ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 98, 1, 2019
УДК 595.715.1
ОБЗОР РАСПРОСТРАНЕНИЯ И ФИЛОГЕНЕТИЧЕСКИХ
ОТНОШЕНИЙ ЩЕТИНОХВОСТОК РОДА
CHARIMACHILIS WYGODZ. (ARCHAEOGNATHA,
MACHILIDAE) С ОПИСАНИЕМ ЛИЧИНОК
CH. CAUCASICA KAPL. И НОВОГО ВИДА
ИЗ БЕЛГОРОДСКОЙ ОБЛАСТИ
© 2019 г. В. Г. Каплин
Всероссийский научно-исследовательский институт защиты растений
шоссе Подбельского, 3, С.-Петербург-Пушкин, 196608 Россия
e-mail: ctenolepisma@mail.ru
Поступила в редакцию 30.11.2017 г.
После доработки 6.08.2018 г.
Принята к публикации 6.08.2018 г.
Впервые выполнен обзор распространения, особенностей внешнего строения, филогенетиче-
ских отношений и направлений эволюции щетинохвосток рода Charimachilis; описаны личинки
второго и третьего возрастов. По внешним морфологическим признакам взрослых особей и ли-
чинок щетинохвостки рода Charimachilis должны быть отнесены к подсем. Machilinae, где этот
род вместе с Turquimachilis образуют группу родов Charimachilis, занимающую обособленное
положение и близкую к подсем. Petrobiinae. Род Charimachilis, вероятно, обособился в низко-
горьях восточного и северо-восточного побережья Средиземного моря в зоне жестколистных
вечнозеленых лесов и кустарников. Прослежены пути распространения его представителей
в глубь европейского континента вдоль южного и западного побережий Черного моря на север
до 50° с. ш. Вероятно, в связи с аридизацией климата и ухудшением условий обитания повсе-
местно получили распространение уменьшение размеров тела и переход щетинохвосток рода
Charimachilis к партеногенезу; обоеполые виды сохранились лишь в условиях влажных реликто-
вых сообществ на Кавказе. Новый вид Charimachilis morozovi sp. n. по отсутствию латеральных
копательных зубцов на передних гонапофизах наиболее близок к Ch. palaestinensis, от которого
отличается строением яйцеклада и стернитов брюшка.
Ключевые слова: Charimachilis, личинки, распространение, морфологические признаки, груп-
па родов, филогения.
DOI: 10.1134/S0367144519010106
Цель исследования - зоогеографический, сравнительно-морфологический и фило-
генетический анализ рода Charimachilis Wygodzinsky, 1939 на основании обобщения
первоначальных и более полных описаний видов и сведений об их распространении
в работах, указанных в списке публикаций, а также материалов автора, для выяснения
их филогенетических отношений и направлений эволюции.
105
При изучении щетинохвосток, собранных автором в Краснодарском крае и в Белго-
родской обл., выявлены личинки младших возрастов и новый вид рода Charimachilis,
описание которых приведено в конце обзора рода. Голотип и все паратипы нового вида
хранятся в коллекции Зоологического института РАН в Санкт-Петербурге.
Положение рода Charimachilis в системе отряда Archaeognatha остается недоста-
точно выясненным. По признакам строения яйцеклада он отнесен к группе родов не-
определенного положения (incertae sedis) в надсем. Machiloidea (Sturm, Bach de Roca,
1993). Мендес (Mendes, 1990) на основании особенностей распределения чешуек
на теле взрослых особей поместил этот род в подсем. Machilinae сем. Machilidae.
При описании рода Charimachilis самцы его представителей не были известны, и
их признаки не учтены в родовой характеристике (Wygodzinsky, 1939). Расширенное
описание рода было опубликовано в 1957 г. (Janetschek, 1957) с добавлением признаков
самца Ch. relicta insularis Janetschek, 1957, на самом деле относящегося, скорее всего,
к роду Lepismachilis. К сожалению, ошибочно указаные Янечеком (Janetschek, 1957)
признаки самца приведены в описании рода Charimachilis, выполненном Мендесом
(Mendes, 1990). Самцы рода Charimachilis были впервые обнаружены и описаны лишь
у кавказских видов Ch. caucasica и Ch. abchasica (Каплин, 1999, 2017).
Дополненное краткое описание рода приводится ниже.
Род CHARIMACHILIS Wygodzinsky, 1939
Типовой вид: Praemachilis orientalis Silvestri, 1908.
Брюшные стерниты хорошо развиты; все тело, включая жгутики усиков, покрыто
чешуйками, что относится к отличительным признакам видов подсем. Machilinae.
Глаза округлые, реже расширенные (Ch. armata, Ch. ukrainensis); парные глазк΄
подошвообразные, расположены перед глазами. Грудных грифельков 2 пары на сред-
них и задних тазиках. Брюшные грифельки на кокситах II-IX сегментов брюшка;
II-VII кокситы брюшка с 1 парой выпячивающихся мешочков. Опорные шипы на кон-
цах брюшных грифельков длинные, составляют 0.35-0.60 длины грифельков (без опор-
ных шипов). Вершинный угол брюшных стернитов острый, прямой или тупой. Лапка
и предлапка хорошо развиты, без скопулы. Передние бедра самцов без сенсорных по-
лей, лапки и голени самцов и самок с игловидными щетинками или без них. Послед-
ний членик нижнегубных щупиков самцов и самок удлиненный. Нижнечелюстные и
нижнегубные щупики и ноги самцов без специализированных щетинок. Генитальный
аппарат самца с длинными 2-4-члениковыми изогнутыми парамерами на IX сегменте
брюшка. Парамеры выступают за вершину пениса и за вершины кокситов IX сегмента.
Пенис крупный, расширенный, достигает вершины кокситов IX сегмента брюшка или
слегка выступает за нее. Яйцеклад членистый, включает 12-20 члеников, утолщенный,
склеротизованный в вершинной части, полностью прикрыт кокситами IX сегмента,
не достигает их вершин. Расширенные в дистальной части передние гонапофизы
яйцеклада с 3-6 склеротизованными латеральными копательными зубцами или без них,
апикальный членик гонапофизов с 1 или 2 выступами, реже без них. Задние гонапофи-
зы с изогнутым, склеротизованным апикальным рогом. Передние и задние гонапофизы
с конечными иглами, их членики с длинными тонкими, редко - с короткими щетинками.
Положение рода Charimachilis в надсем. Machiloidea неясно прежде всего из-за осо-
бенностей строения яйцеклада (Sturm, Bach de Roca, 1993). Описание полового аппа-
106
рата самца (Каплин, 1999) также свидетельствует о высоком своеобразии этого рода.
Высказано предположение о его близости к сравнительно недавно описанному из се-
верной Турции роду Turquimachilis Bach de Roca et al., 2013 (типовой вид T. mendesi
Bach de Roca et al., 2013), отнесенному вместе с Charimachilis к группе с неопределен-
ным положением в сем. Machilidae (Bach de Roca et al., 2013). Самки этих родов сходны
в своеобразном строении яйцеклада, самцы с 1 парой парамер на IX сегменте брюшка,
без сенсорных полей на передних бедрах, специализированных щетинок на ногах и
щупиках; последний членик нижнегубных щупиков самцов и самок удлиненный. Од-
нако II-V кокситы брюшка T. mendesi с 2 парами выпячивающихся мешочков, пара-
меры самцов нечленистые, заметно длиннее более узкого пениса, в дистальной части
изогнутые, утолщенные и почти соприкасающиеся. Передние гонапофизы яйцеклада
T. mendesi не имеют латеральных копательных зубцов. Очевидно, что род Turquimachilis
более специализированный по сравнению с родом Charimachilis, что выражается пре-
жде всего в появлении у его представителей второй пары выпячивающихся мешочков
на брюшных кокситах.
Строение задних гонапофизов яйцеклада у родов Charimachilis и Turquimachilis
с апикальным склеротизованным изогнутым рогом сходно с их строением у представи-
телей юго-западноевропейских родов Catamachilis Silvestri и Promesomachilis Silvestri,
относящихся к группе Promesomachilis с парными глазками, расположенными субме-
диально по отношению к соприкасающимся глазам, с 1 парой парамер на IX сегменте
брюшка самцов и 1 парой грудных грифельков на задних тазиках. Это сходство кон-
вергентное, очевидно, связанное со сходными особенностями условий обитания и от-
кладки яиц (Janetschek, 1991).
Половой аппарат самцов рода Charimachilis сходен с половым аппаратом самцов
рода Kerkiratrobius Bach de Roca et al., 2010 (типовой вид K. rosanae Bach de Roca
et al., 2010), описанного с о. Корфу по самцам и относящегося к подсем. Petrobiinae
(Machilidae) (Bach de Roca et al., 2010). Жгутик усиков K. rosanae без чешуек, пар-
ные глазки΄ подошвообразные, лежат перед глазами, брюшные кокситы с одной па-
рой грифельков, лапки и голени без игловидных щетинок, передние бедра самцов
без сенсорных полей, ноги и щупики без специализированных щетинок, вершин-
ный угол брюшных стернитов острый или прямой, отношение длины брюшных гри-
фельков (без опорных шипов) к длине кокситов V сегмента брюшка 0.43, VIII - 0.92,
IX - 0.63, опорные шипы на грифельках V и VI брюшных кокситов длинные, составля-
ют 0.47-0.58 длины грифельков (без опорных шипов). Почти все перечисленные при-
знаки характерны также для самцов рода Charimachilis, за исключением наличия че-
шуек на жгутиках усиков. Однако у личинок рода Charimachilis третьего возраста, тело
которых покрыто чешуйками, жгутики усиков также лишены чешуек. К сожалению,
самки рода Kerkiratrobius не описаны, и строение их яйцеклада остается неизвестным.
На основании особенностей строения взрослых особей и личинок щетинохвостки рода
Charimachilis отнесены нами здесь к подсем. Machilinae (Machilidae), где этот род вме-
сте с Turquimachilis образует группу Charimachilis, занимающую обособленное поло-
жение и близкую к подсем. Petrobiinae.
Строение полового аппарата самцов отряда Archaeognatha (= Microcoryphia) в зна-
чительной мере связано с особенностями их полового поведения и оплодотворения,
а строение яйцеклада - с особенностями оплодотворения и откладки яиц, которые из-
учены недостаточно. Передние и задние гонапофизы яйцеклада Machiloidea гомоло-
107
гичны выпячивающимся мешочкам (втяжным пузырькам) предыдущих сегментов тела
и служат прежде всего для захвата капель спермы или сперматофоров и втягивания
спермы в половые пути (Silvestri, 1905; Беккер, 1925; Sturm, Machida, 2001; Емелья-
нов, 2014, и др.). У Archaeognatha различают три основных типа оплодотворения. При
первом из них самцы помещают одну порцию половых продуктов (одну или несколь-
ко капель спермы или сперматофоров) на тонкую нить, образованную из выделений
специальных желез на парамерах. Нить прикреплена одним концом к почве, другим
к концу брюшка самца. Самец держит нить в натянутом положении и изгибает тело
вокруг переднего конца тела самки. Самка прикрепляется к самцу с помощью ниж-
нечелюстных щупиков, выдвигает яйцеклад из кокситов IX сегмента, захватывает им
сперматофоры или капли спермы и втягивает сперму. Такой тип оплодотворения отме-
чен у Machilis germanica Janetschek, Lepismachilis y-signata Kratochvil, Dilta insulicola
Wygodzinsky (Machilinae), Pedetontus unimaculatus Machida (Petrobiinae) (Sturm, 1955,
1992, 1993). У самцов родов Machilis, Lepismachilis и Dilta 2 пары, у Pedetontus -
1 пара членистых парамер, пенис маленький или средних размеров, открытый; пенис
и парамеры полностью прикрыты кокситами IX сегмента. Второй тип оплодотворения
описан у Petrobius maritimus Leach (Petrobiinae): самец приближается к самке, выде-
ляет сперматофор вблизи ее яйцеклада, и самка захватывает его и втягивает сперму.
Парамер 1 пара, они сплошные, короткие, значительно короче пениса; пенис длинный,
открытый, почти доходит до вершин кокситов; яйцеклад длинный, тонкий, членистый.
Третий тип оплодотворения описан у самцов Neomachilellus scandens Wygodzinsky
(Meinertellidae). Пенис у самцов сем. Meinertellidae вида маленький, парамеры от-
сутствуют. Самцы откладывают на поверхность почвы сперматофоры, которые самка
захватывает длинным тонким членистым яйцекладом. Особенности оплодотворения
у обоеполых видов рода Charimachilis не изучены.
Строение яйцеклада Archaeognatha в значительной мере также связано со способом
и местом откладки яиц. При откладке яиц махилиды отводят гонапофизы до перпенди-
кулярного к продольной оси тела положения (Sturm, Machida, 2001). Первичной счита-
ется откладка ими яиц в трещины и щели в субстрате, куда они вводят длинный тонкий
членистый яйцеклад (Емельянов, 2014). Откладка в лабораторных условиях яиц самка-
ми Petrobius brevistylis (Petrobiinae), имеющими длинный тонкий членистый яйцеклад,
и расположение их яйцекладок в природных условиях в Юго-Западной Англии деталь-
но описаны Делани (Delany, 1959). Размеры отложенных яиц P. brevistylis 1.0-1.3 ×
0.9 мм. Размеры и форма яиц отличаются значительной изменчивостью, их оконча-
тельная форма зависит от места откладки. При проходе через яйцеклад яйцо сужается
и вытягивается, принимая вновь широкоовальную форму после выхода из яйцекла-
да. В естественных условиях зимующие яйца были обнаружены в феврале и марте.
Яйцекладки компактные, черные, располагались в узких трещинах шириной 1-1.5 мм
в песчанике на глубине редко более 1 дюйма (25 мм), группами от 3 до нескольких
сотен яиц. Яйцекладки располагались изолированно или соприкасались друг с дру-
гом, образуя в трещинах непрерывные полосы. Самки откладывают яйца, засовывая
яйцеклад в трещины в песчанике. Свежеотложенные яйца покрыты утолщенной бле-
стящей оболочкой, вскоре после откладки они приобретают черный цвет. Укорочен-
ный и утолщенный яйцеклад с копательными шипами в дистальной части адаптирован
к откладке яиц в почву и рыхлые субстраты. У щетинохвосток рода Charimachilis
в откладке яиц, очевидно, участвуют задние гонапофизы с апикальным склеротизо-
ванным рогом, имеющимся у всех описанных видов, и передние гонапофизы с опор-
108
ными выступами на их апикальных члениках и боковыми копательными зубцами на
следующих 4-7 члениках, имеющихся у самок большинства описанных видов, однако
субстрат и процесс откладки яиц остаются неизученными.
ОБЗОР ВИДОВ РОДА CHARIMACHILIS И ИХ ФИЛОГЕНЕТИЧЕСКИХ ОТНОШЕНИЙ
В юго-западнопалеарктическом роде Charimachilis Wygodzinsky, 1939 известно
13 видов. Charimachilis egatensis был описан как подвид Ch. relicta (Ch. relicta egatensis)
(Bach de Roca, 1982). После детального анализа дополнительных сборов описанного
с о. Мальта (Stach, 1958a) Ch. melitensis, включавших более 30 самок, и их сравнения
с Ch. relicta, главным образом, по строению яйцеклада, статус Ch. melitensis был по-
нижен до подвида Ch. relicta melitensis (Mendes, 1980). Сравнение особенностей стро-
ения органов зрения, брюшных стернитов и кокситов, нижнечелюстных и нижне-
губных щупиков послужило основанием для повышения статуса подвида Ch. relicta
egatensis до видового (Ch. egatensis) и восстановления видового статуса Ch. melitensis
(Kaplin, 2010). Отношение длины глаза к его ширине составляет у Ch. relicta 0.93-0.96,
у Ch. egatensis 0.88, а у Ch. melitensis - 0.86; отношение длины линии контакта глаз
к длине глаз соответственно 0.51-0.58, 0.75 и 0.58; отношение расстояния между вну-
тренними краями парных глазков к общей ширине глаз - 0.27, 0.15 и 0.12-0.13; отно-
шение длины грифельков (без опорных шипов) к длине кокситов V сегмента брюшка
- 0.37-0.48, 0.58-0.61 и 039-0.45, к длине кокситов VIII сегмента - 0.47, 0.81 и 0.49,
IX - 0.49-0.54, 0.89-0.92 и 0.39-0.47; отношение длины последнего членика нижнече-
люстных щупиков к длине предпоследнего членика - 0.9, 1.1 и 1.2; длины последнего
членика нижнегубных щупиков к его ширине - 2.1, 2.2 и 1.8; вершинный угол брюш-
ных стернитов у Ch. relicta острый, а у Ch. egatensis и Ch. melitensis он прямой или
тупой. Подвиды Ch. relicta insularis Janetschek, 1957 и Ch. relicta meridionalis Janetschek,
1957 были описаны из Греции очень кратко (Janetschek, 1957), и их статус требует
уточнения после изучения дополнительного материала.
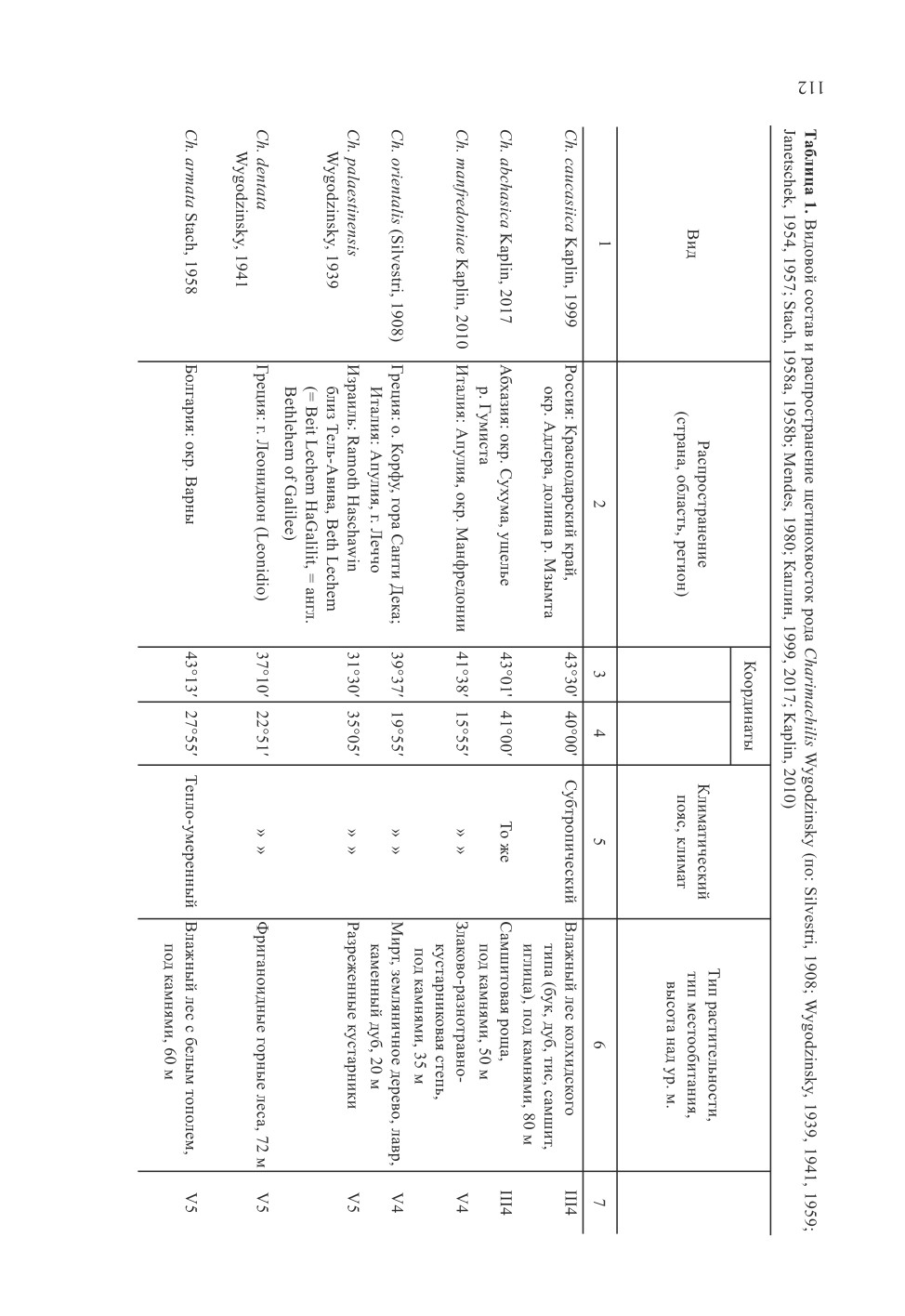
Виды рода Charimachilis распространены в горных, преимущественно лесных и от-
крытых кустарниковых каменистых ландшафтах в субтропическом и умеренном поя-
сах между 31° и 50° с. ш. (от Израиля на юге до Белгородской обл. на севере) и между
11° и 41° в. д. (от Инсбрука (Австрия, Штубайские Альпы) на западе до Абхазии на
востоке) в условиях умеренно-теплого с равномерным увлажнением (30.8 % видов);
умеренно-теплого с сухим и очень сухим летом (средиземноморского) (46.1 %); уме-
ренно-холодного (континентального) климата с равномерным увлажнением (15.4 %)
в Западносредиземноморской и Восточносредиземноморской (по 30.8 % видов), Вос-
точноевропейской (7.7%), Евксинской (горной) (15.4 %) провинциях и в переходной
зоне между Европейской (неморальной) и Скифской (степной) областями (7.6 %). Один
вид (Ch. relicta) с тремя подвидами распространен в Среднеевропейской и Восточно-
средиземноморской провинциях в условиях умеренно-теплого и умеренно-холодного
климатов с равномерным увлажнением и умеренно-теплого климата с сухим и очень
сухим летом. Среднегодовая температура воздуха в пределах ареала рода Charimachilis
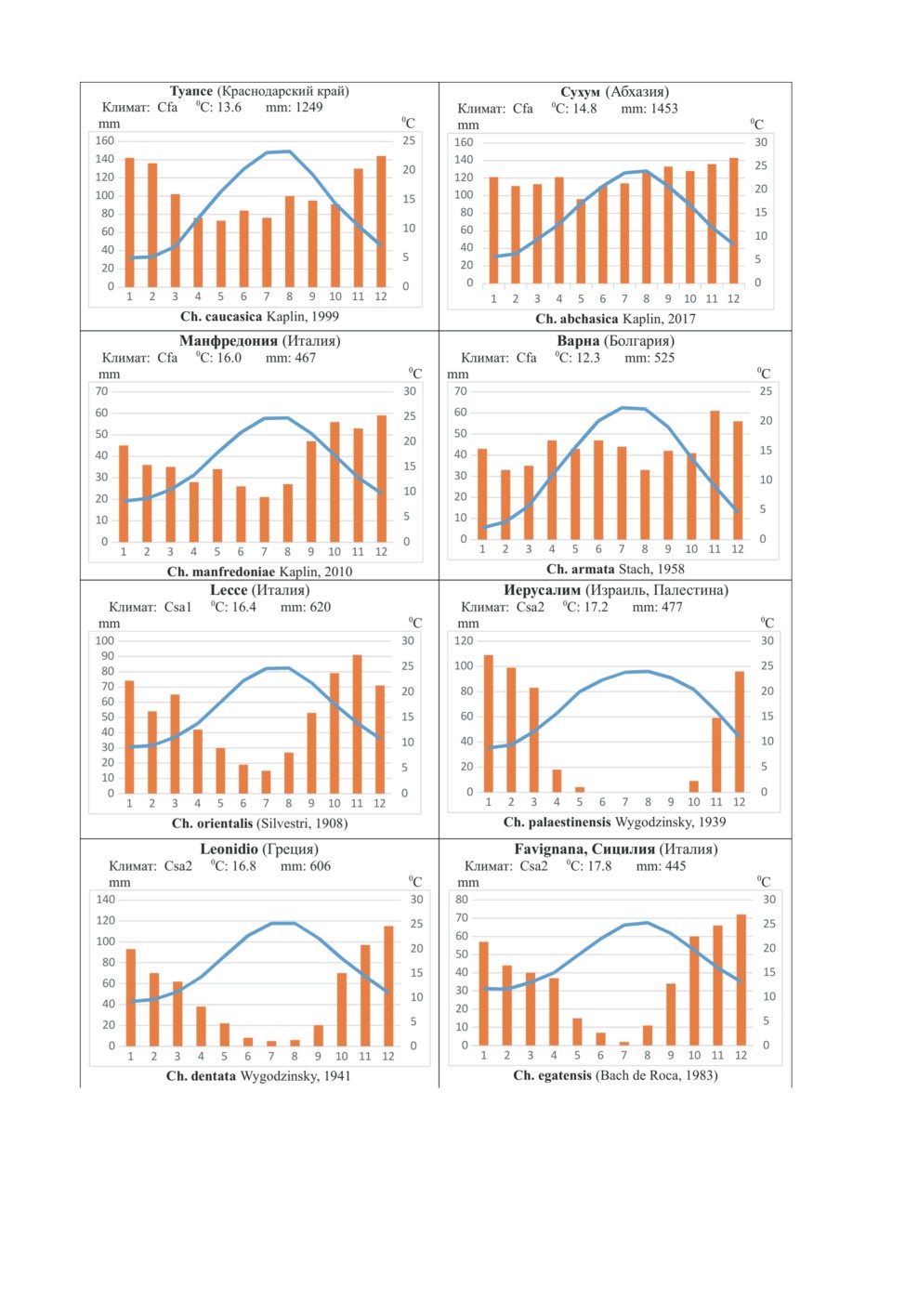
составляет 7-19 °С, среднегодовая сумма осадков -445-1450 мм (табл. 1, рис. 1). Все
виды предпочитают каменистые, меловые, известняковые, часто реликтовые лес-
ные местообитания с повышенной влажностью. Два вида обоеполые (Ch. caucasica,
Ch. abchasica), остальные - партеногенетические. Обоеполые виды известны лишь
из Евксинской горной провинции, отличаются наиболее крупными размерами (дли-
на тела 10.6-14 мм), живут в условиях умеренно-теплого климата с равномерным ув-
109
Рис. 1. Климадиаграммы в местах обитания щетинохвосток рода Charimachilis Wygodz.
Климаты: Cfa - умеренно-теплый с равномерным увлажнением; Csa - умеренно-теплый с сухим (Csa1)
и очень сухим (Csa2) летом (средиземноморский); Dfb - умеренно-холодный (континентальный)
с равномерным увлажнением; 1-12 - месяцы (по: Climate-Data.org).
110
Рис. 1 (продолжение).
111
широта (N)
долгота (E)
Зоогеографическая
область, провинция
лажнением и годовой суммой осадков более 1200 мм. Оптимальные условия для них
складываются в реликтовых тисо-самшитовых ассоциациях Большого Кавказа. Длина
тела партеногенетических видов 7-10 мм, годовая сумма осадков в местах их обитания
составляет 445-810 мм. Наибольшее их видовое разнообразие наблюдается в Западно-
средиземноморской и Восточносредиземноморской провинциях. Географический пар-
теногенез получил сравнительно широкое распространение у щетинохвосток семейств
Machilidae и Meinertellidae и других насекомых; в более благоприятных условиях оби-
тания, обычно в центральных частях ареалов вида или рода, популяции обоеполые, а в
менее благоприятных условиях в периферических частях ареалов виды представлены
только самками, размножающимися партеногенетически (Morgan-Richards et al., 2010;
Notario-Muñoz et al., 2013).
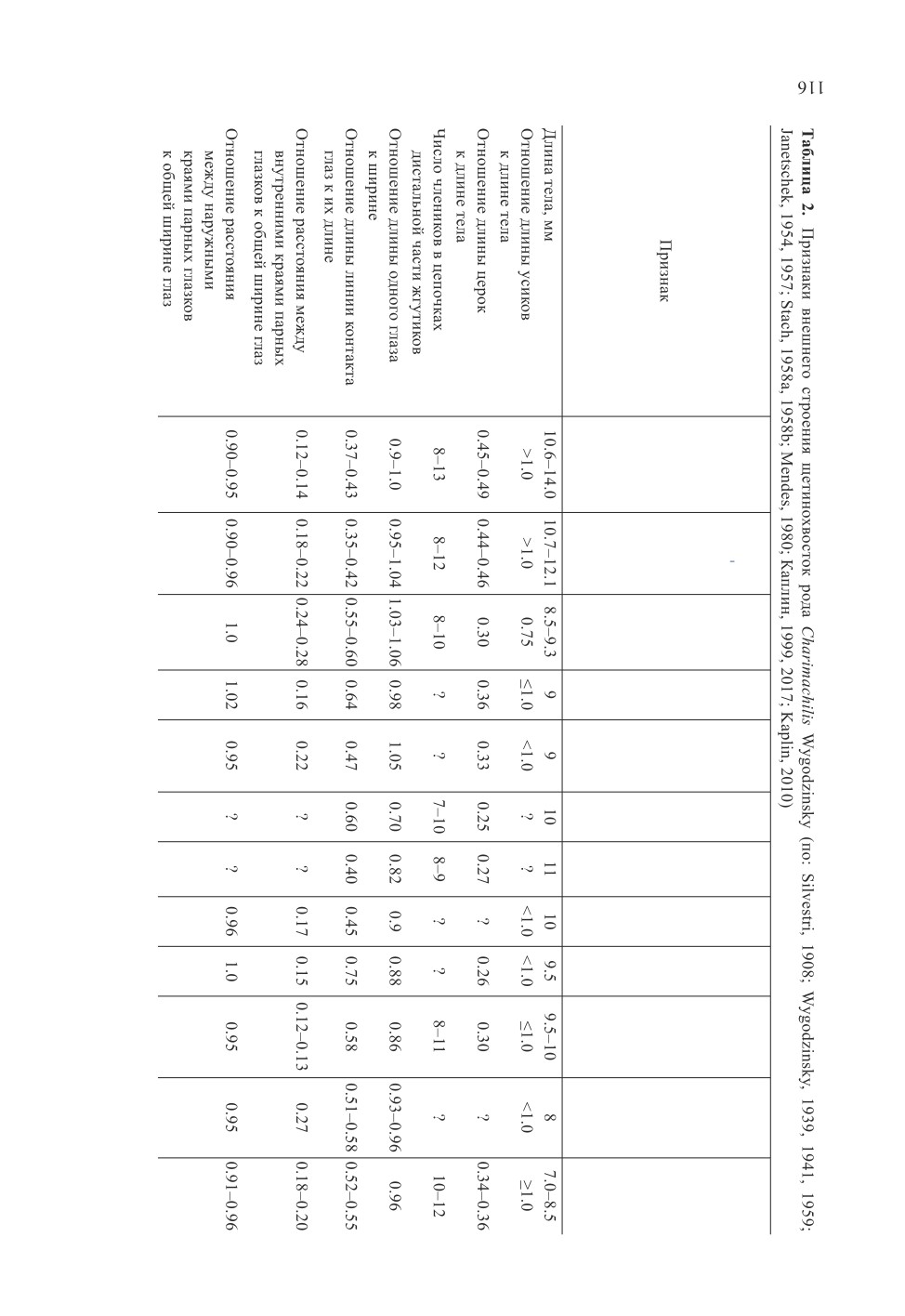
При анализе филогенетических отношений видов рода Charimachilis учтены основ-
ные таксономические признаки видов и их состояния (табл. 2, 3). Необходимо отме-
тить, что такие плезиоморфные признаки, как лежащие перед глазами подошвообраз-
ные парные глазки΄ , наличие 1 пары втяжных пузырьков на I-VII брюшных кокситах,
отсутствие сенсорных полей на передних бедрах самцов, специализированных щети-
нок на их лапках, голенях, нижнечелюстных и нижнегубных щупиках, удлиненный
апикальный членик нижнегубных щупиков у самок и самцов характерны для всех опи-
санных видов рода Charimachilis.
Гла з а и п а р ные гл а зки΄. У видов рода Charimachilis известны 4 типа окра-
ски глаз в спирте. У всех видов, в описании которых указан цвет глаз, их окраска
однотонная. У Ch. armata, Ch. relicta и Ch. manfredoniae глаза темные или черные,
у Ch. palaestinensis - темно-бордовые, у Ch. caucasica - темно-коричневые, у Ch. ab-
chasica и Ch. morozovi - красновато-коричневые, у Ch. wahrmani из Турции - серые.
У турецкого вида Turquimachilis mendesi глаза также серые, двуцветные с темными пят-
нами. Парные глазки΄ черные (Ch. orientalis), темные со светлым пятном (Ch. egatensis),
коричневато-черные (Ch. armata), черновато-коричневые (Ch. morozovi), темно-ко-
ричневые (Ch. manfredoniae), темно-красные (Ch. wahrmani), красновато-коричневые
(Ch. relicta, Ch. caucasica, Ch. abchasica), коричневатые (Ch. ukraniensis), светло-
коричневатые (Ch. melitensis) с узким белым ободом. Иными словами, в окраске глаз
у обитателей открытых ландшафтов преобладает темная, у обитателей лесных место-
обитаний - коричневая, красновато-коричневая, в окраске глазков - коричневая окра-
ска.
У большинства видов рода Charimachilis глаза округлые, отношение длины глаза к
ширине составляет 0.7-1.1. Наиболее расширенные глаза со средним отношением их
длины к ширине 0.7 у восточносредиземноморского Ch. armata. У Ch. ukraniensis этот
показатель составляет около 0.8, у Ch. wahrmani и Ch. egatensis - около 0.9, у осталь-
ных видов глаза округлые или слегка продольно вытянутые (Ch. manfredoniae). Отно-
шение длины линии контакта глаз к их длине в роде Charimachilis меняется от 0.35 до
0.75. Наибольшая относительная длина линии контакта глаз у западносредиземномор-
ского вида Ch. egatensis, наименьшая (около 0.4) - у кавказских и украинского видов
Ch. caucasica, Ch. abchasica и Ch. ukraniensis, у остальных видов она составляет около
0.5-0.6.
Отношение расстояния между внутренними краями парных глазков к общей ши-
рине глаз составляет 0.12-0.28 общей ширины глаз. Сильнее всего сближены глазк΄
у Ch. caucasica, Ch. melitensis и Ch. egatensis, а сравнительно широко расставлены -
у Ch. relicta и Ch. manfredoniae (табл. 2).
114
Особенности строения органов зрения щетинохвосток рода Charimachilis связаны
с природными условиями их местообитаний, что, вероятно, обусловлено сравнительно
небольшими размерами ареала рода (см. рис. 1). Корреляционный анализ показал, что
чем больше годовая сумма осадков и чем больше их выпадает в теплый период года,
а также чем восточнее расположен ареал вида, тем слабее сближены глаза (меньше от-
носительная длина линии контакта глаз) с коэффициентами корреляции соответствен-
но -0.648, -0.534 и -0.756. Парные глазки΄ тем сильнее сближены, чем больше сред-
немесячная температура воздуха в период активности (апрель-октябрь) (коэффициент
корреляции -0.467). Кроме того, чем сильнее расширены глаза (меньше отношение
длины глаза к его ширине), тем сильнее сближены парные глазки΄ (меньше отношение
расстояния между внутренними краями парных глазков к общей ширине глаз) (коэф-
фициент корреляции 0.623).
С учетом анализа морфологических особенностей строения личинок младших воз-
растов в сравнении с имаго к плезиоморфным состояниям следует отнести более уд-
линенные глаза с отношением длины глаза к его ширине более 1.0, наибольшую отно-
сительную длину линии контакта глаз, наибольшее относительное расстояние между
внутренними и наименьшее - между наружными краями парных глазков (табл. 3).
Ус и ки и цер к и. К обычно указываемым систематическим признакам при опи-
сании видов сем. Machilidae относятся число члеников в цепочках дистальной части
жгутика усиков, относительная длина усиков и церок, особенности строения вершины
церок. При этом в пределах рода от примитивных форм к более продвинутым наблю-
дается увеличение числа члеников в цепочках как проявление эволюционного процес-
са полимеризации, способствовавшего увеличению длины усиков в целом. У видов
рода Charimachilis усики обычно короче тела, их длина составляет 0.75-1.2 длины
тела. Число члеников в цепочках вершинной части жгутика меняется незначительно и
у большинства видов Charimachilis составляет 7-13. Наименьшее количество члеников
в цепочках усиков махилид обычно составляет 7-9. Коэффициент корреляции между
длиной усиков и количеством члеников в цепочках жгутика усиков в роде Charimachilis
составляет 0.818.
Хвостовая нить и церки играют важную роль при передвижении щетинохвосток
семейства Machilidae, выполняя опорную функцию. У обоеполых кавказских видов
Ch. caucasica и Ch. abchasica апикальные членики церок двухвершинные, у парте-
ногенетических видов Ch. armata, Ch. egatensis, Ch. manfredoniae и Ch. morozovi они
также двухвершинные, но с более короткой боковой вершиной. Церки Ch. morozovi
10-12-члениковые. Отношение длины церок к длине тела составляет 0.25-0.49, наи-
большее оно (0.44-0.49) у крупных обоеполых видов, а у партеногенетических видов
составляет 0.25-0.36. В процессе эволюции щетинохвосток увеличивается относи-
тельная длина груди и церок. Это подтверждается также при сравнении относительной
длины церок у личинок и имаго. У личинок третьего возраста Ch. caucasica она состав-
ляет 0.21-0.24, а у имаго - 0.40-0.45.
Н иж неч е люс тн ые и н иж не губн ые щ у п ик и имеют обычное для ма-
хилид строение. Дорсальная поверхность 5-7-го члеников нижнечелюстных щупи-
ков с бесцветными зубьевидными хетами. Их количество составляет на 5-м членике
от 1-4 (Ch. morozovi, Ch. ukraniensis, Ch. caucasica) до 2-5 (Ch. abchasica, Ch. egatensis)
и 5-7 (Ch. armata, Ch. manfredoniae), на 6-м членике - от 4 или 5 (Ch. morozovi) до 9-15
(Ch. armata, Ch. manfredoniae, Ch. caucasica, Ch. ukraniensis) и 13-17 (Ch. abchasica,
115
Ch. caucasica Kaplin
Ch. abchasica Kaplin
Ch. manfredoniae
Kaplin (самка)
Ch. orientais
(Silvestri) (самка)
Ch. palaestinensis
Wygodzinsky (самка)
Ch. armata Stach (самка)
Ch. ukrainensis Stach
(самка)
Ch. wahrmani
Wygodzinsky (самка)
Ch. egatensis (Bach
de Roca) (самка)
Ch. melitensis Stach
(самка)
Ch. relicta Janetschek
(самка)
Ch. morozovi sp. n.
(самка)
Таблица 3. Плезиоморфное и апоморфное состояния основных морфологических признаков
в роде Charimachilis Wygodzinsky
Состояние
Признак
плезиоморфное
апоморфное
1. Число члеников в цепочках дистальной
1.1. 6-9
1.2. До 8-10
части жгутиков
1.3. 10-13
2. Длина усиков
2.1. Короче тела
2.2. Длиннее тела
3. Окраска глаз
3.1. Светлая
3.2.Черная, темная
4. Отношение длины церок к длине тела
4.1. 0.25-0.30
4.2. 0.31-0.40
4.3. > 0.4
5. Отношение длины одного глаза к ширине
5.1. < 1.0
5.2. ≥ 1.0
6.Отношение длины линии контакта глаз
6.1. > 0.5
6.2. < 0.5
к длине глаза
7. Отношение расстояния между
8.1. > 0.20
8.2. < 0.2
внутренними краями глазков к общей
ширине глаз
8. Отношение длины последнего членика
8.1. 0.9-1.1
8.2. > 1.1
нижнечелюстных щупиков к длине
предпоследнего членика
9. Количество зубьевидных хет
9.1. < 10
9.2. > 10
на 6-м и 7-м члениках
нижнечелюстных щупиков
10. Отношение длины апикального членика
10.1. 1.8-2.0
10.2. > 2.0
нижнегубного щупика к его ширине
11. Утолщенные игловидные щетинки
11.1. Отсутствуют
11.2. Имеются
на лапках и голенях
12. Отношение длины стернитов II-VI
12.1 0.5-0.6
12.2. > 0.6
сегментов брюшка к длине кокситов
13. Вершинный угол стернитов II-V
13.1. Тупой
13.2. Острый
сегментов брюшка
14. Отношение длины грифельков
14.1. > 0.5
14.2. < 0.5
(без опорных шипов) к длине кокситов V
сегмента брюшка
15. Отношение длины грифельков
15.1. 0.6-0.8
15.2. < 0.6
(без опорных шипов) к длине кокситов
15.3. > 0.8
VIII сегмента брюшка
16. Отношение длины грифельков
16.1. 0.6-0.8
16.2. 0.4-0.6
(без опорных шипов) к длине кокситов IX
сегмента брюшка
17. Латеральные макрохеты на наружной
17.1. Имеются
17.2. Отсутствуют
стороне кокситов IX сегмента брюшка
18. Латеральные макрохеты
18.1. Имеются
18.2. Отсутствуют
на кокситах I-VIII сегментов брюшка
19. Латеральные копательные шипы
19.1. Имеются
19.2. Отсутствуют
в дистальной части передних
гонапофизов яйцеклада
118
Ch. egatensis); на 7-м - от 5 или 6 (Ch. morozovi) до 14-17 (Ch. ukraniensis, Ch. armata,
Ch. manfredoniae), 18-22 (Ch. caucasica и Ch. abchasica) и 22-26 (Ch. egatensis). К од-
ному из выраженных отличительных признаков видов рода Charimachilis может от-
носиться отношение длины последнего членика нижнечелюстных щупиков к длине
их предпоследнего членика, меняющееся от 0.9-1.0 (Ch. relicta, Ch. manfredoniae) до
1.2-1.4 (Ch. melitensis, Ch. ukraniensis, Ch. caucasica, Ch. abchasica) (см. табл. 2). Оче-
видно, что к плезиоморфным состояниям этих признаков относятся умеренное коли-
чество бесцветных зубьевидных хет на 6-м и 7-м члениках нижнечелюстных щупиков,
примерно одинаковая длина их последнего и предпоследнего члеников у самцов и са-
мок. Последний членик нижнечелюстных щупиков у самцов и самок описанных видов
удлиненный, отношение его длины к ширине составляет 1.8-2.8. Специализированные
щетинки на нижнечелюстных и нижнегубных щупиках самцов отсутствуют.
Ноги. Бедра и голени самцов и самок рода Charimachilis немного расширены. Иг-
ловидные щетинки имеются на лапках и голенях у 66.7 % видов, у остальных видов
они отсутствуют (см. табл. 2). Сенсорных полей на передних бедрах и специализиро-
ванных щетинок на ногах самцов нет.
Ст е р нит ы и ко кс ит ы б р ю шка. К основным признакам в строении брюш-
ных стернитов щетинохвосток сем. Machilidae относятся их относительная длина и
размер вершинного угла. На основании изучения строения брюшных стернитов у ли-
чинок младших возрастов Ch. caucasica было установлено, что в процессе эволюции
щетинохвосток рода Charimachilis длина стернитов брюшка возрастала, а размер вер-
шинного угла уменьшался. Следовательно, к плезиоморфным признакам у них отно-
сятся относительно короткий и широкий стернит брюшка и тупой его вершинный угол.
Отношение длины II-V брюшных стернитов к длине кокситов известно всего у 5 ви-
дов, у которых оно составляет 0.50-0.66, наименьшее у Ch. palaestinensis и наибольшее
у Ch. relicta. Вершинный угол V и VI стернитов брюшка меняется в широких пределах
от 50-70° (Ch. armata, Ch. ukraniensis) до 105-110° (Ch. palaestinensis) (см. табл. 2).
У щетинохвосток рода Charimachilis кокситы II-IX сегментов брюшка с парой гри-
фельков; кокситы I-VII сегментов брюшка с 1 парой выпячивающихся мешочков. От-
ношение длины грифельков (без опорных шипов) к длине кокситов V и VI сегментов
брюшка меняется незначительно и составляет у Ch. ukraniensis 0.6-0.7, у остальных ви-
дов этого рода около 0.4-0.5; VIII сегмента - около 0.9 у Ch. ukraniensis и Ch. melitensis,
0.8 - у Ch. morozovi и Ch. egatensis, 0.5-0.7 - у остальных видов; IX сегмента - 0.6-0.8
у Ch. morozovi, Ch. caucasica, Ch. abchasica и Ch. ukraniensis и 0.4-0.5 - у остальных
видов (см. табл. 2). У имаго (в сравнении с личинками младших возрастов) незначи-
тельно уменьшается отношение длины грифельков по отношению к длине кокситов
брюшка. Вероятно, к плезиоморфному состоянию этого признака относятся сравни-
тельно более длинные брюшные грифельки.
Для щетинохвосток рода Charimachilis характерны сравнительно длинные опорные
шипы на брюшных грифельках. Отношение их длины к длине грифельков (без шипов)
составляет на II-VII сегментах брюшка 0.35-0.60. Опорные макрохеты расположены
главным образом на кокситах IX сегмента брюшка. На внутренней стороне этих кок-
ситов 3-11 латеральных макрохет, больше всего их у Ch. palaestinensis. У Ch. morozovi,
Ch. ukraniensis и Ch. armata они есть (в числе 1-3) также на наружной латеральной
стороне кокситов. На IV-VI кокситах брюшка Ch. palaestinensis 0-1 + 0-1, VII - 2-4 +
2-4, VIII - 4-5 + 4-5 сублатеральных макрохет. Наличие макрохет на I-VIII кокситах
брюшка относится к плезиоморфным признакам.
119
Я й це кла д. У видов рода Charimachilis яйцеклад членистый, укороченный, утол-
щенный, не достигает вершин кокситов IX сегмента брюшка. Задние гонапофизы
у всех видов с изогнутым и развитым в разной степени склеротизованным апикаль-
ным рогом. Передние гонапофизы в дистальной части расширенные, их апикальный
членик обычно с 2 выступами, 2-7-й членики с 3-6 латеральными копательными шипа-
ми, реже без них (Ch. morozovi, Ch. orientalis, вероятно, Ch. palaestinensis). Передние го-
напофизы 12-17-, задние - 11-20-члениковые. Апикальные членики передних и задних
гонапофизов с недлинными конечными иглами. Большинство члеников гонапофизов
с длинными волосковидными щетинками, отсутствующими лишь у одного из опи-
санных видов, у которого щетинки на члениках яйцеклада сравнительно короткие
(Ch. orientalis). Наибольшее число члеников в яйцекладах европейских и восточно-
средиземноморских видов Ch. caucasica, Ch. morozovi и Ch. armata, наименьшее -
у самок западносредиземноморских видов Ch. manfredoniae, Ch. melitensis, Ch. egatensis
и Ch. relicta.
Ге ни т а л ьный апп ар ат с ам ца. У самцов рода Charimachilis 1 пара длин-
ных, выходящих за вершины кокситов, в апикальной части суженных и изогнутых
2-4-членистых парамер на IX сегменте. В связи с тем, что парамеры самцов махилид
гомологичны гонапофизам самок, вероятно, исходным состоянием следует считать на-
личие членистых парамер на VIII и IX сегментах, а более продвинутым - их наличие
лишь на IX сегменте. Пенис широкий, доходит до вершин кокситов IX сегмента, не-
много короче парамер.
Анализ состояний основных морфологических признаков позволил предложить схе-
му филогенетических отношений видов рода Charimachilis мировой фауны, которую
следует рассматривать в значительной мере как гипотетическую ввиду отсутствия па-
леонтологических данных и большой однотипности строения видов; схема основана
в значительной мере на сравнении особенностей внешнего строения личинок младших
возрастов и имаго Ch. caucasica и других махилид (рис. 2).
В роде Charimachilis наибольшее число плезиоморфных признаков в строении
тела характерно для восточносредиземноморского вида Ch. palaestinensis (табл. 4).
Обособление рода произошло, по всей вероятности, в горных ландшафтах восточного
и северо-восточного побережья Средиземного моря от Сицилии и Южной Италии до
Израиля (от 14-16 до 35-36° восточной долготы) в зоне жестколистных вечнозеленых
лесов и кустарников. Нет находок из Испании, с юга Франции, северо-запада Италии,
с Сардинии, Корсики, северного побережья Африки. От этой области, скорее всего
с побережий Адриатического моря, произошло распространение представителей рода
вглубь европейского континента до Австрии (Ch. relicta); с побережий Эгейского моря
в одной ветви - в сторону южного побережья Черного моря до северо-западной Тур-
ции (Ch. wahrmani), в другой - от Греции (Ch. dentata) вдоль западного побережья
Черного моря до северо-востока Болгарии (Ch. armata). Группа видов Ch. dentata и
Ch. armata, по-видимому, была исходной для становления восточноевропейского цен-
тра видообразования, включающего районы Большого Кавказа вдоль восточного по-
бережья Черного моря (Ch. caucasica, Ch. abchasica) и континентальные районы Вос-
точной Европы до 50° с. ш. на Украине (Ch. ukraniensis) и на западе Белгородской обл.
(Ch. morozovi; табл. 3, 4; рис. 2). Виды из средиземноморского и восточноевропейского
центров видообразования различаются особенностями строения яйцеклада. У видов
из средиземноморского центра количество члеников в передних и задних гонапофизах
120
Рис. 2. Филогенетические отношения видов в роде Charimachilis Wygodz.
1-19 - см. табл. 8, 9.
121
Признак
Ch. palae-
stinensis
Ch. orientalis
Ch. manfre-
doniae
Ch. egatensis
Ch. melitensis
Ch. armata
Ch. wahrmani
Ch. relicta
Ch. cau-
casiica
Ch. abchasica
Ch. morozovi
sp. n.
Ch. ukrain-
ensis
яйцеклада составляет соответственно 12-16 и 11-14, а у видов из восточноевропей-
ского центра - 14-20 и 15-20. Вершинный угол брюшных стернитов тупой у 16.7 %
восточноевропейских и 42.9 % средиземноморских видов (табл. 1, 2). Обоеполые виды
известны лишь из влажных реликтовых сообществ восточноевропейского центра на
Кавказе. Вероятно, в связи с аридизацией климата и ухудшением условий обитания в
северо-восточной части ареала уменьшались размеры тела и происходил переход ще-
тинохвосток рода Charimachilis к партеногенезу.
ОСОБЕННОСТИ СТРОЕНИЯ ЛИЧИНОК РОДА CHARIMACHILIS
При выяснении вопросов филогении и эволюции насекомых большое значение
имеет изучение изменений строения в процессе постэмбрионального развития, осо-
бенно строения личинок младших возрастов. Сведения о строении личинок рода
Charimachilis в литературе отсутствуют и впервые приводятся здесь на материале по
западнокавказскому виду Charimachilis caucasica Kaplin.
Charimachilis caucasica Kaplin, 1999.
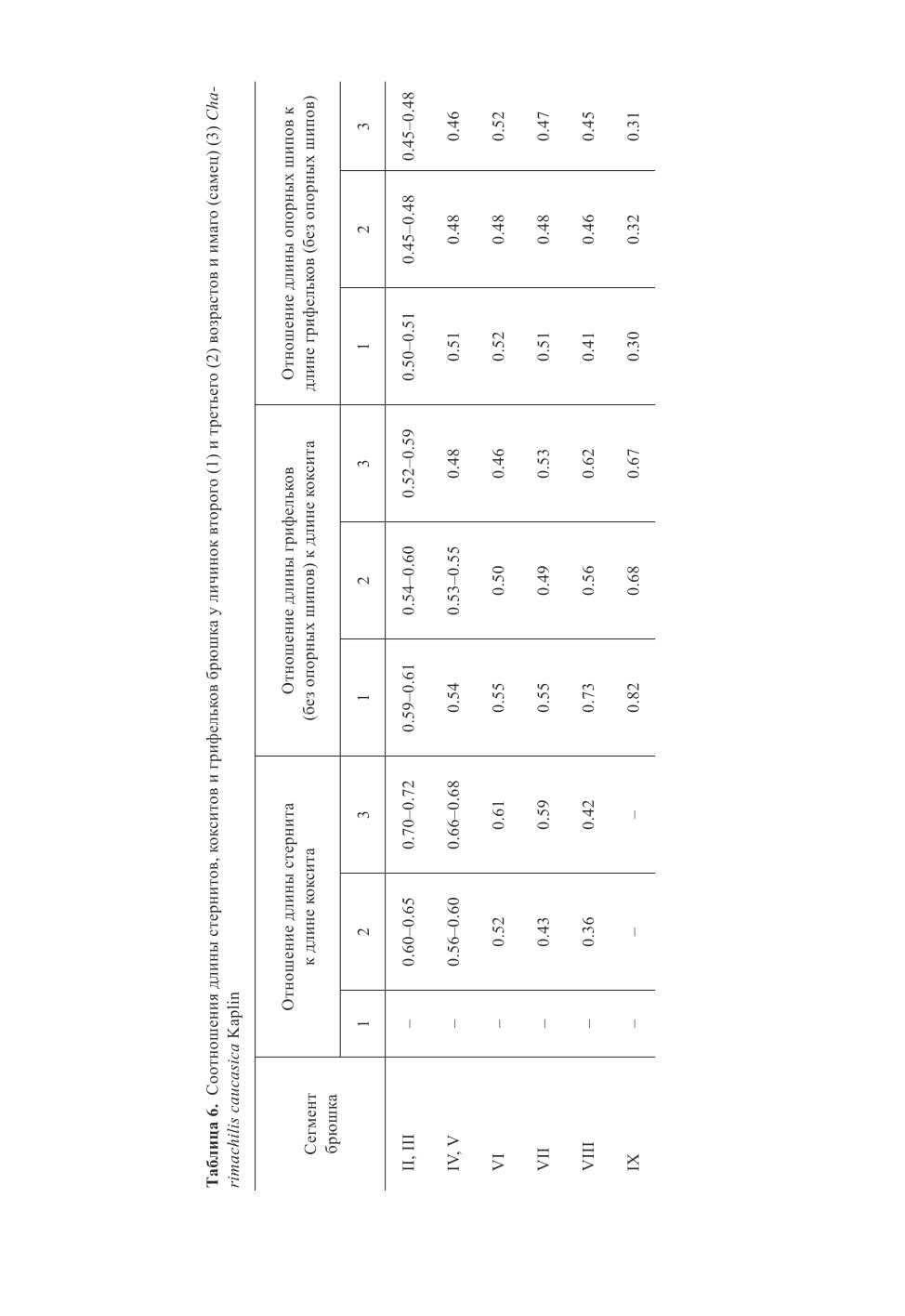
Л ичин ка 2 - го во з р ас т а (рис. 4, 6, 7, 9, 11, 13, 15-23, 26, 27).
М ат ер иа л. Россия. Краснодарский край: окр. Туапсе, ущелье р. Паук, дубово-грабовый лес
с кустарниками, под камнями, 7.V.2017 (В. Г. Каплин), 7 личинок (две личинки в препаратах);
2 ♀, 4 ♂ (1 ♀ и 1 ♂ в препаратах).
Длина тела 4.0-4.1, ширина тела 0.8-0.9, ширина головы 0.65-0.70 мм. Усики немного
длиннее тела, отношение их длины к длине тела 1.02-1.05. Длина церок 0.55-0.60 мм,
или 0.13-0.14 длины тела. Тело без чешуек, темно-коричневое, что, вероятно, обуслов-
лено темной окраской чешуек, формирующихся в новой кутикуле под старой. Наибо-
лее интенсивно окрашены грудные и брюшные тергиты, церки, каудальный филамент,
латеральная сторона бедер. Менее интенсивно окрашены основной членик усиков,
нижнечелюстные щупики, лапки и вентральная сторона брюшка. Цепочки вершинной
части жгутика усиков 6-члениковые. В двух члениках цепочек по 1, в остальных - по
2 ряда щетинок. Все членики цепочек также с 1 округлой, гроздевидной сенсиллой в
средней части с двух сторон, по 1 зерновидной сенсилле ближе к первому ряду щети-
нок тоже с двух сторон, отдельные членики с 1 удлиненной сенсиллой в апикальной
части с одной стороны (рис. 7). Длина основного членика усиков в 1.6 раза больше его
ширины, в базальной части членика имеются очень мелкие сенсорные щетинки. Цер-
ки 10- или 11-члениковые, несут по 1 поперечному ряду щетинок в дистальной части.
Апикальный членик церок с остроконической вершиной (рис. 15). У основания кону-
са по 2 длинных макрохеты, в 1.8 раза длиннее конуса и по 2 сравнительно крупных
сенсиллы. Вблизи других рядов щетинок также имеются по одной мелкой округлой
гроздевидной сенсилле. Внутренние боковые макрохеты церок, направленные к кау-
дальному филаменту, более утолщенные, прямые, немного короче остальных.
Нижнечелюстные щупики с простыми удлиненными щетинками, 7-члениковые. По-
следний членик в 1.1 раза длиннее предпоследнего, 4-й членик в 1.5-1.6 раза короче
5-го. Число дорсальных зубьевидных хет на 7-м членике 5 или 6, на 6-м - 4 или 5, на
5-м членике их нет. Наиболее крупная зубьевидная хета на вершине апикального чле-
ника щупиков, ее длина составляет около 0.20 длины членика. Основной членик щу-
пиков со слабо развитым дорсальным выступом (рис. 11). Вершины верхних челюстей
123
узкие, 4-зубчатые (рис. 9). Вершина лацинии нижней челюсти с 3 хорошо развитыми
зубцами.
Нижнегубные щупики со сравнительно короткими простыми щетинками. Длина
апикального членика в 1.9 раза больше его ширины. Вершина членика с сенсорным
полем, включающим 6 или 7 хорошо развитых сенсорных конусов (рис. 6).
Глаза крупные, соприкасающиеся, в спирте темные. Их длина 0.30-0.31, ширина
0.25-0.26 мм. Отношение длины глаза к его ширине 1.16-1.20. Длина линии контакта
глаз составляет 0.52-0.54 длины глаза. Парные глазки΄ красновато-коричневые, подо-
швообразные, без ободка, лежат перед глазами (рис. 4). Отношение расстояния между
их внутренними краями к общей ширине глаз 0.21-0.22, расстояния между наружными
краями - 0.85-0.86. Ширина глазка в 2.8 раза больше его длины, ширина глазка во вну-
тренней части «подошвы» в 1.6-1.7 раза больше его ширины в наружной части.
Средние бедра, голени и лапки немного расширены по сравнению с передними и
задними (табл. 5). Средние голени с 1, средние бедра с 0-1 игловидными щетинка-
ми. Грифельки на средних тазиках отсутствуют, на задних тазиках едва заметны их
формирующиеся бугорки. Коготки и предлапка хорошо развиты. Лапки 2-члениковые
(рис. 13).
I-VII кокситы брюшка с 1 парой выпячивающихся мешочков. Грифельки на II-IX
кокситах брюшка хорошо развиты. Кокситы IX сегмента брюшка сросшиеся в основ-
ной и средней частях (рис. 27). Стерниты II-VIII сегментов брюшка слабо обособлены,
их границы едва заметны, вершинный угол тупой, в дистальной части с 2-5 хорошо
развитыми щетинками (рис. 26). Все кокситы брюшка с множеством сравнительно рав-
номерно расположенных щетинок средних размеров. Их примерное число на кокситах
I сегмента брюшка 25-30 + 25-30, II-VIII сегментов - 35-45 + 35-45. Кокситы IX сег-
мента с 6 или 7 + 6 или 7 по наружному, 3 + 3 по внутреннему боковому краю и 30-35 +
30-35 щетинками по поверхности кокситов. Отношение длины грифельков (без опор-
ных шипов) к длине кокситов II и III сегментов брюшка около 0.59-0.61, III-VII сег-
ментов - 0.54-0.55, VIII - 0.73, IX - 0.82 (табл. 6). Грифельки с длинными опорными
шипами, отношение длины шипов к длине грифельков (без опорных шипов) на II-VII
кокситах 0.50-0.52, на VIII коксите - около 0.40, на IX - 0.30. Зачатки половых органов
на VIII и IX сегментах отсутствуют.
Тергиты грудных и брюшных сегментов с множеством средних и мелких щетинок,
распределение которых показано на рис. 16-23. Наиболее крупные щетинки образу-
ют поперечный ряд на заднем крае тергитов средне- и заднегруди, а также тергитов
брюшка. Выемка на тергите переднегруди неглубокая, боковые и срединный выступы
находятся на одном уровне.
Задняя кишка вскрытых личинок не содержала остатков пищи. По лабораторным ис-
следованиям постэмбрионального развития щетинохвосток рода Petrobius (Machilidae,
Petrobiinae) и других Archaeognatha, их личинки первых двух возрастов лишены чешу-
ек (Delany, 1959; Sturm, Messner, 1995). Личинки первого возраста оранжево-коричне-
вые, не питаются. В лабораторных условиях продолжительность их жизни составляет
3-5 дней. Личинки второго возраста темные, вероятно, в связи с формированием новой
кутикулы с чешуйками под старой, не имеют грудных грифельков, что в значительной
мере совпадает с особенностями окраски тела и строения вышеописанной личинки.
Отсутствие грудных грифельков у ранних возрастов личинок отмечено также у щети-
124
переднее
среднее
заднее
передняя
средняя
задняя
передняя
средняя
задняя
Рис. 3-15. Charimachilis caucasica Kaplin (личинки).
3, 4 - глаза и глазки΄ ; 5, 6 - нижнегубной щупик и часть нижней губы; 7 - членик цепочки дистальной части
жгутика усиков; 8, 9 - дистальная часть верхней челюсти; 10, 11 - нижнечелюстной щупик;
12, 13 - задняя нога; 14, 15 - вершинный членик церки (3, 5, 8, 10, 12, 14 - личинка 3-го возраста;
4, 6, 7, 9, 11, 13, 15 - личинка 2-го возраста).
Масштабная линейка - 0.1 мм.
127
нохвосток родов Lepismachilis Verhoeff (Verhoeff, 1910) и Machilis Latreille (Machilidae,
Machilinae) (Baer, 1912).
Лич инка 3 - го во зр ас т а (рис. 3, 5, 8, 10, 12, 14, 24, 25, 28).
Мат е ри а л. Россия. Краснодарский край: окр. Туапсе, ущелье р. Паук, дубово-грабовый лес
с кустарниками, под камнями, 7.V.2017 (В. Г. Каплин), 30 личинок (3 личинки в препаратах).
Лич инка с а м ца. Длина тела 4.4-5.0, ширина тела 1.0-1.1, ширина головы
0.80-0.86 мм. Усики длиннее тела, отношение их длины к длине тела 1.2-1.3. Длина
церок 1.0-1.1 мм, или 0.21-0.24 длины тела. Тело покрыто чешуйками, за исключени-
ем жгутика усиков, лишенного чешуек. Чешуйки от темно-бурых до коричневато- и
светло-бурых. Общая окраска тела беловатая или светло-желтоватая. Тело с коричне-
вато-фиолетовым пигментом, собранным в виде двух широких продольных полос на
грудных и брюшных тергитах, вокруг оснований усиков, глазков, задней части головы
вблизи глаз, на основной части верхних челюстей. Цепочки вершинной части жгути-
ка усиков 6-9-, в основном 7-члениковые. В 3 или 4 члениках цепочек по одному, в
остальных - по два ряда щетинок. Все членики цепочек также с 1 округлой, грозде-
видной сенсиллой в средней части с двух сторон, с 1 зерновидной сенсиллой ближе
к первому ряду щетинок тоже с двух сторон, 1 удлиненной сенсиллой в апикальной
части вершинного членика цепочки с одной стороны. Длина основного членика усиков
в 1.5-1.6 раза больше его ширины. Вблизи основания ножки имеются очень мелкие
сенсорные щетинки.
Церки 17- или 18-члениковые, с 1 или 2 поперечными рядами щетинок и чешуек.
Апикальный членик церок с остроконической вершиной (рис. 14). У основания конуса
по 3 длинных макрохеты, в 3.2-3.4 раза более длинных, чем конус, и по 2 сравнительно
крупных сенсиллы. 1-3-й от вершины членики церок без опорных макрохет. 4, 6, 8, 10,
12, 14-16-й от вершины членики с 1 крупной утолщенной бесцветной опорной вну-
тренне-боковой макрохетой в дистальной части члеников. Длина макрохеты в 1.4-1.5
раза больше ширины членика, а ее ширина составляет 0.20-0.25 его ширины.
Нижнечелюстные щупики с простыми удлиненными щетинками, 7-члениковые. По-
следний членик в 1.2 раза длиннее предпоследнего, 4-й членик в 1.4-1.5 раза короче
5-го. Число дорсальных зубьевидных хет на 7-м членике 5 или 6, на 6-м - 4 или 5, на
5-м членике - 0-1. Наиболее крупная зубьевидная хета на вершине апикального члени-
ка щупиков, ее длина составляет около 0.15-0.16 длины членика. 1-й членик щупиков
с хорошо развитым дорсальным выступом (рис. 10). Верхние челюсти в дистальной
части расширенные, с 4-зубчатой вершиной (рис. 8). Вершина лацинии нижней челю-
сти с 3 хорошо развитыми зубцами.
Нижнегубные щупики со сравнительно короткими простыми щетинками. Длина
апикального членика в 1.8 раза больше его ширины (рис. 5). Вершина членика с сен-
сорным полем, включающим 6 или 7 хорошо развитых сенсорных конусов.
Глаза крупные, соприкасающиеся, в спирте темные. Их длина 0.32-0.33, ширина 0.28-
0.30 мм. Отношение длины глаза к его ширине 1.08-1.10. Длина линии контакта глаз
составляет 0.50-0.54 их длины. Парные глазки΄ красновато-коричневые, подошвооб-
разные, без обода, лежат перед глазами (см. рис. 3). Отношение расстояния между их
внутренними краями к общей ширине глаз 0.20-0.22, между наружными краями -
128
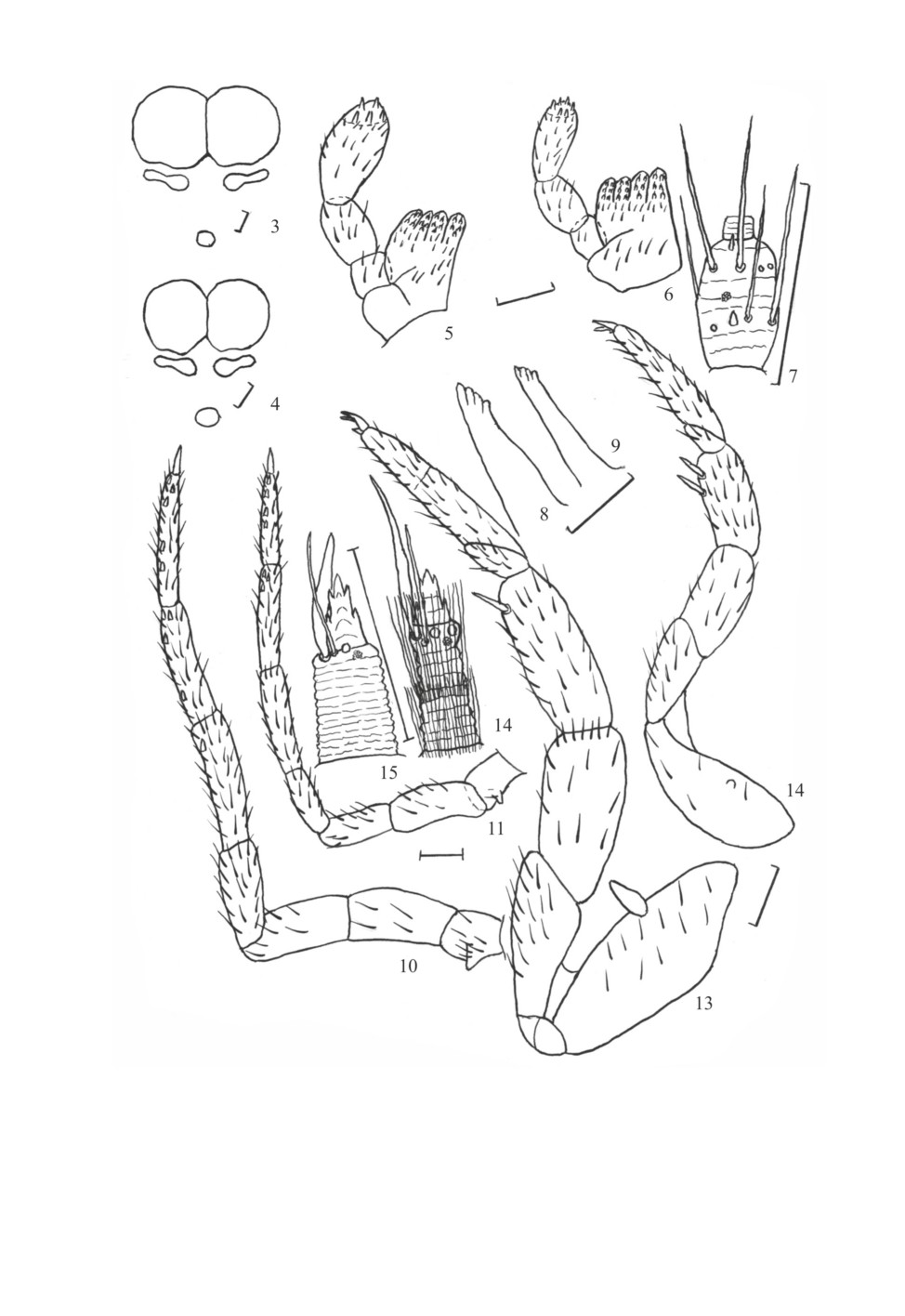
Рис. 16-28. Charimachilis caucasica Kaplin (личинки).
16, 24 - тергит переднегруди; 17, 18 - тергиты средне- и заднегруди; 19-21 - тергиты I-III сегментов
брюшка; 22, 23 - тергиты V и VI сегментов брюшка; 25 - тергит IX сегмента брюшка; 26 - кокситы V
сегмента брюшка; 27 - стернит и кокситы IX сегмента брюшка; 28 - кокситы IX сегмента брюшка с
зачатками полового аппарата самца (16-23, 26, 27 - личинка 2-го возраста; 24, 25, 28 - личинка
3-го возраста).
Масштабная линейка - 0.1 мм.
129
0.82-0.86. Ширина глазка в 2.8-3.0 раза больше его длины, ширина глазков во внутрен-
ней части «подошвы» в 1.6-1.7 раза больше ширины в наружной части.
Ноги хорошо развиты. Средние бедра, голени и лапки немного расширены по срав-
нению с передними и задними (табл. 5). Средние и задние голени с 1 или 2, бедра
с 1 игловидной щетинкой. Грифельки на средних и задних тазиках хорошо заметны,
конусовидные, без щетинок и чешуек (рис. 12); отношение их длины к ширине на сред-
них тазиках 0.25-0.31, на задних - 0.53-0.56. Лапки 3-члениковые, граница между 2-м
и 3-м члениками едва заметна. Отношение длины 3-го членика лапки к общей длине
лапки около 0.39. Коготки и предлапка хорошо развиты.
У личинок 3-го возраста по сравнению с личинками 2-го возраста бедра длиннее
в 1.34-1.40, шире - в 1.42-1.47 раза; голени, соответственно, в 1.28-1.34 и 1.40-1.48
раза; длина лапок больше в 1.19-1.23, ширина передних и задних лапок в 1.11-1.16,
средних - в 1.24 раза.
I-VII кокситы брюшка с одной парой выпячивающихся мешочков. Грифельки на II-
IX и стерниты на II-VIII кокситах хорошо развиты. Грифельки с длинными опорны-
ми шипами. Отношение длины стернитов к длине кокситов II-V сегментов 0.56-0.65,
VI сегмента - 0.52, VII - 0.43, VIII - 0.36; длины грифельков (без опорных шипов)
к длине кокситов II сегмента брюшка около 0.60, III-VII сегментов - 0.50-0.55,
VIII - 0.56, IX сегмента - 0.68. Отношение длины опорных шипов к длине грифельков
(без опорных шипов) II-VIII сегментов 0.45-0.48, IX сегмента - 0.32 (табл. 6). Вер-
шинный угол II-VII стернитов тупой, составляет 104-114°. Все стерниты и кокситы
II-VIII сегментов без щетинок. Кокситы IX сегмента с 2 или 3 внутренними латераль-
ными макрохетами. У личинки самца IX сегмент с зачатками полового аппарата в виде
2 широких треугольных лопастей с закругленными вершинами, отсутствующих
на VIII сегменте (рис. 28).
У личинки самки VIII и IX сегменты брюшка с зачатками передних и задних гонапо-
физов. Зачатки передних гонапофизов в 4-5 раз короче зачатков задних.
Тергиты груди без щетинок, выполняющих механические функции (рис. 24). Тергит
среднегруди в передней трети со значительным числом сенсорных щетинок: напротив
боковых выступов тергита их 12 или 13, напротив срединного выступа - 6. Тергиты I
и II сегментов брюшка с 1 или 2 + 1 или 2, III-X - 2 или 3 + 2 или 3 сублатеральными
макро- и мезохетами (рис. 25).
У щетинохвосток рода Petrobius грудные грифельки отсутствуют у личинок первых
трех возрастов и появляются в виде небольших конусов в 4-м возрасте. У личинок
самцов и самок Petrobius brevistylis Carpenter зачатки полового аппарата появляются
в 4-м возрасте (Delany, 1959).
У личинок 3-го возраста рода Charimachilis происходят существенные морфофи-
зиологические изменения по сравнению с личинками 2-го возраста. Прежде всего,
в 3-м возрасте личинки приступают к питанию, их тело (за исключением жгутика уси-
ков и грифельков) покрыто чешуйками. У личинок 2-го возраста тело покрыто мно-
гочисленными щетинками, без чешуек (рис. 16-23, 26, 27). Как известно, чешуйки
у Archaeognatha по происхождению представляют собой видоизмененные щетинки
(Sturm, Messner, 1995). От личинок второго к личинкам третьего возраста увеличива-
ются размеры тела, число члеников в цепочках жгутиков возрастает от 6 до 7-9, что
130
приводит к относительному увеличению длины усиков, увеличиваются относительная
длина церок от 0.13-0.14 до 0.21-0.24 и количество в них члеников от 10 или 11 до 17
или 18; появляются грудные грифельки на средних и задних тазиках (см. рис. 12), за-
чатки наружных половых органов у личинок самцов и самок (рис. 28); происходит обо-
собление брюшных стернитов, уменьшение относительной длины брюшных грифель-
ков; резко уменьшается число щетинок на брюшных и грудных тергитах, брюшных
кокситах, по-видимому, в связи с замещением их чешуйками; уменьшается отношение
длины глаза к его ширине (см. рис. 12, 13). Практически не меняются количество сен-
сорных конусов на вершине 3-го членика нижнегубных щупиков, зубьевидных хет на
6-м и 7-м члениках нижнечелюстных щупиков (вероятно, в связи с ранним их форми-
рованием), отношения длины линии контакта глаз к длине глаза, расстояний между
внутренним и наружными краями парных глазков к общей ширине глаз.
От личинок 3-го возраста к взрослым особям глаза становятся более широкими,
уменьшаются отношения длины линии контакта глаз к длине глаза от 0.50 до 0.35, рас-
стояния между внутренними краями парных глазков к общей ширине глаз от 0.20-0.22
до 0.12-0.17, а отношение расстояния между их наружными краями к общей ширине
глаз увеличивается от 0.85-0.86 до 0.98-1.05 в связи с увеличением размеров глазков;
увеличивается относительная длина брюшных стернитов, в связи с этим их вершин-
ный угол становится острым (у личинок третьего возраста он тупой); появляются че-
шуйки на жгутике усиков, щетинки и чешуйки на грудных грифельках, значительно
увеличивается их длина, количество члеников в цепочках вершинной части жгутиков
усиков возрастает от 6-9 до 8-13, число зубьевидных хет на 6-м и 7-м члениках ниж-
нечелюстных щупиков - от 4-6 до 10-22, относительная длина церок - от 0.21-0.24
до 0.40-0.45 длины тела; из одновершинных церки становятся двухвершинными. Не-
значительно уменьшается отношение длины грифельков к длине кокситов брюшка;
практически не меняются длина опорных шипов на вершинах грифельков, количество
и расположение макрохет на тергитах груди, брюшка и кокситах брюшка.
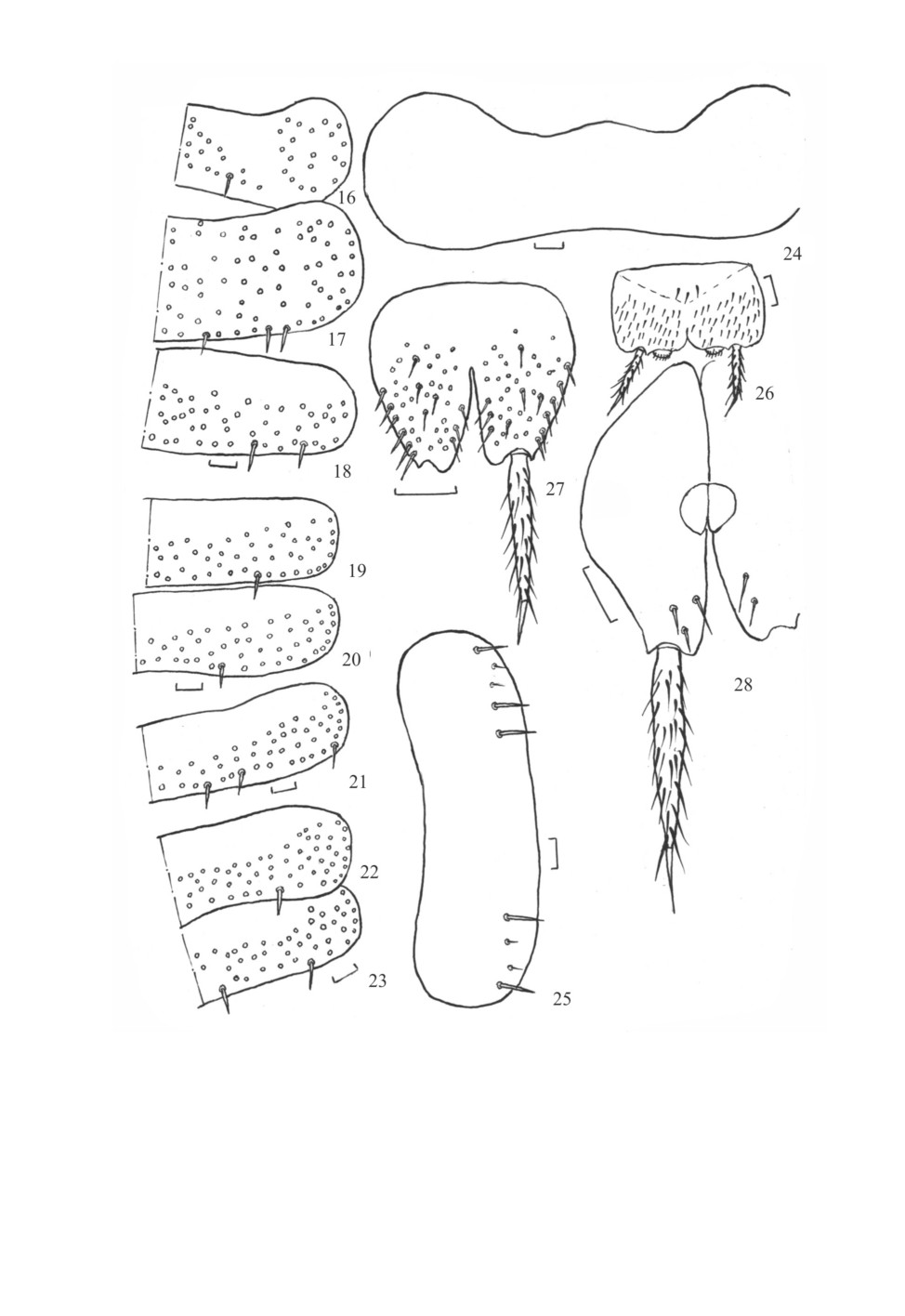
Charimachilis morozovi Kaplin, sp. n. (рис. 29-39).
М ат ер иа л. Россия. Белгородская обл.: Шебекинский р-н, окр. с. Маломихайловка, выходы
меловых пород, разнотравный сосняк, под камнями, 15.VII.2017 (В. Г. Каплин), 1 ♀, голотип
(в препаратах), 2 ♀ (1 ♀ в препаратах), паратипы.
Длина тела 7.0-8.5, ширина - 1.8-2.1 мм. Общая окраска тела беловатая с гипо-
дермальным пигментом вокруг оснований усиков, на лбу, затылке, висках, вокруг не-
парного глазка, по бокам наличник и на нижней губе. Тело полностью покрыто в ос-
новном очень темными чешуйками. Усики равны по длине телу или немного длиннее
его. Цепочки вершинной части жгутика 10-12-члениковые (рис. 30). Членики цепочек
с 1 или 2 рядами щетинок. Длина церок 2.5-2.9 мм, или 0.34-0.36 длины тела. Апи-
кальные членики церок, по-видимому, двухвершинные с короткой, легко обламываю-
щейся боковой вершиной (рис. 35).
Глаза в спирте темные, красновато-коричневые, одноцветные. Общая ширина глаз
0.73-0.80, длина 0.35-0.38 мм. Отношение длины глаза к его ширине 0.96. Длина ли-
нии контакта глаз составляет 0.52-0.55 длины глаза. Парные глазки΄ подошвообраз-
ные, черновато-коричневые с белым ободком, расположены перед глазами; их разме-
ры 0.13-0.14 × 0.31-0.36 мм (рис. 29). Максимальная длина глазков в лобной части
в 1.5-1.7 раза больше их максимальной длины в периферической части. Расстояние
131
Рис. 29-39. Charimachilis morozovi sp. n. (голотип, самка).
29 - глаза и парные глазки΄ ; 30 - цепочка дистальной части жгутика усиков; 31 - нижнечелюстной щупик;
32 - дистальная часть верхней челюсти; 33 - нижнегубной щупик; 34 - передняя нога;
35 - вершинный членик церки; 36 - стернит и кокситы VII сегмента брюшка; 37 - яйцеклад самки
с IX кокситом брюшка; 38 - передний гонапофиз яйцеклада; 39 - задний гонапофиз яйцеклада.
Масштабная линейка - 0.1 мм.
132
между внутренними краями глазков составляет 0.18-0.20, между наружными краями -
0.91-0.96 общей ширины глаз.
Последний и предпоследний членики нижнечелюстных щупиков равной длины
(рис. 31). Дорсальная поверхность 6-го и 7-го члеников нижнечелюстных щупиков
с 8-10, 5-го - с 2 или 3 бесцветными зубьевидными хетами. Последний членик ниж-
негубных щупиков овально-треугольный, его длина в 2.4-2.5 раза больше ширины
(рис. 33). Нижнечелюстные и нижнегубные щупики самцов без специализированных
щетинок. Вершины верхних челюстей 4-зубчатые (рис. 32).
Передние бедра и средние голени расширенные (табл. 7, рис. 34). Наиболее короткие
ноги - средние. Средняя лапка короче передней и задней в 1.23 раза; средняя голень,
соответственно в 1.13 и 1.36 раза, а среднее бедро - в 1.03 и 1.06 раза. Отношение дли-
ны 3-го членика задней лапки к общей длине лапки ~0.35. Все лапки и голени, а также
средние и задние бедра с утолщенными игловидными щетинками (табл. 8). Средние и
задние тазики с грифельками. Длина грифельков на средних тазиках 0.6, задних - око-
ло 0.5 мм. Отношение длины грифельков к ширине среднего тазика составляет око-
ло 1.6, заднего тазика - 1.5. Дистальная часть грифельков с утолщенными опорными
макрохетами.
Вершинный угол II-III стернитов брюшка составляет 75-78°, IV-VII стернитов -
69-73°. I-VII брюшные кокситы с 1 + 1 выпячивающимися мешочками (рис. 36). Соот-
ношения длины грифельков, стернитов и кокситов брюшка приведены в табл. 9.
Грудные, I-V и X брюшные тергиты, брюшные стерниты и I-V брюшные кокситы
без макрохет. Тергиты VI сегмента брюшка с 1 + 1, VII и VIII сегментов - c 1 или
2 + 1 или 2, IX - c 2 или 3 + 2 или 3 сублатеральными макрохетами. Кокситы VI сегмен-
та брюшка с 0-1 + 0-1, VII - с 1-3 + 1-3, VIII - c 1-2 + 1-2 сублатеральными макро-
хетами, IX сегмента с 1-3 + 1-3 наружными и 5-7 + 5-7 внутренними латеральными
макрохетами (рис. 37).
Таблица 7. Отношение длины частей ног к их ширине у самки Charimachilis morozovi sp. n.
Пара ног
Часть ноги
Передняя
Средняя
Задняя
Бедро
1.97-2.00
2.20-2.25
2.34-2.36
Голень
2.18
2.02-2.04
2.86-2.88
Лапка
5.32-5.36
4.84-4.88
5.92-6.00
Таблица 8. Количество игловидных щетинок на ногах самки Charimachilis morozovi sp. n.
Пара ног
Часть ноги
передняя
средняя
задняя
Членик лапки
1-й
1
2 или 3
5
2-й
1 или 2
6
8
3-й
0
1
1 или 2
Голень
1
4 или 5
7
Бедро
0
1
1
133
Таблица 9. Соотношения длины стернитов, кокситов и грифельков брюшка самки Charimachilis
morozovi sp. n.
Отношение длины
Отношение длины
Отношение длины опорных
Сегмент
стернита к длине
грифельков (без опорных
шипов к длине грифельков
брюшка
коксита
шипов) к длине коксита
(без опорных шипов)
II
0.70
0.53
0.47
III-VI
0.60-0.64
0.45-0.46
0.50-0.52
VII
0.66
0.50
0.52
VIII
-
0.82
0.46
IX
-
0.56-0.57
0.36-0.38
Яйцеклад слабо склеротизованный, утолщенный, полностью скрыт кокситами IX
сегмента, типичный для рода Charimachilis (рис. 37). Длина яйцеклада 0.8-1.1 мм.
Передние гонапофизы яйцеклада 16-, задние - 19- или 20-члениковые (рис. 38, 39).
Апикальный членик передних гонапофизов с предвершинной иглой и 2 вершинными
боковыми выступами, 1-й и 2-й членики также с мелкими сенсорными щетинками; 3-й
и 4-й членики от вершины с 1 мелкой простой щетинкой, 5-7-й - с 3 более длинными,
8-12-й - с 4, 13-15-й - с 5 или 6 длинными, 16-й - с 3-5 мелкими щетинками. Длина
конечной иглы равна длине двух вершинных члеников вместе взятых. Копательные
латеральные шипы на передних гонапофизах отсутствуют. Задние гонапофизы с не-
большим изогнутым апикальным рогом и утолщенной предвершинной иглой, длина
которой равна длине двух вершинных члеников, взятых вместе. Три или четыре вер-
шинных и два или три базальных членика задних гонапофизов без щетинок, остальные
членики с 2 сравнительно длинными щетинками.
Самец неизвестен. По-видимому, вид партеногенетический, как и большинство
в этом роде.
Дифф е р е нциа л ьн ый д иа гн о з. Charimachilis morozovi sp. n. по строению
яйцеклада наиболее сходен с Ch. palaestinensis: среди описанных видов только у са-
мок Ch. morozovi sp. n. и Ch. palaestinensis передние гонапофизы яйцеклада лишены
копательных шипов. Задние гонапофизы яйцеклада Ch. morozovi sp. n. включают около
20 члеников, у Ch. palaestinensis - 14. Вершинный угол стернитов брюшка Ch. morozovi
sp. n. отчетливо острый (менее 75°), а у Ch. palaestinensis он тупой (около 105°).
Эт имоло гия. Вид назван именем директора ООО «Агробиотехнология-Шебеки-
но» Дениса Олеговича Морозова, оказавшего помощь в организации сборов щетино-
хвосток в Белгородской обл.
Я искренне благодарен Б. А. Коротяеву (Зоологический институт РАН) за помощь
в подготовке статьи к печати.
СПИСОК ЛИТЕРАТУРЫ
Беккер Э. Г. 1925. К строению и происхождению наружных половых придатков Thysanura и Hymenoptera.
Труды НИИ Зоологии МГУ 1: 157-206.
Емельянов А. Ф. 1974. Предложения по классификации и номенклатуре ареалов. Энтомологическое обозре-
ние 53 (3): 497-522.
Емельянов А. Ф. 2014. Эволюционная роль и судьба первичного яйцеклада насекомых. Энтомологическое
обозрение 93 (1): 91-130. (Английский перевод: Emeljanov A. F. 2014. The evolutionary role and fate
of the primary ovipositor in insects. Entomological Review 94 (3): 367-396).
134
Каплин В. Г. 1999. Новые виды щетинохвосток семейств Machilidae и Lepismatidae (Thysanura) из евро-
пейской части России и Узбекистана. Зоологический журнал. 78 (1): 16-30. (Английский перевод:
Kaplin V. G. 1999. New species of bristletails of the families Machilidae and Lepismatidae (Thysanura) from
European Russia and Uzbekistan. Entomological Review 79 (3): 310-324).
Каплин В. Г. 2017. Новые виды щетинохвосток семейства Machilidae (Insecta, Microcoryphia) из Абхазии.
Энтомологическое обозрение 96 (1): 117-143. (Английский перевод: Kaplin V. G. 2017. New species
of the bristletail family Machilidae (Insecta, Microcoryphia) from Abkhazia. Entomological Review 97 (2):
207-229).
Климат: Туапсе, Сухум, Манфредония, Варна… - Климатический график, График температуры… [Интер-
нет-документ] [Climate-Data.org.] .
Bach de Roca C. 1982. Contribución al conocimiento de los Microcoryphia de Italia (Insecta, Apterygota). Bollettino
del Museo civico di Storia naturale di Verona 9: 523-629.
Bach de Roca C., Gaju-Ricart M., Molero-Baltanás R., Mendes L. F. 2010. A new genus of Petrobiinae (Insecta,
Microcoryphia, Machilidae) from Greece. Zoosystema 32 (2): 259-265.
Bach de Roca C., Fanciulli P. P., Cicconardi F., Molero-Baltanas R., Gaju-Ricart M. 2013. Description of a new
genus and a new species of Machilidae (Insecta: Microcoryphia) from Turkey. Soil Organisms 85 (1): 31-39.
Baer H. 1912. Beiträge zur Kenntnis der Thysanuren. Jenaische Zeitschrift für Naturwissenschaft 48: 1-92.
Delany M. J. 1959. The life histories and ecology of two species of Petrobius Leach, P. brevistylis and P. maritimus.
Transactions of the Royal Society of Edinburgh 63 (3): 501-533.
Janetschek H. 1954. Ein neues inneralpines Nunatakrelikt aus einer für die Alpen neuer Gattung (Insecta, Thysanura).
Sitzungsberichte der Akademie der Wissenschaften in Wien. Mathematisch-Naturwissenschaftliche Klasse)
I (163): 541-548.
Janetschek H. 1957. Über Felsenspringer aus Kreta und den Balkanländern (Thysanura, Machilidae). Acta Zoologica
Cracoviensia 2 (7): 151-191.
Janetschek H. 1991. Konvergenzen beim Bau des Legeapparates von Felsenspringern (Insecta, Apterygota,
Microcoryphia: Machilidae). Berichte des naturwissenschaftlichen-medizinischen Verein Innsbruck 78:
97-102.
Kaplin V. G. 2010. Contribution towards the knowledge of bristletails (Microcoryphia Machilidae) of Italy. Bulletin
of Insectology 63 (1): 1-11.
Mendes L. F. 1980. New data on the thysanurans from Malta (Apterygota: Microcoryphia and Zygentoma). Bolletino
della Societá Entomologica Italiana 112 (4-6): 94-98.
Mendes L. F. 1990. An annotated list of generic and specific names of Machilidae (Microcoryphia, Insecta) with
identification keys for the genera and geographical notes. Estudos, Ensaios e Documentos, Instituto de
Investigação Cientifica Tropical, Lisboa 155: 1-127.
Morgan-Richards M., Trewick S. A., Stringer I. A. 2010. Geographic parthenogenesis and common tea-tree stick
insect of New Zealand. Molecular Ecology 19 (6): 1227-1238.
Notario-Muñoz M. J., Molero-Baltanás R., Bach de Roca C., Gaju-Ricart M. 2013. New Data on the distribution
and biology of Machiloides tenuicornis Stach, 1930 (Microcoryphia: Meinertellidae) in the Iberian Peninsula
(western Palaearctic). Soil Organisms 85 (1): 23-29.
Silvestri F. 1905. Thysanura. Zoologische Jahrbücher 3 (6): 773-786.
Silvestri F. 1908. Materiali per lo studio dei Tisanuri. X: Su alcuni Tisanuri di Corfú. Bollettino del Laboratorio de
Entomologia generale e agraria delle Reale Scuola superiore d’Agricultura in Portici 2: 381-396.
Stach J. 1958a. The Machilidae (Thysanura) of Bulgaria. Acta Zoologica Cracoviensia 3 (1): 1-48.
Stach J. 1958b. Two new species of the genus Charimachilis Wyg. (Thysanura, Machilidae). Acta Zoologica
Cracoviensia 3 (2): 49-66.
Sturm H. 1955. Beiträge zur Ethologie einiger mitteldeutscher Machiliden. Zeitschrift für Tierpsychologie 12 (3):
337-363.
Sturm H. 1992. Mating behaviour and sexual dimorphism in Promesomachilis hispanica Silvestri, 1923 (Machilidae,
Archaeognatha, Insecta). Zoologischer Anzeiger 228 (1-2): 60-73.
Sturm H. 1993. Beziehungen zwischen Paarungsbiologie und Taxonomie bei Felsenspringern (Machiloidea,
Archaeognatha, Insecta). Verhandlungen Westdeutscher Entomologentag, pp. 131-137.
Sturm H., Bach de Roca C. 1993. On the systematics of the Archaeognatha (Insecta). Entomologia Generalis 18
(1/2): 55-90.
Sturm H., Machida R. 2001. Archaeognatha. In: N. P. Kristensen, R. G. Beutel (eds). Handbuch der Zoologie.
Arthropoda: Insecta 4 (37). Berlin: de Gruyter, pp. 63-115.
Sturm H., Messner B. 1995. Chaetotaxy of tergites in the Archaeognatha (Insecta) and its taxonomic significance.
Zoologischer Anzeiger 234: 85-100.
Verhoeff K. W. 1910. Über Felsenspringer, Machiloidea. 3 Aufsatz: die Entwicklungsstufen. Zoologischer Anzeiger
36 (24): 385-399.
135
Wygodzinsky P. W. 1939. Beitrag zur Kenntnis der Thysanuren Palaestinas. Bulletin de la Société Fouad Premier
d’Entomologie 23: 73-85.
Wygodzinsky P. W. 1941. Zur Kenntnis einiger europäischer Dipluren und Thysanuren. Verhandlungen der
Naturforschenden Gesellschaft in Basel 52: 63-100.
Wygodzinsky P. 1959. Beitrag zur Kenntnis der Machilida und Thysanura der Türkei. Opuscula Entomologica
24 (1-2): 36-54.
REVIEW OF DISTRIBUTION AND PHYLOGENETIC RELATIONSHIPS
OF BRISTLETAILS OF THE GENUS CHARIMACHILIS WYGODZ.
(ARCHAEOGNATHA, MACHILIDAE) WITH THE DESCRIPTIONS
OF LARVAE OF CH. CAUCASICA KAPL. AND A NEW SPECIES
FROM BELGOROD PROVINCE
V. G. Kaplin
Key words: Charimachilis, larvae, distribution, morphological characters, group of genera,
phylogeny.
SUMMARY
A review of phylogenetic relationships of species of the genus Charimachilis based on comparative
analysis of zoogeographic distribution of the species, plesiomorphic and apomorphic states of their
main morphological features is performed for the first time. Larvae of the 2nd and 3rd instars of
Charimachilis caucasica are described. The genus Charimachilis belongs to the subfam. Machilinae
(Machilidae), where it forms the Charimachilis group of genera that includes also genus Turkimachilis
and occupies an isolated position in Machilidae close to the subfamily Petrobiinae. Genus Charimachilis
probably originated in the mountainous areas of the Eastern and Northeastern coast of the Mediterranean
Sea in the zone of evergreen forests and shrubs. Patterns of its representatives distribution deep into
the European continent along the southern and western coast of the Black Sea, which have resulted
in the formation of the Eastern European center of speciation, including the Greater Caucasus and
continental regions of Eastern Europe up to 50° N, are traced. Bisexual species are spread only in the
humid environments of relict communities in the Caucasus. Probably everywhere became widespread
body size reduction and the transition of bristletails of the genus Charimachilis to parthenogenesis in
connection with the aridity of the climate and the deterioration of living conditions. The new species
Charimachilis morozovi sp. n. in the structure of the ovipositor is closest to Ch. palaestinensis sharing
the absence of lateral digging teeth on the anterior gonapophyses, but differs from it in the structure of
ovipositor and urosternites.
136