ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 98, 4, 2019
УДК 595.771: 591.813(575.3)
МОРФОЛОГИЯ ЛИЧИНКИ И СТРУКТУРА КАРИОТИПА ЗВОНЦА
CHIRONOMUS SP. (DIPTERA, CHIRONOMIDAE)
C АРХИПЕЛАГА НОВАЯ ЗЕМЛЯ (РОССИЯ)
© 2019 г. Петрова Н. А., 1*
Жиров С. В., 1* Крашенинников А. Б. 2**
1 Зоологический институт РАН
Университетская наб., 1, С.-Петербург, 199034 Россия
*e-mail: chironom@zin.ru
2 Институт биологических проблем Севера ДВО РАН
Портовая ул., 18, Магадан, 685000 Россия
**e-mail: krasheninnikov2005@yandex.ru
Поступила в редакцию 13.04.2017 г.
После доработки 5.06.2018 г.
Принята к публикации 5.06.2018 г.
Описаны строение личинок и особенности кариотипа звонца Chironomus sp. с о. Южный в ар-
хипелаге Новая Земля. 2n = 6: AB, CD, FEG. Последняя хромосома возникла в результате присо-
единения плеча малой хромосомы G к плечу E и генетического закрепления этой мутации. По-
казано, что этот вид близок к Ch. tuvanicus, который был описан из Тувы (Сибирь), принадлежит
к группе riihimakiensis и относится к числу примитивных видов этого рода.
DOI: 10.1134/S0367144519040099
Архипелаг Новая Земля расположен между Баренцевым и Карским морями на край-
нем северо-востоке Европы и состоит из Северного и Южного островов, разделенных
проливом Маточкин Шар. Общая площадь архипелага около 83 000 км. На большей ча-
сти территории доминируют арктические тундры, на севере распространены полярные
пустыни, на крайнем юге - типичные тундры (Боярский и др., 1994).
Кариологические исследования фауны хирономид в высоких широтах Арктики вы-
явили несколько видов хирономусов (Wülker, 1991; Шобанов, Петрова, 1995; Беляни-
на, Филинкова, 1996; Кикнадзе и др., 1996; Shobanov et al., 2002; Petrova, Zhirov, 2008).
Предварительный анализ морфологии личинок, куколок и имаго с полярных терри-
торий России показал, что род Chironomus представлен в этом регионе 15-20 вида-
ми, среди которых есть ряд неописанных (Зеленцов, Шилова, 1996, Макарченко и др.,
1998; Зеленцов, 2007; Крашенинников, 2013). Большинство этих видов слабо изучено,
но почти у всех личинки относятся к морфотипу salinarius, который характеризуется
отсутствием вентральных и латеральных отростков на VII и VIII брюшных сегмен-
тах. Известно 18 видов с личинками морфотипа salinarius (Shobanov et al., 2002). 9 из
них населяют пресные водоемы в северном полушарии - цитокомплекс thummi, другие
761
6 - пресные водоемы в южном полушарии. В большинстве своем это виды, относящи-
еся к цитокомплексу pseudothummi. Оставшиеся 3 вида населяют морские биотопы.
Chironomus sp., обнаруженный нами на Новой Земле, оказался очень близким
к Сh. tuvanicus, относящимся к группе riihimakiensis, который был найден в Туве и тоже
имеет 2n = 6 (Kiknadze et al., 1992). В данной статье мы описываем личинку звонца
с Новой Земли и структуру его кариотипа.
МАТЕРИАЛ И МЕТОДИКА
Закрытым постановлением № 1559-699 Совета Министров СССР от 31 июля 1954 г. на архи-
пелаге Новая Земля был создан Северный полигон для испытаний ядерного оружия. С момента
создания до 24 октября 1990 г. на полигоне было произведено 130 ядерных испытаний (Думик
и др., 1998). В 1993 г. архипелаг перешел под юрисдикцию администрации Архангельской об-
ласти, став центром сообщества «Культура и Мировой океан» (Боярский и др., 1994), и после
длительного перерыва началось активное изучение биоты Новой Земли отечественными специ-
алистами.
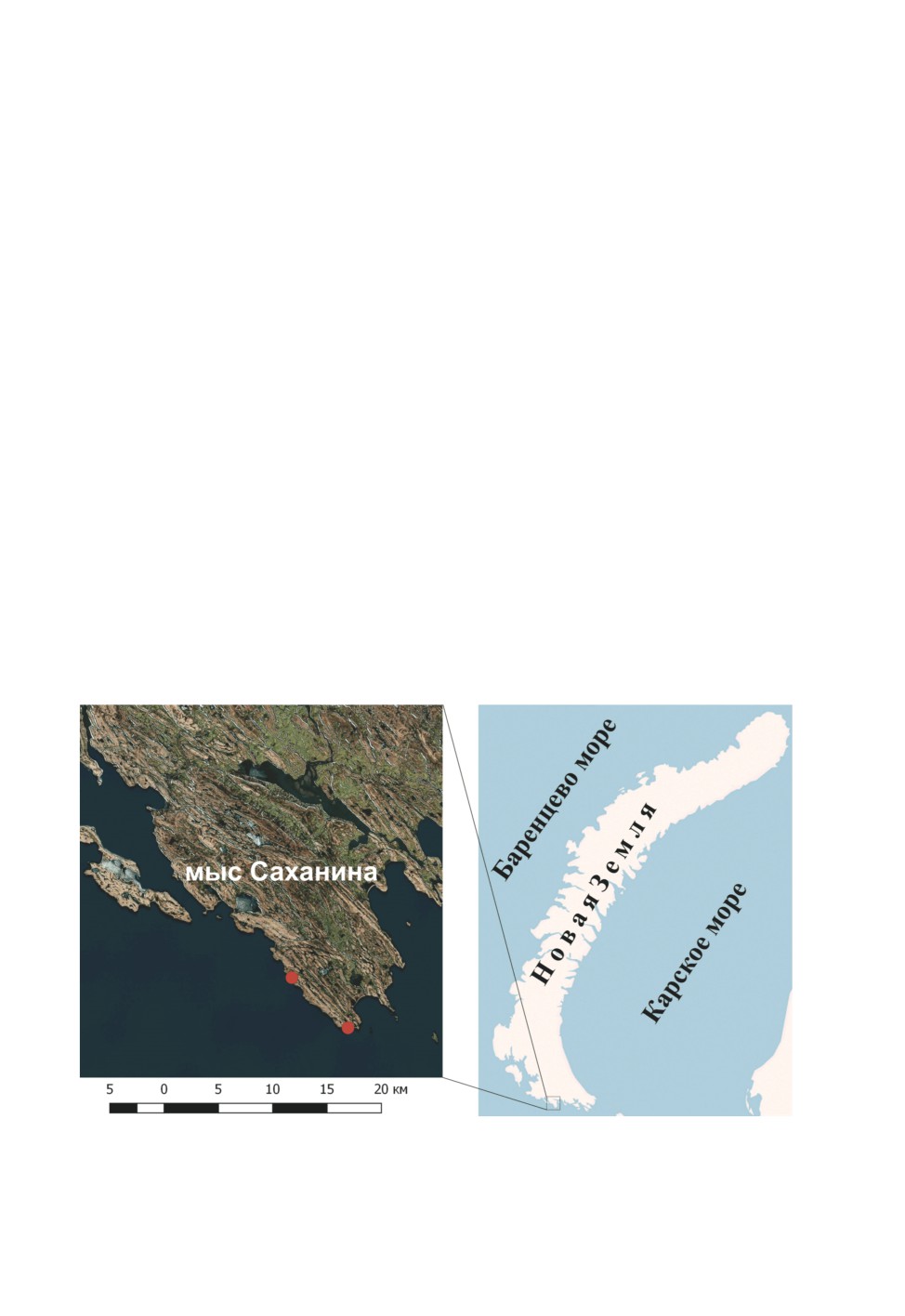
В июле и августе 2015 г. в окрестностях мыса Саханина о. Южный работала комплексная экс-
педиция, организованная Научно-экспедиционным центром Ассоциации «Морское наследие».
В ходе этой экспедиции в двух внутренних пресных водоемах о. Южный был собран вид, услов-
но названый Chironomus sp. и оказавшийся близким к Ch. tuvanicus. Материал собирался 28 июля
2015 г. в окрестностях мыса Саханина в небольшом озере около избы Белой (5 м над ур. м.)
и 5 августа 2015 г. в озере около балков (35 м над ур. м.) (сбор А. Б. Крашенинникова). Приве-
дена карта-схема мест сбора (pис. 1), созданная в программе QGIS 2.14.3-Essen с привлечением
Тонкий слой ила взмучивался с подлежащего слоя крупного щебня на глубине менее 0.5 м
и помещался в скребок. Пробы сразу разбирались на берегу. Личинок фиксировали на месте сбо-
Рис. 1. Места сбора Chironomus sp. (красные кружки), мыс Саханина, о. Южный,
архипелаг Новая Земля.
762
ра в жидкости Кларка (1 часть ледяной уксусной кислоты и 3 части 96%-ного этилового спирта).
Изучено 28 личинок.
Морфологические препараты частей головной капсулы готовили в жидкости Фора. Опреде-
ление проводили в лабораторных условиях по морфологическим признакам с использованием
общих ключей (Панкратова, 1983; Pinder, Reiss, 1983; Kiknadze et al., 1992, 1994). При описании
личинок применялась обычная терминология (Saether, 1979; Cranston, Reiss, 1983). Препара-
ты политенных хромосом готовили по ацетоорсеиновой методике (Чубарева, Петрова, 1982).
Кариотип Сh. tuvanicus использовали как стандарт для картирования хромосом Chiromous sp.
При кариологическом сравнении эти виды сокращенно обозначены как «Ch. sp.» и «tuv».
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Chironomus sp.
Личинка 4-го возраста
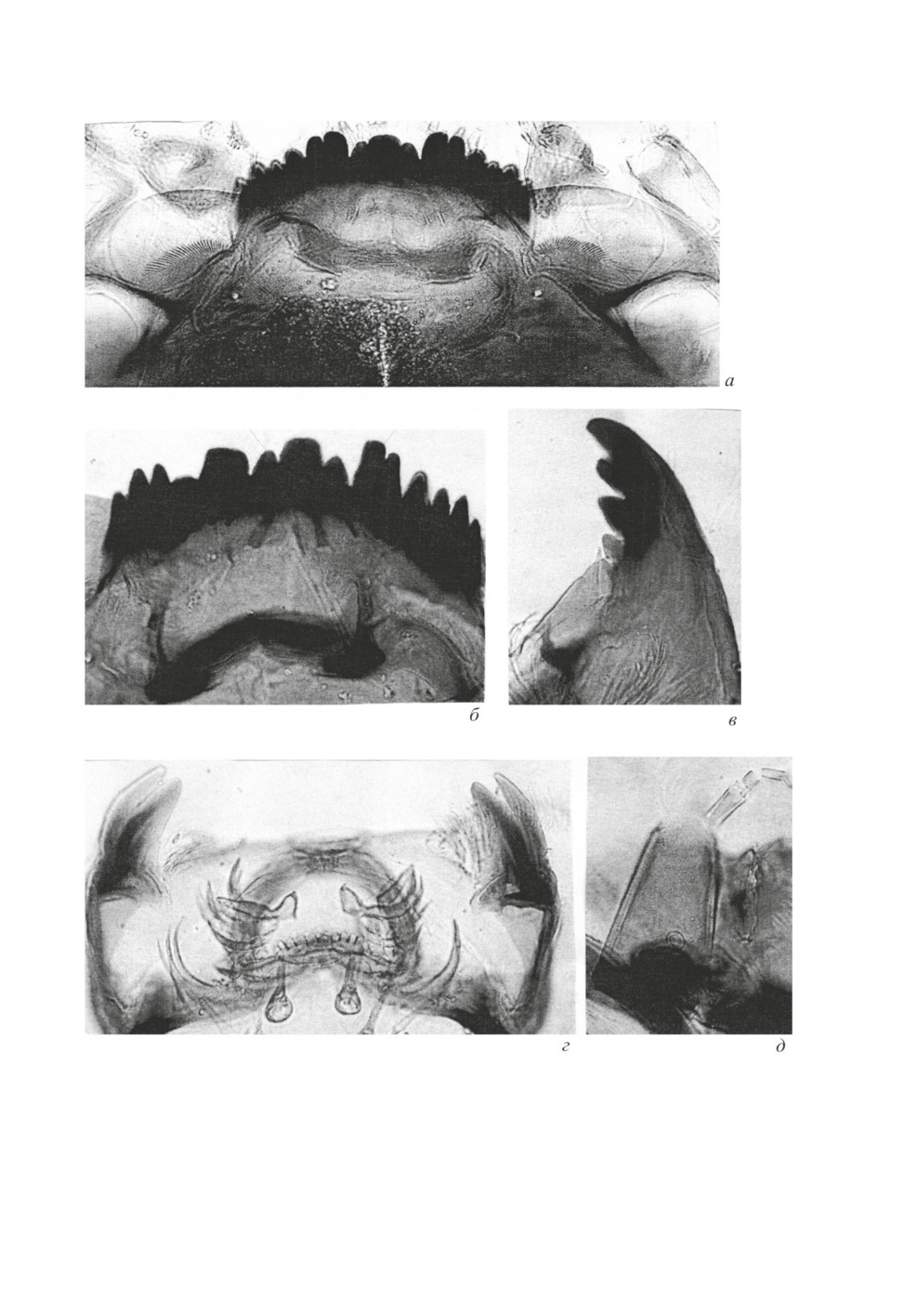
Тело (рис. 2, а-д) длиной 12-14 мм, красного цвета. Вентральные и латеральные отростки
на брюшке отсутствуют (морфотип salinarius).
Головная капсула светло-желтая. Фронтоклипеальная апотома светло-коричневая, с отчет-
ливыми коричневыми продольными полосами. Глазные пятна небольшие, верхнее округлое
и простое, нижнее - сложное. Гулярный склерит темно-коричневый. Внутренняя поверхность
вентральных пластинок бороздчатая. Антенна короткая, 5-члениковая, 1-й членик массивный,
остальные значительно тоньше и короче. В первой трети базального членика располагается
кольцевой орган. Щетинка антенны достигает основания 5-го членика, AR - 1.43. Лаутерборно-
вы органы мелкие, почти незаметные.
Ментум состоит из тройного срединного зубца и 6 пар боковых. Зубцы ментума черные, не-
много закругленные. Срединный зубец широкий, значительно ниже 1-го бокового, придаточ-
ные - почти вдвое ниже. 1-е и 2-е боковые зубцы слиты в верхней четверти. Высота боковых
зубцов ментума постепенно уменьшается от середины к краям. На рис. 2, б зубцы 2-4, по-види-
мому, обломаны. 4-я пара зубца ментума почти вдвое ниже соседних, - это характерный признак
группы видов riihimakiensis. Вентральные пластинки серо-желтые, их внутренние края достига-
ют 2-й боковой пары зубцов, передний край гладкий (рис. 2, б).
Мандибула с 4 постепенно уменьшающимися к основанию зубцами; первые 3 темно-коричне-
вые, 4-й - светло-коричневый с желтизной. Внешний зубец одиночный, небольшой. Субденталь-
ная щетинка ланцетовидная, достигает середины 4-го зубца. У внутренней щетинки сложное
ветвистое строение.
Премандибула с 2 зубцами, внутренний зубец намного шире внешнего, узкого и тонкого. Эпи-
фарингеальный гребень состоит из 8 зубцов.
Имаго и куколка Chironomus sp. в месте сбора не обнаружены.
ЭКОЛОГИЯ
Личинки Chironomus sp. обитают в водоеме с медленным течением на небольших
глубинах (0.5-1.0 м). Озеро в окрестностях избы Белой подвержено периодическим
забросам соленой воды из Баренцева моря. Соленость в самом озере неравномерна
и определялась нами органолептически. Наибольшее количество личинок хирономид
найдено в пресной зоне.
КАРИОТИП
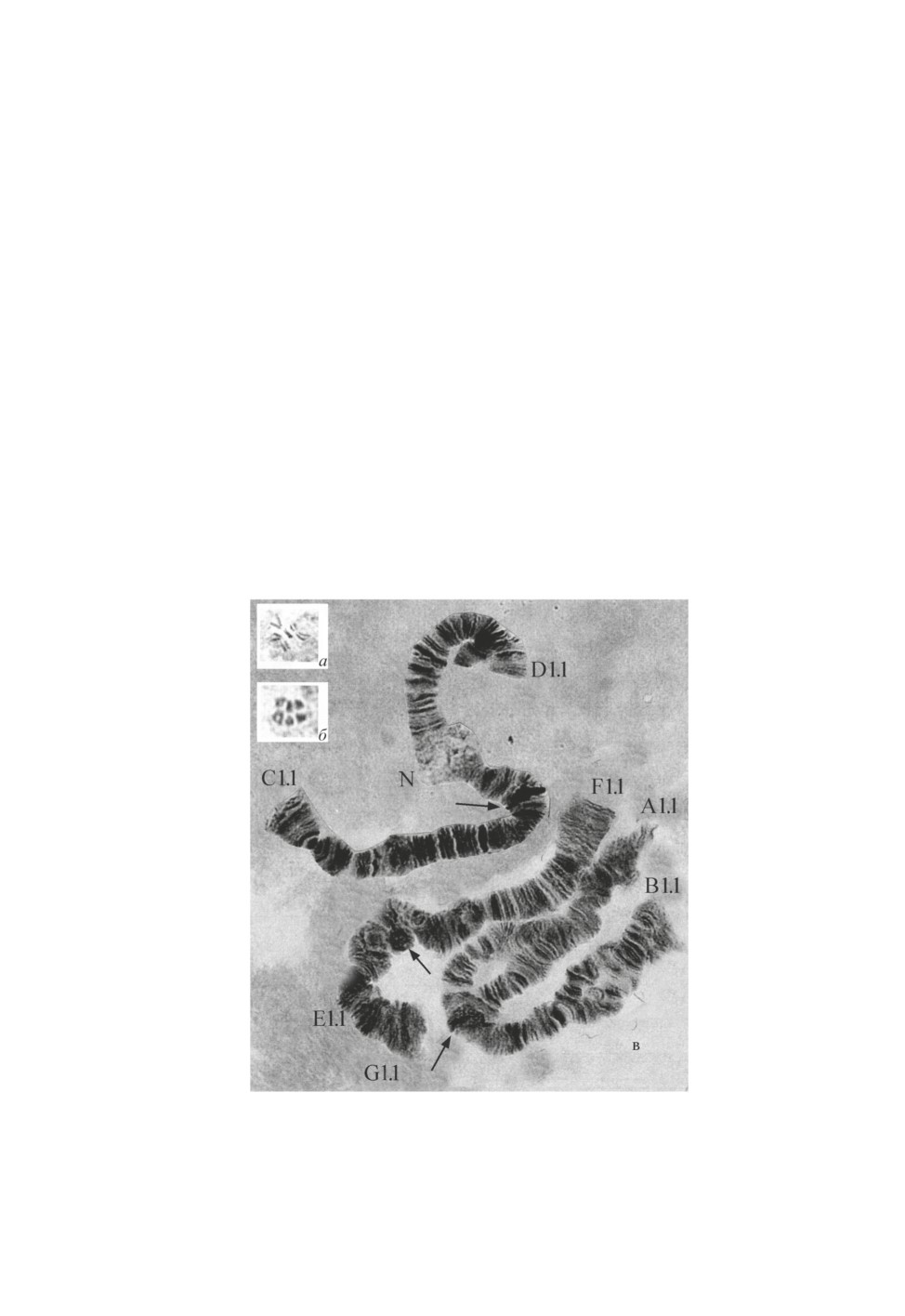
2n = 6. На метафазных и анафазных пластинках различимы 3 пары митотических
хромосом (рис. 3, а, б), Клетки слюнных желез содержат 3 пары плотно сконъюгиро-
763
Рис. 2. Chironomus sp., личинка.
а, б - мандибула с вентральными пластинками; в - мандибула; г - премандибулы и эпифарингеальный
гребень; д - усик.
764
ванных гомологов (рис. 3, в). Комбинация хромосомных плеч AB, CD, FEG. По длине
первые две хромосомы равны, хромосома FEG несколько длиннее: AB = CD ˂ FEG
(рис. 3). Центромерные диски хромосом хорошо выражены в виде массивных гете-
рохроматиновых блоков, часто меняющих свои очертания и образующих рыхлые взду-
тия. На месте прикрепления плеча G к хромосоме EF присутствовало второе ядрышко
(рис. 4, плечо E + G). У одной личинки в разных хромосомах представлены разные
типы центромер, которые хорошо отличаются друг от друга (рис. 3, 4). В хромосоме
FEG иногда наблюдалась гетерозиготность по содержанию прицентромерного гете-
рохроматина. Хромосомы AB и CD метацентрические, FEG - субметацентрическая;
два ядрышка локализованы в плече D и в хромосоме FEG. Это четкий маркер видов
группы riihimakiensis. В середине плеча G обычно функционирует единственное коль-
цо Бальбиани.
Наблюдается полиморфизм по гетерозиготным хромосомным перестройкам, кото-
рые затрагивают как небольшие, так и значительные участки хромосом.
Сравнение рисунков дисков политенных хромосом у Chironomus sp.
и Ch. tuvanicus
Chironomus tuvanicus относится к примитивной группе riihimakiensis рода Chironomus
и имеет 2n = 6 (Kiknadze et al., 1992, 2016). Плечо G присоединилось к теломере пле-
Рис. 3. Кариотип Сhironomus sp. (2n = 6): а, б - анафазные и в - политенные (A1.1, B1.1,
D1.1, C1.1, D1.1, E + G1.1, F1.1) хромосомы; BR - кольцо Бальбиани; N - ядрышко.
Стрелками показаны центромеры.
765
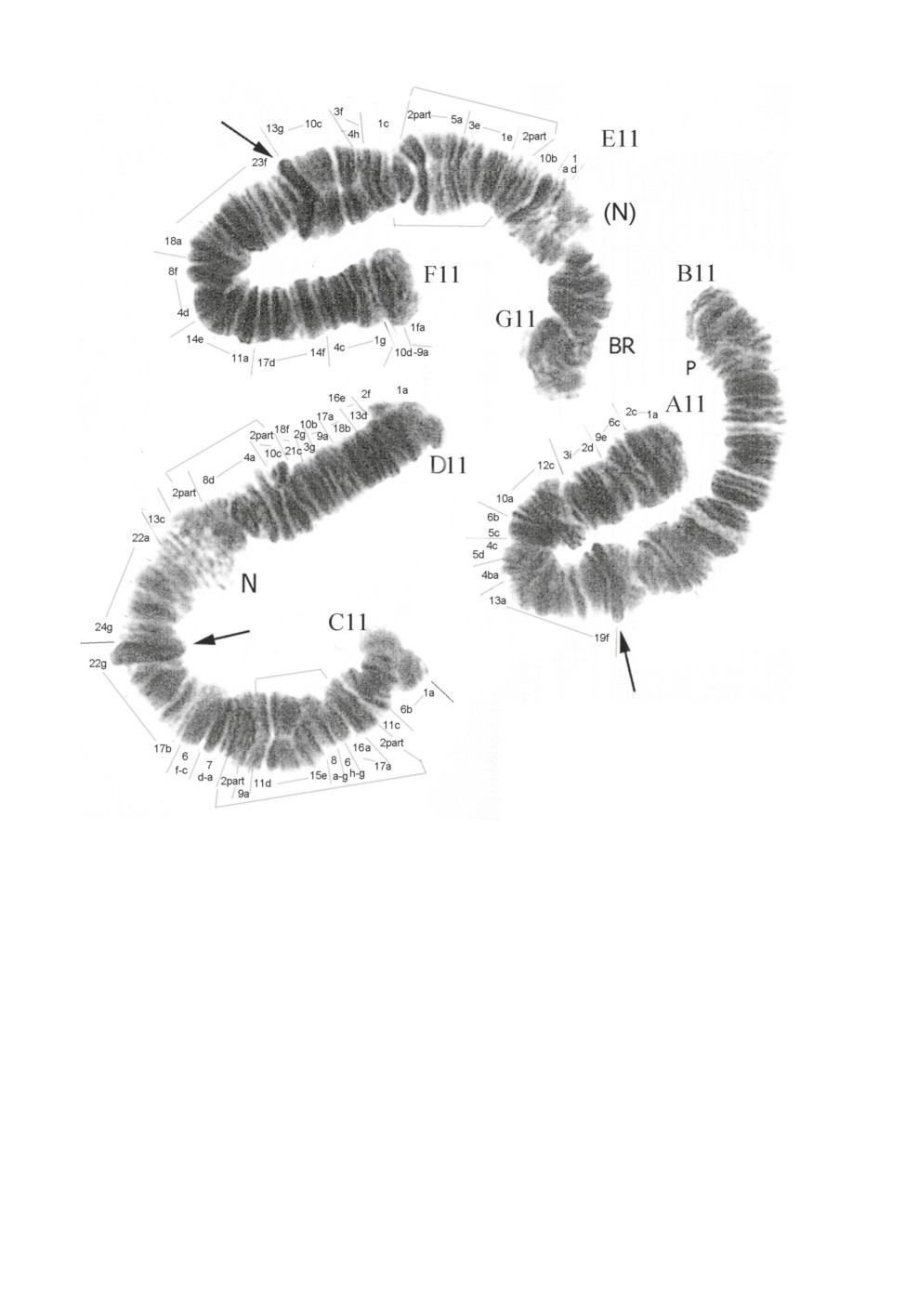
Рис. 4. Chironomus sp., политенные хромосомы.
Арабскими цифрами обозначены районы хромосом, остальные обозначения как на рис. 3.
ча Е, на месте этого объединения работает ядрышко. Второе ядрышко локализовано
в плече D. В кариотипе 2 кольца Бальбиани.
Последовательность дисков плеча A у Chironomus sp. и Ch. tuvanicus идентична.
Ch. sp. A1.1
1a-2c 6c 9e 2d 3i 12c-10a 6d 5e 4c-5d 4ba 13a-19f
Поскольку картирование плеча B у Chironomus sp. представляет трудности, авторы
отказались от него.
766
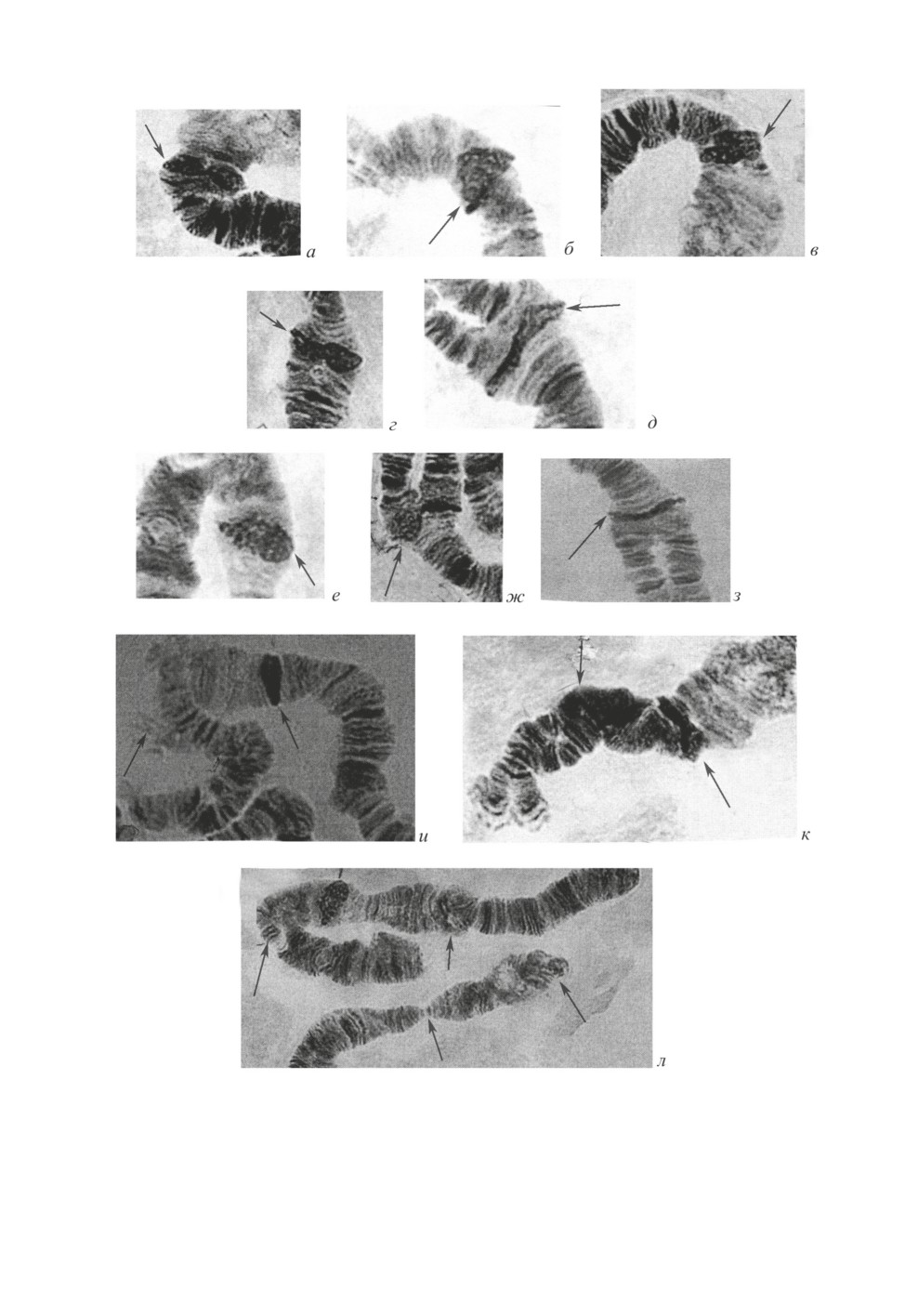
Рис. 5. Центромерные диски политенных хромосом и гетерозиготные инверсии (показаны
стрелками) в популяции Сhironomus sp.
а-в - центромерные диски в хромосоме АВ; г, д - центромерные диски в хромосоме СD;
е-з - центромерные диски в хромосоме GEF; и - гетерозиготная инверсия в плече D;
к - гетерозиготная инверсия в плече С; л - гетерозиготные инверсии в плечах E и G.
767
Последовательность дисков плеча С у Сhironomus sp. отличается от последователь-
ности дисков плеча С у Сh. tuvanicus большой гомозиготной инверсией (эта инверсия
и все последующие подчеркнуты прямой линией).
Ch. sp. C1.1
1a-6b 11c 2part 17a-16a 6hg 8g-a 15с-11d 9a 2part 7d-a 6f-c 17b-22g
tuv C1.1
1a-6b 11c-9a 11d-15e 8a-g 6gh 17a-16a 7d-a 6f-c 17b-22g
Последовательность дисков плеча D у Chironomus sp. очень трудно было картировать,
но мы все же решили сравнить ее с последовательностью в плече D у Ch. tuvanicus.
У Chironomus sp. в плече D есть гомозиготная инверсия
Сh. sp. D1.1
1a-2f 13d-16c 18b-17a 9a-10b 3g-2g 21f-18c 2part-10c 4a-8d 2part-
13c 22a-24g
tuv D1.1
1a-2f 13d-16e 18b-17a 9a-10b 3g-2g 21f-18c 8d-4a 10c-13c 22a-24g
Последовательность дисков плеча Е у Chironomus sp. отличается гомозиготной
инверсией от последовательности дисков плеча Е Ch. tuvanicus. Второе ядрышко
у Chironomus sp. не всегда активно. Диски, кольца Бальбиани и перетяжка не карти-
рованы, они представляют собой малую хромосому G, которая является частью хро-
мосомы FEG. С ядерной оболочкой хромосома соединена очень слабо; освобождаясь
от нее, она сразу попадает в протоплазму (Kiknadze et al., 1992, 1994, 2016).
Ch. sp. E1.1 Диски-BR-перетяжка-1a-d 10b 2part 1e-3e 5a-2part 1с 4h-3f 10c-13g
tuv E 1.1
Диски-BR-перетяжка-N 1a-d 10b-5a 3c-1e 4h-3f 10c-13g
Последовательность дисков плеч F хромосомы FEG Chironomus sp. идентична после-
довательности в плече F Ch. tuvanicus.
Ch. sp. F1.1
1a-f 9a-10d 1g-4c 14f-17d 11a-14e 4d-8f 18f-23f
Кариотип Chironomus sp. по дисковым последовательностям сходен с кариотипом
Ch. tuvanicus, но отличается фиксированными гомозиготными перестройками.
Хромосомный полиморфизм
На 28 особей только 3 (около 11 %) не имели инверсий, остальные 89 % были гете-
розиготны (см. таблицу). Среднее число гетерозиготных инверсий на одну особь со-
ставило 1.2. Больше всего инверсий отмечено в плечах C, D и E. В плече D инверсия
возникла сразу за ядрышком (Inv. 2part-10c); инверсия в С самая большая, она захваты-
вает почти все плечо (Inv. 16a-2part), инверсия в плече Е расположена у его середины
(Inv. 10a-2part).
У Ch. tuvanicus инверсии найдены только в 3 плечах: B, C и F.
Хромосомный полиморфизм личинок Chironomus sp.
Плечо хромосом
Доля личинок с гетерозиготными инверсиями (%)
C1.1
14.2
D1.1
35.2
E1.1
21.5
768
ОБСУЖДЕНИЕ
Сбор материала на архипелаге представляет собой большие трудности из-за отсут-
ствия постоянного гражданского сообщения с Новой Землей, а экспедиции туда крат-
ковременны и не предполагают длительных стационарных работ. Выведение имаго ко-
маров из личинки очень затруднительно из-за краткости теплого периода, и установить
связь личинки с имаго конкретного вида порой невозможно.
Морфологические признаки личинок Chironomus sp. и Ch. tuvanicus очень сходны:
оба относятся к одной группе видов (riihimakiensis) и имеют ее характерные признаки
(Kiknadze et al., 1992, 1994, 2016): зубцы ментума 4-й пары почти вдвое ниже соседних,
виды имеют 2 ядрышка и очень сходны по последовательности дисков хромосом.
Центромерные диски хромосом Chironomus sp. крупные, рыхлые, имеют разные
очертания у разных особей. У Ch. tuvanicus центромеры как у Ch. plumosus, в виде
небольших дисков. У Chironomus sp., как и у Ch. tuvanicus, 2 ядрышка, но второе ра-
ботает не всегда. У Chironomus sp. в бывшем плече G функционирует только 1 кольцо
Бальбиани, а у Ch. tuvanicus - 2, одно работает постоянно, а второе - периодически
(Kiknadze et al., 1992, 1994).
Плечи А и F полностью сохранили одинаковую последовательность дисков у обо-
их видов. Хромосомы C, D и Е Сhironomus sp., наоборот, отличаются от хромосом
Ch. tuvanicus гомозиготными инверсиями, притом что плечо С приобрело достаточно
крупную перестройку.
Таким образом, при большом морфологическом и кариологическом сходстве с одной
стороны, и значительных различиях между этими двумя формами с другой, решить
вопрос о видовой самостоятельности Chironomus sp. пока трудно и нужно подождать
следующей экспедиции на архипелаг Новая Земля для сбора материала по другим ста-
диям этой формы.
В тех же озерах был собран еще один представитель рода - Ch. albimaculatus Shobanov,
Wülker et Kiknadze, 2002. Морфология новоземельских личинок этого вида (n = 33)
и их кариотип идентичны таковым популяции хирономусов из Усть-Ленского заповед-
ника (Shobanov et al., 2002). Принципиальное отличие заключатся в том, что якутская
популяция характеризуется свободнолежащей IV хромосомой, тогда как в популяции
с Новой Земли эта хромосома облигатно участвует в образовании хромоцентра. Раз-
личия такого рода обычно относят к межпопуляционным (Жиров и др., 2016). Таким
образом, на Новой Земле обитают 2 вида, относящиеся к группе riihimakiensis и оба
имеющие 2n = 6.
БЛАГОДАРНОСТИ
Авторы приносят глубокую благодарность Н. С. Хабазовой (ЗИН) за помощь
в оформлении рукописи.
ФИНАНСИРОВАНИЕ
Экспедиция на Новую Землю поддержана Научно-экспедиционным центром
Ассоциации «Морское наследие». Мы выражаем признательность Марии Влади-
славовне Гаврило за организацию экспедиции. Работа выполнена в рамках гостемы
№ АААА-А19-119020790106-0, а также при финансовой поддержке Программы
фундаментальных исследований Президиума РАН № 41 «Биоразнообразие природных
систем и биологические ресурсы России».
769
СПИСОК ЛИТЕРАТУРЫ
Белянина С. И., Филинкова Т. Н. 1996. Морфология и кариотипы трех симпатрических видов рода Chironomus
(Diptera, Chironomidae) из Заполярья. Зоологический журнал 75 (5): 701-710.
Боярский П. В., Гусев С. В., Евсеев Н. Ф., Захаров Ю. С., Калякин В. Н., Корякин В. С., Мазуров Ю. Л.,
Серебрянный Л. Р. 1994. Концепция формирования системы особо охраняемых природных и истори-
ко-культурных территорий на Новой Земле. В кн.: П. В. Боярский (ред.). Новая Земля. Труды Морской
арктической комплексной экспедиции. Вып. IV. Том 3. М.: Российский научно-исследовательский ин-
ститут культурного и природного наследия, с. 13-37.
Думик В. П., Золотухин Г. Е., Матушенко А. М. 1998. Новая Земля - самый северный полигон. Взгляд изну-
три. В кн.: П. В. Боярский (ред.). Новая Земля. Природа. История. Археология. Культура. Труды Мор-
ской арктической комплексной экспедиции (МАКЭ). Кн. 2, ч. 1. Культурное наследие. Радиоэкология.
М.: Российский научно-исследовательский институт культурного и природного наследия, с. 210-220.
Жиров С. В., Петрова Н. А., Крашенинников А. Б. 2016. Chironomus albimaculatus и Chironomus tuvanicus
(Diptera, Chironomidae) с архипелага Новая Земля. Бюллетень медицинских интернет-конференций 6
(9): 1490.
Зеленцов Н. И. 2007 Фауна хирономид (Diptera, Chironomidae) архипелагов Новая Земля и Северная Земля.
Биология внутренних вод 4: 15-19.
Зеленцов Н. И., Шилова А. И. 1996. Фауна хирономид (Diptera, Chironomidae) Усть-Ленского государствен-
ного заповедника. Биология внутренних вод 1: 54-61.
Крашенинников А. Б. 2013. Предварительные данные по фауне и распространению комаров-звонцов
(Diptera, Chironomidae) островов российского сектора Арктики. Вестник Пермского университета,
сер. Биология 1: 32-36.
Макарченко Е. А., Макарченко М. А., Вехов Н. В. 1998. Предварительные данные по фауне хирономид
(Diptera, Chironomidae) архипелага Новая Земля. В кн.: П. В. Боярский (ред.). Новая Земля. Природа.
История. Археология, Культура. Кн. 1. Природа. Труды Морской арктической комплексной экспеди-
ции (МАКЭ). М.: Российский научно-исследовательский институт культурного и природного насле-
дия, с. 262-267.
Кикнадзе И. И., Истомина А. Г., Гундерина Л. И., Салова Т. А., Айманова К. Г., Саввинов Д. Д. 1996. Карио-
фонды хирономид криолитозоны Якутии: триба Chironomini. Новосибирск: Наука, 166 с.
Панкратова В. Я. 1983. Личинки и куколки комаров подсемейства Chironominae фауны СССР (Diptera,
Chironomidae = Tendipedidae). Л.: Наука, 309 с. (Определители по фауне СССР, издаваемые Зоологи-
ческим институтом АН СССР, вып. 134).
Чубарева Л. А., Петрова Н. А. 1982. Цитогенетический метод анализа хромосом у хирономид. В кн.: Мето-
дическое пособие по изучению хирономид. Душанбе: Дониш, с. 64-73.
Шобанов Н. А., Петрова Н. А. 1995. Особенности кариотипа Chironomus saxatilis Wülker, Reser, Scholl
1981 из Сибирского Заполярья и возможное возникновение неоцентромер в хромосоме АЕ (цитоком-
плекс pseudothummi). Цитология 37 (7): 586-592.
Шобанов Н. А., Шилова А. И., Белянина С. И. 1996. Объем и структура рода Chironomus (Diptera,
Chironomidae): обзор мировой фауны. В кн.: Н. А. Шобанов, Т. Д. Зинченко (ред.). Экология, эволю-
ция и систематика хирономид. Тольятти: Институт экологии Волжского бассейна РАН, с. 44-96.
Cranston P. S., Reiss F. 1983. The larvae of Chironomidae (Diptera) of the Holarctic Region. Key to subfamilies
II. Entomologica Scandinavica. Supplement 19: 11-15.
Kiknadze I. I., Siirin M. T., Wülker W. 1992. Siberian species of the riihimakiensis-group in the genus Chironomus
(Diptera, Chironomidae). I. Karyotypes and morphology. Netherlands Journal of Aquatic Ecology 26 (2-4):
163-171.
Kiknadze I. I., Siirin M. T., Wülker W. 1994. Siberian species of the riihimakiensis group in the genus Chironomus.
II. Inversion polymorphism and cytophylogeny. Spixiana. Supplement 20: 115-113.
Kiknadze I. I., Istomina A., Golygina V., Gunderina L. 2016. Karyotypes of Palearctic and Holarctic Species of the
Genus Chironomus. Novosibirsk: Academic Publishing House «GEO», 489 p.
Petrova N. A., Zhirov S. V. 2008. Polytene chromosomes of salivary glands of chironomids (Diptera: Chironomidae)
from the Wrangel Island (Russia). Comparative Cytogenetics 2: 127-130.
Pinder L. C. V., Reiss F. 1983. The larvae of Chironominae (Diptera, Chironomidae) of the Holarctic Region - Keys
and diagnoses. Entomologica Scandinavica. Supplement 19: 293-436.
Saether O. A. 1980. Glossary of chironomid morphology terminology (Diptera: Chironomidae). Entomologica
Scandinavica. Supplement 14: 1-51.
Shobanov N. A., Wülker W. F., Kiknadze I. I. 2002. Chironomus albimaculatus sp. n. and C. trabicola sp. n. (Diptera,
Chironomidae) from Polar Russia. Aquatic Insects 24 (3): 169-188.
Wülker W. 1991. Chironomus fraternus n. sp. and Ch. beljaninae n. sp., sympatric sibling species in Fennoscandian
reservoirs. Entomologica Fennica 2: 97-109.
770
LARVAL MORPHOLOGY AND KARYOTYPE STRUCTURE
OF CHIRONOMUS SP. (DIPTERA, CHIRONOMIDAE)
FROM NOVAYA ZEMLYA ARCHIPELAGO (RUSSIA)
N. A. Petrova,
S. V. Zhirov, A. B. Krasheninnikov
Key words: Chironomus sp., larvae morphology, karyotype, 2n = 6, riihimakiensis group,
Polar area.
SUMMARY
Larval morphology and karyotype characters of Chironomus sp. from Yuzhnyi Island of the Novaya
Zemlya Archipelago (70º32ʹ N, 55º10ʹ E) are described. This species is close to Ch. tuvanicus described
from Tuva (Siberia, Russia) and belongs to the riihimakiensis group. Its chromosome set is 2n = 6:
AB, CD, FEG, the latter chromosome has derived from joining of the small chromosome G with the
arm E and genetic fixation in the karyotype. Chironomus sp. has the Arctic distribution, belongs to the
riihimakiensis group and is a primitive member of the genus.
771