БИОХИМИЯ, 2023, том 88, вып. 9, с. 1606 - 1619
УДК 577.24
ВЛИЯНИЕ РАСПОЛОЖЕНИЯ И ОРИЕНТАЦИИ ГЕНЕТИЧЕСКОГО
ТОКСИН-АНТИТОКСИНОВОГО ЭЛЕМЕНТА hok/sok НА УРОВЕНЬ
БИОСИНТЕЗА ФАРМАЦЕВТИЧЕСКИ ЗНАЧИМЫХ БЕЛКОВ
В БАКТЕРИАЛЬНОЙ СИСТЕМЕ ЭКСПРЕССИИ
© 2023 Ю.А. Ходак, Р.Р. Шайфутдинов, Д.С. Хасанов,
Н.А. Орлова, И.И. Воробьев*
ФИЦ Биотехнологии РАН, Институт биоинженерии,
117312 Москва, Россия; электронная почта: ptichman@gmail.com
Поступила в редакцию 20.04.2023
После доработки 15.08.2023
Принята к публикации 15.08.2023
Генетический токсин-антитоксиновый элемент hok/sok из плазмиды R1 Escherichia coli обеспечи-
вает сегрегационную стабильность плазмид. Бактериальные клетки, потерявшие все копии плаз-
миды, кодирующей короткоживущий антитоксин, гибнут под действием долгоживущего токсина.
Элемент hok/sok в составе векторных плазмид для бактериальной экспрессии может увеличивать
продуктивное время биосинтеза рекомбинантных белков, замедляя накопление в популяции
непродуцирующих клеток, лишенных целевой плазмиды. В настоящей работе были исследованы
различные варианты расположения и ориентации элемента hok/sok в составе стандартной плаз-
миды pET28a с индуцибельным промотором T7lac и геном устойчивости к канамицину. Было об-
наружено, что элемент hok/sok сохраняет функциональную активность вне зависимости от распо-
ложения на плазмиде и ориентации, бактериальные клетки сохраняли плазмиды с hok/sok после
4-х дней культивирования без антибиотика и теряли контрольную плазмиду без данного элемента.
На примере трех целевых белков - аспаргиназы E. coli тип II, гормона роста человека и нуклео-
протеина вируса SARS-CoV-2 было продемонстрировано, что для цитоплазматических целевых
белков максимальная продуктивность бактерий сохраняется только при расположении элемента
hok/sok на плазмиде выше промотора целевого гена. В случае периплазматической локализации
белка продуктивность бактерий уменьшается для всех вариантов расположения hok/sok при куль-
тивировании с антибиотиком, а при периодическом культивировании бактерий без антибиотика
продуктивность лучше сохраняется также при расположении элемента hok/sok выше промотора
целевого гена. Данный вариант векторной плазмиды pEHU позволяет увеличить биосинтез нерас-
творимого в цитоплазме бактерий гормона роста человека более, чем в 2 раза при культивирова-
нии бактерий без антибиотика, а также поддерживать биосинтез аспарагиназы при периодическом
культивировании без антибиотика в течение 4-х дней на уровне не менее 10 мг/литр. Разработан-
ный сегрегационно стабилизированный плазмидный вектор может быть использован для получе-
ния в клетках E. coli различных рекомбинантных белков без применения антибиотиков.
КЛЮЧЕВЫЕ СЛОВА: гетерологичная экспрессия рекомбинантных белков, системы токсин-антитоксин,
аспарагиназа, нуклеопротеин, соматотропин.
DOI: 10.31857/S0320972523090129, EDN: WUOWEN
ВВЕДЕНИЕ
обеспечивает бактериальной клетке значи-
тельное преимущество в скорости деления.
Обеспечение сегрегационной устойчиво-
Вследствие этого даже при времени индукции
сти плазмид в бактериальных системах экс-
промотора целевого гена 4-8 ч популяция бак-
прессии генов остается актуальной задачей
терий-продуцентов обычно замещается непро-
биофармацевтики. Удаление всех копий плаз-
дуцирующими клетками, что приводит к огра-
миды, кодирующей ген гетерологичного белка, ничению общей продуктивности системы [1].
Принятые сокращения: ИПТГ - изопропил-β-D-1-тиогалактопиранозид; ОРС - открытая рамка считывания;
ТА - токсин-антитоксин; ASN - L-аспарагиназа; HGH - гормон роста человека; NP - нуклеопротеин варианта
«Ухань» коронавируса SARS-CoV-2.
* Адресат для корреспонденции.
1606
ВЛИЯНИЕ ЭЛЕМЕНТА hok/sok НА ЭКСПРЕССИЮ БЕЛКОВ Е. coli
1607
Базовую защиту бактериальных клеток от по-
ратной химической реакции для пары DarT/
тери плазмиды обеспечивает ген устойчивости
DarG [9]; системы типа V - на селективном
к действию антибиотика, однако для плот-
разрушении мРНК токсического белка белко-
ных бактериальных культур концентрация
вым антитоксином, как в паре GhoT/GhoS [10].
селективного антибиотика в ростовой среде
В типе VI систем ТА, разновидности систем
постепенно падает под действием инактиви-
типа II, белковый антитоксин SocB является
рующих его ферментов и становится слишком
протеолитическим адаптером, обеспечиваю-
низкой [1]. Возможен второй уровень защи-
щим быстрый протеолиз токсина SocA кле-
ты бактериальных клеток от потери целевых
точной протеазой ClpXP [11]. Недавно был
плазмид - это введение в них природных или
выделен новый VII-тип систем, включающий
модифицированных пар генов токсин-анти-
антитоксины, ферментативно модифицирую-
токсин (ТА), обеспечивающих немедленную
щие токсины. В отличие от систем типа II, в
смерть бактериальной клетки, потерявшей все
данном случае модификации вносятся в про-
активные копии гена антитоксина.
цессе кратковременного, а не постоянного
Генетические элементы ТА присутствуют в
взаимодействия между токсином и антиток-
большом числе копий в геномах прокариот [2],
сином [12] В последнем описанном VIII-типе
плазмидах [3] и бактериофагах [4]; для неко-
систем в качестве токсина выступают малые
торых свободноживущих бактерий описывают
РНК, блокирующиеся антисмысловыми РНК,
более
40 таких генетических элементов [2].
которые кодируются в том же локусе и высту-
Данные системы классифицируют на основа-
пают в качестве антитоксина [13].
нии способа нейтрализации токсина антиток-
Первые системы ТА были открыты при
сином. Традиционно их разделяли на 6 типов,
изучении природных плазмид E. coli и, по ис-
однако в последние годы было выделено еще
ходным предположениям исследователей, их
2 новых типа, таким образом, их общее число
биологической функцией было обеспечение
достигло 8 [5]. Наиболее изученным и рас-
сегрегационной стабильности таких плазмид.
пространенным является тип I, состоящий из
В дальнейшем было обнаружено, что систе-
гена стабильного токсичного белка и малоста-
мы ТА вовлечены в обеспечение персистен-
бильной антисмысловой РНК, блокирующей
ции бактерий [14], ответа на факторы стресса
трансляцию мРНК токсина [2]. Исследуемый
и защиту от бактериофагов [15]. Несмотря на
в настоящей работе локус hok/sok (host killing/
крайне высокое разнообразие генетических
suppressor of killing) относится к первому типу
оперонов систем ТА, их известные токсины
систем ТА, механизм его действия будет по-
атакуют очень небольшой набор биохимичес-
дробно описан ниже.
ких процессов бактериальной клетки - транс-
Тип II систем ТА представляет собой пару
ляцию мРНК, репликацию хромосомной ДНК,
генов, кодирующих стабильный белковый ток-
рост клеточной стенки и поддержание мем-
син и нестабильный белковый антитоксин.
бранного потенциала.
Типичный пример системы ТА типа II - ло-
Рассматриваемый в настоящей работе ге-
кус ccd, содержащий ген токсичного для бак-
нетический локус hok/sok относится к систе-
териальной клетки ингибитора ДНК-гиразы
мам ТА типа I. Он был впервые обнаружен в
CcdB и ген малостабильного ингибитора CcdA,
плазмиде R1 E. coli [16], исходно обозначен как
подверженного быстрому распаду под действи-
ParB и описан как оперон двух цистронов hok
ем протеазы Lon [6]. При потере всех копий
и sok. Матричная РНК гена hok кодирует ток-
плазмиды, кодирующей локус ccd, концентра-
сичный полипептид размером 52 аминокисло-
ция белка CcdA быстро уменьшается и сво-
ты, который нарушает поляризацию внутрен-
бодный белок CcdB блокирует активность
ней мембраны бактерий за счет формирования
ДНК-гиразы E. coli в достаточной мере для
пор, что приводит к стазису и/или гибели бак-
прекращения клеточного деления и индукции
териальных клеток [17]. Ген sok обладает силь-
SOS-ответа [7].
ным промотором и кодирует антисмысловую
Остальные типы систем ТА распростране-
РНК, связывающуюся с участком мРНК hok и
ны значительно реже и характеризуются дру-
обеспечивающую быструю деградацию мРНК
гими способами взаимодействия токсина и
hok под действием РНКазы III. Несмотря на
антитоксина. Системы типа III основаны на
кажущуюся очевидность механизма действия
прямом взаимодействии антитоксической РНК
генов токсина и антитоксина в паре hok/sok, их
с белковой молекулой токсина [8]; системы
фактическое взаимодействие является доста-
типа IV - на блокировании функции токсина
точно сложным и многоступенчатым процес-
без прямого белок-белкового взаимодействия,
сом, подробно описанным в обзорной рабо-
например, путем катализа антитоксином об-
те Gerdes [18].
БИОХИМИЯ том 88 вып. 9 2023
1608
ХОДАК и др.
Вкратце, большая часть мРНК hok обра-
тери клетками E. coli плазмид в неселективных
зуется в форме трансляционно неактивного
условиях. В случае плазмиды с индуцибель-
продукта увеличенной длины, который может
ным промотором T7 и селективным маркером
быть описан как продукт гена mok, перекры-
устойчивости к действию тетрациклина было
вающего ген hok. В данной мРНК с размером
заявлено сохранение у 100% клеток устойчи-
~400 п.о. 5′- и 3′-концевые области образуют
вости к антибиотику после 20 ч индукции це-
дуплекс, блокирующий ее связывание с рибо-
левого гена, при этих условиях контрольную
сомами и с антисмысловой РНК Sok. Ста-
плазмиду сохраняли только 17% клеток [26],
бильная и неактивная форма мРНК (400 п.о.)
уровень биосинтеза целевого белка - пролак-
медленно превращается в активную форму раз-
тина человека, был увеличен в 2,3 раза, хотя и
мером 360 п.о. в результате удаления 3′-фраг-
оставался достаточно низким.
мента под действием РНКазы II. Активная
В случае токсичной для бактериальных
мРНК hok становится доступной для связы-
клеток фосфолипазы Д из Streptomyces anti-
вания антисмысловой РНК Sok, сайт связы-
bioticus введение элемента hok/sok в состав
вания которой расположен вблизи первого
плазмид с промотором T7lac и геном устойчи-
кодона AUG [8]. Возникающий гетеродуплекс
вости к ампициллину привело к исчезновению
РНК быстро гидролизуется РНКазой III [19, 20].
клеток, терявших плазмиду при ведении куль-
Если в клетке нет достаточных количеств РНК
туры до индукции и позволило удвоить общий
Sok, то стартует трансляция мРНК hok, выра-
уровень биосинтеза ферментативно активной
батывается белок Hok, который формирует
фосфолипазы, однако быстрое исчезновение
мембранные поры, что приводит к переходу
клеток с плазмидой после начала индукции
клеток в персистирующее состояние, а при
было зафиксировано как для контрольных
дальнейшем росте концентрации Hok возни-
плазмид, так и для плазмид с локусом hok/sok
кает состояние стазиса и клеточной смерти.
и другими локусами ТА [27].
Предположительно, уровень антисмысло-
Генетический элемент hok/sok увеличи-
вой РНК Sok в клетке определяется не только
вает устойчивость плазмид не только в E. coli,
активностью копий ее гена, но и механизмом
но и в иных видах бактерий, в частности, в
отрицательной обратной связи, включающим
Salmonella enterica (серовар Typhi) [28] и Vib-
мембрано-ассоциированную РНКазу E, актив-
rio cholerae [29]. В случае сальмонеллы при
ность которой снижается при деполяризации
24-часовой культивировании в селективной
мембраны [18], таким образом, антитоксиче-
среде плазмида с локусом hok/sok сохранялась
ское действие Sok сохраняется даже при недо-
у 48% клеток, контрольная плазмида - у 13%
статочном уровне транскрипции гена, однако
клеток. При культивировании холерного ви-
при потере всех копий гена sok данный меха-
бриона с плазмидами, содержащими локус hok/
низм уже не может действовать.
sok, в течение 4-х дней все клетки сохраняли
В геноме базового лабораторного штам-
способность экспрессировать ген люцифера-
ма K12 E. coli выявляется до 5 локусов типа hok/
зы, кодируемый плазмидами, однако в данном
sok [21], при этом 4 из них, по мнению груп-
исследовании не использовались контрольные
пы Gerdes, инактивированы мутациями вблизи
плазмиды, не содержащие локуса hok/sok.
открытой рамки считывания (ОРС) hok [22],
Несмотря на то что локус hok/sok изучен
а единственный активный локус hokB/sokB,
значительно лучше, чем другие локусы ТА, и
по-видимому, участвует в регуляции входа и
применялся для стабилизации плазмид раз-
выхода клеток в состояние персистенции за
личных типов, в литературе отсутствуют дан-
счет процессинга белка HokB протеазами пе-
ные о его фактическом влиянии на свойства
риплазмы [23]. Также локус hokB/ sokB опре-
широко распространенных в биофармацев-
деляет, наряду с другими локусами ТА, повы-
тике экспрессионных плазмид с индуцибель-
шение устойчивости бактериальных клеток к
ным сильным промотором T7lac и селекци-
стрессовым условиям [24] путем увеличения
онным маркером устойчивости к канамицину.
продолжительности лаг-фазы и последующего
Мы предположили, что инсерция генетиче-
увеличения скорости роста на экспоненци-
ского локуса hok/sok в плазмиды данного типа
альной фазе.
может в достаточной мере увеличить их сегре-
Увеличение сегрегационной стабильности
гационную устойчивость в процессе индук-
искусственных, т.е. обычных лабораторных
ции промотора целевого гена и, как следствие,
плазмид при включении в их состав локуса hok/
значимо увеличить продуктивное время ин-
sok было продемонстрировано во множестве
дукции, т.е. общую продуктивность бактери-
работ. В частности, в работе Pecota et al. [25]
альной системы экспрессии гетерологичных
наблюдали 9-кратное замедление процесса по-
белков.
БИОХИМИЯ том 88 вып. 9 2023
ВЛИЯНИЕ ЭЛЕМЕНТА hok/sok НА ЭКСПРЕССИЮ БЕЛКОВ Е. coli
1609
Последовательности праймерных олигонуклеотидов
Название
Сайт
Нуклеотидная последовательность
праймера
рестрикции
AS-HOS-F5
ACACGTGGATCCAACAAACTCCGGGAGGCAGCGTGATGCGG
PspCI
AS-HOS-R5
TCACGTGCATGCACAACATCAGCAAGGAGAAAGGGGCTACCGG
PspCI
AD-Asp-NcoF
ACCATGGAGTTTTTCAAAAAGACGGCACTT
NcoI
AD-Asp-HindR
TAAGCTTCTATTAGTACTGATTGAAGATCTGCTGG
HindIII
M13-dir
GTTGTAAACGACGGCCAGTG
T7prom
TAATACGACTCACTATAGGG
T7t
GCTAGTTATTGCTCAGCGG
МАТЕРИАЛЫ И МЕТОДЫ
(«Евроген»), секвенировали со стандартных
праймеров T7prom и M13rev (таблица).
Получение векторных плазмид pEHU,
ОРС нуклеопротеина (NP) варианта «Ухань»
pEHD, pEHR. Участок hok/sok природной
коронавируса SARS-CoV-2 и экспресссионные
плазмиды R1 E. coli (GenBank X05813), фланки-
конструкции на ее основе получали, как опи-
рованный сайтами узнавания эндонуклеаз ре-
сано в статье Kolesov et al. [30].
стрикции PspCI, BamHI и SphI синтезировали
Оптимизированную для экспрессии в
в «Synbio Technologies» (Китай) и клонировали
E. coli последовательность ОРС гормона роста
в плазмиду pUC57. Плазмидy pUC57-HS сек-
человека (HGH) с метионином [-1] и двумя
венировали со стандартных праймеров M13,
стоп-кодонами синтезировали в ООО «Кло-
после чего элемент клонировали в плазми-
нинг Фасилити» (Россия), клонировали в
ду pET28а (+) («Novagen», США) двумя разны-
pAL2-Т, секвенировали со стандартных прай-
ми способами. Для получения вектора pEHU,
меров к вектору.
где элемент hok/sok расположен до целевого
Экспрессионные векторы pET28a, pEHU,
гена, реципиентную плазмиду pET28а рас-
pEHD, pEHR рестрицировали по сайтам NcoI
щепляли эндонуклеазами BglII и SphI и лиги-
и HindIII и лигировали с фрагментами, содер-
ровали с элементом hok/sok, расщепленным
жащими ОРС целевых белков, вырезанными
BamHI и SphI. Для получения векторов pEHD
по сайтам NcoI и HindIII в случае аспараги-
и pEHR, где элемент hok/sok расположен по-
назы, или PciI и HindIII - в случае нуклеопро-
сле целевого гена, реципиентную плазмиду
теина и гормона роста человека.
pET28а расщепляли эндонуклеазой PsiI и ли-
Полученные плазмиды pET28-ASN,
гировали с элементом hok/sok, расщепленным
pEHU-ASN, pEHD-ASN, pEHR-ASN;
эндонуклеазой PspCI. Методом ПЦР опре-
pET28-NP, pEHU-NP, pEHD-NP, pEHR-
деляли ориентацию вставки с использова-
NP; pET28-HGH, pEHU-HGH, pEHD-
нием праймеров T7prom и AS-HOS-F5 или
HGH, pEHR-HGH секвенировали с прайме-
AS-HOS-R5 (таблица). Плазмида pEHD содер-
ров T7prom и T7t и использовали для трансфор-
жала вставку в прямой ориентации, плазмида
мации клеток BL21[DE3] («Евроген»); транс-
pEHR - в обратной ориентации.
фицированные клетки далее были использо-
Получение экспрессионных плазмид. ОРС
ваны в опытах по биосинтезу целевых белков.
природного гена L-аспарагиназы (ASN) E. сoli
Плазмидную ДНК выделяли набором Plas-
получали методом ПЦР с адапторных прайме-
mid Miniprep («Евроген»), очистку рестрикци-
ров AD-Asp-NcoF и AD-Asp-HindR (таблица),
онных фрагментов и продуктов ПЦР из агароз-
содержащих сайты узнавания эндонуклеаз
ного геля проводили набором Cleanup Standard
рестрикции для последующего клонирования,
(«Евроген»). Использовали эндонуклеазы ре-
используя геномную ДНК E. coli штамма
стрикции производства ООО
«СибЭнзайм»
BL21[DE3] («Novagen») в качестве матрицы и
(Россия) и Т4-ДНК-лигазу («Евроген»).
набор и Encyclo Plus PCR kit («Евроген», Рос-
Секвенирование ДНК проводили в Меж-
сия). Продукт ПЦР выделяли из 1%-ного ага-
институтском Центре коллективного пользо-
розного геля, клонировали в Т-вектор pAL2-T
вания «ГЕНОМ» ИМБ РАН.
БИОХИМИЯ том 88 вып. 9 2023
1610
ХОДАК и др.
Культивирование штаммов-продуцентов и по-
50 мМ Tris-HCl (pH 7,4), 10 мкг/мл лизоцима и
лучение образцов для ДСН-ПААГ. Выращива-
0,1% (v/v) Triton X-100, и инкубировали 30 мин
ние бактерий вели в среде либо 2х Yeast extract
на льду. Разрушали геномную ДНК клеток уль-
and Tripton (2х YT) состава (из расчета на литр):
тразвуковым диспергатором («Misonix», США)
казеиновый пептон - 16 г, дрожжевой экс-
пульсами по 10 с до исчезновения повышен-
тракт - 10 г, NaCl - 5 г, pH среды - 7,0 ± 0,2;
ной вязкости суспензии. Отделяли осадки цен-
либо в среде Terrific broth (TB) состава (из рас-
трифугированием (10 мин при 12 200 g). Фрак-
чета на литр): триптон - 12 г, дрожжевой экс-
цию нерастворимых белков (телец включе-
тракт - 24 г, гидрофосфат калия - 12,54 г, ди-
ния) повторно ресуспендировали в растворе
гидрофосфат калия - 2,31 г, pH среды - 7,0 ± 0,2.
(1/10 объема исходной аликвоты), содержа-
Посевную культуру получали в среде 2x YT,
щем 50 мМ Tris-HCl (pH 7,4) и 500 мМ NaCl,
содержащей 30 мкг/мл канамицина и 1% глю-
и отделяли супернатант центрифугированием
козы, в течение 14-18 ч при 37 °С и 160 об./мин,
(10 мин при 12 200 g). Проводили ДСН-ПААГ-
если не указано иное. Инокулировали полу-
анализ белковых фракций
(12%-ный ДСН-
ченным посевным материалом в соотноше-
ПААГ), гель окрашивали коллоидным рас-
нии 1 : 1000 250 мл сред 2x YT или Terrific broth,
твором Кумасси синий («Fermentas», Литва),
дополненных 30 мкг/мл канамицина и 0,1% глю-
сканировали при помощи планшетного ска-
козы, растили культуру на качалке в тече-
нера в просвечивающем режиме. Получаемое
ние 3,5 ч при 37 °С, добавляли в качестве ин-
изображение геля с глубиной цвета 16 бит/ка-
дуктора ИПТГ до конечной концентрации
нал анализировали с помощью программы
0,2-1 мМ и растили в течение 3-30 ч при тем-
TotalLab, в качестве калибратора интенсивно-
пературе 30-37 °C.
сти окраски полос использовали бычий сыво-
Культивирование штаммов с плазмидами,
роточный альбумин («Диа-М»), по 0,25-2 мкг
кодирующими ASN, HGH и NP, для электро-
на дорожку.
форетического анализа проводили следующим
Определение доли бактериальных клеток,
образом: стартовую культуру получали в среде
способных к колониеобразованию в присутствии
2x YT, содержащей 30 мкг/мл канамицина и
канамицина. Готовили посевную культуру штам-
1% глюкозы, в течение 14-18 ч при 37 °С. Ино-
мов, как указано выше. Вели выращивание
кулировали полученным посевным материалом
культур без канамицина - переносили 1 мл
в соотношении 1 : 50 5 мл среды 2x YT, содер-
посевной культуры в стерильную микропро-
жащей 0,1% глюкозы с 30 мкг/мл канамицина;
бирку, и отделяли среду с остатками антибио-
растили культуру на качалке в течение 1 ч при
тика центрифугированием, ресуспендировали
37 °С, затем добавляли ИПТГ до конечной кон-
осадок клеток в 1 мл 2x YT. Инокулировали
центрации 1 мМ и растили в течение 18 ч при
полученной суспензией в соотношении 1 : 100
температуре 30 °C (NP) и 37 °C (HGH и ASN).
5 мл 2x YT, содержащей 0,1% глюкозы, но без
Осадок биомассы отделяли центрифугирова-
канамицина, растили культуру 2 ч, после чего
нием (30 c 12 200 g), ресуспендировали в буфере
добавляли ИПТГ до 0,2-2 мМ и продолжали
(1/5 объема исходной аликвоты) для образцов
культивирование в течение 8-24 ч. Отбирали
(25 мМ Tris-HCl (pH 6,8), 50 мМ ДТТ, 1% ДСН,
аликвоты тестируемых культур, при помощи
5% глицерина и 0,05% бромфенолового синего),
стерильной среды 2x YT делали последователь-
прогревали 5 мин при 95 °C. Проводили элек-
ные 100-кратные разведения. Из разведений
трофоретический анализ (12%-ный ДСН-ПААГ)
культур с кратностью 10 тыс и 1 млн высевали
белковых фракций, гель окрашивали колло-
по 250 мкл бактериальных суспензий на чашки
идным раствором Кумасси синий («Диа-М»,
Петри с агаризованной средой 2х YT, содержа-
Россия), сканировали при помощи планшет-
щей 1% глюкозы, с добавлением и без добав-
ного сканера в просвечивающем режиме Epson
ления 30 мкг/мл канамицина. Инкубировали
Perfection V700 Photo («Epson», Япония). По-
при 37 °С 14-18 ч и подсчитывали число колоний.
лучаемое изображение геля с глубиной цвета
Измерение ферментативной активности ас-
16 бит/канал анализировали с помощью про-
парагиназы. Отделяли бактерии от культураль-
граммы TotalLab («Nonlinear Dynamics», Вели-
ной среды и проводили их лизис с Triton X-100
кобритания).
и лизоцимом, как указано выше. Для измере-
Анализировали распределение целевого
ния активности использовали аликвоты сус-
белка по фракциям растворимых и нерас-
пензии лизированных бактерий, типичное
творимых белков. Для этого отделяли оса-
разведение - 100 раз. Исследуемые образцы
док биомассы центрифугированием, бакте-
(≤100 мкл), содержащие не более 0,3 МЕ ас-
рии ресуспендировали в растворе для лизиса
парагиназы, смешивали с
20 мкл раствора
(1/10 объема исходной аликвоты), содержащем
Tris-HCl (pH 8,5) до конечной концентрации
БИОХИМИЯ том 88 вып. 9 2023
ВЛИЯНИЕ ЭЛЕМЕНТА hok/sok НА ЭКСПРЕССИЮ БЕЛКОВ Е. coli
1611
по Tris - 50 мМ, добавляли 17 мкл 100 мМ L-ас-
транскрипции не создавало антисмысловую
парагина и инкубировали ровно 10 мин при
мРНК для целевого гена. В случае плазмид
+37 °С. Аналогично готовили раствор отри-
pEHD и pEHR локус hok/sok располагался
цательного контрольного образца, в который
downstream от промотора T7lac, после терми-
вместо лизата добавляли 100 мкл буферного
натора транскрипции целевого гена, т.е. уров-
раствора для лизиса, не содержащего аспара-
ни мРНК генов, кодирующих Hok и Sok, могли
гиназной активности, также проводили его
изменяться при неполной терминации тран-
инкубацию (10 мин при +37 °C). Активность
скрипции как целевого гена c индуцибельным
аспарагиназы в лизате клеток, отобранных до
промотором, так и гена устойчивости к дей-
внесения индуктора, составляла менее 1% от
ствию канамицина (KanR) с конститутивным
типичной активности аспарагиназы в индуци-
промотором. Плазмиды pEHD и pEHR разли-
рованных клетках.
чались ориентацией локуса hok/sok - в плазми-
Реакцию гидролиза останавливали добав-
де pEHD неполная терминация транскрипции
лением 100 мкл 50%-ного раствора трихлор-
целевого гена могла приводить к возрастанию
уксусной кислоты в воде. При появлении
уровня антитоксической РНК Sok, а неполная
видимого осадка его отделяли центрифугиро-
терминация транскрипции гена KanR - к воз-
ванием (10 мин, 14 100 g). Супернатант полно-
растанию уровня мРНК hok. В случае плазми-
стью переносили в новую пробирку.
ды pEHR влияние неполной терминации было
В пробирку калибратора вносили 137 мкл
обратным - промотор целевого гена мог уве-
раствора 0,5 мМ сульфата аммония (1 мМ ионов
личивать уровень мРНК токсина.
аммония), затем вносили
100 мкл раство-
Таким образом, в плазмиде pEHU уровни
ра ТХУ. Во все пробирки вносили по 650 мкл
транскрипции hok и sok не должны были изме-
разведенного реактива Несслера, полученно-
няться при индукции промотора основного
го смешиванием
550 мкл очищенной воды
гена; в плазмиде pEHD возможно возрастание
(«Milli-Q», Германия) и 100 мкл готового реак-
уровня транскрипции sok при индукции целе-
тива Несслера («Уральский завод химической
вого гена, а в плазмиде pEHR - возрастание
продукции», Россия). Растворы перемешивали
уровня транскрипции hok, что потенциально
на вортексе и выдерживали ~10 мин при ком-
может приводить к токсическому поражению
натной температуре. Измеряли поглощение
клеток при индукции целевого гена.
при длине волны 480 нм при помощи спек-
Для сравнения фактической сегрегаци-
трофотометра, рассчитывали концентрацию
онной стабильности полученных плазмид и
ионов аммония по поглощению калибровоч-
влияния их структуры на уровень экспрессии
ного образца (0,5 мМ сульфата аммония).
генов целевых белков в них клонировали ОРС
Статистическая обработка результатов. Ста-
трех типичных рекомбинантных белков - при-
тистическую обработку производили в про-
родную ОРС аспарагиназы E. coli типа II, как
грамме Microsoft Excel при помощи модуля
нетоксичного для E. coli растворимого белка с
Data Analysis. Первичные данные и статистиче-
периплазматической локализацией; кодон-оп-
ские расчеты приведены в Приложениях 1-6.
тимизированную ОРС гормона роста человека,
как малотоксичного для E. coli нерастворимого
в цитоплазме белка; а также кодон-оптимизи-
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
рованную ОРС нуклеопротеина коронавируса
SARS-CoV-2, как пример умеренно токсично-
Вследствие того, что в плазмидах типа pET
го для клеток преимущественно растворимого
действуют не менее трех достаточно сильных
в цитоплазме белка. Электрофореграммы, де-
промоторов - T7lac, lac, KanR, мы предпо-
монстрирующие распределение указанных бел-
ложили, что уровни транскрипции смысловой
ков по фракциям, приведены на рис. 1, б.
и антисмысловой РНК Hok (Mok) и Sok могут
Для плазмид с ОРС аспарагиназы была
быть существенно изменены из-за активности
проверена сегрегационная стабильность при пе-
близлежащих сильных промоторов. Для про-
риодическом культивировании в жидкой среде
верки влияния позиционного и ориентацион-
без антибиотика и без индуктора экспрессии
ного эффектов для генетического локуса hok/
целевого гена. Для всех вариантов расположе-
sok были получены 3 плазмиды - pEHU, pEHD
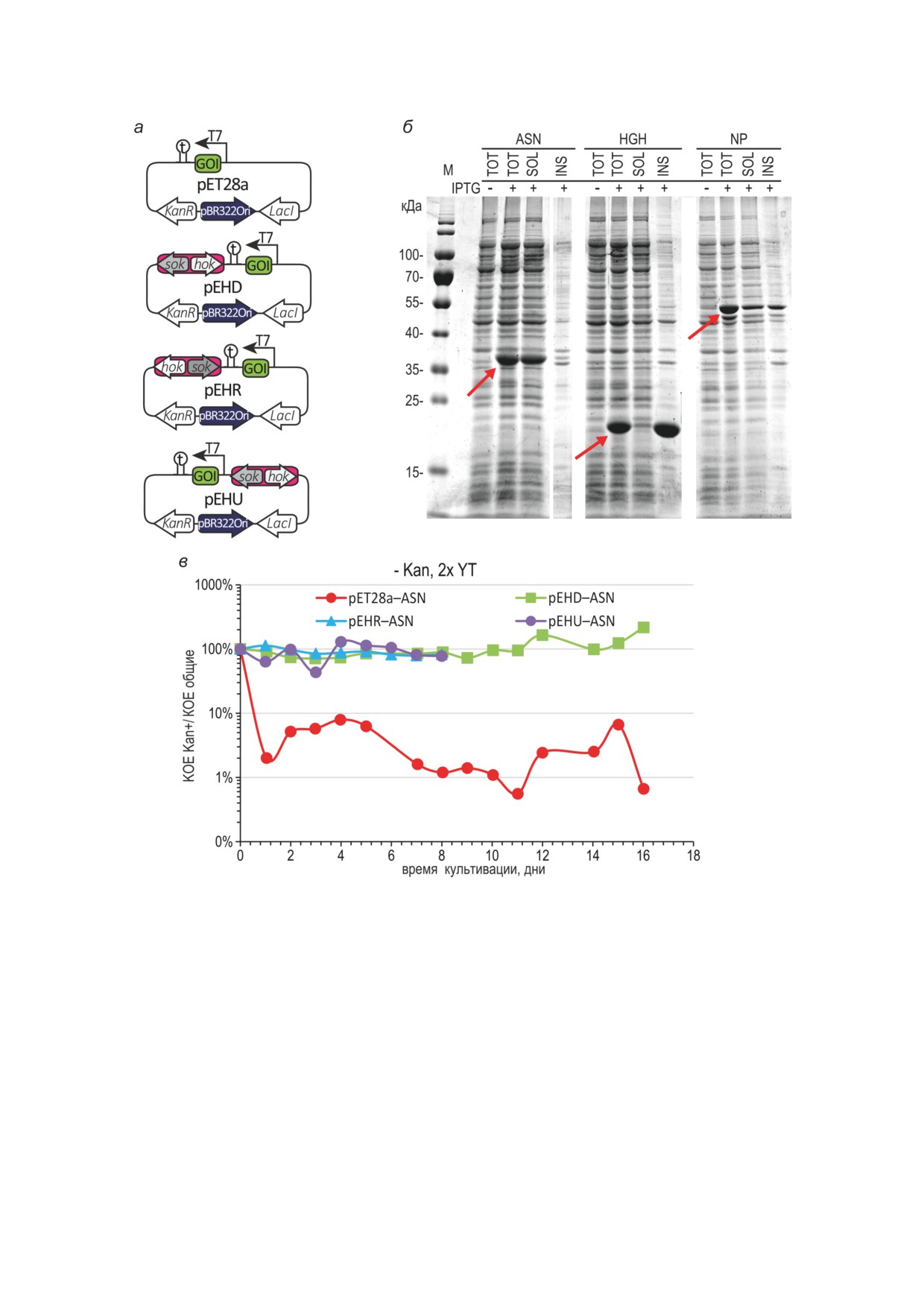
ния hok/sok (конструкции pEHD-ASN, pEHR-
и pEHR (рис. 1, а). В случае плазмиды pEHU
ASN и pEHU-ASN) плазмиды сохранялись в
локус hok/sok был помещен upstream от силь-
течение как минимум 8 дней, а доля клеток,
ного индуцибельного промотора T7lac и ори-
сохранивших контрольную плазмиду pET28a-
ентирован таким образом, чтобы продолже-
ASN, составила менее 10% через 1 день куль-
ние транскрипции гена hok за его терминатор
тивирования, т.е. приблизительно через 20 ге-
БИОХИМИЯ том 88 вып. 9 2023
1612
ХОДАК и др.
Рис. 1. Схемы вариантов сегрегационно-стабилизированных векторных плазмид, электрофореграммы белковых
фракций штаммов-продуцентов, динамика элиминации плазмид. а - Схема экспрессионных векторных плазмид,
обозначения: T7 - промотор целевого гена; GOI - область ОРС целевого белка; t - терминатор транскрипции
целевого гена; KanR - ген устойчивости к канамицину NPT II/Neo; pBR322Ori - вариант области начала репли-
кации плазмид ColE1; LacI - ген белка-репрессора LacI; hok и sok - гены локуса hok/sok. Стрелками обозначены
направления транскрипции генов. б - Электрофореграмма тотального белка (TOT), фракций растворимых (SOL)
и нерастворимых (INS) белков клеток штаммов BL21[DE3]/рET28a-ASN, BL21[DE3]/рET28a-HGH, BL21[DE3]/
pET28a-NP при индукции 1 мМ ИПТГ 3 ч, +30 °C. Обозначения: М - маркер молекулярных масс. Молекуляр-
ные массы полос маркера указаны в кДа. Положения целевых белков указаны стрелками. в - Динамика элими-
нации плазмид при периодическом культивировании штаммов в среде 2x YT без канамицина, без ИПТГ, +37 °С;
кратность пересева - 1000×; пересев 1 раз в 24 ч; в дни 6 и 13 пересев для штаммов BL21[DE3]/pET28a-ASN
и BL21[DE3]/pEHD-ASN проводился, но измерение числа КОЕ не проводилось. Не менее 25 колоний для чашек
без канамицина. Для всех дней, кроме дня 0, по критерию Пирсона (хи-квадрат) доля канамицин-устойчивых КОЕ
для плазмиды pET28a-ASN достоверно отличается от долей канамицин-устойчивых KOE для остальных плазмид;
P < 0,001
нераций клеток (рис. 1, в). При индукции экс-
скорость накопления целевого белка была сход-
прессии гена аспарагиназы в присутствии анти-
ной для всех вариантов плазмиды (рис 2, а),
биотика канамицина при помощи 1 мМ ИПТГ
таким образом, в этих условиях генетический
БИОХИМИЯ том 88 вып. 9 2023
ВЛИЯНИЕ ЭЛЕМЕНТА hok/sok НА ЭКСПРЕССИЮ БЕЛКОВ Е. coli
1613
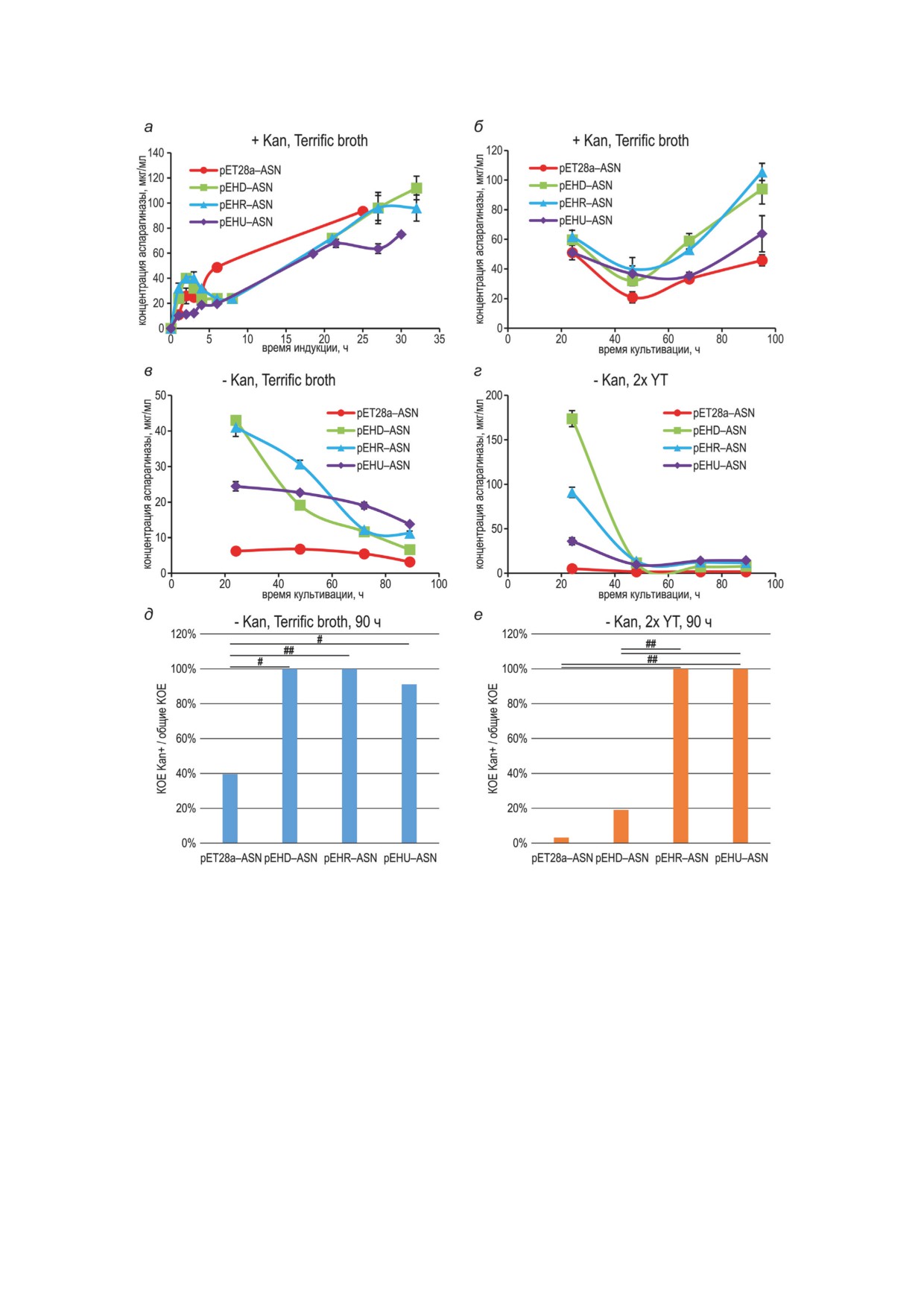
Рис. 2. Динамика накопления аспарагиназы и элиминации плазмид при культивировании штаммов в различных
условиях. Концентрацию аспарагиназы измеряли, как ее ферментативную активность в лизате клеток, условия
культивирования обозначены над соответствующими панелями. а - Культивирование без пересевов. б-г - Пере-
севы с кратностью 1 × 1000 каждые 24 ч. Планки погрешностей соответствуют стандартным отклонениям, n = 2,
биологические повторы. д и е - Определение КОЕ для культур, соответствующих точкам 90 ч на панелях в и г.
В случае, когда для одного и того же образца количество колоний на чашках с антибиотиком превышало таковое
на чашках без антибиотика, за 100% принимали отношение числа колоний на чашках с антибиотиком к числу
колоний на чашках без антибиотика. Не менее 65 колоний на чашках без канамицина. Статистически значимые
различия в долях канамицин-устойчивых колоний по критерию Пирсона (хи-квадрат) обозначены горизонталь-
ными чертами - # P < 0,05; ## P < 0,001
элемент hok/sok не влиял существенным обра-
сутствии 1 мМ ИПТГ и канамицина (рис. 2, б).
зом на продуктивность клеток. Поскольку про-
Было обнаружено, что концентрация аспараги-
мышленное культивирование продуцентов ре-
назы падает на втором цикле пересева, однако
комбинантных белков часто ведется в фермен-
затем возрастает на следующих двух пассажах,
терах в течение нескольких суток с подпиткой,
этот эффект был более выражен для плазмид
мы оценили изменение продуктивности клеток
pEHD-ASN и pEHR-ASN, содержащих hok/
при периодическом культивировании в при-
sok-элемент downstream от целевого гена.
БИОХИМИЯ том 88 вып. 9 2023
1614
ХОДАК и др.
При периодическом культивировании бак-
вызывалось не мутациями в плазмидах, а из-
терий без канамицина, но с 1 мМ ИПТГ, в
менениями в бактериальных клетках. Сегре-
буферизованной среде Terrific broth наблюдали
гационная стабилизация плазмид, в случае
практически полное прекращение накопления
аспарагиназы, существенно увеличила время
аспарагиназы в случае контрольной плазмиды
накопления целевого белка, однако не позволи-
pET28a-ASN и постепенное падение уровня
ла получить штаммы-продуценты, постоянно
аспарагиназы для остальных плазмид, наи-
накапливающие целевой белок при культиви-
менее выраженное для плазмиды pEHU-ASN
ровании без канамицина вследствие возникно-
(рис. 2, в). Такой режим культивирования со-
вения изменений в бактериях-хозяевах.
ответствует текущей практике выращивания
Влияние расположения и ориентации эле-
продуцентов без антибиотиков. Прекращение
мента hok/sok на уровень биосинтеза целевых
накопления аспарагиназы в случае плазмиды
белков было также изучено для локализован-
pET28a-ASN могло быть объяснено ее поте-
ных в цитоплазме бактерий белков HGH и NP,
рей у большей части клеток (рис. 2, д), в то же
накапливающихся только в нерастворимой
время все 3 варианта плазмид с элементом hok/
форме (HGH) или в частично растворимой
sok полностью сохранялись в клетках после
форме (NP). При выращивании культур в сре-
4-х дней культивирования, а продукция целе-
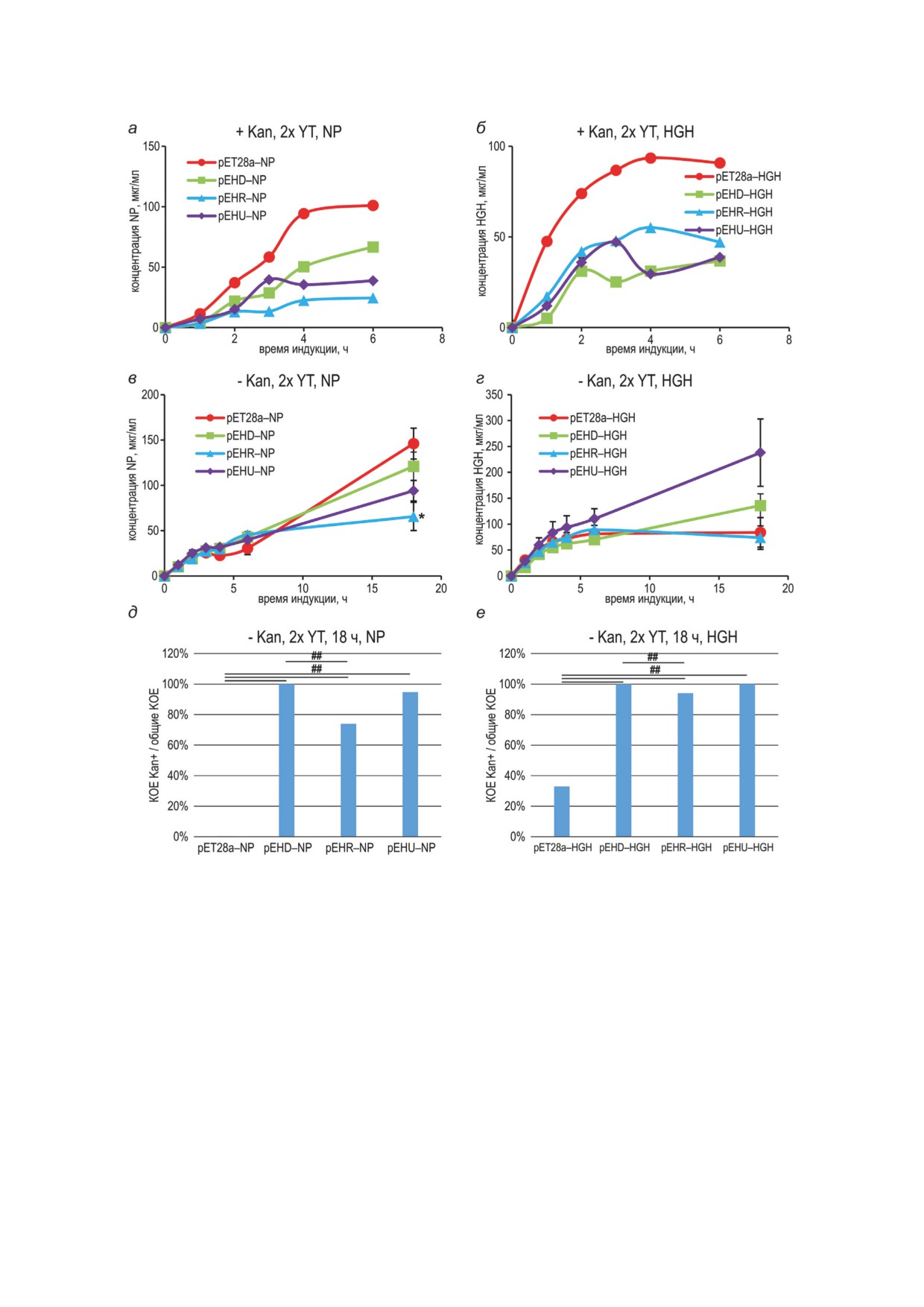
де 2x YT с канамицином и 1 мМ ИПТГ уровень
вого белка многократно падала. Данный эф-
биосинтеза NP и HGH снижался в 2-4 раза
фект
- прекращение накопления целевого
для всех вариантов плазмид с элементом hok/
белка при сохранении плазмид в клетках - был
sok (рис. 3, а и б). При культивировании штам-
еще сильнее выражен при культивировании в
мов без канамицина при индукции экспрессии
среде 2x YT, не обладающей буферной емко-
1 мМ ИПТГ для NP общая продуктивность оста-
стью (рис. 2, г и е). Уже после второго пассажа
валась максимальной для контрольной плаз-
уровень продукции аспарагиназы многократ-
миды pET28a-NP (рис. 3, в), а в случае HGH
но падал для всех вариантов плазмид, а после
общая продуктивность была значительно выше
4-х пассажей наблюдали практически полную
для плазмиды pEHU-HGH, чем для контроль-
потерю контрольной плазмиды pET28a-ASN
ной плазмиды и других вариантов расположе-
клетками, сохранение плазмиды pEHD-ASN -
ния элемента hok/sok (рис 3, г). Все варианты
у 20% клеток и полное сохранение плазмид
плазмид с hok/sok сохранялись после 18 ч ин-
pEHR-ASN и pEHU-ASN - у всех клеток.
дукции, контрольная плазмида сохранялась
Таким образом, при продолжительном культи-
только у 33% клеток для HGH и менее 1% -
вировании бактерий с 1 мМ ИПТГ без канами-
для NP (рис. 3, д и е).
цина элемент hok/sok обеспечивает сохранение
Таким образом, для трех различных сверх-
плазмид клетками, но не обеспечивает сохра-
экспрессируемых белков мы обнаружили, что
нение экспрессии гена аспаргиназы. Прекра-
токсин-антитоксиновый генетический элемент
щение синтеза клетками ферментативно ак-
hok/sok поддерживает сегрегационную стабиль-
тивной аспарагиназы могло быть вызвано му-
ность плазмид при любом положении относи-
тациями в плазмидах или генетическими изме-
тельно промотора целевого гена, однако уве-
нениями в самих бактериальных клетках.
личивает продуктивность бактерий только при
Для двух случайно отобранных колоний
определенном положении в плазмиде - down-
клеток с плазмидой pEHD-ASN, выращен-
stream от целевого гена в случае аспарагиназы
ных на агаризованной среде с канамицином
и upstream - в случае гормона роста человека.
из бактериальной культуры, полученной после
4-х дней культивирования в среде 2x YT без
канамицина с 1 мМ ИПТГ, провели выращи-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
вание в жидкой среде с канамицином, выде-
лили плазмиды и трансформировали ими по-
Несмотря на то что эффекты позиции эле-
вторно клетки BL21[DE3]. Для трех случайно
мента hok/sok и его ориентации в лабораторных
отобранных колоний провели индукцию экс-
плазмидах непосредственно не исследовались
прессии целевого гена 1 мМ ИПТГ в среде с
ранее, возможность влияния окружающих генов
канамицином, и во всех случаях наблюдали
на функционирование цистронов hok/sok уже
уровень накопления аспарагиназы, аналогич-
отмечалась. В частности, Gerdes et al. [31] было
ный исходному штамму с плазмидой pEHD-
сделано предположение о том, что элемент hok/
ASN (данные не приводятся).
sok в составе плазмиды с индуцибельным про-
Таким образом, прекращение накопления
мотором может быть окружен двумя терминато-
клетками ферментативно активной аспара-
рами транскрипции и, таким образом, изолиро-
гиназы при периодическом культивировании
ван от влияния близкорасположенных генов.
БИОХИМИЯ том 88 вып. 9 2023
ВЛИЯНИЕ ЭЛЕМЕНТА hok/sok НА ЭКСПРЕССИЮ БЕЛКОВ Е. coli
1615
Рис. 3. Динамика накопления нуклеопротеина и гормона роста при культивировании штаммов в присутствии
и при отсутствии канамицина. Концентрацию целевых белков измеряли денситометрией электрофореграмм, усло-
вия культивирования обозначены над соответствующими панелями. а, б - Приведены данные одного репрезен-
тативного эксперимента из трех. На панелях в и г планки погрешностей соответствуют стандартным отклонениям,
n = 2, независимые биологические повторы. Для времени индукции 18 ч различия определяли с помощью одно-
параметрического дисперсионного анализа (ANOVA) с апостериорным критерием Тьюки; * p < 0.05. д, е - Опре-
деление числа КОЕ для культур, соответствующих точкам 18 ч для панелей в и г, не менее 16 колоний на каждой
анализируемой чашке. В случае, когда для одного и того же образца количество колоний на чашках с антибио-
тиком превышало таковое на чашках без антибиотика, за 100% принимали отношение числа колоний на чашках
с антибиотиком к числу колоний на чашках без антибиотика. Статистически значимые различия в долях канамицин-
устойчивых колоний по критерию Пирсона (хи-квадрат) обозначены горизонтальными чертами; ## P < 0,001.
Первичные данные приведены в Приложении 7
В ходе настоящего исследования было
по себе полная сегрегационная устойчивость
установлено, что элемент hok/sok сохраняет
плазмид с генами целевых белков недоста-
свою способность предотвращать сегрегацию
точна для поддержания высокого уровня био-
плазмиды при расположении как upstream,
синтеза соответствующих полипептидов при
так и downstream от гена целевого белка. Од-
продолжительном культивировании без анти-
новременно с этим мы обнаружили, что сама
биотиков.
БИОХИМИЯ том 88 вып. 9 2023
1616
ХОДАК и др.
При культивировании бактерий в присут-
ных плазмид с элементом hok/sok [32] - вместо
ствии канамицина для плазмид с элементом
перемещения генетического элемента hok/sok
hok/sok уровень биосинтеза периплазматиче-
в разные позиции экспрессионных конструк-
ского белка аспарагиназы не изменялся значи-
ций авторы получили один вариант плазмиды
мым образом, а уровни биосинтеза цитоплазма-
pPHB01, аналогичный pEHR, констатирова-
тических целевых белков падали примерно вдвое
ли токсичность такой плазмиды для бактерий
по сравнению с контрольными плазмидами.
вследствие избыточного уровня синтеза Hok и
Количество аспарагиназы в основном по-
провели изоляцию элемента hok/sok от целе-
вышается во временном диапазоне
6-35 ч.
вого оперона phaCAB при помощи дополни-
Возможно, волнообразное изменение коли-
тельного ρ-независимого терминатора тран-
чества аспарагиназы в течение первых 6 ч с
скрипции. При конструировании плазмиды,
прохождением через максимум в районе 2-3 ч
кодирующей ген metL под контролем про-
может быть связано с адаптацией клеток к де-
мотора Ptac, также были рассмотрены 2 ва-
лению в присутствии в периплазме больших
рианта размещения элемента hok/sok
- до
количеств аспарагиназы.
терминатора транскрипции целевого гена и
При культивировании бактерий без кана-
после терминатора, ориентация элемента в
мицина для всех тестовых белков наблюдали
обоих случаях аналогична pEHR [33]. Только
сильное падение продуктивности в случае кон-
вариант плазмиды с перемещением элемента
трольных плазмид и сохранение продуктивно-
hok/sok за терминатор транскрипции целе-
сти для некоторых плазмид с элементом hok/
вого гена обеспечивал сохранение нормаль-
sok. По-видимому, оптимальное положение и
ной при выращивании штамма-продуцента
ориентация элемента hok/sok в плазмидах на
L-гомосерина в ферментере без добавления
основе pET28 не универсальны, а должны быть
антибиотиков. Следует также отметить, что
подобраны под целевой белок, поскольку для
другие варианты токсин-антитоксиновых эле-
трех исследованных в настоящей работе белков
ментов, например, использованный в работе
были выявлены разные варианты оптималь-
Liao et al. [32] alp7, не демонстрируют необ-
ных плазмид: pEHD и pEHR - для аспараги-
ходимой сегрегационной стабилизации плаз-
назы, pEHD - для NP и pEHU - для нераство-
мид с промотором T7 целевого гена. По-ви-
римого HGH. Мы предполагаем, что уровень
димому, элемент hok/sok на настоящий мо-
экспрессии мРНК токсина Hok оказывается
мент остается единственным вариантом обес-
максимален для плазмиды pEHR при индук-
печения необходимой сегрегационной ста-
ции целевого гена, минимален для pEHD и
билизации плазмид семейства pET для их
не изменяется от базального уровня для плаз-
промышленного культивирования без анти-
миды pEHU, вследствие чего при индукции
биотиков.
экспрессии гена, кодирующего HGH, наиболь-
шая продуктивность фиксируется для плаз-
миды pEHU, а в случае NP и ASN - для плаз-
ЗАКЛЮЧЕНИЕ
миды pEHD. Токсичность целевых белков для
клетки может быть установлена как доля бак-
Обычно промышленное получение реком-
терий, сохраняющих контрольные плазми-
бинантных белков для медицинского примене-
ды после индукции в среде без антибиотика.
ния в E. coli предполагает использование силь-
В случае HGH 33% клеток сохраняли плазмиду
ных индуцибельных промоторов целевого гена
pET28a-HGH, а плазмиды pET28a-ASN и
и относительно небольшое время индукции.
pET28a-NP сохранялись у 2% и 0,3% клеток
Для таких систем экспрессии генов 2-крат-
соответственно. Исходя из полученных данных
ное повышение общего уровня биосинтеза
можно заключить, что изначальные представ-
целевого белка, достигаемое за счет введения
ления относительно токсичности указанных
в состав плазмиды небольших генетических
белков оказались неточными. Поскольку при
локусов, представляет значительный практи-
длительном культивировании (18 ч индукции)
ческий интерес. Для случая хорошо изучен-
плазмиды без hok/sok почти не сохраняются
ной токсин-антитоксиновой системы hok/sok
в случае NP и аспарагиназы, данные белки
нами было обнаружено, что варьирование по-
можно признать относительно токсичными.
ложения и ориентации hok/sok относительно
В случае HGH плазмида без hok/sok сохраня-
целевого гена позволяет подобрать вариант
ется у 33% клеток, поэтому был сделан вывод о
сегрегационно стабилизированной плазмиды,
том, что этот белок менее токсичен для клеток.
сохраняющий продуктивность бактерий при
Сходные выводы были сделаны при кон-
их культивировании без антибиотиков в тече-
струировании сегрегационно стабилизирован-
ние продолжительного времени индукции.
БИОХИМИЯ том 88 вып. 9 2023
ВЛИЯНИЕ ЭЛЕМЕНТА hok/sok НА ЭКСПРЕССИЮ БЕЛКОВ Е. coli
1617
Разработанные нами плазмидные векторы
Финансирование. Работа выполнена в рам-
pEHD, pEHD и pEHU могут быть использова-
ках госзадания ФИЦ Биотехнологии РАН.
ны для получения в клетках E. coli различных
Благодарности. Л.А. Усакину, В.О. Шендер
рекомбинантных белков без применения анти-
(ИБХ РАН); А.А. Пискаревой (МГУ имени
биотиков.
Ломоносова).
Конфликт интересов. Авторы заявляют об
Вклад авторов. И.И. Воробьев - концепция
отсутствии конфликта интересов.
и руководство работой; Н.А. Орлова, Ю.А. Хо-
Соблюдение этических норм. Настоящая
дак, Р.Р. Шайфутдинов, Д.С. Хасанов - прове-
статья не содержит описания каких-либо ис-
дение экспериментов; И.И. Воробьев, Н.А. Ор-
следований с участием людей или животных
лова, Ю.А. Ходак
- обсуждение результатов
в качестве объектов.
исследования; И.И. Воробьев, Н.А. Орлова -
Дополнительные материалы. Приложе-
написание текста; Ю.А. Ходак - редактирова-
ние к статье опубликовано на сайте журнала
ние текста статьи.
СПИСОК ЛИТЕРАТУРЫ
1.
Collins, T., Azevedo-Silva, J., da Costa, A., Branca, F.,
Proc. Natl. Acad. Sci. USA, 106, 894-899, doi: 10.1073/
Machado, R., and Casal, M. (2013) Batch production
pnas.0808832106.
of a silk-elastin-like protein in E. coli BL21(DE3):
9.
Jankevicius, G., Ariza, A., Ahel, M., and Ahel, I.
key parameters for optimisation, Microb. Cell Fact., 12,
(2016) The toxin-antitoxin system DarTG catalyzes
21, doi: 10.1186/1475-2859-12-21.
reversible ADP-ribosylation of DNA, Mol. Cell, 64,
2.
Pandey, D. P., and Gerdes, K. (2005) Toxin-antitoxin
1109-1116, doi: 10.1016/j.molcel.2016.11.014.
loci are highly abundant in free-living but lost from
10.
Wang, X., Lord, D. M., Hong, S. H., Peti, W.,
host-associated prokaryotes, Nucleic Acids Res., 33,
Benedik, M. J., Page, R., and Wood, T. K. (2013)
966-976, doi: 10.1093/nar/gki201.
Type II toxin/antitoxin MqsR/MqsA controls type V
3.
Gerdes, K., Rasmussen, P. B., and Molin, S. (1986)
toxin/antitoxin GhoT/GhoS, Environ. Microbiol., 15,
Unique type of plasmid maintenance function:
1734-1744, doi: 10.1111/1462-2920.12063.
postsegregational killing of plasmid-free cells, Proc.
11.
Aakre, C. D., Phung, T. N., Huang, D., and Laub,
Natl. Acad. Sci. USA, 83, 3116-3120, doi: 10.1073/
M. T. (2013) A bacterial toxin inhibits DNA replication
pnas.83.10.3116.
elongation through a direct interaction with the beta
4.
Lehnherr, H., Maguin, E., Jafri, S., and Yar-
sliding clamp, Mol. Cell, 52, 617-628, doi: 10.1016/
molinsky, M. B. (1993) Plasmid addiction genes of
j.molcel.2013.10.014.
bacteriophage P1: doc, which causes cell death on
12.
Wang, X., Yao, J., Sun, Y. C., and Wood, T. K. (2021)
curing of prophage, and phd, which prevents host
Type VII toxin/antitoxin classification system for
death when prophage is retained, J. Mol. Biol., 233,
antitoxins that enzymatically neutralize toxins, Trends
414-428, doi: 10.1006/jmbi.1993.1521.
Microbiol., 29, 388-393, doi: 10.1016/j.tim.2020.12.001.
5.
Singh, G., Yadav, M., Ghosh, C., and Rathore, J. S.
13.
Choi, J. S., Kim, W., Suk, S., Park, H., Bak, G.,
(2021) Bacterial toxin-antitoxin modules: classifi-
Yoon, J., and Lee, Y. (2018) The small RNA, SdsR, acts
cation, functions, and association with persistence,
as a novel type of toxin in Escherichia coli, RNA Biol.,
Curr. Res. Microb. Sci., 2, 100047, doi: 10.1016/
15, 1319-1335, doi: 10.1080/15476286.2018.1532252.
j.crmicr.2021.100047.
14.
Gerdes, K., and Maisonneuve, E. (2012) Bacterial
6.
Van Melderen, L., Thi, M. H., Lecchi, P., Gottes-
persistence and toxin-antitoxin loci, Annu. Rev.
man, S., Couturier, M., and Maurizi, M. R. (1996)
Microbiol., 66, 103-123, doi: 10.1146/annurev-micro-
ATP-dependent degradation of CcdA by Lon protease.
092611-150159.
Effects of secondary structure and heterologous
15.
Yamaguchi, Y., and Inouye, M. (2011) Regulation of
subunit interactions, J. Biol. Chem., 271, 27730-27738,
growth and death in Escherichia coli by toxin-antitoxin
doi: 10.1074/jbc.271.44.27730.
systems, Nat. Rev. Microbiol., 9, 779-790, doi: 10.1038/
7.
Michel, B. (2005) After 30 years of study, the bacterial
nrmicro2651.
SOS response still surprises us, PLoS Biol., 3, e255,
16.
Gerdes, K., Larsen, J. E., and Molin, S. (1985) Sta-
doi: 10.1371/journal.pbio.0030255.
ble inheritance of plasmid R1 requires two differ-
8.
Fineran, P. C., Blower, T. R., Foulds, I. J., Hum-
ent loci, J. Bacteriol., 161, 292-298, doi: 10.1128/
phreys, D. P., Lilley, K. S., and Salmond, G. P.
JB.161.1.292-298.1985.
(2009) The phage abortive infection system, Tox-
17.
Pecota, D. C., Osapay, G., Selsted, M. E., and Wood,
IN, functions as a protein-RNA toxin-antitoxin pair,
T. K. (2003) Antimicrobial properties of the Escherichia
12
БИОХИМИЯ том 88 вып. 9 2023
1618
ХОДАК и др.
coli R1 plasmid host killing peptide, J. Biotechnol.,
locus, Gene,
119,
91-93, doi:
10.1016/0378-1119
100, 1-12, doi: 10.1016/s0168-1656(02)00240-7.
(92)90070-6.
18.
Gerdes, K. (2016) Hypothesis: type I toxin-antitoxin
27.
Mishima, N., Mizumoto, K., Iwasaki, Y., Nakano, H.,
genes enter the persistence field-a feedback mechanism
and Yamane, T. (1997) Insertion of stabilizing loci in
explaining membrane homoeostasis, Philos. Trans. R.
vectors of T7 RNA polymerase-mediated Escherichia
Soc. Lond. B Biol. Sci., 371, 20160189, doi: 10.1098/
coli expression systems: a case study on the plasmids
rstb.2016.0189.
involving foreign phospholipase D gene, Biotechnol.
19.
Unterholzner, S. J., Poppenberger, B., and Rozhon, W.
Prog., 13, 864-868, doi: 10.1021/bp970084o.
(2013) Toxin-antitoxin systems: Biology, identifica-
28.
Galen, J. E., Nair, J., Wang, J. Y., Wasserman, S. S.,
tion, and application, Mob. Genet. Elements, 3, e26219,
Tanner, M. K., Sztein, M. B., and Levine, M. M.
doi: 10.4161/mge.26219.
(1999) Optimization of plasmid maintenance in
20.
Van Melderen, L. (2010) Toxin-antitoxin systems: why
the attenuated live vector vaccine strain Salmonella
so many, what for? Curr. Opin. Microbiol., 13, 781-785,
typhi CVD 908-htrA, Infect. Immun., 67, 6424-6433,
doi: 10.1016/j.mib.2010.10.006.
doi: 10.1128/IAI.67.12.6424-6433.1999.
21.
Pedersen, K., and Gerdes, K. (1999) Multiple hok
29.
Morin, C. E., and Kaper, J. B. (2009) Use of stabi-
genes on the chromosome of Escherichia coli, Mol.
lized luciferase-expressing plasmids to examine in
Microbiol., 32, 1090-1102, doi: 10.1046/j.1365-2958.
vivo-induced promoters in the Vibrio cholerae vac-
1999.01431.x.
cine strain CVD 103-HgR, FEMS Immunol. Med.
22.
Pedersen, K., Christensen, S. K., and Gerdes, K.
Microbiol.,
57,
69-79, doi:
10.1111/j.1574-695X.
(2002) Rapid induction and reversal of a bacterio-
2009.00580.x.
static condition by controlled expression of tox-
30.
Kolesov, D. E., Sinegubova, M. V., Safenkova, I. V.,
ins and antitoxins, Mol. Microbiol.,
45,
501-510,
Vorobiev, I. I, and Orlova, N. A. (2022) Antigenic
doi: 10.1046/j.1365-2958.2002.03027.x.
properties of the SARS-CoV-2 nucleoprotein are
23.
Wilmaerts, D., Dewachter, L., De Loose, P. J.,
altered by the RNA admixture, PeerJ, 10, e12751,
Bollen, C., Verstraeten, N., and Michiels, J. (2019)
doi: 10.7717/peerj.12751.
HokB monomerization and membrane repolarization
31.
Gerdes, K., Jacobsen, J. S., and Franch, T. (1997).
control persister awakening, Mol. Cell, 75, 1031-1042.
Plasmid Stabilization by Post-Segregational Killing,
e4, doi: 10.1016/j.molcel.2019.06.015.
in Genetic Engineering (Setlow, J. K., ed), Vol. 19,
24.
Chukwudi, C. U., and Good, L. (2015) The role of the
Springer, Boston, MA, doi:
10.1007/978-1-4615-
hok/sok locus in bacterial response to stressful growth
5925-2_3.
conditions, Microb. Pathog., 79, 70-79, doi: 10.1016/
32.
Liao, Y. C., Saengsawang, B., Chen, J. W., Zhuo,
j.micpath.2015.01.009.
X. Z., and Li, S. Y. (2022) Construction of an anti-
25.
Pecota, D. C., Kim, C. S., Wu, K., Gerdes, K., and
biotic-free vector and its application in the metabolic
Wood, T. K. (1997) Combining the hok/sok, parDE,
engineering of Escherichia coli for polyhydroxybutyr-
and pnd postsegregational killer loci to enhance
ate production, Front. Bioeng. Biotechnol., 10, 837944,
plasmid stability, Appl. Environ. Microbiol.,
63,
doi: 10.3389/fbioe.2022.837944.
1917-1924, doi: 10.1128/AEM.63.5.1917-1924.1997.
33.
Sun, B. Y., Wang, F. Q., Zhao, J., Tao, X. Y., Liu, M.,
26.
De Moerlooze, L., Struman, I., Renard, A., and
and Wei, D. Z. (2023) Engineering Escherichia coli
Martial, J. A. (1992) Stabilization of T7-promoter-
for l-homoserine production, J. Basic Microbiol., 63,
based pARHS expression vectors using the parB
168-178, doi: 10.1002/jobm.202200488.
LOCATION AND ORIENTATION OF THE GENETIC
TOXIN-ANTITOXIN ELEMENT hok/sok IN THE PLASMID
AFFECTS THE EXPRESSION LEVEL OF PHARMACEUTICALLY
SIGNIFICANT PROTEINS
Yu. A. Khodak, R. R. Shaifutdinov, D. S. Khasanov, N. A. Orlova, and I. I. Vorobiev*
Institute of Bioengineering, FRC Biotechnology, Russian Academy of Sciences,
117312 Moscow, Russia; e-mail: ptichman@gmail.com
The genetic toxin-antitoxin element hok/sok from the natural Escherichia coli R1 plasmid ensures the
segregation stability of the plasmids. Bacterial cells that have lost all copies of the plasmid encoding the
short-lived antitoxin die under the action of the long-lived toxin. The hok/sok element in vector plasmids
for bacterial expression can increase the productive time of biosynthesis of recombinant proteins, slowing
БИОХИМИЯ том 88 вып. 9 2023
ВЛИЯНИЕ ЭЛЕМЕНТА hok/sok НА ЭКСПРЕССИЮ БЕЛКОВ Е. coli
1619
down the accumulation of non-producing cells lacking the target plasmid in the population. In this work,
we studied various variants of the position and orientation of the hok/sok element in the standard plas-
mid pET28a with the inducible T7lac promoter and the kanamycin resistance gene. It was found that the
hok/sok element retained functional activity regardless of location on the plasmid and orientation, bacterial
cells retained hok/sok plasmids after four days of cultivation without antibiotics and lost the control plas-
mid without this element. Using the example of three target proteins - E. coli type II asparginase, human
growth hormone, and the SARS-CoV-2 virus nucleoprotein, it was demonstrated that for cytoplasmic target
proteins, the maximum productivity of bacteria is maintained only when the hok/sok element is located
on the plasmid upstream of the target gene promoter. In the case of periplasmic localization of the protein,
the productivity of bacteria decreases for all variants of the hok/sok location during cultivation with an an-
tibiotic, and in the case of periodic cultivation of bacteria without an antibiotic, productivity is also better
preserved when the hok/sok element is located upstream of the target gene promoter. This variant of the
pEHU vector plasmid makes it possible to more than double the biosynthesis of human growth hormone,
which is insoluble in the cytoplasm of bacteria, when bacteria are cultivated without antibiotics, and also
to maintain asparaginase biosynthesis during periodic cultivation without antibiotics for four days at a level
of at least 10 mg/liter. The developed segregation-stabilized plasmid vector can be used to obtain various
recombinant proteins in E. coli cells without the use of antibiotics.
Keywords: heterologous expression of recombinant proteins, toxin-antitoxin systems, asparaginase, nucleoprotein,
somatotropin
БИОХИМИЯ том 88 вып. 9 2023
12*