЯ, 2023, том 88, вып. 8, с. 1378 - 1391
УДК 576.522
ЦЕЛОСТНОСТЬ И БАРЬЕРНАЯ ФУНКЦИЯ МАКРОСОСУДИСТОГО
ЭНДОТЕЛИЯ НЕ НАРУШАЮТСЯ ПРИ ДЛИТЕЛЬНОЙ
ЭКСПЕРИМЕНТАЛЬНОЙ ГИПЕРГЛИКЕМИИ in vitro
© 2023 А.Ю. Хапчаев*, О.А. Антонова, О.А. Казакова, М.В. Самсонов,
А.В. Воротников, В.П. Ширинский
ФГБУ «Национальный медицинский исследовательский центр кардиологии имени академика Е.И. Чазова»
Минздрава России, Институт экспериментальной кардиологии имени академика В.Н. Смирнова,
121552 Москва, Россия; электронная почта: AYKhapchaev@cardio.ru
Поступила в редакцию 13.01.2023
После доработки 08.07.2023
Принята к публикации 11.07.2023
Гипергликемия является отличительной чертой сахарного диабета 2-го типа (СД2), связанной
с дисфункцией эндотелия сосудов и сердечно-сосудистыми осложнениями. Многие исследования
in vitro обнаружили апоптоз эндотелиальных клеток как ранний результат экспериментальной
гипергликемии и обозначили гибель клеток как ключевой фактор повреждения сосудов при СД2.
Однако в условиях in vivo апоптоз не наблюдается вплоть до поздних стадий развития СД2. В дан-
ной работе мы исследовали долгосрочные (до 4 недель) эффекты высокой концентрации глюко-
зы (HG, 30 мМ) на эндотелиальные клетки пупочной вены человека (HUVEC). HG не изменяла
морфологию эндотелиального монослоя, уровень продукции АФК и NO и оказывала лишь незна-
чительное влияние на маркеры апоптоза. Реакции эндотелиального барьера на различные воз-
действия не отличались от реакции клеток, культивируемых при физиологическом уровне глю-
козы (LG, 5 мМ). Фокусируясь на ключевых регуляторах сократительной активности цитоскелета
и эндотелиального барьера, мы не выявили различий в индуцированных гистамином изменени-
ях внутриклеточного Ca2+ и уровня фосфорилирования регуляторных легких цепей миозина или
фосфатазы легких цепей миозина. Полученные нами результаты показывают, что эндотелиаль-
ные клетки сосудов могут хорошо переносить относительно длительное воздействие HG, и нужны
дальнейшие исследования для изучения механизмов развития сосудистых нарушений при прогрес-
сирующем СД2.
КЛЮЧЕВЫЕ СЛОВА: HUVEC, эндотелиальный барьер, гипергликемия, инсулин, АФК, NO, РЛЦ
миозина, MYPT1.
DOI: 10.31857/S0320972523080079, EDN: IJZJHE
ВВЕДЕНИЕ
функции, вызванной гипергликемией [8-10].
Поскольку эндотелиальные клетки сосудов
Гипергликемия является основным при-
малодоступны in vivo, их изучают in vitro путем
знаком сахарного диабета 2-го типа (СД2),
выделения и культивирования в монослое [11,
связанным с дисфункцией клеток эндотелия
12]. Эндотелиальные клетки пупочной вены
сосудов [1, 2] и сердечно-сосудистыми ослож-
человека (HUVEC) являются наиболее рас-
нениями [3-6]. Макрососудистые осложнения
пространенной моделью сосудистого эндо-
являются основной причиной заболеваемости
телия in vitro [13, 14]. Они часто используются
и смертности при диабете [7]. Предполага-
при изучении влияния экспериментальной ги-
ется, что окислительный стресс является объ-
пергликемии на функции эндотелия (см. обзор
единяющим механизмом эндотелиальной дис- Vorotnikov et al. [15]). Многие авторы отмечают
Принятые сокращения: АФК - активные формы кислорода; РЛЦ - регуляторные легкие цепи миозина; СД2 - сахар-
ный диабет 2-го типа; EBM - базальная среда для культивирования клеток эндотелия; EGM - среда для роста эндо-
телиальных клеток; HG - высокая концентрация глюкозы; HUVEC - эндотелиальные клетки пупочной вены человека;
LG - нормальная концентрация глюкозы; MLCK - киназа легкой цепи миозина; MYPT1 - миозин-связывающая субъ-
единица-1 миозинфосфатазы; ROCK - RhoA-ассоциированная протеинкиназа; TER - трансэндотелиальное электри-
ческое сопротивление.
* Адресат для корреспонденции.
1378
ГИПЕРГЛИКЕМИЯ in vitro НЕ НАРУШАЕТ ЭНДОТЕЛИАЛЬНЫЙ БАРЬЕР
1379
апоптоз и гибель клеток как ранний и общий
Предиабетическая резистентность к инсу-
ответ клеток на высокий уровень глюкозы во
лину ведет к развитию гиперинсулинемии.
внешней среде (HG) [8, 16]. Однако в отсут-
Однако, в отличие от известных неблагоприят-
ствие сопутствующих патологий HG может
ных эффектов HG на эндотелиальный барьер,
длительно сохраняться у диабетиков без выражен-
инсулин оказывает вазопротекторное действие
ного апоптоза эндотелиальных клеток [17-19].
при воспалении [38] и ослабляет сердечно-со-
Такое несоответствие требует изучения с ис-
судистые осложнения у диабетиков [39]. В эн-
пользованием протоколов длительного культи-
дотелиальных клетках микрососудов in vitro
вирования эндотелиальных клеток in vitro.
инсулин стабилизирует эндотелиальный барь-
До сих пор в большинстве эндотелиальных
ер за счет инактивации сократительного аппа-
протоколов in vitro использовалась кратковре-
рата клеток [40]. Эта работа, по-видимому,
менная (обычно от 1-2 до 7 дней) обработка
является единственным исследованием влия-
клеток HG (см. обзор Vorotnikov et al. [15]) с
ния инсулина на фосфорилирование миозина
идеей, что чрезмерно высокий уровень глю-
в эндотелиальных клетках.
козы (25-30 мМ) отражает результат длитель-
В настоящем исследовании мы использо-
ного воздействия более низких концентраций
вали долгосрочные протоколы культивирова-
глюкозы in vivo. Продолжительное (7-14 дней)
ния HUVEC с постоянной HG для изучения
культивирование HUVEC было использовано
изменений в морфологии монослоя HUVEC,
в лаборатории Ceriello [20, 21] для изучения
маркеров воспаления и апоптоза, барьерных
более выраженных эффектов осцилляций глю-
свойств, развития окислительного стресса, ре-
козы по сравнению с постоянной HG. Тем не
акций на инсулин и другие стимулы, вызываю-
менее даже в этом случае (через 14 дней) посто-
щие транзиторную проницаемость, а также
янная HG существенно не повышала уровня
для оценки влияния HG на фосфорилирова-
маркеров апоптоза. Применение краткосроч-
ние миозина и фосфатазы миозина. В целом,
ных протоколов при отсутствии морфологиче-
мы не обнаружили существенного влияния
ских наблюдений in vitro не позволяет оценить
экспериментальной гипергликемии на функ-
способность эндотелиального монослоя про-
циональные свойства монослоя HUVEC.
тивостоять апоптотическому повреждению.
Физический барьер между кровью и приле-
гающими тканями является одной из основных
МАТЕРИАЛЫ И МЕТОДЫ
функций эндотелия сосудов [22]. Различные
патологические состояния (например, СД2) и
Материалы. Общие реагенты, желатин и
стрессоры, такие как активные формы кисло-
человеческий инсулин («Sigma», США) ис-
рода (АФК) или медиаторы воспаления (на-
пользовали, как описано ранее [41, 42]. Стан-
пример, гистамин), могут нарушать целост-
дартизованная базовая среда для поддержания
ность эндотелиального монослоя [23-25], что
эндотелиальных клеток (EBM) и среда для
ведет к повышению проницаемости сосудов
роста эндотелиальных клеток (EGM) были
и развитию отека. Сокращение эндотелиаль-
получены от «Cell Application» (США, #210-
ных клеток и образование межклеточных за-
500); глутамин, пенициллин и стрептомицин -
зоров зависит от активности миозина II [26],
от «Gibco» (США); гистамин - от «Sigma»;
которая регулируется фосфорилированием его
Fluo-4 AM (#F14201) - от «Life Technolo-
регуляторных легких цепей (РЛЦ) с участием
gies» (США); DAF-FM-DA (4-амино-5-метил-
ионов Са2+ [27] или Са2+-независимых меха-
амино-2′,7′-дифлуоресцеин диацетат)
- от
низмов [28, 29]. Основными киназами РЛЦ
«Invitrogen» (США) и DCF-DA (2′,7′-дихлоро-
являются Ca2+-зависимая киназа легких цепей
флуоресцеин диацетат)
- от
«Sigma». Для
миозина (MLCK) и Ca2+-независимая RhoA-
вестерн-блоттинга использовали следующие
ассоциированная киназа (ROCK) [30, 31]. Они
антитела: фосфо-eNOS(Ser1177)
(#612392)
совместно противодействуют активности фос-
и eNOS (#N30020L14)
- «BD Biosciences»
фатазы легких цепей миозина (MLCP) [32].
(США); моно-фосфо-РЛЦ (Ser19)
(#3675),
Благодаря своей регуляторной миозин-связы-
ди-фосфо-РЛЦ (S19/T18)
(#3674) и анти-
вающей субъединице MYPT1, MLCP высоко
тела козы против иммуноглобулинов кро-
специфична к миозину и является единствен-
лика, конъюгированные с пероксидазой хре-
ной известной фосфатазой миозина в эндо-
на (HRP) (#7074) - «Cell Signaling» (США);
телиальных клетках [33, 34]. Тогда как роль
анти-MYPT1 (#sc-25618), фосфо-MYPT1(S695)
MLCK, ROCK и MLCP в регуляции гиперпро-
(#sc-337531), фосфо-MYPT1(T696) (#sc-17556),
ницаемости эндотелия доказана [34-37], их
фосфо-MYPT1(T853) (#sc-17432), анти-ROCK1
реакции на HG не исследованы.
(#sc-5560) и анти-ROCK2 (#sc-5561) - «Santa
БИОХИМИЯ том 88 вып. 8 2023
7*
1
380
ХАПЧАЕВ и др.
Cruz Biotechnology» (США); анти-винкулин
среды EGM с добавлением 2 мМ глутамина,
(«Abcam», США; #ab18058), анти-ГАФД (гли-
100 Ед./мл пенициллина и 100 мкг/мл стрепто-
церальдегидфосфатдегидрогеназа)
(«Merck»,
мицина. Непрерывный мониторинг TER на-
США; #MAB374), анти-РЛЦ («ProteinTech»,
чинали сразу в многочастотном режиме, ко-
США; #10324-1-AP), антитела кролика против
торый переключали на 16 кГц через 10 дней,
иммуноглобулинов мыши («Sigma»; #A9044),
чтобы регистрировать быстрые реакции на
фосфо-ROCK2(S1366) («Invitrogen»; #PA5-34895).
гистамин. После формирования прочного мо-
Биотинилированные моноклональные анти-
нослоя (значение TER 15-16 кОм) в питатель-
тела 10F3B2 против молекулы клеточной ад-
ную среду дополнительно добавляли 25 мМ
гезии ICAM-1 [43], набор для определения
D-глюкозы до конечной концентрации 30 мМ
аннексина Annexin-V:FITC («Bio-Rad», США;
(экспериментальная гипергликемия, HG) и/или
#ANNEX100F) и набор для определения ак-
1 нМ инсулина (экспериментальная гиперин-
тивной каспазы-3 (Active Caspase-3 FITC Mab
сулинемия); контрольные клетки культиви-
Apoptosis Kit I, «BD Biosciences»; #550480) ис-
ровали в среде EGM (5 мМ D-глюкозы, LG).
пользовали для проточной цитофлуориметрии.
Значения TER регистрировали в течение
Для иммунохимии использовали препараты
трех недель. Среду заменяли через день, под-
Alexa-fluor 488-конъюгированный фаллоидин
держивая указанные концентрации глюкозы
(«Invitrogen»; #A-12379), DAPI (4',6-диамиди-
и/или инсулина.
но-2-фенилиндол; «Molecular Probes» (США);
Измерение продукции NO. Клетки HUVEC
#D-1306), первичные антитела против VE-кад-
высевали в 96-луночные микропланшеты с
герина («MyBioSource», США; #MBS176401) и
черными стенками и плоским дном (μclear
конъюгированные с Alexa-fluor 594 вторичные
black, Cellstar, TC, F-bottom; «Greiner Bio-One»,
антитела против иммуноглобулинов кролика
Австрия), покрытые
0,2%-ным желатином,
(«Invitrogen»; #R37119). Набор ингибиторов про-
по 0,5 × 105 клеток на лунку, в 100 мкл среды
теаз и фосфатаз «Halt Protease and phosphatase
EGM с добавлением 2 мМ глутамина, 100 Ед./мл
inhibitor cocktail» («Thermo Fisher Scientific»,
пенициллина и
100 мкг/мл стрептомицина.
США) использовали при лизисе клеток, Clarity
После формирования монослоя клеткам со-
ECL («Bio-Rad») - для получения изображе-
здавали условия гипергликемии и/или гипер-
ний после вестерн-блоттинга. HUVEC были
инсулинемии, как это описано выше. Для из-
получены от трех здоровых доноров (от каждо-
мерения уровня NO клетки инкубировали
го донора объединяли равное количество кле-
30 мин в 100 мкл свежей среды EBM, содер-
ток), и во всех экспериментах использовали
жащей 2 мкМ DAF-FM-DA, 1 мМ пробене-
клетки третьего пассажа.
цид («Invitrogen») и 15 мМ HEPES. Затем среду
Поддержание монослоев HUVEC. HUVEC
заменяли на 100 мкл среды EBM, содержащей
культивировали в покрытых 0,2%-ным жела-
только 1 мМ пробенецида и 15 мМ HEPES.
тином 24-луночных планшетах («SPL», Юж-
Флуоресценцию DAF измеряли на микро-
ная Корея) в среде роста EGM с добавлением
скопе AxioVert
200M, снабженном камерой
2 мМ глутамина, 100 Ед./мл пенициллина и
High Speed AxioCam HSm cooled CCD («Zeiss»,
100 мкг/мл стрептомицина, как описано ра-
Германия). Базальную флуоресценцию DAF
нее [42]. Среду роста заменяли через день.
в нестимулированных клетках считали фоновой
Фазово-контрастную микроскопию прово-
флуоресценцией. В соответствующие ячейки
дили сразу после смены среды с использова-
добавляли инсулин до конечной концентра-
нием инвертированного микроскопа Nikon
ции
10 нМ. Сигналы флуоресценции DAF
Diaphot («Nikon», Япония), снабженного ка-
были получены при 10-кратном увеличении и
мерой Nikon D40 («Nikon»). Полученные изо-
включали 200-400 клеток в поле зрения. Изо-
бражения обрабатывали с помощью бес-
бражения получали каждые 10 мин в течение
платного программного обеспечения ImageJ
40-60 мин при минимальной интенсивности
(NIH, США).
освещения, используя время экспозиции 0,8 с,
Проницаемость монослоя HUVEC. Для
чтобы свести к минимуму эффекты обесцве-
определения проницаемости монослоя HUVEC
чивания. Все значения флуоресценции норма-
измеряли трансэндотелиальное электрическое
лизовали к средней фоновой флуоресценции,
сопротивление (TER), как описано ранее
которую определяли перед добавлением инсу-
[41, 42], с использованием прибора ECIS-z
лина, и к количеству клеток. Подтверждали
(«Applied Biophysics», США). Клетки высева-
линейный характер кинетики накопления
ли в лунки планшетов 8W10E («Applied Bio-
флуоресценции DAF, и по ее линейным участ-
physics»), предварительно покрытых 0,2%-ным
кам рассчитывали значения тангенса, соответ-
желатином, по 1,8 × 105 клеток на лунку в 400 мкл
ствующего скорости выделения NO [42].
БИОХИМИЯ том 88 вып. 8 2023
ГИПЕРГЛИКЕМИЯ in vitro НЕ НАРУШАЕТ ЭНДОТЕЛИАЛЬНЫЙ БАРЬЕР
1381
Измерение продукции АФК. HUVEC куль-
увеличении, изображения обрабатывали с по-
тивировали в 96-луночных микропланшетах,
мощью бесплатного программного обеспече-
как указано выше, с добавлением или без
ния LAS X 3.7.6.25997 («Leica Microsystems CMS
25 мМ глюкозы и/или 1 нМ инсулина. Перед из-
GmbH», Германия), GIMP 2.10.34 и ImageJ.
мерением АФК среду заменяли на EBM на 1 ч,
Проточный цитофлуориметр FACSCanto II
затем на EBM, содержащую 10 мкМ DCF-DA,
(«Becton Dickinson», США) использовали для
на 60 мин, затем на EBM на 30 мин, после чего
определения экспозиции на поверхности кле-
клетки помещали на 30 мин в раствор Хэнка,
ток провоспалительного маркера (ICAM-1),
содержащий 15 мМ HEPES. Интенсивность флуо-
или раннего маркера апоптоза (фосфатидил-
ресценции DCF измеряли на планшетном ри-
серин) [44, 45], или внутриклеточного специ-
дере Victor X3 («Perkin Elmer», США). Парал-
фичного маркера апоптоза (активированной
лельно определяли фоновую флуоресценцию
каспазы-3) [46]. Через 2 дня после последней
в клетках, не обработанных DCF-DA; эти зна-
замены среды в каждой пробе анализировали
чения вычитали из значений флуоресценции
не менее 3000 клеток. Для этого собирали кле-
клеток, обработанных DCF-DA. Результаты вы-
точную среду с открепившимися клетками и
ражали как разницу флуоресценции DCF в клет-
осаждали клетки путем низкоскоростного цен-
ках, подвергнутых воздействию HG и/или инсу-
трифугирования. Прикрепленные клетки сни-
лина, и в контрольных клетках, нормализован-
мали раствором трипсин/ЭДТА (0,02%/0,05%).
ную к таковой в контрольных клетках [42].
Для каждой лунки эти две клеточные фракции
Определение концентрации внутриклеточ-
объединяли и делили на три части: 1) для опре-
ного Ca2+. HUVEC культивировали в 8-луноч-
деления ICAM-1, 2) для определения фосфати-
ных камерах («Ibidi», Германия), предвари-
дилсерина (путем связывания аннексина V) и
тельно покрытых 0,2%-ным желатином, как
окрашивания пропидий иодидом, 3) для опре-
описано выше. В среду добавляли Fluo-4 AM
деления активированной каспазы-3. После-
на 20 мин до конечной концентрации 1 мкМ,
дующие процедуры выполняли в соответствии
затем добавляли 10 мМ HEPES. Флуоресцен-
с инструкциями производителя и как это было
цию Fluo-4 измеряли, как указано выше, при
описано ранее [47].
минимальной интенсивности освещения на
Вестерн-блоттинг. Клетки HUVEC выра-
микроскопе AxioVert 200M, оснащенном вы-
щивали до достижения монослоя в чашках
сокоскоростной охлаждаемой камерой и встро-
диаметром 35 мм, предварительно покрытых
енным термостатом, отрегулированным до 37 °C.
0,2%-ным желатином в среде EGM, как опи-
Изображения получали с 10-секундными ин-
сано выше, в условиях LG или HG, без или с
тервалами. Через 2 мин после начала записи
добавлением инсулина. Клетки стимулировали
к клеткам вносили 10 мкМ гистамина. Полу-
гистамином (10 мкМ) в течение разного време-
ченные данные обрабатывали с помощью мо-
ни, после чего клеточную среду удаляли, клет-
дуля Physiology программного обеспечения
ки быстро промывали ледяным PBS и лизиро-
AxioVision 4.8.2 («Zeiss») и с помощью бес-
вали в 2-кратном буфере Лэммли для образцов,
платного программного обеспечения ImageJ.
содержащем также набор ингибиторов протеаз
Иммунофлуоресценция и проточная цито-
и коктейль с ингибиторами фосфатаз. Лизаты
флуориметрия. HUVEC культивировали в по-
клеток 10 раз пропускали через иглу шпри-
крытых желатином
8-луночных камерах
ца 30G для разрушения ДНК, нагревали при
(«Ibidi») в условиях LG или HG в течение
56 oC в течение 30 мин и центрифугировали при
10 дней при смене среды роста клеток через
16 000 g (4 °C, 10 мин). Вестерн-блоттинг прово-
день. Клетки фиксировали в 3,7%-ном форм-
дили, как описано ранее [41, 42], с использова-
альдегиде в фосфатном солевом растворе (PBS)
нием прибора Criterion («Bio-Rad») при 0,4 A·ч
в течение 5 мин и обрабатывали 0,1%-ным
для переноса белков на поливинилиденфто-
Triton X-100 в PBS в течение 5 мин. Фиксиро-
ридные мембраны («Millipore», США). Мем-
ванные клетки блокировали в 1%-ном БСА в
браны инкубировали с сайт-специфическими
течение 1 ч и инкубировали сначала с первич-
или независимыми от фосфорилирования ан-
ными антителами кролика против VE-кадге-
тителами, а затем со вторичными антителами,
рина, затем со вторичными антителами, конъ-
конъюгированными с пероксидазой. Разные
югированными с Alexa-fluor 594 (для окраски
формы РЛЦ визуализировали последовательно
VE-кадгерина), и с фаллоидином, конъюгиро-
на одной и той же мембране после стрипиро-
ванным с Alexa-fluor 488 (для окраски F-акти-
вания антител, как описано ранее [48]. Изо-
на); для визуализации клеточных ядер исполь-
бражения получали, обрабатывали и анали-
зовали DAPI. Флуоресценцию регистрировали
зировали с помощью реагентов Clarity ECL,
на микроскопе Leica Stellaris 5 при 40-кратном
прибора Fusion-SL 3500WL («Vilber Lourmat»,
БИОХИМИЯ том 88 вып. 8 2023
1
382
ХАПЧАЕВ и др.
Франция) и программы ImageJ; уровни содер-
ческой значимости. Уровень АФК, измеренный
жания общего белка в образцах нормализовали
на 5-й день воздействия HG, также был выше,
по винкулину и ГАФД. Для сравнения разных
но статистически не отличался от такового
наборов образцов в гели загружали эталонные
в контрольных клетках, культивированных
образцы.
в LG (рис. 1, е). Наконец, ни базальная, ни
Статистическая обработка результатов. По-
стимулированная инсулином продукция NO
лученные данные анализировали с использова-
не изменялись под действием HG (рис. 1, ж).
нием двустороннего U-критерия Манна-Уит-
В целом, эти результаты показывают, что
ни с уровнем значимости 0,05. Для проведения
длительная экспериментальная гипергликемия
множественных сравнений применяли кри-
не нарушает целостность монослоя HUVEC, а
терий Крускала-Уоллиса, а затем - апосте-
также не увеличивает продукцию АФК или NO
риорный критерий Данна с использованием
в течение срока воздействия HG, обычно ис-
поправки Бонферрони. Данные представлены
пользуемого в других работах. Далее мы иссле-
в виде диаграмм «ящик с усами» (box-and-
довали барьерные свойства монослоя HUVEC
whisker) или среднего значения ± стандартное
и его реакции на различные стрессоры.
отклонение (M ± SD), как указано в подпи-
HG не влияет на эндотелиальный барьер
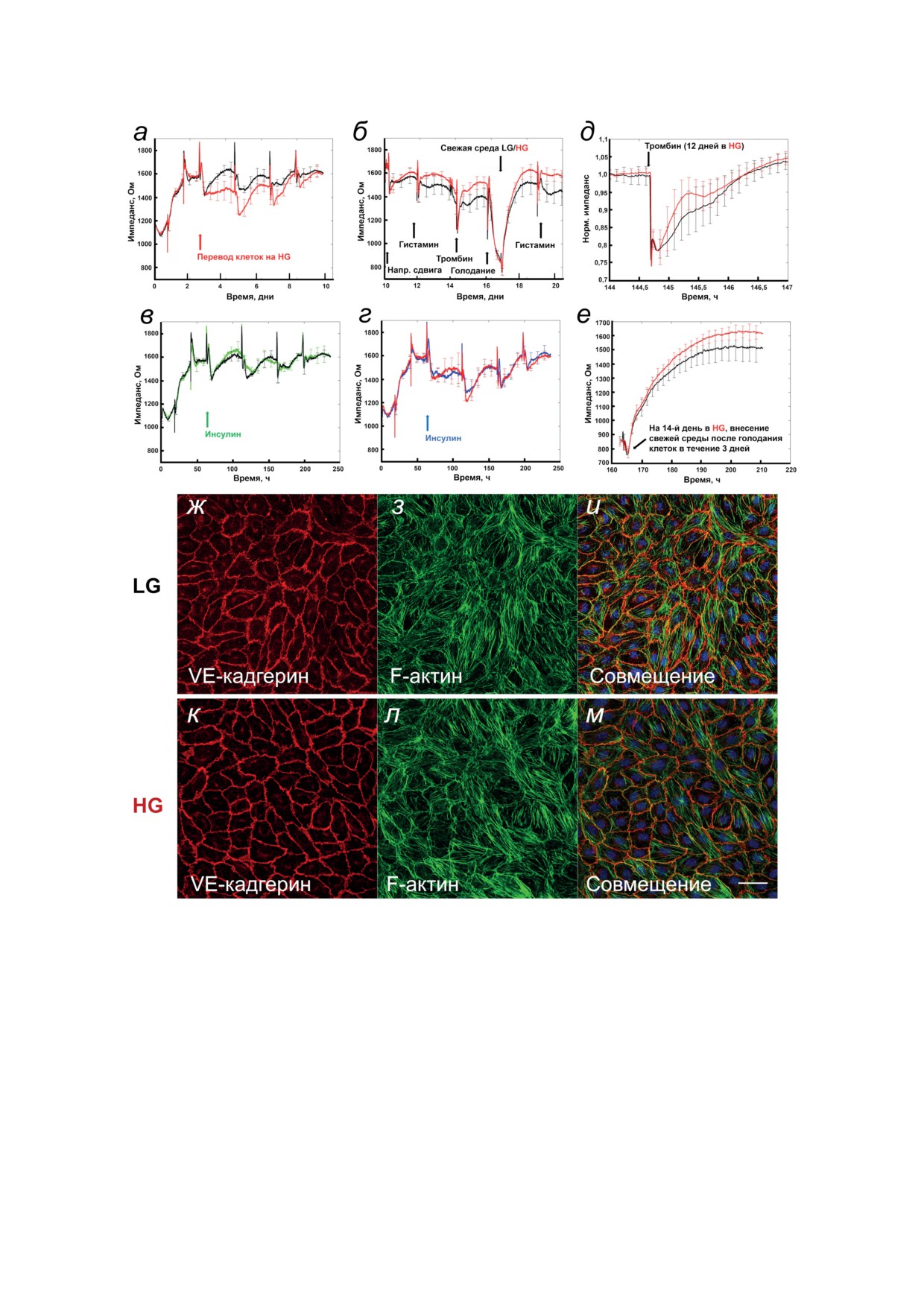
сях к рисункам. Эксперименты проводили не
HUVEC. Эндотелиальный барьер регистри-
менее чем в двух повторах и повторяли n раз,
ровали непрерывно по трансэндотелиаль-
как указано в подписях к рисункам.
ному электрическому сопротивлению (TER).
На рис. 2, а и б показаны репрезентативные за-
писи TER, полученные в течение первых 10 дней
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
после посева клеток и последующих 10 дней
соответственно. Через 2 дня барьер полностью
В работе были использованы долгосрочные
укрепился и значения TER стабилизировались
протоколы культивирования HUVEC без пере-
на уровне 16-18 кОм. После этого иницииро-
сева клеток и с заменой питательной среды
вали экспериментальную гипергликемию и/или
через день. Для сравнения с результатами дру-
гиперинсулинемию. В отдельных эксперимен-
гих исследований, измерения NO и АФК, мар-
тах, показанных на рис. 2, а, HG умеренно сни-
керов апоптоза и уровня фосфорилирования
жала уровень TER примерно на 10%, но затем
белков проводили через 5 дней после воздей-
TER постепенно восстановился до значений,
ствия HG на клетки. Остальные эксперименты
наблюдаемых до инициации HG и в LG. В дру-
проводили, используя длительные протоколы
гих экспериментах (не показано) начальное
(три и более недели), для мониторинга состоя-
падение уровня TER не было очевидным.
ния клеточного монослоя, эндотелиального
Во всех культурах HUVEC наблюдалось
барьера и кальциевых ответов.
резкое падение значений TER после каждой
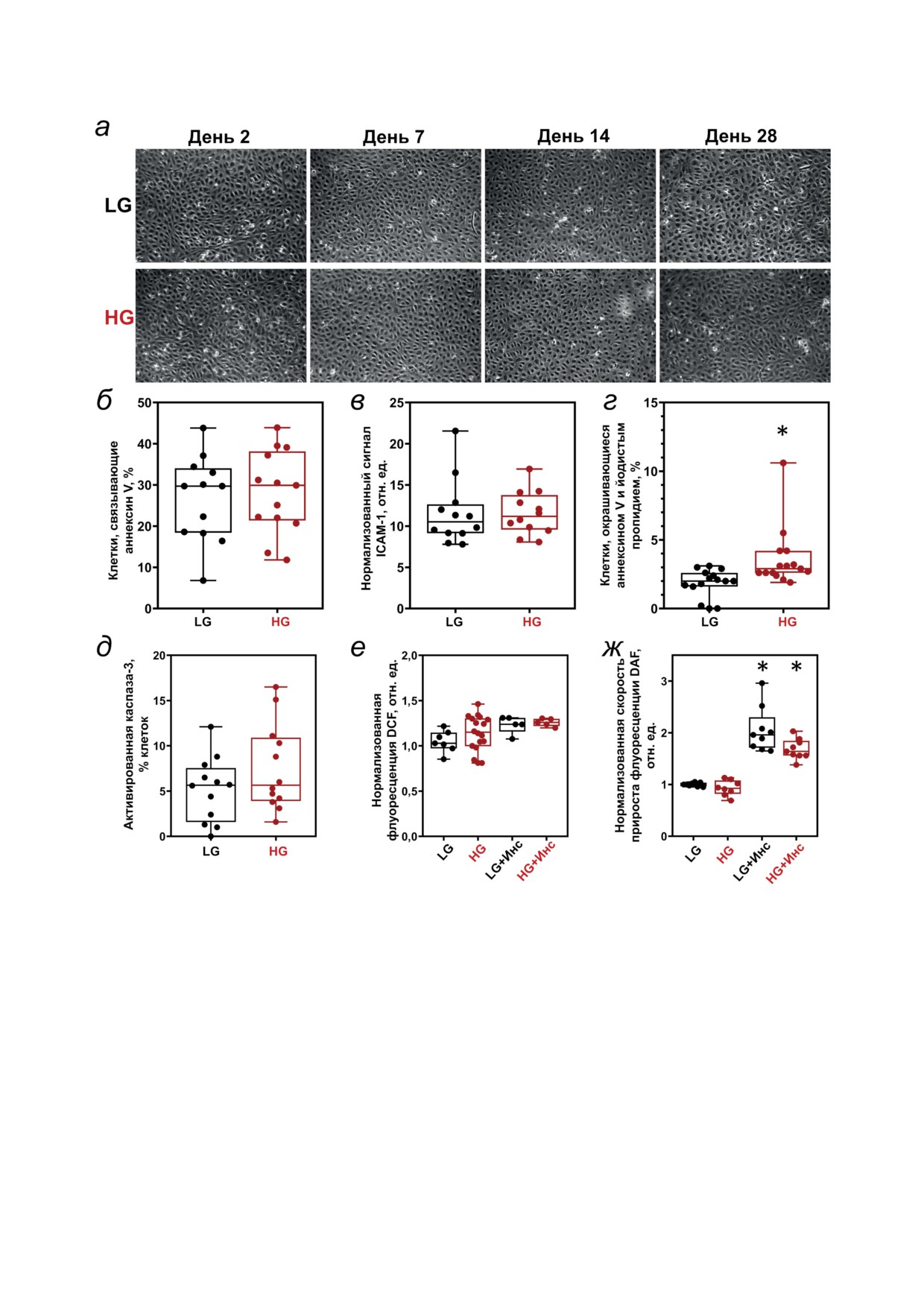
HG не влияет на морфологию монослоя
смены среды, после чего уровень TER восста-
HUVEC и продукцию АФК и NO. Световая мик-
навливался до исходных значений в течение по-
роскопия показала, что эндотелиальный моно-
следующих нескольких часов. Эти изменения,
слой сохранял классический вид «булыжной
вызванные свежей средой, были более выра-
мостовой», который не изменялся при воздей-
жены в клетках, находящихся в условиях HG,
ствии HG в течение 4 недель (рис. 1, а). Мы не
но на 10-й день их амплитуда сравнялась с
обнаружили существенных изменений в экспо-
таковой в контрольных клетках. После этого
зиции провоспалительного фосфатидилсерина
номинальные значения TER клеток в услови-
(рис. 1, б) или молекулы клеточной адгезии
ях HG демонстрировали тенденцию к превы-
ICAM-1 (рис. 1, в) на поверхности клеток через
шению показателей контрольных (LG) клеток
5 дней воздействия HG. Значимое различие
при сохранении схожей динамики (рис. 2, б).
по сравнению с контрольными клетками (хотя
В течение этого периода мы исследовали реак-
всего лишь 3,00 ± 0,99% против 2,08 ± 0,78%)
ции TER на различные стрессоры, такие как
было обнаружено для дважды положительных
напряжение сдвига, вызванное повторным
по аннексину V и йодиду пропидия клеток
пипетированием среды, стимуляцию гиста-
(рис. 1, г), что указывает на инициацию апоп-
мином (10 мкМ) (рис. 2, б), или α-тромбином
тоза в этих клетках [44]. Хотя количество кле-
(100 нМ) (рис. 2, д), или клеточным голоданием
ток, положительных по активированной кас-
(среду не меняли 3 дня) (рис. 2, е). После каждо-
пазе-3, истинному маркеру апоптоза [46], было
го воздействия проводили замену среды на све-
заметно выше в условиях HG (рис. 1, д), в сред-
жую для удаления остаточных эффектов стрес-
нем оно не превышало 6% от всей клеточной по-
сорного агента. Очевидно, что HG существенно
пуляции, и эти различия не достигали статисти-
не изменяла ответы TER на напряжение сдвига,
БИОХИМИЯ том 88 вып. 8 2023
ГИПЕРГЛИКЕМИЯ in vitro НЕ НАРУШАЕТ ЭНДОТЕЛИАЛЬНЫЙ БАРЬЕР
1383
Рис. 1. Влияние экспериментальной гипергликемии на морфологию монослоя HUVEC (а), провоспалительные
маркеры (б и в), маркеры апоптоза (г и д), продукцию АФК (е) и NO (ж). Клетки культивировали в обычной сре-
де EGM (LG) или EGM с добавлением 25 мМ глюкозы (HG) в течение 4 недель (а) или 5 дней (б-ж), как указано.
а - Репрезентативные фазово-контрастные изображения клеточных монослоев, полученные после замены клеточ-
ной среды. Провоспалительное состояние клеток определяли с помощью проточной цитофлуориметрии через 2 дня
после последней замены культуральной среды: б - по экспозиции фосфатидилсерина (связывание аннексина V);
в - по экспрессии молекул адгезии ICAM-1 на внешней поверхности плазматической мембраны. Апоптоз HUVEC
измеряли по доле клеток, дважды положительных по аннексину V и йодистому пропидию (PI) (г) или положительных
по активированной каспазе-3 (д). Данные представлены в виде диаграмм, показывающих межквартильный диапа-
зон (б-ж) и выраженных в процентах клеток, положительных по соответствующему маркеру (б, г, д), или в виде сигнала
флуоресценции, нормализованного относительно сигнала в контрольных (LG) образцах (в, е, ж). Инсулин (1 нМ) при-
сутствовал в течение всего периода (е) или был внесен однократно на 30 мин (ж), где это указано (+ Ins). * p = 0,0056 (г),
p = 0,00012 (ж, LG + Ins), p = 0,0015 (ж, HG + Ins) по сравнению с соответствующими контролями, n = 4-5
гистамин или α-тромбин, восстановление по-
ции TER ни контрольных клеток (рис. 2, в), ни
сле голодания, т.е. ответы были сходными по
клеток в условиях HG (рис. 2, г). Окрашивание
длительности и амплитуде. Экспериментальная
VE-кадгерина (рис. 2, ж и к) и актинового ци-
гиперинсулинемия также не повлияла на реак-
тоскелета (рис. 2, з и л) в клетках, находившихся
БИОХИМИЯ том 88 вып. 8 2023
1384
ХАПЧАЕВ и др.
Рис. 2. Длительная экспериментальная гипергликемия и/или гиперинсулинемия существенно не влияют на базовое
электрическое сопротивление эндотелиального барьера HUVEC и его реакции на стресс. Трансэндотелиальное элек-
трическое сопротивление (TER) монослоя HUVEC регистрировали непрерывно при частоте 16 кГц в условиях низких
(LG, 5 мМ) или высоких концентраций глюкозы (HG, 30 мМ) и/или инсулина (1 нМ). Смену среды проводили через
день или сразу после получения реакции на стрессовый стимул. а - Записи TER в условиях HG (красная кривая) и LG
(черная кривая) за первые 10 дней после посева клеток; HG инициировали через 62 ч после посева (красная стрел-
ка). б - Записи TER для последующих 10 дней, как продолжение записи на панели (а). Стрелками отмечены реакции
на указанные стрессоры. в и г - Записи TER за первые 10 дней после посева клеток в присутствии 1 нМ инсули-
на (момент добавления отмечен стрелкой на 62 ч) для имитации гиперинсулинемии в LG (в), зеленая кривая, или
в HG (г), красная кривая. д и е - Клетки культивировали в условиях LG (черная кривая) или HG (красная кривая)
и измеряли ответы TER на воздействие 100 нМ α-тромбина (д) или как восстановление после голодания (е) в моменты
времени, отмеченные соответствующими стрелками на панели (б). Чтобы упростить сравнение, на панели (д) при-
ведены нормализованные значения TER, где фактические значения TER составляли ~14 кОм как для условий LG,
так и для HG. Каждая кривая представляет собой среднее электрическое сопротивление монослоя с планками по-
грешностей, указывающими стандартные отклонения на основе 8 (а, б, д, е) или 4 (в, г) измерений в параллельных
лунках. ж-м - Флуоресцентные изображения HUVEC, культивированных в течение 10 дней в условиях LG (ж-и)
или HG (к-м) и окрашенных антителами к VE-кадгерину (панели ж и к; красный) или фаллоидином на F-актин
(з и л; зеленый), а также DAPI для визуализации ядер клеток (и и м; синий). Объединенные изображения представ-
лены на панелях и и м. Шкала - 50 мкм
БИОХИМИЯ том 88 вып. 8 2023
ГИПЕРГЛИКЕМИЯ in vitro НЕ НАРУШАЕТ ЭНДОТЕЛИАЛЬНЫЙ БАРЬЕР
1385
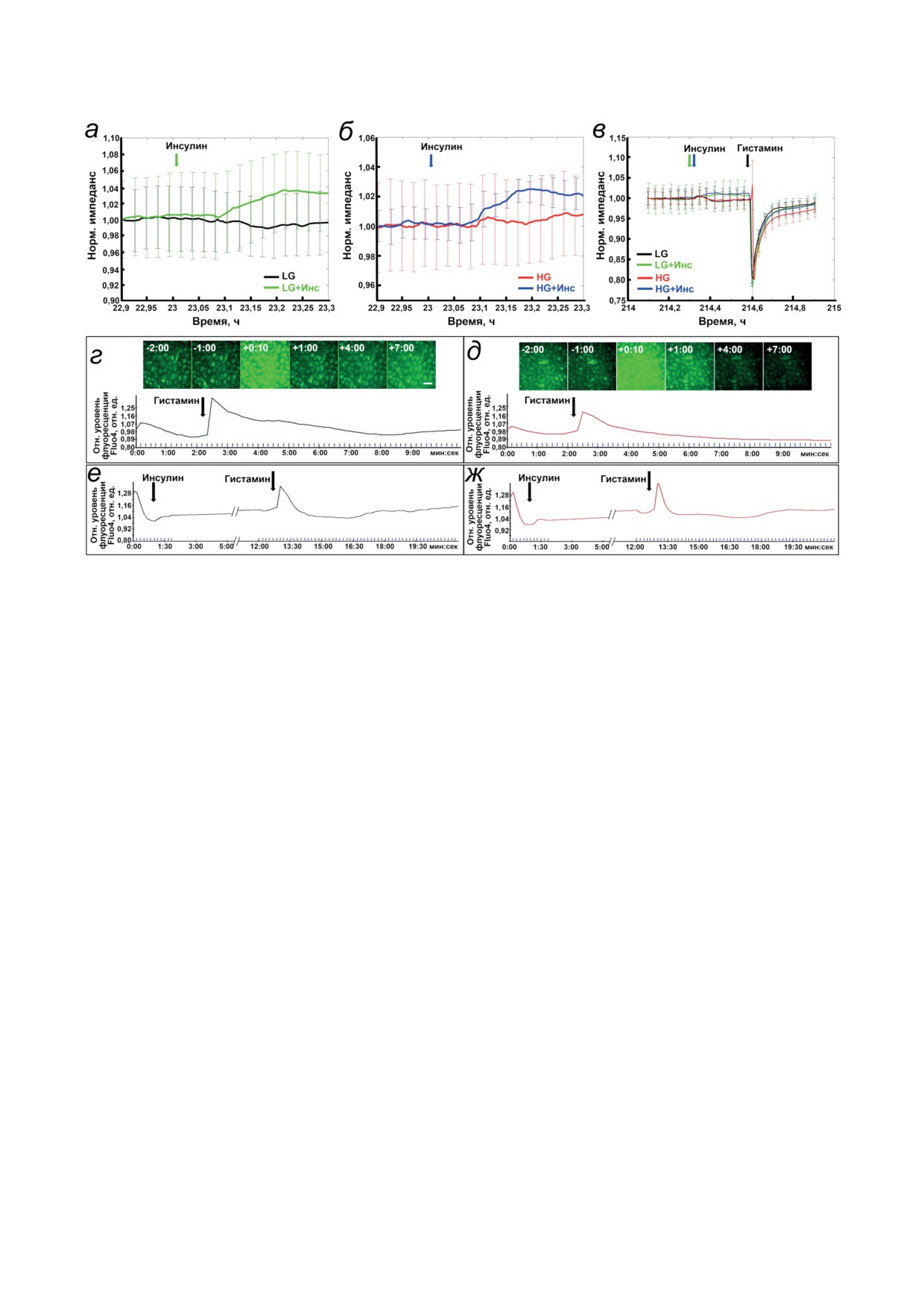
Рис. 3. HG и инсулин не влияют на транзиторные реакции [Ca2+]in и эндотелиального барьера HUVEC в ответ на гиста-
мин. а-в - Клетки культивировали в состоянии монослоя в условиях LG (a) или HG (б) и стимулировали на 11-й день
10 нМ инсулина, демонстрируя отсутствие различий между LG и HG. После такой же однократной стимуляции кле-
ток инсулином на 20-й день проводили последующую стимуляцию гистамином (10 мкМ), которая вызывала одина-
ковые транзиторные ответы TER (в). Показаны репрезентативные нормализованные записи TER; его фактические
значения составляли ~14-15 кОм для всех кривых. Каждая кривая представляет среднее электрическое сопротивле-
ние монослоя с планками погрешностей, указывающими значения стандартной ошибки среднего (SEM) для четырех
параллельных измерений. г-ж - HUVEC культивировали в состоянии монослоя в условиях LG (г и е) или HG (д и ж)
в течение 10 дней, инкубировали с Fluo-4 AM и стимулировали гистамином (10 мкМ) в момент времени 0:00 (мин:с)
или инсулином (1 нМ), как указано стрелками. На верхних панелях (г и д) показаны репрезентативные необработан-
ные изображения флуоресценции Fluo-4 в одном поле зрения в указанные моменты времени. Кривые в нижней части
панелей показывают динамику флуоресценции Fluo-4
в условиях LG или HG в течение 10 дней, также
ни инсулин, ни HG не влияли на повыше-
не показало повреждающего эффекта HG на
ние [Ca2+]in, вызванное гистамином. Типичный
межклеточные контакты. В целом, эти резуль-
профиль [Са2+]in, регистрируемый c помощью
таты не выявили существенного влияния дли-
зеленого флуоресцентного кальциевого зонда
тельной HG на целостность барьера и функцию
Fluo-4 (рис. 3, г и е), оставался неизменным для
монослоя HUVEC in vitro.
всех клеток независимо от HG или инсулина
Инсулин не влияет на изменения внутрикле-
(рис. 3, е и ж). Примечательно, что спонтанные
точного уровня Ca2+ и эндотелиальный барьер
колебания [Ca2+]in происходили случайным об-
HUVEC в ответ на действие гистамина. Тогда
разом в отдельных нестимулированных клетках.
как длительная экспериментальная гиперин-
Гистамин увеличивал флуоресценцию Fluo-4
сулинемия не влияла на эндотелиальный
одновременно во всех клетках в течение 10-20 с,
барьер HUVEC (рис. 2, в и г), мы проверили
после чего интенсивность флуоресценции воз-
гипотезу о том, влияет ли инсулин на мобили-
вращалась к исходному уровню. В целом, эти
зацию внутриклеточного Ca2+ ([Ca2+]in) и про-
результаты показывают, что длительная HG не
ницаемость монослоя HUVEC в ответ на ги-
влияет на всплески [Ca2+]in и проницаемость
стамин. Однократная стимуляция инсулином
барьера HUVEC в ответ на гистамин.
вызывала небольшие изменения TER, пока-
HG не изменяет содержания и уровня фос-
зывающие, что клетки реагировали на инсу-
форилирования цитоскелетных мишеней. Кроме
лин (рис. 3, а-в). Однако инсулин не изменял
быстрого Ca2+-зависимого воздействия на TER,
транзиторной реакции TER на гистамин, неза-
гистамин вызывает в эндотелиальных клетках
висимо от того подвергались ли клетки воздей-
более длительные сигнальные реакции, которые
ствию HG или нет (рис. 3. в). Соответственно,
могут влиять на барьерную функцию клеток [28].
БИОХИМИЯ том 88 вып. 8 2023
1386
ХАПЧАЕВ и др.
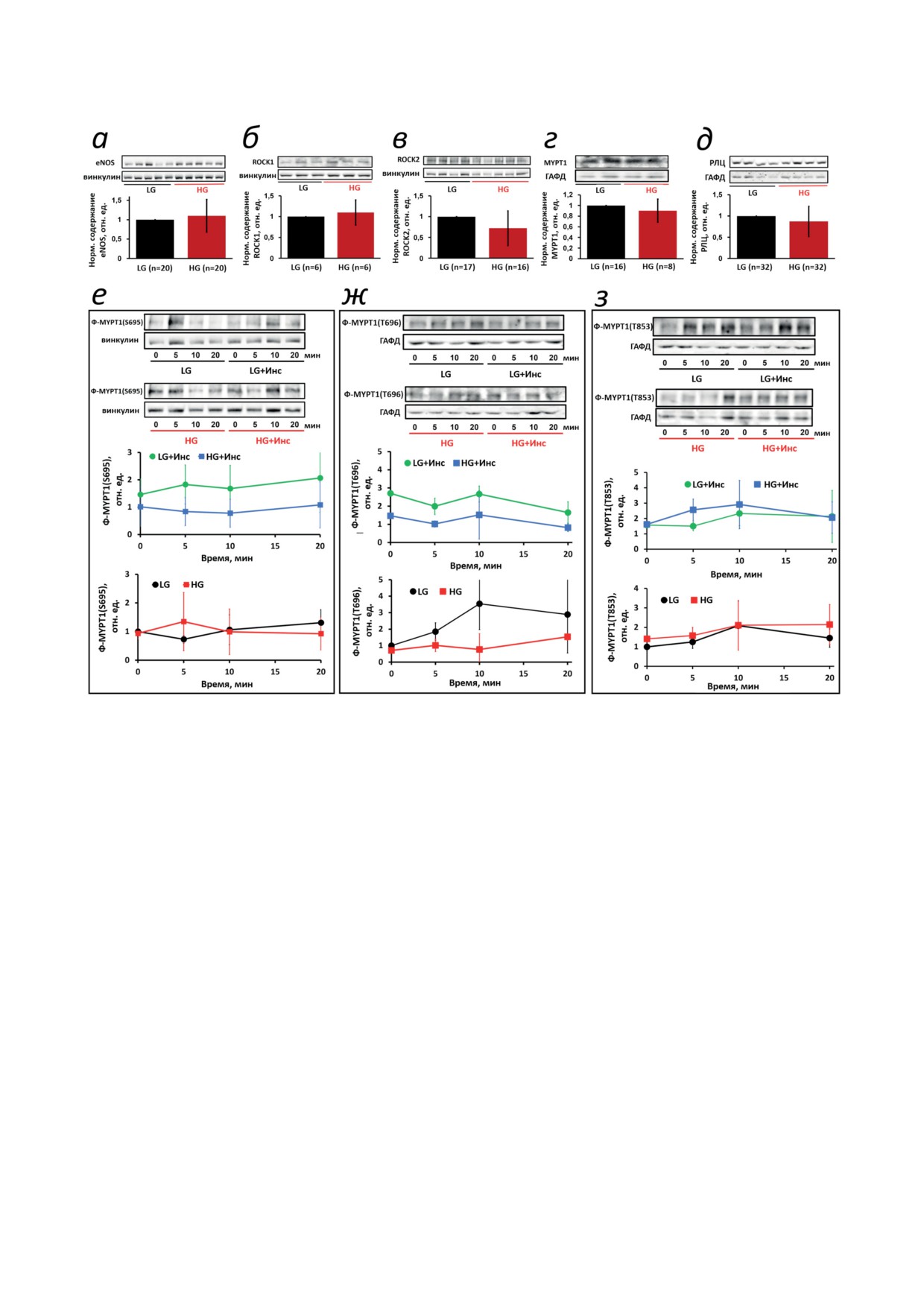
Рис. 4. HG и инсулин не влияют на содержание регуляторных белков и гистамин-зависимое фосфорилирование
MYPT1 в HUVEC. Клетки культивировали в течение 5 дней в присутствии 5 мМ (LG) или 30 мМ (HG) глюкозы и
стимулировали 10 мкМ гистамина без или с предварительной стимуляцией 10 нМ инсулина в течение 10 мин (+Инс).
а-д - Содержание белка анализировали с помощью вестерн-блоттинга в лизатах клеток, культивированных в услови-
ях LG (черные столбцы) или HG (красные столбцы). Полученные значения нормализовали относительно винкулина
(для eNOS, ROCK1/2) или ГАФД (для MYPT1, РЛЦ), а затем - относительно содержания соответствующего белка
в контрольных клетках (условия LG). Под каждым столбцом указано число независимых измерений, типичные
изображения мембран иммуноблота показаны над соответствующими столбцами. е-з - Динамика фосфорилиро-
вания белков в ответ на добавление гистамина в момент времени 0 мин. Все значения нормализованы относитель-
но базового уровня фосфорилирования Ser695 (е), Thr696 (ж) или Thr853 (з) в условиях LG без добавки инсулина.
Все данные представлены как средние значения ± стандартное отклонение по результатам по крайней мере двух неза-
висимых экспериментов, каждый из которых был проведен в трех повторах
Мы обнаружили, что через 5 дней пребывания
форилирования Ser695 и Thr696 (рис. 4, е и ж)
HUVEC в условиях HG содержание обеих изо-
и не изменяла уровень фосфорилирования
форм ROCK (ROCK1 и ROCK2), eNOS, MYPT1
Thr853 (рис. 4, з).
и РЛЦ в клетках не изменилось (рис. 4, а-д).
Экспериментальная гипергликемия не ока-
Далее, мы проверили влияние HG на гистамин-
зывала существенного влияния ни на базовый,
зависимое фосфорилирование РЛЦ и MYPT1
ни на стимулированный гистамином уровень
до или после предварительной стимуляции кле-
фосфорилирования РЛЦ (рис. 5). Поскольку ди-
ток инсулином. Фосфорилирование MYPT1 по
намика монофосфорилирования РЛЦ (Ф-РЛЦ)
сайтам, регулирующим фосфатазную актив-
(рис. 5, б и в) в основном отражает активность
ность (Ser695, Thr696 и Thr853), было одинако-
Са2+-зависимой MLCK [32], фактическое отсут-
вым на базовом уровне и незначительно изме-
ствие реакции фосфорилирования в минутном
нялось после стимуляции HUVEC гистамином
диапазоне согласуется с быстрыми изменения-
как в условиях LG, так и HG. Предварительная
ми [Са2+]in в секундном диапазоне (~30 с) в ответ
стимуляция инсулином вызывала умеренное,
на гистамин (рис. 3). Хотя просматривается тен-
но незначительное повышение уровней фос-
денция к снижению уровня Ф-РЛЦ в клетках,
БИОХИМИЯ том 88 вып. 8 2023
ГИПЕРГЛИКЕМИЯ in vitro НЕ НАРУШАЕТ ЭНДОТЕЛИАЛЬНЫЙ БАРЬЕР
1387
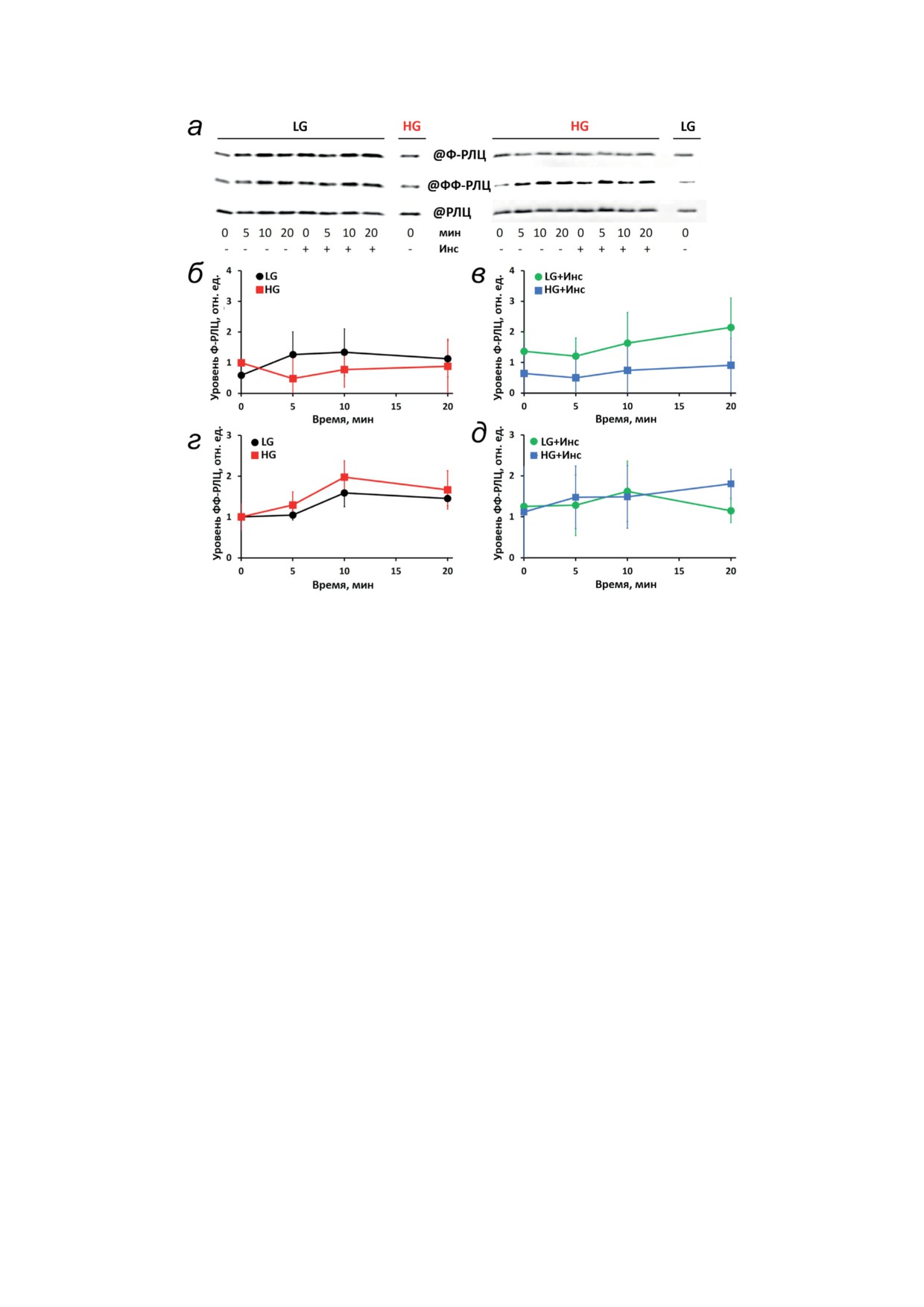
Рис. 5. HG и инсулин не влияют на индуцированное гистамином фосфорилирование РЛЦ в HUVEC. Клетки под-
вергали стимуляции, как описано на рис. 4, и анализировали с помощью вестерн-блоттинга на монофосфорилирова-
ние (Ф-РЛЦ), дифосфорилирование (ФФ-РЛЦ) и содержание регуляторных легких цепей миозина (РЛЦ). Окраши-
вание мембран проводили последовательно после стрипирования предыдущих антител. а - Типичные изображения
мембран, показывающие динамику монофосфорилирования РЛЦ по Ser19 (Ф-РЛЦ, верхние панели), дифосфори-
лирования РЛЦ по Ser19/Thr18 (ФФ-РЛЦ, средние панели) и содержания РЛЦ (нижние панели) в клетках, культиви-
рованных в условиях LG или HG, как указано. б-д - Графики, отражающие динамику Ф-РЛЦ (б и в) или ФФ-РЛЦ
(г и д) в клетках без (б и г) и после (в и д) предварительной стимуляции 10 нМ инсулина. Данные представлены как
нормализованные средние значения ± стандартное отклонение по результатам по крайней мере четырех независимых
экспериментов, каждый из которых был выполнен в двух повторах
культивированных в HG и предварительно сти-
к увеличению маркеров апоптоза через 5 дней
мулированных инсулином, это различие не было
в HG, ни повышения накопления АФК, ни
статистически значимым (рис. 5, в). Динамика
изменения скорости продукции NO не наблю-
дифосфорилирования РЛЦ (ФФ-РЛЦ) пока-
далось. Экспериментальная гиперинсулине-
зала ~2-кратное увеличение через 10 мин после
мия не влияла на эндотелиальный барьер как
стимуляции гистамином (рис. 5, г), но ни HG,
в HG, так и в LG. Эти результаты находятся в
ни присутствие инсулина на изменения не влия-
кажущемся противоречии со многими сообще-
ли (рис. 5, д). Увеличение уровня ФФ-РЛЦ мо-
ниями о том, что HG быстро повышает продук-
жет отражать активацию RhoA гистамином [28],
цию АФК и NO в HUVEC и других эндотели-
что согласуется с основным вкладом ROCK в
альных клетках, вызывая окислительный стресс
дифосфорилирование РЛЦ [32].
и апоптоз (см. обзор [15] и ссылки в нем).
При этом различия в апоптозе между клетками,
культивированными в HG и LG, были ста-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
тистически достоверными, но небольшими,
оставаясь в пределах нескольких процентов
Мы обнаружили, что целостность и внеш-
(2-10% при измерении разными методами):
ний вид монослоя HUVEC не изменяются при
3,5% [49], 10% [50], 5% [51], 6-8% [52], 5% [53],
длительном воздействии HG на клетки. Это по-
2,5% [7] и 2% даже после длительного воздей-
зволяет предположить, что нормальные функ-
ствия HG на HUVEC в течение 2 недель [21].
ции клеток также сохраняются, что хорошо
В целом, полученные нами значения находятся
согласуется с сохранением барьерных свойств
в том же диапазоне. Воспалительная реакция, о
монослоя HUVEC. За исключением тенденции
которой можно судить по экспозиции ICAM-1,
БИОХИМИЯ том 88 вып. 8 2023
1
388
ХАПЧАЕВ и др.
не развивалась. Усиление апоптотической ре-
лином показала только тенденцию к увеличе-
акции клеток на HG (по результатам двойного
нию фосфорилирования MYPT1 по остаткам
окрашивания аннексином V и йодидом про-
Ser695 и Thr696. Если для Ser695 известно, что
пидия) было достоверным, но также оставалось
его фосфорилирование опосредовано eNOS и
в пределах нескольких процентов. Более того,
cGMP-зависимой протеинкиназой [54], то за
истинный маркер апоптоза, активированная
фосфорилирование Thr696, скорее всего, отве-
каспаза-3, показывал лишь тенденцию к повы-
чает ROCK, активируемая RhoA [28]. Однако
шению без статистически достоверных разли-
фосфорилирование этих остатков оказывает
чий. Эти результаты свидетельствуют, что если
противоположное воздействие на активность
HG и усиливает апоптоз, то его доля от общей
фосфатазы [54], что согласуется с отсутствием
популяции клеток низка и постоянна, а целост-
изменений общего уровня фосфорилирова-
ность эндотелиального монослоя поддержива-
ния РЛЦ.
ется за счет пролиферации и распластывания
В целом, наши результаты указывают, что
соседних клеток. Этот феномен не мог быть
сосудистый эндотелий устойчив к длительному
замечен в предыдущих работах, поскольку в
воздействию HG. Отсутствие ранних ответов
них использовались краткосрочные протоколы
на HG в виде повышения уровня АФК и изме-
воздействия HG. С этим выводом согласуется
нения продукции NO, которые часто наблюда-
отсутствие существенных изменений эндотели-
лись в предыдущих исследованиях, моделирую-
ального барьера HUVEC в условиях HG (рис. 2).
щих гипергликемию in vitro [15], не означает,
Вопрос о том, могут ли колебания HG [20, 21]
что они не могут развиться на более поздних
снижать устойчивость клеток и давать каче-
стадиях СД2. Вполне вероятно, что повреждаю-
ственно иной результат, представляется важ-
щие эффекты гипергликемии у больных СД2
ным и требует повторного исследования.
развиваются на фоне дислипидемии, которая
Отсутствие повреждающих эффектов HG
обычно предшествует гипергликемии. Полу-
на HUVEC дополнительно подтверждается от-
ченные нами ранее данные подтверждают это
сутствием различий в реакциях эндотелиаль-
предположение [42]. Кроме хорошо известного
ного барьера на стрессовые воздействия, в меж-
окислительного стресса [8-10], эффекты HG,
клеточных контактах и актиновом цитоскелете.
вероятно, включают гликирование и гликози-
В частности, HG не изменяла реакции TER в
лирование белков [55, 56]. Очевидно, что не-
ответ на гистамин, α-тромбин или инсулин.
обходимы дальнейшие исследования для выяс-
Известно, что ответы на гистамин слабые и за-
нения молекулярных механизмов сосудистых
висят от ионов Са2+ [27, 29]. Ни инсулин, ни HG
повреждений при развитии СД2.
не влияли на повышение [Ca2+]in, вызванное ги-
стамином. Тем не менее остается возможным,
Вклад авторов. Концептуализация и мето-
что HG может влиять на отсроченные реакции
дология
- А.Ю. Хапчаев, А.В. Воротников,
эндотелия на стимулы, задействующие Ca2+-не-
В.П. Ширинский; проведение исследований -
зависимые механизмы в дополнение к класси-
А.Ю. Хапчаев, О.А. Антонова, М.В. Самсонов,
ческим Ca2+-зависимым.
О.А. Казакова, А.В. Воротников; обработ-
Индуцированное гистамином фосфорили-
ка полученных результатов - А.Ю. Хапчаев,
рование белков цитоскелета согласуется с ре-
А.В. Воротников; написание манускрипта -
акцией эндотелиального барьера на этот аго-
А.Ю. Хапчаев, А.В. Воротников; руководство -
нист. Сократимость цитоскелета, регулируемая
А.Ю. Хапчаев, А.В. Воротников, В.П. Ширин-
Са2+-зависимыми и независимыми механизма-
ский; получение финансирования - А.Ю. Хап-
ми, обеспечивает тонкую настройку реакций
чаев, В.П. Ширинский.
эндотелиального барьера на внешние сигна-
Финансирование. Работа поддержана Россий-
лы [28, 29]. Она зависит от фосфорилирования
ским научным фондом (грант № 19-15-00361)
РЛЦ миозина [26], регулируемого киназами
в части экспериментов по жизнеспособности
РЛЦ и фосфатазой [30, 34-37]. Ни содержание
HUVEC и долгосрочных экспериментов ECIS и
этих ключевых регуляторов, ни их фосфори-
Российским фондом фундаментальных иссле-
лирование, ни фосфорилирование самих РЛЦ
дований (грант № 20-015-00565) в проведении
в ответ на гистамин существенно не изменя-
других экспериментах.
лись под действием HG. Возможность того,
Конфликт интересов. Авторы заявляют об
что эти пути могут активироваться инсулином
отсутствии конфликта интересов.
и изменять реакции на гистамин, скорее всего,
Соблюдение этических норм. Настоящая
можно исключить, поскольку предварительная
статья не содержит описания каких-либо ис-
стимуляция клеток инсулином практически
следований с участием людей или животных в
не влияла на реакцию TER. Стимуляция инсу-
качестве объектов.
БИОХИМИЯ том 88 вып. 8 2023
ГИПЕРГЛИКЕМИЯ in vitro НЕ НАРУШАЕТ ЭНДОТЕЛИАЛЬНЫЙ БАРЬЕР
1389
СПИСОК ЛИТЕРАТУРЫ
1.
Giugliano, D., Ceriello, A., and Paolisso, G. (1996)
Trends Biotechnol., 35, 265-277, doi: 10.1016/j.tibtech.
Oxidative stress and diabetic vascular complica-
2016.09.007.
tions, Diabetes Care,
19,
257-267, doi:
10.2337/
14.
Medina-Leyte, D. J., Domínguez-Pérez, M.,
diacare.19.3.257.
Mercado, I., Villarreal-Molina, M. T., and Jacobo-
2.
Goligorsky, M. S. (2017) Vascular endothelium in dia-
Albavera, L. (2020) Use of human umbilical vein
betes, Am. J. Physiol. Renal Physiol., 312, F266-F275,
endothelial cells (HUVEC) as a model to study
doi: 10.1152/ajprenal.00473.2016.
cardiovascular disease: a review, Appl. Sci., 10, 938,
3.
Bakker, W., Eringa, E. C., Sipkema, P., and van
doi: 10.3390/app10030938.
Hinsbergh, V. W. (2009) Endothelial dysfunction and
15.
Vorotnikov, A. V., Khapchaev, A. Yu., Nickashin,
diabetes: roles of hyperglycemia, impaired insulin
A. V., and Shirinsky, V. P. (2022) In vitro modeling
signaling and obesity, Cell Tissue Res., 335, 165-189,
of diabetes impact on vascular endothelium: Are
doi: 10.1007/s00441-008-0685-6.
essentials engaged to tune metabolism? Biomedicines,
4.
Del Turco, S., Gaggini, M., Daniele, G., Basta, G.,
10, 3181, doi: 10.3390/biomedicines10123181.
Folli, F., Sicari, R., and Gastaldelli, A. (2013) Insulin
16.
Van den Oever, I. A., Raterman, H. G., Nurmohamed,
resistance and endothelial dysfunction: a mutual
M. T., and Simsek, S. (2010) Endothelial dysfunction,
relationship in cardiometabolic risk, Curr. Pharm. Des.,
inflammation, and apoptosis in diabetes mellitus, Media-
19, 2420-2431, doi: 10.2174/1381612811319130010.
tors Inflamm., 2010, 792393, doi: 10.1155/2010/792393.
5.
Jansson, P. A. (2007) Endothelial dysfunction in insu-
17.
Gill, S. E., Rohan, M., and Mehta, S. (2015) Role of
lin resistance and type 2 diabetes, J. Intern. Med., 262,
pulmonary microvascular endothelial cell apoptosis in
173-183, doi: 10.1111/j.1365-2796.2007.01830.x.
murine sepsis-induced lung injury in vivo, Respir. Res.,
6.
Knapp, M., Tu, X., and Wu, R. (2019) Vascular
16, 109, doi: 10.1186/s12931-015-0266-7.
endothelial dysfunction, a major mediator in diabetic
18.
Mallat, Z., and Tedgui, A. (2000) Apoptosis in the vas-
cardiomyopathy, Acta Pharmacol. Sin.,
40,
1-8,
culature: mechanisms and functional importance, Br. J.
doi: 10.1038/s41401-018-0042-6.
Pharmacol., 130, 947-962, doi: 10.1038/sj.bjp.0703407.
7.
Varma, S., Lal, B. K., Zheng, R., Breslin, J. W.,
19.
Winn, R. K., and Harlan, J. M. (2005) The role
Saito, S., Pappas, P. J., Hobson, R. W., 2nd, and
of endothelial cell apoptosis in inflammatory and
Durán, W. N. (2005) Hyperglycemia alters PI3k and
immune diseases, J. Thromb. Haemost., 3, 1815-1824,
Akt signaling and leads to endothelial cell proliferative
doi: 10.1111/j.1538-7836.2005.01378.x.
dysfunction, Am. J. Physiol. Heart Circ. Physiol., 289,
20.
Quagliaro, L., Piconi, L., Assaloni, R., Martinelli, L.,
H1744-H1751, doi: 10.1152/ajpheart.01088.2004.
Motz, E., and Ceriello, A. (2003) Intermittent high glu-
8.
Evans, J. L., Goldfine, I. D., Maddux, B. A., and
cose enhances apoptosis related to oxidative stress in
Grodsky, G. M. (2002) Oxidative stress and stress-
human umbilical vein endothelial cells: the role of pro-
activated signaling pathways: a unifying hypothesis
tein kinase C and NAD(P)H-oxidase activation, Diabe-
of type
2 diabetes, Endocr. Rev.,
23,
599-622,
tes, 52, 2795-2804, doi: 10.2337/diabetes.52.11.2795.
doi: 10.1210/er.2001-0039.
21.
Risso, A., Mercuri, F., Quagliaro, L., Damante, G.,
9.
Green, K., Brand, M. D., and Murphy, M. P. (2004)
and Ceriello, A. (2001) Intermittent high glucose en-
Prevention of mitochondrial oxidative damage as
hances apoptosis in human umbilical vein endothelial
a therapeutic strategy in diabetes, Diabetes,
53,
cells in culture, Am. J. Physiol. Endocrinol. Metab., 281,
S110-S118, doi: 10.2337/diabetes.53.2007.s110.
E924-E930, doi: 10.1152/ajpendo.2001.281.5.E924.
10.
Shah, M. S., Brownlee, M. (2016) Molecular and
22.
Komarova, Y., and Malik, A. B. (2010) Regulation of
cellular mechanisms of cardiovascular disorders in
endothelial permeability via paracellular and transcel-
diabetes, Circ. Res., 118, 1808-1829, doi: 10.1161/
lular transport pathways, Annu. Rev. Physiol., 72, 463-
CIRCRESAHA.116.306923.
493, doi: 10.1146/annurev-physiol-021909-135833.
11.
Jiménez, N., Krouwer, V. J., and Post, J. A. (2013)
23.
Cahill, P. A., and Redmond, E. M. (2016) Vascular
A new, rapid and reproducible method to obtain high
endothelium - Gatekeeper of vessel health, Athero-
quality endothelium in vitro, Cytotechnology, 65, 1-14,
sclerosis, 248, 97-109, doi: 10.1016/j.atherosclerosis.
doi: 10.1007/s10616-012-9459-9.
2016.03.007.
12.
Navone, S. E., Marfia, G., Invernici, G., Cristini, S.,
24.
Di, A., Mehta, D., and Malik, A. B. (2016) ROS-
Nava, S., Balbi, S., Sangiorgi, S., Ciusani, E., Bosutti, A.,
activated calcium signaling mechanisms regulating
Alessandri, G., Slevin, M., and Parati, E. A. (2013)
endothelial barrier function, Cell Calcium, 60, 163-171,
Isolation and expansion of human and mouse brain
doi: 10.1016/j.ceca.2016.02.002.
microvascular endothelial cells, Nat. Protoc.,
8,
25.
Rajendran, P., Rengarajan, T., Thangavel, J., Nishiga-
1680-1693, doi: 10.1038/nprot.2013.107.
ki, Y., Sakthisekaran, D., Sethi, G., and Nishigaki, I.
13.
Hauser, S., Jung, F., and Pietzsch, J. (2017) Human
(2013) The vascular endothelium and human diseases,
endothelial cell models in biomaterial research,
Int. J. Biol. Sci., 9, 1057-1069, doi: 10.7150/ijbs.7502.
БИОХИМИЯ том 88 вып. 8 2023
1
390
ХАПЧАЕВ и др.
26.
Goeckeler, Z. M., Bridgman, P. C., and Wysolmerski,
Shirinsky, V., Jia, Y., Haiech, J., Van Eldik, L. J.,
R. B. (2008) Nonmuscle myosin II is responsible for
and Watterson, D. M. (2003) Protein kinase involved
maintaining endothelial cell basal tone and stress
in lung injury susceptibility: evidence from enzyme
fiber integrity, Am. J. Physiol. Cell Physiol., 295,
isoform genetic knockout and in vivo inhibitor
C994-C1006, doi: 10.1152/ajpcell.00318.2008.
treatment, Proc. Natl. Acad. Sci. USA, 100, 6233-6238,
27.
Dalal, P. J., Muller, W. A., and Sullivan, D. P. (2020)
doi: 10.1073/pnas.1031595100.
Endothelial cell calcium signaling during barrier
38.
Langouche, L., Vanhorebeek, I., Vlasselaers, D.,
function and inflammation, Am. J. Pathol., 190, 535-
Vander Perre, S., Wouters, P. J., Skogstrand, K.,
542, doi: 10.1016/j.ajpath.2019.11.004.
Hansen, T. K., and Van den Berghe, G.
(2005)
28.
Kugelmann, D., Rotkopf, L. T., Radeva, M. Y.,
Intensive insulin therapy protects the endothelium of
Garcia-Ponce, A., Walter, E., and Waschke, J. (2018)
critically ill patients, J. Clin. Invest., 115, 2277-2286,
Histamine causes endothelial barrier disruption
doi: 10.1172/JCI25385.
via Ca2+-mediated RhoA activation and tension at
39.
Nathan, D. M., Cleary, P. A., Backlund, J. Y., Genuth,
adherens junctions, Sci. Rep., 8, 13229, doi: 10.1038/
S. M., Lachin, J. M., Orchard, T. J., Raskin, P.,
s41598-018-31408-3.
Zinman, B., and Diabetes control and complications
29.
Van Nieuw Amerongen, G. P., Draijer, R., Vermeer,
trial/epidemiology of diabetes interventions and
M. A., and van Hinsbergh, V. W. (1998) Transient
complications (DCCT/EDIC) study research group
and prolonged increase in endothelial permeability
(2005) Intensive diabetes treatment and cardiovascular
induced by histamine and thrombin: role of protein
disease in patients with type 1 diabetes, N. Engl. J.
kinases, calcium, and RhoA, Circ. Res., 83, 1115-1123,
Med., 353, 2643-2653, doi: 10.1056/NEJMoa052187.
doi: 10.1161/01.res.83.11.1115.
40.
Gunduz, D., Thom, J., Hussain, I., Lopez, D.,
30.
Khapchaev, A. Y., and Shirinsky, V. P. (2016) Myosin
Hartel, F. V., Erdogan, A., Grebe, M., Sedding, D.,
light chain kinase MYLK1: anatomy, interactions,
Piper, H. M., Tillmanns, H., Noll, T., and Aslam, M.
functions, and regulation, Biochemistry (Moscow), 81,
(2010) Insulin stabilizes microvascular endothelial barri-
1676-1697, doi: 10.1134/S000629791613006X.
er function via phosphatidylinositol 3-Kinase/Akt-me-
31.
Shimokawa, H., Sunamura, S., and Satoh, K.
diated Rac1 activation, Arterioscler. Thromb. Vasc. Biol.,
(2016) RhoA/Rho-kinase in the cardiovascular
30, 1237-1245, doi: 10.1161/ATVBAHA.110.203901.
system, Circ. Res.,
118,
352-366, doi:
10.1161/
41.
Samsonov, M. V., Podkuychenko, N. V., Lankin,
CIRCRESAHA.115.306532.
V. Z., Vorotnikov, A. V., and Shirinsky, V. P. (2021)
32.
Kazakova, O. A., Khapchaev, A. Y., and Shirinsky,
Malondialdehyde but not methylglyoxal impairs insulin
V. P.
(2020) MLCK and ROCK mutualism in
signaling, NO production, and endothelial barrier,
endothelial barrier dysfunction, Biochimie, 168, 83-91,
Biochemistry (Moscow) Suppl. Series A Memb. Cell
doi: 10.1016/j.biochi.2019.10.010.
Biol., 15, 195-200, doi: 10.1134/S1990747821030089.
33.
Ito, M., Nakano, T., Erdodi, F., and Hartshorne,
42.
Samsonov, M. V., Podkuychenko, N. V., Khapchaev,
D. J. (2004) Myosin phosphatase: structure, regulation
A. Y., Efremov, E. E., Yanushevskaya, E. V., Vlasik,
and function, Mol. Cell. Biochem., 259, 197-209,
T. N., Lankin, V. Z., Stafeev, I. S., Skulachev, M. V.,
doi: 10.1023/b:mcbi.0000021373.14288.00.
Shestakova, M. V., Vorotnikov, A. V., and Shirinsky,
34.
Kim, K. M., Csortos, C., Czikora, I., Fulton, D.,
V. P.
(2022) AICAR protects vascular endothelial
Umapathy, N. S., Olah, G., and Verin, A. D. (2012)
cells from oxidative injury induced by the long-term
Molecular characterization of myosin phosphatase
palmitate excess, Int. J. Mol. Sci., 23, 211, doi: 10.3390/
in endothelium, J. Cell. Physiol., 227, 1701-1708,
ijms23010211.
doi: 10.1002/jcp.22894.
43.
Printseva, O., Peclo, M. M., and Gown, A. M. (1992)
35.
Beckers, C. M., Knezevic, N., Valent, E. T., Tauseef, M.,
Various cell types in human atherosclerotic lesions
Krishnan, R., Rajendran, K., Hardin, C. C., Aman, J.,
express ICAM-1. Further immunocytochemical and
van Bezu, J., Sweetnam, P., van Hinsbergh, V. W.,
immunochemical studies employing monoclonal anti-
Mehta, D., and van Nieuw Amerongen, G. P. (2015)
body 10F3, Am. J. Pathol., 140, 889-896.
ROCK2 primes the endothelium for vascular hyper-
44.
Segawa, K., and Nagata, S. (2015) An apoptotic “eat
permeability responses by raising baseline junctional
me” signal: phosphatidylserine exposure, Trends Cell
tension, Vascul. Pharmacol., 70, 45-54, doi: 10.1016/
Biol., 25, 639-650, doi: 10.1016/j.tcb.2015.08.003.
j.vph.2015.03.017.
45.
Bui, T. M., Wiesolek, H. L., and Sumagin, R. (2020)
36.
Birukova, A. A., Smurova, K., Birukov, K. G.,
ICAM-1: a master regulator of cellular responses in
Kaibuchi, K., Garcia, J. G., and Verin, A. D. (2004)
inflammation, injury resolution, and tumorigenesis,
Role of Rho GTPases in thrombin-induced lung vas-
J. Leukoc. Biol.,
108,
787-799, doi:
10.1002/
cular endothelial cells barrier dysfunction, Microvasc.
JLB.2MR0220-549R.
Res., 67, 64-77, doi: 10.1016/j.mvr.2003.09.007.
46.
Porter, A. G., and Janicke, R. U. (1999) Emerging
37.
Wainwright, M. S., Rossi, J., Schavocky, J., Crawford, S.,
roles of caspase-3 in apoptosis, Cell Death Differ., 6,
Steinhorn, D., Velentza, A. V., Zasadzki, M.,
99-104, doi: 10.1038/sj.cdd.4400476.
БИОХИМИЯ том 88 вып. 8 2023
ГИПЕРГЛИКЕМИЯ in vitro НЕ НАРУШАЕТ ЭНДОТЕЛИАЛЬНЫЙ БАРЬЕР
1391
47. Antonova, O. A., Loktionova, S. A., Romanov, Y. A.,
52. Sheu, M. L., Ho, F. M., Yang, R. S., Chao, K. F.,
Shustova, O. N., Khachikian, M. V., and Mazurov,
Lin, W. W., Lin-Shiau, S. Y., and Liu, S. H. (2005)
A. V. (2009) Activation and damage of endothelial cells
High glucose induces human endothelial cell
upon hypoxia/reoxygenation. Effect of extracellular
apoptosis through a phosphoinositide
3-kinase-
pH, Biochemistry (Moscow), 74, 605-612, doi: 10.1134/
regulated cyclooxygenase-2 pathway, Arterioscler.
s0006297909060030.
Thromb. Vasc. Biol.,
25,
539-545, doi:
10.1161/
48. Kazakova, O. A., Khapchaev, A. Y., Ragimov, A. A.,
01.ATV.0000155462.24263.e4.
Salimov, E. L., and Shirinsky, V. P. (2019) Western
53. Pal, P. B., Sonowal, H., Shukla, K., Srivastava, S. K.,
blotting-based quantitative measurement of myosin II
and Ramana, K. V. (2019) Aldose reductase regulates
regulatory light chain phosphorylation in small
hyperglycemia-induced HUVEC death via SIRT1/
amounts of non-muscle cells, Biochemistry (Moscow),
AMPK-α1/mTOR pathway, J. Mol. Endocrinol.,
84, 11-19, doi: 10.1134/S0006297919010024.
63, 11-25, doi: 10.1530/JME-19-0080.
49. Baumgartner-Parzer, S. M., Wagner, L., Pettermann, M.,
54. Kiss, A., Erdődi, F., and Lontay, B. (2019) Myosin
Grillari, J., Gessl, A., and Waldhausl, W.
(1995)
phosphatase: unexpected functions of a long-known
High-glucose - triggered apoptosis in cultured en-
enzyme, Biochim. Biophys. Acta Mol. Cell Res.,
dothelial cells, Diabetes, 44, 1323-1327, doi: 10.2337/
1866, 2-15, doi: 10.1016/j.bbamcr.2018.07.023.
diab.44.11.1323.
55. Federici, M., Menghini, R., Mauriello, A., Hribal,
50. Fan, W., Han, D., Sun, Z., Ma, S., Gao, L., Chen, J.,
M. L., Ferrelli, F., Lauro, D., Sbraccia, P., Spagnoli,
Li, X., Li, X., Fan, M., Li, C., Huc, D., Wanga, Y.,
L. G., Sesti, G., and Lauro, R. (2002) Insulin-
and Cao, F. (2017) Endothelial deletion of mTORC1
dependent activation of endothelial nitric oxide
protects against hindlimb ischemia in diabetic
synthase is impaired by O-linked glycosylation
mice via activation of autophagy, attenuation of
modification of signaling proteins in human
oxidative stress and alleviation of inflammation,
coronary endothelial cells, Circulation, 106, 466-472,
Free Radic. Biol. Med., 108, 725-740, doi: 10.1016/
doi: 10.1161/01.cir.0000023043.02648.51.
j.freeradbiomed.2017.05.001.
56. Lenin, R., Nagy, P. G., Jha, K. A., and
51. Ido, Y., Carling, D., and Ruderman, N.
(2002)
Gangaraju, R. (2019) GRP78 translocation to the
Hyperglycemia-induced apoptosis in human umbilical
cell surface and O-GlcNAcylation of VE-Cadherin
vein endothelial cells: inhibition by the AMP-activated
contribute to ER stress-mediated endothelial per-
protein kinase activation, Diabetes,
51,
159-167,
meability, Sci. Rep., 9, 10783, doi: 10.1038/s41598-
doi: 10.2337/diabetes.51.1.159.
019-47246-w.
LONG-TERM EXPERIMENTAL HYPERGLYCEMIA
DOES NOT IMPAIR MACROVASCULAR ENDOTHELIAL BARRIER
INTEGRITY AND FUNCTION in vitro
A. Y. Khapchaev*, O. A. Antonova, O. A. Kazakova, M. V. Samsonov,
A. V. Vorotnikov, and V. P. Shirinsky
Institute of Experimental Cardiology, E. I. Chazov National Medical Research Center for Cardiology,
121552 Moscow, Russia; e-mail: AYKhapchaev@cardio.ru
Hyperglycemia is a hallmark of type 2 diabetes implicated in vascular endothelial dysfunction and car-
diovascular complications. Many in vitro studies identified endothelial apoptosis as an early outcome of
experimentally modeled hyperglycemia emphasizing cell demise as a significant factor of vascular injury.
However, endothelial apoptosis has not been observed in vivo until the late stages of type 2 diabetes.
Here, we studied the long-term (up to 4 weeks) effects of high glucose (HG, 30 mM) on human umbilical
vein endothelial cells (HUVEC) in vitro. HG did not alter HUVEC monolayer morphology, ROS levels,
NO production, and exerted minor effects on the HUVEC apoptosis markers. The barrier responses to var-
ious clues were indistinguishable from those by cells cultured in physiological glucose (5 mM). Tackling
the key regulators of cytoskeletal contractility and endothelial barrier revealed no differences in the his-
tamine-induced intracellular Ca2+ responses, nor in phosphorylation of myosin regulatory light chain or
myosin light chain phosphatase. Altogether, these findings suggest that vascular endothelial cells may well
tolerate HG for relatively long exposures and warrant further studies to explore mechanisms involved in
vascular damage in advanced type 2 diabetes.
Keywords: HUVEC, endothelial barrier, hyperglycemia, insulin, ROS, NO, myosin RLC, MYPT1
БИОХИМИЯ том 88 вып. 8 2023