ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 98, 4, 2019
УДК 595.722
НОВЫЙ ВИД ТАРАКАНОВ РОДА MACROPHYLLODROMIA
SAUSSURE ET ZEHNTNER, 1893 (DICTYOPTERA, ECTOBIIDAE),
С ЗАМЕЧАНИЯМИ О ТАКСОНОМИЧЕСКОМ
ПОЛОЖЕНИИ РОДА
© 2019 г. Л. Н. Анисюткин
Зоологический институт РАН
Университетская наб., 1, С.-Петербург, 199034 Россия
e-mail: leonid.dictyoptera@gmail.com, Leonid.Anisyutkin@zin.ru
Поступила в редакцию 10.12.2018 г.
После доработки 10.1.2019 г.
Принята к публикации 10.1.2019 г.
Описан новый вид рода Macrophyllodromia, M. obscura sp. n. из Перу. Предлагается новый диа-
гноз рода, основанный на признаках строения гипандрия и гениталий самцов. Кратко рассматри-
вается таксономическая структура сем. Ectobiidae и положение в нем рода Macrophyllodromia.
Ключевые слова: тараканы, морфология, новый вид, Plectopterinae.
DOI: 10.1134/S0367144519040130
Род Macrophyllodromia Saussure et Zehntner, 1893 включает 13 видов (вместе с опи-
сываемым ниже), распространенных в Центральной и Южной Америке (Vidlička,
2013; Beccaloni, 2014). 8 из 13 видов были описаны после 2007 г., поэтому не вызывает
сомнений, что количество известных в нем видов будет расти и далее. Морфология
представителей рода хорошо изучена по сравнению со многими родами тропических
эктобиид, тем не менее даже принадлежность рода к какому-либо подсемейству сем.
Ectobiidae Brunner von Wattenwyl, 1865 не бесспорна, что объясняется запутанной над-
родовой системой эктобиид.
В данной статье описывается новый вид рода Macrophyllodromia и делается попытка
проанализировать таксономическое положение рода в сем. Ectobiidae.
МАТЕРИАЛ И МЕТОДИКА
Единственный известный экземпляр был собран в 70%-ный этанол и хранится в нем в насто-
ящее время. Автор следует ранее описанной методике работы с материалом (Анисюткин, 2012).
Номенклатура жилкования надкрылий и крыльев принята по работе Рена (Rehn, 1951), во-
оружение передненижнего края передних бедер описывается по схеме Бей-Биенко (1950) и Рота
(Roth, 2003). Номенклатура склеритов гениталий самцов приводится по Классу (Klass, 1997),
в скобках дается номенклатура по Гранколасу (Grandcolas, 1996).
Голотип хранится в коллекции Зоологического института РАН в Санкт-Петербурге (ЗИН).
804
РЕЗУЛЬТАТЫ
Сем. ECTOBIIDAE Brunner von Wattenwyl, 1865
Подсем. PLECTOPTERINAE Saussure et Zehntner, 1893
Род MACROPHYLLODROMIA Saussure et Zehntner, 1893
Тип ово й вид: Pseudophyllodromia maximiliani Saussure, 1873, по первоначальной моно-
типии.
Ди а гно з. В качестве диагностических для рода в настоящее время указывается
комплекс признаков внешнего строения, из которых можно выделить следующие: от-
носительно крупные для сем. Ectobiidae размеры; наличие продольной темной полосы
на лицевой части головы и пары продольных темных полос на пронотуме; расстояние
между глазами меньше, чем между основаниями антенн; скошенное медио-кубиталь-
ное поле надкрылья (область ветвей M и CuA), обычно именуемое «discoidal sector»;
отсутствие тергальных желез у самцов; асимметричный гипандрий (Rocha e Silva
Albuquerque, 1962; Roth, Naskrecki, 2001; Lopes, Oliveira, 2006; Anisyutkin, 2007, 2012;
Vidlička, 2013). Ни один из перечисленных признаков не является уникальным и не
может рассматриваться в качестве аутапоморфии рода.
По моему мнению, род Macrophyllodromia характеризуется своеобразным строе-
нием гипандрия, на признаках строения которого, равно как и на признаках генита-
лий самцов, может быть основан диагноз рода. Среди этих признаков следует искать
и аутапоморфии рода.
В этой работе я предлагаю предварительный диагноз рода, основанный на признаках
строения гипандрия и гениталий самцов. Гипандрий широкий, с хорошо выраженны-
ми заднебоковыми углами, часто оттянутыми в склеротизованные выросты; по задне-
му краю с 2-4 склеротизованными выростами сложной формы; стилусы отсутствуют
или маленькие, мембранозные [возможно, что некоторые выросты гипандрия явля-
ются преобразованными стилусами (Anisyutkin, 2007)]. Правая фалломера крупная и
сильно склеротизованная, склерит R2 сложного строения, склерит R3 с крупным рез-
ко изогнутым выростом («anterior protrusion», по: Anisyutkin, 2011). Вершинная часть
склерита L2D обычно вздута и сильно склеротизована.
Окончательно диагноз рода может быть сформулирован после детального изучения
структур генитального комплекса большинства видов, включая типовой, а также близ-
ких родов.
Важность признаков строения правой фалломеры для диагностики рода
Macrophyllodromia ранее уже отмечалась Ротом и Наскрецким (Roth, Naskrecki, 2001).
Эта структура может быть особенно важной для реконструкции филогении тараканов
на надродовом уровне (Anisyutkin, Yushkova, 2017).
С о с т а в. 12 видов согласно Беккалони (Beccaloni, 2014) и M. obscura sp. n.
Macrophyllodromia obscura Anisyutkin, sp. n. (рис. 1-16).
М ат ер иа л. Перу, регион Укаяли (Ucayali), провинция Аталайя (Atalaya), ~ 35 км NWW от
г. Аталайя (Atalaya), вблизи дер. Сапани (Sapani), h ~ 300 м, в лесу, 26-31.X.2008 (А. Горохов,
Л. Анисюткин, М. Березин, Е. Ткачева, В. Изерский), 1 ♂ (голотип).
805
Са ме ц (голотип). Общая окраска темно-коричневая. Голова и ее придатки коричневые; глаза
черные; глазки и дистальный участок лабрума белесые; проксимальные 7 или 8 члеников антенн
светло-коричневые, далее членики более темные, почти черные, а в дистальной части, примерно
после 35-го членика, снова более светлые - коричневые. Центральная часть пронотума черная,
боковые части желтые. Надкрылья, грудь, ноги и брюшко сверху коричневые; голени и лапки
немного более светлые; брюшко снизу почти черное.
Покровы гладкие и блестящие, не скульптурированы и не пунктированы; опушение выражено
только на антеннах, дистальнее примерно 20-го членика.
Голова приблизительно треугольной формы, поперечная (рис. 1, 2); глазки маленькие; мини-
мальное расстояние между глазами составляет примерно 1.7 длины глаза; расстояние между
антеннальными впадинами примерно 1.5 длины скапуса (~ 0.6 мм); соотношение длин 3 апи-
кальных члеников максиллярных щупиков (от 3-го к 5-му) примерно 1.2 : 1.0 : 1.3, апикальный
членик увеличен.
Пронотум поперечно-овальный (рис. 2). Надкрылья и крылья полностью развиты, общая кар-
тина жилкования надкрылий и крыльев сходна с таковой представителей рода Euphyllodromia
Shelford, 1908 (см.: Anisyutkin, 2011). Надкрылья слабо склеротизованные, с хорошо выражен-
ными жилками. Sc простая, длиной примерно в 1/3 надкрылья, утолщена в основании (хорошо
заметно с нижней стороны крыла); R проходит примерно посередине надкрылья, достигая его
вершины, с многочисленными, иногда ветвящимися передними ветвями; M и CuA не разделены;
CuP резко изогнута перед впадением в задний край надкрылья.
Передненижние края передних бедер вооружены по типу A, с 11 или 12 шипами, включая
2 вершинных. Передние голени не утолщены. Вооружение голеней и бедер хорошо развито.
В задних лапках метатарзус немного длиннее остальных члеников вместе взятых, вдоль ниж-
него края с 2 примерно одинаковыми рядами шипиков; все эуплантулы маленькие, вершинные,
с 0-2 дополнительными шипиками с внешней и внутренней сторон. 2-й и 3-й членики с корот-
кими рядами шипиков по нижнему краю; аролиум по длине примерно равен половине коготка;
коготки симметричные, простые. Передние и средние лапки сходны с задними, но их метатарзус
короче; в средних лапках 3-й, а в передних 3-й и 4-й членики без рядов шипиков по нижнему
краю.
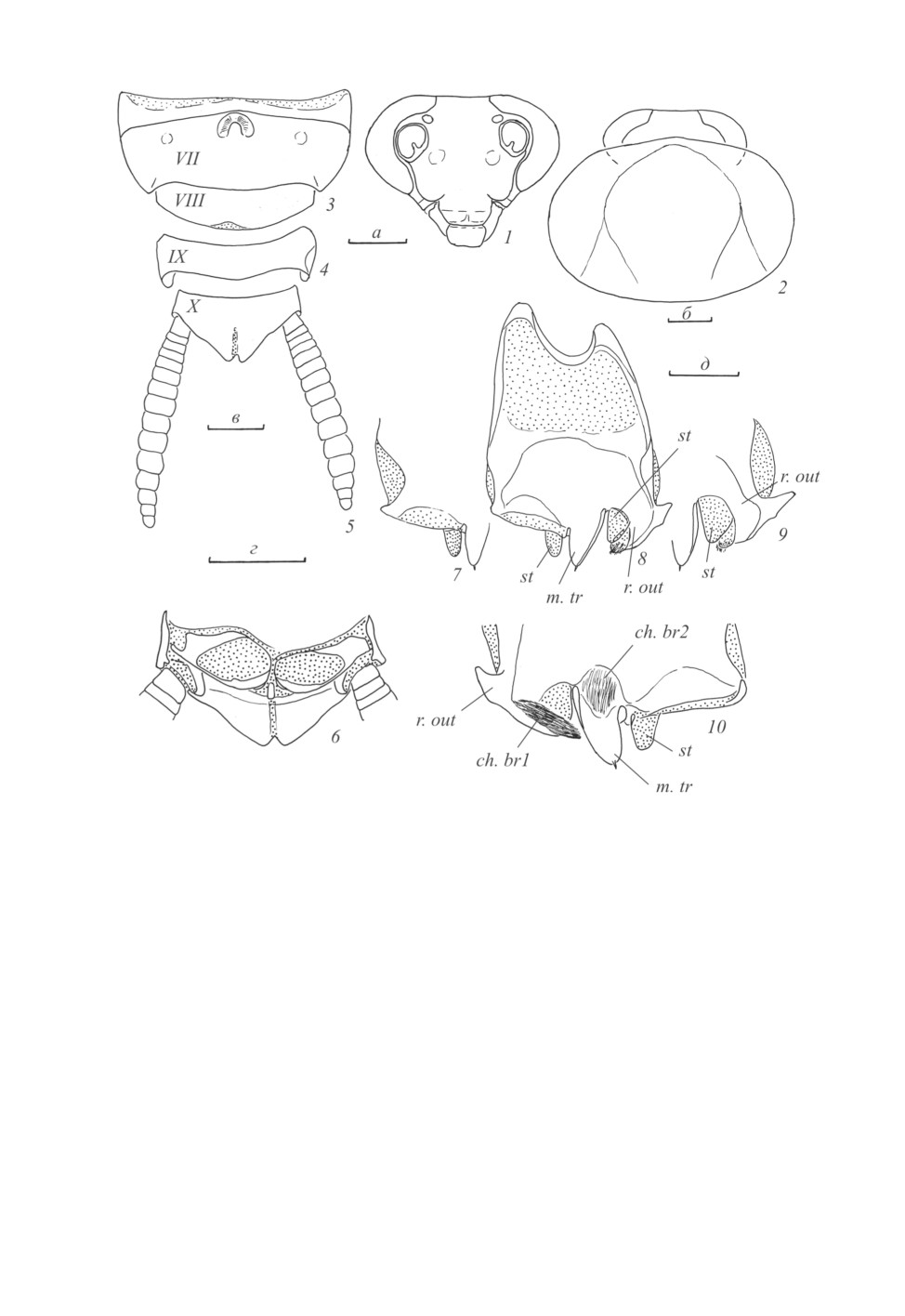
VII тергит брюшка с медиальным вдавлением у переднего края, во вдавлении располагается
округленный вырост, обрамленный щетинками; латерально располагается пара нечетких вдавле-
ний (рис. 3). VI и VII тергиты брюшка с заостренными и оттянутыми назад заднебоковыми
углами (рис. 3). IX тергит в виде поперечной пластинки с округленным задним краем (рис. 4).
Анальная пластинка треугольная, каудально выступающая, с небольшим вырезом по заднему
краю и продольной мембранозной полосой. Церки длинные, отчетливо сегментированные, по-
следние членики апикально закруглены (рис. 5). Парапрокты симметричные, без вооружения, в
срединной части мембранозные; склериты Pv крупные, склеротизованные (рис. 6). Гипандрий
асимметричный, приблизительно прямоугольной формы; правый переднебоковой вырост боль-
ше левого (рис. 8); левый заднебоковой угол каудально оттянут в апикально раздвоенный вырост
(рис. 8-10, r. out), сверху на нем располагается пучок щетинок (рис. 10, ch. br1); медиальнее
выроста располагается стилус (рис. 9, st); посередине заднего края гипандрия располагается
срединный треугольник с единственным шипиком на вершине (рис. 8, 10, m. tr), в основании
срединного треугольника на его верхней стороне располагается еще один, менее склеротизо-
ванный пучок щетинок (рис. 10, ch. br2); правее срединного треугольника располагается второй
стилус (рис. 7, 10, st); правый заднебоковой угол гипандрия почти прямоугольный, с небольшим
выростом (рис. 7); стилусы мембранозные, не имеют выраженного сочленения с гипандрием.
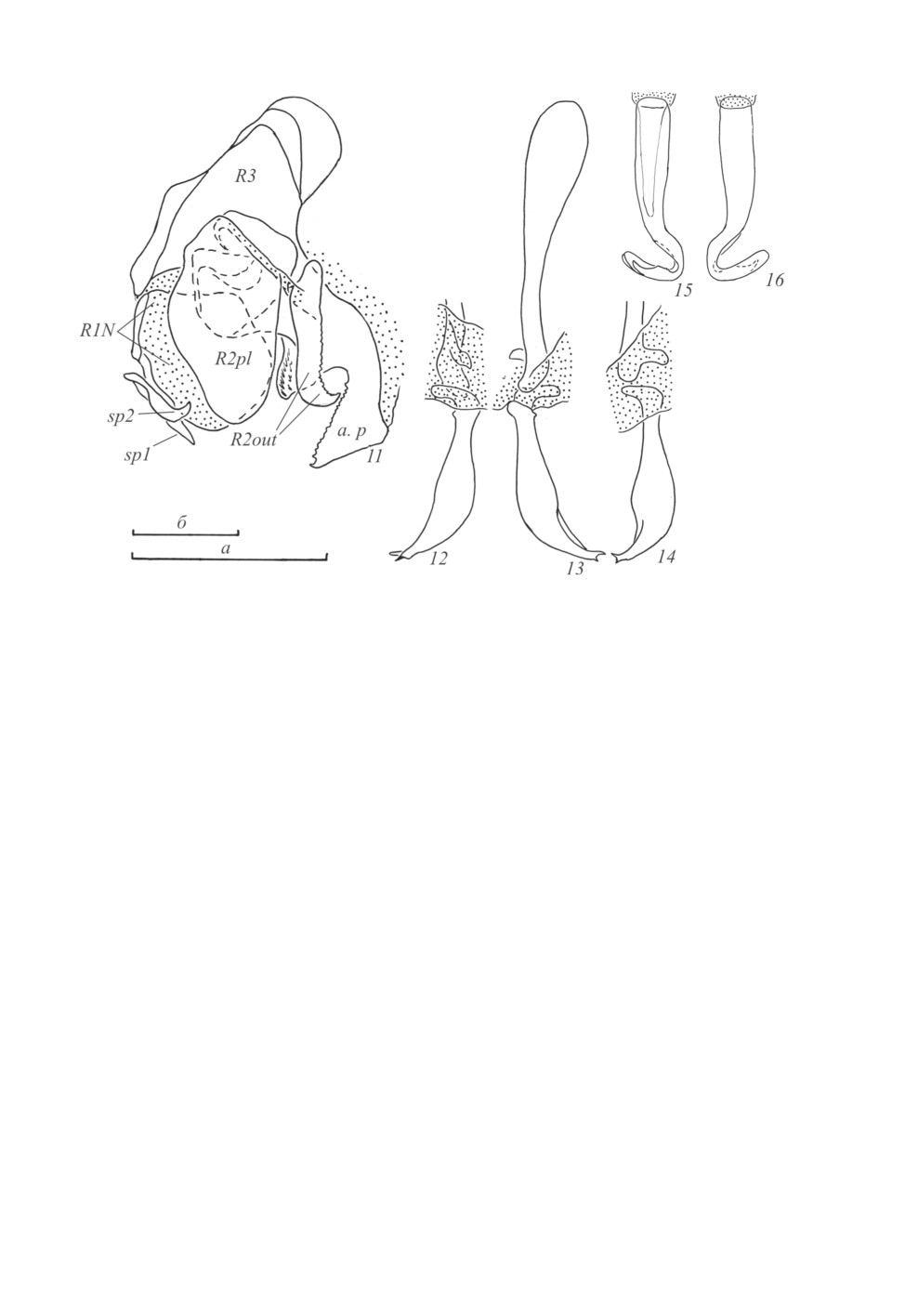
Гениталии самца (рис. 11-16). Правая фалломера (комплекс склеритов R + N - здесь и да-
лее в скобках дается номенклатура по: Grandcolas, 1996) (рис. 11): склерит R1N широкий, рас-
полагается преимущественно на нижней части фалломеры, каудолатерально несет уплощен-
ный вырост с парой шипов (рис. 11, sp1, sp2); склерит R2 сложной формы, сверху с широким
806
Рис. 1-10. Macrophyllodromia obscura sp. n., самец, голотип.
1 - лицевая часть головы; 2 - голова и пронотум сверху; 3, 4 - VII, VIII (3) и IX (4) тергиты брюшка сверху;
5 - анальная пластинка и церки сверху; 6 - вершина брюшка снизу, гипандрий и гениталии удалены;
7, 9 - левый (7) и правый (9) заднебоковые углы гипандрия, вид снизу и сбоку; 8 - гипандрий снизу;
10 - задний край гипандрия сверху. Пунктировкой показаны мембранозные участки.
Обозначения морфологических структур см. в тексте.
Масштабная линейка - 1 мм: а - к рис. 1; б - к рис. 2; в - к рис. 3-5; г - к рис. 6, 10; д - к рис. 7-9.
пластинчатым выростом (рис. 11, R2pl), медиально - с изогнутым зазубренным выростом
(рис. 11, R2out), противолежащим выросту склерита R3; склерит R3 большой и хорошо скле-
ротизованный, с крупным, резко изогнутым выростом («anterior protrusion», по: Anisyutkin,
2011) (рис. 11, a. p). Склерит L2D (L1) разделен на вершинную и базальную части (рис. 12-14);
базальная часть краниально расширена, сильно склеротизована, каудально, у вершины, несет
дорсально направленный вырост; вершинная часть массивная, краниально с боковым выростом,
апикально с 2 зубчиками (рис. 12-14). Склерит L3 (L2d) (рис. 15, 16) удлиненный, апикально
с бороздой hge («subapical incision», по: Roth, 1970a). Какие-либо другие структуры из лабильно-
го комплекса («labile complex», по: Anisyutkin, 2011) не выражены.
807
Рис. 11-16. Macrophyllodromia obscura sp. n., самец, голотип.
11 - правая фалломера сверху; 12, 14 - вершина склерита L2D сверху (12) и сбоку (14); 13 - склерит L2D
снизу; 15, 16 - склерит L3. Пунктировкой показаны мембранозные участки.
Обозначения морфологических структур см. в тексте.
Масштабная линейка - 1 мм: а - к рис. 11; б - к рис. 12-16.
Са мка неизвестна.
Р азме ры (мм). Длина головы - 2.6, ширина - 3.1; длина пронотума - 3.6, ширина - 5.4;
длина надкрылья - 16.1, ширина - 5.1.
С ра в ни те л ьны е за меч а ни я. Macrophyllodromia obscura sp. n. хорошо отли-
чается от большинства видов рода Macrophyllodromia темными, без рисунка головой
и центральной частью пронотума, которые у большинства представителей рода светло
окрашены с продольными темными полосами - 1 на лицевой части головы и 2 на про-
нотуме (Rocha e Silva Albuquerque, 1962; Roth, Naskrecki, 2001; Lopes, Oliveira, 2006;
Anisyutkin, 2007, 2012; Vidlička, 2013). Новый вид сходен с M. nigrigena Hebard, 1927 и
M. nobile Anisyutkin, 2007 темноокрашенной головой, но для M. nigrigena и M. nobile
характерно присутствие пары продольных полос на пронотуме (Hebard, 1927, pl. XII,
fig. 18; Anisyutkin, 2007, fig. 10), они хорошо отличаются от нового вида также строе-
нием заднего края гипандрия, а именно отсутствием двух изогнутых шиповидных вы-
ростов (ср. рис. 7-10 и pl. XII, Fig. 19 в: Hebard, 1927 и Fig. 14, 15 в: Anisyutkin, 2007).
Эт имоло гия. Название вида - латинское прилагательное женского рода («тем-
ная»).
З а меча н ия. Macrophyllodromia obscura sp. n. занимает обособленное положение
в роде не только благодаря нетипичной окраске (отсутствие контрастных полос на го-
лове и пронотуме), но и из-за наличия железы на VII тергите брюшка (см. рис. 3), что
808
не соответствует ранее данным диагнозам рода (Rocha e Silva Albuquerque, 1962; Roth,
Naskrecki, 2001; Lopes, Oliveira, 2006; Anisyutkin, 2007, 2012; Vidlička, 2013). Нельзя,
однако, исключать вероятность того, что небольшая железа могла быть просто не об-
наружена при работе с сухим материалом, а на самом деле есть и у ранее описанных
видов.
ОБСУЖДЕНИЕ
В настоящее время род Macrophyllodromia относят к подсем. Pseudophyllodromiinae
Hebard, 1929 (Roth, Naskrecki, 2001; Roth, 2003; Beccaloni, 2014), однако диагноз это-
го подсемейства явно недостаточен (см. далее), из чего следует неопределенность его
состава.
В сем. Ectobiidae выделяют 4 подсемейства: Ectobiinae Brunner von Wattenwyl, 1865,
Pseudophyllodromiinae, Blattellinae Karny, 1908 и Nyctiborinae Brunner von Wattenwyl,
1893 (Beccaloni, 2014). Эктобиины и никтиборины - хорошо очерченные группы,
к которым Macrophyllodromia явно не имеет отношения. Для Ectobiinae характер-
но специфическое строение тергальных желез брюшка и гипандрия (Roth, 2003),
для Nyctiborinae - опушенные покровы и специализированные гениталии самцов
(Anisyutkin, 2009; Vélez-Bravo, 2013; Анисюткин, 2017).
Различия между крупнейшими подсемействами эктобиид, Pseudophyllodromiinae
и Blattellinae, заключаются в инвертированности, т. е. различных типах зеркальной сим-
метрии, левой или правой, гениталий самцов, и в отсутствии ротации оотеки у псевдо-
филлодромиин (Roth, 2003). Отсутствие ротации оотеки, т. е. положение, в котором
ее киль направлен вверх, и неинвертированность гениталий самцов являются плезио-
морфиями (Roth, 1970b, 2003). Таким образом, подсемейства Pseudophyllodromiinae и
Blattellinae в настоящее время диагностируются одним признаком каждое: инвертиро-
ванностью гениталий и ротацией оотеки соответственно. Сем. Blaberidae, к которому
принадлежат наиболее продвинутые тараканы (Roth, 1970b, 2003), имеет инвертиро-
ванные гениталии, сходно с псевдофиллодромиинами, и ротированную оотеку, сходно
с блаттеллинами, а по строению правой фалломеры семейство сходно с никтиборинами
(Анисюткин, 2017). Очевидно, что мы имеем дело с гомоплазиями, а эволюция экто-
биид и блаберид в значительной степени была параллельной.
Насколько стабилен признак инвертированности гениталий самцов? Показана его
вариабельность у некоторых родов подсем. Ectobiinae (Bohn, 1987). У богомолов
случаи реверсии более многочисленны (Balderson, 1978; Anisyutkin, Gorochov, 2005;
Анисюткин, 2005; Holwell et al., 2015; Lombardo et al., 2016). Возможно, что у экто-
биид реверсии более многочисленны, но остаются нераспознанными, так как близкие
таксоны просто разносят по разным подсемействам в соответствии с типом симметрии
гениталий.
Название Pseudophyllodromiinae (исходно Pseudophyllodromiae - Hebard,
1929)
было введено Виккери и Кеваном (Vickery, Kevan, 1983) как приоритетное перед
Plectopterinae Saussure et Zehntner, 1893, которому они приписали авторство МакКит-
трик (McKittrick, 1964).
Представители родов Plectoptera Saussure, 1864 и Pseudophyllodromia Brunner von
Wattenwyl, 1865 очень непохожи по признакам внешнего строения. Для Plectoptera
характерны скорее жукообразный габитус, укороченные и уплотненные надкрылья и
809
крылья с подворачивающимся сочленовным полем, гипандрий со срединным треуголь-
ником и короткими массивными стилусами (Saussure et al., 1893; Rehn, Hebard, 1927;
Pruna, 1974; Rocha e Silva, de Aguiar, 1977; Anisyutkin, 2009). Род Pseudophyllodromia
характеризуется обычным «тараканьим» габитусом - удлиненные, умеренно склеро-
тизованные надкрылья; крылья без сочленовного поля, гипандрий без срединного тре-
угольника, стилусы немного укороченные, веретеновидные (Roth, 1996).
Гениталии самцов этих родов слабо изучены. Все имеющиеся описания (Pruna, 1974;
Rocha e Silva, de Aguiar, 1977; Roth, 1996; Anisyutkin, 2009) делают возможным опре-
деление видов, но недостаточны для гомологизации структур и выяснения филогене-
тических отношений между таксонами. Тем не менее, заметно сходство в строении
правой фалломеры, а именно присутствие каудально направленного выроста склерита
R3 («anterior protrusion», по: Anisyutkin, 2011) (ср. Fig. 2F, a. p в: Anisyutkin, 2009 и
Figs 16 и 21 в Roth, 1996).
Роды Plectoptera, Macrophyllodromia и Euphyllodromia имеют ряд общих признаков
в строении генитальных комплексов: наличие срединного треугольника на гипандрии,
короткие и массивные стилусы и каудально направленный вырост склерита R3 пра-
вой фалломеры. Резкие габитуальные различия между «жукообразной» Plectoptera и
имеющими обычный «тараканий» облик Macrophyllodromia и Euphyllodromia могут
объясняться принадлежностью к разным жизненным формам, поэтому до получения
более полной информации о Pseudophyllodromia и Plectoptera я предпочитаю относить
Macrophyllodromia именно к Plectopterinae.
ФИНАНСИРОВАНИЕ
Работа выполнена в рамках гостемы AAAA-A19-119020690101-6 и при финансовой
поддержке Российского фонда фундаментальных исследований (грант № 16-04-01143
A).
СПИСОК ЛИТЕРАТУРЫ
Анисюткин Л. Н. 2005. Новые данные по роду Haania Sauss. (Dictyoptera, Mantidae, Thespinae) Индокитая
с описанием нового вида из Вьетнама. Труды Русского энтомологического общества 76: 15-20.
Анисюткин Л. Н. 2012. Описание нового вида тараканов рода Prosoplecta Saussure, 1864 (Dictyoptera,
Ectobiidae) из Южного Вьетнама. Энтомологическое обозрение 91 (4): 742-756. (Anisyutkin L. N. 2013.
A description of a new species of the cockroach genus Prosoplecta Saussure, 1864 (Dictyoptera, Ectobiidae)
Анисюткин Л. Н. 2017. Тараканы подсемейства Epilamprinae (Dictyoptera, Blaberidae): таксономическое по-
ложение и состав. В кн.: Материалы Юбилейной отчетной научной сессии, посвященной 185-летию
Зоологического института РАН. СПб.: Зоологический институт РАН, с. 25-27.
Бей-Биенко Г. Я. 1950. Насекомые таракановые. Фауна СССР, новая серия № 40. М.; Л.: Издательство
Академии наук СССР, 343 с.
Anisyutkin L. N. 2007. New species of the genus Macrophyllodromia Saussure & Zehntner, 1893 (Dictyoptera:
Blattina: Blattellidae) from Ecuador. Cockroaches Studies 2: 27-41.
Anisyutkin L. N. 2009. A new species of the genus Plectoptera Saussure and redescription of Eushelfordia pica
Walker (Dictyoptera: Blattellidae) from Peru. Proceedings of the Zoological Institute RAS 313 (4): 440-446.
Anisyutkin L. N. 2011. A review of the genus Euphyllodromia Shelford, 1908 (Dictyoptera: Ectobiidae), with
description of three new species. Proceedings of the Zoological Institute RAS 315 (4): 369-398.
Anisyutkin L. N. 2012. Additional data on the genus Macrophyllodromia Saussure & Zehntner, 1893 (Dictyoptera:
Blattina: Ectobiidae). Cockroach Studies 3: 14-21.
Anisyutkin L. N., Gorochov A. V. 2005. Haania doroshenkoi, a new species of mantises from Cambodia (Mantina:
Mantidae: Thespinae) and a case of mirror symmetry in the structure of the male genitalia of mantises.
Russian Entomological Journal 13 (3) (for 2004): 119-122.
810
Anisyutkin L. N., Yushkova O. V. 2017. New data on cockroaches of the subfamily Epilamprinae (Dictyoptera:
Blaberidae) from India and Sri Lanka, with description of new species and the genital complex of Aptera
Balderson J. 1978. Reversal of the phallic complex in the genera Ciulfina Giglio-Tos and Stenomantis Saussure
(Mantodea: Mantidae: Iridopteryginae). Journal of the Australian Entomological Society 17: 235-239.
Beccaloni G. W. 2014. Cockroach Species File Online. Version 5.0/5.0. World Wide Web electronic publication.
Bohn H. 1987. Reversal of the right-left asymmetry in male genitalia of some Ectobiinae (Blattaria: Blattellidae)
and its implications on sclerite homologization and classification. Entomologica Scandinavica 18: 293-303.
Grandcolas P. 1996. The phylogeny of cockroach families: a cladistic appraisal of morpho-anatomical data. Canadian
Journal of Zoology 74: 508-527.
Hebard M. 1927. The Blattidae of French Guiana. Proceedings of the Academy of Natural Sciences of Philadelphia
78 (for 1926): 135-244.
Hebard M. 1929. Studies in Malayan Blattidae (Orthoptera). Proceedings of the Academy of Natural Sciences of
Philadelphia 81: 1-109.
Holwell G. I., Kazakova O., Evans F., O’Hanlon J. C., Barry K. L. 2015. The functional significance of chiral
genitalia: Patterns of asymmetry, functional morphology and mating success in the praying mantis Ciufina
Klass K.-D. 1997. The external male genitalia and the phylogeny of Blattaria and Mantodea. Bonner Zoologische
Monographien 42: 1-341.
Lombardo F., Stiewe M. B. D., Ippolito S. 2016. Chirality of male genitalia in Otomantis casaica Giglio-Tos, 1915
(Mantodea: Hymenopodidae, Acromantinae). Zootaxa 4105 (2): 198-200.
Lopes S. M., Oliveira de E. H. 2006. Duas espécies novas de Macrophyllodromia do Estado do Acre, Brasil
(Blattaria, Blattellidae) coletadas em ninhos de vespas. Iheringia 96 (2): 257-260.
McKittrick F. A. 1964. Evolutionary studies of cockroaches. Cornell University Agricultural Experiments Station
Memoir 389: 1-197.
Pruna P. M. 1974. Las Especies Cubanas del Género Plectoptera Saussure (Blattariae: Anaplectidae). Poeyana 124:
1-18.
Rehn J. W. H. 1951. Classification of the Blattaria as indicated by their wings (Orthoptera). Memoirs of the American
Entomological Society 14: 1-134.
Rehn J. A. G., Hebard M. 1927. The Orthoptera of the West Indies. Number 1. Blattidae. Bulletin of the American
Museum of Natural History 54: 1-320.
Rocha e Silva I., Aguiar de G. M. 1977. Blattariae da Amazônia. Familia Blaberidae da localidade Sinop, Estado
de Mato Grosso, Brasil (Dictyoptera). Boletim do Museu Paraense Emilio Goeldi, Nova Série. Zoologia 88:
1-30.
Rocha e Silva Albuquerque I. 1962. Synopsis of the Neotropical cockroach genus Macrophyllodromia (Orthoptera:
Blattoidea, Epilampridae). Proceedings of the United States National Museum 113 (3461): 421-428.
Roth L. M. 1970a. The male genitalia of Blattaria. II. Poeciloderrhis spp. (Blaberidae: Epilamprinae). Psyche 77:
104-119.
Roth L. M. 1970b. Evolution and taxonomic significance of reproduction in Blattaria. Annual Review of Entomology
15: 75-96.
Roth L. M. 1996. The cockroach genera Sundablatta Hebard, Pseudophyllodromia Brunner, and Allacta Saussure
& Zehntner (Blattaria: Blattellidae, Pseudophyllodromiinae). Tijdschrift voor Entomologie 139: 215-242.
Roth L. M. 2003. Systematics and phylogeny of cockroaches (Dictyoptera: Blattaria). Oriental Insects 37: 1-186.
Roth L. M., Naskrecki P. 2001. Trophobiosis between a blattellid cockroach (Macrophyllodromia spp.) and fulgorids
(Enchophora and Copidocephala spp.) in Costa Rica. Journal of Orthoptera Research 10 (2): 189-194.
Saussure de H., Zehntner L., Pictet A. 1893. Biologia Centrali-Americana. Insecta, Orthoptera. Vol. 1. London:
Vélez-Bravo A. 2013. One new genus of cockroach in the Neotropical subfamily Nyctiborinae (Dictyoptera:
Blattodea: Ectobiidae). Zootaxa 3681 (1): 79-84.
Vickery V. R., Kevan D. K. McE. 1983. A monograph of the orthopteroid insects of Canada and adjacent regions.
Lyman Entomological Museum and Research Laboratory. Memoir 13 (1): 1-679.
Vidlička L. 2013. New species of Macrophyllodromia (Blattaria, Blattellidae) from Ecuador and a key to males of
the genus. Zootaxa 3635 (2): 185-193.
811
A NEW SPECIES OF THE COCKROACH GENUS MACROPHYLLODROMIA
SAUSSURE ET ZEHNTNER, 1893 (DICTYOPTERA, ECTOBIIDAE),
WITH NOTES ON THE TAXONOMIC POSITION OF THE GENUS
L. N. Anisyutkin
Key words: cockroaches, morphology, new species, Plectopterinae.
SUMMARY
A new species of the genus Macrophyllodromia, M. obscura sp. n., is described from Peru.
An emended diagnosis of the genus Macrophyllodromia is given based on the characters of hypandrium
and male genitalia. The taxonomy of the family Ectobiidae and the place of Macrophyllodromia in it
are briefly analyzed.
812