БИОХИМИЯ, 2023, том 88, вып. 8, с. 1412 - 1422
УДК 577.12
ЗАЩИТНЫЕ ЭФФЕКТЫ ПЕРОКСИРЕДОКСИНА 6
ПРИ МОДЕЛИРОВАНИИ ПРОВОСПАЛИТЕЛЬНОГО ОТВЕТА
С ИСПОЛЬЗОВАНИЕМ МАКРОФАГОВ RAW 264.7
© 2023 С.Б. Парфенюк1*, О.В. Глушкова1, М.Г. Шарапов1, М.О. Хренов1,
С.М. Лунин1, А.А. Кузекова1, Э.К. Мубаракшина1, Т.В. Новоселова1,
Д.А. Черенков2, Е.Г. Новоселова1
1 Институт биофизики клетки ФИЦ ПЦБНИ РАН,
142290 Пущино, Московская обл., Россия; электронная почта: lana_kras2@rambler.ru
2 ФГБОУ ВО Воронежский государственный университет инженерных технологий,
394036 Воронеж, Россия
Поступила в редакцию 05.06.2023
После доработки 14.07.2023
Принята к публикации 15.07.2023
Целью работы было изучение действия пероксиредоксина 6 (PRDX6), рекомбинантного белка-
антиоксиданта, на уровень провоспалительных ответов, вызванных воздействием эндотоксина
на макрофаги RAW 264.7. Добавление липополисахаридов (ЛПС) в среду культивирования клеток
RAW 264.7 ожидаемо увеличивало продукцию фактора некроза опухоли-альфа (TNF-α), а добавле-
ние PRDX6 привело к достоверному снижению его продукции на 15-20%. Уровень продукции дру-
гого провоспалительного цитокина, IL-1β, значительно повышенный эндотоксином, под воздей-
ствием PRDX6 полностью нормализовался. Кроме того, добавление PRDX6 снижало продукцию
активных форм кислорода, индуцированную эндотоксином, а также препятствовало сверхэкс-
прессии гена Nos2 в клетках RAW 264.7. Результаты показали, что PRDX6 оказывает подавляющее
действие на экспрессию гена Nfe2l2 и продукцию фактора транскрипции NRF-2 в течение первых
6 ч культивирования клеток. Добавление эндотоксина вызывает активацию сигнальных каскадов
NF-κB и SAPK/JNK, при этом в присутствии PRDX6 происходит снижение активности этих сиг-
нальных каскадов. Известно, что провоспалительный ответ клеток, вызванный бактериальным
ЛПС, приводит к активации апоптоза и элиминации повреждённых клеток. Нашими исследова-
ниями это подтверждается, поскольку ЛПС приводит к активации гена Trр53, маркера апоптоза.
Добавление PRDX6 в первые часы развития острого провоспалительного ответа приводит к подав-
лению экспрессии гена Trр53, что указывает на защитный эффект PRDX6, снижающий апоптоз
в макрофагах RAW 264.7.
КЛЮЧЕВЫЕ СЛОВА: пероксиредоксин 6, воспаление, цитокины, экспрессия генов, сигнальные каскады.
DOI: 10.31857/S0320972523080092, EDN: IKLJLS
ВВЕДЕНИЕ
ких (абсцесс, гангрена), а также хронических
неспецифических заболеваний лёгких (хрони-
Воспаление является универсальной за-
ческая обструктивная болезнь лёгких (ХОБЛ),
щитной реакцией на любое повреждение тка-
бронхиальная астма и другие) [1]. По этой
ни, однако важное значение имеет уровень
причине в настоящее время необходимы ис-
провоспалительного ответа. Известно, что его
следования, направленные на изучение внут-
высокий уровень может вызывать множество
риклеточных механизмов развития различных
критических состояний, включая сепсис, де-
заболеваний, в частности болезней лёгких.
структивный панкреатит и другие. Воспале-
Альвеолярные макрофаги - клетки, иг-
нию отводится ключевая роль в патогенезе
рающие центральную роль в воспалении и
большинства лёгочных заболеваний, таких как
регулирующие сложные межклеточные взаи-
пневмония, деструктивные заболевания лёг- модействия. При активации альвеолярных
Принятые сокращения: АФК - активные формы кислорода; ЛПС - липополисахарид; NF-κB - ядерный фактор-κB;
PRDX6 - пероксиредоксин 6; TNF-α - фактор некроза опухоли-альфа.
* Адресат для корреспонденции.
1412
ЗАЩИТНЫЕ ЭФФЕКТЫ ПЕРОКСИРЕДОКСИНА 6
1413
макрофагов реализуются чрезвычайно важные
леваниях. С одной стороны, фосфолипазный
события врождённого иммунитета, в частно-
активный центр (aiPLA2) необходим для акти-
сти, фагоцитоз, презентация антигена Т-лим-
вации провоспалительной NADPH-оксидазы.
фоцитам, секреция лизоцима и активных фер-
С другой стороны, пероксидазная активность
ментов, активация системы комплемента [2].
PRDX6 имеет решающее значение для проти-
Именно поэтому исследование молекулярно-
водействия повышенному образованию АФК
клеточных механизмов активации альвеоляр-
и для восстановления окисленных клеточных
ных макрофагов имеет большое значение для
мембран после окислительного стресса [7]. В
понимания механизмов развития воспаления
настоящее время рецептор PRDX6 достоверно
лёгочной ткани.
не определён, однако в работе Sharapov et al. [8]
Воспалительные реакции, вызванные ли-
показано, что трансмембранный транспорт
пополисахаридами (ЛПС) грамотрицательных
PRDX6 осуществляется при участии рецептора
бактерий, характеризуются мощным иммун-
TLR4. Гомеостаз окислительно-восстановитель-
ным ответом с участием Toll-подобных ре-
ных реакций жизненно важен для регуляции
цепторов (TLR). TLR относятся к семейству
воспалительных состояний, и нарушение его
паттерн-распознающих рецепторов (PRRs)
регуляции может влиять на патогенез воспа-
и осуществляют распознавание молекуляр-
лительных заболеваний дыхательных путей.
ных структур патогенов экзогенных (PAMPs,
Известно, что PRDX6 является важным внутри-
патоген-ассоциированные молекулярные пат-
клеточным ферментом-антиоксидантом, спо-
терны) и ряда эндогенных лигандов (DAMPs,
собным к значительной экспрессии в дыха-
ассоциированные с повреждениями молеку-
тельных путях и лёгких здоровых людей [9].
лярные паттерны - молекулы, которые выс-
В настоящее время имеется достаточное ко-
вобождаются в ответ на повреждение клеток),
личество сведений, которые дают основания
обеспечивая быструю реакцию клетки. Среди
полагать, что пероксиредоксины в перспекти-
TLR первым был идентифицирован TLR4,
ве будут применяться в терапии заболеваний,
который участвует в распознавании ЛПС. Сиг-
патогенез которых связан с окислительным
нальный путь TLR4, опосредованный узна-
стрессом. К примеру, в ряде работ показана
ванием внеклеточного эндотоксина, также
защитная роль пероксиредоксинов при раз-
включает белок миелоидной дифференциров-
личных патологиях: кожи [10, 11], лёгких [12],
ки 2 (MD2) и CD14 [3]. Взаимодействие ЛПС
глаз [13], нервной системы [14], поджелудоч-
с поверхностными рецепторами макрофагов
ной железы [15].
активирует ядерный фактор-κB (NF-κB), ко-
Исходя из актуальности проблемы, в на-
торый участвует в провоспалительном ответе.
стоящей работе с использованием линии мак-
Известно, что значимую роль в развитии вос-
рофагоподобных клеток мыши RAW
264.7
паления играют активные формы кислорода
были исследованы эффекты PRDX6 при сти-
(АФК). Несмотря на то что АФК являются
мулировании клеток ЛПС грамотрицатель-
важными регуляторными молекулами на всех
ных бактерий. Концентрация ЛПС 1 мкг/мл
этапах воспаления, их чрезмерная продук-
используется в мировой практике как приво-
ция при несвоевременной или недостаточной
дящая к острому лёгочному воспалению [16].
работе антиоксидантных систем приводит к
Добавление ЛПС к макрофагам RAW 264.7
переходу воспаления в хроническую стадию,
моделирует условия воспаления дыхательных
которая, в свою очередь, лежит в основе раз-
путей, вызванного бактериальной инфекцией,
личных заболеваний [4].
а также может рассматриваться в качестве мо-
Пероксиредоксин 6 (PRDX6) - предста-
дели активации макрофагальных клеток при
витель 1Cys-пероксиредоксинов, содержащих
различных неинфекционных поражениях ды-
единственный активный остаток цистеина
хательной системы.
(Cys47) - был обнаружен во всех клетках мле-
Целью работы было изучение эффектов
копитающих, однако его наибольшие количе-
PRDX6, рекомбинантного белка-антиоксидан-
ства выявлены в цитоплазме эпителиальных
та, на уровень провоспалительных ответов, вы-
клеток: лёгких, дыхательных путей, желудоч-
званных воздействием эндотоксина на макро-
но-кишечного тракта и ротовой полости, а
фаги RAW 264.7.
также в клетках печени и поджелудочной желе-
зы [5]. Исследования рекомбинантных белков
PRDX6 человека и крысы подтвердили нали-
МАТЕРИАЛЫ И МЕТОДЫ
чие у них как пероксидазной, так и фосфоли-
пазной активности [6]. PRDX6, по-видимому,
Культивирование клеток RAW 264.7. Клетки
играет двойную роль в воспалительных забо-
RAW 264.7 растили в культуральных флаконах
БИОХИМИЯ том 88 вып. 8 2023
1414
ПАРФЕНЮК и др.
в среде для культивирования, состоящей из
значение. Усреднённые значения по четырём
смеси RPMI : DMEM («ПанЭко», Россия) в со-
экспериментам использовали для определе-
отношении 1 : 1, с добавлением 10%-ной (v/v)
ния достоверности различий между группами
эмбриональной телячьей сыворотки, 2,04 мМ
(n = 4). В качестве контролей использовали
L-глютамина («ПанЭко») и раствора антибио-
клетки, не подвергавшиеся обработке.
тика-антимикотика («Sigma», США) в концен-
Определение уровня продукции АФК в клет-
трации, рекомендованной производителем,
ках. Уровень генерации АФК в клетках изме-
при 37 °C и 5% СО2. Пересев (пассаж) клеток
ряли с помощью диацетилового производного
производили после достижения плотности
5,6-карбокси-2′,7′-дихлордигидрофлуоресцеи-
монослоя ~80%. В экспериментах использо-
на (карбокси-H2DCFDA, «Invitrogen», США).
вались клетки после 4-8 пассажей. Жизнеспо-
Карбокси-H2DCFDA - химически восстанов-
собность клеток RAW 264.7 оценивалась визу-
ленный аналог флуоресцеина, который исполь-
ально, с использованием инвертированного
зуют в качестве общего индикатора окисли-
микроскопа LEICA DMIL HC («Leica», Герма-
тельного стресса в клетках. Клетки RAW 264.7
ния) с применением окрашивания 0,01%-ным
предварительно культивировали в течение 24 ч
(m/v) раствором трипанового синего («Appli-
в 96-луночном планшете (2,5 × 104 клеток на
Chem», Германия) в физиологическом рас-
лунку) в 100 мкл среды DMEM, промывали
творе
(0,9%-ный (m/v) NaCl) и подсчётом
PBS и обрабатывали свежеприготовленным
живых клеток в камере Горяева. Количество
раствором карбокси-H2DCFDA («Invitrogen») в
жизнеспособных макрофагальных клеток со-
стерильном ДМСО в конечной концентрации
ставляло 93-97%. Провоспалительный ответ
2,5 мкМ в среде с 2%-ной (v/v) эмбриональной
индуцировали ЛПС, выделенным из стенок
телячьей сывороткой. Далее клетки инкубиро-
Escherichia coli O55:B5 («Sigma») в концентра-
вали с карбокси-H2DCFDA в темноте в тече-
ции 1 мкг/мл, экзогенный PRDX6 добавляли в
ние 1 ч. Одновременно с добавлением карб-
концентрации 150 мкг/мл одновременно с ин-
окси-H2DCFDA добавляли ЛПС (1 мкг/мл) и
дукцией провоспалительных ответов. Ген, ко-
PRDX6 (150 мкг/мл).
дирующий PRDX6 дикого типа, был клони-
В качестве контроля использовали сигнал
рован и экспрессирован в клетках E. coli BL21
флуоресценции от необработанных клеток
(DE3). Белок был очищен с помощью металло-
RAW 264.7. Фоновый сигнал культуральной
аффинной хроматографии (Ni-NTA-агароза)
среды вычитали из соответствующих сигна-
в соответствии с ранее описанной процеду-
лов экспериментальных образцов. Флуорес-
рой [17]. Для определения продукции цито-
ценцию измеряли с помощью флуоресцент-
кинов клетки снимали с поверхности культу-
ного планшетного ридера Infinite 200 («Tecan»,
рального флакона с помощью скребков для
Австрия) при возбуждении длиной волны
клеток («Corning», США), избавляли от старой
480 нм и с длиной волны поглощения 530 нм
среды путём промывания, разводили в свежей
(апроксимальное возбуждение флуоресценции
среде и рассевали с плотностью 106 клеток в
(Ex)/испускание (Em) = Ex/Em = 485/535 нм),
1 мл в 24-луночные планшеты. По истечении
как описано ранее [18]. Проводили три неза-
24 ч в каждую лунку добавляли исследуемые
висимых эксперимента, в каждом из которых
вещества в заданной концентрации. Завершив
было по 3 повтора на каждую эксперименталь-
72-часовую инкубацию, клетки лизировали
ную группу (n = 9 в каждой группе).
3 раза замораживанием-оттаиванием, при этом
Определение продукции цитокинов. Приме-
лизаты использовали немедленно либо хра-
няли наборы для ИФА для определения мы-
нили при -20 °С. Для определения экспрес-
шиного TNF-α, IL-10 и IL-1β («PeproTech»,
сии генов и продукции сигнальных и стрес-
США). Для визуализации связывания исполь-
совых белков клетки рассевали в культураль-
зовали зелёный краситель ABTS («Sigma») -
ные флаконы площадью 25 см2 с плотностью
100 мкл, растворённые в 0,05 М цитратном
100 × 106 клеток на флакон. После прикреп-
буфере (pH 5,0) с добавлением
0,01%-ного
ления клеток в течение 24 ч добавляли иссле-
пероксида водорода - и измеряли оптическую
дуемые вещества по приведённой выше схеме
плотность при 405 нм с помощью спектрофо-
и инкубировали 6 ч. Далее клетки снимали с
тометра для микропланшетов (Multiskan EX,
поверхности флакона, отмывали от старой
«Thermo Electron Corporation», США).
среды и разводили в 1 мл физиологического
Ds-Na-ПААГ-электрофорез и иммуноблот-
раствора для дальнейшего выделения РНК или
тинг. Для приготовления белковых образцов
белков. В пределах каждого независимого экс-
из клеток RAW 264.7 клеточную суспензию,
перимента характеристики образцов измеряли
предварительно отмытую от культуральной
параллельно в 6-9 повторах, получая среднее
среды 0,9%-ного (m/v) NaCl, подвергали трёх-
БИОХИМИЯ том 88 вып. 8 2023
ЗАЩИТНЫЕ ЭФФЕКТЫ ПЕРОКСИРЕДОКСИНА 6
1415
кратному замораживанию-оттаиванию. Перед
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
циклами замораживания-оттаивания к суспен-
зии добавляли ингибиторы протеаз и фосфа-
В настоящее время известно, что PRDX6
таз (100×) по рекомендациям производителя
вносит существенный вклад в защиту орга-
(«Thermo Fisher Scientific», США). Далее до-
нов дыхания от воспаления, вызванного ЛПС.
бавляли солюбилизирующий буфер для элек-
Новоселов в 2012 г. [21] показал увеличение
трофореза и β-меркаптоэтанол (2-5%), кипя-
секреции PRDX6 в эпителии трахеи и брон-
тили 5 мин. Концентрацию белка измеряли с
хов сразу после аппликации ЛПС и сохране-
помощью спектрофотометра NanoDrop2000
ние высокого уровня секреции на протяжении
(«NanoDrop», США), образцы наносились
всего периода воспаления. Между тем внутри-
по 10 мкл в каждую лунку с 10%-ным (m/v)
клеточные механизмы влияния экзогенного
Ds-Na-ПААГ [19]. Наличие белков в образцах
PRDX6 на процесс ЛПС-индуцированной ак-
определяли методом Вестерн-блот анализа с
тивации альвеолярных макрофагов до сих пор
использованием наборов следующих антител:
остаются малоизученными.
кроличьи антитела к общему NF-κB (total-
Влияние PRDX6 на продукцию цитокинов
NF-κB) и фосфорилированному по Ser536
клетками RAW 264.7 при воздействии эндоток-
(ph-NF-κB (Ser536)), p53, phSAPK/JNK, NRF-2,
сина. Известно, что в очаге воспаления проис-
TLR4, глицеральдегид-3-фосфатдегидрогеназе
ходит увеличение пролиферации макрофагов.
(GAPDH) («Cell Signaling», США). Для выяв-
Это является защитной функцией организма,
ления белков использовали систему ECL («GE
направленной на борьбу с воспалительными
Healthcare», Швеция). Фотографии полос ре-
агентами разной природы, в частности бакте-
гистрировали с помощью трансиллюминатора
риальной. С другой стороны, чрезмерное уве-
TFX 35.WL («Vilber Lourmat», Франция). Коли-
личение количества альвеолярных макрофагов
чественную оценку белков после денситоме-
в тканях лёгких, вырабатывающих факторы,
трии проводили с использованием программы
участвующие в формировании фиброзной
Qapa (Ver. 3.7). Было проведено по 3 независи-
ткани, такие как фибронектин, IL-1 и другие,
мых эксперимента (используя клетки от разных
приводит к необратимым повреждениям орга-
пассажей) для каждого белка. Полученные ци-
на. Необходимо отметить, что стойкое увели-
фровые данные нормировали к соответствую-
чение количества альвеолярных макрофагов
щему контролю нагрузки (полосам GAPDH) и
лежит в основе формирования хронических
выражали в относительных единицах.
воспалительных заболеваний дыхательных пу-
Оценка изменений экспрессии генов. Экс-
тей за счёт увеличения количества рецепторов
прессию генов анализировали в клетках
IgE, продукции хемоаттрактантов, привлекаю-
RAW 264.7 методом ОТ-ПЦР в режиме реаль-
щих нейтрофилы и лимфоциты, в свою оче-
ного времени. После воздействия исследуе-
редь стимулирующие макрофаги [22].
мых веществ, а именно экзогенного PRDX6
Важным маркером развития провоспа-
(150 мкг/мл), ЛПС (1 мкг/мл) или их совмест-
лительного ответа является продукция ряда
ного добавления в среду культивирования кле-
про- и антивоспалительных цитокинов. В ра-
ток через 6 ч инкубации были исследованы
боте были измерены уровни продукции про-
гены, кодирующие ряд ферментов антиокси-
воспалительных цитокинов, таких как TNF-α
дантного ответа (Sod3, Prdx1, Nos2), апоптоз
и IL-1β, а также продукция антивоспалитель-
(Trp53), факторы транскрипции (Nfe2l2, Nfκb1,
ного цитокина IL-10 (рис. 1). Параллельно ис-
Ap1m1), рецептор (Tlr4), цитокин (Tnfsf18).
следовали уровень экспрессии гена Tnfsf18 на
Процедура выделения тотальной РНК,
первичном этапе провоспалительного ответа
синтеза кДНК и организация ПЦР в режиме
(таблица).
реального времени подробно описана нами
Добавление ЛПС в среду культивирования
ранее [20].
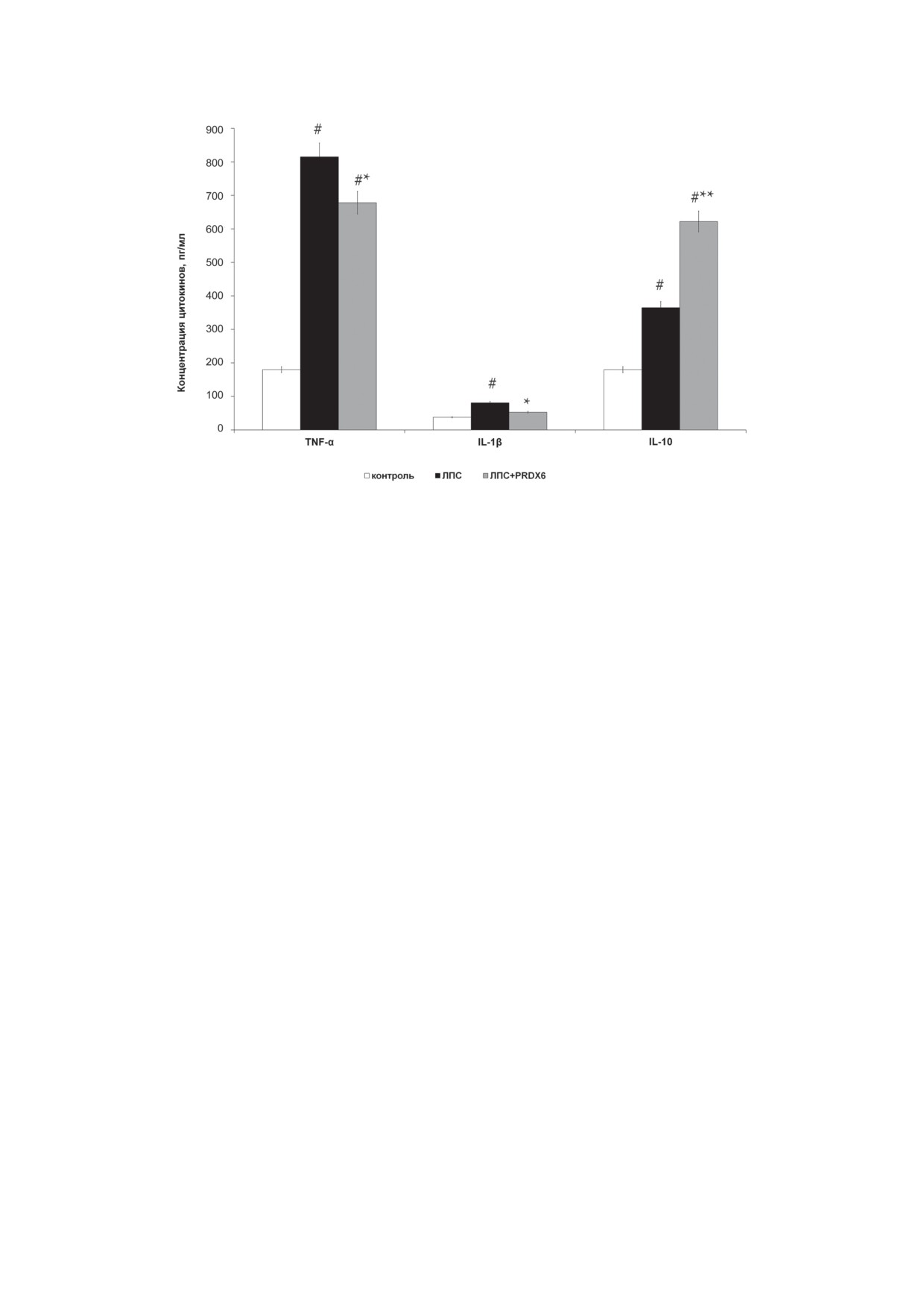
клеток RAW 264.7 ожидаемо увеличило про-
Статистический анализ проводили с ис-
дукцию TNF-α до 450% от контрольных зна-
пользованием программного обеспечения
чений, а добавление PRDX6 привело к незна-
Statistica 6.0 («StatSoft», США). Для определе-
чительному, но достоверному снижению его
ния достоверности различий применяли одно-
продукции на 15-20%. Путём исследования
факторный дисперсионный анализ (ANOVA) с
уровня экспрессии гена Tnfsf18 было показано,
апостериорным критерием Тьюки. Значимыми
что экзогенный PRDX6 в первые 6 ч способен
считали различия при p < 0,05. Для проверки
удерживать уровень экспрессии данного гена
нормальности распределения использовали
на уровне контроля.
критерий Шапиро-Уилка. Значение W во всех
Уровень продукции другого провоспали-
случаях было незначимым (p > 0,05).
тельного цитокина, IL-1β, значительно акти-
БИОХИМИЯ том 88 вып. 8 2023
1416
ПАРФЕНЮК и др.
Рис. 1. Влияние экзогенного PRDX6 на продукцию цитокинов в макрофагах RAW 264.7. Средние значения (± стандарт-
ная ошибка) изменения уровней продукции цитокинов представлены по отношению к уровням в клетках RAW 264.7
в нормальных условиях. # p < 0,05, изменения статистически значимы для групп «ЛПС + PRDX6» и «ЛПС» отно-
сительно группы «контроль»; ** р < 0,01, * p < 0,05, изменения статистически значимы для группы «ЛПС + PRDX6»
относительно группы «ЛПС»
вированный эндотоксином, под воздействием
времени (от 0 до 8 ч). При этом добавление
PRDX6 практически возвращается к контроль-
PRDX6 полностью блокирует его увеличение,
ному значению.
удерживая концентрацию АФК на уровне та-
Антивоспалительные цитокины на ранних
ковой для клеток, не подвергающихся ника-
этапах развития провоспалительного ответа
кому воздействию (рис. 2). Таким образом,
ограничивают повреждение здоровой ткани
было установлено, что PRDX6 стабилизирует
и обеспечивают баланс между физиологиче-
антиоксидантный статус клетки, нарушенный
ской реакцией и патологическим воспалени-
в условиях провоспалительного ответа макро-
ем. Ключевым антивоспалительным цитоки-
фагов RAW 264.7, индуцированного ЛПС.
ном является IL-10, который противодействует
При изучении экспрессии гена Nos2 было
эффекту основных провоспалительных цито-
показано, что в условиях воспаления в первые
кинов. Положительным эффектом в борьбе с
6 ч PRDX6 препятствует его сверхэкспрессии
воспалением можно считать тот факт, что до-
(таблица).
бавление PRDX6 двукратно увеличивает про-
Влияние экзогенного PRDX6 на экспрессию
дукцию IL-10.
генов системы антиоксидантной защиты (Sod3,
Полученные нами данные по увеличению
Prdx1). При взаимодействии клеток RAW 264.7
продукции TNF-α, IL-1β и IL-10 подтвер-
с бактериальным ЛПС происходит активация
ждают тот факт, что в условиях воспаления,
ферментных систем антиоксидантной защиты.
вызванного воздействием ЛПС, активация мак-
Наблюдается повышение экспрессии генов та-
рофагов проходит по так называемому класси-
ких ферментов, как внеклеточная супероксид-
ческому пути с приобретением фенотипа М1
дисмутаза 3. Этот фермент является участни-
(провоспалительный). Результаты показали,
ком системы быстрой защиты, и поэтому его
что экзогенный рекомбинантный PRDX6 яв-
максимальная активность проявляется в пер-
ляется перспективным антивоспалительным
вые часы развития провоспалительного ответа
агентом, способным частично снижать уровень
при резком повышении АФК [23]. Экзоген-
патологических реакций, связанных с чрезмер-
ный PRDX6 препятствует активации экспрес-
ным уровнем продукции провоспалительных
сии гена супероксиддисмутазы 3. Вероятно,
цитокинов.
что PRDX6, добавленный в среду культиви-
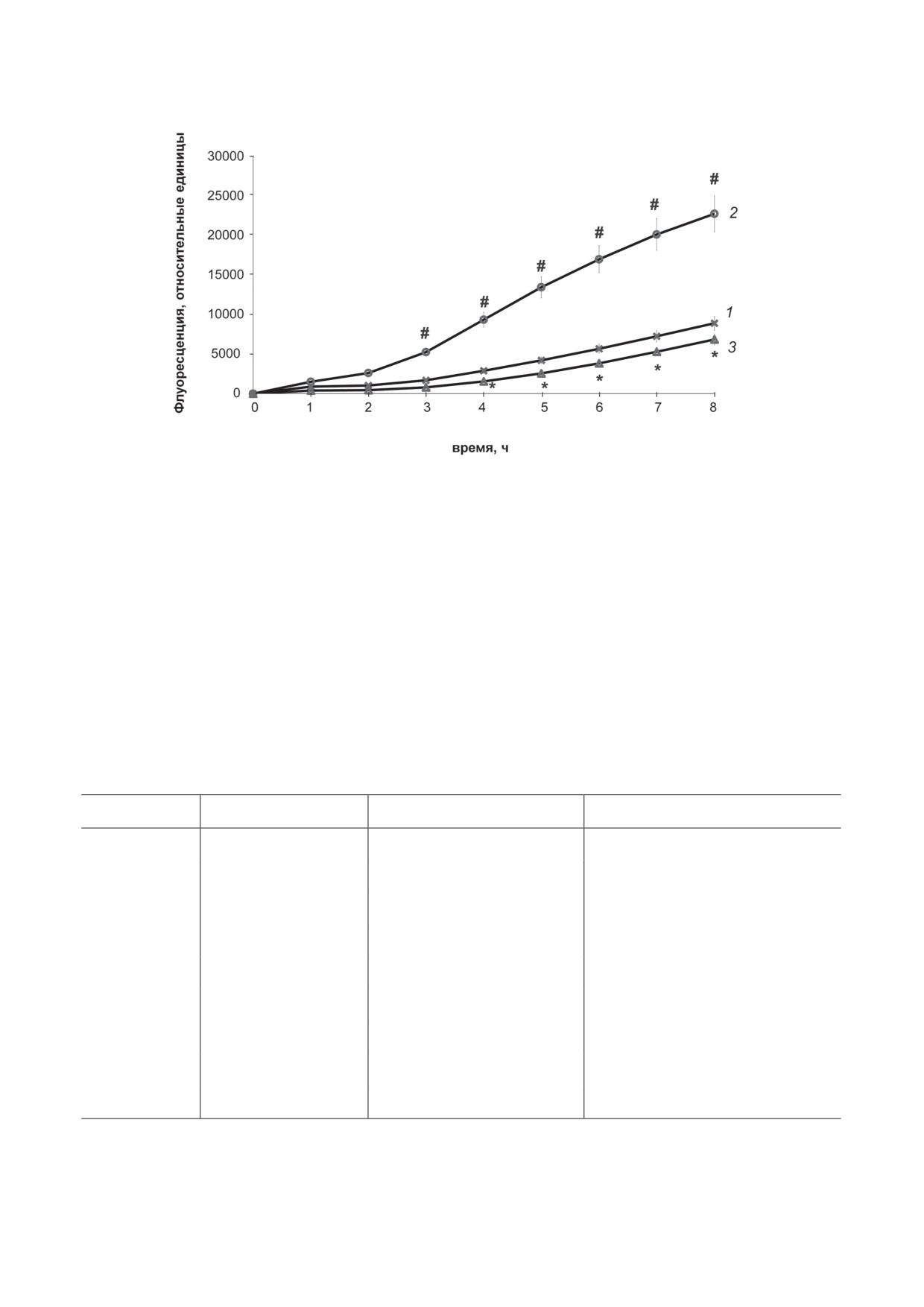
Влияние PRDX6 на генерацию АФК в
рования активированных ЛПС макрофагов,
клетках RAW
264.7, стимулированных ЛПС.
является достаточным для антиоксидантной
В условиях воспаления показан высокий уро-
защиты клетки, защищая её от повреждения и
вень АФК, который увеличивается с течением
апоптоза, вызванных гиперпродукцией АФК.
БИОХИМИЯ том 88 вып. 8 2023
ЗАЩИТНЫЕ ЭФФЕКТЫ ПЕРОКСИРЕДОКСИНА 6
1417
Рис. 2. Влияние PRDX6 на продукцию АФК клетками RAW 264.7. Средние значения (± стандартная ошибка) изме-
нения уровня флуоресценции карбокси-H2DCFDA в клетках RAW 264.7. 1 - Нормальные клетки, группа «контроль»;
2 - клетки, стимулированные ЛПС, группа «ЛПС»; 3 - клетки, культивированные в среде с одновременным добавле-
нием ЛПС и PRDX6, группа «ЛПС + PRDX6». # p < 0,05, изменения статистически значимы для группы «ЛПС» отно-
сительно группы «контроль»; * p < 0,05, изменения статистически значимы для группы «ЛПС + PRDX6» относительно
группы «ЛПС»
Это подтверждается и количеством АФК в
положить, что добавленный в начале культи-
клетках при добавлении PRDX6, которое не
вирования экзогенный PRDX6 используется
повышается в присутствии ЛПС.
клетками для защиты от АФК, в свою очередь
Исследовав экспрессию гена Prdx1, мы
не позволяя PRDX1 проявить свои провоспа-
пришли к выводу, что ЛПС вызывает актива-
лительные свойства (таблица).
цию его экспрессии. Добавление экзогенного
Влияние PRDX6 на внутриклеточную сигна-
PRDX6 снижает активацию экспрессии гена
лизацию в макрофагах RAW 264.7 при воздей-
Prdx1 в первые 6 ч инкубации. Логично пред-
ствии эндотоксина. Интересные данные были
Изменение уровня экспрессии (в отн. ед.) некоторых маркерных генов в нормальных и стимулированных эндотоксином
клетках RAW 264.7 через 6 ч инкубации с PRDX6
Гены
Контроль
ЛПС
ЛПС + PRDX6
Tnfsf18
1 ± 0,087
4,95 ± 0,361#
1,2 ± 0,088**
Nos2
1 ± 0,059
8 ± 0,688#
3,6 ± 0,259#**
Nfκb1
1 ± 0,051
3,7 ± 0,316#
0,8 ± 0,054**
Tlr4
1 ± 0,095
2,4 ± 0,195#
0,25 ± 0,011#**
Nfe2l2
1 ± 0,072
1,2 ± 0,086
0,4 ± 0,024#*
Ap1m1
1 ± 0,058
6,5 ± 0,498#
0,25 ± 0,021#**
Trp53
1 ± 0,089
8,5 ± 0,615#
0,2 ± 0,017#**
Sod3
1 ± 0,062
5,2 ± 0,337#
0,3 ± 0,024#**
Prdx1
1 ± 0,077
2,48 ± 0,156#
1,2 ± 0,121*
Примечание. Средние значения (± стандартная ошибка) изменения уровней экспрессии генов представлены по отно-
шению к уровням в клетках RAW 264.7 в нормальных условиях. # p < 0,05, изменения статистически значимы для
группы «ЛПС + PRDX6» и «ЛПС» относительно группы «контроль»; ** р < 0,01, * p < 0,05, изменения статистически
значимы для группы «ЛПС + PRDX6» относительно группы «ЛПС».
БИОХИМИЯ том 88 вып. 8 2023
1418
ПАРФЕНЮК и др.
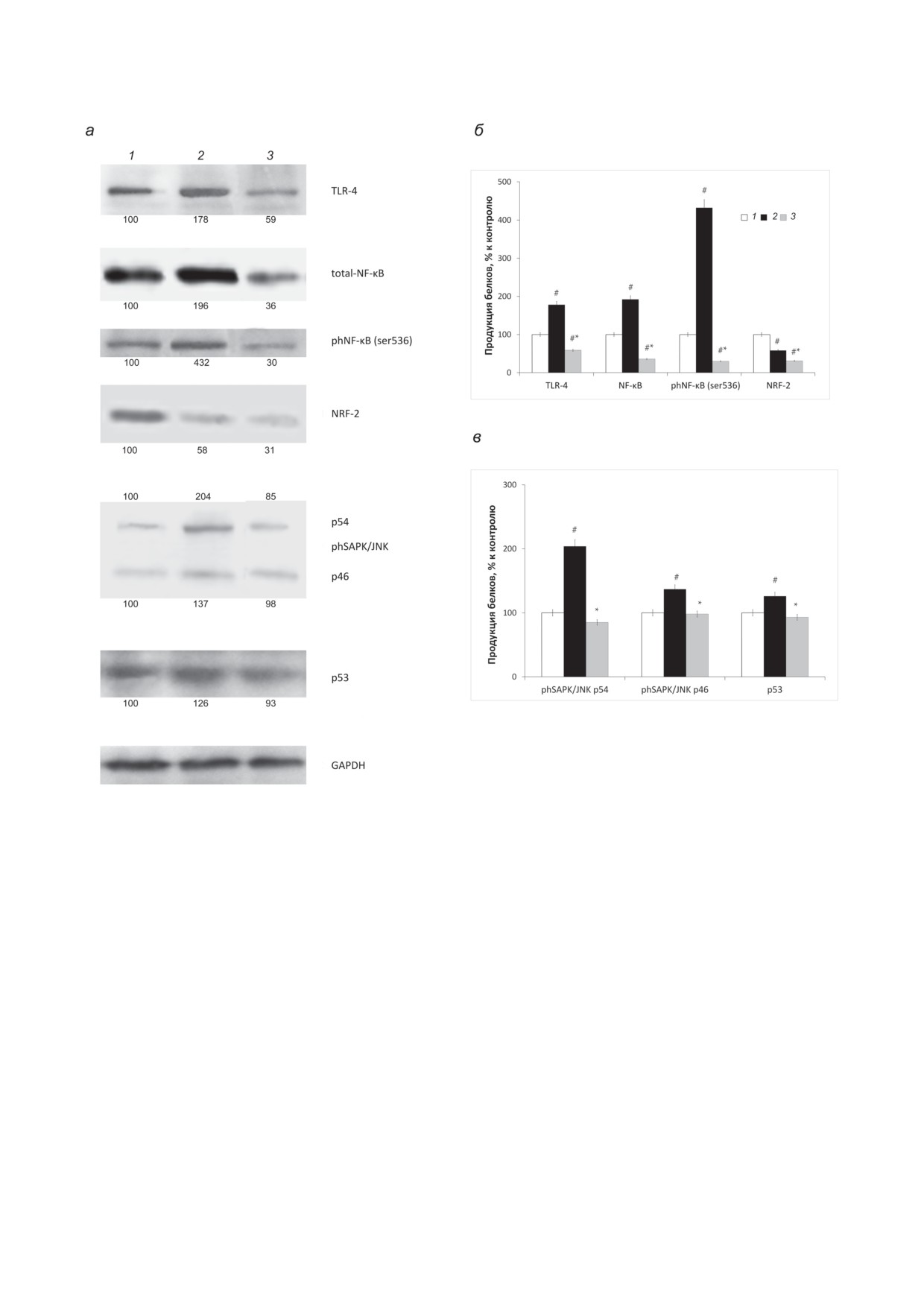
Рис. 3. Влияние PRDX6 на продукцию TLR4, NF-κB, phNF-κB (Ser536), NRF-2, phSAPK/JNK, р53 и референсного
белка GAPDH. Числовые значения внизу - данные денситометрического анализа, средние значения (± стандарт-
ная ошибка) изменения уровней продукции цитокинов представлены по отношению к уровням в клетках RAW 264.7
в нормальных условиях. а - Репрезентативные фотографии результатов Вестерн-блот анализа с использованием спе-
цифических антител к данным белкам; б, в - графическое представление результатов денситометрического измерения
количества белка (в % от контроля). Условные обозначения: 1 - нормальные клетки, группа «контроль»; 2 - клет-
ки, стимулированные ЛПС, группа «ЛПС»; 3 - клетки, культивированные в среде с одновременным добавлением
ЛПС и PRDX6, группа «ЛПС + PRDX6». Каждое значение - среднее от трёх независимых экспериментов, на рисунке
представлены результаты одного из трёх экспериментов. # p < 0,05, изменения статистически значимы для группы
«ЛПС + PRDX6» и «ЛПС» относительно группы «контроль»; * p < 0,05, изменения статистически значимы для группы
«ЛПС + PRDX6» относительно группы «ЛПС»
получены при изучении экспрессии гена Tlr4
характерного для провоспалительного ответа
рецептора TLR4 (таблица) и непосредствен-
(рис. 3).
но продукции самого рецептора (рис. 3). Так,
ЛПС-Активация рецептора TLR4 при-
было показано, что ЛПС стимулирует экспрес-
водит к последующей активации сигнального
сию Tlr4 (широко известный факт), а экзоген-
каскада NF-κB. Было установлено значитель-
ный PRDX6 полностью снимает эту стимуля-
ное увеличение экспрессии гена Nfkb1 в ответ
цию. C использованием Вестерн-блот анализа
на присутствие ЛПС. Интересно, что в первые
мы продемонстрировали аналогичные резуль-
6 ч добавление PRDX6 удерживает уровень
таты. Так, через 6 ч инкубации с PRDX6 клет-
экспрессии гена Nfkb1 в клетках активирован-
ки, стимулированные ЛПС, показали резкое
ных ЛПС на контрольном уровне (таблица).
снижение продукции TLR4 до 30% от уровня,
Результаты Вестерн-блот анализа показали,
БИОХИМИЯ том 88 вып. 8 2023
ЗАЩИТНЫЕ ЭФФЕКТЫ ПЕРОКСИРЕДОКСИНА 6
1419
что через 6 ч после добавления ЛПС присут-
это подтверждается, поскольку ЛПС приводит
ствие PRDX6 снижает продукцию как общего,
к активации гена Trр53, маркера апоптоза.
так и фосфорилированного по Ser536 NF-κB
Добавление PRDX6 в первые часы развития
до контрольных значений. Известно, что фос-
острого провоспалительного ответа приводит
форилирование NF-κB по Ser536 активирует
к подавлению экспрессии гена Trр53 (таблица).
клеточный апоптоз, таким образом, PRDX6
Аналогичные эффекты нами показаны с помо-
проявляет себя как антиапоптозный агент. Эти
щью Вестерн-блот анализа. Так, продукция
результаты свидетельствуют о том, что PRDX6
белка p53 в присутствии PRDX6 остаётся на
обладает ингибирующим влиянием на сиг-
уровне контроля в макрофагах RAW 264.7, сти-
нальный каскад NF-κB. Его действие заметно
мулированных эндотоксином (рис. 3). Можно
по экспрессии соответствующего гена и про-
предположить, что эффект PRDX6 заключа-
дукции как общего, так и фосфорилирован-
ется в том, чтобы не дать в первые часы раз-
ного по Ser536 белка (рис. 3).
виться массовому апоптозу макрофагов, необ-
NRF-2 участвует в сложной регуляторной
ходимых для местной защиты ткани лёгких, но
сети и выполняет плейотропную роль в регу-
при «взятии под контроль» воспалительного
ляции метаболизма, воспаления, аутофагии,
процесса данный фермент способствует эли-
протеостаза, митохондриальных, физиологи-
минации повреждённых макрофагов, которые
ческих и иммунных ответов [24]. Интересно,
уже не могут быть «реанимированы».
что продукция NRF-2 при действии ЛПС
почти в 2 раза ниже, чем в контрольных клет-
ках. Это согласуется с тем, что NRF-2 и NF-κB
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
действуют друг на друга по принципу отрица-
тельной обратной связи. И так как в «класси-
Исследование ряда патологий, сопряжён-
ческих» условиях воспаления активен каскад
ных с окислительным стрессом, показало, что
NF-κB, то количество NRF-2 в этих условиях
защитные свойства PRDX6 связаны как с его
уменьшается, при этом наблюдается повыше-
пероксидазной активностью, так и с его спо-
ние уровня АФК в клетках, культивированных
собностью выступать в роли сигнально-регу-
с ЛПС. Результаты исследования показали,
ляторной молекулы [20]. Ранее было показано,
что PRDX6 проявляет подавляющее действие
что PRDX6 ингибирует воспалительные заболе-
на экспрессию гена Nfe2l2 и продукцию фак-
вания путём подавления повреждения, вызван-
тора транскрипции NRF-2 в течение первых
ного свободными радикалами и митохондри-
6 ч инкубации. В присутствии PRDX6 не было
альной генерацией H2O2, а также препятствует
выявлено описанных для ЛПС регуляторных
воспалительным заболеваниям посредством
механизмов между NRF-2 и NF-κB, что может
активации NF-κB/AP-1 в сочетании с путём
говорить о том, что фермент-антиоксидант так
JNK [25]. В своей работе, в отличие от предыду-
влияет на эти сигнальные каскады, что они
щих исследований, мы изучали влияние экзо-
теряют свою способность к регулированию
генного PRDX6, тогда как большинство ранее
по принципу отрицательной обратной связи
опубликованных работ было направлено на ис-
(рис. 3).
следование влияния экспрессии собственного
В условиях индукции провоспалительного
PRDX6. Гиперпродукция провоспалительных
ответа активен и сигнальный каскад SAPK/
цитокинов, таких как TNF-α, IL-1β и IL-6,
JNK. Показано, что при этом продукция
является ключевым фактором в развитии пато-
phSAPK/JNK гораздо выше контрольного уров-
генеза многих лёгочных патологий, таких как
ня. Обе изоформы р54 и р46 активны через 6 ч
бронхиальная астма, ХОБЛ, респираторный
инкубации клеток с ЛПС (рис. 3). Добавление
дистресс-синдром взрослых, идиопатический
PRDX6 полностью снимает активацию данного
фиброз лёгких [26]. Поэтому полученные нами
каскада и приводит продукцию phSAPK/JNK к
данные о том, что добавление PRDX6 снижает
норме. Сигнальный каскад SAPK/JNK закан-
продукцию TNF-α и IL-1β, а также увеличива-
чивается активацией гена Ap1m1. Нами было
ет продукцию антивоспалительного цитокина
показано, что PRDX6, добавленный к активи-
IL-10 в макрофагальных клетках, стимулиро-
рованным ЛПС клеткам, практически полно-
ванных ЛПС, являются актуальными для даль-
стью ингибирует экспрессию этого транскрип-
нейшего исследования антивоспалительных
ционного фактора (таблица).
свойств PRDX6. В работе Zhao et al. [27] были
Известно, что провоспалительный ответ
исследованы эффекты всех 6 типов перокси-
клеток, вызванный бактериальным ЛПС, при-
редоксинов и показано, что PRDX1, PRDX2 и
водит к активации апоптоза и элиминации по-
PRDX4 обладают выраженным провоспали-
вреждённых клеток. Нашими исследованиями
тельным действием на макрофаги RAW 264.7,
БИОХИМИЯ том 88 вып. 8 2023
1420
ПАРФЕНЮК и др.
тогда как PRDX6 влияет на них незначи-
Местная клеточная среда играет решающую
тельно. Работа была проведена с использо-
роль в характере этих видов регуляции [32].
ванием нативных клеток, тогда как в своей
Высокая продукция NO действует как про-
работе мы исследовали влияние PRDX6 на
апоптотический модулятор, активируя протеазы
макрофаги, активированные бактериальным
семейства каспаз через высвобождение мито-
эндотоксином. Исходя из полученных нами
хондриального цитохрома c в цитозоль, повы-
результатов, можно сделать вывод о том, что
шенную регуляцию белка р53, активацию сиг-
PRDX6 снижает провоспалительные эффекты
нального пути SAPK/JNK, изменяя экспрессию
эндотоксина на макрофагоподобных клетках
апоптоз-ассоциированных белков семейства
RAW 264.7.
Bcl-2. Тем не менее, низкие или физиологиче-
Сигнальные пути NF-κB и MAPK регули-
ские концентрации NO препятствуют апоп-
руют воспаление и иммунные ответы, контро-
тозу клеток, вызванному Fas и цитокинами.
лируя экспрессию генов провоспалительных
Антиапоптотический механизм связан с тран-
факторов, таких как TNF-α и IL-6. Yuan et al.
скрипцией генов защитных белков (Вcl-2, ци-
в 2019 г. показали, что ядерная транслокация
клооксигеназа-2) и прямым ингибированием
фосфорилированного р65, ингибирование ки-
эффекторных каспаз путём S-нитрозилирова-
назы IκB (IKK) в сигнальном пути NF-κB и
ния тиоловой группы цистеина в их каталити-
фосфорилирование ERK, JNK и p38 MAPK
ческом сайте. Dubey et al. [33] доказали решаю-
(т.е. активация сигнального пути MAPK) мо-
щую роль индуцибельной NO-синтазы (iNOS) в
жет контролировать выработку IgE и IL-4 и
апоптозе нейтрофилов посредством усиленного
ингибировать медиаторы воспаления, харак-
генерирования АФК и опосредованной кас-
терные для бронхиальной астмы [28]. Ранее
пазой-8 активации пути гибели митохондрий.
нами было показано, что PRDX6 активирует
В своей работе мы показали, что экзогенный
каскад NF-κB в фибробластах 3Т3 и стаби-
PRDX6 препятствует гиперэкспрессии гена Nos2
лизирует до контрольного уровня активность
и повышению концентрации АФК в макрофа-
данного сигнального каскада при его гипер-
гах, что, в конечном счёте, оказывает влияние
активации в условиях воздействия ионизирую-
на апоптоз. Подавление экспрессии гена Trp53
щего излучения [29]. В настоящей работе мы
экзогенным PRDX6 подтверждает его анти-
подтвердили способность PRDX6 выступать в
апоптозный эффект на клетки RAW 264.7, акти-
роли регуляторной молекулы, показав его спо-
вированные эндотоксином.
собность снижать гиперактивацию транскрип-
Таким образом, настоящая работа допол-
ционного фактора NF-κB при развитии про-
няет наши более ранние исследования и вы-
воспалительного ответа, вызванного ЛПС.
двинутое предположение о том, что экзо-
Zhang et al. [30] показали, что нокдаун
генный PRDX6 выступает в роли регулятор-
TLR4 приводит к блокировке выработки АФК,
ной молекулы. В частности, PRDX6 является
ингибированию экспрессии p53 и каспазы-3.
антивоспалительным агентом, способным
В своей статье авторы предполагают, что TLR4
защитить активированные ЛПС макрофаги
является перспективной мишенью для профи-
RAW 264.7 от чрезмерного провоспалительно-
лактики и лечения заболеваний, связанных с
го ответа и не допустить развития патологиче-
апоптозом, в частности атеросклероза. Наши
ских состояний, вызванных хроническим вос-
результаты, полученные при исследовании
палением лёгких.
экспрессии и продукции TLR4, показали, что
экзогенное влияние PRDX6 на активирован-
Вклад авторов. С.Б. Парфенюк, Е.Г. Ново-
ные ЛПС макрофаги приводит к блокировке
селова - концепция и руководство работой;
данного рецептора и всех нижележащих сиг-
С.Б. Парфенюк, О.В. Глушкова, М.Г. Шарапов,
нальных путей, что в конечном итоге приводит
М.О. Хренов, Э.К. Мубаракшина, Т.В. Ново-
к ингибированию апоптоза, вероятно, через
селова, А.А. Кузекова
- проведение экспе-
ингибирование экспрессии р53.
риментов; С.Б. Парфенюк, Е.Г. Новоселова,
Оксид азота, синтезируемый из L-арги-
С.М. Лунин, О.В. Глушкова
- обсуждение
нина с помощью NO-синтаз, представляет
результатов исследования; С.Б. Парфенюк,
собой небольшую липофильную диффунди-
Е.Г. Новоселова - написание текста; С.М. Лу-
рующую высокореактивную молекулу с дихо-
нин, Д.А. Черенков, О.В. Глушкова, М.Г. Ша-
томической регуляторной ролью во многих
рапов - редактирование текста статьи.
биологических событиях в физиологических и
Благодарности. В работе использовали обо-
патологических условиях [31]. NO может спо-
рудование (планшет-ридер Infinite 200 («Tecan»))
собствовать апоптозу в одних клетках и инги-
центра коллективного пользования Пущин-
бировать апоптоз в других.
ского научного центра.
БИОХИМИЯ том 88 вып. 8 2023
ЗАЩИТНЫЕ ЭФФЕКТЫ ПЕРОКСИРЕДОКСИНА 6
1421
Финансирование. Работа выполнена при
Соблюдение этических норм. Настоящая
поддержке Российского научного фонда, грант
статья не содержит описания выполненных
№ 23-24-00041.
авторами исследований с участием людей или
Конфликт интересов. Авторы заявляют об
использованием животных в качестве объек-
отсутствии конфликта интересов.
тов.
СПИСОК ЛИТЕРАТУРЫ
1.
Barnes, P. J.
(2017) Cellular and molecular
regulates acute and chronic cigarette smoke-mediated
mechanisms of asthma and COPD, Clin. Sci. (Lond),
lung inflammatory response and injury, Exp. Lung
13, 1541-1558, doi: 10.1042/CS20160487.
Res., 36, 451-462, doi: 10.3109/01902141003754128.
2.
Joshi, N., Walter, J. M., and Misharin, A. V. (2018)
13.
Fisher, A. B. (2011) Peroxiredoxin 6: a bifunctional
Alveolar macrophages, Cell Immunol., 330, 86-90,
enzyme with glutathione peroxidase and phospholipase
doi: 10.1016/j.cellimm.2018.01.005.
A2 activities, Antioxid. Redox Signal., 15, 831-844,
3.
Lu, X. J., Ning, Y. J., Liu, H., Nie, L., and
doi: 10.1089/ars.2010.3412.
Chen, J. (2018) A novel lipopolysaccharide recognition
14.
Power, J. H., Asad, S., Chataway, T. K., Chegini, F.,
mechanism mediated by internalization in teleost
Manavis, J., et al. (2008) Peroxiredoxin 6 in human
macrophages, Front. Immunol., 9, 2758, doi: 10.3389/
brain: molecular forms, cellular distribution and
fimmu.2018.02758.
association with Alzheimer’s disease pathology, Acta
4.
Челомбитько М. А. (2018) Роль активных форм
Neuropathol.,
115,
611-622, doi:
10.1007/s00401-
кислорода в воспалении, Вестн. Моск. Унив., 73,
008-0373-3.
242-246.
15.
Novoselova, E. G., Glushkova, O. V., Parfenuyk, S. B.,
5.
Sharapov, M. G., Ravin, V. K., and Novoselov, V. I.
Khrenov, M. O., Lunin, S. M., et al. (2019) Protective
(2014) Peroxyredoxins as multifunctional enzymes,
effect of peroxiredoxin 6 against toxic effects of
Mol. Biol. (Mosk),
48,
600-628, doi:
10.1134/
glucose and cytokines in pancreatic RIN-m5F β-cells,
S0026893314040128.
Biochemistry (Moscow), 84, 637-643, doi: 10.1134/
6.
Hanschmann, E. M., Godoy, J. R., Berndt, C.,
S0006297919060063.
Hudemann, C., and Lillig, C. H. (2013) Thioredox-
16.
Chen, S., Hu, Y., Zhang, J., and Zhang, P. (2021)
ins, glutaredoxins, and peroxiredoxins - molecular
Anti-inflammatory effect of salusinβ knockdown
mechanisms and health significance: from cofactors
on LPS-activated alveolar macrophages via NF-κB
to antioxidants to redox signaling, Antioxid. Redox Sig-
inhibition and HO-1 activation, Mol. Med. Rep., 23,
nal., 19, 1539-1605, doi: 10.1089/ars.2012.4599.
127, doi: 10.3892/mmr.2020.11766.
7.
Arevalo, J. A., and Vázquez-Medina, J. P. (2018) The
17.
Sharapov, M. G., Novoselov, V. I., and Ravin, V. K.
role of peroxiredoxin 6 in cell signaling, Antioxidants
(2009) Cloning, expression and comparative analysis
(Basel), 7, 172, doi: 10.3390/antiox7120172.
of peroxiredoxine 6 from different species, Mol. Biol.
8.
Sharapov, M. G., Goncharov, R. G., Parfenyuk,
(Mosk), 43, 505-511, doi: 10.1134/s0026893309030194.
S. B., Glushkova, O. V., and Novoselov, V. I. (2022)
18.
Wu, D., and Yotnda, P.
(2011) Production and
The role of phospholipase activity of peroxiredoxin
detection of reactive oxygen species (ROS) in cancers,
6 in its transmembrane transport and protective
J. Vis. Exp., 57, 3357, doi: 10.3791/3357.
properties, Int. J. Mol. Sci., 23, 152-165, doi: 10.3390/
19.
Glushkova, O. V., Khrenov, M. O., Novoselova, T. V.,
ijms232315265.
Lunin, S. M., Parfenyuk, S. B., et al. (2015) The role
9.
Chuchalin, A. G., Novoselov, V. I., Shifrina,
of the NF-κB, SAPK/JNK, and TLR4 signalling
O. N., Soodaeva, S. K., Yanin, V. A., et al. (2003)
pathways in the responses of RAW 264.7 cells to
Peroxiredoxin VI in human respiratory system, Respir.
extremely low intensity microwaves, Int. J. Radiat. Biol.,
Med., 97, 147-151, doi: 10.1053/rmed.2003.1429.
91, 321-328, doi: 10.3109/09553002.2014.996261.
10.
Kümin, A., Huber, C., Rülicke, T., Wolf, E., and
20.
Sharapov, M. G., Novoselov, V. I., and Gudkov,
Werner, S. (2006) Peroxiredoxin 6 is a potent cytopro-
S. V. (2019) Radioprotective role of peroxiredoxin 6,
tective enzyme in the epidermis, Am. J. Pathol., 169,
Antioxidants (Basel), 8, 15, doi: 10.3390/antiox8010015.
1194-1205, doi: 10.2353/ajpath.2006.060119.
21.
Новоселов В. И. (2012) Роль пероксиредоксинов
11.
Kümin, A., Schäfer, M., Epp, N., Bugnon, P., Born-
при окислительном стрессе в органах дыхания,
Berclaz, C., et al. (2007) Peroxiredoxin 6 is required
Пульмонология, 83-87.
for blood vessel integrity in wounded skin, J. Cell.
22.
Ross, E. A., Devitt, A., and Johnson, J. R. (2021)
Biol., 179, 747-760, doi: 10.1083/jcb.200706090.
Macrophages: the good, the bad, and the gluttony,
12.
Sundar, I. K., Chung, S., Hwang, J. W., Arunachalam,
Front. Immunol., 12, 708186, doi: 10.3389/fimmu.
G., Cook, S., et al. (2010) Peroxiredoxin 6 differentially
2021.708186.
БИОХИМИЯ том 88 вып. 8 2023
1422
ПАРФЕНЮК и др.
23. Fukai, T., and Ushio-Fukai, M. (2011) Superoxide
MAPK/NF-κB inflammatory signaling, Phytomedi-
dismutases: role in redox signaling, vascular function,
cine, 57, 305-314, doi: 10.1016/j.phymed.2018.12.043.
and diseases, Antioxid. Redox. Signal., 15, 1583-1606,
29. Sharapov, M. G., Glushkova, O. V., Parfenyuk, S. B.,
doi: 10.1089/ars.2011.3999.
Gudkov, S. V., Lunin, S. M., et al. (2021) The role of
24. Zhang, X., Yu, Y., Lei, H., Cai, Y., Shen, J., et al.
TLR4/NF-κB signaling in the radioprotective effects
(2020) The Nrf-2/HO-1 signaling axis: a ray of hope
of exogenous PRDX6, Arch. Biochem. Biophys., 15,
in cardiovascular diseases, Cardiol. Res. Pract., 30,
108830, doi: 10.1016/j.abb.2021.108830.
5695-5723, doi: 10.1155/2020/5695723.
30. Zhang, Y., Xia, G., Zhang, Y., Liu, J., Liu, X., et al.
25. Park, M. H., Jo, M., Kim, Y. R., Lee, C. K., and
(2017) Palmitate induces VSMC apoptosis via toll like
Hong, J. T. (2016) Roles of peroxiredoxins in can-
receptor (TLR)4/ROS/p53 pathway, Atherosclerosis,
cer, neurodegenerative diseases and inflammatory
263, 74-81, doi: 10.1016/j.atherosclerosis.2017.06.002.
diseases, Pharmacol. Ther., 163, 1-23, doi: 10.1016/
31. Duran, X., Vilahur, G., and Badimon, L.
(2009)
j.pharmthera.2016.03.018.
Exogenous in vivo NO-donor treatment preserves
26. Webster, J. M., Kempen, L. J. A. P., Hardy, R. S., and
p53 levels and protects vascular cells from apop-
Langen, R. C. J. (2020) Inflammation and skeletal
tosis, Atherosclerosis,
205,
101-106, doi:
10.1016/
muscle wasting during cachexia, Front. Physiol., 11,
j.atherosclerosis.2008.11.016.
597-675, doi: 10.3389/fphys.2020.597675.
32. Lee, M., Rey, K., Besler, K., Wang, C., and Choy, J.
27. Zhao, L. X., Du, J. R., Zhou, H. J., Liu, D. L., Gu,
(2017) Immunobiology of nitric oxide and regulation
M. X., et al. (2016) Differences in proinflammatory
of inducible nitric oxide synthase, Results. Probl.
property of six subtypes of peroxiredoxins and anti-
Cell Differ.,
62,
181-207, doi:
10.1007/978-3-319-
inflammatory effect of ligustilide in macrophages,
54090-0_8.
PLoS One,
11, e0164586, doi:
10.1371/journal.
33. Dubey, M., Nagarkoti, S., Awasthi, D., Singh, A. K.,
pone.0164586.
Chandra, T., et al. (2016) Nitric oxide-mediated
28. Yuan, F., Liu, R., Hu, M., Rong, X., Bai, L., et al.
apoptosis of neutrophils through caspase-8 and
(2019) JAX2, an ethanol extract of Hyssopus cuspida-
caspase-3-dependent mechanism, Cell Death Dis., 7,
tus Boriss, can prevent bronchial asthma by inhibiting
e2348, doi: 10.1038/cddis.2016.248.
PROTECTIVE EFFECTS OF PEROXIREDOXIN 6 IN MODELING
PROINFLAMMATORY RESPONSE USING RAW 264.7 MACROPHAGES
S. B. Parfenyuk1*, O. V. Glushkova1, M. G. Sharapov1, M. O. Khrenov1,
S. M. Lunin1, A. A. Kuzekova1, E. K. Mubarakshina1, T. V. Novoselova1,
D. A. Cherenkov2, and E. G. Novoselova1
1 Institute of Cell Biophysics of the Russian Academy of Sciences,
Pushchino Scientific Center for Biological Research of the Russian Academy of Sciences,
142290 Pushchino, Moscow Region, Russia; e-mail: lana_kras2@rambler.ru
2 Voronezh State University of Engineering Technologies, 394036 Voronezh, Russia
The aim of the work was to study the effects of peroxiredoxin 6 (PRDX6), a recombinant antioxidant pro-
tein, on the level of proinflammatory responses caused by endotoxin exposure to RAW 264.7 macrophages.
The addition of LPS to the RAW 264.7 cell culture medium expectedly increased the production of TNF-α,
and the addition of PRDX6 led to a significant decrease in its production by 15-20%. The level of produc-
tion of another proinflammatory cytokine, IL-1β, significantly activated by endotoxin, was completely nor-
malized under the PRDX6 action. In addition, the addition of PRDX6 reduced the production of reactive
oxygen species (ROS) induced by endotoxin and also prevented overexpression of the iNos gene in RAW
264.7 cells. The results showed that PRDX6 has a suppressive effect on the expression of the Nfe2l2 gene
and the production of the transcription factor NRF-2 during the first 6 h of cell culture. The addition of
endotoxin causes the activation of the NF-κB and SAPK/JNK signaling cascades, while in the presence
of PRDX6, the activity of these signaling cascades decreases. It is known that the proinflammatory re-
sponse of cells caused by bacterial LPS leads to the activation of apoptosis and the elimination of damaged
cells. Our studies confirm this, since LPS leads to the activation of the Trp53 gene, a marker of apoptosis.
The addition of peroxiredoxin 6 in the first hours of the development of an acute proinflammatory response
leads to the suppression of Trp53 gene expression, which indicates a protective effect of PRDX6 that reduces
apoptosis in RAW 264.7 macrophages.
Keywords: peroxiredoxin 6, inflammation, cytokines, gene expression, signaling cascades
БИОХИМИЯ том 88 вып. 8 2023