БИОХИМИЯ, 2023, том 88, вып. 8, с. 1423 - 1440
УДК 577.112.7;577.113.7;577.22
КОМПЛЕКСЫ И ВЫСОКОМОЛЕКУЛЯРНЫЕ АССОЦИАТЫ
ДОДЕЦИЛ-СОДЕРЖАЩИХ ОЛИГОНУКЛЕОТИДОВ
С СЫВОРОТОЧНЫМ АЛЬБУМИНОМ
© 2023 А.С. Павлова*, В.В. Илющенко, М.С. Купрюшкин, Т.Д. Жарков, Е.С. Дюдеева,
И.А. Бауэр, А.С. Чубаров, Д.В. Пышный, И.А. Пышная*
Институт химической биологии и фундаментальной медицины СО РАН,
630090 Новосибирск, Россия; электронная почта: pyshnaya@niboch.nsc.ru, pavlova@niboch.nsc.ru
Поступила в редакцию 26.04.2023
После доработки 26.06.2023
Принята к публикации 04.07.2023
Сывороточный альбумин, имеющий в своей молекуле несколько сайтов связывания различных со-
единений, в том числе липофильных олигонуклеотидных производных, является объектом биоме-
дицинских исследований как перспективная платформа для создания многокомпонентных само-
организующихся систем. В данной работе мы исследовали методом задержки в геле стехиометрию
связывания бычьего сывороточного альбумина и сывороточного альбумина человека с додецил-
содержащими олигонуклеотидами. Результаты исследования свидетельствуют об образовании
в условиях, близких к физиологическим, комплексов со стехиометрией альбумин : олигонуклео-
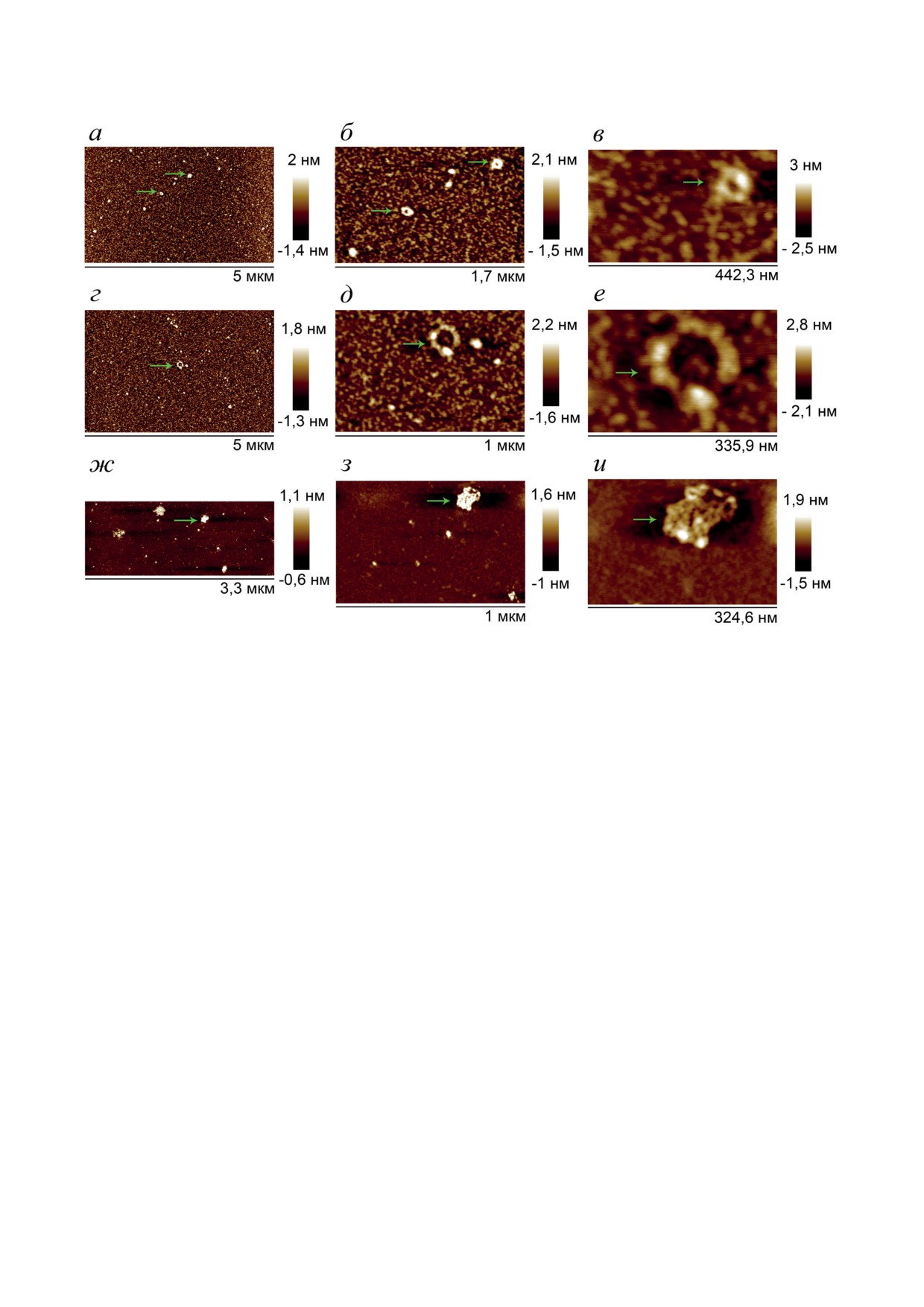
тидное производное, равной 1 : (1,25 ± 0,25). Методом атомно-силовой микроскопии показано,
что результатом взаимодействия сывороточного альбумина человека с дуплексом комплементар-
ных додецил-содержащих олигонуклеотидов являются высокомолекулярные ассоциаты округ-
лой формы диаметром 165,5 ± 94,3 нм и высотой 28,9 ± 16,9 нм, а с комплексами полиадениловой
кислоты и додецил-содержащего олиготимидилата - супрамолекулярные ассоциаты размером
315,4 ± 70,9 и 188,3 ± 43,7 нм. Полученные данные позволяют рассматривать додецил-содержащие
олигонуклеотиды и альбумин как перспективные составляющие для разработки самоорганизую-
щихся систем для решения задач молекулярной биологии и биомедицины, в частности, создания
уникальных тераностиков направленного действия.
КЛЮЧЕВЫЕ СЛОВА: сывороточный альбумин, додецил-содержащие олигонуклеотиды, белково-нуклеи-
новые комплексы, самоорганизация биополимеров, супрамолекулярные ассоциаты, доставка нуклеино-
вых кислот.
DOI: 10.31857/S0320972523080109, EDN: IKOTTQ
ВВЕДЕНИЕ
факторов, сдерживающих потенциал примене-
ния тНК in vivo, является проблема их направ-
Терапевтические нуклеиновые кислоты
ленной доставки к внутриклеточным мише-
(тНК), их синтетические аналоги и нанокон-
ням [1-2]. Помимо этого, существуют другие
струкции на их основе являются в настоящее
сложности в применении тНК, связанные с та-
время объектами исследований для решения
кими аспектами, как быстрая деградация тНК
различных задач в областях персонализиро-
под действием внеклеточных и внутриклеточ-
ванной медицины и тераностики [1-4]. Восем-
ных ферментов, выведение тНК из организ-
надцать лекарственных препаратов на осно-
ма, плохая способность тНК к преодолению
ве тНК получили разрешение на применение
барьеров ретикулоэндотелиальной системы,
в терапии различных заболеваний человека и
низкая эффективность проникновения тНК
более 100 находятся на разных этапах доклини-
внутрь клеток и выхода из эндосом [3, 8-10].
ческих испытаний [3, 5-7]. Одним из важных Имеющиеся достижения в области разработки
Принятые сокращения: АСМ - атомно-силовая микроскопия; БСА - бычий сывороточный альбумин; ДСО - доде-
цил-содержащий олигонуклеотид; ЖК - жирные кислоты; НК - нуклеиновые кислоты; тНК - терапевтические НК;
поли(dA) - полиадениловая кислота; СА - сывороточный альбумин; ЧСА - сывороточный альбумин человека;
ЧСАм - очищенная мономерная фракция ЧСА; экв. - эквивалент; FAM - остаток флуоресцентного красителя
6-карбоксифлуоресцеина.
* Адресат для корреспонденции.
1423
1424
ПАВЛОВА и др.
синтетических (модифицированных) аналогов
использованием элементов НК-архитектони-
тНК позволили существенно нивелировать их
ки [41-45]. Несомненным преимуществом та-
деградацию [2, 8, 11-17]. Поэтому в настоящее
ких сложноорганизованных систем доставки
время особое внимание уделяют изучению
является возможность включения в их состав
влияния модификаций тНК-конструкций на
нескольких, в том числе направляющих, тера-
их взаимодействие с белками и другими ком-
певтических либо диагностических, а также
понентами как крови, так и внутриклеточны-
стимул-чувствительных компонентов [45-49].
ми, поскольку это может прямым или косвен-
Ранее было показано, что производные
ным образом отразиться как на доставке тНК,
олигонуклеотидов, содержащие 3 додецильных
так и на эффективности оказываемого тНК
остатка в составе ненуклеотидного блока в
биологического эффекта в организме
[1, 8,
своей структуре, способны эффективно про-
18-23]. Присоединение гидрофобных остат-
никать in vitro в клетки аденокарциномы лёгко-
ков к тНК, в частности, холестерина, жирных
го A549 и эпидермоидной карциномы KB-8-5
кислот (ЖК) и липидов, а также других алкил-
человека в отсутствие трансфецирующих аген-
содержащих группировок, способствует повы-
тов [50]. Инкубирование додецил-содержащих
шению эффективности проникновения таких
олигонуклеотидов (ДСО) в присутствии почти
производных тНК через мембрану внутрь
2-кратного избытка бычьего сывороточного
клеток, а также улучшению их фармакокине-
альбумина (БСА) приводит к формированию
тических характеристик in vivo [8, 10, 24-29].
белково-нуклеиновых комплексов [51]. Однако
Среди данных остатков особый интерес пред-
известно, что молекула СА содержит как ми-
ставляют группировки, имеющие повышенное
нимум 7 сайтов связывания остатков жирных
сродство к связыванию с сывороточным аль-
кислот, транспорт которых белок осуществля-
бумином (СА) [20, 25, 30-32]. СА, основной
ет в организме [52-54].
белковый компонент сыворотки крови, за счёт
В данной работе методом задержки в геле
низкой иммуногенности, продолжительного
мы исследовали связывание БСА и сыворо-
времени полужизни, способности накапли-
точного альбумина человека (ЧСА) с олиго-
ваться в тканях, подверженных онкотранс-
нуклеотидными производными, содержащими
формации и воспалению, а также наличию в
1, 2 или 3 додецильных остатка. Отдельное
своей структуре сайтов связывания различ-
внимание было уделено оценке возможности
ных лигандов, является объектом многочис-
комплементарного взаимодействия олигонук-
ленных исследований в мире, направленных
леотидов в составе комплексов НК-белок.
на разработку систем доставки широкого
Получено, что ДСО с тремя додецильными
спектра терапевтических и диагностических
остатками формируют с альбумином ста-
средств [33-38]. В свою очередь, связыва-
бильные в условиях электрофореза ком-
ние СА с олигонуклеотидами и их производ-
плексы, стехиометрия образования которых
ными рассматривается как один из способов
в эквивалентах (экв.) белок : ДСО составила
для создания тНК-содержащих конструкций
1 : (1,25 ± 0,25). Методом атомно-силовой мик-
с целью увеличения их времени циркуляции,
роскопии (АСМ) была изучена морфология ас-
повышения биодоступности и стабильности,
социатов в образцах ЧСА с комплементарными
снижения иммуногенности и, как следствие,
ДСО и ЧСА с полиадениловой кислотой
уменьшения вероятности неспецифических по-
(поли(dA)) и олиготимидилатным ДСО (далее:
бочных эффектов и улучшения эффективно-
высокомолекулярных или супрамолекулярных
сти действия [20, 30-32]. Вследствие эффекта
ассоциатов соответственно). Зафиксировано
повышенной проницаемости и удерживания
наличие высокомолекулярных ассоциатов ЧСА
(enhanced permeability and retention, EPR-эф-
с дуплексом комплементарных ДСО округ-
фект), характерного для опухоли и её окру-
лой формы диаметром 165,5 ± 94,3 нм и вы-
жения, а также наличия специфических к СА
сотой 28,9 ± 16,9 нм, а также двух популяций
рецепторов, количество которых повышено в
супрамолекулярных ассоциатов ЧСА с поли(dA)
некоторых видах рака [37-40], ожидается, что
и олиготимидилатным ДСО размером 315,4 ±
создание альбумин-связанных конструкций бу-
± 70,9 и 188,3 ± 43,7 нм соответственно.
дет способствовать улучшению накопления те-
рапевтических средств в тканях опухоли in vivo.
Помимо относительно простых конструк-
МАТЕРИАЛЫ И МЕТОДЫ
ций с присоединением липофильных остатков
к тНК, актуально направление по созданию
Материалы. Акриламид («AppliChem GmbH»,
самоорганизующихся (и, как правило, опре-
Германия); бис-акриламид, Кумасси брилли-
делённым образом упорядоченных) систем с
антовый голубой G-250 («Amresco», США);
БИОХИМИЯ том 88 вып. 8 2023
СВОЙСТВА ДОДЕЦИЛ-СОДЕРЖАЩИХ ОЛИГОНУКЛЕОТИДОВ
1425
ксиленцианоловый голубой, бычий сывороточ-
Стоковые растворы СА, ДСО и поли(dA)
ный альбумин (А7030), натриевая соль поли(2′-
разбавляли до указанных концентраций, со-
дезокси)адениловой кислоты (81342), 1 М MgCl2
ответствующих соотношению смешивания
(«Sigma», США); сывороточный альбумин
СА : ДСО в экв. За 1 экв. принимали количе-
человека (фракция V;
«Reanal», Венгрия);
ство моль СА при указанной концентрации и
для выделения мономера использовали ЧСА
конечном объёме проб 20 мкл. При смешива-
(P06-26050, «PAN Biotech UK Ltd», Велико-
нии комплементарных нуклеотидных после-
британия); ацетон (о.с.ч.) («Реахим», Россия);
довательностей и СА для анализа методом
LiClO4, NaCl, (NH4)2S2O8 (98-99,85%, «Acros
задержки в ПААГ пробы готовили двумя спо-
Organics», США); N,N,N′,N′-тетраметилэтилен-
собами. В первом случае сперва добавляли
диамин («Bio-Rad Laboratories», США); Tris
НК-компоненты, инкубировали (3 мин 95 °С),
(«Fisher Scientific», США); AcOH (ледяная);
охлаждали до 37 °С, затем добавляли альбумин.
деионизованная H2O 18 MΩ, полученная в си-
Во втором случае сперва готовили комплексы
стеме очистки Simplicity 185 («Millipore», США).
альбумина с одним из ДСО, смешивая в одной
Состав буферных растворов. В работе ис-
пробирке белок с ДСО 3R, а в другой - белок
пользовали следующие буферные растворы:
с 3L, отдельно инкубировали их 30 мин при
ТАN - 100 мМ NaCl, 50 мМ Tris-Ac (рН 7,5);
37 °С, а затем объединяли в одну пробирку.
TAM - 15 мМ MgCl2, 50 мМ Tris-Ac (рН 7,5).
При приготовлении образцов для исследова-
Все растворы фильтровали через
0,22 мкм
ния методом АСМ сперва смешивали НК-ком-
шприцевые фильтры Millipore Syringe Filter
поненты, инкубировали (3 мин 95 °С), охлаж-
units («Merck», Германия).
дали до 37 °С, затем добавляли альбумин.
Синтез олигонуклеотидных производных с
Исследование методом задержки в 8%-ном
остатками додецил-содержащего ненуклеотид-
нативном ПААГ (акриламид : бисакриламид =
ного мономерного звена D и флуоресцентно-
= 29 : 1) проводили с использованием камеры
го красителя 6-карбоксифлуоресцеина (FAM)
Owl™ Dual-Gel Vertical Electrophoresis System
проводили твердофазным амидофосфитным
P10DS («Owl separation Systems Inc.», США)
методом на автоматическом НК-синтезаторе
при термостатировании системы с помощью
АСМ-800 («Биоссет», Россия) с использова-
циркуляционного термостата F12 («Julabo Inc.»,
нием реагентов и протоколов, в том числе
США). Приготовленные, как описано выше,
постсинтетической очистки, описанных ранее
пробы инкубировали 30 мин при 37 °С без пе-
[50-51, 55].
ремешивания, затем добавляли 5 мкл водно-
Концентрации растворов ДСО, СА и поли(dA)
го раствора 30% (v/v) глицерина с 0,05% (w/v)
определяли по измерению поглощения (А) при
ксиленцианоловым голубым, и
5 мкл из
комнатной температуре с использованием
каждой пробы наносили на ПААГ. Отно-
спектрофотометра NanoVue™
(«GE Health-
сительную подвижность проб анализиро-
care», США). Молярные коэффициенты по-
вали при постоянной мощности 6 Вт и тер-
глощения ε260 ДСО рассчитывали, используя
мостатировании камеры для электрофоре-
значения ε260 для моно- и динуклеотидов, при-
за при 37 °С. Документирование результатов
ведённые в работе Dunn и Hall [56], ε260 остат-
анализа проводили с помощью сканирования
ка FAM считали равным 20 900 М-1·см-1 (Glen
гелей (Ex/Em = 488/530 нм) с использова-
Research Technical Bulletin; «Glen Research»,
нием системы VersaDoc™ MP 4000 Molecular
США). Молярные коэффициенты поглоще-
Imager® System («Bio-Rad Laboratories», США;
ния ДСО полагали равными сумме ε260 со-
(ОЦГПМИ) ИХБФМ СО РАН). Интенсив-
ответствующего олигонуклеотида и остат-
ность флуоресценции от полос образцов на
ка FAM, вклад ненуклеотидного додецил-со-
электрофореграммах оценивали с помощью
держащего звена в суммарную величину ε260
программного пакета GelPro
4.0
(«Media
ДСО не учитывали. Значения ε280 альбуминов
Cybernetics L.P.», США). Выход кривых на
использовали: 43 824 М-1·см-1 - для БСА и
плато определяли с помощью аппроксима-
37 000 М-1·см-1 - для ЧСА [57]. Значение кон-
ции данных методом нелинейной фильтра-
центрации поли(dA) рассчитывали, полагая
ции в программном обеспечении Origin®
ε260 аденина равным 12 300 М-1·см-1.
(«OriginLab», США).
Мономерная фракция сывороточного альбу-
Исследование методом АСМ проводили
мина человека была выделена, как описано в
с использованием сканирующего зондового
работе Chubarov et al. [58].
микроскопа MultiMode 8™ («Bruker», США),
Формирование комплексов альбумин-ДСО
подключённого к контроллеру NanoScope® V
проводили в растворе TAN (для анализа мето-
(«Veeco», США). Приготовленные, как описа-
дом задержки в геле) либо ТАМ (для АСМ).
но выше, пробы инкубировали 15 ч при 15 °С
10
БИОХИМИЯ том 88 вып. 8 2023
1426
ПАВЛОВА и др.
без перемешивания, затем 7 мкл каждой про-
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
бы наносили на свежесколотую поверхность
слюды площадью 1 × 1 см2, спустя 1 мин про-
Исследование проводили на модельной си-
мывали 3 раза по 1 мл H2O 18 MΩ, сушили
стеме, состоящей из серий олигонуклеотидных
в токе аргона и исследовали методом АСМ.
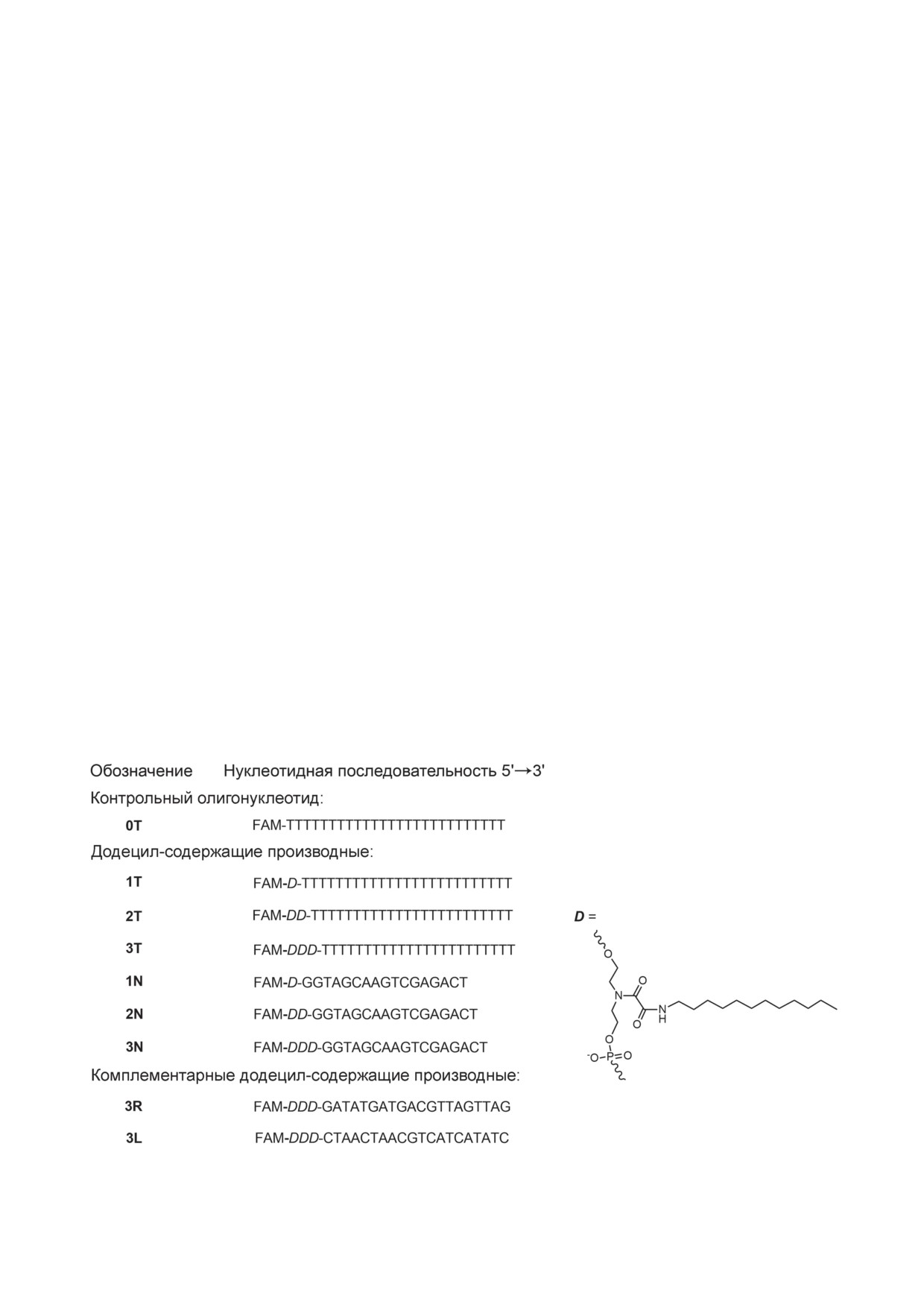
производных гомотимидилатного (1Т, 2Т, 3Т)
Остатки проб хранили при 4 °С, и дальнейшее
и гетеронуклеотидного состава (1N, 2N, 3N),
разбавление проводили буферным раство-
имеющих на
5′-конце остаток
6-карбокси-
ром ТАМ такой же температуры. Изображения
флуоресцеина и разное число (1-3) последова-
поверхности получали в режиме полуконтакт-
тельно расположенных остатков додецил-со-
ной моды Soft tapping микроскопа в атмосфер-
держащего ненуклеотидного звена D (рис. 1).
ных условиях с помощью алмазоподобного
Исследование связывания додецил-содер-
кантилевера NSG10_DLC («NT-MDT Spectrum
жащих олигонуклеотидов с сывороточными аль-
Instruments», Россия), имеющего параметры
буминами методом задержки в геле. Для оценки
радиуса кривизны
- 1-3 нм, резонансной
стехиометрии образования комплексов альбу-
частоты - 190-325 кГц и силовой постоян-
мин-ДСО анализировали степень ассоциации
ной - 5,5-22,5 Н/м. Полученные изображе-
олигонуклеотидных производных с белком в
ния обрабатывали и анализировали с помо-
зависимости от соотношения СА : ДСО. Для
щью программного обеспечения Nanoscope
этого провели серию экспериментов, добав-
Analysis 1.40 («Bruker»).
ляя к 0,1 нмоль альбумина (1 экв.) ДСО, коли-
Статистическая значимость полученных
чество которого варьировали от 0,5 до 3 экв.
данных. Приведённые в работе электрофоре-
по отношению к белку. Электрофоретическая
граммы и изображения АСМ исследованных
подвижность не связанного с белком ДСО
образцов являются результатами одного ти-
значительно выше, чем подвижность СА. По-
пичного эксперимента из серии не менее трёх
этому при добавлении альбумина к ДСО на
аналогичных. Размерные характеристики ком-
электрофореграмме можно наблюдать появ-
плексов ЧСА с ДСО и с поли(dA) на изобра-
ление ДСО-содержащих полос с более низ-
жениях АСМ определяли с помощью функций
кой подвижностью в ПААГ, соответствующих
Section и Particle analysis, производя анализ
образованию альбумин-олигонуклеотидных
не менее трёх типичных изображений одина-
комплексов СА••ДСО (здесь и далее знак ••
кового масштаба из разных мест поверхности
будет обозначать связывание СА с ДСО) [51].
слюды с нанесённым образцом. Результат при-
Область подвижности данных комплексов
ведён в виде среднего значения и стандартного
далее на представленных электрофореграм-
отклонения (М ± SD).
мах обозначена красной квадратной скобкой,
Рис.
1. Структуры олигодезоксирибонуклеотидных производных. FAM
- остаток
6-карбоксифлуоресцеина;
D - структура ненуклеотидного додецил-содержащего звена в составе олигонуклеотида
БИОХИМИЯ том 88 вып. 8 2023
СВОЙСТВА ДОДЕЦИЛ-СОДЕРЖАЩИХ ОЛИГОНУКЛЕОТИДОВ
1427
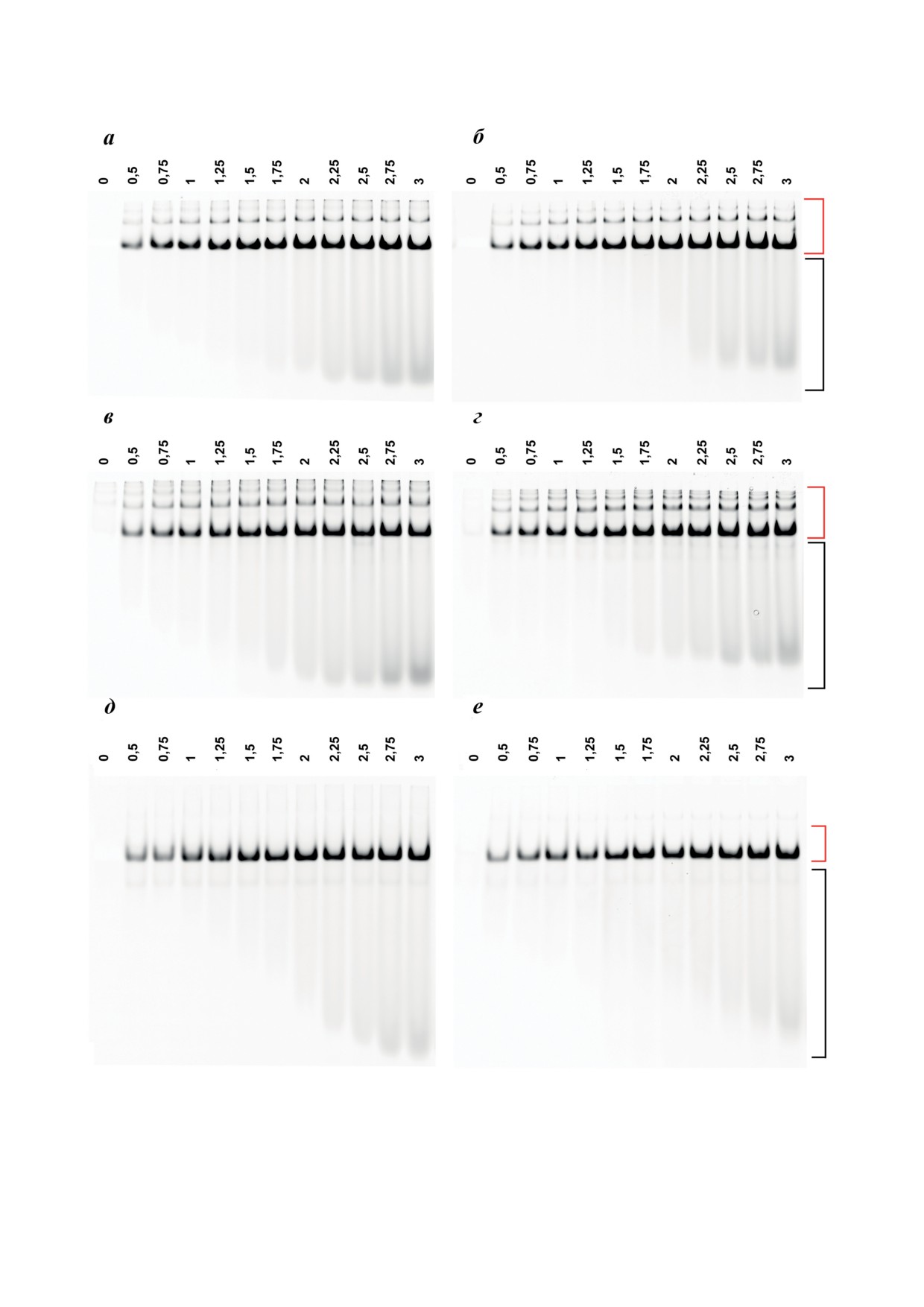
Рис. 2. Электрофореграмма относительной подвижности комплексов БСА (а и б), ЧСА (в и г) и мономерной фрак-
ции ЧСАм (д и е) с ДСО 3R (а, в, и д) и 3L (б, г, и е) в 8%-ном ПААГ в неденатурирующих условиях. Каждая проба
содержала 5 мкМ белка в растворе TAN, объём пробы - 20 мкл. Цифры над дорожками соответствуют эквивален-
там ДСО в составе пробы. За 1 экв. было принято количество вещества альбумина, равное 0,1 нмоль. Пробы гото-
вили и анализировали, как описано в разделе «Материалы и методы». Красной квадратной скобкой обозначена об-
ласть, соответствующая подвижности комплексов белка с ДСО в данных условиях, чёрной - область подвижности
свободного ДСО
БИОХИМИЯ том 88 вып. 8 2023
10*
1428
ПАВЛОВА и др.
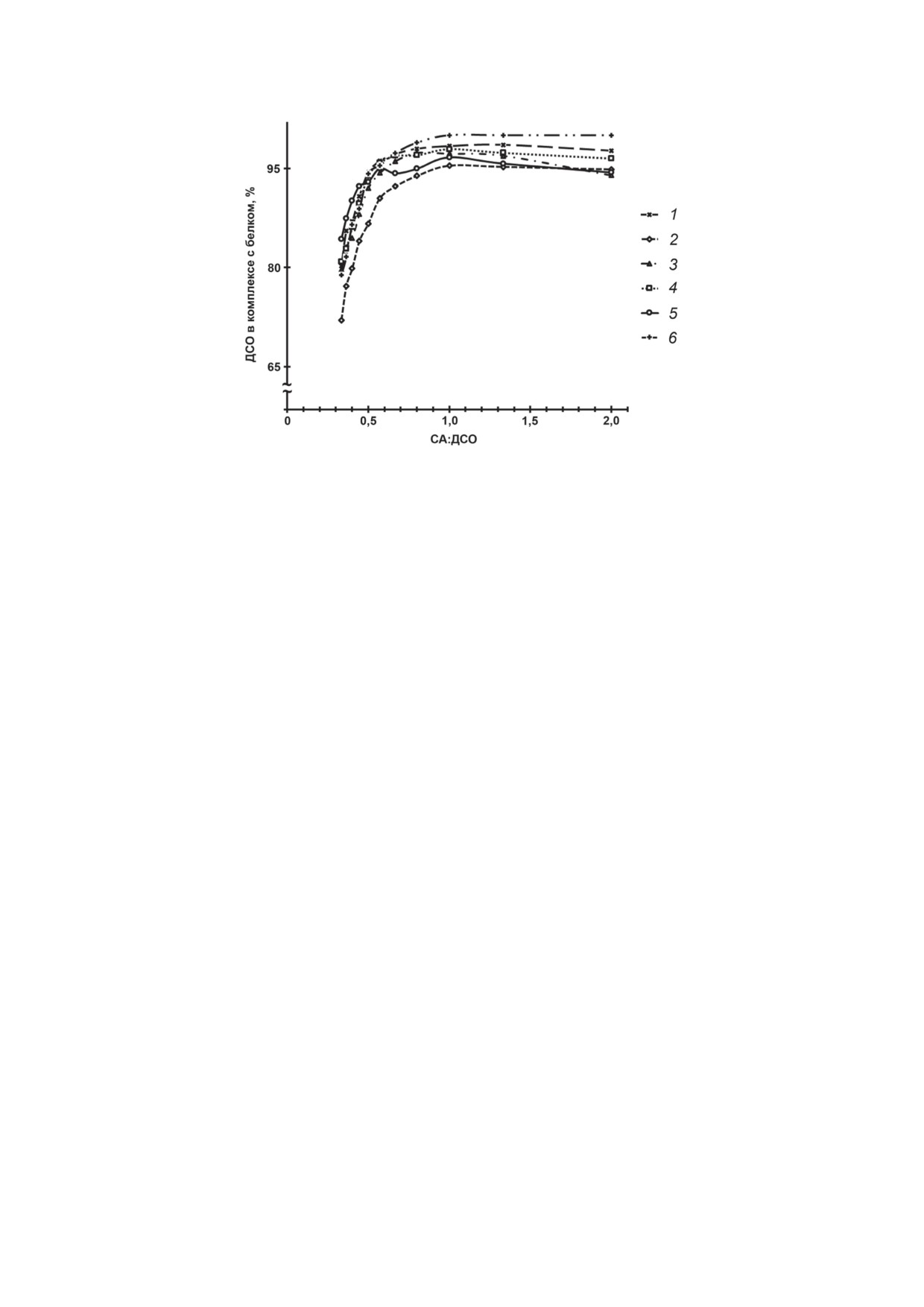
Рис. 3. Зависимость количества ДСО в комплексе с белком (%) от значения соотношения СА к ДСО (в экв.). За 1 экв.
было принято количество вещества n(СА), равное 0,1 нмоль. 1 - БСА••3L; 2 - ЧСА••3R; 3 - ЧСА••3L; 4 - ЧСАм••3R;
5 - ЧСАм••3L; 6 - БСА••3R
а область подвижности не связанного с бел-
Сравнительное исследование связывания
ком ДСО - чёрной (рис. 1П в Приложении 1;
ДСО с гомологичными альбуминами БСА
рис. 2). Полос, соответствующих комплексам
и ЧСА, а также очищенной мономерной
СА••ДСО, несколько, так как сывороточный
фракцией ЧСА (ЧСАм) показало, что ДСО 3R
альбумин, помимо мономера, также суще-
и 3L (рис. 1) образуют стабильные комплек-
ствует в виде димера и других олигомерных
сы со всеми указанными вариантами СА
форм [58-59], с которыми, как было нами ра-
(рис. 2, а-г). Полученные электрофореграм-
нее показано, ДСО также связывается [51].
мы были использованы для расчёта долей свя-
Исследование влияния количества вве-
занного с белком и свободного ДСО, исходя из
дённых в олигонуклеотид додецильных остат-
интенсивности флуоресценции соответствую-
ков на степень сродства ДСО к связыванию
щих полос для каждой из проб. Зависимости
с альбумином показало, что ДСО с одним
количества связанного с белком ДСО от зна-
(1Т, 1N) и один из ДСО с двумя остатками (2Т)
чения соотношения СА : ДСО имеют вид кри-
не образуют с БСА комплексов, устойчи-
вых с выходом на плато в диапазоне 0,67-1 по
вых в условиях электрофоретического ана-
оси абсцисс (рис. 3), что соответствует стехио-
лиза (рис. 1П, а, б и г в Приложении 1). Дру-
метрии образующихся ассоциатов в среднем
гое производное с двумя (2N) и производные
1 : (1,25 ± 0,25) (СА : ДСО, в экв.).
с тремя додецильными остатками (3Т, 3N)
Олигонуклеотидные производные 3R и 3L
способны формировать с БСА комплексы с
являются комплементарными (рис. 1) и могут
более низкой электрофоретической подвиж-
образовывать дуплекс
(3R/3L), температура
ностью в сравнении со свободными ДСО
плавления которого в исследуемых условиях,
(рис. 1П, в, д и е в Приложении 1). При этом в
согласно данным OligoAnalyzer [60], состав-
дорожках с комплексами БСА••2N (рис. 1П, д
ляет ~ 52 °C, что свидетельствует о его стабиль-
в Приложении
1) наблюдается размывание
ности. Было показано, что как при добавлении
флуоресцентного сигнала ДСО вне зависимо-
любого из вариантов исследуемых альбуми-
сти от его количества, добавляемого к белку,
нов к преформированному дуплексу 3R/3L,
что не позволяет оценить стехиометрию фор-
так и при смешивании растворов преформи-
мирования данного комплекса. Стехиометрия
рованных комплексов СА••3R с комплекса-
образования комплексов БСА••3Т и БСА••3N
ми СА••3L наблюдается образование белково-
составила не более 1 : 1,5 (БСА : ДСО), так как
олигонуклеотидных ассоциатов, отличных по
на полученных электрофореграммах при соот-
своей подвижности от ранее зафиксирован-
ношении 1 : 1,5 и выше (рис. 1П, в и е в Прило-
ных (рис. 4). Флуоресцентный сигнал от фор-
жении 1) мы наблюдали накопление флуорес-
мирующихся в образцах, содержащих СА, 3R
центного сигнала от свободного, не связанного
и 3L, комплексов регистрируется на электро-
с белком, ДСО.
фореграмме, в том числе в области кармана
БИОХИМИЯ том 88 вып. 8 2023
СВОЙСТВА ДОДЕЦИЛ-СОДЕРЖАЩИХ ОЛИГОНУКЛЕОТИДОВ
1429
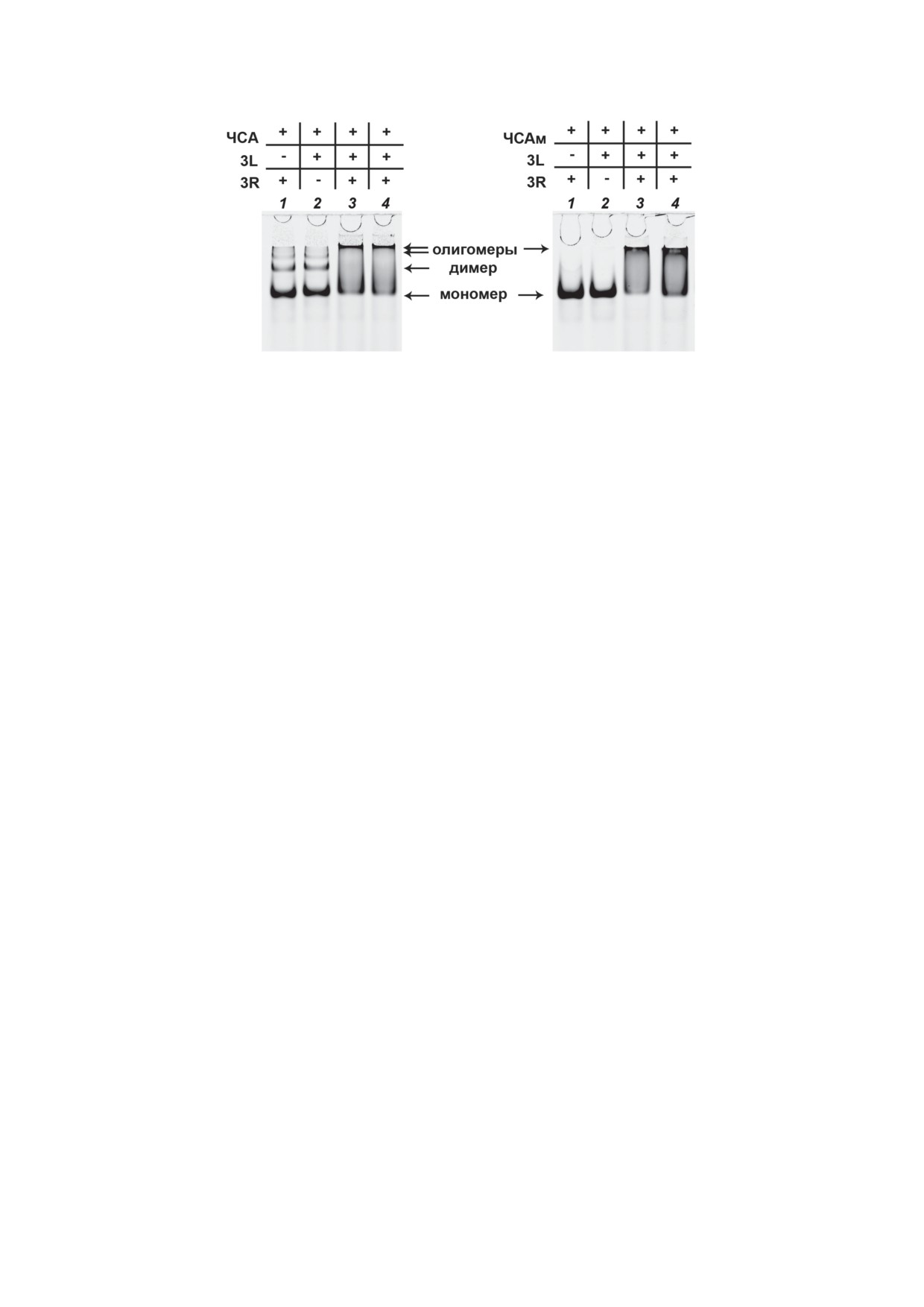
Рис.
4. Электрофореграмма относительной подвижности комплексов ЧСА (слева) и мономерной фракции
ЧСАм (справа) с ДСО 3R и 3L в 8%-ном ПААГ в неденатурирующих условиях. Каждая проба содержала 0,1 нмоль бел-
ка в растворе TAN, 0,175 нмоль 3R и/или 3L (указано знаком «+»), объём проб - 20 мкл. Цифрами указаны номера
дорожек. Методики приготовления проб для нанесения на дорожки 1-4 отличались. 1 и 2 - Готовили разбавлением
стоковых растворов СА и ДСО до необходимых концентраций. 3 - Готовили, добавив в буферный раствор сперва
ДСО 3R и 3L, после чего инкубировали 3 мин при 95 °С, охлаждали до 37 °С, затем добавляли альбумин и инкубирова-
ли 30 мин при 37 °С. 4 - Готовили последовательно: сперва в двух отдельных пробирках смешивали по 0,05 нмоль СА
с 0,175 нмоль 3R и 0,05 нмоль СА с 0,175 нмоль 3L так, чтобы конечный объём в каждой пробирке составил 10 мкл,
после чего данные пробирки инкубировали 30 мин при 37 °С, чтобы образовался комплекс СА••ДСО, а затем содержимое
пробирок объединяли в одну и инкубировали ещё 30 мин при 37 °С. По прошествии времени инкубации пробы наноси-
ли на ПААГ и анализировали, как описано в разделе «Материалы и методы». Стрелками указана область подвижности
соответствующих форм сывороточного альбумина, связанного с ДСО
на границе буферный раствор-ПААГ (рис. 4).
более 3 нм (рис. 1П и 2П в Приложении 2). По-
Данная область является характерной для по-
скольку мелкие частицы образовывали плот-
движности олигомеров сывороточного альбу-
ное покрытие на поверхности, мы предполо-
мина (рис. 4). Полученные результаты свиде-
жили, что в данных условиях при концентрации
тельствуют о частичном образовании высо-
ЧСА 0,4 мкМ это может свидетельствовать о
комолекулярных ассоциатов, вероятно, морфо-
перенасыщении поверхности слюды белком
логия (форма и/или размер) которых препят-
вследствие его хорошей адсорбции. Разбавив
ствует их движению в 8%-ном ПААГ.
образец (3R/3L + ЧСА) в 10 раз, мы снова на-
Таким образом, результаты, полученные с
блюдали редко представленные крупные ча-
помощью метода задержки в геле, свидетель-
стицы аналогичного размера (рис. 3П и 4П
ствуют о способности ДСО связываться с БСА
в Приложении 2). Данные частицы характе-
и ЧСА, включая его очищенную мономер-
ризовались однородной структурой без ка-
ную фракцию, в условиях, близких к физио-
ких-либо отдельно просматривающихся ком-
логическим, с образованием как комплек-
понентов внутри (рис. 5П в Приложении 2).
сов со стехиометрией белок : ДСО в среднем
Слой мелких частиц стал менее плотным
1 : (1,25 ± 0,25) (в экв.), так и высокомолеку-
(рис. 3П и 6П в Приложении 2) и было отмече-
лярных ассоциатов, формирование которых
но, что некоторые из мелких частиц располага-
может быть обусловлено комплементарным
лись на поверхности парами на минимальном
взаимодействием ДСО, связанных с белком.
расстоянии друг от друга (рис. 6П в Прило-
Исследование ассоциатов альбумина с ком-
жении 2).
плементарными додецил-содержащими произ-
В контрольном образце дуплекса 3R/3L
водными методом атомно-силовой микроскопии.
при концентрации ДСО 0,12 мкМ были обна-
Для изучения морфологии полученных высо-
ружены крупные частицы диаметром 101,8 ±
комолекулярных ассоциатов образец, содер-
± 31,6 нм и высотой 30,8 ± 10,7 нм (рис. 7П и 8П
жащий преформированный дуплекс 3R/3L и
в Приложении 2) и единично представленные
ЧСА, был исследован методом АСМ. Было об-
мелкие частицы, диаметр и высота которых не
наружено, что при концентрации ЧСА 0,4 мкМ
превышали 10 и 3 нм соответственно (рис. 9П
(и ДСО - 1,2 мкМ) поверхность слюды с нанесён-
в Приложении 2).
ным образцом (3R/3L + ЧСА) содержит круп-
В контрольном образце 0,04 мкМ ЧСА без
ные частицы округлой формы, диаметром
добавления ДСО мы наблюдали только мно-
165,5 ± 94,3 нм и высотой 28,9 ± 16,9 нм, рас-
жество низких (высотой до 1,5 нм) мелких
положенные на слое значительно более мелких
(диаметром до 20 нм) частиц (рис. 10П и 11П
частиц диаметром 12,0 ± 2,3 нм и высотой не
в Приложении 2).
БИОХИМИЯ том 88 вып. 8 2023
1430
ПАВЛОВА и др.
Исследование ассоциатов альбумина с ком-
ложении 2). Размер частиц округлой формы в
плексами поли(dA) и олиготимидилатными оли-
образце (поли(dA) + 3T + ЧСА), по данным
гонуклеотидами методом атомно-силовой мик-
АСМ, соответствовал таковому для контроль-
роскопии. Для исследования способности свя-
ного образца ЧСА (рис. 12П-14П в Приложе-
зывания СА с ДСО в составе супрамолекуляр-
нии 2). Мы предположили, что в данных усло-
ных ассоциатов методом АСМ был исследован
виях (концентрация ЧСА - 1 мкМ) это снова
образец, состоящий из предварительно сфор-
может свидетельствовать о перенасыщении по-
мированных комплексов поли(dA) с олиго-
верхности слюды альбумином из-за повышен-
тимидилатным ДСО 3Т, к которым затем был
ного сродства именно белка к неорганической
добавлен ЧСА: (поли(dA) + 3T + ЧСА).
поверхности. Поэтому были исследованы об-
На первом этапе были изучены типичные
разцы (поли(dA) + 3T + ЧСА), разбавленные
топографические изображения слюды с нане-
в 100 раз (концентрация ЧСА - 10 нМ). На ти-
сёнными на её поверхность контрольными об-
пичных топографических изображениях слю-
разцами сравнения - ЧСА, а также комплексов
ды (рис. 33П, 35П и 37П в Приложении 2;
(поли(dA) + 3Т) и (поли(dA) + 0Т) (рис. 12П-
рис. 5, ж-и), как и ожидалось, отсутствовал
22П в Приложении 2). Образец (поли(dA) +
ранее наблюдаемый белковый слой, и мы фик-
+ 0Т) был выбран в качестве образца сравне-
сировали крупные (отдельные - до 500 нм в
ния, поскольку известно, что СА может взаи-
диаметре) образования из хорошо различи-
модействовать с нативными НК, не содер-
мых нитевидных составляющих (рис. 5, и).
жащими липофильные остатки [61]. Видно,
Среди данных образований можно выделить
что образец ЧСА визуализируется на поверх-
две популяции крупного
(315,4 ± 70,9 нм)
ности в виде слоя типичных округлых частиц,
(рис. 41П в Приложении 2) и более мелко-
большинство из которых менее 20 нм, но
го (188,3 ± 43,7 нм) размера. Образования в
присутствуют также более крупные частицы
большинстве случаев имели форму близкую
диаметром ~ 25-35 нм (рис. 12П-14П в При-
к округлой. Толщина отдельной нитевидной
ложении 2). Поверхность слюды с образцом
составляющей внутри этих образований была
(поли(dA) + 3Т) (рис. 15П, 17П, 19П и 21П
~ 11 нм (рис. 39П в Приложении 2), что соот-
в Приложении 2) представлена как дцДНК-
ветствует типичным дцДНК-структурам, на-
подобными обрывочными фрагментами ли-
блюдаемым методом АСМ в контрольных экс-
нейной формы, так и эллипсоид-подобными
периментах (рис. 21П и 22П в Приложении 2).
фрагментами (40 × 15 нм; рис. 21П в Приложе-
Полученные изображения для образца
нии 2). В контрольном образце (поли(dA) + 0Т)
(поли(dA) + 0T + ЧСА) при концентрации ЧСА
видна сеть из линейных дцДНК-цепей, кото-
1 мкМ (рис. 24П, 26П и 28П в Приложении 2)
рые, предсказуемо, из-за реализации связы-
были схожими с изображениями контрольного
вания гомо(олиго/поли)-нуклеотидных после-
образца 1 мкМ ЧСА (рис. 12П-14П в Прило-
довательностей способны формировать раз-
жении 2) и для образца (поли(dA) + 3T + ЧСА).
ветвлённые формы ДНК (рис. 16П, 18П, 20П
Однако в образце (поли(dA) + 0T + ЧСА) не
и 22П в Приложении 2). В отличие от образца
было найдено кольцеобразных структур как
(поли(dA) + 3Т), в образце (поли(dA) + 0Т) на-
до (рис. 24П, 26П, 28П в Приложении 2), так
блюдаются более протяжённые цепи (рис. 20П
и после его разбавления (рис. 34П, 36П, 38П
и 22П в Приложении 2). Анализ показывает,
в Приложении 2). После разбавления образца
что в случае (поли(dA) + 0Т) на типичном поле
(поли(dA) + 0T + ЧСА) мы наблюдали нали-
наблюдения масштабом 1 мкм2 визуализируется
чие главным образом линейных и в значитель-
20 ± 1 точек ветвления дцДНК, тогда как для
ной степени укороченных фрагментов дцДНК
(поли(dA) + 3Т) - всего 1-2 точки (рис. 19П
(рис. 38П в Приложении 2), содержащих округ-
и 20П в Приложении 2).
лые, белок-подобные возвышения/утолще-
На поверхности слюды, содержащей
ния как внутри, так и на концах цепи ДНК
(поли(dA) + 3T + ЧСА) мы наблюдали частицы
(рис. 40П в Приложении 2). Похожие округ-
округлой формы и редко представленные коль-
лые возвышения/утолщения, напоминающие
цеобразные структуры (рис. 23П, 25П, 27П, 29П,
частицы белка, мы наблюдали в структурах
30П и 32П в Приложении 2; рис. 5, а, б, г и д).
зафиксированных образований в образце
Внешний диаметр кольцеобразных структур
(поли(dA) + 3T + ЧСА) (рис. 5, и; рис. 39П и 42П
варьировал от 65 до 200 нм (рис. 31П и 32П
в Приложении 2).
в Приложении 2; рис. 5, в и е). Следует отметить,
Таким образом, с помощью метода АСМ
что каждая из зафиксированных нами кольцевых
было обнаружено, что в указанных условиях
структур типично имела несколько утолщённых
результатом взаимодействия ЧСА с дуплек-
участков (рис. 5, в и е; рис. 31П и 32П в При-
сом комплементарных ДСО могут являться
БИОХИМИЯ том 88 вып. 8 2023
СВОЙСТВА ДОДЕЦИЛ-СОДЕРЖАЩИХ ОЛИГОНУКЛЕОТИДОВ
1431
Рис. 5. Топографические изображения слюды, полученные методом АСМ, с нанесённым на неё образцом (поли(dA) +
+ 3Т + ЧСА) в растворе ТАМ до разбавления (а-е) и после (ж-и). Концентрация поли(dA) - 23 мкМ (а-е) и 0,23 мкМ
(ж-и); ДСО - 0,5 мкМ (а-е) и 5 нМ (ж-и); ЧСА - 1 мкМ (а-е) и 10 нМ (ж-и). Пробы готовили и анализировали,
как описано в разделе «Материалы и методы»
округлые высокомолекулярные ассоциаты
кинетические характеристики и эффектив-
диаметром 165,5 ± 94,3 нм и высотой 28,9 ±
ность действия [20, 29-31], а эффективность
± 16,9 нм, а результатом взаимодействия ЧСА
действия антисмысловых олигонуклеотидов,
с комплексами поли(2′-дезокси)адениловой
содержащих остатки жирных кислот, коррели-
кислоты и олиготимидилатного ДСО могут
рует со степенью их сродства к СА [25]. Пара-
являться супрамолекулярные ассоциаты, са-
метры связывания липофильных производных
моорганизованные главным образом в виде
тНК с альбумином, например, стабильность
субмикронных структур. Такие структуры
и стехиометрия формирующихся комплексов,
представлены двумя основными популяциями
зависят от типа модификаций в составе тНК,
размером 315,4 ± 70,9 и 188,3 ± 43,7 нм. При
её последовательности и вторичной структу-
этом их высота не превышала 2,5 нм.
ры [20, 30-32].
В данной работе мы изучили олигонуклео-
тидные производные, содержащие от 1 до 3 ли-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
пофильных додецильных остатков D. После-
довательное увеличение числа остатков D по-
Возможность связывания альбумина с оли-
зволило нам отследить, как повышение гид-
гонуклеотидами продемонстрирована как для
рофобности олигонуклеотида влияло на его
нативных НК [61], так и для их синтетических
сродство к СА. В качестве олигонуклеотидных
аналогов, модифицированных по углеводно-
составляющих ДСО мы использовали либо
фосфатному остову [19], а также для содержа-
гомотимидилатные, либо гетеронуклеотидные
щих липофильные группировки олигонуклео-
последовательности, значительно отличаю-
тидных производных [20, 25, 30-32]. Было
щиеся по таким физико-химическим характе-
отмечено, что липофильные производные
ристикам, как стэкинг азотистых оснований
тНК, обладающие высоким сродством к аль-
внутри цепи и образование вторичных струк-
бумину, демонстрируют улучшенные фармако-
тур [62]. Наше исследование демонстрирует,
БИОХИМИЯ том 88 вып. 8 2023
1432
ПАВЛОВА и др.
что степень сродства ДСО к альбумину сим-
транспортировать в случае напряжённых фи-
батна количеству введённых гидрофобных
зических нагрузок или адренергической сти-
додецильных остатков (рис. 1П в Приложе-
муляции [64]. Установленные сайты связыва-
нии 1). Наличие одного додецильного остатка
ния остатков ЖК в молекуле ЧСА обсуждались
в составе олигонуклеотидов (на примере 1T
ранее [52, 65-68]. В экспериментах in vitro при
и 1N) не обеспечивало стабильности образую-
инкубации СА в присутствии значительных
щихся комплексов БСА••1T и БСА••1N, по
избытков ЖК удалось получить кристалличе-
данным электрофоретического анализа. В слу-
ские структуры ЧСА, в которых c молекулой
чае введения двух остатков D мы регистри-
белка связано 7-8 молекул ЖК [65]. Так, ме-
ровали формирование частично устойчивых
тодом РСА была установлена структура ЧСА в
комплексов БСА••2N, то есть для производно-
комплексе с восемью молекулами лауриновой
го с гетеронуклеотидной последовательностью
(C11H23COOH) кислоты (рис. 6, а), алифатиче-
(рис 1П, д в Приложении 1). В то же время
ский остаток которой близок по длине к доде-
для 2T нам удалось зафиксировать, по всей ви-
цильному (C12H24-) в составе ДСО. Отмечается,
димости, факт диссоциации соответствующего
что сайты связывания 2, 4 и 5 характеризуются
комплекса ДСО с альбумином в процессе ана-
высоким сродством к ЖК, а сайты связыва-
лиза: чётких полос, характеризующих подвиж-
ния 1, 3 и 6 - средним сродством (рис. 6, а) [65,
ность комплексов БСА••2T, не регистрируется.
68]. Сайт 7, по некоторым данным, может яв-
Полученные данные указывают, что нуклео-
ляться одним из главных сайтов связывания
тидная последовательность ДСО может оказы-
для короткоцепочечных ЖК [65]. Мы пред-
вать влияние на стабильность их комплексов
полагаем, что связывание ДСО с альбумином
с альбумином. Разное сродство к связыванию
происходит в местах расположения вышеопи-
с СА в зависимости от нуклеотидной после-
санных высокоаффинных сайтов связывания
довательности ранее было описано для тио-
остатков ЖК, что обеспечивает стабильность
фосфатных аналогов олигонуклеотидов [19].
комплексов СА••ДСО в условиях электрофо-
В нашем случае, вероятно, после закрепления
ретического анализа. Вероятно, именно ком-
алифатических остатков ДСО в сайтах пред-
бинация сайтов связывания, занимаемых до-
полагаемого гидрофобного взаимодействия
децильными остатками первого ДСО, может
с молекулой белка олигонуклеотидная цепь в
определять возможность взаимодействия с СА
зависимости от её последовательности имеет
второй молекулы ДСО. Альбумин является
разные возможности для реализации сети до-
белком, подверженным аллостерической мо-
полнительных низкоспецифических контактов
дуляции, когда связывание некоторых лиган-
белок-НК. При этом наиболее гидрофобные
дов может влиять на взаимодействие других
производные ДСО, содержащие 3 додециль-
лигандов в других сайтах молекулы СА [69],
ных остатка, образовывали стабильные ком-
в том числе делая такое взаимодействие не-
плексы со всеми вариантами образцов СА,
возможным. Также популяции СА••(ДСО)1 и
используемых в работе.
СА••(ДСО)2 могут иметь различную устойчи-
Нами была установлена стехиометрия об-
вость в условиях электрофоретического разде-
разования стабильных комплексов СА••ДСО
ления, в частности, более «нагруженный» ком-
для трёх различных препаратов сывороточ-
плекс белка с ДСО может диссоциировать, не
ных альбуминов: выделенного мономера из
позволяя зафиксировать его наличие методом
фракции V по методу Кона ЧСА, фракции V
задержки в геле.
по методу Кона ЧСА и фракции V по методу
В отличие от ЧСА, о сайтах связывания
Кона БСА (без жирных кислот). Показано,
БСА с ЖК в литературе представлено мень-
что в среднем с молекулой белка любого из
ше данных. Известно, что БСА и ЧСА имеют
исследованных видов СА может связаться
высокую гомологию, оба состоят из 3 (I-III)
1,25 ± 0,25 молекул ДСО с тремя додециль-
также гомологичных доменов, разделённых на
ными остатками. Мы полагаем, что данный
2 субдомена (А и В) [64, 70]. Наглядно сравне-
факт свидетельствует о присутствии в образ-
ние структур данных белков приведено в рабо-
цах комплексов альбумина с ДСО по крайней
те Golianová et al. [71], где также обозначены
мере двух популяций ассоциатов: СА••(ДСО)1,
основные сайты связывания гидрофобных со-
когда с белком связана 1 молекула ДСО, и
единений в БСА. В нашей работе стехиометрия
СА••(ДСО)2 - когда связаны 2 молекулы. Со-
образования комплексов альбумина с ДСО
гласно литературным данным, в физиологи-
отличалась незначительно в случаях с БСА,
ческих условиях молекула ЧСА осуществляет
ЧСА или его мономерной фракции ЧСАм
транспорт до 2 молекул ЖК [63-64]. Большее
(рис. 3), что свидетельствует о реализации схо-
количество ЖК (до 4 молекул) альбумин может
жих механизмов связывания рассматриваемых
БИОХИМИЯ том 88 вып. 8 2023
СВОЙСТВА ДОДЕЦИЛ-СОДЕРЖАЩИХ ОЛИГОНУКЛЕОТИДОВ
1433
Рис. 6. Схематичное изображение: а - структуры ЧСА в комплексе с лауриновой кислотой (С11H23COOH); б-г - пред-
полагаемых комплексов ЧСА с ДСО. Тёмно-зелёным цветом выделена структура белка (а-г), светло-зелёным -
молекулы лауриновой кислоты (а), оранжевым цветом - карбоксильные группы в С11H23COOH (а), волнистой лини-
ей красного цвета - 3R (б-г), волнистой линией синего цвета - 3L (б-г). Арабскими цифрами (а) показаны сайты
связывания жирной кислоты: 1-7 - основные, 2′ - дополнительный, согласно данным Bhattachary et al. [65]. При-
ведённая структура ЧСА (а) опубликована в работе Bhattachary et al. [65] и в банке данных трёхмерных структур бел-
ков и НК PDB (RCSB.org), ID 1E7F. Римские цифры (б-г) указывают на демонстрацию теоретически возможного (I)
и полученного (II) вида расположения полос образца ЧСА (5 мкМ) в комплексе с 3R (15 мкМ) и 3L (15 мкМ) при ана-
лизе методом задержки в геле. Стрелками (б-г) указаны полосы, предположительно соответствующие подвижности
изображённых схематично комплексов белка с ДСО
производных с белками, где превалирующее,
первой популяции, когда с СА связана 1 моле-
гидрофобное, взаимодействие осуществляется
кула ДСО, обусловливает одну из двух наблю-
между додецильными остатками и соответ-
даемых нами полос в образце СА с двумя ком-
ствующими участками в структурах белков.
плементарными ДСО (рис. 4, дорожки 3 и 4) -
Данные об образовании комплексов альбу-
с высокой электрофоретической подвижно-
мина с 18-звенным антисмысловым олигонук-
стью, сопоставимой с таковой для комплексов
леотидом, содержащим на 5′-конце лиганд D1
мономерной формы СА••ДСО (рис. 4, дорож-
(не путать с додецильным остатком D в теку-
ки 1 и 2). В образце, содержащем 2 комплемен-
щем исследовании) для связывания с белком,
тарных ДСО и белок, данная полоса может со-
были опубликованы в работе Lacroix et al. [20].
ответствовать комплексам как состава СА••3R,
Как и в нашем случае, сродство к связыва-
СА••3L, так и дуплексу, связанному с моно-
нию с белком для олигонуклеотидов, содер-
мерной формой белка СА••3R/3L, поскольку
жащих лиганд D1, сохранялось вне зависимо-
в данных условиях подвижность указанных
сти от вида сывороточного альбумина - ЧСА
комплексов отличается незначительно, так
или БСА. Кроме того, авторами было проде-
как молекулярная масса ЧСА (~ 66,5 кДа [54])
монстрировано аналогичное сродство к свя-
существенно превышает молекулярную массу
зыванию D1-содержащих олигонуклеотидов с
данных ДСО (~ 7,5 кДа). При этом вторая по-
мышиным СА.
пуляция комплексов типа СА••(ДСО)2, несу-
Важной частью работы стало исследование
щих по 2 молекулы ДСО, имеет возможность
возможности реализации Уотсон-Криковских
и реализует её в образце СА с комплементар-
взаимодействий между олигонуклеотидными
ными ДСО в виде формирования структур,
цепями двух комплементарных ДСО, связан-
молекулярность которых по белковому ком-
ных в комплексах с альбумином. Полученные
поненту явно превышает значение как мини-
нами данные (рис. 4) по изучению подвиж-
мум в 3 молекулы, что видно при анализе рас-
ности ассоциатов, образованных комплемен-
положения на электрофореграмме второй
тарными ДСО 3R и 3L с альбумином, также
полосы (рис. 4, дорожки 3 и 4) - с очень низ-
свидетельствуют в пользу наличия в комплек-
кой подвижностью, характерной для области
сах СА с одним ДСО как минимум двух по-
подвижности олигомеров альбумина в кон-
пуляций, СА••(ДСО)1 и СА••(ДСО)2. Наличие
трольных пробах (рис. 4, дорожки 1 и 2) [59].
БИОХИМИЯ том 88 вып. 8 2023
1434
ПАВЛОВА и др.
Значимое количество флуоресцентно мечен-
при добавлении даже
6-кратного избытка
ной ДНК в этом случае регистрируется лока-
одного из ДСО их формирования не наблю-
лизованной вблизи дна кармана для нанесения
далось (рис. 2П в Приложении 1; дорожка 1),
образца.
что позволяет сделать вывод о вкладе имен-
На рис.
6, б-г под соответствующими
но Уотсон-Криковских взаимодействий в их
римскими цифрами приведены теоретически
организацию.
возможные (I) и экспериментально наблю-
Для изучения морфологии образующихся
даемые (II) изображения электрофореграмм,
высокомолекулярных ассоциатов мы исполь-
характеризующих подвижность ассоциатов с
зовали метод АСМ. Было обнаружено, что в
участием ЧСА, 3R и 3L (рис. 4). Заметим, если
образце
(3R/3L + ЧСА) присутствуют круп-
предположить, что с СА связывается 1 молеку-
ные по диаметру и высоте округлые частицы
ла 3R или 3L, тогда образующиеся комплексы
(рис. 1П, 3П-5П в Приложении 2), структура
СА••3R и СА••3L (схематично представлены
которых ввиду своей однородной плотности
на рис. 6, б) при реализации комплементарного
не позволяет однозначно заключить, участвует
взаимодействии должны привести к «сцепке»
ли в их образовании белок или нет. С одной
2 молекул белка (СА••3R/3L••СА) при усло-
стороны, похожие крупные частицы были за-
вии, что связанные молекулы ДСО способны
фиксированы нами в контрольном образце ду-
реализовать такое взаимодействие. Подвиж-
плекса 3R/3L (рис. 7П и 8П в Приложении 2),
ность такого ассоциата (СА••3R/3L••СА) (схе-
где они, по всей видимости, являются мицел-
матично представлен на рис. 6, в) не должна
лами, сформированными ДСО за счёт нали-
быть ниже подвижности олигомеров альбу-
чия липофильных додецильных остатков по
мина, состоящих из более чем 2 молекул бел-
обоим концам дуплекса. Образование мицелл
ка (рис. 4), а должна быть близка к подвижно-
характерно как для ДСО по ранее получен-
сти димера СА. Таких полос при соотношении
ным нами данным [51], так и для других про-
СА : 3R : 3L = 1 : 1,5 : 1,5 (в экв.) и времени
изводных олигонуклеотидов с липофильными
инкубации проб 30 мин мы не наблюдали
остатками [31], при этом размер мицелл зави-
(рис. 4; рис. 6, в). Однако при соотношении
сит от условий экспериментов (концентрации
ЧСАм : 3R : 3L = 1 : 0,25 : 0,25 (в экв.) и време-
ДСО, рН среды, вида и концентрации солей,
ни инкубации 15 мин нам удалось зафикси-
температуры, времени инкубации). С другой
ровать ассоциат (рис. 2П в Приложении 1; до-
стороны, электрофоретическая подвижность
рожка 8 на панелях а и б, обозначен стрелкой
образцов 3R/3L и (3R/3L + ЧСА) значительно
красного цвета), по всей видимости, состава
отличается (рис. 3П в Приложении 1). Обра-
(ЧСАм••3R/3L••ЧСАм), поскольку его подвиж-
зец 3R/3L при концентрации каждого из ДСО
ность была сопоставима с подвижностью
15 мкМ имеет вид нечёткой растянутой поло-
димера ЧСА и комплексов димера ЧСА с од-
сы в геле, что свидетельствует в пользу фор-
ним ДСО (рис. 2П в Приложении 1; дорожка 2
мирования мицелл дуплексом ДСО. Однако
на панели б и дорожки 6, 7 на панелях а и б).
при добавлении к дуплексу 3R/3L белка мы
При увеличении количества комплементар-
не наблюдали подобного вида полос в образце
ных ДСО мы наблюдали постепенное умень-
(3R/3L + ЧСА), часть которого визуализиру-
шение подвижности данной полосы (рис. 2П
ется ближе к области кармана для нанесения.
в Приложении 1; дорожки 9-12 на панелях
Это может свидетельствовать о том, что фор-
а и б), вероятно, вызванное участием ассоциата
мирование высокомолекулярных ассоциатов
(ЧСАм••3R/3L••ЧСАм) в дальнейшем образо-
в данном случае происходит уже с участием
вании более высокомолекулярных ассоциатов.
белка, молекулярная масса которого значи-
Если же с молекулой белка способно свя-
тельно превышает таковую для ДСО и может
заться 2 и более молекул ДСО, то в этом слу-
приводить к существенному снижению элек-
чае возможно образование более сложно
трофоретической подвижности образующихся
организованных высокомолекулярных ассо-
ассоциатов. Мы предположили, что размер ду-
циатов (схематично представлены на рис. 6, г)
плекса 3R/3L (~ 7 нм из расчёта 0,34 нм на одно
и их подвижность может быть значительно
нуклеотидное звено) слишком мал в сравнении
ниже мономерной и димерной форм бел-
с данными о размере ЧСА (~ 6-13 нм) [38, 72],
ка (рис. 4). В нашем случае при соотношении
чтобы было возможным отличить с помощью
СА : 3R : 3L = 1 : 1,5 : 1,5 (в экв.) и времени ин-
метода АСМ олигонуклеотидные цепи в соста-
кубации проб 30 мин мы наблюдали именно
ве высокомолекулярных ассоциатов от частиц,
это (рис. 4; рис. 6, б-г). Образование данных
являющихся молекулами белка, и оценить ко-
высокомолекулярных ассоциатов происходит
личество его молекул, входящее в их состав.
только в присутствии комплементарных ДСО;
Поэтому далее мы исследовали образец ЧСА
БИОХИМИЯ том 88 вып. 8 2023
СВОЙСТВА ДОДЕЦИЛ-СОДЕРЖАЩИХ ОЛИГОНУКЛЕОТИДОВ
1435
с комплексами (поли(dA) + 3Т). Теоретически
тимидилата с полиадениловой кислотой. При
с молекулой полиадениловой кислоты может
этом более высокое сродство додецильных
связаться несколько молекул олиготимиди-
остатков к белку послужило причиной самоор-
лата с образованием протяжённых дуплексов,
ганизации разбросанных до этого по поверх-
которые также могут соединяться по типу фор-
ности обрывочных фрагментов ДНК в образ-
мирования конкатамеров в случае связыва-
це (поли(dA) + 3Т) в сравнительно крупные
ния олиготимидилата с разными молекулами
округлые образования, состоящие из отдель-
поли(dA). Мы предположили, что такая систе-
ных нитевидных компонентов (рис. 5, ж-и),
ма будет более пригодна для сравнительного
в образце (поли(dA) + 3Т + ЧСА). Толщина дан-
исследования связывания альбумина с ДСО с
ных «нитей» близка к таковой для фрагмента
помощью метода АСМ по аналогии примене-
ДНК и составила ~ 11 нм (рис. 39П в При-
ния данного метода для визуализации связан-
ложении
2). Следует отметить, что в дан-
ных с протяжёнными ДНК других белков [73].
ных структурах были различимы округлые
Полученные нами данные исследования
возвышения/утолщения, близкие по раз-
супрамолекулярных ассоциатов методом АСМ
меру к частицам белка (рис. 5, и; рис. 42П
свидетельствуют, что без альбумина как натив-
в Приложении 2). Вероятно, реализующиеся
ный 0T, так и додецил-содержащий 3T олиго-
взаимодействия СА••ДСО в качестве «скре-
тимидилаты образовывали с поли(dА)-матри-
пок» могли способствовать объединению ком-
цей конкатамерные ДНК-подобные комплек-
плексов (поли(dA) + 3Т) в зафиксированные
сы (рис. 15П-22П в Приложении 2): в обоих
более сложные структуры в образце (поли(dA) +
образцах наблюдали фрагменты высотой до
+ 3Т + ЧСА). Данные популяции округлых
1,5 нм и шириной в местах без утолщений/воз-
или близких к овальной форме крупных
вышений - 10-15 нм (рис. 21П и 22П в При-
(315,4 ± 70,9 нм) и менее крупных
(188,3 ±
ложении 2). Наблюдаемые более короткие фраг-
± 43,7 нм) структур, сформированных из пере-
менты и существенно меньшее количество
плетённых «нитей», таким образом, можно
точек их разветвления (рис. 19П и 20П в При-
квалифицировать как прототип наногелей
ложении 2) в образце (поли(dA) + 3Т) в срав-
с элементами НК-архитектоники по анало-
нении с (поли(dA) + 0Т) позволяют предполо-
гии с работами Thelu et al [41], Li et al [46],
жить, что наличие додецильных остатков в
Zhang и Tung [78] и Lacroix et al [79]. При
ДСО способствует ограничению длины обра-
этом зарегистрированные нами кольцеобраз-
зующихся конкатамерных комплексов в об-
ные структуры (рис. 5, а-е) в этом же образце
разце (поли(dA) + 3Т). Вероятно, додецильные
(поли(dA) + 3Т + ЧСА) до его разбавления
остатки могут препятствовать кооперативному
могли являться лишь «выпетливаниями» дан-
связыванию содержащих их олиготимидила-
ных образований в избыточно присутствую-
тов на поли(dA)-матрице, поэтому образец
щем белковом слое.
(поли(dA) + 3Т) имел вид неупорядоченно рас-
С учётом известной способности СА к
положенных укороченных фрагментов (рис. 19П
желированию с образованием гелей, отличаю-
в Приложении 2). Либо алифатический доде-
щихся фазовым состоянием
[80], а также
цильный остаток способствует сворачива-
стремительного развития направления ДНК-
нию/изгибу НК-цепи за счёт образования
оригами и дизайна основанных на данном
гидрофобных скрепок ДСО, расположенных
принципе систем доставки для тНК, получен-
на одной или разных поли(dА)-матрицах, как
ные результаты свидетельствуют о высоком
было показано ранее [74-75]. Изображение же
потенциале ДСО и СА как соединений-состав-
образца (поли(dA) + 0Т) соответствовало виду
ляющих для создания новых многокомпонент-
«классических» конкатамерных комплексов
ных самоорганизующихся систем, в том числе
(рис. 18П и 20П в Приложении 2) по аналогии
обладающих свойствами биосовместимых на-
с работой Виноградовой и соавт. [76]. Нали-
ночастиц или наногелей. Важность создания
чие утолщений/возвышений в обоих образцах
подобных систем для решения задач молеку-
(рис. 21П и 22П в Приложении 2) также может
лярной биологии и биомедицины отмечается
быть обусловлено возможностью образования
ведущими научными изданиями [2]. При этом
триплексов в данных образцах [77].
следующим этапом подтверждения потенциала
Добавление альбумина к контрольному об-
систем доставки на основе СА и ДСО должны
разцу (полиd(A) + 0Т) привело к уменьшению
стать исследования их активности, возможных
наблюдаемой длины конкатамерных комплек-
токсических эффектов и предполагаемых ме-
сов, по нашему предположению, из-за срод-
ханизмов действия с использованием не толь-
ства НК к альбумину [19, 61], что также может
ко клеточных культур, но и объектов in vivo,
нарушать кооперативность связывания олиго-
например, специальных линий мышей.
БИОХИМИЯ том 88 вып. 8 2023
1436
ПАВЛОВА и др.
ЗАКЛЮЧЕНИЕ
М.С. Купрюшкин, Е.С. Дюдеева - синтез и
постсинтетическое выделение ДСО и контроль-
Полученные в представленном исследова-
ных олигонуклеотидов; И.А. Бауэр, А.С. Чу-
нии результаты, наряду с достаточно быстрым
баров - выделение и очистка мономерной фрак-
связыванием ДСО с сывороточным альбуми-
ции ЧСА; Д.В. Пышный, И.А. Пышная - кон-
ном в условиях, близких к физиологическим, а
цепция и руководство работой, редактирование
также ранее продемонстрированными факта-
текста статьи.
ми, что связывание ДСО с СА на 30% снижает,
Финансирование. Исследование проводи-
но не предотвращает их проникновения внутрь
лось в рамках государственного задания
клеток in vitro [51] и отсутствием цитотоксич-
ИХБФМ СО РАН № 121031300042-1.
ности в рабочем диапазоне концентраций [50],
Благодарности. А.С.П. выражает благодар-
делают привлекательным использование таких
ность к.ф.-м.н., зав. ЛСТБ ИХБФМ СО РАН
олигонуклеотидных производных для разра-
Ломзову А.А. и к.ф.-м.н., н.с. ЛСТБ ИХБФМ
ботки препаратов-транспортёров тНК, в том
СО РАН Голышеву В.М. за полезные консуль-
числе на основе аутологичной крови паци-
тации по использованию метода АСМ. В ра-
ента. На следующих этапах необходимо про-
боте использовали оборудование VersaDocTM
должить исследования, чтобы изучить другие
MP 4000 Molecular Imager® System («Bio-Rad
свойства ДСО, способные оказать влияние
Laboratories», США) Объединённого Центра
на их фармакодинамику и фармакокинетику
геномных, протеомных и метаболомных ис-
в организме.
следований (ОЦГПМИ) ИХБФМ СО РАН.
Конфликт интересов. Авторы заявляют об
Вклад авторов. А.С. Павлова - реализация
отсутствии конфликта интересов.
концепции исследования, постсинтетическое
Соблюдение этических норм. Настоящая
выделение ДСО, проведение экспериментов
статья не содержит описания каких-либо ис-
(метод задержки в ПААГ и АСМ), обработка
следований с участием людей или животных
и анализ полученных данных, написание тек-
в качестве объектов.
ста статьи; В.В. Илющенко - проведение экс-
Дополнительные материалы. Приложение
периментов (метод задержки в ПААГ), анализ
к статье опубликовано на сайте журнала
и обработка первичных данных; Т.Д. Жарков,
СПИСОК ЛИТЕРАТУРЫ
1. Smith, C. I. E., and Zain, R. (2019) Therapeutic oligo-
6. Mullard, A. (2023) 2022 FDA approvals, Nat. Rev Drug.
nucleotides: state of the art, Annu. Rev. Pharmacol.
Discov., 22, 83-88, doi: 10.1038/d41573-023-00001-3.
Toxicol., 59, 605-630, doi: 10.1146/annurev-pharmtox-
7. Egli, M., and Manoharan, M. (2023) Chemistry,
010818-021050.
structure and function of approved oligonucleotide
2. Roberts, T. C., Langer, R., and Wood, M. J. A. (2020)
therapeutics, Nucleic Acids Res.,
51,
2529-2573,
Advances in oligonucleotide drug delivery, Nat. Rev
doi: 10.1093/nar/gkad067.
Drug. Discov., 19, 673-694, doi: 10.1038/s41573-020-
8. Juliano, R. L. (2016) The delivery of therapeutic
0075-7.
oligonucleotides, Nucleic Acids Res., 44, 6518-6548,
3. Gökirmak, T., Nikan, M., Wiechman, S., Prakash,
doi: 10.1093/nar/gkw236.
T. P., Tanowitz, M., and Seth, P. P. (2021) Overcoming
9. Juliano, R. L. (2021) Manipulation of the endosome
the challenges of tissue delivery for oligonucleotide
trafficking machinery: implications for oligonucleotide
therapeutics, Trends Pharmacol. Sci., 42, 588-604,
delivery, Biomedicines,
9,
512, doi:
10.3390/
doi: 10.1016/j.tips.2021.04.010.
biomedicines9050512.
4. Bakowski, K., and Vogel, S. (2022) Evolution of
10. Tran, P., Weldemichael, T., Liu, Z., and Li, H.-Yu
complexity in non-viral oligonucleotide deliv-
(2022) Delivery of oligonucleotides: efficiency with
ery systems: from gymnotic delivery through bio-
lipid conjugation and clinical outcome, Pharmaceutics,
conjugates to biomimetic nanoparticles, RNA
14, 342, doi: 10.3390/pharmaceutics14020342.
Biol.,
19,
1256-1275, doi:
10.1080/15476286.2022.
11. Kupryushkin, M. S., Pyshnyi, D. V., and Stetsenko,
2147278.
D. A. (2014) Phosphoryl guanidines: A new type of
5. Crooke, S. T., Baker, B. F., Crooke, R. M., and Liang,
nucleic acid analogues, Acta Naturae, 6, 116-118,
X.-H. (2021) Antisense technology: an overview and
10.32607/20758251-2014-6-4-116-118.
prospectus, Nat. Rev Drug. Discov., 20, 427-453,
12. Lomzov, A. A., Kupryushkin, M. S., Shernyukov, A. V.,
doi: 10.1038/s41573-021-00162-z.
Nekrasov, M. D., Dovydenko, I. S., Stetsenko, D. A.,
БИОХИМИЯ том 88 вып. 8 2023
СВОЙСТВА ДОДЕЦИЛ-СОДЕРЖАЩИХ ОЛИГОНУКЛЕОТИДОВ
1437
and Pyshnyi, D. V. (2019) Diastereomers of a mo-
Characterization of the interactions of chemically-
no-substituted phosphoryl guanidine trideoxyribo-
modified therapeutic nucleic acids with plasma
nucleotide: isolation and properties, Biochem. Bio-
proteins using a fluorescence polarization assay,
phys. Res. Commun.,
513,
807-811, doi:
10.1016/
Nucleic Acids Res., 47, 1110-1122, doi: 10.1093/nar/
j.bbrc.2019.04.024.
gky1260.
13.
Miroshnichenko, S. K., Patutina, O. A., Burakova,
20.
Lacroix, A., Fakih, H. H., and Sleiman, H. F. (2020)
E. A., Chelobanov, B. P., Fokina, A. A., Vlassov,
Detailed cellular assessment of albumin-bound
V. V., Altman, S., Zenkova, M. A., and Stetsenko,
oligonucleotides: Increased stability and lower non-
D. A.
(2019) Mesyl phosphoramidate antisense
specific cell uptake, J. Control. Release, 324, 34-46,
oligonucleotides as an alternative to phosphorothioates
doi: 10.1016/j.jconrel.2020.04.020.
with improved biochemical and biological properties,
21.
Crooke, S. T., Seth, P. P., Vickers, T. A., and Liang,
Proc. Natl. Acad. Sci. USA,
116,
1229-1234,
X.-H. (2020) The interaction of phosphorothioate-
doi: 10.1073/pnas.1813376116.
containing RNA targeted drugs with proteins is a
14.
Pavlova, A. S., Yakovleva, K. I., Epanchitseva, A. V.,
critical determinant of the therapeutic effects of
Kupryushkin, M. S., Pyshnaya, I. A., Pyshnyi, D. V.,
these agents, J. Am. Chem. Soc., 142, 14754-14771,
Ryabchikova, E. I., and Dovydenko, I. S. (2021) An
doi: 10.1021/jacs.0c04928.
influence of modification with phosphoryl guanidine
22.
Kim, W., Ly, N. K., He, Y., Li, Y., Yuan, Zh., and
combined with a 2′-O-methyl or 2′-fluoro group on
Yeo, Y. (2023) Protein corona: friend or foe? Co-
the small-interfering-RNA effect, Int. J. Mol. Sci., 22,
opting serum proteins for nanoparticle delivery,
9784, doi: 10.3390/ijms22189784.
Adv. Drug Deliv. Rev., 192, 114635, doi: 10.1016/
15.
Anderson, B. A., Freestone, G. C., Low, O., De-
j.addr.2022.114635.
Hoyos, C. L., Drury, III, W. J., Østergaard, M. E.,
23.
Chen, Z., Chen, X., Huang, J., Wang, J., and
Migawa, M. T., Fazio, M., Wan, W. B., Berdeja, A.,
Wang, Z.
(2022) Harnessing protein corona for
Scandalis, E., Burel, S. A., Vickers, T. A., Crooke,
biomimetic nanomedicine design, Biomimetics, 7, 126,
S. T., Swayze, E. E., Liang, X., and Seth, P. P. (2021)
doi: 10.3390/biomimetics7030126.
Towards next generation antisense oligonucleotides:
24.
Chernikov, I. V., Gladkikh, D. V., Meschaninova,
mesylphosphoramidate modification improves thera-
M. I., Ven’yaminova, A. G., Zenkova, M. A., et al.
peutic index and duration of effect of gapmer antisense
(2017)
Cholesterol-containing nuclease-resistant
oligonucleotides, Nucleic Acids Res., 49, 9026-9041,
siRNA accumulates in tumors in a carrier-free mode
doi: 10.1093/nar/gkab718.
and silences MDR1 gene, Mol. Ther., 6, 209-220,
16.
Kandasamy, P., Liu, Y., Aduda, V., Akare, S., Alam, R.,
doi: 10.1016/j.omtn.2016.12.011.
Andreucci, A., Boulay, D., Bowman, K., Byrne, M.,
25.
Prakash, T. P., Mullick, A. E., Lee, R. G., Yu, J.,
Cannon, M., Chivatakarn, O., Shelke, J. D., Iwamoto, N.,
Yeh, S. T., Low, A., Chappell, A. E., Østergaard,
Kawamoto, T., Kumarasamy, J., Lamore, S., Le-
M. E., Murray, S., Gaus, H. J., Swayze, E. E., and
maitre, M., Lin, X., Longo, K., Looby, R., Marappan, S.,
Seth, P. P. (2019) Fatty acid conjugation enhances
Metterville, J., Mohapatra, S., Newman, B., Paik,
potency of antisense oligonucleotides in muscle,
I.-H., Patil, S., Purcell-Estabrook, E., Shimizu, M.,
Nucleic Acids Res., 47, 6029-6044, doi: 10.1093/
Shum, P., Standley, S., Taborn, K., Tripathi, S.,
nar/gkz354.
Yang, H., Yin, Y., Zhao, X., Dale, E., and Vargeese, S.
26.
Benizri, S., Gissot, A., Martin, A., Vialet, B.,
(2022) Impact of guanidine-containing backbone link-
Barthelemy, P., and Grinstaff, M. W. (2019) Bioconju-
ages on stereopure antisense oligonucleotides in the
gated oligonucleotides: recent developments and ther-
CNS, Nucleic Acids Res., 50, 5401-5423, doi: 10.1093/
apeutic applications, Bioconjugate Chem., 30, 366-383,
nar/gkac037.
doi: 10.1021/acs.bioconjchem.8b00761.
17.
Hall, J.
(2023) Future directions for medicinal
27.
Kupryushkin, M. S., Zharkov, T. D., Ilina, E. S.,
chemistry in the field of oligonucleotide therapeutics,
Markov, O. V., Kochetkova, A. S., Akhmetova, M. M.,
RNA, 29, 423-433, doi: 10.1261/rna.079511.122.
Lomzov, A. A., Pyshnyi, D. V., Lavrik, O. A., and
18.
Godfrey, C., Desviat, L. R, Smedsrød, B., Piétri-
Khodyreva, S. N. (2021) Triazinylamidophosphate
Rouxel, F., Denti, M. A., Disterer, P., Lorain, S.,
oligonucleotides: synthesis and study of their
Nogales-Gadea, G., Sardone, V., Anwar, R.,
interaction with cells and DNA-binding proteins,
Andaloussi, S. E., Lehto, T., Khoo, B., Brolin, C.,
Russ. J. Bioorganic Chem., 47, 719-733, doi: 10.1134/
van Roon-Mom, W. M. C., Goyenvalle, A., Aartsma-
S1068162021030110.
Rus, A., and Arechavala-Gomeza, V. (2017) Delivery
28.
Brown, K. M., Nair, J. K., Janas, M. M., Anglero-
is key: lessons learnt from developing splice-switching
Rodriguez, Y. I., Dang, L. T. H., Peng, H., Theile,
antisense therapies, EMBO Mol. Med., 9, 545-557,
C. S., Castellanos-Rizaldos, E., Brown, C., Foster, D.,
doi: 10.15252/emmm.201607199.
Kurz, J., Allen, J., Maganti, R., Li, J., Matsuda, S.,
19.
Gaus, H. J., Gupta, R., Chappell, A. E., Østergaard,
Stricos, M., Chickering, T., Jung, M., Wassarman, K.,
M. E., Swayze, E. E., and Seth, P. P.
(2019)
Rollins, J., Woods, L., Kelin, A., Guenther, D. C.,
БИОХИМИЯ том 88 вып. 8 2023
1438
ПАВЛОВА и др.
Mobley, M. W., Petrulis, J., McDougall, R., Racie, T.,
39.
Merlot, A. M., Kalinowski, D. S., and Richardson,
Bombardie, J., Cha, D., Agarwal, S., Johnson, L.,
D. R. (2014) Unraveling the mysteries of serum
Jiang, Y., Lentini, S., Gilbert, J., Nguyen, T., Chigas, S.,
albumin - more than just a serum protein, Front.
LeBlanc, S., Poreci, U., Kasper, A., Rogers, A. B.,
Physiol., 5, 299, doi: 10.3389/fphys.2014.00299.
Chong, S., Davis, W., Sutherland, J. E., Castonero, A.,
40.
Zhao, P., Wang, Y., Wu, A., Rao, Y., and Huang, Y.
Milstein, S., Schlegel, M. K., Zlatev, I., Charisse, K.,
(2018) Roles of albumin-binding proteins in can-
Keating, M., Manoharan, M., Fitzgerald, K., Wu,
cer progression and biomimetic targeted drug de-
J.-T., Maier, M. A., and Jadhav, V. (2022) Expanding
livery, ChemBioChem, 19, 1796-1805, doi: 10.1002/
RNAi therapeutics to extrahepatic tissues with
cbic.201800201.
lipophilic conjugates, Nat. Biotechnol., 40, 1500-1508,
41.
Thelu, H. V. P., Atchimnaidu, S., Perumal, D.,
doi: 10.1038/s41587-022-01334-x.
Harikrishnan, K. S., Vijayan, S., and Varghese, R.
29.
Biscans, A., Caiazzi, J., McHugh, N., Hariharan, V.,
(2019) Self-assembly of an aptamer-decorated, DNA-
Muhuri, M., and Khvorova, A. (2021) Docosanoic
protein hybrid nanogel: a biocompatible nanocarrier
acid conjugation to siRNA enables functional and safe
for targeted cancer therapy, ACS Appl. Bio Mater.,
delivery to skeletal and cardiac muscles, Mol. Ther.,
2, 5227-5234, doi: 10.1021/acsabm.9b00323.
29, 1382-1394, doi: 10.1016/j.ymthe.2020.12.023.
42.
Huang, J., Ma, W., Sun, H., Wang, H., He, X.,
30.
Sarrett, S. M., Werfel, T. A., Lee, L., Jackson, M. A.,
Cheng, H., Huang, M., Lei, Y., and Wang, K. (2020)
Kilchrist, K. V., Brantley-Sieders, D., and Duvall,
Self-assembled DNA nanostructures-based nano-
C. L. (2017) Lipophilic siRNA targets albumin in
carriers enabled functional nucleic acids delivery,
situ and promotes bioavailability, tumor penetration,
ACS Appl. Bio Mater., 3, 2779-2795, doi: 10.1021/
and carrier-free gene silencing, Proc. Natl. Acad.
acsabm.9b01197.
Sci. USA,
114, E6490-E6497, doi:
10.1073/pnas.
43.
Harris, M. A., Kuang, H., Schneiderman, Z., Shiao,
1621240114.
M. L., Crane, A. T., Chrostek, M. R., Tăbăran,
31.
Jin, C., Zhang, H., Zou, J., Liu, Y., Zhang, L.,
A.-F., Pengo, T., Liaw, K., Xu, B., Lin, L., Chen,
Li, F., Wang, R., Xuan, W., Ye, M., and Tan, W. (2018)
C. C., O’Sullivan, M. G., Kannan, R., Low, W. C.,
Floxuridine homomeric oligonucleotides “Hitchhike”
and Kokkoli, E. (2021) ssDNA nanotubes for selec-
with albumin in situ for cancer chemotherapy,
tive targeting of glioblastoma and delivery of doxo-
Angew. Chem. Int. Ed., 57, 8994-8997, doi: 10.1002/
rubicin for enhanced survival, Sci. Adv., 7, eabl5872,
anie.201804156.
doi: 10.1126/sciadv.abl5872.
32.
Chappell, A. E., Gaus, H. J., Berdeja, A., Gupta, R.,
44.
Liu, J., Chen, L., Zhai, T., Li, W., Liu, Y., and
Jo, M., Prakash, T. P., Oestergaard, E. E., Swayze,
Gu, H. (2022) Programmable assembly of amphiphilic
E. E., and Seth, P. P. (2020) Mechanisms of palmit-
DNA through controlled cholesterol stacking,
ic acid-conjugated antisense oligonucleotide distri-
J. Am. Chem. Soc., 144, 16598-16603, doi: 10.1021/
bution in mice, Nucleic Acids Res., 48, 4382-4395,
jacs.2c06610.
doi: 10.1093/nar/gkaa164.
45.
Ma, W., Zhan, Y., Zhang, Y., Mao, C., Xie, X.,
33.
Sleep, D. (2015) Albumin and its application in
and Lin, Y. (2021) The biological applications of
drug delivery, Expert Opin. Drug Deliv., 12, 793-812,
DNA nanomaterials: current challenges and future
doi: 10.1517/17425247.2015.993313.
directions, Signal Transduct. Target. Ther., 6, 351,
34.
Hoogenboezem, E. N., and Duvall, C. L. (2018)
doi: 10.1038/s41392-021-00727-9.
Harnessing albumin as a carrier for cancer therapies,
46.
Li, J., Zheng, C., Cansiz, S., Wu, C., Xu, J., Cui, C.,
Adv. Drug Deliv. Rev., 130, 73-89, doi: 10.1016/
Liu, Y., Hou, W., Wang, Y., Zhang, L., Teng, I-T.,
j.addr.2018.07.011.
Yang, H.-H., and Tan, W.
(2015) Self-assembly
35.
Varanko, A., Saha, S., and Chilkoti, A. (2020) Recent
of DNA nanohydrogels with controllable size and
trends in protein and peptide-based biomaterials for
stimuli-responsive property for targeted gene therapy,
advanced drug delivery, Adv. Drug Deliv. Rev., 156,
J. Am. Chem. Soc., 137, 1412-1415, doi: 10.1021/
133-187, doi: 10.1016/j.addr.2020.08.008.
ja512293f.
36.
Prajapati, R., and Somoza, Á.
(2021) Albumin
47.
Raniolo, S., Unida, V., Vindigni, G., Stolfi, C.,
nanostructures for nucleic acid delivery in cancer:
Iacovelli, F., Desideri, A., and Biocca, S.
(2021)
current trend, emerging issues, and possible solutions,
Combined and selective miR-21 silencing and doxo-
Cancers, 13, 3454, doi: 10.3390/cancers13143454.
rubicin delivery in cancer cells using tailored DNA
37.
Chubarov, A. S. (2022) Serum albumin for magnet-
nanostructures, Cell Death Dis., 12, 7, doi: 10.1038/
ic nanoparticles coating, Magnetochemistry, 8, 13,
s41419-020-03339-3.
doi: 10.3390/magnetochemistry8020013.
48.
Wang, Y., Cheng, J., Zhao, D., Liu, Y., Luo, T.,
38.
Hu, H., Quintana, J., Weissleder, R., Parangi, S.,
Zhong, Y.-F., Mo, F., Kong, X.-Y., and Song, J. (2020)
and Miller, M. (2022) Deciphering albumin-directed
Designed DNA nanostructure grafted with erlotinib
drug delivery by imaging, Adv. Drug Deliv. Rev., 185,
for non-small-cell lung cancer therapy, Nanoscale,
114237, doi: 10.1016/j.addr.2022.114237.
12, 23953, doi: 10.1039/d0nr06945k.
БИОХИМИЯ том 88 вып. 8 2023
СВОЙСТВА ДОДЕЦИЛ-СОДЕРЖАЩИХ ОЛИГОНУКЛЕОТИДОВ
1439
49.
Wang, H., Xiao, H., Zhu, X., Liu, Y., Fu, Z., Li, C.,
and physico-chemical properties, Jordan J. Chem., 2,
Lu, C., and Yang, H. (2021) A cyanine-mediated self-
169-182.
assembly system for the construction of a two-in-one
60.
Owczarzy, R., Tataurov, A. V., Wu, Y., Manthey, J. A.,
nanodrug, Angew. Chem. Int. Ed., 60, 21226-21230,
McQuisten, K. A., Almabrazi, H. G., Pedersen, K. F.,
doi: 10.1002/anie.202108393.
Lin, Y., Garretson, J., McEntaggart, N. O., Sailor,
50.
Markov, O. V., Filatov, A. V., Kupryushkin, M. S.,
C. A., Dawson, R. B., and Peek, A. S. (2008) IDT
Chernikov, I. V., Patutina, O. A., Strunov, A. A.,
SciTools: a suite for analysis and design of nucleic
Chernolovskaya, E. L., Vlassov, V. V., Pyshnyi, D. V.,
acid oligomers, Nucleic Acids Res., 36, W163-W169,
and Zenkova, M. A. (2020) Transport oligonucle-
doi: 10.1093/nar/gkn198.
otides - a novel system for intracellular delivery of an-
61.
Alinovskaya, L. I., Sedykh, S. E., Ivanisenko, N. V.,
tisense therapeutics, Molecules, 25, 3663, doi: 10.3390/
Soboleva, S. E., and Nevinsky, G. A. (2018) How
molecules25163663.
human serum albumin recognizes DNA and RNA,
51.
Pavlova, A. S., Dovydenko, I. S., Kupryushkin, M. S.,
Biol. Chem., 399, 347-360, doi: 10.1515/hsz-2017-0243.
Grigor’eva, A. E., Pyshnaya, I. A., and Pyshnyi, D. V.
62.
Bar-Ziv, R., and Libchaber, A. (2001) Effects of
(2020) Amphiphilic “like-a-brush” oligonucleotide con-
DNA sequence and structure on binding of RecA to
jugates with three dodecyl chains: self-assembly features
single-stranded DNA, Proc. Natl. Acad. Sci. USA, 98,
of novel scaffold compounds for nucleic acids delivery,
9068-9073, doi: 10.1073/pnas.15124289.
Nanomaterials, 10, 1948, doi: 10.3390/nano10101948.
63.
Fredrickson, D. S., and Gordon, R. S.
(1958)
52.
Fasano, M., Curry, S., Terreno, E., Galliano, M.,
The metabolism of albumin-bound C14-labeled
Fanali, G., Narciso, P., Notari, S., and Ascenzi, P.
unesterified fatty acids in normal human sub-
(2005) The extraordinary ligand binding properties
jects, J. Clin. Invest., 37, 1504-1515, doi: 10.1172/
of human serum albumin, IUBMB Life, 57, 787-796,
JCI103742.
doi: 10.1080/15216540500404093.
64.
Peters, Jr., T. (1995) All about Albumin. Biochemistry,
53.
Fanali, G., di Masi, A., Trezza, V., Marino, M.,
Genetics, and Medical Applications, Academic Press,
Fasano, M., and Ascenzi, P. (2012) Human serum
San Diego, doi: 10.1016/B978-0-12-552110-9.X5000-4.
albumin: from bench to bedside, Mol. Aspects Med.,
65.
Bhattacharya, A. A., Grüne, T., and Curry, S. (2000)
33, 209-290, doi: 10.1016/j.mam.2011.12.002.
Crystallographic analysis reveals common modes
54.
Knudsen Sand, K. M., Bern, M., Nilsen, J., Noordzij,
of binding of medium and long-chain fatty acids to
H. T., Sandlie, I., and Andersen, J. T. (2015) Un-
human serum albumin, J. Mol. Biol., 303, 721-732,
raveling the interaction between FcRn and albu-
doi: 10.1006/jmbi.2000.4158.
min: opportunities for design of albumin-based
66.
Curry, S. (2003) Plasma albumin as a fatty acid carrier,
therapeutics, Front. Immunol., 5, 682, doi: 10.3389/
in Advances in Molecular and Cell Biology, Lipobiology
fimmu.2014.00682.
(van der Vusse, G. J., eds) Elsevier, vol. 33, pp. 29-46.
55.
Kupryushkin, M. S., Nekrasov, M. D., Stetsenko,
67.
Fujiwara, S.-i., and Amisaki, T. (2008) Identification
D. A., and Pyshnyi, D. V. (2014) Efficient function-
of high affinity fatty acid binding sites on human
alization of oligonucleotides by new achiral non-
serum albumin by MM-PBSA method, Biophys. J.,
nucleosidic monomers, Org. Lett., 16, 2842-2845,
94, 95-103, doi: 10.1529/biophysj.107.111377.
doi: 10.1021/ol500668n.
68.
Van der Vusse, G. J. (2009) Albumin as fatty acid
56.
Dunn, D. B., and Hall, R. H. (1975) Purines, pyrim-
transporter, Drug Metab. Pharmacokinet., 24, 300-307,
idines, nucleosides and nucleotides: physical con-
doi: 10.2133/dmpk.24.300.
stants and spectral properties, in Handbook of Bio-
69.
Ascenzi, P., Bocedi, A., Notari, S., Fanali, G., Fesce, R.,
chemistry and Molecular Biology (Fasman, G. D., eds)
and Fasano, M. (2006) Allosteric modulation of drug
CRC Press, Cleveland, Vol. 1, pp. 65-215.
binding to human serum albumin, Mini. Rev. Med.
57.
Dobrynin, S., Kutseikin, S., Morozov, D., Krum-
Chem., 6, 483-489, doi: 10.2174/138955706776361448.
kacheva, O., Spitsyna, A., Gatilov, Y., Silnikov, V.,
70.
Agudelo, D., Bourassa, P., Bruneau, J., Bérubé, G.,
Angelovski, G., Bowman, M. K., Kirilyuk, I., and
Asselin, É., and Tajmir-Riahi, H.-A. (2012) Probing
Chubarov, A. (2020) Human serum albumin labelled
the binding sites of antibiotic drugs doxorubicin
with sterically-hindered nitroxides as potential
and N-(trifluoroacetyl) doxorubicin with human
MRI CONTRAST Agents, Molecules,
25,
1709,
and bovine serum albumins, PLoS One, 7, e43814,
doi: 10.3390/molecules25071709.
doi: 10.1371/journal.pone.0043814.
58.
Chubarov, A., Spitsyna, A., Krumkacheva, O.,
71.
Golianová, K., Havadej, S., Verebová, V., Uličný, J.,
Mitin, D., Suvorov, D., Tormyshev, V., Fedin, M.,
Holečková, B., and Staničová, J. (2021) Interaction
Bowman, M. K., and Bagryanskaya, E. (2021) Revers-
of conazole pesticides epoxiconazole and prothio-
ible dimerization of human serum albumin, Molecules,
conazole with human and bovine serum albumin
26, 108, doi: 10.3390/molecules26010108.
studied using spectroscopic methods and molecular
59.
Atmeh, R. F., Arafa, I. M., and Al-Khateeb, M.
modeling, Int. J. Mol. Sci., 22, 1925, doi: 10.3390/
(2007) Albumin aggregates: hydrodynamic shape
ijms22041925.
БИОХИМИЯ том 88 вып. 8 2023
1440
ПАВЛОВА и др.
72. Bisker, G., Ahn, J., Kruss, S., Ulissi, Z. W., Salem,
Вестник НГУ. Серия: Биология, клиническая меди-
D. P., and Strano, M. S. (2015) A mathematical
цина, 9, 109-117.
formulation and solution of the CoPhMoRe
77. Pyne, A. L. B., Noy, A., Main, K. H. S., Velasco-
inverse problem for helically wrapping polymer
Berrelleza, V., Piperakis, M. M., Mitchenall, L. A.,
corona phases on cylindrical substrates, J. Phys.
Cugliandolo, F. M., Beton, J. G., Stevenson, C. E. M.,
Chem. C, 119, 13876-13886, doi: 10.1021/acs.jpcc.
Hoogenboom, B. W., Bates, A. D., Maxwell, A., and
5b01705.
Harris, S. A. (2021) Base-pair resolution analysis of
73. Beckwitt, E., Kong, M., and Van Houten, B. (2018)
the effect of supercoiling on DNA flexibility and ma-
Studying protein-DNA interactions using atomic
jor groove recognition by triplex-forming oligonucle-
force microscopy, Semin. Cell Dev. Biol., 73, 220-230,
otides, Nat. Commun., 12, 1053, doi: 10.1038/s41467-
doi: 10.1016/j.semcdb.2017.06.028.
021-21243-y.
74. Pyshnaya, I. A., Pyshnyi, D. V., Lomzov, A. A.,
78. Zhang, W., and Tung, C.-H.
(2017) Sequence-
Zarytova, V. F., and Ivanova, E. M. (2004) The influ-
independent DNA nanogel as a potential drug carrier,
ence of the non-nucleotide insert on the hybridiza-
Macromol. Rapid Commun., 38, 1700366, doi: 10.1002/
tion properties of oligonucleotides, Nucleosides Nu-
marc.201700366.
cleotides Nucleic Acids, 23, 1065-1071, doi: 10.1081/
79. Lacroix, A., Edwardson, T. G. W., Hancock, M. A.,
NCN-200026073.
Dore, M. D., and Sleiman, H. F. (2017) Development
75. Виноградова О. A., Еремеева Е. В., Ломзов A. A.,
of DNA nanostructures for high-affinity binding
Пышная И. А., Пышный Д. В. (2009) Изогнутые
to human serum albumin, J. Am. Chem. Soc., 139,
дцДНК с заданными геометрическими характери-
7355-7362, doi: 10.1021/jacs.7b02917.
стиками на основе комплексов мостиковых оли-
80. Arabi, S. H., Aghelnejad, B., Schwieger, C., Meister, A.,
гонуклеотидов, Биоорг. химия, 35, 384-396.
Kerth, A., and Hinderberger, D. (2018) Serum albu-
76. Виноградова О. A., Щеглов Д. В., Латышев A. В.,
min hydrogels in broad pH and temperature ranges:
Пышный Д. В. (2011) Изучение структурной орга-
characterization of their self-assembled structures and
низации конкатемерных комплексов на основе
nanoscopic and macroscopic properties, Biomater.
нативных и модифицированных дцДНК-блоков,
Sci., 6, 478, doi: 10.1039/c7bm00820a.
COMPLEXES AND SUPRAMOLECULAR ASSOCIATES
OF DODECYL-CONTAINING OLIGONUCLEOTIDES
WITH SERUM ALBUMIN
A. S. Pavlova*, V. V. Ilyushchenko, M. S. Kupryushkin, T. D. Zharkov, E. S. Dyudeeva,
I. A. Bauer, A. S. Chubarov, D. V. Pyshnyi, and I. A. Pyshnaya*
Institute of Chemical Biology and Fundamental Medicine, Siberian Branch of Russian Academy of Sciences,
630090 Novosibirsk, Russia; e-mail: pyshnaya@niboch.nsc.ru, pavlova@niboch.nsc.ru
Serum albumin is currently in the focus of biomedical research as a promising platform for the creation of
multicomponent self-assembling systems due to the presence of several sites with a high binding affinity
of various compounds in its molecule, including lipophilic oligonucleotide conjugates. In this work,
we investigated the stoichiometry of the dodecyl-containing oligonucleotides binding to bovine and human
serum albumins via electrophoretic mobility shift assay. The results testify to the formation of albumin-oli-
gonucleotide complexes with a stoichiometry of about 1 : (1.25 ± 0.25) under physiological-like conditions.
Using atomic force microscopy, it was found that interaction of human serum albumin with a duplex of
complementary dodecyl-containing oligonucleotides resulted in associates of round shape with a diameter
of 165.5 ± 94.3 nm and 28.9 ± 16.9 nm in height, and with polydeoxyadenylic acid and dodecyl-containing
oligothymidylate also resulted in formation of supramolecular associates with a size of about 315.4 ± 70.9
and 188.3 ± 43.7 nm. The obtained data allow considering dodecyl-containing oligonucleotides and albu-
min as potential components for the development of methods to design self-assembled systems for solv-
ing problems of molecular biology, biomedicine, and the development of unique theranostics with targeted
action.
Keywords: serum albumin, dodecyl-containing oligonucleotides, protein-nucleic acids complexes, biopolymers self-
assembly, supramolecular associates, nucleic acids delivery
БИОХИМИЯ том 88 вып. 8 2023