ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 98, 1, 2019
УДК 595.794:591.563 (477.75)
ЖАЛЯЩИЕ ПЕРЕПОНЧАТОКРЫЛЫЕ (HYMENOPTERA,
ACULEATA), ЗАСЕЛЯЮЩИЕ ГНЕЗДА-ЛОВУШКИ В КРЫМУ
© 2019 г. С. П. Иванов,1* А. В. Фатерыга,2** В. Ю. Жидков3, 4***
1 Таврическая академия Крымского федерального университета им. В. И. Вернадского
пр. Академика Вернадского, 4, Симферополь, 295007 Россия
*e-mail: spi2006@list.ru
2 Карадагская научная станция им. Т. И. Вяземского - природный заповедник РАН
ул. Науки, 24, пгт Курортное, Феодосия, 298188 Россия
**e-mail: fater_84@list.ru
3 Управление особо охраняемыми территориями Республики Крым
ул. Киевская, 77/4, Симферополь, 295034 Россия
***e-mail: aravar@list.ru
4 Национальный природный парк «Тарханкутский»
ул. Кирова, 43, пгт Черноморское, 296400 Россия
Поступила в редакцию 6.04.2018 г.
После доработки 25.01.2019 г.
Принята к публикации 25.01.2019 г.
В течение 15 лет (2002-2016 гг.) в 80 пунктах Крыма выставляли гнезда-ловушки для жа-
лящих перепончатокрылых. Всего было использовано более 500 гнезд-ловушек, содержащих
около 25 000 полостей. В результате было получено 6 895 гнезд перепончатокрылых 97 видов из
9 семейств: Pompilidae (5 видов / 118 гнезд), Vespidae (27 / 1201), Ampulicidae (1 / 6), Sphecidae
(2 / 8), Crabronidae (10 / 591), Colletidae (7 / 202), Megachilidae (37 / 4619), Apidae (3 / 9) и For-
micidae (5 / 141). Обсуждаются интенсивность заселения гнезд-ловушек разными видами и
группами жалящих перепончатокрылых, привлекательность гнезд-ловушек для этих насекомых
в связи с особенностями их биологии (строение гнезд, выбор субстрата гнездования, число поко-
лений в году, трофические связи и филопатрия). Выявлена сильная положительная связь между
интенсивностью заселения гнезд-ловушек и количеством осадков, выпавших в первой полови-
не соответствующего сезона в данной местности. Сообщается о гнездах некоторых необычных
поселенцев гнезд-ловушек, в том числе Eumenes papillarius (Christ) и Eumenes sp. Сравнивается
число полученных гнезд представителей сем. Megachilidae с числом коллекционных экземпля-
ров соответствующих видов; корреляция между этими показателями существует, но выражена
довольно слабо. Два вида ос и 9 видов пчел оценены как перспективные для искусственного
разведения.
Ключевые слова: гнезда-ловушки, ульи Фабра, Pompilidae, Vespidae, Ampulicidae, Sphecidae,
Crabronidae, Colletidae, Megachilidae, Apidae, Formicidae, Крым.
DOI: 10.1134/S0367144519010076
Гнезда-ловушки для изучения жалящих перепончатокрылых (прежде всего одиноч-
ных ос и пчел) впервые применил Ж.-А. Фабр (см.: Фабр, 1993), используя деревян-
70
ные ящики с вставленными в них стеблями тростника. В честь этого выдающегося
энтомолога Малышев (1963) предложил назвать такие конструкции «ульями Фабра».
В настоящее время в качестве гнезд-ловушек, как правило, используют более простые
конструкции. Обычно они представляют собой связки отрезков пустотелых стеблей
растений и трубочек из различных материалов, деревянные бруски с просверленны-
ми в них отверстиями (Krombein, 1967). Ульи Фабра чаще используются для искус-
ственного разведения одиночных пчел, но и как гнезда-ловушки они также по-преж-
нему применяются (Иванов и др., 2009). Разнообразные модификации гнезд-ловушек
(Collins, Jennings, 1984; Potts et al., 2005; Taki et al., 2008) применяют для выявления
видового состава отдельных групп одиночных жалящих перепончатокрылых, обитаю-
щих на определенной территории (Budrys et al., 2009), оценки состояния экосистем
(Tscharntke et al., 1998; Klein et al., 2002; Kruess, Tscharntke, 2002, и др.), а в отдельных
случаях для увеличения разнообразия, сохранения и поддержания популяций одиноч-
ных ос и пчел в местах их естественного обитания (Иванов, 1984; Мариковская и др.,
2001; Иванов и др., 2005; Gaston et al., 2005, и др.). Однако чаще всего гнезда-ловушки
используют для получения гнезд одиночных ос и пчел с целью изучения биологии
гнездования этих насекомых. Опубликована не одна сотня работ, в которых описаны
гнезда разных видов, полученные из гнезд-ловушек. Прежде всего, это фундаменталь-
ная монография Кромбайна (Krombein, 1967). Гнезда-ловушки применяют также для
выявления видового состава паразитов, связанных с гнездящимися в них видами ос и
пчел (Parker, Bohart, 1966, 1968; Мартынова, Фатерыга, 2015; Pärn et al., 2015, и др.).
Из гнезд-ловушек было выведено и описано несколько новых для науки видов (Parker,
1976, 1977; Griswold, 2001). Кроме того, гнезда-ловушки находят применение в сель-
ском и лесном хозяйстве для поддержания популяций опылителей и энтомофагов -
агентов биологического контроля фитофагов (Мариковская, Щербакова, 1989; Wearing,
Harris, 1999, 2005, и др.).
Цель настоящей работы - выявить видовой состав жалящих перепончатокрылых,
заселяющих гнезда-ловушки в Крыму, оценить влияние факторов среды на интенсив-
ность их заселения, установить связь между биологическими особенностями видов и
степенью привлекательности для них гнезд-ловушек, а также выявить виды, перспек-
тивные для искусственного разведения.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили в Крыму в 2002-2016 гг. Гнезда-ловушки выставляли во
всех ландшафтных зонах полуострова (по: Выработка приоритетов, 1999): редколесья
южнобережья - 12 пунктов установки; леса южного макросклона - 3, горные луга и
степи яйл - 4; леса северного макросклона - 5; предгорные лесостепи - 16, настоящие
степи - 19, полупустынные степи и солончаки - 21 (рис. 1).
В качестве гнезд-ловушек в большинстве случаев использовали связки полых сте-
блей тростника [Phragmites australis (Cav.) Trin. ex Steud. (Poaceae)] (рис. 2), реже -
ульи Фабра различной конструкции. Последние чаще всего представляли собой ящи-
ки, заполненные связками стеблей тростника (рис. 3). В некоторых ульях Фабра вместе
со связками стеблей тростника использовали такие же стебли, вставленные в верти-
кальные щитки, а также стебли растений с мягкой сердцевиной и гнездовые блоки
из деревянных желобчатых пластин (рис. 4). Такая конструкция обеспечивала пригод-
ность улья для заселения большим числом видов ос и пчел (см.: Иванов и др., 2009).
71
Наиболее частое применение нашли гнезда-ловушки в виде стандартных связок из
20-35 отрезков стеблей тростника длиной 15-25 см и диаметром 4-12 мм, обернутые
плотной бумагой и поверх (при необходимости в момент установки) - полиэтиленовой
пленкой (рис. 5).
Стебли тростника заготавливали зимой на одном из водохранилищ Крыма после
того, как лед достигал достаточной толщины (рис. 6). С помощью совковой лопаты
стебли срубали на уровне поверхности льда, связывали в большие охапки, перемеща-
ли на сушу и оставляли до наступления положительной температуры воздуха. Перед
нарезкой стебли смачивали водой. Затем их разрезали на отдельные отрезки острым
ножом (косыми срезами) таким образом, чтобы полученные отрезки с одного конца
были открыты, а с другого - закрыты узлом, над которым делался срез. В результа-
те «дно» глухого конца отрезка стебля получалось правильной закругленно-вогнутой
формы. Если срез делать под узлом, то «дно» полости стебля будет выпуклым и менее
привлекательным, как для ос, так и для пчел. Полученные отрезки стеблей связывали в
пучки с помощью полосок эластичной ткани, нарезанных из колготок. Это обеспечива-
ло прочность связок в условиях меняющейся влажности воздуха - набухания стеблей
во влажную погоду и последующего высыхания в сухую.
В большинстве из 80 отмеченных на карте пунктов (см. рис. 1) гнезда-ловушки уста-
навливали однократно, но в некоторых местах их выставляли в течение 2, 3 и боль-
шего числа последовательных сезонов. Наибольшее число раз их устанавливали в
Карадагском природном заповеднике - 12 сезонов, в том числе 8 сезонов подряд (2009-
2016 гг.). В первые годы исследований (2002-2009 гг.) выставление гнезд-ловушек в
Крыму проводили с охватом как можно большего числа пунктов при небольшом числе
конструкций в каждом из них. Такая тактика позволила выявить наиболее подходящие
для целей исследования биотопы и отработать особенности методики. Наибольшее
число пунктов установки гнезд-ловушек было использовано в 2009 г. - 44. В последу-
ющие годы число пунктов их ежегодной установки было уменьшено до 15-25. Всего за
время исследований было выставлено более 500 гнезд-ловушек, содержащих в сумме
около 25 000 стеблей тростника.
Гнезда-ловушки устанавливали в местах естественного обитания ос и пчел
(рис. 7-12), выбирая места с богатой, прежде всего энтомофильной растительностью.
В лесу это были опушки вдоль больших полян или 2-3-летние горельники (рис. 7),
в степи - участки с выходами скал и балками, заросшими кустарником (рис. 9), на
морском побережье - участки вдоль склонов с хорошо сохранившейся естественной
растительностью (рис. 11). Связки стеблей тростника прикрепляли к веткам деревьев
или кустарников, стволам деревьев (рис. 8), помещали в щелях скал или среди камней
(рис. 10), привязывали к корягам и стволам плавника на берегу моря (рис. 12), вставля-
ли в отверстия в лессовых обрывах, закрепляли на стенах строений или под крышами
навесов (см. рис. 2). Во всех случаях гнезда-ловушки старались устанавливать так,
чтобы обеспечить одновременно и скрытность от глаз человека, и доступность для ос
и пчел; только таким образом удавалось обеспечить их сохранность и избежать разру-
шения местными жителями или туристами. Ульи Фабра, используемые как гнезда-ло-
вушки, устанавливали только на заповедных и других охраняемых территориях. Разме-
щая гнезда-ловушки в заповедниках, старались обеспечить их защиту от повреждения
дикими животными, в частности кабанами.
Гнезда-ловушки устанавливали ранней весной. Опыт первых лет показал, что
конструкции, установленные в начале апреля, в некоторых пунктах уже к середине
72
мая часто оказывались полностью заселенными одним видом пчел - Osmia bicornis
(Linnaeus), поэтому в дальнейшем большую часть гнезд-ловушек в таких пунктах уста-
навливали позднее - в конце апреля или в начале мая. В конце сезона гнезда-ловушки
разбирали на отдельные стебли тростника, заселенные стебли отделяли от незаселен-
ных и подсчитывали.
Отобранные стебли с гнездами вскрывали, раскалывая тростинку с помощью ножа.
Каждое вскрытое гнездо целиком помещали в стеклянную трубку, закрывали с обеих
сторон плотными ватными тампонами и снабжали этикеткой с номером гнезда-ловуш-
ки и сведениями о пункте ее установки. При необходимости гнезда разбирали на от-
дельные ячейки для более подробного изучения. Ячейки разобранных гнезд помещали
отдельно в короткие отрезки стеклянных трубок или небольшие пробирки. Иденти-
фикацию видов, заселивших гнезда-ловушки, обычно проводили на следующий год,
после выхода имаго из ячеек гнезд и коллектирования части особей (по 1 самке и
1 самцу из каждого гнезда). Большую часть вышедших из гнезд ос и пчел отпускали.
В отдельных случаях видовую принадлежность гнездящихся насекомых устанавлива-
ли уже в период их гнездования, в ходе наблюдений за самками, строящими и прови-
антирующими гнезда (рис. 13-16). Иногда из гнезд-ловушек извлекали свежезапеча-
танные гнезда для попутного изучения формы и консистенции хлебца, положения и
состава жертв, способов прикрепления яйца и других особенностей строения и состава
гнезд, которые можно выявить только при исследовании свежих гнезд.
В отдельных случаях гнезда перепончатокрылых были обнаружены в гнездах-ло-
вушках между стеблями тростника, под оберткой связки стеблей, или в полости кор-
пуса улья Фабра. Эти гнезда прибавляли к общему числу заселенных стеблей. Для
бивольтинных видов учитывали гнезда обоих поколений, поскольку довольно часто
наблюдалось гнездование только одного из них. Заселение гнезд-ловушек некоторыми
видами ос и пчел [Ancistrocerus nigricornis (Curtis), Ceratina cyanea (Kirby) и Xylocopa
valga (Gerstaecker)] можно было установить только в ходе наблюдений, проведенных в
период их гнездования. Это связано с тем, что молодые особи данных видов покидают
материнские гнезда после отрождения и очень редко остаются в них на зимовку.
Привлекательность гнезд-ловушек для представителей отдельных видов оцени-
вали по общему числу гнезд данного вида, обнаруженных во всех пунктах установ-
ки гнезд-ловушек в Крыму за все время исследований. Интенсивность заселения
гнезд-ловушек, установленных в отдельных местностях Крыма в отдельные годы, оце-
нивали по процентной доле заселенных полостей от общего числа полостей во всех
гнездах-ловушках, установленных в данном пункте в этот год. Это связано с необходи-
мостью нивелировать влияние различий в числе гнездовых конструкций, выставлен-
ных в разных пунктах в разные годы, и их различий по числу полостей.
Была проведена оценка возможности искусственного разведения некоторых видов
перепончатокрылых. В тех случаях, когда после разбора гнезд-ловушек на отдельные
гнезда, их идентификации и изучения оставался запас невскрытых гнезд идентифи-
цированных видов, такие гнезда помещали в ульи Фабра и после окончания сезона
гнездования производили подсчет вторично заселенных материнских гнезд и числа
вновь построенных. Виды, повторно заселившие ульи, оценивали как перспективные
для искусственного разведения.
73
В работе использованы коэффициенты корреляции Спирмена (rS) (Лакин, 1990) и
выравненности видов по обилию (Песенко, Боголюбов, 1979). Данные по количеству
осадков в Карадагском природном заповеднике получены на станции фонового эколо-
гического мониторинга Карадагской научной станции им. Т. И. Вяземского - природ-
ного заповедника РАН.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Видовой состав жалящих перепончатокрылых, заселивших гнезда-ловушки
За 15 лет исследований в Крыму гнезда-ловушки заселили 97 видов жалящих пере-
пончатокрылых из 39 родов 9 семейств (табл. 1). Среди них 45 видов ос (семейства
Pompilidae, Vespidae, Ampulicidae, Sphecidae и Crabronidae), 47 видов пчел (семейства
Colletidae, Megachilidae и Apidae) и 5 видов муравьев (сем. Formicidae). В общей слож-
ности из гнезд-ловушек было получено 6 895 гнезд жалящих перепончатокрылых (для
муравьев за число гнезд принимали число заселенных полостей).
Обращает на себя внимание большой разброс числа полученных гнезд как между
разными видами одного семейства, так и между представителями разных семейств
в целом. По соотношению число гнезд / число видов лидирует сем. Megachilidae (в сред-
нем 125 гнезд на один вид). Далее следуют Crabronidae (59), Vespidae (44), Colletidae
(29), Formicidae (28) и Pompilidae (24). При этом сем. Crabronidae представлено глав-
ным образом гнездами одного рода Trypoxylon, сем. Colletidae также представлено
одним родом Hylaeus, a среди муравьев в гнездах-ловушках преобладает один вид
Crematogaster schmidti (Mayr). Три остальных семейства (Ampulicidae, Sphecidae и
Apidae) имеют значительно меньшие значения показателя «число гнезд / число видов».
Это свидетельствует о том, что тип укрытий, имитируемых гнездами-ловушками,
в природе занимают главным образом именно пчелы-мегахилиды. Впрочем, отдель-
ные представители сем. Vespidae также построили большое число гнезд в гнездах-ло-
вушках. Среди них Syneuodynerus egregius (Herrich-Schäffer), Euodynerus posticus
(Herrich-Schäffer), Ancistrocerus antilope (Panzer) и A. nigricornis, намного опережаю-
щие по числу гнезд другие виды этого семейства. Среди Crabronidae помимо видов
рода Trypoxylon в большом числе обнаружены гнезда Psenulus fuscipennis (Dahlbom).
Можно с уверенностью заключить, что все эти виды, а также представители
сем. Megachilidae с числом гнезд больше 100 и в природе выбирают для строитель-
ства своих гнезд полости, сходные с полостями в гнездах-ловушках. Наш опыт обна-
ружения гнезд данных видов в природе в полной мере подтверждает это заключение.
С другой стороны, небольшое число полученных гнезд некоторых видов необязатель-
но свидетельствует о «неприятии» ими гнезд-ловушек и может быть связано, напри-
мер, с редкостью этих видов в природе (см. ниже).
Следует особо отметить случаи заселения гнезд-ловушек видами, которые в нор-
ме не заселяют готовые полости, а строят свободные полнокомпонентные ячейки
на поверхности субстрата, либо самостоятельно выгрызают гнездовой канал в древе-
сине. К первым относятся осы рода Eumenes из сем. Vespidae и Sceliphron curvatum
(F. Smith) из сем. Sphecidae. Нами получены гнезда Eumenes papillarius (Christ) из
ульев Фабра, установленных в Симферополе и Карадагском природном заповеднике
(рис. 17, 18), и Eumenes sp. из связок стеблей тростника, установленных в националь-
ном природном парке «Тарханкутский» (рис. 19). Гнезда первого вида размещались,
помимо полостей стеблей тростника, также и на поверхности связки этих стеблей
74
Таблица 1. Таксономический состав жалящих перепончатокрылых, заселивших гнезда-ло-
вушки в Крыму в 2002-2016 гг., и число полученных гнезд
Семейство
Вид
Число гнезд
(число видов/гнезд)
Pompilidae (5/118)
Auplopus carbonarius (Scopoli, 1763)
21
Agenioideus cinctellus (Spinola, 1808)
17
Dipogon bifasciatus (Geoffroy, 1785)
29
D. subintermedius (Magretti, 1886)
49
D. variegatus (Linnaeus, 1758)
2
Vespidae (27/1201)
Discoelius dufourii Lepeletier de Saint-Fargeau, 1841
26
D. zonalis (Panzer, 1801)
10
Gymnomerus laevipes (Shuckard, 1837)
26
Alastor mocsaryi (André, 1884)
10
Leptochilus regulus (de Saussure, 1855)
2
Syneuodynerus egregius (Herrich-Schäffer, 1839)
244
Euodynerus dantici (Rossi, 1790)
19
Eu. disconotatus (Lichtenstein, 1884)
56
Eu. fastidiosus (de Saussure, 1853)
9
Eu. posticus (Herrich-Schäffer, 1841)
253
Eu. quadrifasciatus (Fabricius, 1793)
11
Eu. velutinus Blüthgen, 1951
6
Stenodynerus bluethgeni van der Vecht, 1971
10
S. chevrieranus (de Saussure, 1855)
3
S. steckianus (Schulthess, 1897)
3
Allodynerus rossii (Lepeletier de Saint-Fargeau, 1841)
6
Parodontodynerus ephippium (Klug, 1817)
2
Symmorphus bifasciatus (Linnaeus, 1761)
1
S. crassicornis (Panzer, 1798)
2
Ancistrocerus antilope (Panzer, 1798)
185
A. auctus (Fabricius, 1793)
17
A. gazella (Panzer, 1798)
53
A. nigricornis (Curtis, 1826)
161
A. parietinus (Linnaeus, 1761)
60
A. trifasciatus (Müller, 1776)
16
Eumenes papillarius (Christ, 1791)
6
Eumenes sp.
4
Ampulicidae (1/6)
Ampulex fasciata Jurine, 1807
6
Sphecidae (2/8)
Sceliphron curvatum (F. Smith, 1870)
5
Isodontia mexicana (de Saussure, 1867)
3
Crabronidae (10/591)
Psenulus fuscipennis (Dahlbom, 1843)
117
Ps. pallipes (Panzer, 1798)
8
Pison atrum (Spinola, 1808)
23
Trypoxylon deceptorium Antropov, 1991
33
T. figulus (Linnaeus, 1758)
338
T. kolazyi Kohl, 1893
3
T. medium de Beaumont, 1945
6
T. scutatum Chevrier, 1867
50
Solierella compedita (Piccioli, 1869)
8
Crossocerus vagabundus (Panzer, 1798)
5
75
Таблица 1 (продолжение)
Семейство
Вид
Число гнезд
(число видов/гнезд)
Colletidae (7/202)
Hylaeus annularis (Kirby, 1802)
1
H. communis Nylander, 1852
4
H. gibbus S. Saunders, 1850
163
H. gracilicornis (Morawitz, 1867)
5
H. leptocephalus (Morawitz, 1871)
4
H. punctulatissimus Smith, 1842
2
H. signatus (Panzer, 1798)
23
Megachilidae
Lithurgus chrysurus Fonscolombe, 1834
26
(37/4619)
L. cornutus (Fabricius, 1787)
5
Anthidium florentinum (Fabricius, 1775)
20
Anthidium manicatum (Linnaeus, 1758)
132
A. oblongatum (Illiger, 1806)
2
Pseudoanthidium nanum (Mocsáry, 1881)
48
Ps. sp. aff. nanum (Mocsáry, 1881)
12
Ps. tenellum (Mocsáry, 1881)
1
Chelostoma mocsaryi Schletterer, 1889
116
Ch. rapunculi (Lepeletier de Saint-Fargeau, 1841)
5
Heriades crenulata Nylander, 1856
9
H. truncorum (Linnaeus, 1758)
5
Protosmia tiflensis (Morawitz, 1876)
2
Hoplitis manicata Morice, 1901
66
H. tridentata (Dufour et Perris, 1840)
10
Osmia bicornis (Linnaeus, 1758)
1028
O. bidentata (Morawitz, 1876)
46
O. brevicornis (Fabricius, 1798)
55
O. caerulescens (Linnaeus, 1758)
1569
O. cerinthidis Morawitz, 1876
14
O. cornuta (Latreille, 1805)
17
O. dimidiata (Morawitz, 1870)
348
O. leaiana (Kirby, 1802)
9
O. melanogaster Spinola, 1808
4
O. niveata (Fabricius, 1804)
11
O. signata Erichson, 1835
143
Megachile apicalis Spinola, 1808
552
M. centuncularis (Linnaeus, 1758)
160
M. ericetorum Lepeletier de Saint-Fargeau, 1841
28
M. lefebvrei Lepeletier de Saint-Fargeau, 1841
1
M. maritima (Kirby, 1802)
9
M. octosignata Nylander, 1852
1
M. pilicrus Morawitz, 1878
30
M. pilidens Alfken, 1924
5
M. rotundata (Fabricius, 1787)
26
M. versicolor Smith, 1844
102
M. willughbiella (Kirby, 1802)
2
76
Таблица 1 (продолжение)
Семейство
Вид
Число гнезд
(число видов/гнезд)
Apidae (3/9)
Xylocopa valga (Gerstaecker, 1872)
4
Ceratina cyanea (Kirby, 1802)
2
Anthophora furcata (Panzer, 1798)
3
Formicidae (5/141)
Camponotus fallax (Nylander, 1856)
1
C. lateralis (Olivier, 1792)
3
Crematogaster schmidti (Mayr, 1853)
121
Temnothorax tauricus Ruzsky, 1902
12
T. unifasciatus (Latreille, 1798)
4
П р и м е ч а н и е . Для сем. Formicidae за число гнезд принимали число заселенных полостей.
(рис. 17). Гнезда второго вида были найдены только в полостях стеблей. Опреде-
лить второй вид не удалось, так как все его потомство, находящееся в ячейках гнезд,
погибло. Однако можно с большой долей вероятности предположить, что это был
либо Eumenes dubius de Saussure, либо Eu. sareptanus André. Помимо этих двух ви-
дов на территории парка отмечен еще только Eu. coarctatus (Linnaeus), который стро-
ит более мелкие и шарообразные ячейки, обычно прилепляя их к стеблям растений
(см.: Фатерыга, Жидков, 2012). Гнезда ос рода Eumenes, построенные в цилиндриче-
ских полостях, ранее были отмечены только для Eu. micado Cameron (= Eu. samuray
von Schulthess), который может также сооружать их и на поверхности субстрата
(см.: Iwata, 1976).
Гнезда Sceliphron curvatum состоят из отдельных бутылкообразных ячеек, чаще все-
го расположенных группами, в которых ячейки скреплены друг с другом боковыми
сторонами. В природе они располагаются в различных укрытиях (Фатерыга, Ковблюк,
2013). В гнездах-ловушках ячейки этого вида отмечены нами как в полостях стеблей
тростника, где они располагались одна за другой в линию, так и под крышей корпуса
улья Фабра, где ячейки располагались, как обычно, группами.
К видам, самостоятельно выгрызающим гнездовой канал в древесине, но способ-
ным также заселять и готовые полости подходящих параметров, относятся отмечен-
ные нами в гнездах-ловушках Crossocerus vagabundus (Panzer) из сем. Crabronidae,
Lithurgus chrysurus Fonscolombe, L. cornutus (Fabricius), а также Heriades truncorum
(Linnaeus) из сем. Megachilidae и, кроме того, все 3 вида сем. Apidae: Ceratina cyanea,
Xylocopa valga и Anthophora furcata (Panzer). Все остальные осы и пчелы, зарегистри-
рованные в гнездах-ловушках в Крыму, в норме заселяют готовые полости.
Для некоторых видов сем. Megachilidae, отмеченных нами в гнездах-ловушках, ха-
рактерно гнездование в полостях, расположенных в земле или другом субстрате, не
сходном со стеблями тростника (см. ниже).
Интенсивность заселения гнезд-ловушек и оценка перспективности видов
в отношении возможностей их искусственного разведения
Доля полостей, заселенных разными группами жалящих перепончатокрылых, как и
общий процент заселения гнезд-ловушек, сильно варьировали в разных пунктах иссле-
дований и даже в пределах одного пункта в разные годы (табл. 2). Эти данные свиде-
77
Таблица 2. Заселение гнезд-ловушек разными группами жалящих перепончатокрылых в от-
дельных местностях Крыма в разные годы
Доля полостей, заселенных разными
группами жалящих перепончатокрылых, %
Местность
Год
Ленинский р-н, Опукский
2005
26
650
2.2
2.8
0.2
25.8
0.0
0.0
30.9
природный заповедник
2011
14
378
0.3
0.0
0.0
26.7
0.0
0.0
27.0
Бахчисарайский р-н,
2011
25
625
0.3
8.0
5.9
11.4
0.2
1.4
27.2
яблоневые сады
Симферопольский р-н,
2009
6
210
1.0
0.0
1.0
8.1
0.0
0.0
10.0
окр. с. Краснолесье
2010
5
175
24.0
0.6
0.0
14.9
0.0
0.0
39.4
2011
4
120
6.7
20.8
0.0
12.5
0.0
0.0
40.0
Окр. г. Ялта, Никитский
2011
19
457
7.4
0.0
2.3
2.3
17.3
1.5
30.7
ботанический сад
Окр. г. Феодосия,
2009
4
2280
0.1
0.0
0.2
5.3
0.0
2.4
7.9
Карадагский природный
2010
8
2607
0.5
0.0
0.0
28.1
0.0
1.5
30.0
заповедник
2011
14
2257
1.3
0.1
0.0
9.9
0.0
0.1
11.3
2012
7
788
0.9
0.0
0.0
0.3
0.0
0.0
1.1
2013
19
1814
0.8
0.4
0.0
3.7
0.0
0.4
5.4
2014
19
1105
1.4
0.0
0.0
2.0
0.2
0.2
3.7
2015
9
1432
2.5
0.7
0.3
24.9
0.0
0.3
28.7
2016
12
573
1.9
0.9
0.2
55.8
0.0
0.7
59.5
тельствуют о том, что на заселение гнезд-ловушек оказывают существенное влияние
характерные условия местности и погода.
Наблюдения показали, что более интенсивное заселение гнезд-ловушек в отдельных
пунктах в некоторые годы происходит на фоне массового цветения энтомофильных
растений, большинство из которых служит кормовой базой для пчел, заселяющих гнез-
да-ловушки. Хорошее состояние растительности, видимо, благоприятно сказывается
и на численности личинок насекомых-фитофагов - объектов охоты ос сем. Vespidae.
Основным фактором, определяющим (а часто лимитирующим) рост и развитие рас-
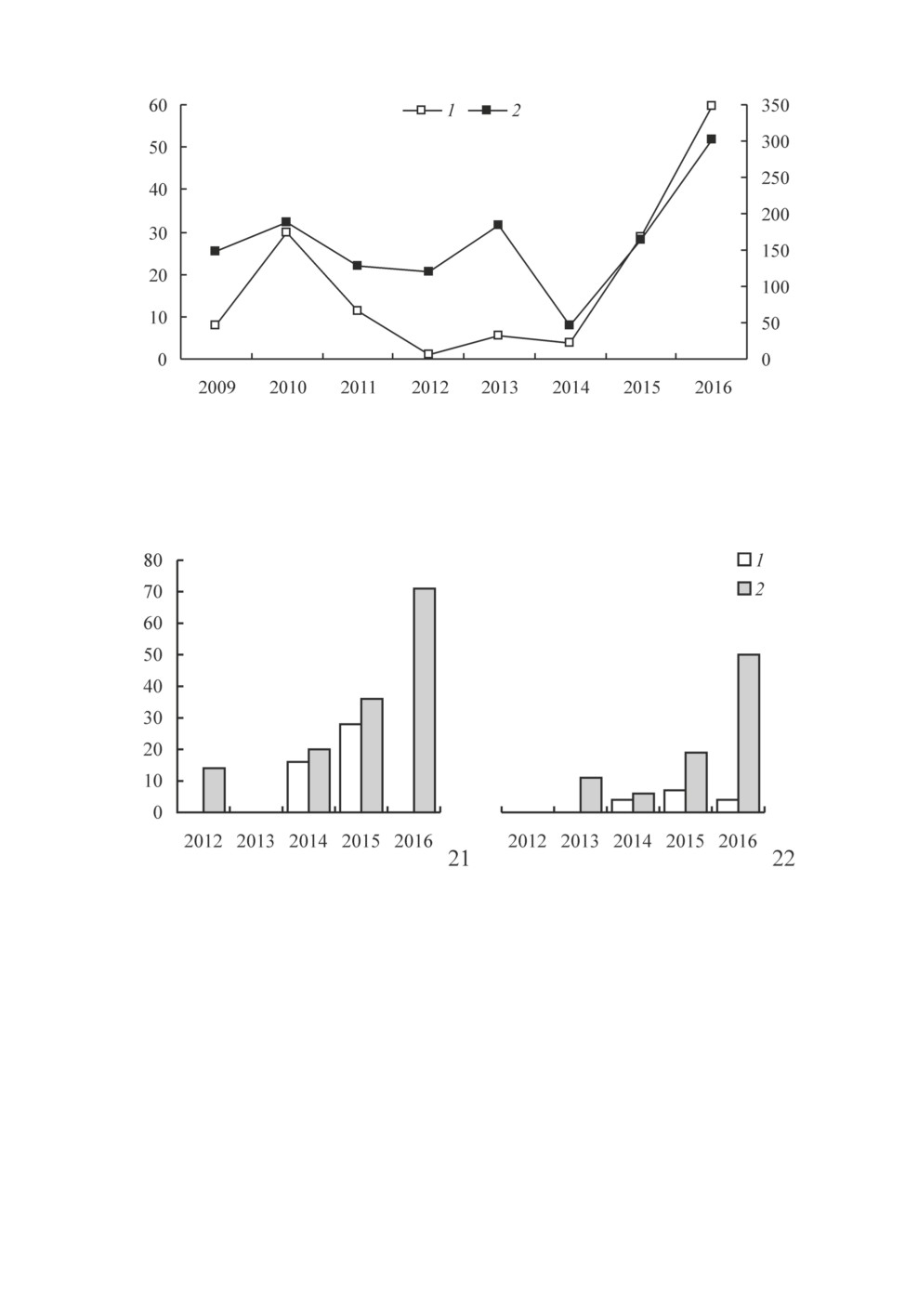
тений в Крыму, является количество осадков. На примере Карадагского природного
заповедника, где исследования были проведены в течение 8 лет подряд, мы проана-
лизировали связь между количеством осадков и заселенностью гнезд-ловушек. Наи-
более сильная связь (rS = 0.86) была выявлена между процентной долей заселения
гнезд-ловушек и количеством осадков, выпавших в период с марта по июнь (рис. 20).
В несколько меньшей степени (rS = 0.83) связь выражена между долей заселения и
количеством осадков, выпавших с января по июнь. Оба коэффициента корреляции до-
стоверны при p ≤ 0.05. Связь процента заселения с количеством осадков, выпавших за
другие месяцы, оказалась заметно меньшей либо недостоверной.
Колебание интенсивности заселения гнезд-ловушек в разные годы определяет-
ся прежде всего сильными изменениями числа гнезд таких видов, как Ancistrocerus
78
Рис. 1. Пункты установки гнезд-ловушек в Крыму.
Основные ландшафтные зоны (по: Выработка приоритетов, 1999): 1 - полупустынные степи и солончаки;
2 - настоящие степи; 3 - предгорные лесостепи; 4 - леса северного макросклона; 5 - горные луга
и степи яйл; 6 - леса южного макросклона; 7 - редколесья южнобережья.
nigricornis, Psenulus fuscipennis, Osmia caerulescens (Linnaeus) и Megachile apicalis
Spinola. Эти виды наиболее часто заселяют гнезда-ловушки в Крыму. Крайняя нерав-
номерность заселения ими гнезд-ловушек на протяжении ряда последовательных лет
в отдельных пунктах позволяет предположить, что они не привязаны к месту своего
отрождения из материнских гнезд и каждый год при выборе места для гнездования
ориентируются в первую очередь на наличие достаточной кормовой базы.
Использование такой своеобразной тактики выбора места гнездования (по обилию
кормовой базы) подтверждается анализом заселения гнезд-ловушек первым и вто-
рым поколениями пчел Osmia caerulescens и Megachile apicalis. В целом ряде случа-
ев в отдельных пунктах установки гнезд-ловушек мы наблюдали полное отсутствие
гнезд первого поколения этих видов при наличии второго, или такое соотношение
числа гнезд первого и второго поколений, которое можно объяснить только прилетом
в данную местность дополнительного числа пчел, отродившихся из гнезд первого по-
коления в других пунктах (рис. 21, 22). Отсутствие второго поколения при наличии
первого мы отметили только в одном случае из 50. Среднее соотношение числа гнезд
пчел первого и второго поколений (в тех пунктах, где обнаружены гнезда обоих поко-
лений) для O. caerulescens составило 1 : 1.2, а для M. apicalis - 1 : 4.6. Возможно, это
означает, что второй вид более мобилен в поиске мест гнездования. С другой стороны,
отсутствие первого поколения у O. caerulescens было отмечено в 1.6 раза чаще, чем
у M. apicalis, что может рассматриваться и как свидетельство обратного. Что касается
осы Ancistrocerus nigricornis, то данный вид дает одно поколение в году, но зимует
на стадии имаго вне гнезд (Фатерыга, 2009). Очевидно, что самки этого вида перед
тем как приступить к гнездованию могут мигрировать на значительные расстояния,
останавливаясь в итоге в местах с наиболее богатой кормовой базой. Оса Psenulus
79
Рис. 2-6. Установка и изготовление гнезд-ловушек.
2 - связка стеблей тростника под крышей строения (вход в гнездо-ловушку показан стрелкой);
3, 4 - ульи Фабра разной конструкции; 5 - связки стеблей тростника, подготовленные к установке;
6 - заготовка тростника на замерзшем пруду.
fuscipennis охотится на тлей, численность которых в отдельных местностях крайне не-
стабильна и подвержена сильным колебаниям от года к году. Поэтому данный вид так-
же не обладает привязанностью к материнских гнездам, которые он, к тому же, никогда
не заселяет вторично.
Для некоторых видов пчел имеет значение не только обильное цветение энтомофиль-
ной растительности, но наличие в ее составе определенных видов растений, напри-
мер, из сем. Brassicaceae для Osmia brevicornis (Fabricius), Lamiaceae для Anthidium
manicatum (Linnaeus) или Asteraceae для Osmia leaiana (Kirby) и O. niveata (Fabricius).
80
Рис. 7-12. Места установки гнезд-ловушек и способы их закрепления.
7, 8 - горельник в Ялтинском горно-лесном природном заповеднике; 9, 10 - каменистая степь с выходами
скал в Опукском природном заповеднике; 11, 12 - побережье в Опукском природном заповеднике.
Вход в гнезда-ловушки показан стрелками.
Еще в большей степени это касается некоторых моно- и узко олиголектических видов,
например, Osmia cerinthidis Morawitz [монолект на Cerinthe minor L. (Boraginaceae)],
Chelostoma mocsaryi Schletterer [олиголект на Ornithogalum (Asparagaceae)] и Hoplitis
manicata Morice [олиголект на Echium (Boraginaceae)]. Для этих трех видов еще одним
фактором, сдерживающим активное заселение гнезд-ловушек, является их очевидная
привязанность к материнским гнездам (филопатрия), которые они способны заселять
многократно (см. ниже). Несмотря на относительно большую численность этих ви-
дов в природе и приемлемость для них гнезд-ловушек в целом, эти пчелы заселяли их
лишь в отдельных пунктах и всегда в небольшом числе.
81
Рис. 13-16. Пчелы и осы, заселившие гнезда-ловушки.
13 - Osmia cerinthidis Morawitz, 14 - Megachile rotundata (Fabricius), 15 - Alastor mocsaryi (André),
16 - Euodynerus posticus (Herrich-Schäffer).
Рис. 17-19. Гнезда Eumenes spp. в гнездах-ловушках.
17 - ячейки гнезда Eumenes papillarius (Christ), построенные на поверхности связки стеблей тростника
в улье Фабра; 18 - гнездо E. papillarius в полости стебля тростника; 19 - гнездо Eumenes sp.
в полости стебля тростника.
82
Рис. 20. Динамика заселения гнезд-ловушек и количества осадков за март-июнь в Карадагском
природном заповеднике в 2009-2016 гг.
По горизонтальной оси - годы наблюдений; по левой вертикальной оси - доля полостей, заселенных
разными группами жалящих перепончатокрылых, %; по правой вертикальной оси - количество
осадков, мм. 1 - доля заселенных полостей; 2 - количество осадков.
Рис. 21, 22. Число гнезд первого и второго поколений двух видов пчел в гнездах-ловушках,
установленных в национальном природном парке «Тарханкутский» в 2012-2016 гг.
21 - Osmia caerulescens (Linnaeus), 22 - Megachile apicalis Spinola. По горизонтальным осям - годы
наблюдений; по вертикальной оси - число гнезд. 1 - первое поколение; 2 - второе поколение.
Как отмечено выше, за все время исследований в гнезда-ловушки удалось привлечь
97 видов жалящих перепончатокрылых. Однако число видов, привлеченных в каж-
дый из сезонов, было значительно меньшим и колебалось от 14 до 42 при среднем
значении 26 (рис. 23). На рис. 23 отражено ежегодное увеличение общего числа ви-
дов, зарегистрированных в гнездах-ловушках. Наиболее быстрый прирост его отме-
чен в первые 3 года (в среднем 15 новых видов в год). В последующие 9 лет средний
ежегодный прирост общего числа видов снизился до 5, а в последние 3 года - до 2.
К видам, отмеченным в гнездах-ловушках в течение наибольшего числа лет, отно-
83
Рис. 23. Динамика заселения гнезд-ловушек жалящими перепончатокрылыми в Крыму
в 2002-2016 гг.
По горизонтальной оси - годы наблюдений; по вертикальной оси - число видов, заселивших гнезда-
ловушки. 1 - число видов, загнездившихся в конкретном году наблюдений; 2 - кумулята накопления
числа загнездившихся видов.
сятся Osmia caerulescens (все 15 лет), Trypoxylon figulus (Linnaeus) и Hoplitis manicata
(по 12 лет), Dipogon subintermedius (Magretti), Euodynerus posticus, Ancistrocerus
parietinus (Linnaeus) и Megachile apicalis (по
11 лет), Syneuodynerus egregius,
A. nigricornis и O. bicornis (по 10 лет).
Более или менее регулярно повторно заселяли ульи Фабра (после размещения в них
гнезд, полученных из гнезд-ловушек) 2 вида ос: Syneuodynerus egregius и Euodynerus
posticus и 9 видов пчел: Chelostoma mocsaryi, Heriades crenulata Nylander, Hoplitis
manicata, Osmia bicornis, O. bidentata (Morawitz), O. cerinthidis, O. dimidiata (Morawitz),
O. cornuta (Latreille) и Megachile rotundata (Fabricius). Все эти виды можно считать
перспективными для искусственного разведения в Крыму. В то же время стоит отме-
тить, что филопатрия у каждого из этих видов проявляется в разной степени и форме,
что необходимо учитывать при разработке технологии их искусственного разведения.
Связь числа полученных гнезд с относительной численностью видов
в природе на примере пчел сем. Megachilidae
Как уже было отмечено выше, пчелы-мегахилиды представлены наибольшим числом
видов среди всех жалящих перепончатокрылых, заселивших гнезда-ловушки в Крыму
(37 из 97), а гнезда их составили 2/3 от всех полученных нами. В связи с этим нами
предпринята попытка выявить связь между числом полученных гнезд отдельных ви-
дов пчел-мегахилид и относительной численностью этих видов в природе. По данным
пяти крупнейших энтомологических коллекций из Крыма известно 8 907 экземпляров
пчел-мегахилид (Fateryga et al., 2018). На основании ограниченной сверху 5-балльной
логарифмической шкалы (Песенко, 1982), виды пчел, заселившие гнезда-ловушки,
можно отнести к редким (8 видов), обычным (25) и фоновым (4 вида). Очень редкие
виды (представленные в коллекциях менее чем 7 экз.) не заселили гнезда-ловушки,
84
Рис. 24-26. Виды пчел сем. Megachilidae, ранжированные по числу полученных гнезд (24, 25)
и коллекционных экземпляров (26).
По горизонтальным осям - виды пчел; по вертикальным осям - число полученных гнезд (24)
или коллекционных экземпляров (25, 26) (число коллекционных экземпляров
по: Fateryga et al., 2018).
85
а массовых видов (представленных в коллекциях более чем 1 445 экз.) в Крыму не
выявлено.
Число коллекционных экземпляров пчел, на наш взгляд, достаточно адекватно от-
ражает относительную численность большинства видов в природе. С другой стороны,
число гнезд, полученных из гнезд-ловушек, в целом пропорционально числу самок
пчел, привлеченных в эти конструкции из природы. Поэтому следует ожидать, что
между числом полученных нами гнезд и числом коллекционных экземпляров пчел-ме-
гахилид должна существовать положительная связь. Такая связь действительно имеет-
ся, однако она довольно слабая (rS = 0.41), хоть и достоверная при p ≤ 0.05. Так, среди
4 видов, построивших в гнездах-ловушках наибольшее число гнезд (более 300), лишь
2 фоновых вида (Osmia caerulescens и Megachile apicalis). Еще один фоновый вид,
M. centuncularis (Linnaeus), отстроил в гнезда-ловушках значительно меньшее число
гнезд (160), а наиболее многочисленный фоновый вид M. pilidens Alfken - всего 5 гнезд
(рис. 24, 25). В то же время среди пчел, построивших в гнездах-ловушках наибольшее
число гнезд, представлены два обычных (т. е., менее многочисленных) вида: Osmia
bicornis и O. dimidiata. Отмеченные случаи несоответствия, на наш взгляд, объясняют-
ся особенностями биологии гнездования отдельных видов пчел-мегахилид.
Так, несоответствие числа полученных гнезд Osmia bicornis численности пчел этого
вида в природе можно объяснить тем, что оценка численности по коллекционным сбо-
рам оказалась заниженной. На самом деле относительная численность пчел этого вида
в природе выше. Это объясняется тем, что сроки массового лёта O. bicornis в Крыму
приходятся на конец апреля и первую половину мая, в то время как большая часть фау-
нистических сборов пчел приходится на более поздние сроки. Относительно большое
число гнезд O. dimidiata в гнездах-ловушках объясняется склонностью самок этого
вида к совместному гнездованию в местах скопления полостей и их активному поиску.
Изучение строения гнезд O. dimidiata (Иванов и др., 2013) показало, что самки данного
вида в природе склонны к заселению брошенных гнезд других видов пчел, очевидно,
предпочитая виды, образующие колонии (агрегации). Гнезда-ловушки хорошо имити-
руют такие агрегации, в особенности, если в них ко времени начала лёта O. dimidiata
уже поселились самки других видов.
Противоположной тактики при выборе места для гнездования придерживаются
самки пчел Anthidium manicatum и Megachile centuncularis. Они относительно часто
и повсеместно заселяют гнезда-ловушки, но, как правило, единично. Наблюдения
показали, что самки этих видов не ищут конструкции с плотным расположением
полостей, а возможно, даже избегают их. Мы часто находили гнезда A. manicatum и
M. centuncularis в одиночно расположенных полостях в непосредственной близости
от ульев Фабра с большим числом свободных гнездовых каналов. Возможно, это явля-
ется причиной меньшего числа полученных нами гнезд этих видов, чем можно было
бы ожидать, исходя из числа собранных коллекционных экземпляров.
Часть видов, относительно редко заселяющих гнезда-ловушки по сравнению с их
численностью в природе, не проявляют особого интереса к заселению гнезд-ловушек
по причине несоответствия последних потребностям этих видов в определенном суб-
страте для гнездования. Так, довольно обычный вид Megachile octosignata Nylander и
наиболее многочисленный фоновый вид M. pilidens предпочитают заселять полости в
почве. Находки их гнезд в гнездах-ловушках можно считать результатом отклонения
от нормы у отдельных самок этих видов в отношении выбора субстрата гнездования.
86
То же относится к Anthidium oblongatum (Illiger), обычно заселяющему широкие ще-
левидные полости между камнями, и к Megachile lefebvrei Lepeletier de Saint-Fargeau,
который использует для строительства ячеек неглубокие выемки в камнях, как прави-
ло, соизмеримые с величиной ячейки. Это можно отнести также к M. maritima (Kirby)
и M. willughbiella (Kirby), обычно выбирающим для строительства гнезд отверстия
в рыхлой древесине, щели под корой и другие подобные полости. Среди остальных
обычных видов обращает на себя внимание относительно малое число гнезд двух ви-
дов пчел из рода Heriades. Для H. crenulata причиной этого является очевидная привя-
занность самок к месту отрождения из материнских гнезд, которые они способны засе-
лять многократно (см. выше). Причина относительно низкого числа гнезд H. truncorum
в гнездах-ловушках, видимо, заключается в отсутствии дефицита гнездового субстрата
данного вида в природе (сухих стеблей растений с мягкой сердцевиной).
Основная масса пчел сем. Megachilidae, заселивших гнезда-ловушки, относятся
к обычным видам, однако использование гнезд-ловушек позволило получить и гнезда
3 довольно редких в Крыму видов: Pseudoanthidium tenellum (Mocsáry), Hoplitis triden-
tata (Dufour et Perris) и Protosmia tiflensis (Morawitz). Гнезда последнего вида отмечены
в гнездах-ловушках впервые.
Ранжированный по числу коллекционных экземпляров ряд видов пчел-мегахилид,
заселивших гнезда-ловушки (рис. 26), обладает заметно большей выравненностью по
обилию (0.78), чем ряд этих же видов, ранжированный по числу полученных гнезд
(0.38). Столь существенные различия между этими рядами по выравненности (рис. 26
в сравнении с рис. 24), на наш взгляд, свидетельствуют о том, что дефицит естествен-
ных полостей, пригодных для гнездования, испытывает в природе лишь небольшая
часть видов пчел-мегахилид. Активное заселение гнезд-ловушек только этим неболь-
шим числом видов является, на наш взгляд, основной причиной меньшей выравненно-
сти видов по обилию гнезд, чем по общему обилию в природе.
ЗАКЛЮЧЕНИЕ
Использование гнезд-ловушек - эффективный метод получения гнезд жалящих пе-
репончатокрылых (прежде всего, одиночных ос и пчел), необходимых для исследова-
ния особенностей их биологии и оценки возможности их искусственного разведения.
Прежде всего, данный метод эффективен для пчел сем. Megachilidae и ос сем. Vespidae.
Предыдущие подсчеты показали, что число гнезд одиночных ос сем. Vespidae, полу-
ченное из связок стеблей тростника и ульев Фабра (использованных как гнезда-ловуш-
ки) за 12 лет, почти в 6 раз превышает число гнезд, найденных за это время в естествен-
ных условиях (Мартынова, Фатерыга, 2015).
Несмотря на абсолютное преобладание видов пчел сем. Megachilidae в гнездах-ло-
вушках в Крыму, общее число видов ос (45) оказалось примерно равным общему числу
видов пчел (47). Для сравнения: многолетние исследования в различных районах США
выявили 75 видов ос и 43 вида пчел, заселяющих гнезда-ловушки (Krombein, 1967).
Другое близкое по объему исследование, проведенное в течение 14 лет в Литве, по-
зволило получить гнезда 26 видов ос и всего 7 видов пчел (Budrienė et al., 2004). В то
же время, в одном из районов Канады выявлено гнездование 9 видов ос (Fye, 1965a) и
10 видов пчел (Fye, 1965b), на Ямайке из гнезд-ловушек получено 6 видов ос и 5 видов
пчел (Jayasingh, Freeman, 1980), а в Свердловской обл. России - 14 видов ос и 4 вида
пчел (Рудоискатель и др., 2010), однако все эти данные основаны на значительно мень-
87
шем числе использованных гнезд-ловушек и менее продолжительных исследованиях.
Анализ роста числа видов, обнаруженных нами в гнездах-ловушках, с течением лет
(см. рис. 23) позволяет заключить, что для выявления основной массы видов, заселя-
ющих эти конструкции, необходимо устанавливать их на определенной территории в
течение не менее чем 10 лет.
БЛАГОДАРНОСТИ
Авторы признательны А. Г. Радченко (Институт зоологии им. И. И. Шмальгаузена
НАН Украины, Киев) за определение видовой принадлежности муравьев, Ю. В. Про-
ценко (Киевский национальный университет им. Тараса Шевченко, Киев) за помощь в
определении апоидных ос, А. С. Кумпаненко (Институт эволюционной экологии НАН
Украины, Киев) за определение дорожных ос, М. Ю. Прощалыкину (Федеральный на-
учный центр биоразнообразия наземной биоты Восточной Азии ДВО РАН, Владиво-
сток) за подтверждение определения пчел рода Hylaeus и В. А. Лапченко (Карадагская
научная станция им. Т. И. Вяземского - природный заповедник РАН, Феодосия) за пре-
доставление данных по количеству осадков в Карадагском природном заповеднике.
Работа А. В. Фатерыги выполнена в рамках темы государственного задания (№ АА-
АА-А19-119012490044-3).
СПИСОК ЛИТЕРАТУРЫ
Выработка приоритетов: новый подход к сохранению биоразнообразия в Крыму. 1999. Вашингтон:
Biodiversity Support Program, 257 с.
Иванов С. П. 1984. Природоохранные аспекты использования искусственных гнездилищ для диких пчел-ме-
гахилид. В кн.: Л. Г. Апостолов (ред.). Природные комплексы Крыма, их оптимизация и охрана. Сим-
ферополь: Симферопольский государственный университет, c. 105-109.
Иванов С. П., Жидков В. Ю., Фатерыга А. В. 2005. Поддержка популяций диких пчел-мегахилид
(Hymenoptera: Megachilidae) в местах естественного гнездования в Крыму. Фальцфейнівські читання.
Т. 1. Херсон: Терра, c. 209-213.
Иванов С. П., Фатерыга А. В., Жидков В. Ю. 2009. Использование гнезд-ловушек и ульев Фабра для изуче-
ния фауны и биологии гнездования одиночных видов ос и пчел (Hymenoptera: Aculeata) в Карадагском
природном заповеднике. В кн.: А. В. Гаевская, А. Л. Морозова (ред.). Карадаг - 2009. Севастополь:
ЭКОСИ-Гидрофизика, c. 215-222.
Иванов С. П., Фатерыга А. В., Кобецкая М. А. 2013. Биология гнездования пчелы Osmia dimidiata Morawitz,
1871 (Hymenoptera, Megachilidae) в Крыму. Энтомологическое обозрение 92 (1): 35-61.
Лакин Г. Ф. 1990. Биометрия. М.: Высшая школа, 352 с.
Малышев С. И. 1963. Дикие опылители на службе человека. М.; Л.: Наука, 68 с.
Мариковская Т. С., Есенбекова П. А., Казенас В. Л. 2001. Сохранение разнообразия жалящих перепонча-
токрылых (Hymenoptera) в антропогенных биотопах юго-восточного Казахстана с помощью искус-
ственных приманочных гнездилищ. В кн.: А. Е. Пахомов, В. Л. Булахов (ред.). Структура и функ-
циональная роль животного населения в природных и трансформированных экосистемах. Тезисы I
международной конференции, Днепропетровск, 17-20 сентября 2001 г. Днепропетровск: Днепропе-
тровский национальный университет, c. 80-82.
Мариковская Т. С., Щербакова Т. И. 1989. Осы в искусственных гнездовьях. Защита растений 1989 (6): 29-31.
Мартынова Е. В., Фатерыга А. В. 2015. Хризидиды (Hymenoptera, Chrysididae) - паразиты ос-эвменин
(Hymenoptera, Vespidae, Eumeninae) в Крыму. Энтомологическое обозрение 94 (2): 379-396.
Песенко Ю. А. 1982. Принципы и методы количественного анализа в фаунистических исследованиях. М.:
Наука, 287 с.
Песенко Ю. А., Боголюбов А. Г. 1979. Оценка выравненности видов по обилию и сравнительный анализ
основных индексов разнообразия. Журнал общей биологии 15 (1): 104-117.
Рудоискатель П. В., Фадеев К. И., Николаенкова А. В. 2010. Использование искусственных гнезд для из-
учения биологии одиночных ос и пчел (Hymenoptera: Vespidae, Pompilidae, Crabronidae, Colletidae,
Megachilidae, Apidae) в окрестностях Екатеринбурга. Труды Русского энтомологического общества
82 (2): 160-164.
88
Фабр Ж. А. 1993. Инстинкт и нравы насекомых. И. Я. Шевырев (ред.). Т. 1. М.: Терра, ix + 608 с.
Фатерыга А. В. 2009. Фенология лёта складчатокрылых ос (Hymenoptera: Vespidae) в Крыму. Известия Харь-
ковского энтомологического общества 16 (1-2): 57-63.
Фатерыга А. В., Жидков В. Ю. 2012. Складчатокрылые осы (Hymenoptera, Vespidae) национального природ-
ного парка «Чаривна гавань». Заповідна справа в Україні 18 (1/2): 81-87.
Фатерыга А. В., Ковблюк Н. М. 2013. Экология гнездования роющей осы Sceliphron curvatum (F. Smith, 1870)
(Hymenoptera, Sphecidae) на Украине. Евразиатский энтомологический журнал 12 (3): 309-314.
Budrienė A., Budrys E., Nevronytė Ž. 2004. Solitary Hymenoptera Aculeata inhabiting trap-nests in Lithuania:
nesting cavity choice and niche overlap. Latvijas Entomologs 41: 19-31.
Budrys E., Budrienė A., Nevronytė Ž. 2009. Check-list of Eumeninae wasps (Hymenoptera: Vespidae) collected
in Lithuania using trap-nests. New and Rare for Lithuania Insect Species. Records and Descriptions 21:
140-149.
Collins J. A., Jennings D. T. 1984. A simplified holder for eumenid nesting blocks (Hymenoptera: Eumenidae).
Entomological News 95 (2): 58-62.
Fateryga A. V., Ivanov S. P., Filatov M. A. 2018. Megachilid-bees (Hymenoptera: Megachilidae) of the Crimean
Peninsula. Entomofauna 39 (1): 235-283.
Fye R. E. 1965a. The biology of the Vespidae, Pompilidae and Sphecidae (Hymenoptera) from trap nests in
northwestern Ontario. Canadian Entomologist 97 (7): 716-744.
Fye R. E. 1965b. Biology of Apoidea taken in trap nests in northwestern Ontario (Hymenoptera). Canadian
Entomologist 97 (8): 863-877.
Gaston K. J., Smith R. M., Tompson K., Warren P. H. 2005. Urban domestic gardens (II): experimental tests of
methods for increasing biodiversity. Biodiversity and Conservation 14: 395-413.
Griswold T. 2001. Two new species of trap-nesting Anthidiini (Hymenoptera: Megachilidae) from Sri Lanka.
Proceedings of the Entomological Society of Washington 103 (2): 269-273.
Iwata K. 1976. Evolution of Instinct. Comparative Ethology of Hymenoptera. New Delhi: Amerind Publishing
Company, xii + 536 p.
Jayasingh D. B., Freeman B. E. 1980. Trap-nesting solitary aculeates (Insecta: Hymenoptera) in St. Catherine
Jamaica. Caribbean Journal of Science 15 (3/4): 69-78.
Klein A.-M., Steffan-Dewenter I., Buchory D., Tscharntke T. 2002. Effects of land-use intensity in tropical
agroforestry system on coffee flower-visiting and trap-nesting bees and wasps. Conservation Biology 16 (4):
1003-1014.
Krombein K. V. 1967. Trap-Nesting Wasps and Bees: Life Histories, Nests, and Associates. Washington: Smithsonian
Press, vi + 570 p.
Kruess A., Tscharntke T. 2002. Grazing intensity and the diversity of grasshoppers, butterflies, and trap-nesting bees
and wasps. Conservation Biology 16 (6): 1570-1580.
Parker F. D. 1976. A new Proteriades reared from trap stems, its biology and nest associates (Hymenoptera:
Megachilidae). Pan-Pacific Entomologist 52 (1): 73-80.
Parker F. D. 1977. A new Proteriades (Xenosmia) from New Mexico with biological notes. Journal of the Kansas
Entomological Society 50 (3): 317-323.
Parker F. D., Bohart R. M. 1966. Host-parasite associations in some twig-nesting Hymenoptera from Western North
America. Pan-Pacific Entomologist 42 (2): 91-98.
Parker F. D., Bohart R. M. 1968. Host-parasite associations in some twig-nesting Hymenoptera from Western North
America. Part II. Pan-Pacific Entomologist 44 (1): 1-6.
Pärn M., Soon V., Vallisoo T., Hovi K., Luig J. 2015. Host specificity of the tribe Chrysidini (Hymenoptera:
Chrysididae) in Estonia ascertained with trap-nesting. European Journal of Entomology 112 (1): 91-99.
Potts S. G., Kevan P. G., Boone J. W. 2005. Conservation in pollination: collecting, surveying and monitoring.
In: A. Dafni, P. Kevan, C. Husband (eds). Practical Pollination Biology. Cambridge (Ontario): Enviroquest,
pp. 401-434.
Taki H., Kevan P. G., Viana B. F., Silva F. O., Buck M. 2008. Artificial covering on trap nests improves the
colonization of trap-nesting wasps. Journal of Applied Entomology 132 (3): 225-229.
Tscharntke T., Gathmann A., Steffan-Dewenter I. 1998. Bioindication using trap-nesting bees and wasps and their
natural enemies: community structure and interactions. Journal of Applied Ecology 35 (5): 708-719.
Wearing C. H., Harris A. C. 1999. Evaluation of the predatory wasp, Ancistrocerus gazella, for biological control of
leafrollers in Otago fruit crops. I. Prey composition, nest structure and wasp productivity from artificial nests.
Biocontrol Science and Technology 9 (3): 315-325.
Wearing C. H., Harris A. C. 2005. Evaluation of the predatory wasp, Ancistrocerus gazella, for biological control
of leafrollers in Otago fruit crops. II. Wasp phenology and seasonal changes in prey composition. Biocontrol
Science and Technology 15 (3): 281-298.
89
ACULEATE HYMENOPTERA (HYMENOPTERA, ACULEATA)
INHABITING TRAP-NESTS IN THE CRIMEA
S. P. Ivanov, A. V. Fateryga, V. Yu. Zhidkov
Key words: trap-nests, Fabre’s hives, Pompilidae, Vespidae, Ampulicidae, Sphecidae, Cra-
bronidae, Colletidae, Megachilidae, Apidae, Formicidae, Crimea.
SUMMARY
Trap-nests for aculeate Hymenoptera were exposed in 80 localities of the Crimea during
15 years (2002-2016). A total of more than 500 trap-nests containing about 25,000 cavities were used.
The result was 6,895 obtained nests of 97 species of Hymenoptera in 9 families: Pompilidae (5 species /
118 nests), Vespidae (27 / 1201), Ampulicidae (1 / 6), Sphecidae (2 / 8), Crabronidae (10 / 591), Colletidae
(7 / 202), Megachilidae (37 / 4619), Apidae (3 / 9), and Formicidae (5 / 141). The rate of colonization
of the trap-nests by different species and groups of the aculeate Hymenoptera is discussed. The
attractiveness of the trap-nests for these insects is discussed in relation to their bionomics (nest structure,
choice of the nesting substrate, number of generations per year, trophic relations, and philopatry).
A strong correlation between the rate of colonization of the trap-nests and the amount of rains fallen
during the beginning of the corresponding season in the given locality is discovered. Nests of some
unusual inhabitants of the trap nests, including Eumenes papillarius (Christ) and Eumenes sp., are
reported. Number of obtained nests of representatives of the family Megachilidae is compared with
the number of museum specimens of corresponding species; the correlation between these parameters
exists but it is relatively low. Two species of wasps and 9 species of bees are considered as suitable for
breeding.
90