Остатки динозавров встречаются в нескольких местонахождениях бостобинской свиты (верхний мел, сантон–кампан) в Северо-Восточном При- аралье (Кызылординская область, Казахстан), наиболее богатым из которых является Шах-Шах [1, 2]. Здесь отмечены завроподы, тероподы Tyrannosauroidea indet., Ornithomimidae indet., Therizinosauroidea indet., Caenagnathidae? indet., Dromaeosauridae indet. и Troodontidae indet., анкилозавры и гадрозавроиды Aralosaurus tuberiferus Rozhdestvensky, 1968 [2–11]. Предварительное определение Caenagnathidae основано на зубной кости из Шах-Шаха, которая первоначально была отнесена к черепахам, но может принадлежать овирапторозаврам [6, 12, 13]. Дромеозавриды из Шах-Шаха были известны по когтевым фалангам [11] и изолированным зубам [6]. Недавно все когтевые фаланги, описанные Ю.В. Сусловым из Шах-Шаха [11], были отнесены к Therizinosauridae [2], однако по крайней мере часть из них действительно принадлежат дромеозавридам. В данной работе описываются две изолированные плюсневые кости хищных динозавров из Шах-Шаха из сборов экспедиции Палеонтологического института АН СССР под руководством А.К. Рождественского 1957 г. Одна из этих костей может быть отнесена к овирапторозаврам семейства Caenagnathidae, вторая – к Dromaeosauridae. Первая находка подтверждает существование ценагнатид в комплексе позвоночных бостобинской свиты. Материал хранится в коллекции Палеонтологического института им. А.А. Борисяка РАН (ПИН), г. Москва.
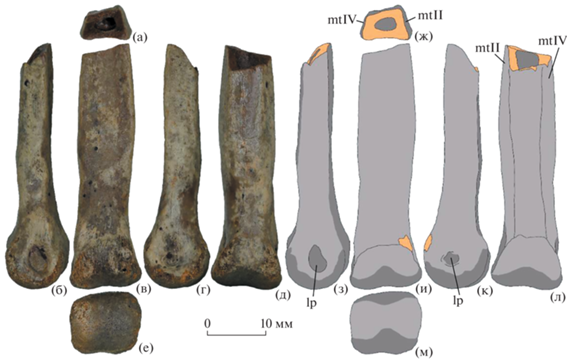
Экз. ПИН, № 2229/7 представляет собой дистальный фрагмент правой III пястной кости (метатарзалии III: рис. 1). Диафиз кости расширен медиолатерально близ проксимального конца фрагмента и сужается в проксимальном и дистальном направлениях. В наиболее широкой части диафиз лишь немного уступает по ширине дистальному эпифизу. Медиальный край диафиза более выпуклый, чем латеральный. Экстенсорная поверхность диафиза плоская близ дистального эпифиза и вогнутая проксимальнее. Свободная флексорная поверхность диафиза, не перекрываемая боковыми метатарзалиями, вогнутая. Ее ширина составляет примерно половину всей флексорной поверхности диафиза. Фасетка для контакта с метатарзалией II немного больше, чем фасетка для метатарзалии IV. Диафиз кости полый, с толстыми костными стенками (рис. 1 а, ж). При виде с медиальной или латеральной стороны диафиз кости изогнут проксимальнее дистального эпифиза (рис. 1 б, г, з, к). При виде с экстенсорной или флексорной стороны дистальный эпифиз отогнут немного медиально, из-за чего латеральный мыщелок выдается более дистально по сравнению с латеральным мыщелком (рис. 1 в, д, и, л). Дистальный эпифиз слабо гинглимоидный, с очень неглубокой бороздкой, разделяющей медиальный и дистальный мыщелки. Дистальный эпифиз субпрямоугольный при виде с дистального конца. Его медиолатеральная ширина (12.4 мм) больше дорсовентрального диаметра (11.0 мм). Мыщелки округлой формы при виде с медиальной или латеральной стороны. Сочленовная поверхность медиального мыщелка на экстенсорной и флексорной сторонах простирается более проксимально по сравнению с латеральным мыщелком. Связочная ямка латерального мыщелка крупная и глубокая, с хорошо очерченными краями. Она расположена в центре латерального мыщелка. На медиальном мыщелке связочная ямка также крупная, но не такая глубокая и не имеет четко очерченного края.
Рис. 1.
Caenagnathidae indet., экз. ПИН, № 2229/7, правая метатарзалия III, фотографии (а–е) и пояснительные рисунки (ж–м), вид с проксимального конца (а, ж), с латеральной стороны (б, з), с экстенсорной стороны (в, и), с медиальной стороны (г, к), с флексорной стороны (д, л) и с дистального конца (е, м). Шах-Шах, Кызылординская область, Казахстан; бостобинская свита, верхний мел (сантон–кампан). Обозначения: lp, связочная ямка; mtII, фасетка для метатарзалии II; mtIV, фасетка для метатарзалии IV.
Метатарзалия III, экз. ПИН, № 2229/7 относится к субарктометатарзальному типу стопы, при котором боковые метатарзалии (II и IV) не полностью закрывают среднюю метатарзалию (III) с вентральной (флексорной) стороны. Поперечное сечение диафиза метатарзалии III при таком типе стопы имеет вид трапеции (рис. 1 а, ж). При арктометатарзальном типе стопы, характерном для Tyrannosauridae, Ornithomimidae, Avimimidae, Alvarezsauridae и Troodontidae, боковые метатарзалии полностью перекрывают среднюю метатарзалию вентрально, и ее диафиз имеет в сечении треугольную форму [14]. Субарктометатарзальный тип стопы встречается у овирапторозавров семейства Caenagnathidae и некоторых базальных дромеозаврид или авиал (Microraptorinae, Unenlagiinae) [15]. В частности, экз. ПИН, № 2229/7 довольно сходен по строению с метатарзалией III уненлагиина Neuquenraptor argentinus Novas et Pol, 2005 из коньяка Аргентины ([16]: рис. 7). Экз. ПИН, № 2229/7 отличается большей шириной свободной флексорной поверхности диафиза и менее глубокой бороздой, разделяющей дистальные мыщелки. Все уненлагиины известны только из Южной Америки, поэтому отнесение экз. ПИН, № 2229/7 к этой группе представляется маловероятным. Еще большее сходство экз. ПИН, № 2229/7 обнаруживает с метатарзалией III ценагнатид, известных из верхнего мела Азии и Северной Америки. От метатарзалии III Elmisaurus rarus Osmólska, 1981 из маастрихта Монголии [17] экз. ПИН, № 2229/7 отличается немного более узкой свободной вентральной стороной диафиза и связочными ямками, которые у E. rarus глубокие на обоих мыщелках. От метатарзалии III Citipes elegans (Parks, 1933) из кампана Канады ([18]: рис. 2) экз. ПИН, № 2229/7 отличается лишь более выпуклым медиальным краем диафиза и сочленовной поверхностью дистального мыщелка, который в меньшей степени распространяется на вентральную сторону. Такое значительное сходство позволяет уверенно отнести экз. ПИН, № 2229/7 к представителю семейства Caenagnathidae.
Экз. ПИН, № 2229/8 является проксимальным фрагментом левой IV плюсневой кости (метатарзалии IV) вместе с приросшей предплюсневой костью (дистальной тарзалией III; рис. 2). Швы между костями различимы на медиальной и проксимальной сторонах (рис. 2). Дистальная тарзалия не полностью покрывает проксимальную сторону метатарзалии, не доходя до ее дорсального (экстенсорного) края. При виде с медиальной или латеральной стороны диафиз метатарзалии IV значительно расширен на проксимальном конце и немного расширяется в дистальном направлении. При виде с дорсальной или вентральной стороны диафиз плавно сужается в дистальном направлении. Проксимальная поверхность плоская и имеет форму буквы D при виде с проксимального конца, с прямым медиальным краем (рис. 2 а, ж). На медиальной стороне проксимальный конец занимает крупная фасетка треугольной формы для проксимального конца метатарзалии III. На латеральной стороне имеется существенно более мелкая фасетка треугольной формы для метатарзалии V. Экстенсорная поверхность диафиза вогнутая в поперечной плоскости. Вдоль ее медиального края имеется плоская продольная фасетка для диафиза метатарзалии III, которая отделена небольшим перерывом от проксимальной фасетки. Продольная фасетка расширяется в дистальном направлении, так что близ сохранившегося дистального конца ее латеральный край находится посередине экстенсорной поверхности диафиза. В этом месте диафиз кости имеет треугольную форму поперечного сечения (рис. 2 е, м). Вентральный (флексорный) край фасетки для метатарзалии III образует острый гребень. Флексорная поверхность диафиза кости плоская. Экстенсорная и флексорная поверхности диафиза сходятся к ее латеральному краю, образуя мощный латеральный гребень вдоль всего сохранившегося диафиза. Медиолатеральная ширина проксимального эпифиза – 11.4 мм, его дорсовентральный диаметр – 14.3 мм.
Рис. 2.
Dromaeosauridae indet., экз. ПИН, № 2229/8, левые дистальная тарзалия III и метатарзалия IV, фотографии (а–е) и пояснительные рисунки (ж–м), вид с проксимального конца (а, ж), с медиальной стороны (б, з), с экстенсорной стороны (в, и), с латеральной стороны (г, к), с флексорной стороны (д, л) и с дистального конца (е, м). Шах-Шах, Кызылординская область, Казахстан; бостобинская свита, верхний мел (сантон–кампан). Обозначения: dtIII, дистальная тарзалия III; lc, латеральный гребень; mtIII, фасетка для метатарзалии III; mtV, фасетка для метатарзалии V; s, шов между дистальной тарзалией III и метатарзалией IV.

Экз. ПИН, № 2229/8 практически идентичен по строению метатарзалии IV дромеозаврида Velociraptor mongoliensis Osborn, 1924 из кампана Монголии [19]. У велоцираптора дистальная тарзалия III также не полностью покрывает проксимальную поверхность метатарзалии IV и имеется небольшая метатарзалия V, контактирующая с проксимальным концом метатарзалии IV ([19]: рис. 15, 16). Кроме того, у велоцираптора, как и у экз. ПИН, № 2229/8, имеется мощный гребень вдоль латерального края метатарзалии IV ([19]: рис. 14). У большинства теропод, включая других дромеозаврид, латеральная поверхность метатарзалии IV плоская. У Balaur bondoc Csiki et al., 2010 из маастрихта Румынии метатарзалии толстые вдоль одного края и утончаются к противоположному краю, образуя гребень вдоль него [20]. Однако у Balaur на метатарзалии IV гребень идет вдоль медиального, а не латерального края, противоположно экз. ПИН, № 2229/8. Экз. ПИН, № 2229/8 можно уверенно отнести к Dromaeosauridae indet. на основании отмеченного сходства с метатарзалией IV велоцираптора.