ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 98, 4, 2019
УДК 632.754.1
ФЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ОКРАСКИ ТЕЛА
В ПОПУЛЯЦИЯХ КЛОПОВ-ЧЕРЕПАШЕК (EURUGASTER
LAPORTE, HETEROPTERA, SCUTELLERIDAE) В ПОСЕВАХ
ЗЕРНОВЫХ ЗЛАКОВЫХ КУЛЬТУР И ОБУСЛОВЛИВАЮЩИЕ
ЕЕ ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
© 2019 г. В. Г. Каплин, 1* Г. А. Бурлака2**
1Всероссийский научно-исследовательский институт защиты растений
шоссе Подбельского, 3, С.-Петербург-Пушкин, 196608 Россия
*e-mail: ctenolepisma@mail.ru
2Самарский государственный аграрный университет
ул. Учебная, 2, г. Кинель, пгт. Усть-Кинельский, Самарская обл., 446442 Россия
**e-mail: gaburlaka@mail.ru
Поступила 10.09.2018 г.
После доработки 8.11.2019 г.
Принята к публикации 8.11.2019 г.
Для клопов-черепашек рода Eurygaster - вредителей зерновых злаковых культур в Европе и
Азии - характерна хорошо выраженная модификационная (фенотипическая) изменчивость осо-
бей в популяциях по окраске щитка. Окраска тела клопов-черепашек выполняет защитную функ-
цию и бывает маскировочной, покровительственной, гармонирующей с фоном, либо расчленяющей.
К основным факторам ее формирования относятся существование в неоднородных условиях от-
крыто на надземных органах кормовых растений при питании, открыто или полуоткрыто на по-
верхности и в верхних слоях почвы и подстилки при суточных миграциях, скрыто во вторичных
укрытиях при сезонных миграциях на зимовку и в период летней засухи, а также взаимоотноше-
ния с кормовыми растениями, хищниками и паразитами. Эти факторы способствуют становлению
ряда фенотипов по окраске тела клопов в зависимости от климатических и почвенно-эдафических
условий их обитания. К основным экологическим факторам, оказывающим влияние на формирование
окраски тела клопов-черепашек, относятся климатический, в особенности гидротермические условия
в период их развития; особенности почвы, в частности, ее тип и цвет; характер растительного покрова;
состав и биологические особенности преобладающих кормовых растений, доминирующих энтомо-
фагов и взаимоотношения с ними. В умеренном поясе с возрастанием широты и долготы местно-
сти, снижением температура воздуха и увеличением количества осадков в мае-августе, переходом от
степной зоны к лесостепной, от более светлых каштановых почв к черноземам в популяциях вредной
черепашки возрастает доля особей с однотонно окрашенным темным, серым или серо-коричневым
щитком и снижается участие в популяциях особей со светлым щитком, доля которых, напротив, уве-
личивается в горах и предгорьях Передней и Средней Азии. В лесостепи Самарской обл. в попу-
ляциях клопов-черепашек в агроценозах озимой и яровой пшеницы выявлены особи 5, в агро-
ценозах ячменя - 3 фенотипов по окраске щитка. В посевах пшеницы и ячменя в популяциях
Eu. integriceps и Eu. maura в среднем доминируют особи 1-го фенотипа, довольно часто встреча-
706
ются особи 2-го и 3-го фенотипов. Наибольшее сходство популяций Eu. integriceps и Eu. maura
по окраске тела отмечено в посевах озимой и яровой пшеницы.
Ключевые слова: вредная черепашка, черепашка-мавр, популяция, фенотип, внутривидовая
изменчивость, факторы изменчивости, фитофаги, озимая пшеница, яровая пшеница, ячмень.
DOI: 10.1134/S036714451904004X
Голарктический род Eurygaster Laporte включает 15 видов, из которых 5 описаны из
Северной Америки (Göllner-Scheiding, 2006). В Евразии среди них к вредителям зер-
новых злаковых культур относятся 3 вида: черепашка-мавр Eu. maura (L.), австрийская
Eu. austriaca (Schrank) и особенно вредная черепашка Eu. integriceps (Puton) (Paulian,
Popov, 1980); к потенциальным вредителям - влаголюбивая черепашка Eurygaster
testudinaria (Geoffr.). В России и Казахстане вредная черепашка включена в список
особо опасных вредителей пшеницы.
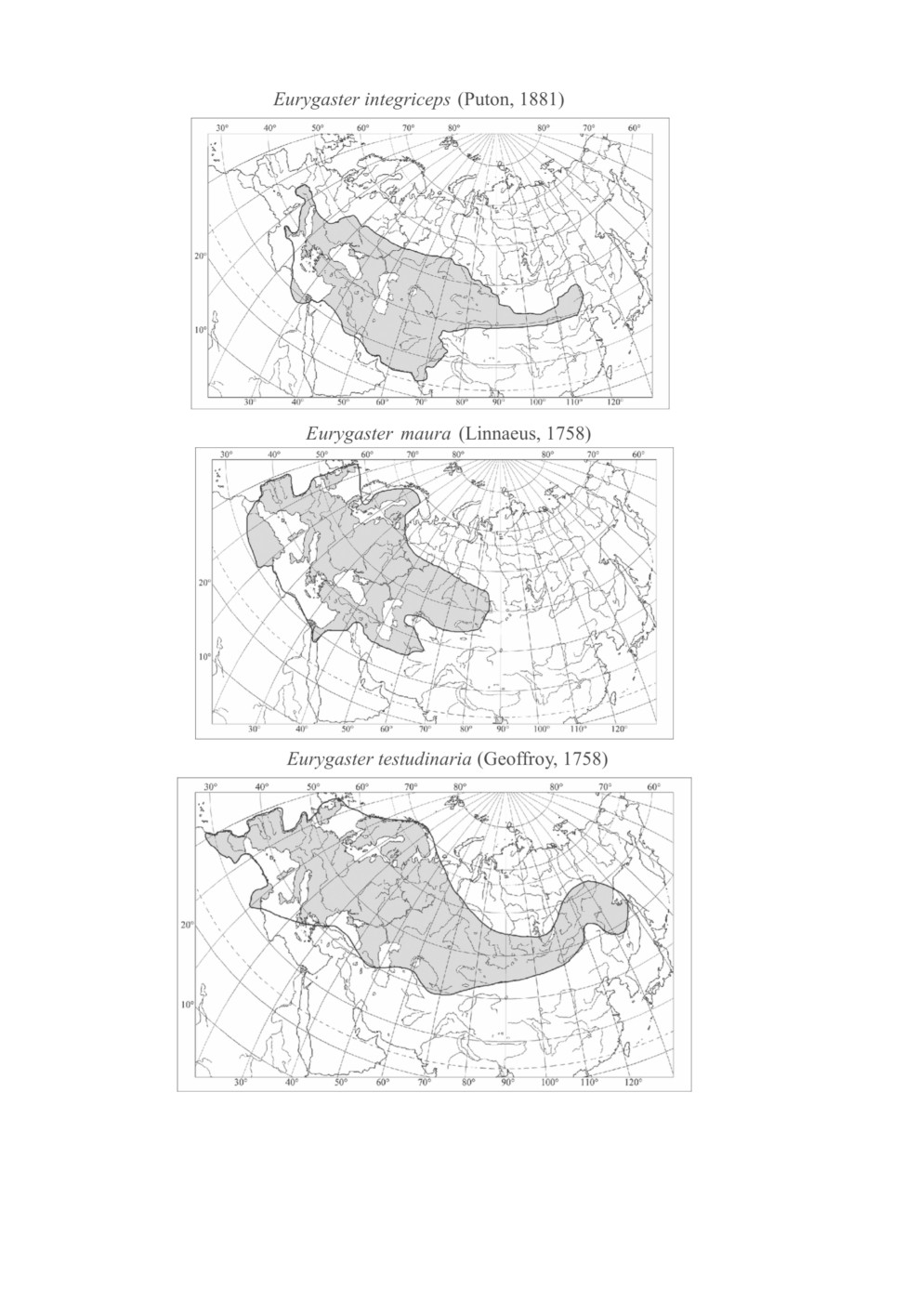
Ареал вредной черепашки охватывает основные районы возделывания пшеницы
в теплоумеренном и субтропическом поясах северного полушария, степной и лесо-
степной зонах, в горных долинах и оазисах в полупустынной и пустынной зонах от
Италии и Хорватии на западе до Восточного Казахстана, Алтайского края, Хакасии
и Северо-Восточного Китая на востоке и от северных областей Украины, Курской,
Липецкой, Тамбовской, Пензенской, Челябинской областей и Башкирии на севере до
Северной Африки (Египет), Ирана и северного Пакистана на юге (Göllner-Scheiding,
2006; Винокуров и др., 2010) (рис. 1). К основным типам почв в местах обитания вред-
ной черепашки относятся черноземы, каштановые, коричневые, серо-коричневые,
реже бурые почвы и сероземы. В наибольшей степени вредная черепашка поврежда-
ет мягкую и твердую яровую и озимую пшеницы, рожь, в меньшей степени ячмень,
овес, кормовые многолетние злаки (Bromopsis inermis (Leys), Agropyron cristatum (L.)
Gaertn., Elytrigia repens (L.) Desv. ex Nevski, Lolium perenne L., Phleum pratense L.,
Dactylis glomerata L., Festuca pratensis Huds. и др.); развивается она также на дико-
растущих многолетних злаках. Дополнительное питание черепашки отмечено на
широком круге двудольных травянистых растений (Павлюшин и др., 2010).
Черепашка-мавр, австрийская и особенно влаголюбивая черепашки имеют бо-
лее широкое распространение по сравнению с вредной черепашкой (рис. 1). Ареалы
Eu. maura и Eu. austriaca - западнопалеарктические, занимают большую часть Евро-
пы, где черепашка-мавр встречается на севере до Великобритании, Швеции и Фин-
ляндии включительно, а австрийская - до Великобритании и Дании. Ареал австрий-
ской черепашки простирается на восток до Восточного Казахстана и Алтайского края,
Eu. maura - до северной Монголии (сомон Баян-Уул, Восточный аймак (аймак Дорнод)).
Южная граница ареала Eu. maura почти совпадает с таковой у вредной черепашки,
у австрийской черепашки она проходит через Алжир, Марокко, Турцию и Иран. Аре-
ал влаголюбивой черепашки транспалеарктический, охватывает Европу от Финлян-
дии, Норвегии, Великобритании и Ирландии на севере до Марокко, Туниса, Турции,
Казахстана, Киргизии, Узбекистана и Таджикистана (Kerzhner, 2003) на юге, на вос-
ток простирается до Приморского края, Кореи и Японии. Австрийская черепашка
и черепашка-мавр развиваются на пшенице, ржи, ячмене, дикорастущих злаках, влаго-
любивая - преимущественно на дикорастущих злаках. В Самарской обл. во влажном
2011 г. единичные особи влаголюбивой черепашки собраны в посевах мягкой яровой
пшеницы (Бурлака, Каплин, 2015).
707
Рис. 1. Распространение видов рода Eurygaster Laporte (по: Göllner-Scheiding, 2006; Винокуров
708
Для всех видов клопов-черепашек рода Eurygaster, питающихся на растениях преи-
мущественно формирующимися, созревающими и зрелыми семенами в светлое время
суток и совершающими регулярные суточные миграции с растений на поверхность
почвы в неблагоприятный по метеоусловиям период, а также сезонные миграции
в жаркие летние месяцы и на зимовку с возвращением на посевы, характерна хорошо
выраженная в популяциях ненаследственная (фенотипическая) изменчивость особей
по окраске щитка и брюшка (рис. 2-4).
Фасулати (2005, 2010) были впервые описаны 4 фенотипа по окраске щитка в популя-
ции вредной черепашки, названные им морфотипами: 1 - щиток с четко выраженным
темно-серо-белым почти без полутонов контрастным узором; 2 - щиток с малокон-
трастным черно-серым или светло-серо-белым полутональным узором; 3 - щиток без
узора, серый, тон окраски темный; 4 - щиток без узора, тон окраски светлый (рис. 4).
Позднее при описании этих фенотипов были добавлены уточнения окраски щитков
для 1-го и 3-го типов - серо-коричневый, 2-го и 4-го - серо-желтый, светло-серо-жел-
тый (Павлюшин и др., 2008, 2010, 2015). Аналогичные фенотипы выделяли Скребцова
(2009) для популяций вредной черепашки из Ставропольского края, Вилкова с соавт.
(Вилкова и др., 2014) из Краснодарского края, Ростовской, Саратовской и Волгоград-
ской областей; Капусткина и Нефедова (2015) для Алтайского края и др. Необходимо
отметить, что в популяциях вредной черепашки окраска тела отличается значительной
изменчивостью. Различия между особями особей 1-го и 2-го фенотипов в окраске щит-
ков нерезкие, узор меняется от наиболее резко контрастного у особей 1-го фенотипа
до слабоконтрастного с сильно размытыми границами у 2-го. У особей 3-го фенотипа
щиток может быть однотонно окрашенный без узора, со слабо заметными осветленны-
ми пятнами в боковых и задней, или только в задней части щитка; с узкой светлой про-
дольной полосой при однотонной окраске основной части щитка или также со слабо
заметными осветленными пятнами. У светлых особей 4-го фенотипа в различной сте-
пени выражен также узор из наиболее осветленных боковых и заднего пятен. Общая
окраска тела, включая щиток, может быть также светлой, коричневатой, красноватой,
желтоватой. Бурлака (2016) по результатам исследований в Самарской обл. добавила
к указанным фенотипам еще один (№ 5), к которому отнесла меланистических особей
черного цвета (см. рис. 4). Снегин и Курносова (2013) в популяциях вредной чере-
пашки в Белгородской обл. выделили 10 фенотипов по степени выраженности рисунка
щитка, переднеспинки и надкрылий: 1 - четко выраженный светлый рисунок, все эле-
менты которого соединяются между собой; 2 - четко выраженный светлый рисунок,
границы между всеми его элементами отчетливы; 3 - четко выражена верхняя часть
рисунка, нижние парные пятна размыты; 4 - неясно выражены нижнее одиночное пят-
но и парные верхние, остальные элементы рисунка отсутствуют; 5 - неясно выраже-
ны верхние элементы рисунка и нижнее непарное пятно, нижних парных пятен нет;
6 - неясно выражены верхние элементы рисунка, слабо заметны нижние парные пятна,
нижнее непарное пятно выражено четко; 7 - окраска темная, контрастно выделяются
верхние парные пятна и нижнее непарное пятно; 8 - черная окраска с коричневато-
кирпичными нечетко выраженными пятнами; 9 - черная окраска без каких-либо свет-
лых пятен; 10 - однотонная светло-коричневая окраска кутикулы без выраженных
пятен. Окраска особей 1-го и 2-го фенотипов в этой классификации соответствует 1-му
фенотипу в классификации Фасулати (2005); 3-го и 6-го - 2-му; 4-го и 5-го - 3-му;
10-го - 4-му; 7-9-го - 5-му фенотипу в классификации Бурлаки (2016). Киль и Исмаи-
лов (2009) в популяциях вредной черепашки в Краснодарском крае выделяют 15 фено-
709
Рис. 2. Eurygaster testudinaria (Geoffr.), изменчивость окраски тела.
org.uk/heteroptera/Scutellerideae/Scutellerideae_images/e_testudinaria_6.jpg;
Scutellerideae_images/e_testudinaria_1.jpg); 2, 3 - Surrey (August 2007) ©Stuart Read;
Scutellerideae_images/e_testudinaria_3.jpg);
arthropoda/insecta/heteroptera/scutelleridae/eurygaster/zzimages/2014_08_12.jpg).
710
Рис. 3. Eurygaster maura (L.), изменчивость окраски тела.
heteroptera/Scutellerideae/Scutellerideae_images/e_maura_9.jpg);
Scutellerideae_images/e_maura_7.jpg); 4 - Jersey (24.VII.2014) © Tim Ransom
711
Рис. 4. Eurygaster integriceps (Put.), изменчивость окраски тела.
1-5 - то же, что в тексте (Фасулати, 2005): 1 - Харьковская обл., Безлюдовка, окр. пос. Удянская, 2.IV.2009
712
типов по окраске щитка (1 - желтый, контрастный; 2 - желтый, малоконтрастный;
3 - желтый, без рисунка; 4 - коричневый, контрастный; 5 - коричневый, малокон-
трастный; 6 - коричневый, без рисунка; 7 - бурый, контрастный; 8 - бурый, мало-
контрастный; 9 - бурый, без рисунка; 10 - темный, серо-коричневый, контрастный;
11 - темный, серо-коричневый, малоконтрастный; 12 - темный, серо-коричневый, без
рисунка; 13 - темно-бурый, малоконтрастный; 14 - темно-бурый, без рисунка; 15 - чер-
ный, без рисунка) и 10 фенотипов по окраске брюшка (1 - оранжевое, рисунок раз-
вит; 2 - оранжевое, без рисунка; 3 - бурое, рисунок имеется; 4 - бурое, без рисунка;
5 - желтое, рисунок развит; 6 - желтое, без рисунка; 7 - темно-серое, рисунок развит;
8 - темно-серое, без рисунка; 9 - коричневое, рисунок развит; 10 - коричневое, без
рисунка). В основу их классификации взяты общая окраска щитка (желтый, корич-
невый, бурый, темный серо-коричневый - по 3 фенотипа, темно-бурый - 2 фенотипа,
и черный - 1 фенотип) и брюшка (оранжевое, бурое, желтое, темно-серое и коричне-
вое), а также развитие на них рисунка (контрастный, слабоконтрастный или отсутству-
ет на щитке; отсутствует или развит на брюшке).
Популяции вредной черепашки были исследованы в лесостепях и степях умеренного
пояса в посевах зерновых злаковых культур на черноземах и каштановых почвах (табл. 1).
Их состав по соотношению особей разных фенотипов различается в зависимости от гео-
графического положения (широты и долготы местности), климатических и почвенных
условий. Чем больше широта и долгота местности, тем ниже в популяциях доля особей
с малоконтрастным узором щитка 2-го фенотипа и выше доля особей с однотонно окра-
шенным темным, серым или серо-коричневым щитком 3-го фенотипа с коэффициен-
тами корреляции по широте соответственно -0.366 и 0.576, по долготе -0.630 и 0.644.
По классификации климатов Кеппена, климаты в исследованных регионах относятся к
умеренно теплому типу с равномерным увлажнением и средней температурой воздуха
в июле 23-28 °C (Краснодарский край); умеренно холодному с равномерным увлажне-
нием и средней температурой воздуха в июле 23-28 °C (Саратовская, Волгоградская,
Ростовская области, Ставропольский край); умеренно холодному с равномерным увлаж-
нением и средней температурой воздуха в июле 18-23 °C (Белгородская, Тамбовская,
Самарская области, Алтайский край). Доля особей со светлым щитком 4-го фенотипа
в популяции тем выше, чем меньше осадков в теплый период года и выше среднемесячная
температура воздуха в мае-августе с коэффициентом корреляции соответственно -0.437
и 0.317. Доля особей с темным щитком без узора 3-го фенотипа, напротив, тем выше,
чем ниже температура воздуха в мае-августе и больше осадков в этот период, с коэф-
фициентами корреляции соответственно -0.719 и 0.270. Доля особей с неясным узором
щитка 2-го фенотипа возрастает при увеличении количества осадков и температуры в
теплый период с коэффициентами корреляции 0.239-0.310. В большинстве популяций
преобладали особи 1-го фенотипа; в популяциях из Тамбовской обл. и Ставропольско-
го края - 2-го. Количество особей 3-го фенотипа было наибольшим в популяциях из
Алтайского края, Самарской и Белгородской областей (20-26 %), 4-го - в популяциях
из Ставропольского края и Волгоградской обл. в посевах пшеницы на светло-каштано-
вых и каштановых почвах (11-15 %). Южнее в горах, предгорьях и оазисах Передней и
Средней Азии доля светлых особей 4-го фенотипа в популяциях возрастает до 19-21 %
(Павлюшин и др., 2013).
Черепашка-мавр, вредная и влаголюбивая черепашки и, вероятно, другие виды
клопов-черепашек рода Eurygaster образуют сходные ряды фенотипической изменчи-
вости окраски тела (см. рис. 2-4). По мнению В. А. Павлюшина с соавт. (Павлюшин
713
и др., 2008), проявление полиморфизма в окраске щитка у родственных видов клопов-
черепашек соответствует закону гомологических рядов наследственной изменчивости
признаков Вавилова (1987). Бурлакой (2016) описаны фенотипы Eu. maura по окрас-
ке щитка, которые оказались такими же, как у Eu. integriceps. Ею проанализированы
показатели 23 морфометрических признаков у этих видов, и различия по ним между
фенотипами оказались недостоверными.
Генетический анализ популяций вредной черепашки из Белгородской обл. проведен
Снегиным и Курносовой (2013) с использованием фенетического подхода и межми-
кросателлитных маркеров ДНК (ISSR). Всего были проанализированы генотипы 121
особи и выявлены 16 локусов по сочетанию доминантных и рецессивных аллельных
генов, оценен уровень их гетерозиготности, проведено сравнение изменчивости ло-
кусов ДНК и окраски особей в популяциях этого вида. Этими авторами установле-
но, что вариации окраски тела обеспечиваются модификационной (фенотипической)
изменчивостью, подверженной действию естественного отбора, а полиморфизм ДНК
в популяциях поддерживается генетико-автоматическими процессами. Это под-
тверждает, что различные типы окраски тела клопов-черепашек обусловлены фено-
типической изменчивостью в связи с приспособлениями к экологическим условиям
среды обитания и их изменениям (Павлюшин и др., 2008; Скребцова, 2009).
Клопы-черепашки рода Eurygaster - сравнительно крупные насекомые с длиной тела
8-13 мм, хорошо, но неохотно летают, имеют лазательно-ходильные ноги, отличаются
слабыми бегательными способностями; питаются открыто на листьях, стеблях и особен-
но на генеративных органах преимущественно злаков в фазы молочной, молочно-воско-
вой, восковой и полной спелости в светлое время суток. К первичным кормовым расте-
ниям вредной черепашки относятся мягкая и твердая яровая и озимая пшеницы, рожь,
ячмень, овес. По всему ареалу этот вид развивается в одном поколении в году с зимовкой
взрослых особей в сухих местах в подстилке в лесополосах, прилежащих к полям лесах
и в садах. Миграция вредной черепашки на поля после зимовки начинается при сред-
несуточной температуре воздуха 10-14 °C в конце апреля - первой половине мая (Ан-
дреев, 1940; Шумаков, Виноградова, 1958; Иванцова, 2013). Весной клопы вначале
заселяют посевы озимой пшеницы, которые находятся в фазе кущения, а затем мигри-
руют на всходы яровых зерновых культур. Основная масса клопов мигрирует с полей в
фазе полной спелости пшеницы во второй половине июля и в начале августа. Суточная
активность имаго вредной черепашки в полевых условиях изучалась в Северном Иране
с помощью круглосуточного слежения в течение 3-4 дней за питавшимися на коло-
сьях 6 взрослыми особями, помеченными изотопом тантала Ta182, в начале июня в фазу
молочной спелости и за другими 6 особями в конце июня в фазу восковой и полной
спелости пшеницы. В начале июня клопы большую часть времени проводили на расте-
ниях, где питались в светлое время суток на колосьях зеленых растений с максимумом
активности в 8 ч. утра; их активность снижалась к середине дня и вновь возрастала
в его второй половине. Питание клопов почти прекращалось на закате, в темноте их
активные движения были редки. В конце июня в фазу восковой и полной спелости
пшеницы клопы питались на колосьях утром и вечером, днем большую часть времени
проводили в трещинах и щелях в почве. К концу периода питания они редко питались
и долгое время находились в щелях почвы, где активно двигались (Banks et al., 1961).
В ночное время и в жаркие часы дня в летний период клопы держатся в нижней части
растений или на поверхности почвы, среди ее комочков. При опасности, имаго черепа-
шек мгновенно падают с колоса на поверхность почвы.
715
Окраска тела черепашек выполняет защитную функцию, делает их менее заметными
в местах обитания, прежде всего на колосьях во время питания и на поверхности почвы,
а также в местах зимовки, является маскировочной, покровительственной, гармонирующей
с фоном (Котт, 1950). Черепашки 1-го фенотипа с наиболее контрастным, в значительной
мере расчленяющим бело-серым или бело-серо-коричневым рисунком щитка малозамет-
ны на стеблях и колосьях пшеницы; светлая окраска тела особей 4-го фенотипа делает их
малозаметными на беловато-желтых колосьях созревающей пшеницы. Однотонная темная,
серая или серо-коричневая окраска щитка особей 3-го фенотипа сходна с цветом поверхно-
сти черноземных почв. Окраска тела клопов-черепашек обусловлена наличием меланинов
в кутикуле и сохраняется после их гибели. Больше всего меланинов в утолщенной кутикуле
щитка, переднеспинки и кожистой части крыльев. Основной цветовой спектр меланинов от
светло-бежевой, охряной, красноватой до коричневой, темно-коричневой и черной окраски
(Росс и др., 1985). В лесостепи Самарской обл. доля особей черного цвета в популяциях
Eu. integriceps и Eu. maura составляет в среднем 1.0-1.3 %, австрийской - 1.5-1.8 % (Бурла-
ка, 2016); в популяциях вредной черепашки в лесостепи Харьковской обл., по многолетним
данным (1969-2001 гг.), в годы депрессий - 0.5-2.0 %, в годы вспышек численности в цен-
тре локальных популяций - до 10-12 %, а по их периферии - до 15.4 % (Белецкий, 2011).
Коэффициент корреляции между процентной долей меланистов в популяциях вредной че-
репашки и коэффициентом ее размножения составил 0.716. В окрестностях Тегерана доля
меланистов в популяции вредной черепашки около 0.3 %; в лабораторных условиях при
24 °C период развития личинок там составлял у черепашек типичного фенотипа 42.1-42.6,
а у меланистов - 44.0-44.2 дня при продолжительности жизни самцов типичной формы
38.1, самок 42.6, а самцов и самок меланистов соответственно 49.1 и 50.2 дня. Самки мела-
нистов откладывали яиц в 1.5 раза меньше, чем самки типичной формы (Mohaghegh, 2008).
Причины естественного меланизма у насекомых исследованы недостаточно.
В естественном отборе фенотипов имаго клопов-черепашек по окраске тела важную
роль играют их взаимоотношения с хищниками и паразитами, которые остаются слабо
изученными. У вредной черепашки наибольшая смертность наблюдается на стадиях
яйца и личинок 1-го и 2-го возрастов. Она значительно снижается у личинок 3-5-го
возрастов и особенно у имаго (Tafaghodinia et al., 2004) весной и осенью в местах их
зимовки, где они становятся объектом добычи крупных жужелиц родов Calosoma
Weber, Carabus Linnaeus и Pterostichus Bonelli, насекомоядных птиц (грачей, сорок,
ворон, скворцов, дроздов и др.) (Благосклонов, 1972; Воронин и др., 1988). На поверх-
ности почвы большое число имаго черепашек уничтожается крупными напочвенными
жужелицами и пауками (Титова, Егорова, 1978).
На взрослых клопах-черепашках паразитируют мухи-фазии (Tachinidae): золотистая
Clytiomyia helluo (F.), серая Allophora subcoleoptrata (L.), пестрая Phasia crassipennis
(F.) и черная Helomyia lateralis (Meig.). Самки 1-го поколения золотистой фазии за-
ражают перезимовавших клопов на растениях, самки 2-го поколения - молодых кло-
пов, реже личинок старших возрастов. Мухи откладывают яйца на их глаза и верхнюю
часть головы, зимуют личинки 2-го возраста в теле черепашек. У серой фазии одно
поколение в году, зимуют пупарии в почве, яйца откладывают в тело клопа, в крыло-
вые мышцы. Заражению фазиями имаго вредной черепашки предшествуют поиск ими
местообитания хозяина, поиск хозяина в пределах местообитания и выбор хозяина.
В поиске местообитания хозяина важная роль принадлежит химическим стимулам,
связанным с кормовыми растениями хозяев (Викторов, 1967, 1976). К механическим
защитным реакциям клопов относятся их попытки избежать заражения: стремление
716
отпугнуть самку фазии работой крыльев, быстро скрыться среди растительности. Если
это не удается, клоп может, резко отцепившись от растения, упасть на почву. Такие по-
веденческие реакции позволяют ему избежать заражения (Аксененко, Гапонов, 2016).
Зараженный клоп первое время ведет себе крайне возбужденно: быстро двигается, ра-
ботает крыльями, пытается всячески сбросить с себя яйцо. Кроме того, у клопов име-
ются механизмы физиологической защиты, связанные с реакциями внутренней среды
организма в ответ на отложенное яйцо или проникновение паразитоида.
На нижней боковой поверхности заднегруди взрослых особей вредной черепашки
открываются протоки пахучих желез. В их выделениях у самцов и самок обнаружены
7 химических веществ: (E)-2-гексанал ((E)-2-Hexenal)) (аттрактант в низких и репел-
лент - в высоких концентрациях, выполняет две основные функции: защитная при
нападении хищников и феромонные контакты между особями при их скоплениях; ток-
сичен для яиц двукрылых, возможно, обеспечивает защиту клопов от заражения яйца-
ми тахинид); 2(5H)-фуранон (2(5H)-Furanone) (препятствуют поражению клопов гриб-
ными и бактериальными инфекциями); 2-гексен-1-ол ацетат (2-Hexen-1-ol, acetate),
лимонен (Limonene) (обладают антибактериальным действием), 5-децин (5-Decyne)
(биологические функции слабо изучены), тридекан (Tridecane) (одно из основных
химических веществ, выделяемых насекомыми при защите от хищников), нонадекан
(Nonadecane) (оказывает раздражающее и репеллентное действие при защите от хищ-
ников). Из них около 95 % приходится на (E)-2-гексанал и тридекан, которые имеют
наибольшее значение в критические периоды жизненного цикла вредной черепашки,
а именно в конце периода питания, когда клопы скапливаются перед уходом на зимов-
ку, и весной после зимовки при массовой концентрация клопов на ближайших полях,
с которых они расселяются на соседние поля (Hassani et al., 2010).
Таким образом, для клопов-черепашек рода Eurygaster характерна модификацион-
ная (фенотипическая) изменчивость особей в популяциях по окраске щитка. У разных
видов этого рода существуют сходные ряды фенотипических форм в связи с суще-
ствованием в одних и тех же или близких условиях и однотипностью их эколого-био-
логических особенностей. Формированию этих форм способствуют существование
в неоднородных условиях (открыто на надземных органах кормовых растений при пи-
тании, открыто или полуоткрыто на поверхности и в верхних слоях почвы и подстилки
при суточных миграциях, скрыто во вторичных укрытиях при сезонных миграциях на
зимовку и в период летней засухи) и взаимоотношения с кормовыми растениями, хищ-
никами и паразитами. Окраска тела клопов-черепашек выполняет защитную функцию, де-
лает их менее заметными в местах их обитания, прежде всего на колосьях во время питания
и на поверхности почвы, а также в местах зимовки и эстивации. Она бывает маскировоч-
ной, покровительственной, гармонирующей с фоном, либо расчленяющей.
МАТЕРИАЛ И МЕТОДИКА
Исследования по фауне и динамике численности клопов-черепашек проводились в 1996-
2018 гг., по внутривидовой изменчивости их популяций в 2002-2004 и 2018 гг. на территории
Кинельского р-на в лесостепной зоне Самарской обл. в окр. пгт. Усть-Кинельский. Внутриви-
довая изменчивость имаго клопов-черепашек учитывалась по фенетической методике, предло-
женной Фасулати (2005, 2010), за основу при выделении фенотипов были взяты рисунок щитка
и его окраска (Бурлака, 2016). Клопы собирались в агроценозах ярового ячменя, озимой и яро-
вой пшеницы в течение всей вегетации культурных растений. Всего было учтено 2873 особи
вредной черепашки и 1297 особей черепашки-мавра. Анализ внутрипопуляционной структуры
717
самцов и самок вредной черепашки и черепашки-мавра проведен в 2002 г. Показатель феноти-
пического сходства популяций клопов-черепашек на разных кормовых культурах (r) определяли
по методике Животовского (1982), формула расчета:
r=√p1q1+√p2q2+...+√pnqn,
где p1, p2, … pn - частоты в выборке в долях от единицы из первой популяции, q1, q2, … qn - из
второй популяции. Статистическая обработка полученных данных проводилась методами дис-
персионного и корреляционного анализов.
Вегетационный период 2002 г. (апрель-сентябрь) в целом можно охарактеризовать как засуш-
ливый, дефицит влаги составил 100.9 мм по сравнению со среднемноголетними данными. Тем-
пература воздуха была близка к норме. Вегетационный период 2003 г. был менее засушливым
с суммой осадков на уровне среднемноголетних значений. Вегетационный период 2004 г. по тем-
пературным условиям и по степени увлажнения был теплым и влажным: температурный режим
периода вегетации был повышен на 1.2 °C, количество осадков превысило норму на 44.4 мм.
2018 г. отличался холодной зимой и влажным прохладным апрелем, что отрицательно сказалось
на выходе черепашек из зимовки. Посев пшеницы был проведен лишь в конце первой декады
мая, а май и июнь были засушливыми - выпало соответственно 16 и 7 мм осадков, что было
неблагоприятно для развития пшеницы, и численность имаго вредной черепашки была низкой.
Однако в июне гидротермические условия для откладки яиц и развития личинок черепашки
были благоприятными, и ко второй половине июля ее численность стала восстанавливаться.
РЕЗУЛЬТАТЫ
В лесостепи Самарской обл. выявлены 4 вида рода Eurygaster: вредная черепашка
(Eu. integriceps), черепашка-мавр (Eu. maura), австрийская (Eu. austriaca) и влаго-
любивая (Eu. testidinaria) черепашки. Во все годы исследований (1996-2018 гг.), за
исключением 1999 г., в посевах пшеницы преобладала вредная черепашка. На ее долю
приходилось в 1999 г. 26 %, в остальные годы - 33-86 % общего количества учтенных
клопов-щитников. Черепашка-мавр входила в состав доминантов в 1996-2004 гг., когда
на ее долю приходилось 13-48 % общего количества клопов-щитников, с максималь-
ным участием в 1999 г. После 2006 г. доля Eu. maura в населении клопов-щитников
в посевах пшеницы и ячменя снизилась до 1-10 %. Наибольшее участие в нем австрий-
ской черепашки отмечено в 2000 г. (36 %). В 1996-1999 и 2002-2009 гг. она относилась
к субдоминантам (2-10 %), а в последующие годы - к второстепенным видам (менее
1 %). В 2018 г. на долю Eu. maura приходилось 2.0, Eu. austriaca - 1.3 % общего коли-
чества учтенных клопов-черепашек. Единичные особи влаголюбивой черепашки со-
браны в посевах мягкой яровой пшеницы во влажном 2011 г. (Бурлака, Каплин, 2015).
В популяциях Eu. integriceps, Eu. maura и Eu. austriaca по окраске щитка были выде-
лены 5 фенотипов, встречались также особи с нечеткими признаками (Бурлака, 2016).
Клопы 1-4-го фенотипов имели окраску щитков, сходную с описанной Фасулати (2005).
Клопы 5-го фенотипа со щитком почти черной окраски без рисунка (меланистическая
форма) (см. рис. 4) наиболее часто встречались в популяции австрийской черепашки.
В агроценозах озимой и яровой пшеницы были выявлены 5 фенотипов клопов-чере-
пашек по окраске щитков, ячменя - 3 фенотипа (табл. 2, 3). В посевах всех кормовых
культур в популяциях Eu. integriceps и Eu. maura преобладали особи первого фенотипа,
значительное участие в них принимали также особи 2-го и 3-го фенотипов; клопы 4-го
718
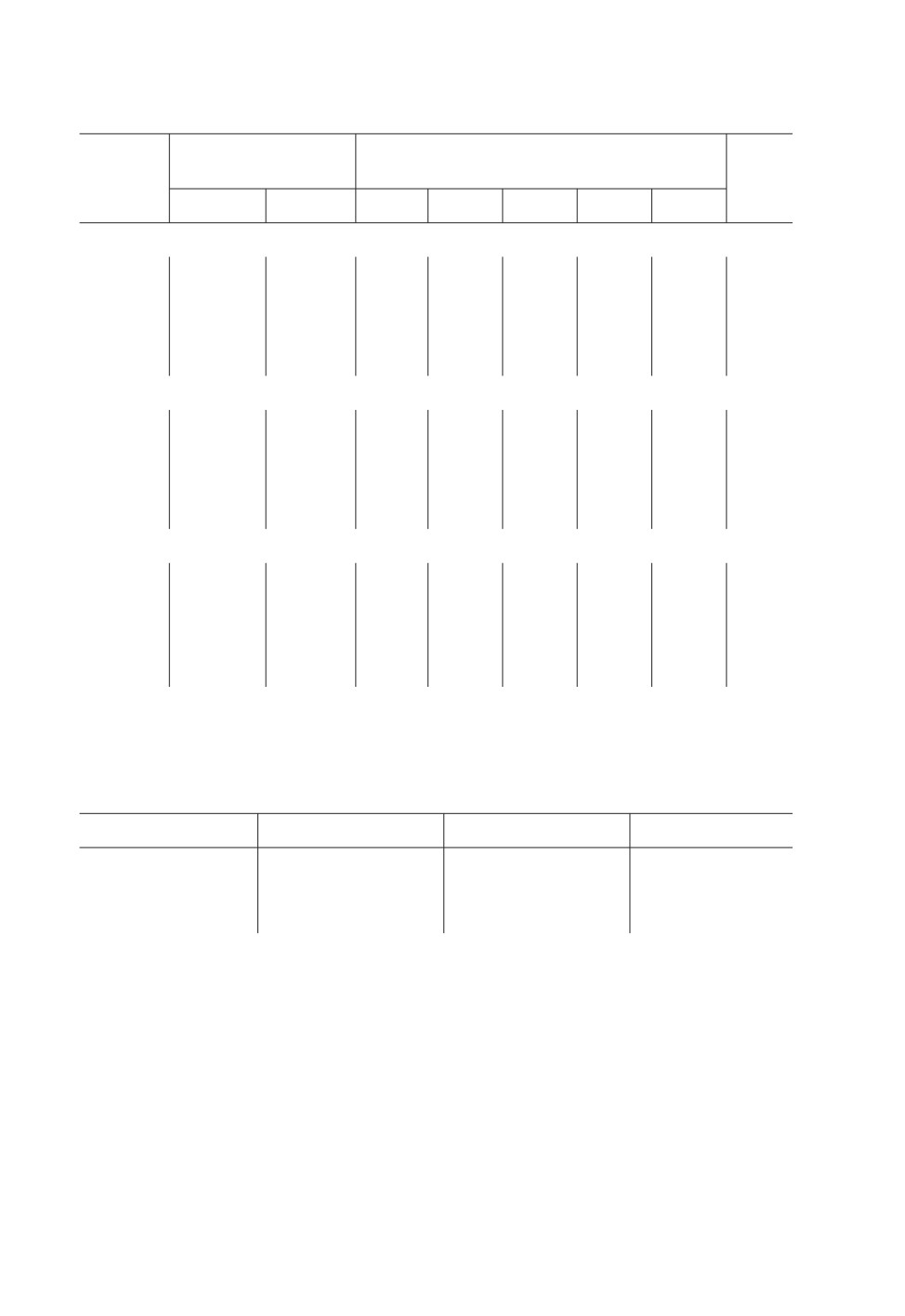
Таблица 2. Фенотипическая структура популяций Eurygaster integriceps (Put.) по окраске щитка
имаго в посевах зерновых злаковых культур
Проанализировано:
Доля особей имаго по каждому фенотипу, %
Год
НСР05
учетов
особей
1
2
3
4
5
Озимая пшеница
2002
8
471
34.4
31.2
33.5
0.2
0.6
16.1
2003
12
251
57.8
18.3
21.5
1.2
1.2
13.4
2004
7
311
60.5
15.4
21.9
0.3
1.9
12.3
Всего
27
1033
50.9
21.6
25.6
0.6
1.2
Яровая пшеница
2002
7
611
47.1
33.6
18.7
0.2
0.5
22.4
2003
11
166
59.0
19.3
19.9
0.6
1.2
17.3
2004
5
577
59.6
17.0
20.6
0.7
2.1
19.2
2018
12
393
51.0
27.2
19.4
2.1
0.3
16.4
Всего
35
1747
54.2
24.3
19.6
0.9
1.0
Ячмень
2002
5
83
38.6
43.4
18.0
0
0
34.2
2003
4
10
80.0
10.0
10.0
0
0
18.5
Всего
9
93
59.3
26.7
14.0
0
0
НСР05
19.3
14.8
9.1
0.7
1.0
П р и м е ч а н и е. Фенотипы 1-5 по: Фасулати, 2005; Бурлака, 2016 (см. текст, с. 709).
и 5-го фенотипов в посевах озимой и яровой пшеницы встречались единично, в посе-
вах ячменя особи этих фенотипов не регистрировались.
Существенных различий в фенотипической структуре популяций Eu. integriceps
и Eu. maura на разных кормовых культурах не выявлено (табл. 4). Статистическая об-
работка данных измерений внешних морфометрических показателей и массы тела у
клопов Eu. integriceps и Eu. maura разных фенотипов показала, что у них нет статисти-
чески достоверных различий (Бурлака, 2016).
Структура популяций клопов-черепашек существенно различалась по годам. В по-
пуляции вредной черепашки наибольшая доля особей 1-го фенотипа в посевах озимой
и яровой пшеницы отмечена в теплом и влажном 2004 г., 2-го - в засушливом 2002 г.,
3-го - в посевах озимой пшеницы в засушливом 2002 г., а в посевах яровой пшени-
цы она в годы исследований была примерно одинаковой. В популяции Eu. maura наи-
большая доля особей 1-го фенотипа в посевах озимой пшеницы была также в теплом
и влажном 2004 г., а в посевах яровой пшеницы - в более засушливом 2003 г.; 2-го
фенотипа - в посевах озимой и яровой пшеницы в засушливом 2002 г.; 3-го - в менее
засушливом 2003 г.
719
Таблица 3. Фенотипическая структура популяций Eurygaster maura (L.) по окраске щитка
имаго в посевах зерновых злаковых культур
Фенотипы и доля особей имаго по каждому из
Проанализировано
них, %
Год
НСР05
учетов
особей
1
2
3
4
5
Озимая пшеница
2002
8
285
34.0
43.9
21.8
0
0.3
14.1
2003
10
191
60.7
16.8
20.4
0.5
1.6
8.8
2004
7
85
74.1
12.9
11.8
0
1.2
10.0
Всего
25
561
56.3
24.5
18.0
0.2
1.0
Яровая пшеница
2002
7
454
55.1
29.7
14.8
0.2
0.2
11.5
2003
11
107
59.8
16.8
22.4
0
0.9
19.3
2004
5
111
53.2
22.5
20.7
0.9
2.7
7.2
Всего
23
672
56.0
23.0
19.3
0.4
1.3
Ячмень
2002
5
50
52.0
38.0
10.0
0
0
18.1
2003
4
14
85.7
7.1
7.1
0
0
20.8
Всего
9
64
68.9
22.6
8.6
0
0
НСР05
27.9
16.8
10.2
0.8
1.7
П р и м е ч а н и е. Столбцы 1-5 - то же, что в табл. 2.
Таблица 4. Показатель фенотипического сходства (r) популяций черепашек Eurygaster
integriceps (Put.) (слева, ниже диагонали) и Eu. maura (L.) (справа, выше диагонали) по окраске
щитка имаго в посевах зерновых злаковых культур
Культура
Озимая пшеница
Яровая пшеница
Ячмень
Озимая пшеница
-
0.999
0.982
Яровая пшеница
0.998
-
0.977
Ячмень
0.979
0.988
-
Анализ внутрипопуляционной фенотипической структуры в зависимости от пола
Eu. integriceps и Eu. maura показал, что существенных отличий между самцами и сам-
ками по этому показателю не наблюдается. Отмечены высокие коэффициенты феноти-
пического сходства между внутривидовой структурой выборок самок и самцов в попу-
ляциях Eu. integriceps и Eu. maura на всех исследуемых кормовых культурах.
Таким образом, в лесостепи Самарской обл. в популяциях клопов-черепашек в агро-
ценозах озимой и яровой пшеницы выявлены особи 5, ячменя - 3 фенотипов по окра-
ске щитков. В посевах пшеницы и ячменя в популяциях Eu. integriceps и Eu. maura
в среднем доминируют особи 1-го фенотипа, большую долю составляют также особи
720
2-го и 3-го фенотипов. Наибольшее сходство популяций вредной черепашки и чере-
пашки-мавра отмечено в посевах озимой и яровой пшеницы.
СПИСОК ЛИТЕРАТУРЫ
Аксененко Е. В., Гапонов С. П. 2016. Репродуктивная стратегия фазиин (Diptera: Tachinidae: Phasiinae) -
паразитоидов полужесткокрылых (Heteroptera). Современные проблемы паразитологии и эпизоото-
логии. Сборник статей IX Всероссийской научно-практической конференции, посвященной 85-летию
создания кафедры паразитологии и зпизоотологии Воронежского ГАУ (Россия, Воронеж, 4 декабря
2015 г.). Воронеж: Воронежский ГАУ, с. 3-14.
Андреев Л. Л. 1940. Методы борьбы с клопами-черепашками. Пятигорск: 1-я государственная типография,
32 с.
Белецкий Е. Н. 2011. Массовые размножения насекомых. История, теория, прогнозирование. Харьков: Май-
дан, 172 с.
Благосклонов К. Н. 1972. Охрана и привлечение птиц. М.: Просвещение, 240 с.
Бурлака Г. А. 2016. Морфотипическая изменчивость популяции клопов-черепашек в лесостепи Самарской
области. Известия Самарской государственной сельскохозяйственной академии 4: 21-25.
Бурлака Г. А., Каплин В. Г. 2015. Биоэкологическое обоснование защиты зерновых злаков от хлебных клопов
(надсемейства Pentatomоidea) в лесостепи Среднего Поволжья. Кинель: РИЦ СГСХА, 145 с.
Вавилов Н. И. 1987. Происхождение и география культурных растений. Л.: Наука, 440 с.
Викторов Г. А. 1967. Проблемы динамики численности насекомых на примере вредной черепашки. М.: На-
ука, 271 с.
Викторов Г. А. 1976. Экология паразитов-энтомофагов. М.: Наука, 152 с.
Вилкова Н. А., Нефедова Л. И., Капусткина А. В. 2014. Внутривидовая структура локальных популяций
вредной черепашки в экосистемах Северо-Кавказского и Нижневолжского регионов России. Вестник
защиты растений 4: 3-7.
Винокуров Н. Н., Канюкова Е. В., Голуб В. Б. 2010. Каталог полужесткокрылых насекомых (Heteroptera)
азиатской части России. Новосибирск: Наука, 323 с.
Воронин К. Е., Шапиро В. А., Пукинская Г. А. 1988. Биологическая защита зерновых культур от вредителей.
М.: Агропромиздат, 197 с.
Животовский Л. А. 1982. Показатели популяционной изменчивости по полиморфным признакам. В. кн.:
А. В. Яблоков (ред.). Фенетика популяций. М.: Наука, с. 38-44.
Иванцова Е. И. Биоэкология клопа вредная черепашка (Eurygaster integriceps Put.) в условиях Нижнего По-
волжья. Вестник Волгоградского государственного университета. Серия 11. Естественные науки 2
(6): 45-52.
Капусткина А. В., Нефедова Л. И. 2015. Внутривидовая структура локальных популяций вредной черепашки
в Алтайском крае. Вестник защиты растений 3 (85): 30-33.
Киль В. И., Исмаилов В. Я. 2009. Идентификация резистентных к инсектицидам генотив в популяции клопа
вредная черепашка по фенам рисунка и RAPD-маркерам. Агрохимия 1: 38-49.
Котт Х. 1950. Приспособительная окраска животных. М.: Издательство иностранной литературы, 543 с.
Павлюшин В. А., Фасулати С. Р., Вилкова Н. А., Сухорученко Г. И., Нефедова Л. И. 2008. Антропогенная
трансформация экосистем и ее фитосанитрные последствия. СПб.: ВИЗР, 120 с.
Павлюшин В. А., Вилкова Н. А., Сухорученко Г. И., Нефедова Л. И. 2010. Вредная черепашка: распростране-
ние, вредоносность, метода контроля. Защита и карантин растений 1 (Приложение): 53-84.
Павлюшин В. А., Вилкова Н. А., Сухорученко Г. И., Нефедова Л. И. 2013. Фитосанитарная дестабилизация
агроэкосистем. СПб.: НППЛ «Родные просторы», 184 с.
Павлюшин В. А., Вилкова Н. А., Сухорученко Г. И., Капусткина А. В. 2015. Вредная черепашка и другие
хлебные клопы. СПб.: ВИЗР, 280 с.
Росс Г., Росс Ч., Росс Д. 1985. Энтомология. М.: Мир, 572 с.
Скребцова Т. И. 2009. Биоэкологические особенности вредной черепашки (Eurygaster integriceps Put.) и со-
вершенствование мер борьбы с ней в Центральном Предкавказье. Автореферат диссертации на соис-
кание ученой степени кандидата сельскохозяйственных наук. М.: РГАУ-МСХА, 20 с.
721
Снегин Э. А., Курносова К. Д. 2013. Генетическая структура популяций клопа Eurygaster integriceps в ус-
ловиях агроценозов юга Среднерусской возвышенности. Научные ведомости. Серия Естественные
науки 7 (160) (24): 56-61.
Титова Э. В., Егорова Н. С. 1978. Оценка трофической связи пауков с вредной черепашкой Eurygaster
integriceps (Hemiptera, Scutelleridae) путем использования серологического метода исследования. Эн-
томологическое обозрение 57 (2): 284-289.
Фасулати С. Р. 2005. Индикация адаптивных процессов у вредной черепашки Eurygaster integriceps Put.
(Heteroptera, Scutelleridae) методами фенетики популяций. Труды Ставропольского НИИСХ. Ставро-
поль: Аргус, с. 68-73.
Фасулати С. Р. 2010. Формирование внутривидовой структуры у насекомых в условиях агроэкосистем на
примерах колорадского жука Leptinotarsa decemlineata Say, 1824 (Coleoptera, Chrysomelidae) и вред-
ной черепашки Eurygaster integriceps Puton, 1881 (Heteroptera, Scutelleridae). Научный вестник Ужго-
родского национального университета. Серия Биология 29: 13-27.
Шумаков Е. М., Виноградова Н. М. 1958. Экология вредной черепашки. Труды Всероссийского института
защиты растений 9: 9-71.
Banks C. J., Brown E. S., Dezfulian A. 1961. Field studies of the daily activity and feeding behaviour of Sunn Pest,
Eurygaster integriceps Put. (Hemiptera, Scutelleridae) on wheat in North Iran. Entomologia Experimentalis
et Applicata. 4 (4): 289-300.
Göllner-Scheiding U. 2006. Family Scutelleridae Leach, 1815 - shield bugs. In: B. Aukema, Ch. Rieger (eds).
Catalogue of the Heteroptera of the Palaearctic Region. Volume 5. Pentatomomorpha II. Amsterdam: The
Netherlands Entomological Society, pp. 190-227.
Hassani S., Pour R. A., Djozan D., Movahedi F. M. 2010. Compounds in metathoracic glands of adults of the
sunn pest, Eurygaster integriceps (Puton) (Heteroptera: Scutelleridae). Munis Entomology and Zoology 5:
232-239.
Kerzhner I. M. 2003. Notes on synonymy, nomenclature, and distribution of some Palaearctic Coreoidea and
Pentatomoidea (Heteroptera). Zoosystematica Rossica 12 (1): 101-107.
Mohaghegh J. 2008. Comparison of development time and reproduction of typical and melanic Eurygaster
integriceps (Heteroptera: Scutelleridae) under laboratory conditions. Journal of Entomological Society of
Iran 27 (2): 109-126.
Paulian F., Popov C. 1980. Sunn Pest or cereal bug. In: E. Hafliger (ed.). Wheat. Ciba-Geigy Ltd, pp. 69-74.
Tafaghodinia B., Tafaghodinia M., Nouri-Ganbalani G. 2004. Population dynamics system of cereal sunn pest
(Eurygaster integriceps Put.). WSEAS Transactions on Biology and Biomedicine 1 (1): 36-40.
PHENOTYPIC VARIABILITY OF BODY COLORATION IN THE POPULATIONS
OF SCUTELLERID BUGS (EURYGASTER LAPORTE; HETEROPTERA,
SCUTELLERIDAE) IN CEREAL CROPS AND ECOLOGICAL FACTORS
RESPONSIBLE FOR IT
V. G. Kaplin, G. A. Burlaka
Key words: Eurygaster integriceps, Eu. maura, population, phenotype, intraspecific vari-
ability, environmental factors, phytophages, winter wheat, spring wheat, barley.
SUMMARY
Eurygaster bugs, which are pests of cereal crops in Europe and Asia, are characterized by a well-
defined modification (phenotypic) inter-individual variability of the scutellum coloration in populations.
The colour of the scutellerid body performs a protective function, is camouflaged or disruptive, helping
the bug blend in with its environment. The main ecological prerequisites for its formation include the
existence of bugs in inhomogeneous conditions on the aboveground organs of food plants, on the surface
and in the upper layers of the soil and litter during daily migrations, hidden in secondary shelters during
seasonal migrations for the winter and during the summer drought, the relationships with food plants,
predators and parasites. This contributes to the formation of a series of phenotypes in the body color
722
of bugs with transitional forms between them, and their dynamics in space and time due to changes in
living conditions. The leading factors determining the body color of scutellerids are climate, especially
humidity and thermal conditions during development; soil, in particular, its type and color; features of
vegetation; composition and biological characteristics of the main host plants, entomophages of bugs
and relationships between them. In the temperate zone the proportion of bugs with uniformly colored
dark, gray or gray-brown scutellum increases and with a light scutellum decreases in populations of
Eu. integriceps with increasing latitude and longitude, precipitation in May-August, and reducing
air temperature, with the transition from the steppe to forest-steppe, from lighter chestnut soils to
chernozems. The proportion of bugs with light scutellum increases in populations of Eu. integriceps in
the mountains and foothills of Western and Middle Asia. In the forest-steppe of Samara Province, there
are 5 and 3 phenotypes in the color of the scutellum in the scutellerid populations in agrocenosis of
wheat and barley, respectively. Bugs of the 1st phenotype, with a large proportion of scutellerids of the
2nd and 3rd phenotypes, dominate in populations of Eu. integriceps and Eu. maura in wheat and barley
crops. Populations of Eu. integriceps and Eu. maura are most similar in the coloration of the body in
the crops of winter and spring wheat.
723
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}