Особенности среды обитания (температура, соленость, кислородный режим и т.д.) формируют у живущих в прибрежье рыб адаптивные реакции, которые в первую очередь затрагивают органы и системы, обеспечивающие газотранспортные функции организма. Условия обитания в прибрежных водах Крыма характеризуются значительными перепадами температуры: от + 2–4°С в зимний период до + 25–28°С в летние месяцы.
Адаптационные приспособления донных рыб имеют разные уровни, среди которых особое место занимают размерные характеристики органов и их функциональная активность. Жаберный аппарат и система крови (сердце, сосуды и форменные элементы) – это самые важные составляющие, обеспечивающие общую энергетику рыб и определяющие их адаптационную основу. Ранее на пресноводных видах показана зависимость морфофизиологических параметров эритроцитов от содержания кислорода в среде обитания рыб (Заботкина и др., 2015), что подтверждает важность определения морфологических показателей этих клеток. Морские рыбы в этом отношении являются недостаточно изученными объектами. Немногочисленные экспериментальные исследования по гипоксии свидетельствуют о четкой реакции организма морских рыб на низкое содержание кислорода, приводящей к изменению физиологических и морфологических параметров крови (Солдатов и др., 1994; Солдатов, Парфенова, 2001; Силкин, Силкина, 2005; Андреева, 2014). Высокие летние температуры в прибрежной акватории приводят к падению кислородного напряжения в воде, вызывая сезонную гипоксию.
Изменение содержания гемоглобина, гликогена и активности экто-АТФазы плазматических мембран эритроцитов при неблагоприятных условиях отражает функциональную активность газотранспортной системы рыб. Гемоглобин обеспечивает емкостные характеристики кислородного транспорта, гликоген – степень метаболической автономности эритроцитов, активность экто-АТФазы – особые условия реологии эритроцитов в капиллярном отделе кровотока. Задачей настоящего исследования явилось изучение особенностей индексов сердца, жабр и размерных характеристик эритроцитов, а также содержания гликогена, гемоглобина и активности экто-АТФазы в красных клетках крови пяти видов морских донных рыб.
МАТЕРИАЛ И МЕТОДИКА
Объектами исследования служили рыбы донных биоценозов черноморского прибрежья Юго-Восточного Крыма: один представитель Chondrichthyes – скат морская лисица Raja clavata, четыре вида Teleostei – черноморская скорпена Scorpaena porcus, пятнистый звездочет Uranoscopus scaber, зеленушка-рулена Crenilabrus tinca и средиземноморский морской налим Gaidropsarus mediterraneus. Отлов производили в октябре–ноябре 2021 г., когда у большинства рыб, кроме налима, заканчивался нерест и начинался нагульный период. Костистых рыб отлавливали в 100 м от берега на глубине 5–7 м ставной сетью с ячеей 40 мм, ската – на глубине 30–50 м специальной снастью с крючками. После отлова рыб помещали в аэрируемые бассейны емкостью 100 л на одну особь. После 3-суточной акклимации рыб кормили рыбьим мясом, в течение 7 суток наблюдали за их поведением и затем брали в опыт. Соматические индексы для сердца и жабр, согласно известным методикам (Смирнов и др., 1972), определяли как отношение сырого веса органа к весу тела без внутренностей и выражали в %.
Все выловленные скаты были 50–60 см в длину и 40–45 см в ширину, длина тела S. porcus составила 20–24 см, U. scaber – 19–21 см, C. tinca – 21–24 см и G. mediterraneus – 17–20 см. Возраст рыб – от трех до пяти лет. Отловленных рыб выбирали по массе так, чтобы можно было получить достаточное количество крови: 29 особей черноморской скорпены и 16 особей пятнистого звездочета массой 150–250 г каждая; 17 экз. зеленушки-рулены массой 200–300 г и 11 экз. средиземноморского налима массой 70–100 г. Полученные данные использовали без учета пола, так как не было отмечено в процессе исследования межполовых различий по анализируемым показателям.
Получение образцов крови
У скатов кровь получали пункцией сердца, у костистых рыб – пункцией хвостовой артерии стеклянной пипеткой, смоченной раствором 5% гепарина. Для отмывания эритроцитов от других элементов крови и плазмы полученную порцию крови помещали в охлажденный физиологический раствор (1:10) следующего состава: для костистых рыб – 180 мМ NaCl + 5 мМ Tрис-HCl (pH 7.4); для ската – 220 мМ NaCl + 300 мМ СО(NH2)2 + 5 мМ Tрис-HCl (pH 7.4). Центрифугирование проводили в течение 10 мин на центрифуге JANETZKI К-23 (Германия) при 1500 об/мин, после удаляли надосадочную жидкость. В полученной суспензии эритроцитов определяли величину гематокрита, которую учитывали в опытах и расчетах при определении активности фермента и количества гликогена. Определение гликогена в эритроцитах проводили по известным цветным реакциям с антроном (Силкин и др., 2021), а активность экто-АТФазы − по накоплению фосфора согласно методике, ранее описанной в нашей статье (Силкин и др., 2018). Количество эритроцитов подсчитывали в камере Горяева, содержание гемоглобина определяли по методу Сали и выражали в г %.
Методы исследования размерных характеристик эритроцитов и их ядер
Использовали “мокрые” и “сухие” препараты клеток крови рыб. Эритроциты исследованных особей имели типичную эллипсоидную форму, отличаясь размерными характеристиками, как самой клетки, так и ядра. Толщину эритроцитов, равную толщине их ядер, определяли на “мокрых” препаратах с помощью светового микроскопа Carl Zeiss Jena (Германия) с объективом L Plan-Apochromat 100×/1.46 (иммерсионное масло Dic III), используя программу Axio Imager Z1.
На сухих мазках крови, фиксированных этиловым спиртом и окрашенных краской Романовского-Гимза, определяли длину и ширину клеток и их ядер. Оптические исследования проводили под бинокулярным микроскопом STUDAR EK PZO (Польша). Для получения снимков использовали цифровую камеру МС-6.3 USB 3.0 (“ЛОМО-МА”, РФ) (разрешение матрицы – 6.3 Мпкс, сенсор – 1/1.8″ SONY с улучшенной светочувствительностью и цветопередачей, размер пикселя – 2.4 × 2.4, разрешение полученных снимков 3072 × 2048 пикс). Измерение эритроцитов и их ядер проводили с помощью программы MCview (“ЛОМО–Микросистемы”, РФ). Расчет объема клетки Vc и ядра Vn эритроцитов осуществляли по формуле для трехосного эллипсоида, которая хорошо соответствует геометрии эритроцитов рыб (рис. 1):
(1)
$V = {4 \mathord{\left/ {\vphantom {4 {3\pi }}} \right. \kern-0em} {3\pi }}(a \times b \times c),$Рис. 1.
Эритроциты донных рыб черноморского прибрежья Крыма: а – ската Raja clavata; б – черноморской скорпены Scorpaena porcus; в – средиземноморского налима Gaidropsarus mediterraneus; г – пятнистого звездочета Uranoscopus scaber; д – зеленушки-рулены Crenilabrus tinca.
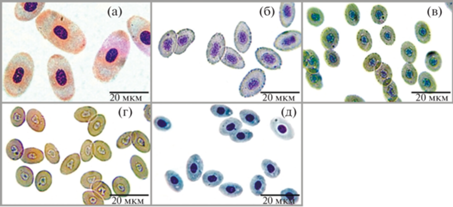
Для расчета площади одиночного эритроцита (Sс) использовали формулу, применяемую для определения площади поверхности трехосного эллипсоида (Косачев и др., 2010). Одновременный учет трех измерений эритроцита позволил обнаружить различия в площадях поверхности клеток у разных видов рыб в соответствии с их размерными характеристиками.
(2)
$S \approx 4\pi {{\left[ {{{\left( {{{a}^{p}} \times {{b}^{p}} + {\text{ }}{{a}^{p}} \times {{c}^{p}} + {\text{ }}{{b}^{p}} \times {{c}^{p}}} \right)} \mathord{\left/ {\vphantom {{\left( {{{a}^{p}} \times {{b}^{p}} + {\text{ }}{{a}^{p}} \times {{c}^{p}} + {\text{ }}{{b}^{p}} \times {{c}^{p}}} \right)} 3}} \right. \kern-0em} 3}{\text{ }}} \right]}^{{1/p}}},$Общую площадь поверхности эритроцитов рыб (RBC × Sc) рассчитывали как произведение числа эритроцитов в 1 мм3 крови (RBC) на площадь поверхности одного эритроцита.
Полученные результаты, обработанные с помощью программы Microsoft Excel, представлены в виде среднего значения и его ошибки (M ± m). Оценку результатов проводили параметрическим методом для несвязанных выборок (t-критерий Стьюдента) при уровне достоверности p ≤ 0.05 (Лакин, 1990).
РЕЗУЛЬТАТЫ
Согласно полученным данным, относящийся к хрящевым рыбам скат Raja clavata по величине сердечного индекса соответствует параметрам представителя Teleostei морского налима Gaidropsarus mediterraneus. Наибольший сердечный индекс отмечен у Scorpaena porcus, его величина несколько выше, чем у G. mediterraneus, и превышает значения сердечного индекса пятнистого звездочета Uranoscopus scaber в 1.7 раз, а зеленушки-рулены Crenilabrus tinca – в 2.5 раза (табл. 1).
Таблица 1.
Величина соматических индексов донных рыб (%)
| Индекс | Вид | ||||
|---|---|---|---|---|---|
| Raja clavata | Scorpaena porcus | Gaidropsarus mediterraneus | Crenilabrus tinca | Uranoscopus scaber | |
| Сердечный | 0.09 ± 0.004* (10) |
0.1 ± 0.001* (17) |
0.09 ± 0.003* (11) |
0.04 ± 0.002** (17) |
0.06 ± 0.001 (16) |
| Жаберный | 1.2 ± 0.1** (10) |
3.3 ± 0.1* (29) |
1.9 ± 0.1** (11) |
2.3 ± 0.2 (16) |
1.2 ± 0.01** (16) |
Жабры у ската развиты слабо, по величине жаберного индекса он, как и пятнистый звездочет Uranoscopus scaber, уступает остальным исследованным Teleostei (табл. 1). Наибольшим этот индекс был у S. porcus, он превышал индексы U. scaber, G. mediterraneus и C. tinca соответственно в 2.7, 1.4 и 1.7 раза. По уменьшению величины индекса жабр исследованные рыбы распределялись следующим образом: S. porcus, C. tinca, G. mediterraneus, R. clavata и U. scaber (табл. 1).
Анализ морфологических показателей эритроцитов и их ядер у исследованных видов рыб (табл. 2) показал, что колебания линейных размеров эритроцитов были пропорциональны изменениям размеров их ядер. Наибольшие размеры эритроцитов и их ядер отмечены у представителя Chondrichthyes. Размеры больших и малых диаметров красных клеток ската R. clavata примерно в 2 раза больше, чем у эритроцитов костистых рыб. Среди представителей Teleostei самые крупные эритроциты отмечены у черноморской скорпены, а самые мелкие – у зеленушки-рулены, у которой длины больших и малых осей эритроцита были в 1.2–1.3 раза меньше, чем у эритроцита S. porcus. Эритроциты остальных видов рыб занимали промежуточное положение. Площадь эритроцита R. clavata в 3 раза больше, чем S. porcus, площадь эритроцитов которой была наибольшей среди исследованных костистых рыб и в 1.7 раза превышала площадь клетки C. tinca. Разность объемов эритроцита и ядра (Vc – Vn), характеризующая величину цитоплазматического пространства была наибольшей у R. clavata (670.2 мкм3). Среди костистых рыб этот показатель был наибольшим у S. porcus (116.6 мкм3), а наименьшим (43.6 мкм3) – у C. tinca (табл. 2).
Таблица 2.
Размеры эритроцитов и их ядер у донных рыб (M ± m)
| Вид | Размерные параметры эритроцита | Размеры ядра эритроцита | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| С1, мкм | С2, мкм | С3, мкм | Sс, мкм2 | Vc, мкм3 | Vc – Vn, мкм3 | Sn/Sс | N1, мкм | N2, мкм | Sn, мкм2 | Vn, мкм3 | |
| Raja clavata | 26.8 ± 1.6 (6) |
14.0 ± 0.9 (6) |
3.6 ± 0.2 (6) |
668.7 ± 9.1 (6) |
728.3 ± 11.7 (6) |
670.2 ± 11.1 (6) |
0.11 ± 0.01 (6) |
6.7 ± 0.4 (6) |
4.6 ± 0.3 (6) |
76.2 ± 2.2 ( 6) |
58.1 ± 1.5 (6) |
| Scorpaena porcus | 13.5 ± 0.9 (61) |
9.1 ± 0.5 (61) |
2.1 ± 0.1 (61) |
212.0 ± 5.6 (61) |
135.1 ± 4.7 (61) | 116.6 ± 5.1 (61) |
0.18 ± 0.02 (61) | 4.8 ± 0.5 (61) | 3.5 ± 0.3 (61) |
37.1 ± 1.9 (61) | 18.5 ± 1.3 (61) |
| Gaidropsarus mediterraneus | 11.7 ± 0.1 (45) | 8.0 ± 0.7 (45) |
1.7 ± 0.1 (45) |
160.0 ± 5.6 (45) | 83.3 ± 3.9 (45) | 68.7 ± 3.1 (45) |
0.21 ± 0.02 (45) | 4.7 ± 0.3 (45) | 3.5 ± 0.3 (45) |
33.7 ± 1.5 (45) | 14.6 ± 0.6 (45) |
| Uranoscopus scaber | 11.5 ± 1.1 (27) | 8.3 ± 0.7 (27) |
1.6 ± 0.1 (27) |
161.8 ± 6.9 (27) | 80.0 ± 4.9 (27) |
67.6 ± 4.2 (27) | 0.19± 0.02 (27) | 4.5 ± 0.4 (27) | 3.3 ± 0.2 (27) |
30.3 ± 1.4 (27) | 12.4 ± 0.7 (27) |
| Crenilabrus tinca | 11.0 ± 0.9 (46) |
6.8 ± 0.6 (46) |
1.4 ± 0.2 (27) |
127.0 ± 6.5 (46) |
54.8 ± 1.7 (46) |
43.6 ± 2.7 (46) | 0.23 ± 0.02 (46) | 4.5 ± 0.4 (46) |
3.4 ± 0.3 (46) |
29.8 ± 1.3 (46) | 11.2 ± 0.5 (46) |
Примечание: С1 и С2 – большой и малый диаметры эритроцита; С3 – толщина эритроцита и ядра; N1 и N2 – большой и малый диаметры ядра; Sc – площадь поверхности эритроцита; Sn – площадь поверхности ядра; Vc – объем эритроцита; Vn – объем ядра эритроцита; Vc – Vn – объем цитоплазмы; в скобках – число рыб в опыте.
Площадь эритроцита варьировала от 668.7 мкм2 у R. clavata до 127.0 мкм2 у C. tinca и ранжировалась в следующем порядке: R. clavata > S. porcus > > U. scaber > G. mediterraneus > C. tinca . Площадь ядра Sn, определенная по снимку мазка, также была самой большой у ската (76.2 мкм2), в 2 раза превышая этот показатель для S. porcus. Однако порядок убывания по площади ядра эритроцита был несколько иной: R. clavata > S. porcus > > G. mediterraneus > U. scaber > C. tinca (табл. 2).
Минимальное отношение Sn/Sс, характеризующее долю площади клетки, занимаемую ядром, отмечено в эритроцитах R. clavata (11%). Среди костистых рыб этот показатель был наименьшим в эритроцитах S. porcus (18%), а наибольшим (23%) – у C. tinca. У G. mediterraneus и U. scaber отношение Sn/Sс было соответственно 21 и 9% (табл. 2).
Согласно результатам физиолого-биохимического исследования, количество эритроцитов в крови у костистых рыб колебалось от 420 тыс. до 1 млн (табл. 3). У представителя Chondrichthyes количество эритроцитов и содержание в них гемоглобина и гликогена было значительно ниже. В крови ската содержалось эритроцитов в 2.85 раза меньше, чем у средиземноморского налима, в 2.5 раза меньше, чем у зеленушки-рулены и пятнистого звездочета, и в 1.2 раза меньше, чем у черноморской скорпены (табл. 3). Размер красных клеток крови S. porcus в 2 раза превышал размер эритроцитов средиземноморского налима.
Таблица 3.
Количество эритроцитов, содержание гемоглобина и гликогена и активность экто-АТФазы в эритроцитах донных видов рыб
| Параметры | Вид | ||||
|---|---|---|---|---|---|
| Raja clavata | Scorpaena porcus | Gaidropsarus mediterraneus | Uranoscopus scaber | Crenilabrus tinca | |
| Количество эритроцитов в 1 мм3 крови × 106 | 0.35 ± 0.04* (10) |
0.42 ± 0.03 (18) |
1.0 ± 0.01** (16) |
0.87 ± 0.01** (16) |
0.85 ± 0.05** (15) |
| Содержание гемоглобина в крови, г % | 4.33 ± 0.42* (10) |
6.08 ± 0.32 (20) |
8.0 ± 0.85 (15) |
8.16 ± 0.38** (15) |
7.67 ± 0.83 (15) |
| Общая площадь поверхности эритроцитов, (RBC × Sc) мм2 в 1 мм3 крови | 234.0 | 89.0 | 160.0 | 141.0 | 108.0 |
| Количество гемоглобина на единицу поверхности эритроцитов, ρг/мм2 | 0.0185* | 0.0683** | 0.0500** | 0.0578** | 0.0710** |
| Содержание гликогена, мг % сырого веса клеток | 142.8 ± 15.1* (10) |
З05.2 ± 35.1** (7) |
272.6 ± 29.1** (5) |
217.0 ± 22.1 (5) |
298.2 ± 30.1** (5) |
| Активность экто-АТФазы эритроцитов, мкмоль Фн/мин/мл эритроцитов | 3.1 ± 0.2* (10) |
4.8 ± 0.3 (7) |
0.4 ± 0.02** (5) |
1.6 ± 0.2** (5) |
0.5 ± 0.02** (5) |
Уровень гемоглобина в клетках крови R. clavata в 1.4–1.9 раза ниже, чем у костистых рыб, среди которых наиболее высокое содержание гемоглобина отмечено у U. scaber. Содержание гемоглобина в эритроцитах S. porcus примерно в 1.3 раза ниже, чем в клетках крови других Teleostei. Низкий уровень гемоглобина у S. porcus сочетается с малым количеством красных клеток крови этого вида (табл. 3).
Несмотря на самый низкий гематокрит, общая площадь поверхности эритроцитов в единице объема крови оказалась самой высокой у ската R. clavata. У представителей костистых рыб этот показатель четко коррелировал с гематокритом и был самым высоким у G. mediterraneus и низким – у S. porcus (табл. 3).
Независимо от количества гемоглобина и колебаний площади эритроцитов в единице объема крови (мм3), отношение содержания гемоглобина к общей площади эритроцитарной поверхности у донных костистых рыб отличалось незначительно, и было в 2.7–3.8 раза больше, чем у ската R. clavata (табл. 3). Из чего следует, что этот показатель у представителей Teleostei можно охарактеризовать как константу, физиологический смысл которой будет рассмотрен несколько ниже.
Основного углеводного энергетического резерва – гликогена в эритроцитах – у ската содержалось в 1.5–2 раза меньше, чем у донных костистых рыб. У других исследованных видов этот показатель отличался незначительно. Крупные эритроциты R. clavata обладали высокой экто-АТФазной активностью, которая составляла 3.1 мкмоль Фн/мин/мл эритроцитов. Величина активности экто-АТФазы в эритроцитах костистых донных рыб колебалась от 0.4 до 4.8 мкмоль Фн/мин/мл эритроцитов (табл. 3). Самая высокая активность фермента среди костистых рыб отмечена в эритроцитах черноморской скорпены; у пятнистого звездочета этот показатель ниже в 3 раза, а у средиземноморского налима и зеленушки-рулены – ниже почти на порядок (табл. 3).
ОБСУЖДЕНИЕ
Все исследованные рыбы обитают в прибрежных акваториях Юго-Восточного Крыма. Скат Raja clavata, который относится к Chondrichthyes, значительно отличается от костистых рыб в плане общего анатомического, морфофункционального, осморегуляторного и биохимического устройства. Представителей Teleostei, выбранных для исследования, в соответствии с их подвижностью можно расположить в порядке убывания: Cre-nilabrus tinca > Gaidropsarus mediterraneus > Scorpaena porcus > Uranoscopus scaber. К постоянно живущим на дне рыбам относятся черноморская скорпена, пятнистый звездочет, средиземноморский налим и скат морская лисица. Зеленушка-рулена – маневренный вид, обитающий в толще воды. В связи с этим было интересно определить, какие морфофизиологические особенности дыхательной и сердечно-сосудистой систем могут обеспечить тот или иной уровень подвижности этих рыб и адаптивные реакции на динамично изменяющиеся условия прибрежной среды, а также выяснить взаимосвязь подвижности рыб с морфологическими и биохимическими параметрами эритроцитов этих видов.
Морфологические показатели жабр и сердца занимают важное место в системе оценки энергетического потенциала рыб. Согласно современным представлениям линейно-весовые показатели жаберного аппарата рыб играют решающую роль в транспорте кислорода (Матей, 1996). Размерно-весовые характеристики жабр позволяют провести межвидовое сравнение способности органа к обеспечению газообмена у рыб. Определение индекса сердца помогает оценить примерную мощность этого органа как насоса в прокачке крови и скорости доставки извлеченного жабрами кислорода к тканям и органам рыб.
Морская лисица R. clavata – рыба с не очень мощным сердцем и со слабым жаберным аппаратом (табл. 1). Индекс жабр R. clavata аналогичен индексу малоподвижного U. scaber, для которого характерны самые низкие значения этого показателя среди костистых рыб. Однако скаты не испытывают проблем с извлечением растворенного в воде кислорода. Это происходит благодаря активной вентиляции жаберного аппарата, осуществляемой путем нагнетания воды парой жаберных щелей (брызгалец), расположенных позади глаз. Кроме того, обмениваться газами со средой скатам помогает кожное дыхание, величина которого у этого вида и других донных рыб может составлять 10–20% в зависимости от степени гипоксии среды (Костоусов, 2018).
У ската, как и у костистых рыб, двухкамерное сердце, но отличительной особенностью его функционирования является активная работа артериального конуса. Сердечный конус скатов состоит из поперечнополосатых мышц, благодаря чему он способен к пульсации, позволяющей увеличивать скорость продвижения крови (Иванов, 2003). Несмотря на скромные характеристики индексов сердца, и жаберного аппарата, R. clavata нельзя отнести к пассивным обитателям морского дна. Скат активно охотится и совершает миграционные перемещения на черноморском шельфе. Благодаря крупным капиллярам, движение крови R. clavata происходит при низком давлении, которого вполне достаточно для обеспечения кислородом тканевых потребностей этих донных рыб. Низкий гематокрит у ската и других донных видов также может быть адаптацией, которая снижает потребление кислорода собственно эритроцитами рыб. Исследователи обращают мало внимания на этот момент, но, в отличие от млекопитающих, эритроциты которых являются строгими анаэробами, ядерные эритроциты рыб для поддержания своего метаболизма активно потребляют кислород и по скорости его потребления вполне сравнимы с метаболизмом белых мышц рыб (Силкин и др., 2017a).
Среди исследованных рыб самые крупные линейные размеры эритроцитов отмечены у ската R. clavata (табл. 2). Движение таких крупных эритроцитов в капиллярах неизбежно должно вызывать определенные сложности ввиду больших деформационных нагрузок на клетки. Тем не менее, они эффективно осуществляют газообмен в тканях. Благодаря большому объему клетки и относительно маленькому объему ядра, эритроциты ската обладают самым большим цитоплазматическим пространством, которое в 5–15 раз больше, чем у костистых рыб. Однако количество гемоглобина и гликогена в таком объеме цитоплазматического пространства оказалось самым низким среди исследованных особей. Низкий уровень содержания гликогена, обеспечивающего относительную метаболическую автономность эритроцитов ската, свидетельствует о достаточно благополучном и разнообразном питании этих рыб. Низкое содержание гемоглобина может указывать на невысокие емкостные характеристики крови (количество кислорода, связываемое единицей объема крови). Однако общие площади поверхностей, как одного эритроцита R. clavata (табл. 2), так и эритроцитов в единице объема крови, были самыми высокими среди исследованных рыб и не могли не сказаться на показателе количества гемоглобина, приходящегося на единицу общей поверхности аналогичного объема крови (табл. 3). У R. clavata этот показатель самый низкий (0.0185 ρг/мм2) и сопоставим с таковым у пресноводных костистых рыб (Maciak, Kostelecka-Myrcha, 2011). Низкий показатель гемоглобина на единицу площади у пресноводных костистых рыб связан с высоким количеством эритроцитов крови, а у ската малые значения этого параметра определяются низким гемоглобином, большой величиной эритроцитарной площади и в 5 раз меньшим числом эритроцитов.
Основываясь на размерных характеристиках органов исследованных рыб (табл. 1), можно заключить, что у малоподвижной S. porcus органы, извлекающие кислород из воды (жабры) и доставляющие его в организм (сердце), обладают несколько большей массой, чем у других донных видов рыб, что дает скорпене бесспорные преимущества. Действительно, S. porcus обладает феноменальными способностями переносить гипоксию, резкие перепады температуры, голодание, гипербарию и другие факторы природного и антропогенного происхождения (Силкин и др., 2018). Газотранспортная система этого вида на тканевом уровне максимально эффективно приспособлена к изменению концентрации кислорода в воде. Это стало возможным благодаря идеальному сочетанию невысокой двигательной активности и достаточно высоко сбалансированной способностью ее газотранспортного комплекса к переносу кислорода. Показано, что у высокоактивной прибрежной рыбы Diplodus vulgaris, весившей 96 г, величина поглощения кислорода на единицу массы составила 107 мл/кг, что трехкратно превышает этот показатель (29 мл/кг) у малоактивной S. porcus весом 98 г (Джелинео, 1966). Автор подчеркнул, что подвижность рыб коррелирует с концентрацией гемоглобина.
У черноморской скорпены размеры эритроцитов и их ядер почти в 2 раза больше, чем у других, более активных, донных видов, количество эритроцитов и гемоглобина существенно меньше, а уровень гликогена – выше. Низкий уровень гемоглобина находится в соответствии с небольшим количеством эритроцитов и низкой двигательной активностью этого вида. Крупные линейные размеры эритроцитов S. porcus свидетельствуют о большей емкостной способности отдельной клетки к переносу кислорода. Однако если судить о потенциальных возможностях кислородного транспорта сердечно-сосудистой системы черноморской скорпены, то ее вряд ли можно считать высокопродуктивной.
Самое высокое содержание гликогена, отмеченное в эритроцитах S. porcus, может свидетельствовать о больших перепадах уровня глюкозы в крови, возникающих из-за нестабильности поступления пищи и длительных перерывов в питании в зимний период, которые могут составлять от 6 до 16 суток (Световидов, 1964). С другой стороны, высокий уровень гликогена может указывать на высокую автономность эритроцитов и их способность использовать внутриклеточный гликоген для своих энергетических потребностей, что особенно актуально при падении уровня глюкозы в крови в случае длительного голодания.
Вторые по величине жабры принадлежат зеленушке-рулене и обеспечивают ей достаточно комфортное существование в прибрежной зоне. Однако C. tinca отличается от черноморской скорпены наличием самого малого индекса сердца (0.04), что указывает на невысокую способность сердечной мышцы к интенсивной работе. Вероятно, по этой причине C. tinca не может быстро плавать и демонстрирует не очень высокую способность к поглощению кислорода. Известно, что скорость поглощения кислорода при спонтанной активности у близкого вида – коричневого губана Labrus merula – не превышает 65 мл/кг (Джелинео, 1966). Это лишь в 2 раза выше, чем у S. porcus, что не позволяет отнести зеленушку-рулену к высокоактивным видам рыб. Среди исследованных рыб самые малые эритроциты, имеющие самые крупные ядра, характерны для зеленушки-рулены (табл. 2). Такое сочетание размерных характеристик клетки и ядра говорит о малом полезном объеме цитозоля в эритроците. Количество эритроцитов у C. tinca в 2 раза выше, чем у S. porcus, а количество гемоглобина больше только на 26% (табл. 3), что свидетельствует о низком содержании гемоглобина в одном эритроците. Как и у черноморской скорпены, высокая концентрация гликогена, найденная в эритроцитах C. tinca, указывает на скудность питания этого вида и способность его эритроцитов к автономному энергетическому существованию.
Согласно нашим исследованиям, средиземноморский налим G. mediterraneus наделен очень сбалансированными величинами индексов жабр и сердца. Индекс сердца у налима практически такой же, как у S. porcus, а индекс жабр на 74% меньше (табл. 1). Вероятно, более низкий индекс жаберного аппарата отражает низкую способность к перенесению гипоксии, которая может компенсироваться незначительной двигательной активностью, свойственной G. mediterraneus, как ночному хищнику. По спонтанной двигательной активности этот вид нельзя отнести к активным формам. Однако низкая двигательная активность G. mediterraneus и подобных ему видов не является признаком их малоподвижности. По меткому выражению В. Беляева и его соавторов, это рыбы, производящие малый объем работы (Беляев и др., 1983). Наличие относительно мощного сердца позволяет им резко нарастить объемы транспортируемого кислорода и кратно увеличить метаболизм в тканях. Высокому уровню доставки кислорода способствуют также самое большое количество эритроцитов и высокий уровень гемоглобина в крови G. mediterraneus (табл. 3). Кроме того, у этого вида рыб хорошо развиты способности к гипоксическому использованию гликогена при совершении мощных и стремительных бросков для захвата жертвы. Такая стратегия обеспечивает им успешную охоту, а малый объем работы позволяет держать на низком уровне общие метаболические расходы. К тому же достаточно высокая концентрация гликогена в клетках обеспечивает стабильность функционирования эритроцитов, несмотря на частую гипогликемию, связанную с сезонной и суточной активностью охоты в прибрежье. Исходя из этих соображений, феномен малоподвижности необходимо рассматривать как особую форму адаптации, которая позволяет этим рыбам успешно выживать в прибрежной акватории моря.
Среди исследованных рыб самые низкие значения жаберного индекса отмечены у пятнистого звездочета U. scaber. Индекс сердца у него также низок и сопоставим с индексом сердца у C. tinca. Такие показатели являются отражением истинной малоподвижности, присущей этой рыбе, которая крайне редко покидает свое убежище – углубление, вырытое в песчаном грунте. Стратегия U. scaber определяется способностью привлекать к себе жертв (в основном мелкую рыбу) электромагнитными импульсами и ложно выбрасываемой изо рта складкой верхней губы, напоминающей полихету, сводя к минимуму движения при захвате жертвы. Спонтанная активность его чрезвычайно низка и в некоторых случаях не превышает 1.4 м/час (наши данные), что совпадает с величиной двигательной активности Arnoglossus boscii (Джелинео, 1966). Потребление кислорода у U. scaber, вероятно, тоже низкое и, возможно, сопоставимо с поглощением кислорода у A. boscii, которое равно 26 мг/кг (Джелинео, 1966). Размеры эритроцитов U. scaber и G. mediterraneus практически одинаковы (табл. 2), и количество эритроцитов у пятнистого звездочета лишь немного уступает количеству красных клеток крови у G. mediterraneus. Однако эти различия компенсируются самым высоким содержанием гемоглобина в клетках крови U. scaber. В этом смысле наши результаты находятся в противоречии с мнением, что высокий гемоглобин свойственен только высокоактивным рыбам (Белокопытин, Ракицкая, 1981). Согласно полученным нами данным, эритроциты пятнистого звездочета отличаются низким содержанием гликогена, что свидетельствует о достаточно регулярном питании, которому, несомненно, способствует “военная” хитрость, применяемая этой рыбой при поимке жертвы.
Количество кислорода, переносимого эритроцитами в единицу времени, зависит от скорости кровотока (Nikinmaa, 2002), а эффективность обмена газов находится под влиянием скорости потока воды через жабры (Wieser, 1991). Этот механизм перекрестного газообмена у рыб столь совершенен, что обеспечивает достаточное время контакта эритроцитов с кислородом независимо от скорости кровотока в жабрах (Kostelecka-Myrcha, 1997; 2002). Благодаря этому у пресноводных рыб, даже приспособленных к разным эколого-физиологическим условиям, количество гемоглобина на единицу поверхности эритроцитов в 1 мм3 крови, которое отражает эффективность насыщения гемоглобина кислородом, поддерживается на относительно постоянном уровне (0.021–0.04 ρг/мм2) (Maciak, Kostelecka-Myrcha, 2011). Согласно нашим данным, у морских костистых рыб соотношение гемоглобина (Hb) на единицу поверхности эритроцитов в 1 мм3 крови было в 2–3 раза выше, чем у пресноводных костистых рыб (табл. 3), и только у R. clavata, представителя Chondrichthyes, совпадало с данными по пресноводным рыбам. Низкие значения этого показателя у пресноводных рыб, возможно, являются свидетельством того, что, по сравнению с океанами и морями, колебания парциального напряжения кислорода в пресных водоемах имеют большую лабильность. Это может вызывать общее увеличение количества эритроцитов в крови. В таком случае более высокие значения отношения содержания гемоглобина к площади эритроцитарной поверхности в 1 мм3 крови (Hb/RBCxSc) (0.05–0.071 ρг/мм2) (табл. 3) у морских донных Teleostei можно объяснить более стабильным содержанием кислорода в морской среде.
Активность экто-АТФазы эритроцитов у C. tinca и G. mediterraneus на порядок ниже, чем у S. porcus, и в 3–4 раза выше, чем у U. scaber (табл. 3). Установлено, что величина активности экто-АТФазы четко коррелировала с размерами эритроцитов рыб. Отмечено, что активность экто-АТФазы у оседлых, малоподвижных рыб (R. clavata, S. porcus и U. scaber), которые сталкиваются в прибрежье с низкими температурами (4–8°С), была выше, чем у других исследованных видов. Ранее на основании результатов широкого скрининга активности экто-АТФазы у 34 видов, относящихся к шести классам позвоночных от рыб до млекопитающих, установлено, что активность экто-фермента в эритроцитах варьировала в широком диапазоне (Bencic et al., 1997). Различия в активности достигали шести порядков. Авторы исследования не установили причин такой большой вариации ферментативной активности поверхностно-локализованной АТФазы. Внимательно проанализировав эти данные, мы обнаружили, что все эритроциты с высокой активностью экто-АТФазы были крупными клетками, а животные с высокой активностью экто-ферментов эритроцитов характеризовались высокой адаптивной способностью к перенесению низких температур. Функциональное назначение экто-АТФаз неизвестно. Мы предположили, что способность некоторых животных переносить низкие температуры может обеспечиваться за счет тепловой энергии гидролиза экто-АТФаз, которая поддерживает минимальную функциональную активность кровотока, обеспечивая преимущественную возможность занимать северные широты, недоступные видам, эритроциты которых обладают невысокими теплопродукционными способностями. Эта тепловая энергия может также лежать в основе эффекта Фареуса-Линдквиста и обеспечивать 20-кратное различие вязкостных характеристик крови в капиллярах диаметром меньше 300 мкм (Katiukhin, 2014). Она же позволяет изменять вязкоэластичные свойства мембраны крупных эритроцитов, давая им возможность легко преодолевать капиллярный отдел кровотока, что обеспечивает эффективность газотранспортной функции крови рыб, особенно обитателей донных биоценозов (Силкин и др., 2014, 2017б; Силкин, Силкина, 2017).
Таким образом, донные рыбы, обитающие в прибрежной акватории Юго-Восточного Крыма, обладают рядом общих и индивидуальных адаптивных приспособлений на морфологическом и физиолого-биохимическом уровнях. Высокие значения жаберного индекса S. porcus и G. mediterraneus свидетельствуют о высоком потенциале к извлечению кислорода при экстремальных состояниях, а высокий индекс сердца указывает на эффективные транспортные возможности кровеносной системы этих рыб. Такое сочетание индексов сердца и жаберного аппарата можно рассматривать как адаптацию, позволяющую успешно осваивать экологические ниши среды обитания недоступные менее приспособленным к гипоксии видам рыб. У других донных рыб низкие возможности дыхания и сердечной мышцы компенсировались крайне низкой спонтанной двигательной активностью, как у пятнистого звездочета, или размеренной активностью, как у ската и зеленушки-рулены.
Эритроциты представителя Chondrichthyes (ската) – самые крупные среди исследованных рыб, они в несколько раз превосходили эритроциты костистых рыб по площади поверхности и по внутриклеточному объему. Несмотря на большое цитоплазматическое пространство, эритроциты у R. clavata и S. porcus содержали низкое количество гемоглобина. Количество гемоглобина на единицу поверхности эритроцита, отражающее эффективность насыщения гемоглобина кислородом, у исследованных костистых рыб варьировало незначительно, и было в 2.7–3.8 раза выше, чем у ската, и в 2–3 раза выше, чем у пресноводных костистых рыб. Низкая насыщенность гемоглобина кислородом в эритроцитах пресноводных костистых рыб может быть связана с большим колебанием парциального напряжения кислорода пресных водоемов. В пользу этого предположения свидетельствует высокий гематокрит крови пресноводных видов рыб, более чем в 2 раза превышающий таковой черноморских донных рыб. Низкий гематокрит у R. clavata и S. porcus также можно рассматривать как адаптацию к гипоксии, направленную на экономию расходов кислородного потребления на собственные потребности крови.
Внутриклеточная концентрация гликогена, характеризующая степень независимости эритроцита от перепадов гликемии, была самой высокой в красных клетках S. porcus, а самой низкой – в эритроцитах R. clavata и U. scaber. Низкий уровень гликогена в эритроцитах характерен для рыб, не испытывающих больших периодов голодания и более удачливых в добыче пищи.
Активность экто-АТФазы в крупных эритроцитах донных рыб R. clavata, S. porcus и U. scaber была выше, чем в мелких красных клетках средиземноморского налима и зеленушки-рулены. Это подтверждает предположение об использовании эритроцитами тепловой энергии гидролиза внеклеточного АТФ для “подогрева” собственной мембраны, позволяющего изменить реологические характеристики крови в капиллярном отделе кровотока.