ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 98, 1, 2019
УДК 591.3 (595.792)
ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИВОСТЬ ИНДУКЦИИ ДИАПАУЗЫ
ПОТОМСТВА У САМОК TRICHOGRAMMA TELENGAI SOR.
(HYMENOPTERA, TRICHOGRAMMATIDAE)
© 2019 г. C. Я. Резник*, Н. Д. Войнович
Зоологический институт РАН
Университетская наб., 1, С.-Петербург, 199034 Россия
*e-mail: reznik1952@mail.ru
Поступила в редакцию 22.01.2019 г.
После доработки 24.01.2019 г.
Принята к публикации 24.01.2019 г.
В лабораторных условиях исследована индивидуальная изменчивость самок партеногене-
тической лабораторной линии Trichogramma telengai Sor. (Hymenoptera, Trichogrammatidae) по
доле диапаузирующего потомства. Материнское поколение развивалось при температуре 20 °С
и длине дня 12, 16 и 18 ч. Потомство инкубировали при низкой температуре (6, 9, 12 или 15 °C)
в течение 15 или 30 дней. Индивидуальная изменчивость оказалась статистически достоверной
в большинстве вариантов опыта, включая и те случаи, когда средняя доля диапаузирующего
потомства была близка к 90 %. Распределение самок по доле диапаузирующего потомства было
унимодальным, а в тех случаях, когда средняя доля диапаузирующего потомства составляла око-
ло 50 %, - близким к нормальному, т. е. индивидуальная изменчивость была не качественной,
а количественной (градуальной). Корреляция между долями диапаузирующих особей в потом-
стве, произведенном одной и той же самкой во время последовательного заражения разных пор-
ций яиц зерновой моли, была достоверно положительной. Следовательно, изменчивость тен-
денции к диапаузе (по крайней мере отчасти) определяется индивидуальными особенностями
трихограмм. Индивидуальная изменчивость самок по доле диапаузирующего потомства должна
учитываться при разработке методов лабораторного исследования и массового разведения три-
хограмм.
Ключевые слова: диапауза, изменчивость, температура, фотопериод, Trichogramma telengai.
DOI: 10.1134/S0367144519010015
Диапауза - одна из важнейших эколого-физиологических адаптаций, необходимая
для синхронизации циклов активности многих видов насекомых с сезонной динами-
кой факторов окружающей среды (Tauber et al., 1986; Danks, 2007). Эта реакция упре-
ждающая, она индуцируется еще до наступления периода неблагоприятных условий
сигнальными факторами: фотопериодом (длиной светового дня), температурой и др.
(Данилевский, 1961; Заславский, 1984; Saunders et al., 2002; Denlinger, 2002; Саулич,
Волкович, 2004; Saunders, 2010). Фотопериодические и температурные реакции, осу-
ществляющие индукцию диапаузы и тем самым обеспечивающие переживание не-
благоприятных периодов, постоянно контролируются и корректируются естествен-
5
ным отбором не только на видовом, но и на внутривидовом (популяционном) уровне.
У большинства исследованных в этом отношении широко распространенных видов
насекомых выявлена более или менее значительная межпопуляционная изменчивость
фотопериодических и температурных реакций, позволяющая адаптироваться к особен-
ностям климата в разных частях ареала. Иногда обнаруживается также и значительная
индивидуальная (внутрипопуляционная) изменчивость, служащая обязательной пред-
посылкой микро- и макроэволюции сезонных адаптаций (Tauber et al., 1986; Саулич,
1999; Denlinger, 2002; Saunders et al., 2002; Danks, 2007; Saunders, 2010; Goto, Numata,
2014). Все случаи изменчивости различных параметров сезонно-циклических реакций
(как и других признаков) можно по внешним проявлениям разделить на две категории:
дискретная (полиморфизм) и градуальная изменчивость (Саулич, Волкович, 2004).
Количественный анализ индивидуальной изменчивости различных параметров тем-
пературных и фотопериодических реакций, контролирующих индукцию диапаузы,
осложняется тем, что конечная реакция носит качественный характер: диапауза либо
индуцируется, либо нет. Для сравнения выборок (в том числе и для семейного анализа)
можно использовать, например, долю особей, диапаузирующих при каких-либо опре-
деленных условиях, но для сравнения отдельных насекомых этот способ неприменим.
Заметим, что изменчивость количественных фотопериодических и температурных ре-
акций (влияние на скорость развития, размер, плодовитость и т. п.) может исследо-
ваться теми же методами, что и изменчивость количественных морфологических при-
знаков. Это же относится и к количественным параметрам диапаузы (длительность,
интенсивность метаболизма, рост устойчивости к экстремальным условиям среды и т.
п.), но не к индукции диапаузы как таковой. В этом отношении особенно интересно так
называемое «материнское влияние», при котором тенденция к диапаузе определяется
факторами, действовавшими на одно или даже несколько предшествующих поколе-
ний (Заславский, 1984; Mousseau, Dingle, 1991; Denlinger, 2002; Saunders et al., 2002;
Саулич, Волкович, 2004). Сравнение материнских особей по доле диапаузирующего
потомства отличается от «классического» семейного анализа тем, что сравниваемые
выборки могут различаться не только генетически, но и по силе материнского влияния,
индуцирующего или ингибирующего диапаузу потомства.
Специальные исследования, посвященные индивидуальной изменчивости самок по
характеру материнского влияния на диапаузу или сезонный полиморфизм потомства,
были проведены на нескольких модельных видах насекомых (Lees ,1959; Saunders,
1965; Виноградова, Резник, 2000). Сильное материнское влияние обнаружено и у мно-
гих видов рода Trichogramma Westw. (Hymenoptera, Trichogrammatidae). Эти мельчай-
шие паразитоиды-яйцееды - естественные враги многих чешуекрылых вредителей
сельского и лесного хозяйства, широко использующиеся для биологической защиты
растений (Smith, 1996; Сорокина, 2011). Кроме того, трихограммы оказались весьма
удобными модельными объектами для изучения механизмов регуляции диапаузы. Из-
вестно, что факультативная зимняя предкуколочная диапауза трихограмм, обитающих
в условиях умеренного климата, регулируется прежде всего температурой, но при око-
лопороговых температурах доля диапаузирующего потомства существенно зависит от
фотопериодических условий развития материнского и (в ряде случаев) предшеству-
ющего ему поколений (Заславский, Умарова, 1981; Май Фу Кви, Заславский, 1983;
Zaslavski, Umarova, 1990; Boivin, 1994; Резник, 2011).
6
Сравнительные исследования выявили межпопуляционную изменчивость темпера-
турных и фотопериодических реакций трихограмм. Как и следовало ожидать, особи
из северных популяций обычно проявляют большую тенденцию к диапаузе (Сороки-
на, 1987; Сорокина, Масленникова, 1987; Сорокина, 2010). Кроме того, существенные
изменения доли диапаузирующего потомства наблюдаются в последовательных поко-
лениях лабораторных линий разных видов трихограмм (Заславский, Умарова, 1981;
Май Фу Кви, Заславский, 1983; Войнович и др., 2013; Резник и др., 2015). Однако
количественный анализ индивидуальной изменчивости трихограмм по силе индукции
диапаузы потомства (сравнение самок одного поколения, происходящих из одной ла-
бораторной линии и развивавшихся в одних и тех же условиях), насколько нам извест-
но, ранее не проводился. Мы исследовали изменчивость самок Trichogramma telengai
Sor. по доле диапаузирующего потомства на фоне воздействия внешних сигнальных
факторов (фотопериода и температуры).
МАТЕРИАЛ И МЕТОДИКА
В работе была использована партеногенетическая линия T. telengai, происходящая от
особей, собранных в Московской обл., и на протяжении многих лет разводимая на яй-
цах зерновой моли Sitotroga cerealella Oliv. (Lepidoptera, Gelechiidae) при константных
лабораторных условиях (температура 20 °C, длина дня 18 ч).
Целью первого опыта было выявление индивидуальной изменчивости самок
T. telengai по доле диапаузирующего потомства при различной силе индукции диапау-
зы (т. е. при различных фотопериодических условиях развития материнского поколе-
ния и различных температурных условиях развития потомства) и определение харак-
тера этой изменчивости (дискретна она или градуальна). В начале каждой повторности
опыта около 1000 яиц зерновой моли, наклеенных клеем ПВА на небольшие картонные
карточки, предоставляли на 24 ч для заражения самкам лабораторной линии. Сразу
после этого зараженные яйца переносили для развития трихограмм в один из двух
фотопериодических режимов, различающихся длиной дня: 12 ч (С : Т = 12 : 12) или
18 ч (С : Т = 18 : 6). Температура в обоих режимах была одинаковой (20 °C). В день
массового вылета имаго этого первого (материнского) поколения самок рассаживали
по одной в маленькие пробирки и предоставляли им для заражения карточки с яйцами
зерновой моли (около 50 яиц на самку). Заражение длилось 2 ч, после чего карточки
распределяли между 8 режимами, различающимися температурой (6, 9, 12 или 15 °C)
и длительностью холодовой экспозиции (15 или 30 дней). Длина дня во всех темпера-
турных режимах развития потомства была 12 ч. После завершения холодовой экспози-
ции все зараженные яйца содержали при температуре 20 °C до вылета имаго активной
(не диапаузирующей) фракции потомства. После вылета имаго активной фракции все
зараженные яйца вскрывали для определения числа активно развивавшегося (имаго и
куколки) и диапаузирующего (предкуколки) потомства. Число вылетевших имаго оце-
нивали по числу пустых зараженных яиц с летными отверстиями. Личинок, погибших
на разных стадиях развития, при определении доли диапаузирующих особей не учиты-
вали. В каждом из 16 вариантов этого опыта (сочетания двух фотопериодических ре-
жимов развития материнского поколения и восьми температурных режимов развития
потомства) было исследовано потомство 50 самок, но для анализа результатов были
использованы только те самки, у которых к моменту завершения опыта (вскрытия за-
раженных яиц) было не менее 5 выживших потомков. Объемы выборок приведены на
рис. 1.
7
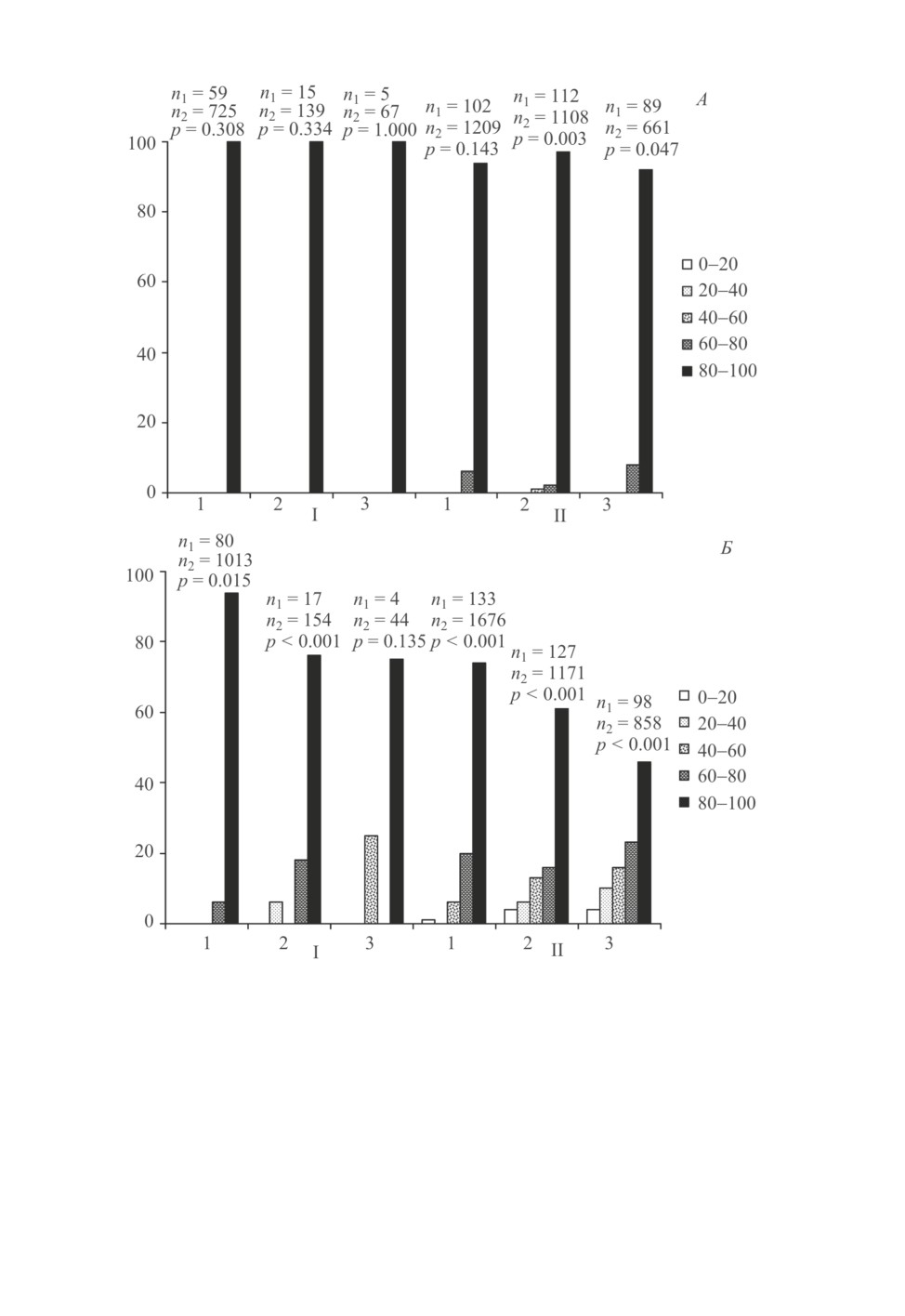
Рис. 1. Изменчивость доли диапаузирующего потомства самок Trichogramma telengai Sor.
в зависимости от фотопериодических условий развития материнского поколения
и температурных условий развития потомства (результаты первого опыта).
По горизонтальной оси - температура во время холодовой экспозиции потомства (°С). По вертикальной
оси - процентное распределение самок по классам, различающимся по доле диапаузирующего потомства
(границы классов указаны на рисунке). Длина дня во время развития материнского поколения:
А и Б - 12 ч, В и Г - 18 ч. Длительность холодовой экспозиции потомства: А, В - 15 дней;
Б, Г - 30 дней. Над каждой гистограммой указаны объемы выборок (n1 - число самок,
n2 - число потомков) и достоверность неоднородности выборки (достоверность
различий между самками), определенная с помощью теста хи-квадрат.
8
Рис. 1 (продолжение).
Целью второго опыта было получение ответа на вопрос: определяется изменчивость
доли диапаузирующего потомства особенностями самок или случайными вариация-
ми каких-либо параметров получаемых ими порций яиц зерновой моли? Кроме того,
задачей опыта был анализ корреляции между тенденцией к диапаузе потомства и пло-
довитостью самок. В опыте были использованы самки, развивавшиеся при длине дня
12, 16 и 18 ч; их потомство во всех вариантах опыта развивалось при температуре
9
14 °C. В ходе опыта одной и той же самке последовательно предоставляли для зара-
жения три порции из 40-50 яиц. Каждая экспозиция (заражение данной порции яиц)
длилась 2 ч. В первом варианте опыта экспозиции следовали одна за другой, во вто-
ром варианте они были разделены двухдневными промежутками, во время которых
самки получали углеводную подкормку (50%-ный водный раствор меда). В обоих
вариантах опыта для каждой порции определяли число зараженных (потемневших)
яиц хозяина и долю диапаузирующего потомства (по той же методике, что и в первом
опыте). В общей сложности в первом варианте опыта было использовано 300, а во
втором - 600 самок, но в обоих вариантах часть их них погибла до завершения опыта,
а некоторые из выживших не заразили ни одного хозяина (особенно часто это наблю-
далось во время второй и третьей экспозиций). Фактические объемы выборок указаны
на рис. 2-5.
Статистическая обработка результатов включала дисперсионный анализ, перед кото-
рым процентные величины были ранжированы. Для оценки гетерогенности выборки
(достоверности различий между отдельными самками) использовался тест хи-квадрат.
РЕЗУЛЬТАТЫ
Многофакторный дисперсионный анализ ранжированных совокупных результатов
первого опыта (n = 1334) показал, что доля диапаузирующих предкуколок T. telengai
существенно зависела как от фотопериодических условий развития материнского по-
коления (критерий Фишера F = 177.4, достоверность влияния p < 0.001), так и от тем-
пературы (F = 914.9, p < 0.001) и длительности (F = 146.5, p < 0.001) холодовой экспо-
зиции потомства, причем эффект попарного взаимодействия всех трех факторов также
был высоко достоверным (p < 0.001), хотя и не таким сильным (F < 50).
Для анализа индивидуальной изменчивости по доле диапаузирующего потомства
были использованы только самки, заразившие не менее 5 яиц хозяина (размеры вы-
борок указаны на рис. 1). Индивидуальная изменчивость оказалась статистически до-
стоверной в большинстве вариантов опыта, включая и те случаи, когда более 80 %
самок относились к одному из классов, т. е. подавляющее большинство особей потом-
ства развивались с диапаузой (например, рис. 1, Г, температура 12 °C) или без диапа-
узы (например, рис. 1, Г, температура 15 °C). С другой стороны, в некоторых случаях
(рис. 1, А, температура 9 °C и рис. 1, Б, температура 15 °C) различия между самками
были статистически недостоверны, хотя доля диапаузирующих особей среди совокуп-
ного потомства была далека и от 0, и от 100 %. Заметим, что распределение самок
по классам во всех вариантах опыта было унимодальным, т. е. гетерогенность, даже
если она была статистически достоверна, имела не качественный, а количественный
характер. В тех же случаях, когда средняя доля диапаузирующего потомства составля-
ла около 50 %, распределение самок по классам было близко к нормальному (рис. 1, А,
температура 6 °C, и рис. 1, В, температуры 9 и 12 °C).
Дисперсионный анализ результатов второго опыта также показал, что доля диапа-
узирующего потомства в хозяевах, зараженных во время первой (n = 703, F = 302.0,
p < 0.001), второй (n = 651, F = 283.8, p < 0.001) и третьей (n = 534, F = 256.4, p < 0.001)
экспозиций, зависела от фотопериодических условий развития предшествующего по-
коления. Индивидуальная изменчивость, для анализа которой, как и в первом опыте,
были использованы только самки, заразившие за время данной экспозиции не менее
5 яиц хозяина, также была весьма существенной (рис. 2). Заметим, что эта изменчи-
вость проявлялась даже в тех вариантах опыта, когда совокупная доля диапаузирующе-
10
го потомства была близка к 90 % (рис. 2, А), но только в хозяевах, зараженных во время
второй и третьей экспозиций и при наличии двухдневного интервала между ними. При
развитии материнского поколения в условиях околопорогового (рис. 2, Б) и длинного
(рис. 2, В) дня индивидуальная изменчивость имела больший размах, но по-прежнему
оставалась унимодальной (оценивая характер распределения, следует принимать во
внимание объем выборки).
Дальнейший анализ результатов второго опыта показал, что корреляция по доле ди-
апаузирующего потомства между порциям яиц хозяина, зараженными одной и той же
самкой во время трех последовательных экспозиций, в среднем была достоверно по-
ложительной: r = 0.125 ± 0.036 (здесь и в дальнейшем приведены средние арифмети-
ческие и их ошибки). На рис. 3 показаны коэффициенты корреляции между долями
диапаузирующих особей в потомстве, произведенном одной и той же самкой во время
трех последовательных экспозиций, подсчитанные отдельно для трех фотопериодиче-
ских режимов развития материнских самок и для вариантов с наличием и отсутствием
двухдневных интервалов между экспозициями. Видно, что большинство коэффици-
ентов корреляции (и все достоверные коэффициенты) положительны. Что же касается
влияния внешних факторов, то дисперсионный анализ (n = 18) показал, что интер-
валы времени между периодами заражения не влияли на силу корреляции (F = 0.06,
p = 0.806): средние коэффициенты корреляции при двухдневном интервале между
периодами заражения и при его отсутствии составили r = 0.134 ± 0.060 и r = 0.116
± 0.042 соответственно. Влияние фотопериода также было недостоверным (F = 0.20,
p = 0.818): при длинах дня 12, 16 и 18 ч средние коэффициенты корреляции соста-
вили, соответственно, r = 0.093 ± 0.055, r = 0.150 ± 0.080 и r = 0.132 ± 0.056. Зато
влияние порядкового номера экспозиции было на грани достоверности (F = 3.59,
p = 0.053): корреляция между второй и третьей порциями (r = 0.242 ± 0.079) была
заметно сильнее, чем между первой и второй (r = 0.079 ± 0.045) и первой и третьей
(r = 0.054 ± 0.021) порциями.
Плодовитость заражавших самок (точнее, число успешно зараженных ими хозяев)
убывала со временем в обоих вариантах второго опыта, хотя двухдневный интервал
между экспозициями несколько замедлял этот процесс (рис. 4). Корреляция между до-
лей диапаузирующего потомства и плодовитостью самки в среднем была отрицатель-
ной (r = -0.091 ± 0.030). На рис. 5 видно, что большинство коэффициентов корреля-
ции (и все статистически достоверные коэффициенты) также отрицательны. При этом
корреляция не зависела от наличия интервалов между периодами заражения (F = 0.28,
p = 0.605): средние коэффициенты корреляции при наличии двухдневного интер-
вала между периодами заражения и при его отсутствии составили, соответственно,
r = -0.107 ± 0.041 и r = -0.075 ± 0.044. Влияние фотопериода также было недостовер-
ным (F = 0.50, p = 0.614): при длинах дня 12, 16 и 18 ч средние коэффициенты корреля-
ции составили r = -0.131 ± 0.046, r = -0.085 ± 0.032 и r = -0.057 ± 0.072, соответственно.
Статистически недостоверными (F = 1.840, p = 0.193) были и различия между экспо-
зициями: средние коэффициенты корреляции между плодовитостью и долей диапау-
зирующего потомства, произведенного за время первой, второй и третьей экспозиций
составили r = -0.086 ± 0.038, r = -0.160 ± 0.062 и r = -0.028 ± 0.044, соответственно.
ОБСУЖДЕНИЕ
Влияние внешних факторов на индукцию диапаузы, выявленное в наших опытах,
ранее было неоднократно продемонстрировано как для T. telengai (Сорокина, Мас-
11
Рис. 2. Изменчивость доли диапаузирующего потомства в яйцах хозяина, зараженных самками
Trichogramma telengai Sor. во время трех последовательных экспозиций в зависимости
от фотопериодических условий развития материнского поколения и наличия временного
интервала между экспозициями (результаты второго опыта).
По горизонтальной оси - порядковый номер экспозиции (периода заражения хозяев). По вертикальной
оси - процентное распределение самок по классам, различающимся по доле диапаузирующего потомства
(границы классов указаны на рисунке). I - заражение во время трех последовательных 2-часовых
экспозиций в течение одного дня, II - заражение во время трех 2-часовых экспозиций, разделенных
2-дневными интервалами. Длина дня во время развития материнского поколения: А - 12 ч, Б - 16 ч,
В - 18 ч. Над каждой гистограммой указаны объемы выборок (n1 - число самок, n2 - число потомков) и
достоверность неоднородности выборки (достоверность различий между самками),
определенная с помощью теста хи-квадрат.
12
Рис. 2 (продолжение).
ленникова, 1986; Сорокина, 1987; Reznik et al., 2008, 2011; Vaghina et al., 2014), так и
для многих других видов трихограмм (Zaslavski, Umarova, 1990; Boivin, 1994; Резник,
2011). Хорошо известно, что материнская фотопериодическая реакция обеспечивает
очень точную регуляцию: даже небольшое уменьшение длины дня, при которой разви-
вались самки предшествующего поколения, приводит к соответствующему росту доли
диапаузирующих предкуколок. Эти изменения в совокупных выборках могут быть
объяснены двумя причинами: 1) при изменении фотопериода происходят соответству-
ющие градуальные изменения в потомстве каждой самки или 2) реакция самок дис-
кретна (т. е. они могут производить либо только активно развивающееся, либо только
диапаузирующее потомство), но доля самок, производящих диапаузирующее потом-
ство, градуально зависит от длины дня. Во всех цитированных выше работах долю
диапаузирующих особей определяли в относительно больших выборках, включающих
потомство десятков и сотен самок трихограмм, поэтому вопрос о характере реакции
отдельных особей оставался открытым. Данное исследование свидетельствует от том,
что реакция самок T. telengai градуальна: распределение отдельных особей по доле
диапаузирующего потомства унимодально, зачастую близко к нормальному. Достовер-
ной бимодальности распределения, которая могла бы свидетельствовать о дискретной
изменчивости, не выявлено. При этом размах индивидуальной изменчивости относи-
тельно велик и обнаружена она в большинстве выборок, включая и те, в которых сово-
купная доля диапаузирующего потомства близка к 90 %.
Самки Nasonia vitripennis (Walker) (Hymenoptera, Pteromalidae), как правило, произ-
водят либо только активно развивающееся, либо только диапаузирующее потомство,
т. е. их индивидуальная изменчивость по доле диапаузирующего потомства дискретна
(Saunders, 1965). Живородящие самки тли Megoura viciae Buckt. (Homoptera, Aphididae)
в условиях длинного дня производят таких же живородящих самок, а в условиях ко-
13
Рис. 3. Корреляция по доле диапаузирующего потомства между порциям яиц хозяина,
зараженными одной и той же самкой Trichogramma telengai Sor. во время трех
последовательных экспозиций в зависимости от фотопериодических условий развития
материнского поколения и наличия временного интервала между экспозициями
(результаты второго опыта).
Под горизонтальной осью - фотопериодические условия развития материнского поколения (С : Т).
По вертикальной оси - коэффициент корреляции Спирмена по доле диапаузирующего потомства между
двумя порциям яиц хозяина, зараженными одной и той же самкой. 1 - корреляция между первой и второй
порциями, 2 - корреляция между первой и третьей порциями, 3 - корреляция между второй и третьей
порциями. А - заражение во время трех последовательных 2-часовых экспозиций в течение одного дня,
Б - заражение во время трех 2-часовых экспозиций, разделенных 2-дневными интервалами.
Над столбцами гистограмм указаны объем выборки (число самок) и
достоверность
коэффициента корреляции: * - p < 0.05, ** - p < 0.01.
14
Рис. 4. Плодовитость (число зараженных яиц хозяина) заражавших самок
Trichogramma telengai Sor. во время трех последовательных экспозиций
в зависимости от наличия временного интервала между ними
(результаты второго опыта).
По горизонтальной оси - порядковый номер экспозиции (периода заражения хозяев). По вертикальной
оси - плодовитость самок (среднее арифметическое и среднее отклонение). I - заражение во время трех
последовательных 2-часовых экспозиций в течение одного дня, II - заражение во время трех 2-часовых
экспозиций, разделенных 2-дневными интервалами. Над каждым столбцом гистограммы указан
объем выборки (число заражавших самок).
роткого дня - яйцекладущих самок и самцов, но при околопороговых фотопериодах
(14-15 ч) почти все особи производят смешанное потомство (Lees, 1959), а самки
синей мясной мухи Calliphora vicina R.-D. (Diptera, Calliphoridae) производят преи-
мущественно смешанное потомство даже в условиях короткого (12-14 ч) и длинного
(16-20 ч) дня (Виноградова, Резник, 2000).
Известно, что тенденция к диапаузе и многие другие биологические параметры
видов рода Trichogramma могут в значительной степени зависеть от свойств хозяев,
в которых они развивались (Масленникова, Сорокина, 1986; Tauber et al., 1986; Boivin,
1994). В частности, у трихограмм было обнаружено влияние непостоянства «качества»
яиц хозяина на колебания доли диапаузирующих особей в последовательных поколе-
ниях лабораторных линий, развивающихся в константных фототермических условиях
(Войнович и др., 2013; Резник и др., 2015). Однако положительная корреляция между
долями диапаузирующих особей в потомстве, произведенном одной и той же самкой
при последовательном заражении трех разных порций яиц зерновой моли, говорит
о том, что отмеченная в наших опытах изменчивость тенденции к диапаузе (по крайней
мере отчасти) определяется индивидуальными особенностями самих паразитоидов,
а не их хозяев.
Опыты показали также, что корреляция по доле диапаузирующих особей между вто-
рой и третьей порциями зараженных яиц хозяина сильнее, чем между первой и второй
и между первой и третьей порциями. Вероятно, различия между самками, влияющие
15
Рис. 5. Корреляция между долей диапаузирующего потомства и плодовитостью
самки Trichogramma telengai Sor. в зависимости от порядкового номера экспозиции,
фотопериодических условий развития материнского поколения и наличия временного
интервала между экспозициями (результаты второго опыта).
Под горизонтальной осью - фотопериодические условия развития материнского поколения (С : Т).
По вертикальной оси - коэффициент корреляции Спирмена между долей диапаузирующего потомства и
плодовитостью самки. 1- первая экспозиция, 2 - вторая экспозиция, 3 - третья экспозиция. А - заражение
во время трех последовательных 2-часовых экспозиций в течение одного дня, Б - заражение во время трех
2-часовых экспозиций, разделенных 2-дневными интервалами. Над столбцами гистограмм указаны объем
выборки (число самок) и
достоверность коэффициента корреляции: * - p < 0.05, ** - p < 0.01.
16
на материнскую индукцию диапаузы, не наследуются, а приобретаются за время жиз-
ни имаго, хотя вполне возможно также, что генетически детерминированные индиви-
дуальные различия увеличиваются с возрастом, который и сам по себе может влиять на
долю диапаузирующего потомства (Виноградова, Резник, 2000).
Как уже упоминалось, изменчивость доли диапаузирующих особей среди потомства
самок, развивающихся в одних и тех же условиях, может объясняться как различиями
в силе материнской (в нашем случае - фотопериодической) индукции диапаузы, так
и различиями в прямой (в нашем случае - температурной) реакции личинок, причем
вполне вероятно и сочетание действия обоих факторов. Имеющиеся данные не позво-
ляют однозначно выбрать между этими альтернативами, но достоверная корреляция
между плодовитостью и долей диапаузирующего потомства свидетельствует о том, что
наблюдаемая изменчивость хотя бы отчасти обусловлена особенностями самок. Ме-
ханизм, лежащий в основе этой корреляции, пока неясен. Известно, что материнская
регуляция диапаузы эмбрионов шелкопряда Bombyx mori L. (Lepidoptera, Bombycidae)
обусловлена изменением содержания в яйце углеводов и других питательных веществ
(Yamashita, 1996; Denlinger, 2002). Возможно, размер самки трихограммы, в значитель-
ной мере определяющий ее плодовитость, влияет также и на размер или химический
состав откладываемых яиц. Как бы то ни было, индивидуальная изменчивость самок
по силе индукции диапаузы потомства, несомненно, заслуживает дальнейшего специ-
ального изучения и, кроме того, должна учитываться при разработке методов лабора-
торного исследования и массового разведения трихограмм.
БЛАГОДАРНОСТИ
За помощь в проведении экспериментов авторы глубоко признательны Т. Я. Умаро-
вой (Зоологический институт РАН). Работа осуществлена при финансовой поддержке
Программы фундаментальных исследований ОБН РАН «Биологические ресурсы Рос-
сии: оценка состояния и фундаментальные основы мониторинга», раздел программы
«Динамика и прогнозирование состояния важнейших видов ресурсов», регистрацион-
ный номер АААА-А18-118042390186-9.
СПИСОК ЛИТЕРАТУРЫ
Виноградова Е. Б., Резник С. Я. 2000. Влияние возраста самок на личиночную диапаузу потомства у си-
ней мясной мухи Calliphora vicina R.-D. (Diptera, Calliphorodae). Энтомологическое обозрение 79 (2):
296-302.
Войнович Н. Д., Резник С. Я., Вагина Н. П. 2013. Изменчивость «спонтанной» динамики тенденции к диапа-
узе в ряду поколений Trichogramma telengai Sor. Энтомологическое обозрение 92 (3): 465-479.
Данилевский А. С. 1961. Фотопериодизм и сезонное развитие насекомых. Л.: Издательство ЛГУ. 244 с.
Заславский В. А. 1984. Фотопериодический и температурный контроль развития насекомых. Л.: Наука. 180 с.
Заславский В. А., Умарова Т. Я. 1981. Фотопериодический и температурный контроль диапаузы у Tri-
chogramma evanescens Westw. (Hymenoptera, Trichogrammatidae). Энтомологическое обозрение 60 (4):
721-731.
Май Фу Кви, Заславский В. А. 1983. Фотопериодические и температурные реакции Trichogramma euproctidis
(Hymenoptera, Trichogrammatidae). Зоологический журнал 62 (11): 1676-1680.
Масленникова В. А., Сорокина А. П. 1986. Влияние физиологии хозяина на диапаузу и интенсивность за-
ражения Trichogramma cacoeciae March., T. evanescens Westw. и T. embryophagum Htg. (Hymenoptera,
Trichogrammatidae). Информационный бюллетень ВПС МОББ 14: 33-38.
Резник С. Я. 2011. Экологические и эволюционные аспекты фототермической регуляции диапаузы у трихо-
грамм. Журнал эволюционной биохимии и физиологии 47 (6): 434-443.
Резник С. Я., Вагина Н. П., Войнович Н. Д. 2015. Изменения тенденции к диапаузе в последовательных по-
колениях лабораторных линий трихограмм (Hymenoptera, Trichogrammatidae): эндогенные или экзо-
генные? Зоологический журнал 94 (4): 446-454.
Саулич А. Х. 1999. Сезонное развитие насекомых и возможности их расселения. СПб.: Издательство СПбГУ,
248 с.
17
Саулич А. Х., Волкович Т. А. 2004. Экология фотопериодизма насекомых. СПб.: Издательство СПбГУ. 276 с.
Сорокина А. П. 1987. Биологическое и морфологическое обоснование видовой самостоятельности
Trichogramma telengai sp. n.(Hymenoptera, Trichogrammatidae). Энтомологическое обозрение 66 (1):
32-46.
Сорокина А. П. 2010. Фототермические реакции, контролирующие диапаузу у трех видов трихограммы
(Hymenoptera, Trichogrammatidae) из Ленинградской области. Вестник защиты растений 3: 51-54.
Сорокина А. П. 2011. Применение трихограммы: прошлое и настоящее. Защита и карантин растений 10:
9-12.
Сорокина А. П., Масленникова В. А. 1986. Особенности фототермических реакций некоторых видов рода
Trichogramma (Hymenoptera, Trichogrammatidae). Вестник Ленинградского государственного универ-
ситета. Сер. 3. 1: 9-14.
Сорокина А. П., Масленникова В. А. 1987. Температурный оптимум формирования диапаузы у видов рода
Trichogramma Westw. (Hymenoptera, Trichogrammatidae). Энтомологическое обозрение 66 (4): 689-699.
Boivin G. 1994. Overwintering strategies of egg parasitoids. In: E. Wajnberg, S. A. Hassan (eds). Biological Control
with Egg Parasitoids. Vallingford, UK: CAB International, pp. 219-244.
Danks H. V. 2007. The elements of seasonal adaptations in insects. The Canadian Entomologist 139 (1): 1-44.
Denlinger D. L. 2002. Regulation of diapause. Annual Review of Entomology 47: 93-122.
Goto S. G., Numata H. 2014. Insect photoperiodism. In: K. H. Hoffmann (ed.). Insect Molecular Biology and
Ecology. Boca Raton, Florida, US: CRC Press, pp. 217-244.
Lees A. D. 1959. The role of photoperiod and temperature in the determination of parthenogenetic and sexual forms
in the aphid Megoura viciae Buckton. - I: The influence of these factors on apterous virginoparae and their
progeny. Journal of Insect Physiology 3 (1): 92-117.
Mousseau T. A., Dingle H. 1991. Maternal effects in insect life histories. Annual Review of Entomology 36:
511-534.
Reznik S. Ya., Vaghina N. P., Voinovich N. D. 2008. Diapause induction in Trichogramma embryophagum Htg.
(Hymenoptera, Trichogrammatidae): the dynamics of thermosensitivity. Journal of Applied Entomology
132 (6): 502-509.
Reznik S. Ya., Vaghina N. P., Voinovich N. D. 2011. Maternal influence on diapause induction in Trichogramma
(Hymenoptera, Trichogrammatidae): the dynamics of photosensitivity. Journal of Applied Entomology
135 (6): 438-445.
Saunders D. S. 1965. Larval diapause of maternal origin: induction of diapause in Nasonia vitripennis (Walk.)
(Hymenoptera: Pteromalidae). Journal of Experimental Biology 42 (3): 495-508.
Saunders D. S. 2010. Photoperiodism in insects: migration and diapause responses. In: R. J. Nelson, D. L. Denlinger,
D. E. Somers (eds). Photoperiodism: the Biological Calendar. Oxford, USA: Oxford University Press,
pp. 218-257.
Saunders D. S., Steel C. G. H., Vafopoulou X., Lewis R. D. 2002. Insect Clocks. Amsterdam: Elsevier, 560 p.
Smith S. M. 1996. Biological control with Trichogramma: advances, successes, and potential of their use. Annual
Review of Entomology 41: 375-406.
Tauber M. J., Tauber C. A., Masaki S. 1986. Seasonal Adaptations of Insects. New York: Oxford University Press,
411 p.
Vaghina N. P., Voinovich N. D., Reznik S. Ya. 2014. Maternal thermal and photoperiodic effects on the progeny
diapause in Trichogramma telengai Sorokina (Hymenoptera: Trichogrammatidae). Entomological Science.
17 (2): 198-206.
Yamashita O. 1996. Diapause hormone of the silkworm, Bombyx mori: structure, gene expression and function.
Journal of Insect Physiology 42 (7): 669-679.
Zaslavski V. A., Umarova T. Ya. 1990. Environmental and endogenous control of diapause in Trichogramma species.
Entomophaga 35 (1): 23-29.
INDIVIDUAL VARIATION IN PROGENY DIAPAUSE INDUCTION
IN TRICHOGRAMMA TELENGAI SOR. FEMALES (HYMENOPTERA,
TRICHOGRAMMATIDAE)
S. Ya. Reznik, N. D. Voinovich
Key words: diapause, variation, temperature, photoperiod, Trichogramma telengai.
SUMMARY
Individual variations in the proportion of diapausing progeny of females from a laboratory
parthernogenetic strain of Trichogramma telengai Sor. were investigated. The maternal generation
18
developed at a temperature of 20 °С and day length of 12, 16, and 18 ч. The progeny generation was
incubated at the low temperature conditions (6, 9, 12, and 15 °C) during 15 or 30 days. Individual
variations were statistically significant in most treatments of the experiment, including also the cases
when the average proportion of diapausing progeny was close to 90 %. The distribution of females
by the proportion of diapausing progeny was unimodal. When the average proportion of diapausing
progeny was about 50 %, the distribution was close to normal, i. e. the individual variation was rather
quantitative (gradual) than qualitative. The correlation in the proportion of diapausing individuals
between the progenies produced by the same female during sequential parasitization of different
portions of the grain moth eggs was significantly positive suggesting that the variability in the tendency
to diapause was (at least partly) determined by individual characteristics of Trichogramma wasps.
Individual variation of females in the proportion of diapausing progeny should be accounted for in the
development of the methods for laboratory investigation and mass rearing of Trichogramma species.
19