ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 99, 1, 2020
УДК 595.76
ЖУЖЕЛИЦЫ ТРИБЫ SPHODRINI LAPORTE, 1834 (COLEOPTERA,
CARABIDAE) ФАУНЫ ВОСТОЧНОЙ ЕВРОПЫ
© 2020 г. А. В. Пучков,1* О. Р. Александрович2**
1Институт зоологии им. И. И. Шмальгаузена НАН Украины
ул. Богдана Хмельницкого, 15, Киев-30, ГСП, 01601 Украина
*e-mail: putchkov@izan.kiev.ua; 1954caradus@gmail.com
2Институт биологии и наук о Земле
ул. Арцишевского, 22Б, г. Слупск, 76-200 Польша
**e-mail: oleg.aleksandrowicz@apsl.edu.pl
Поступила в редакцию 26.09.2019 г.
После доработки 30.01.2020 г.
Принята к публикации 30.01.2020 г.
Для Восточной Европы указано 24 вида из 8 родов трибы Sphodrini Laporte, 1834. Наиболь-
шее число видов отмечено для родов Calathus Bonelli, 1810 (10) и Laemostenus Bonelli, 1810
(6). Род Pseudotaphoxenus Schaufuss, 1865 представлен тремя, а роды Dolichus Bonelli, 1810,
Platyderus Stephens, 1827, Sphodrus Clairville, 1806, Synuchus Gyllenhal, 1810, Taphoxenus
Motschulsky, 1850 - одним видом каждый. Для всех видов приведены очерки с данными по рас-
пространению, биономии и экологии. Даны определительные таблицы 27 видов и подвидов
из 8 родов.
Ключевые слова: Coleoptera, Carabidae, Sphodrini, Восточная Европа, распространение, эколо-
гия, определитель.
DOI: 10.31857/S0367144520010141
Жужелицы трибы Sphodrini Laporte, 1834 (Coleoptera, Carabidae) характеризуются
небольшими или сравнительно крупными (6-30 мм) размерами; уплощенным или вы-
пуклым телом; чаще черной или бурой окраской, иногда двуцветной с металлическим
блеском. От близких групп (например трибы Platynini) виды Sphodrini не всегда четко
отличимы по наружным признакам, но хорошо - по строению гениталий. Классифика-
ция трибы недостаточно разработана и границы ее не вполне четки, особенно пробле-
матичен статус подтрибы Synuchina. В ряде работ (Крыжановский, 1983; Kryzhanovskij
et al., 1995) она включена в трибу Platynini (как и род Platyderus), однако в последнем
каталоге (Hovorka, 2017a, 2017b) Synuchina перенесена в состав трибы Sphodrini, а род
Platyderus отнесен к подтрибе Atranopsina Baehr, 1982. Согласно этим публикациям,
в настоящее время триба Sphodrini в Палеарктике представлена 6 подтрибами, 38 рода-
ми и более чем 730 видами. Для Европы указано почти 200 видов из 13 родов (Casale,
2003, 2017; Hovorka, 2017a, 2017b).
190
Специальных исследований трибы Sphodrini в Восточной Европе не проводили, хотя
во многих публикациях (в том числе и фундаментальных) приведены особенности рас-
пространения ряда видов, встречающихся в этом регионе (Верещагина, 1984; Casale,
1988, 2003, 2017; Kryzhanovskij et al., 1995; Assmann, 2004; Hovorka, 2017a, 2017b).
Отдельные сведения представлены для некоторых стран и регионов - Украины (Поно-
марчук, 1963; Петрусенко О. А., Петрусенко С. В., 1971; Эйдельберг и др., 1988; Хо-
менко, Вакаренко, 1993; Пучков, 1990, 2001, 2012, 2018; Різун, 2003а; 2003b, 2009; Ки-
риченко, Бабко, 2007), Белоруссии и восточной Польши (Александрович, 2014;
Aleksandrowicz et al., 2016), Молдавии (Nekuliseanu, Matalin, 2000; Bacal et al., 2013),
лесной и лесостепной зон России (Феоктистов, Душенков, 1982; Sharova, Denisova,
1995; Филиппов, 2006). Большинство этих публикаций посвящено фаунистике. Напри-
мер, для Украины указано 18, Молдавии - 14, Белоруссии - 7 видов трибы Sphodrini,
но некоторые из них, достоверно отмеченные в Украине или Белоруссии (Пучков,
2012; Aleksandrowicz et al., 2016), в последнем каталоге (Casale, 2017) не приведены.
Наиболее полная сводка по фауне Sphodrini Восточной Европы - аннотированный спи-
сок жужелиц России и сопредельных территорий, в котором для европейской части
этого региона указано 24 вида из 8 родов (Kryzhanovskij et al., 1995).
Несмотря на хорошую фаунистическую изученность трибы в Восточной Европе,
распространение отдельных видов и подвидов в пределах изучаемого региона требует
уточнения, как и определительные таблицы, особенно для трудноразличимых видов.
Наиболее полные определительные таблицы существуют для видов рода Calathus (Ве-
рещагина, 1984) и подтрибы Sphodrina (Casale, 1988), по которым можно идентифици-
ровать все встречающиеся в Восточной Европе виды этих групп. По другим работам
можно определить от 13 (Assmann, 2004) до 15 видов (Крыжановский, 1965) Sphodrini,
распространенных в Восточной Европе. Сведения по биономии и экологии отдельных
видов (особенно из рода Calathus) для восточноевропейских регионов (Sharova,
Denisova, 1995; Петрусенко и др., 1999; Різун, 2003а; Филиппов, 2006; Александрович,
2014; Пучков, 2018), значительно менее полны, чем таковые для Западной и Централь-
ной Европы (Gilbert, 1956; Kůrka, 1972; Dijk, 1973, 1978, 1994, 1996; Georgely, Lovei,
1987; Aukema, 1995).
МАТЕРИАЛ И МЕТОДИКА
Настоящая работа подытоживает результаты многолетних (1978-2017 гг.) полевых наблюдений
и сборов авторов, обработки литературных данных и коллекций Зоологического института РАН
(С.-Петербург, Россия). У ряда проблемных видов рода Calathus (C. cinctus, C. melanocephalus,
C. mollis, C. distinguendus, C. fuscipes) изучены гениталии самцов и уточнено их распростране-
ние в некоторых регионах.
Следует отметить, что понятие Восточная Европа (как и ее границы) считается не вполне од-
нозначным, так как существует достаточно много ее определений. Границы выделения этого
региона, связанные с административно-территориальными, культурными и социальными пока-
зателями, очень усложняют зоогеографические исследования. Ряд таких противоречий проявил-
ся и при составлении каталогов жуков подотряда Adephaga Палеарктики (Catalogue …, 2017),
так как ее территория была разделена в основном по административным границам (за исключе-
нием Китая и, частично, России). При этом в Европу попали как Большой Кавказ, так и все стра-
ны Закавказья, что географически представляется спорным.
Рассматриваемая в данной работе территория (Восточная Европа) включает всю европейскую
часть бывшего СССР, но без Урала, Западного Казахстана и Кавказа. Однако, в очерках неко-
торых проблемных видов дополнительно рассмотрены особенности их распространения в вос-
191
точной Польше (до р. Висла) и Западном Казахстане (на восток до р. Урал). Всего на основе
географического деления (Физико-географическое районирование СССР, 1968) и данных о диф-
ференциации фауны жужелиц (Kryzhanovskij et al., 1995) в пределах региона нами выделено
14 регионов (рис. 1) с дополнениями, уточнениями и изменениями, особенно для Украины (Ма-
ринич и др., 1982; Пучков, 2012, 2018). Северная граница Кавказа проведена по линии Анапа-
Майкоп-Черкесск-Пятигорск-Грозный-Хасавюрт-Махачкала. Большой Кавказ и закавказские
республики - Грузию, Армению и Азербайджан - в пределы Восточной Европы (куда их отнес-
ли в Каталоге жуков Палеарктики) мы не включаем.
В работе использована классификация трибы, представленная в последнем каталоге жуков
Палеарктики (Hovorka, 2017а, 2017b; Casale, 2017), а видовые очерки (в пределах родов) приве-
дены в алфавитном порядке.
Коэффициент фаунистического сходства (по Жаккару) определяли с помощью программы
PAST (Hammer et al., 2001).
РЕЗУЛЬТАТЫ
Всего для территории Восточной Европы достоверно отмечено 24 вида из 8 родов
5 подтриб (см. таблицу), которые охватывают около 3 % видового и 20 % родового со-
става Sphodrini фауны Палеарктики.
Ниже представлены таблицы для определения родов, видов и подвидов Sphodrini,
встречающихся в Восточной Европе, а также эколого-фаунистические очерки всех ви-
дов, отмеченных в исследуемом регионе, с особенностями их распространения и био-
номии. Морфологические описания родов и видов не приведены, так как достаточно
полно даны в ряде работ (Верещагина, 1984; Casale, 1988; Aukema, 1995).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ ТРИБЫ SPHODRINI ФАУНЫ ВОСТОЧНОЙ ЕВРОПЫ
1(14). Лапки сверху голые или с отдельными тонкими и длинными волосками.
2(3). Отросток переднегруди между передними тазиками не окаймлен. Последний
членик губных щупиков топоровидно расширен. Переднеспинка почти диско-
видная (подтриба Synuchina)
1. Synuchus Gyll.
3(2). Отросток переднегруди на вершине окантован. Последний членик губных щупи-
ков не расширен. Задние углы переднеспинки почти прямоугольные или окру-
глены только на вершине (рис. 2-12), но иногда развиты в виде остроугольных
лопастей (рис. 13-16).
4(7). Отросток переднегруди образует острый киль, скошенный к среднегруди. Когот-
ки зазубренные или гребневидные.
5(6). Крупнее: 14-20 мм. Передние лапки самца с почти прямоугольными основными
члениками. Переднеспинка с округленными боками и задними углами (рис. 2).
Полиморфный по окраске вид: более обычна форма с черной головой и передне-
спинкой и большим красно-желтым треугольным пятном на середине основания
надкрылий (подтриба Dolichina)
2. Dolichus Bon.
6(5). Мельче: 8-12 мм, если чуть больше (до 14.5 мм), то тело всегда черное, а перед-
неспинка почти квадратная или слегка трапециевидная, ее основание явственно
шире переднего края (рис. 3-7). Передние лапки самца с треугольно расширен-
ными члениками (подтриба Calathina)
3. Calathus Bon.
192
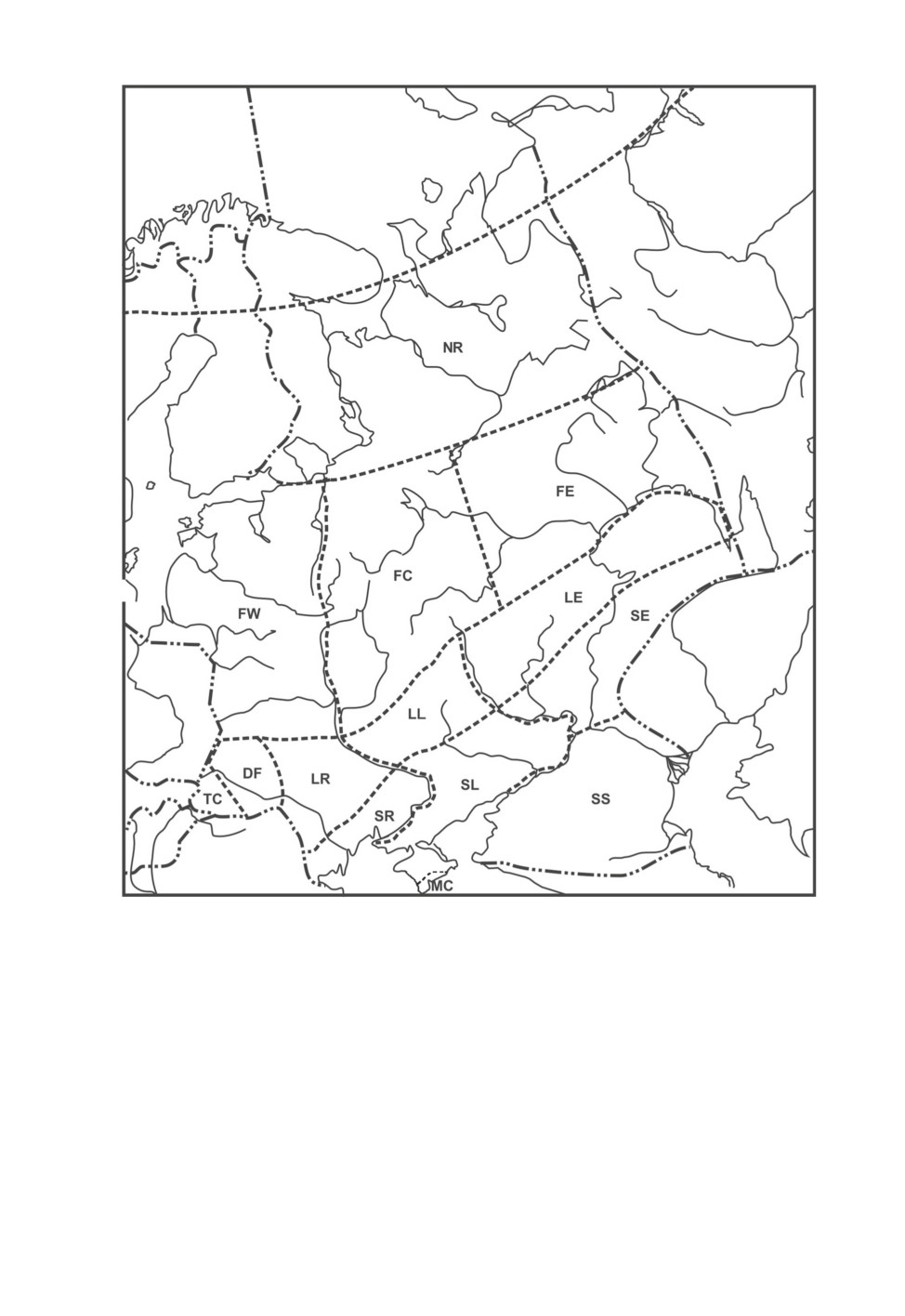
Рис. 1. Карта основных географических регионов Восточной Европы.
▬••▬••▬•• - границы Восточной Европы, ------------ - границы ее отдельных географических регионов.
NR - север Русской равнины, Карелия и Кольский полуостров (кроме зоны арктических пустынь) на юг
до 60° с.. ш.; FW - запад лесной зоны (Прибалтика, Белоруссия, Полесье Украины на восток до рек
Днепр и Ловать); FC - центр лесной зоны (левобережное Полесье Украины и Белоруссии, Нечерноземье
Центральной России на восток до 48° в. д.); FE - восток лесной зоны (от 48° в. д. на восток до Урала);
TC - Карпаты (включая Закарпатскую низменность); DF - подзона широколиственных лесов Украины
и север Молдавии (Кодры); LR - западная Лесостепь (Молдавия, правобережная Лесостепь Украины
на восток до р. Днепр); LL - центральная Лесостепь (левобережная Лесостепь на восток до р. Дон);
LE - восточная Лесостепь (от р. Дон до Урала); SR - правобережная Степь (южная часть Молдавии,
правобережная Степь Украины до р. Днепр); SL - левобережная Степь (от р. Днепр на восток до р. Дон);
SE - восточная Степь (от р. Дон на восток до Урала и Западного Казахстана); SS - юг Русской равнины
(степная и полупустынная зоны, от 47° с. ш. на юг до Предкавказья, на восток до Западного Казахстана
и Каспийского моря); MC - Горный Крым с предгорьями (включая Южный берег Крыма).
193
Видовой состав и распространение видов трибы Sphodrini в Восточной Европе
Подтриба AT R A N O P S I N A Baehr, 1982
Род PLATYDERUS Stephens, 1827
rufus (Duftschmid, 1812)
TC DF (юг) LR LL LE SR SL SE SS
Подтриба C A L AT H I N A Laporte, 1834
Род CALATHUS Bonelli, 1810
Подрод C a l a t h u s Bonelli, 1810
distinquendus Chaudoir, 1846
SR (юг) SL (юг) SE (юг) SR MC
fuscipes fuscipes (Goeze, 1777)
FW FC FE TC DF LR LL LE SR SL SE SS (север)
fuscipes punctipennis Germar, 1823
SR (юг) ?MC
syriacus Chaudoir, 1863
?SL (юг) SS MС
Подрод N e o c a l a t h u s Ball et Negre, 1972
ambiguus (Paykull, 1790)
NR FW FC FE TC DF LR LL LE SR SL SE SS
MC
cinctus (Motschulsky, 1850)
FW TC ?SL ?SS ?MC
erratus (C. R. Sahlberg, 1827)
NR FW FC FE TC DF LR LL LE SR SL SE SS
MC
melanocephalus (Linnaeus, 1758)
NR FW FC FE TC DF LR LL LE SR SL SE SS
MC
metallicus Dejean, 1828
TC DF (север Молдавии)
micropterus (Duftschmid, 1812)
NR FW FC FE TC DF LR (северо-запад)
mollis (Marsham, 1802)
SR SL SE SS MC
Подтриба D O L I C H I N A Audouin et Brullé, 1834
Род DOLICHUS Bonelli, 1810
halensis (Schaller, 1783)
FW FC FE TC DF LR LL LE SR SL SE SS MC
Подтриба S P H O D R I N A Laporte, 1834
Род LAEMOSTENUS Bonelli, 1810
Подрод A n t i s p h o d r u s Schaufuss, 1865
jailensis Breit, 1914
MC
Подрод L a e m o s t e n u s Bonelli, 1810
venustus (Dejean, 1828)
MC
Подрод P r i s t o n y c h u s Dejean, 1828
cimmerius (Fischer von Waldheim, 1823)
MC
sericeus sericeus (Fischer von Waldheim, 1824)
?LE (юг) SS SL
sericeus tauricus Dejean, 1828
SR SL MC
terricola terricola (Herbst, 1784)
NR FW FC FE TC DF LR LL LE SL SE SS MC
terricola punctatus Dejean, 1828
SR (юг)
tichyi Kult, 1946
DF (юг Западного Подолья, Кодры, Товтры)
Род SPHODRUS Clairville, 1806
leucophthalmus (Linnaeus, 1758)
FW FC FE TC DF LR (север) LL (север)
194
Таблица (продолжение)
Род PSEUDOTAPHOXENUS Schaufuss, 1865
rufitarsis (Fischer von Waldheim, 1823)
SR (юг) SL SE SS
angusticollis (Fischer von Waldheim, 1823)
?SL SE SS
horvathi (Jedlička, 1952)
?SE ?SS
Род TAPHOXENUS Motschulsky, 1850
gigas (Fischer von Waldheim, 1823)
LR (юг) LL (юг) LE (юг) SR SL SE SS
Подтриба S Y N U C H I N A Lindroth, 1956
Род SYNUCHUS Gyllenhal, 1810
vivalis (Illiger, 1798)
NR FW FC FE TC DF LR LL LE SR SL SE SS
MC
П р и м еч а н и е. Границы географических регионов те же, что на рис. 1; «?» - находки
таксона в данном регионе маловероятны или требуют подтверждений.
7(4). Отросток переднегруди обычно тупо скошен к среднегруди, без киля на заднем
скате. Коготки гладкие, без зубца даже у основания.
8(9). 3-й членик усиков не длиннее 1-го или 4-го. Передний край переднеспинки посе-
редине со слабым тупоугольным выступом (рис. 12); задние углы переднеспинки
тупоугольные, на вершине округлены. Зубец подбородка на вершине простой,
округленный. Верх тела красно-бурый, усики и ноги светлее. Тело не более 8 мм.
Правая парамера эдеагуса маленькая, изогнутая (подтриба Atranopsina Baehr,
1982)
4. Platyderus Steph.
9(8). 3-й членик усиков почти в 1.4 раза длиннее 1-го или 4-го. Передний край пе-
реднеспинки посредине почти прямой или слегка вогнутый. Зубец подбородка
на вершине раздвоен. Тело крупное (16-40 мм). Правая парамера иного строения
(подтриба Sphodrina).
10(11). Задние вертлуги заострены, у самцов вытянуты в шип
....................................................................................................... 5. Sphodrus Clairv.
11(10). Задние вертлуги на вершине закруглены.
12(13). Надкрылья почти яйцевидные, с нежно, но явственно точечными бороздками.
Крупнее (24-31 мм)
6. Taphoxenus Motsch.
13(12). Надкрылья овально-удлиненные. Точки в бороздках надкрылий едва различи-
мы. Мельче (16-20 мм)
7. Pseudotaphoxenus Schauf.
14(1). Лапки сверху покрыты короткими густыми волосками. Коготки гладкие или заз-
убрены
8. Laemostenus Bon.
Подтриба AT R A N O P S I N A Baehr, 1982
В Палеарктике 8 родов и почти 120 видов, среди которых преобладают представите-
ли рода Platyderus.
195
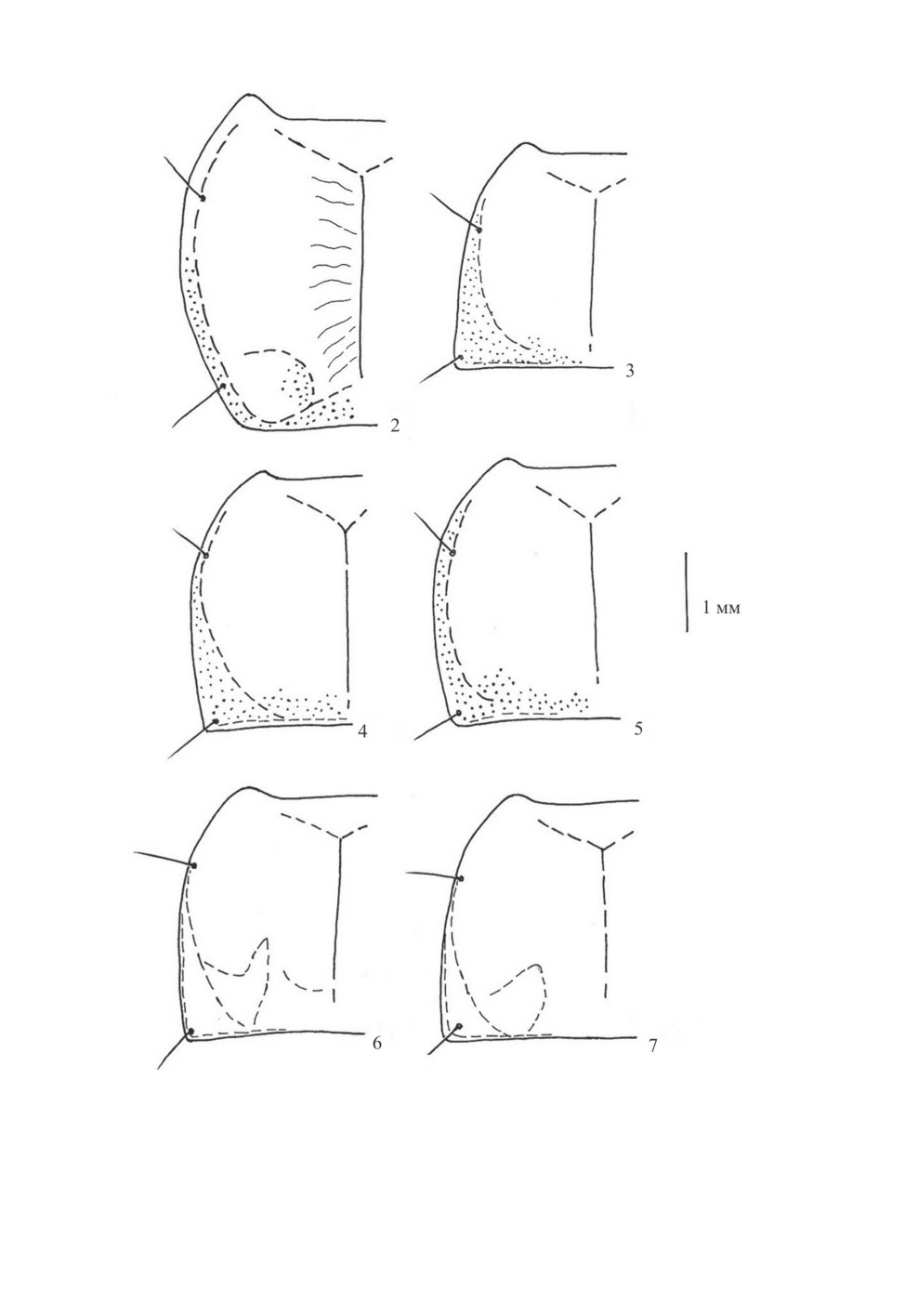
Рис. 2-7. Контур левой половины переднеспинки (по: Верещагина, 1984; Assmann, 2004,
с дополнениями).
2 - Dolichus halensis (Schall.), 3 - Calathus fuscipes (Gz.), 4 - C. distinguendus Chd., 5 - C. syriacus Chd.,
6 - C. erratus (C. R. Sahlb.), 7 - C. ambiguus (Payk.).
196
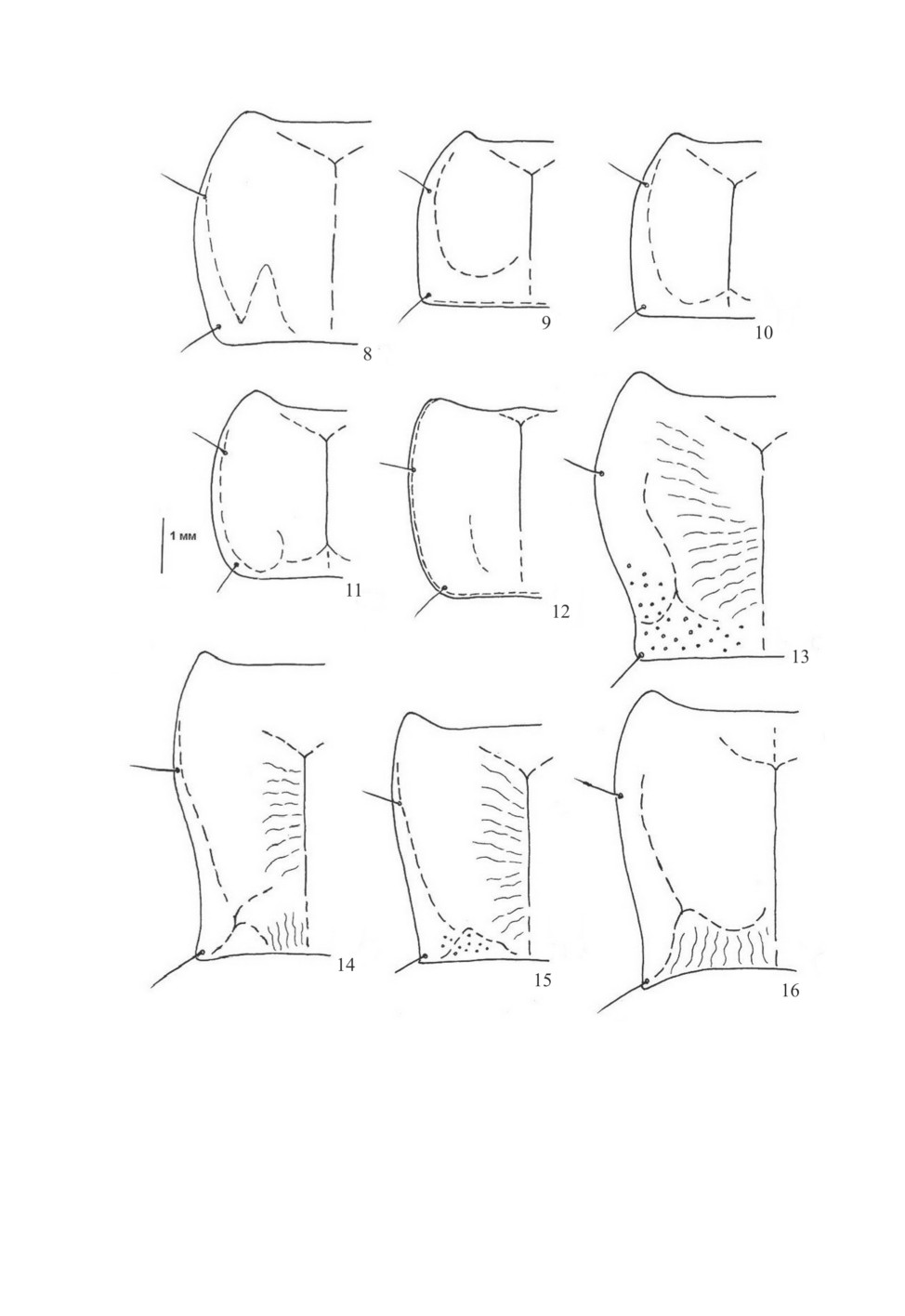
Рис. 8-16. Контур левой половины переднеспинки (по: Assmann, 2004 и ориг.).
8 - Calathus metallicus Dej., 9 - C. melanocephalus (L.), 10 - C. mollis (Marsh.), 11 - C. micropterus (Duft.),
12 - Platyderus rufus (Duft.), 13 - Laemostenus terricola (Hbst.), 14 - L. sericeus tauricus (Dej.),
15 - L. tichyi (Kult), 16 - Pseudotaphoxenus rufitarsis (F.-W.).
197
Род PLATYDERUS Stephens, 1827
Для всей Европы указано более 80 (в Палеарктике около 110) видов. Для территории
Восточной Европы указан единственный вид - P. (s. str.) rufus (Duftschmid, 1812), рас-
пространенный также в Южной и, частично, Центральной Европе.
Подрод P l a t y d e r u s Stephens, 1827
Platyderus rufus (Duftschmid, 1812).
Для Восточной Европы указан номинативный подвид, распространенный только
в западных регионах Украины (Пучков, 2012, 2018), в сопредельной юго-восточной
Польше (Burakowski et al., 1974) и в Молдавии (Nekuliseanu, Matalin, 2000; Bacal et al.,
2013). Кроме того, известен по единственной находке в Литве, на пляже Куршской ко-
сы (Ferenca et al., 2016) (см. таблицу). Другой подвид, P. rufus transalpinus Breit, 1914,
распространен в Альпах.
Мезофил. Встречается нечасто в широколиственных лесах разных типов, а также
в рощах. В Карпатах проникает до пояса буковых лесов, где иногда имеет статус суб-
доминанта (Різун, 2003а). Отмечен как случайный вид в садах, парках и лесополосах.
Жуки активны с конца марта до начала ноября. Зимуют преимущественно имаго. За-
дние крылья редуцированы, не летает (Assman, 2004).
Подтриба C A L AT H I N A Laporte, 1834
В Палеарктике около 150 видов из 3 родов.
Род CALATHUS Bonelli, 1810
В Палеарктике известны представители 8 подродов (почти 145 видов). В Европе за-
регистрировано чуть более 60 видов, а в восточной ее части - 10 видов из 2 подродов
(см. таблицу).
Подрод C a l a t h u s Bonelli, 1810
Calathus distinguendus Chaudoir, 1846.
Балканы, Кавказ, Закавказье, Турция (откуда вид описан), юг Восточной Европы.
В Восточной Европе указан для большинства южных регионов Молдавии (Nekuliseanu,
Matalin, 2000; Bacal et al., 2013), Украины (Пучков, 2012, 2018) и европейской России
(Верещагина, 1984). Для уточнения распространения вида необходимы дальнейшие
исследования в связи со смешением его с близким C. fuscipes. При этом в Украине
C. distinguendus отмечен не только по всей степной зоне, но и на юге левобережной
Лесостепи (Харьковская и Полтавская области). Установление северной границы рас-
пространения вида в правобережной части Украины требует дополнительных исследо-
ваний.
Экология изучена недостаточно, но, по-видимому, C. distinguendus более сухолюбив,
чем сходный с ним по образу жизни и облику C. fuscipes. Отмечен в типичных степных
биотопах, реже на залежах и пастбищах. Местами обычен в предгорьях и на южном
берегу Крыма. Часто встречается в степях Нижнего Поволжья (Калюжная и др., 2000).
Имаго отмечены с мая до конца августа, но чаще в июне.
198
Calathus fuscipes (Goeze, 1777).
Европа, Кавказ, Закавказье, Иран, Ирак, завезен в Северную Америку. В пределах
Восточной Европы (кроме крайнего юга и высокогорий) встречается номинативный
подвид. Распространение C. fuscipes на юге равнинной Украины, Молдавии и России
требует уточнений (см. выше). На юге Одесской обл. встречается подвид С. fuscipes
punctipennis Germar, 1823 (см. таблицу). Еще 3 подвида описаны из Северной Африки
и Турции (Hovorka, 2017a).
Полизональный, политопный мезофильный вид. Встречается в разнообразных от-
крытых биотопах, часто в парках, иногда в негустых лиственных лесах и зарослях ку-
старников, в речных и морских наносах. В Карпатах из предгорий проникает до пояса
буковых лесов (Різун, 2003а). Во всех типах агроценозов (чаще пропашных и зерновых
культурах), а также в парках и в старых садах лесной и лесостепной зон - обычный,
а иногда и массовый вид (Петрусенко и др., 1999; Александрович, 2014; Пучков, 2018).
В агроландшафтах степной зоны встречался реже, чем в Лесостепи, но был обычен
на залежах; нередок и в урбоценозах. Имаго отмечены с конца марта до середины ок-
тября, но чаще в июне (степная зона, Горный Крым) или в июне и июле (Лесостепь).
Самки с яйцами встречались в конце мая - июне (Лесостепь) и в июле - начале августа
(Степь, горы Крыма). Имаго активны ночью, а днем - только ранней весной. Зимуют
преимущественно личинки. Выход молодых жуков чаще наблюдался в июне - нача-
ле июля (юг Лесостепи, Степь) и до конца августа, что сходно с фенологией в других
регионах Европы (Lindroth, 1945; Kůrka, 1972). Копуляция отмечена в июле и сентябре
(северная подзона Степи). Плодовитость колебалась от 100 до 200 яиц (Kůrka, 1972).
Имаго преимущественно короткокрылые (Assmann, 2004), не летают (Lindroth, 1945).
Calathus syriacus Chaudoir, 1863.
Крайний юго-восток Европы, Кавказ, Закавказье, Малая Азия. В пределах Восточ-
ной Европы - крайний юг степной зоны, Крым.
Степной вид, но обычен и в Горном Крыму, особенно в открытых ландшафтах. В по-
левых агроценозах отмечены единичные особи, немного чаще встречался в садах пред-
горий Крыма (Эйдельберг, 1989). Имаго активны с мая до августа (чаще в июне - нача-
ле июля). Ксеромезофил, более сухолюбив, чем другие виды подрода.
Подрод N e o c a l a t h u s Ball et Nègre, 1972
Calathus ambiguus ambiguus (Paykull, 1790).
Вся Европа, Кавказ, Закавказье, Турция (подвид C. ambiguus rugicollis Putzeys, 1873),
Малая и Средняя Азия, Казахстан, юг Западной Сибири. В Восточной Европе - повсе-
местно (кроме высокогорий и крайнего севера).
Политопный мезофильный вид (но тяготеет к суходольным участкам). Встречается
в разнообразных открытых травянистых биотопах, местами обычен и в изреженных
лиственных лесах и кустарниках. Имаго изредка поднимаются на растения, попадают-
ся в речных и морских наносах. Иногда встречался на солонцах и по берегам соленых
водоемов. В Карпатах из предгорий проникает до пояса буковых лесов (Різун, 2003а).
В Польше и Белоруссии предпочитает суходольные луга и полевые агроценозы на лег-
ких почвах (Александрович, 2014). Во всех типах агроландшафтов Украины - обыч-
199
ный вид, но чаще в лесной и лесостепной зонах, где преобладает на пахотных землях
(чаще на пропашных, зерновых культурах и многолетних травах). Нередок в садах,
парках и лесополосах, а также ягодниках и виноградниках (особенно с июля до кон-
ца сентября) (Пучков, 2018). Среди доминантных видов жужелиц агроценозов указан
как один из весьма эффективных энтомофагов колорадского жука (по данным сероло-
гического анализа, доля питавшихся вредителем жужелиц достигала 70 %) (Коваль,
2009).
Имаго встречаются с конца марта (юг) - середины апреля (Полесье) до октября,
но чаще поздним летом. В Карпатах отмечен с начала апреля до конца августа, а в За-
падном Подолье и Приднестровье молодые имаго зарегистрированы с конца мая до
начала июля (Різун, 2003а). Зимуют преимущественно личинки разных возрастов
(в степном Крыму весной обычны и перезимовавшие жуки). В июне - начале июля на-
блюдается массовое появление молодых жуков. Откладка яиц отмечена с середи-
ны июля до второй половины октября (Gilbert, 1956; Kůrka, 1972), плодовитость дости-
гала 40-60 яиц (Kůrka, 1972). Имаго активны преимущественно в сумеречные и ночные
часы. Летает в июле-августе (Lindroth, 1945). Имаго могут питаться семенами одуван-
чика Taraxacum officinale Wiggers, 1780 (Honek et al., 2007).
Calathus cinctus Motschulsky, 1850.
Почти вся Европа, Турция, Иран, Иордания. В Восточной Европе изредка отмечался
на крайнем западе Украины (Закарпатье и западный макросклон предгорий Карпат)
(Kryzhanovskij et al., 1995; Різун, 2003а) и Белоруссии (Александрович, Козулько,
2017), в Литве (Ferenca, 2016) и Латвии (Telnov et al., 2008) (см. таблицу). Указание
для юга Русской равнины и Крыма (Kryzhanovskij et al., 1995) требует дополнительных
исследований, так как этот вид часто смешивают с очень сходными C. melanocephalus
и C. mollis. В результате изучения гениталий самцов с юга степной зоны Восточной
Европы и из Горного Крыма С. cinctus не был обнаружен. Однако этот вид активно
расширяет свой ареал и, например, отмечен уже в северо-восточных регионах Польши
(Aleksandrowicz et al., 2016) и в Калининградской обл. России (30.X.2000, Куршская
коса, серая дюна, в подстилке, 1 ♂; О. Александрович). Вполне достоверны указания
С. cinctus и для Северного (Ставрополье) и Восточного Кавказа (Дагестан) (Сигида,
1993; Абдурахманов и др., 2011; Ильина, Алиев, 2015).
Политопный мезофильный вид. Особенности экологии в Восточной Европе изучены
недостаточно. В Западной Европе нередок на суходольных лугах, зарастающих дюнах,
полях и пустошах, где встречается вместе с C. melanocephalus (Aukema, 1995). Указа-
ния для агроценозов (картофельные поля) степного Крыма (Коваль, 2009) требуют
уточнений. В Нидерландах (Aukema, 1995) и северной Польше (наши наблюдения
в Поморье) отмечен с начала июля до середины ноября с максимумом активности
в конце августа - первой половине сентября. Массовое появление молодых жуков за-
регистрировано во второй половине июня (особенно на открытых участках с легкими
почвами и на прибрежных дюнах). Период откладки яиц длится 9 недель, максималь-
ное среднее число отложенных яиц в неделю достигает 50 (Aukema, 1995). Для С. cinctus
характерен крыловой диморфизм, длиннокрылые особи летают в конце июля - нача-
ле августа (Aukema, 1995; Šustek, 1999).
200
Calathus erratus erratus (C. R. Sahlberg, 1827).
Европа, Кавказ, Турция, Казахстан, Средняя Азия, юг Западной Сибири. В Восточ-
ной Европе - почти повсеместно (кроме крайнего юга степной зоны). Известно
два подвида, из которых в Восточной Европе распространен номинативный (Hovorka,
2017a).
Кустарниково-луговой мезофильный вид (но более влаголюбив, чем C. ambiguus).
Встречается в разнообразных открытых и затененных биотопах, но чаще - на лугах
разных типов. Нередок в пойменных древесных стациях, на заболоченных участках,
в изреженных лиственных лесах и кустарниках. В Польше и Белоруссии предпочитает
молодые сосновые леса и верещатники, реже встречается на суходольных лугах (Алек-
сандрович, 2014). Попадается в речных наносах. В Степи тяготеет к байрачным лесам,
реже отмечен и на прилегающих участках мезофитных лугов. В Карпатах из предгорий
проникает до пояса буковых лесов (Різун, 2003а). Обычен в полевых агроценозах
на песчаных почвах Ленинградской (Гусева, Коваль, 2008) и Московской (Шарова, Со-
болева-Докучаева, 1984) областей. В агроландшафтах Украины в незначительном ко-
личестве зарегистрирован только в лесной зоне и Лесостепи (Пучков, 2018). Иногда
относительно обычен в садах и городских парках (Киев, Харьков), особенно в июне.
Взрослые жуки отмечены с начала апреля до ноября, но чаще в июле-августе. Имаго
активны ночью, а днем - только ранней весной. Зимуют преимущественно личинки,
реже имаго. Окукливание отмечено в апреле и мае (Gilbert, 1956), а появление молодых
особей - с конца мая до середины сентября, но чаще в июне - начале августа. Описан
поливариантный жизненный цикл с летне-осенним размножением - с июня до ноября
(Sharova, Denisova, 1995). При этом наиболее многочисленная часть популяции вида,
зимующая в стадии личинки, размножается в июле-октябре, а меньшая часть жуков,
второго года жизни, - в июне-августе. Плодовитость составляла 105-135 яиц (Kůrka,
1972). Характерен крыловой диморфизм, но длиннокрылые особи не летают (Lindroth,
1945).
Calathus melanocephalus melanocephalus (Linnaeus, 1758).
Западнопалеарктический вид, включает 5 подвидов, из которых в Восточной Европе
повсеместно распространен номинативный (Hovorka, 2017a). Изучение гениталий под-
твердило его распространение в большинстве регионов Украины, а также на юге Рус-
ской равнины и на Кавказе.
Политопный полизональный мезофильный вид, один из наиболее многочисленных
видов жужелиц в регионе. Обитает в большинстве биотопов, как открытых, так и зате-
ненных, часто встречается в изреженных лесах и зарослях кустарников. Может подни-
маться на растения. В Карпатах из предгорий проникает до субальпийского пояса
(Різун, 2003а). Обычный (иногда и доминантный) вид в большинстве агроценозов
(Шарова, Соболева-Докучаева, 1984; Гусева, Коваль, 2008; Александрович, 2014; Пуч-
ков, 2018). В Украине часто отмечается в садах, парках и, особенно, на залежах и паст-
бищах (Пучков, 2018).
Имаго встречаются с середины марта до конца ноября, чаще в июне - начале июля
(Степь, Крым) или в июле-августе (Лесостепь), но нередки и в сентябре-октябре. Жу-
ки активны преимущественно ночью, но весной часто и днем. Выход имаго из куколок
отмечен с июля до конца августа (Лесостепь) или в середине сентября (Степь). В авгу-
201
сте (в степной зоне) у личинок и имаго наблюдается диапауза (Петрусенко, Петрусен-
ко, 1971). Зимуют преимущественно личинки всех возрастов, реже имаго, способные
к повторному размножению (Kůrka, 1972). Например, перезимовавшие жуки постоян-
но встречаются поздней весной в степном Крыму. Развитие куколки длится около 2 не-
дель. Молодые имаго отмечены уже в мае (чаще с середины месяца), обычно с кон-
ца июня до третьей декады июля с максимумом в первой половине этого месяца.
Копуляцию наблюдали весной, а также в конце лета и в сентябре (чаще в первой поло-
вине) (Kůrka, 1972). Характерен крыловой диморфизм (Lindroth, 1945), редко летит
на свет (Šustek, 1999). В Украине и Белоруссии откладка яиц отмечена с конца июля
до октября, хотя в Центральной Европе она наблюдалась и в марте (Gilbert, 1956). От-
кладка яиц длится 10 недель, максимальное среднее число отложенных за неделю
яиц достигает 65 в начале сентября (Aukema, 1995). Плодовитость может составлять
около 200 яиц. Длительность развития яиц в зависимости от температуры колебалась
от 10-12 до 30 дней (Gilbert, 1956).
Calathus metallicus metallicus Dejean, 1828.
Горы Центральной и Юго-Восточной Европы, запад Турции. Известны 4 подвида,
из которых в Восточной Европе (Карпаты) распространен номинативный (Hovorka,
2017a; Casale, 2017). Приведен и для Молдавии (Nekuliseanu, Matalin, 2000), что требу-
ет уточнений, так как это указание касается не только небольших возвышенностей
(Кодры), но и равнинных территорий (долины рек Прут и Днестр) и даже некоторых
засушливых участков (дубовые леса юга Молдавии).
Европейский монтанный мезогигрофильный вид. В Карпатах чаще встречается
в высокогорье (субальпийский и альпийский пояса на высотах 1000-1900 м над ур. м.),
где он иногда составлял около 25 % всех отловленных Carabidae. Иногда (как доми-
нант) отмечен и в верхнем (700-1000 м над ур. м.) поясе еловых лесов северо-западно-
го макросклона массива Черногора (Різун, 2003а). В Центральной и Южной Европе
нижняя граница распространения отмечается от 800 (Чехия, Словакия) до 1200 м
над ур. м. (Балканы) (Kůrka, 1972).
Зимуют, по-видимому, преимущественно имаго, но иногда и личинки старших воз-
растов, так как молодые жуки отмечены уже с начала или середины мая (как и отклад-
ка яиц). Взрослые жуки зарегистрированы до второй декады августа (Kůrka, 1972;
Різун, 2003а). Задние крылья редуцированы, жуки не летают (Aukema, 1995).
Calathus micropterus (Duftschmid, 1812).
Европа, Кавказ, Иран, Афганистан, Урал, Сибирь, Дальний Восток. В Восточной Ев-
ропе - Карпаты, Западное Подолье, лесная зона и почти вся правобережная Лесостепь
Украины.
Лесной полизональный мезофильный вид (встречается иногда и на увлажненных
участках). Обычен в лесах разных типов на всей территории ареала. В Карпатах отме-
чен в поясе хвойных лесов и субальпийском поясе (Різун, 2003а). В трансформирован-
ных ценозах единичные находки отмечены на полях в Московской (Шарова, Соболе-
ва-Докучаева, 1984) и Ленинградской (Гусева, Коваль, 2008) областях, а также
в Белоруссии (Александрович, 2014).
202
Задние крылья редуцированы, жуки не летают (Lindroth, 1945). Имаго активны пре-
имущественно ночью, встречаются с конца апреля до конца июля, но чаще в июне.
Зимуют обычно личинки старших возрастов, но нередко и имаго. Молодые жуки отме-
чены с начала июня до середины августа на высотах до 1300 м над ур. м. (Kůrka, 1972).
В Карпатах самки с яйцами встречались с середины июля до начала августа, а плодо-
витость достигала почти 20 яиц (Kůrka, 1972; Різун, 2003а). Для лесной зоны детально
описан поливариантный жизненный цикл с весенним (зимующими половозрелыми
имаго) и летне-осенним (с зимующими личинками и постгенеративными имаго) раз-
множением с апреля до ноября (Sharova, Denisova, 1995).
Calathus mollis mollis (Marsham, 1802).
Почти вся Европа (кроме северо-западной), Турция, Иран, Израиль, юг Туркмении
(Копетдаг). В Восточной Европе достоверно (на основании изучения гениталий сам-
цов) отмечен на юге степной зоны Украины (Николаевская, Херсонская и юг Донецкой
областей), а также в Горном Крыму и юге Русской равнины. Для Белоруссии не указан.
Находки в Закарпатье, Карпатах, Подолье и Полесье маловероятны. Распространение
требует уточнений в связи с трудностями различения этого вида и близких, очень по-
хожих симпатричных видов - C. cinctus и C. melanocephalus. В Северной Африке рас-
пространен подвид С. mollis atticus Gautier des Cottes, 1867.
Пойменно-луговой полизональный мезофильный вид (более тепло- и сухолюбив,
чем C. melanocephalus). Встречается чаще в открытых стациях и пойменных биотопах
(изреженные древесно-кустарниковые сообщества, заливные и суходольные луга).
В Европе обычен на слабо заросших дюнах, даже на сыпучем песке (Aukema, 1995).
Сообщение о находках на берегах водоемов, а иногда и на солончаках (Верещагина,
1984), требует проверки. Отмечен на пастбищах, реже в старых садах, парках и лесо-
полосах. Единичные находки сделаны также на полевых культурах, однако обитание
вида в агроценозах все же изучено недостаточно (также из-за смешения с близкими
таксонами).
Имаго активны обычно ночью. Встречается с конца мая до начала октября, но чаще
в июне-июле. Выход жуков из куколок отмечен уже в начале июня. Зимуют, по-види-
мому, личинки, реже взрослые жуки. Массовое появление молодых жуков - с середи-
ны июня. Откладка яиц длится 8 недель, максимальное среднее число отложенных
за неделю яиц (в начале сентября) достигает 40 (Aukema, 1995). Характерен крыловой
диморфизм, длиннокрылые особи летают (Aukema, 1995).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА CALATHUS
(ПО: ВЕРЕЩАГИНА, 1984; AUKEMA, 1995; ASSMANN, 2004, С ДОПОЛНЕНИЯМИ)
1(6). Надкрылья с многочисленными порами на 3-м и 5-м промежутках и прилежащих
бороздках. Жуки одноцветно-черные, иногда ноги, основания усиков и низ тела
буро-красные. Переднеспинка с грубой пунктировкой на основании (рис. 3-5).
В Восточной Европе группа из 3 близких видов.
2(5). Метэпистерны короткие и широкие; их длина по внутреннему краю не более
чем в 1.5 раза больше их ширины на основании (рис. 17, 18). Задние углы перед-
неспинки почти прямые или чуть притуплены на вершине (рис. 3-4). В среднем
мельче: 8.5-14.0 мм. Эдеагус не расширен в средней части, его вершина слегка
заострена (рис. 26, 28).
203
3(4). Длина метэпистерн по переднему краю почти в 1.5 больше их ширины на осно-
вании (рис. 17). Переднеспинка к основанию чуть расширена (рис. 3). Дисталь-
ная часть эдеагуса явственнее сужена к вершине (рис. 28). Повсеместно
1. C. (s. str.) fuscipes Gz.
4(3). Метэпистерны короткие и широкие; их длина по внутреннему краю равна
или чуть больше ширины на основании (рис. 18). Бока переднеспинки к осно-
ванию почти параллельные (рис. 4). Дистальная часть эдеагуса слабее и более
плавно сужена (рис. 26). Юг, но распространение в Восточной Европе слабо из-
учено
2. C. (s. str.) distinguendus Chd.
5(2). Метэпистерны узкие, их длина по внутреннему краю почти в 2 раза больше ши-
рины на основании (рис. 19). Переднеспинка, как правило, с более округленными
боками, ее задние углы округло притуплены (рис. 5). Эдеагус расширен в сред-
ней и апикальной частях, его вершина притуплена и слегка изогнута (рис. 27).
В среднем крупнее: 10.5-14.5 мм. Крым, крайний юг степной зоны
3. C. (s. str.) syriacus Chd.
6(1). Надкрылья с хорошо заметными порами только на 3-м промежутке или прилежа-
щих бороздках. Верх одноцветный или двуцветный.
7(10). Задние углы переднеспинки почти прямые, слегка заострены на вершине
(рис. 6, 7).
8(9). Наибольшая ширина переднеспинки посередине, ее длина почти равна ширине,
а основание не шире середины (рис. 6). Черный или смоляно-бурый, надкрылья
самцов иногда с легким зеленоватым блеском. 7.5-11.0 мм. Повсеместно, но ре-
док в степной зоне
4. С. (Neocalathus) erratus C. R. Sahlb.
9(8). Наибольшая ширина переднеспинки в основании, кпереди она постепенно суже-
на; ее длина меньше ширины (рис. 7). Темно-коричневый, реже почти черный,
надкрылья самцов без зеленоватого блеска, у самок матовые. 8-11 мм. Почти
повсеместно
5. С. (Neocalathus) ambiguus Payk.
10(7). Задние углы переднеспинки более или менее закруглены на вершине (рис. 8-11).
11(12). Верх (особенно надкрылья) ярко металлически блестящий: медно-зеленый, ре-
же с синим отливом. Бока переднеспинки почти равномерно округлены (рис. 8).
На 3-м промежутке надкрылий 2-5 пор. Карпаты
6. C. (Neocalathus) metallicus Dej.
12(11). Верх одноцветный или двуцветный, но без металлического блеска или он едва
заметен на надкрыльях.
13(14). Темно-бурый или черный; ноги, ротовые придатки и усики светло-желтые.
Задние углы переднеспинки явственно округлены (рис. 11). Правая парамера
изогнута на вершине (рис. 20). Основные вдавления на переднеспинке не упло-
щены, без пунктировки. Мельче: 6.0-7.5 мм. Почти повсеместно, кроме крайнего
юга, но чаще на западе
7. С. (Neocalathus) micropterus Duft.
14(13). Верх одноцветный или двуцветный. Переднеспинка слабее сужена к основа-
нию, ее задние углы почти прямые, слабо округлены на вершине (рис. 9, 10).
Правая парамера другой формы (рис. 21-23).
204
15(16). Правая парамера расширена апикально, обычно с хорошо заметным крюч-
ком (рис. 21); эдеагус в вершинной части (см. сбоку) почти равномерно изо-
гнут (рис. 25). Плечо на изгибе с маленьким зубчиком. Боковые края надкрылий
(особенно у самок) слабо округлены, почти параллельные в средней части; над-
крылья в 1.21-1.23 раза шире переднеспинки. Переднеспинка не сужена к осно-
ванию, ее боковые края за серединой почти параллельные, отношение ширины
к длине - 1.25-1.29 (рис. 9). Переднеспинка обычно красная или оранжевая, го-
лова почти черная, надкрылья темно-бурые. Иногда переднеспинка коричневая,
бурая (но края просвечивают красным) или почти черная (Горный Крым и Кав-
каз), но обычно все же светлее головы. 5.8-8.2 мм. Повсеместно и часто
..................................................................... C. (Neocalathus) melanocephalus (L.).
16(15). Правая парамера не расширена, апикально с очень маленьким крючком или без
него (рис. 22, 23). Эдеагус в вершинной части (см. сбоку) с легким изгибом возле
вершины. Плечо на изгибе без явственного зубчика. Боковые края надкрылий
(у самок) сильнее округлены в средней части; надкрылья в 0.72-0.73 раза ши-
ре переднеспинки. Верх часто почти одноцветный, но переднеспинка, особенно
по бокам, светлее головы и надкрылий.
17(18). Правая парамера без крючка на вершине, иногда с малозаметным зубчиком
или его следами (рис. 22), слабее сужена дистально. Эдеагус в вершинной части
(см. сбоку) резко сужен. Боковые края надкрылий (у самок) ýже и слабее округ-
лены в средней части. Переднеспинка более узкая и параллельносторонняя,
отношение ширины к длине - 1.20-1.26. Надкрылья темно-коричневые (редко
почти черные). 6.0-8.5 мм. Запад региона. В Восточной Европе редок
............................................................................ 9. C. (Neocalathus) cinctus Motsch.
18(17). Правая парамера с очень маленьким крючком или неявственным бугорком
(рис. 23), сильнее сужена к вершине. Эдеагус в вершинной части (см. сбоку)
почти равномерно сужен к вершине (рис. 24). Боковые края надкрылий (у са-
мок) шире и явственно округлены в средней части. Переднеспинка слегка сужена
к основанию, отношение ширины к длине - 1.24-1.28 (рис. 10). Верх коричневый
или темно-коричневый; бока переднеспинки светлее. 6.5-9.0 мм. Юг региона.
Нечасто
10. С. (Neocalathus) mollis Marsh.
Подтриба D O L I C H I N A Audouin et Brullé, 1834
В Палеарктике 7 родов и около 30 видов. В Европе (в том числе Восточной) отмечен
единственный вид рода Dolichus (в Палеарктике - 2 вида), ранее включенного в род
Calathus (Крыжановский, 1983; Верещагина, 1984; Kryzhanovskij et al., 1995).
Род DOLICHUS Bonelli, 1810
Dolichus halensis (Schaller, 1783).
Транспалеарктический вид. В регионе - почти повсеместно (кроме севера и высоко-
горий), но чаще в лесной зоне и Лесостепи.
Луговой полизональный мезофильный вид. Встречается в разнообразных открытых
биотопах, иногда и в умеренно влажных местах, а также в зарослях кустарников, ре-
же - в парках. На западе Лесостепи предпочитает хорошо прогреваемые стации с изре-
205
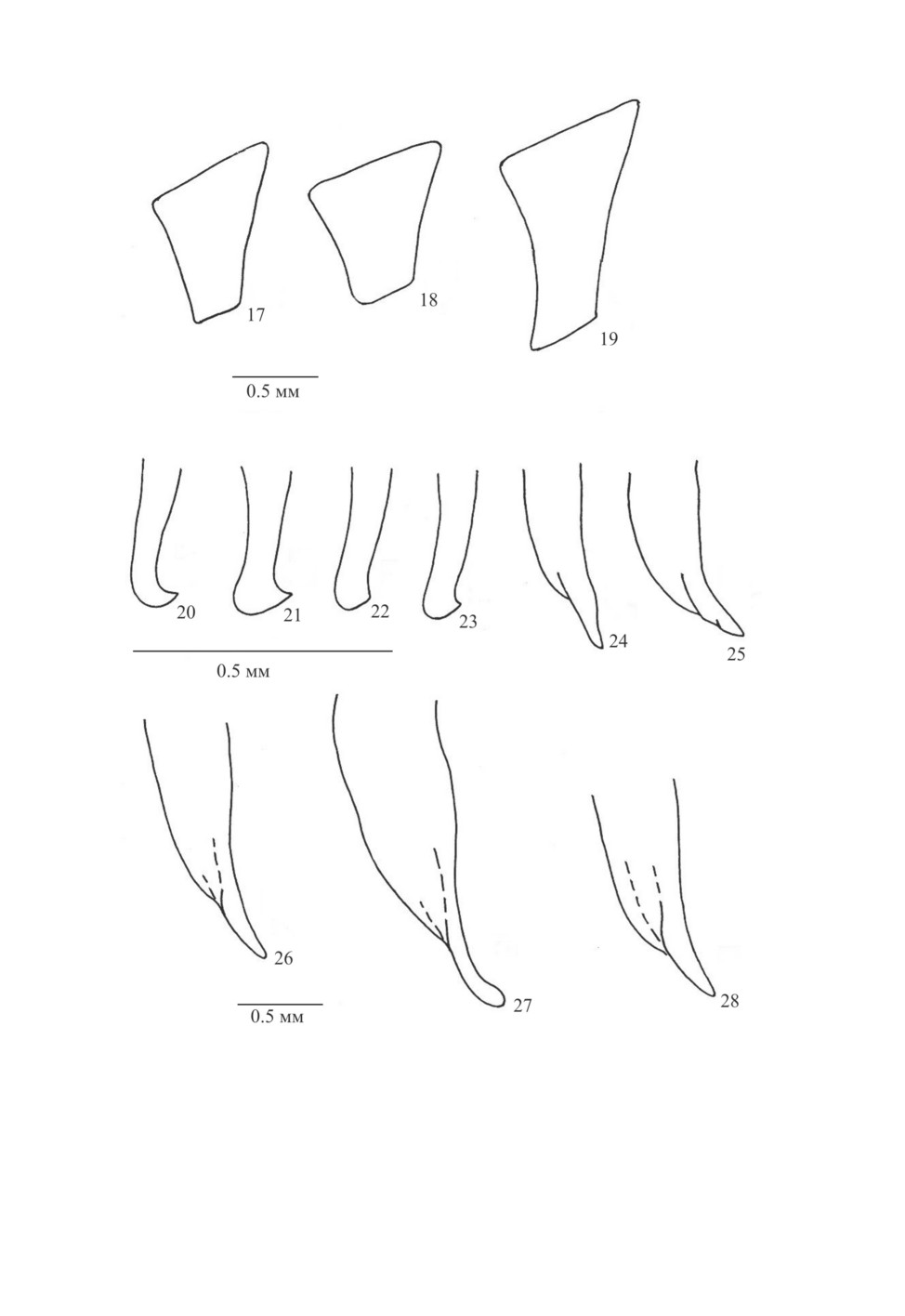
Рис. 17-28. Calathus Bon. (по: Верещагина, 1984; Assmann, 2004, с дополнениями).
17-19 - метэпистерн (17 - C. fuscipes (Gz.), 18 - C. distinguendus Chd., 19 - C. syriacus Chd.),
20-23 - вершинная часть правой парамеры (20 - C. micropterus (Duft.), 21 - C. melanocephalus (L.),
22 - C. cinctus Motsch., 23 - C. mollis (Marsh.)), 24-28 - вершинная часть эдеагуса сбоку (24 - C. mollis
(Marsh.), 25 - С. melanocephalus, 26 - C. distinguendus, 27 - C. syriacus, 28 - C. fuscipes).
206
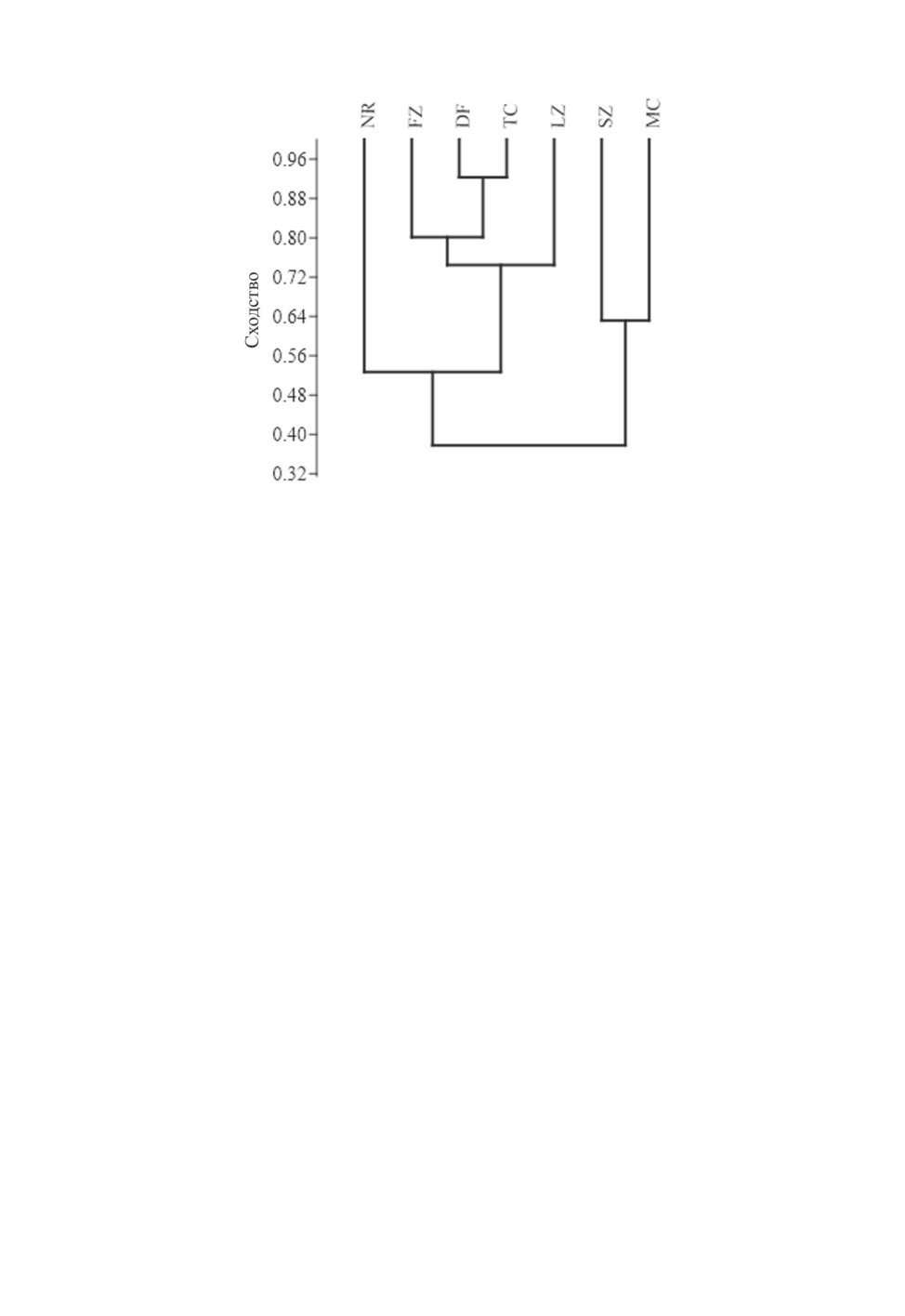
Рис. 29. Дендрограмма сходства фаун трибы Sphodrini основных географических регионов
Восточной Европы (на основе коэффициента Жаккара по программе PAST)
NR - северная часть Русской равнины, FZ - лесная зона (средняя и южная части), DF - подзона
широколиственных лесов, TC - Карпаты, LZ - лесостепная зона, SZ - степная и полупустынная
зоны, MC - Горный Крым.
женным травостоем, южнее держится на участках с густой растительностью, а в степ-
ной зоне обитает преимущественно в пойменных биотопах, на дне степных подов
и балок. Редок на полях Московской (Шарова, Соболева-Докучаева, 1984) и Ленин-
градской (Гусева, Коваль, 2008) областей, Среднего Поволжья (Алейникова, Утробина,
1969) и Литвы (Tamutis et al., 2007). Обычный вид в агроценозах юга Белоруссии (на
легких почвах) и равнинной Украины (кроме юга), где предпочитает пропашные куль-
туры и сады. На полях (особенно со второй половины июня) встречается иногда в боль-
шем количестве, чем в естественных биотопах (Пучков, 2018). На юге Степи чаще от-
мечен на орошаемых участках, но единично - на богаре.
Имаго встречаются с конца мая (степная зона) - середины июня (лесная зона, Карпа-
ты) до начала октября, но чаще в июле-августе. Самки со зрелыми яйцами отмечены
в июле и начале августа. Молодые жуки появляются с середины июня до начала по-
следней декады июля. Зимуют преимущественно личинки, но иногда и имаго. Жизнен-
ный цикл поливариантный, часть самок живет два года и дважды участвует в размно-
жении (Gergely, Lovei, 1987). Жуки активны ночью. Имаго летят на свет в конце
июля - начале августа. Среди массовых видов жужелиц агроценозов указан как один
из весьма эффективных энтомофагов колорадского жука (по данным серологического
анализа, доля питавшихся вредителем жужелиц достигала 90 %) (Коваль, 2009).
Подтриба S P H O D R I N A Laporte, 1834
В Палеарктике зарегистрировано 15 родов и более 370 видов, крупнейшие среди ко-
торых - Laemostenus и Pseudotaphoxenus. В Восточной Европе - 9 видов из 4 родов.
207
Род LAEMOSTENUS Bonelli, 1810
В Европе не менее 60, а в Палеарктике - около 210 видов. В Восточной Европе заре-
гистрированы 6 видов из 3 подродов.
Подрод A n t i s p h o d r u s Schaufuss, 1865
Laemostenus (Antisphodrus) jailensis Breit, 1914.
Эндемик Крыма; горные луга (яйлы), под камнями. Крайне редок. Биология не изу-
чена.
Подрод L a e m o s t e n u s Bonelli, 1810
Laemostenus (s. str.) venustus (Dejean, 1828).
Южная Европа, Малая Азия. Указан и для юга европейской России (Casale, 2017),
что требует подтверждений. На исследуемой территории отмечен только в горах
и предгорьях Крыма.
Лесной вид. Встречается в широколиственных лесах и перелесках, чаще на западном
макросклоне Горного Крыма. Как случайный вид зарегистрирован в садах (Эйдель-
берг, 1989). Мезофил. Активен ночью и в сумерки. Отмечен с середины мая до июля
(чаще в июне). Весенний тип размножения, зимуют имаго.
Подрод P r i s t o n y c h u s Dejean, 1828
Laemostenus (Pristonychus) cimmerius cimmerius (Fischer von Waldheim, 1823).
Юго-Восточная Европа, страны Леванта, Турция (Casale, 2017). Как и предыдущий
вид, указан для юга европейской России (Casale, 2017), что требует подтверждений.
В Восточной Европе нами отмечен только в Крыму (номинативный подвид).
Лесо-кустарниковый мезофильный вид (но встречается и в относительно сухих био-
топах редколесья). Обычен в широколиственных лесах, реже - в перелесках западного
макросклона Горного Крыма. Как случайный вид отмечен в садах (Эйдельберг, 1989).
Жуки встречались с мая до августа, но чаще в июне.
Laemostenus (Pristonychus) sericeus sericeus (Fischer von Waldheim, 1824).
Указан для Украины, Молдавии, юга европейской России, Большого и Малого Кав-
каза (Грузия, Армения), Турции и Ирана (Casale, 2017). Включает 3 подвида, статус
и распространение которых требуют уточнений. Номинативный подвид указан
для Предкавказья, Кавказа (Грузия) и даже Молдавии (но, по-видимому, это указание
относится к следующему подвиду). Laemostenus sericeus tauricus Dejean, 1828 приве-
ден для Украины, Западного и Центрального Кавказа, включая Грузию (Casale, 2017).
По нашим данным, этот подвид обычен в Горном Крыму и на юге Одесской обл. Эк-
земпляры из степной зоны Украины (юг Днепропетровской, Харьковской и Донецкой
областей) сходны с жуками номинативного подвида (Пучков, 2001, 2012, 2018).
Степной мезоксерофильный вид, стратопетробионт. Населяет выходы каменистых
пород, особенно известковых (на юге Одесской обл.). В Крыму зарегистрирован и в
пещерах (на глубине не более 20 м от входа). Иногда отмечался на шахтных отвалах
(Днепропетровская обл., окр. г. Кривой Рог). Имаго спорадично встречаются в пороях
208
грызунов. Как случайный вид зарегистрирован в садах Крыма (Эйдельберг, 1989). Жу-
ки встречаются с июня до сентября. Зимуют как личинки, так и имаго.
Laemostenus (Pristonychus) terricola terricola (Herbst, 1784).
Европа (кроме севера), на восток до Волги и Предкавказья, завезен в Северную Аме-
рику. Почти на всей территории Восточной Европы распространен номинативный под-
вид. В юго-западной и западной частях Украины зарегистрированы находки подвида
L. terricola punctatus (Dejean, 1828), указанного также для Молдавии и балканских
стран (Casale, 1988, 2017; Bacal et al., 2013).
Политопный мезофильный вид, стратоботробионт. Умеренный синантроп: обитает
в погребах и подвалах, в норах грызунов, под различными укрытиями (Bercio,
Folwaczny, 1979; Федоренко, 1988; Клаузницер, 1990; Александрович, 2014). Встреча-
ется также в разнообразных открытых биотопах: на лугах, в степи, иногда в изрежен-
ных лесах и балках. В Карпатах с равнин доходит до пояса буковых и еловых лесов (до
высот 600 м над ур. м.). Изредка как обычный вид (0.35-0.58 % всех жужелиц) был от-
мечен в буковых пралесах Карпатского биосферного заповедника (Різун, 2003а).
На юге встречается (иногда как субдоминант) в городских парках (г. Донецк, Раковка)
и лесонасаждениях (г. Ясиноватая, Донецкая обл.). Как случайный вид отмечен на зер-
новых и пропашных культурах, многолетних травах и в садах (Пучков, 1990, 2018; Ко-
валь, 2009; Сумароков, 2009; Александрович, 2014).
В Карпатах единичные особи отмечены уже с середины марта, иногда - в начале ок-
тября и даже в декабре (Різун, 2003а). В степной зоне жуки встречались чаще в кон-
це июня, а в парках Донецка - в конце мая и июле-начале августа. Молодые имаго от-
мечены в середине августа (Южная Польша) (Різун, 2003а). Зимуют имаго. Задние
крылья не развиты (Lindroth, 1945).
Laemostenus (Pristonychus) tichyi (Kult, 1946).
Эндемик Украины, известный исключительно с территории Тернопольской обл. (За-
падная Украина, Подолье, пос. Грымайлив). Биономия и экология почти неизвестны.
Мезофил. Возможно, синантропный вид. Отмечен в погребах, чаще в июне. Актив-
ность ночная, в это время жуков удавалось ловить и в бытовых помещениях (устное
сообщение В. Б. Ризуна). Возможны находки и в пещерах Тернопольщины (отмечены
остатки имаго возле пещеры «Оптимистическая», Борщевский р-н; устное сообщение
Е. А. Романенко).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ И ПОДВИДОВ РОДА LAEMOSTENUS
ФАУНЫ ВОСТОЧНОЙ ЕВРОПЫ
1(2). Голова крупная, равна по ширине переднеспинке. Задние голени без щетки из во-
лосков в дистальной части. Метэпистерны не удлинены. Переднеспинка слабо
сердцевидная, явственно сужена к основанию. Коричневый, верх без металличе-
ского блеска. 13.5 мм. Яйлы Крыма
1. L. (Antisphodrus) jailensis Breit.
2(1). Голова ýже переднеспинки. Задние голени со щеткой из волосков (иногда сла-
бо выраженной) в дистальной части. Метэпистерны удлинены. Тело от черного
до коричневого, часто с металлическим блеском.
209
3(6). Надкрылья с явственным фиолетовым, синим или зеленым металлическим бле-
ском. Первые 3 или 4 членика передних лапок самца не расширены.
4(5). Голова и мезэпистерны в грубой пунктировке. Надкрылья почти параллельносто-
ронние, слабо выпуклые, с неглубокими бороздками. Метэпистерны сильно
вытянуты, их длина почти в 2 раза больше ширины. Первые 3 или 4 членика
передних лапок самца снизу не расширены, но с волосяной подошвой. Черный
с синим металлическим блеском. 14-15 мм. Крым
2. L. (s. str.) venustus Dej.
5(4). Голова и мезэпистерны в тонких морщинках и точках. Надкрылья широкооваль-
ные, выпуклые, с глубокими пунктированными бороздками. Длина метэпистерн
не более чем в 1.4 раза больше ширины. Черный с синим, фиолетовым или зе-
леным металлическим блеском. Первые 3 или 4 членика передних лапок самца
снизу без волосяной подошвы. 17-20 мм. Крым
...................................................... 3. L. (Pristonychus) cimmerius cimmerius F.-W.
6(3). Надкрылья со слабым металлическим (чаще фиолетово-голубоватым) блеском
или черные (иногда шелковисто-блестящие) либо черно-бурые до коричневых.
Первые 3 или 4 членика передних лапок самца расширены, снизу с волосяными
подошвами.
7(10). Переднеспинка умеренно сердцевидная, ее ширина слегка (в среднем в 1.1 раза)
больше длины (рис. 13, 15). Задние голени со щеткой из золотистых волосков
в дистальной части. 1-й членик средних и задних лапок снизу между 2 рядами
шипиков в густых волосках. Средние голени самца заметно, а самки - слабее
искривлены.
8(9). Надкрылья уплощенные, явственно расширены за серединой, их боковой кант
слегка приподнят. Форма тела весьма изменчива: переднеспинка более или ме-
нее сердцевидная (рис. 13), ее максимальная ширина в 1.10-1.12 раза больше
длины и в 1.24-1.28 больше ширины в основании. Коготки зазубрены или почти
гладкие. Верх коричневый или почти черный, со слабым синим или фиолетовым
блеском, иногда без него. 12-18 мм
4. L. (Pristonychus) terricola Hbst.
В Восточной Европе известны два подвида, распространение которых требует до-
полнительных исследований:
а(б). Коготки почти гладкие или только слегка зубчатые у основания. В среднем мель-
че (12-16 мм). Глаза уплощенные. Основание переднеспинки со слабой пункти-
ровкой. Почти вся Европа (кроме севера и юго-востока)
........................................................... 4а. L. (Pristonychus) terricola terricola Hbst.
б(а). Коготки явственно зубчатые. В среднем крупнее (13-18 мм). Глаза сильнее выпу-
клые. Основание переднеспинки сильнее пунктировано. Юго-Восточная Европа,
в том числе Румыния и Молдавия, возможен и на юге Степи Украины
4б. L. (Pristonychus) terricola punctatus Dej.
9(8). Надкрылья выпуклые, по бокам равномерно округлены, их боковой кант не при-
поднят, плечи явственно выступающие. Переднеспинка перед почти прямыми
задними углами слабо выемчатая; ее максимальная ширина в 1.04-1.07 раза
больше длины и в 1.17-1.20 больше ширины в основании (рис. 15). Коготки сла-
бо зазубрены до середины. Смоляно-бурый или почти черный, надкрылья мато-
210
вые со слабым синеватым отливом, переднеспинка с умеренным блеском. Самец
неизвестен. 13.5-15.0 мм
5. L. (Pristonychus) tichyi Kult.
10(7). Переднеспинка уже, с плавно изогнутыми боками; ее длина равна ширине; за-
дние углы почти прямые, но на вершине слабо притупленные (рис. 14). Средние
голени почти прямые, дополнительное опушение из золотистых волосков в их
дистальной части слабее выражено. 1-й членик средних и задних лапок снизу
между 2 рядами шипиков в редких волосках. Коготки зазубрены у основания,
реже почти гладкие
6. L. (Pristonychus) sericeus F.-W.
Известны 3 подвида, из которых 2 отмечены для Восточной Европы, а L. sericeus
hepaticus Faldermann, 1836 распространен на востоке Большого Кавказа и в Закавказье.
а(б). Верх со слабым голубоватым блеском. Ширина надкрылий в 1.36-1.53 раза боль-
ше максимальной ширины переднеспинки, которая в 1.17-1.22 раза больше ши-
рины переднеспинки в основании. Боковой край переднеспинки слабее изогнут;
ее задние углы почти прямые; основные вдавления иногда явственно пунктиро-
ваны. 12.8-18.3 мм. Юг Русской равнины, Предкавказье
6а. L. (Pristonychus) sericeus sericeus F.-W.
б(а). Верх черный или смоляно-бурый, со слабым коричневатым блеском. Ширина
надкрылий в 1.55-1.64 раза больше максимальной ширины переднеспинки, ко-
торая в 1.15-1.21 раза больше своей ширины у основания. Боковой край перед-
неспинки изогнут сильнее (рис. 14); задние углы слабо заостренные; основные
вдавления с изреженной пунктировкой или без нее. 13.0-17.5 мм. Степная зона
от юга Молдавии до р. Дон, Горный Крым
6б. L. (Pristonychus) sericeus tauricus Dej.
Род PSEUDOTAPHOXENUS Schaufuss, 1865
Для Восточной Европы указаны 3 вида, распространение которых требует уточне-
ний. В Палеарктике известно более 110 видов, встречающихся главным образом в гор-
ных районах Средней Азии.
Pseudotaphoxenus rufitarsis (Fischer von Waldheim, 1823).
От юга Молдавии (Нижнее Приднестровье) до юга Западной Сибири. В Казахстане
и Сибири распространен номинативный подвид, а в Восточной Европе - подвид
Ps. rufitarsis major (Tschitschérine, 1895) (Casale, 2017). В Молдавии он отмечен на край-
нем юге (Nekuliseanu, Matalin, 2000), в Украине - на юго-востоке (юг Донецкой обл.),
в России - на юго-востоке европейской части (Волгоградская и Астраханская области:
Калюжная и др., 2000).
Типичный степной мезоксерофильный вид, стратоботробионт. В регионе везде
встречается единично. Биологические и экологические особенности малоизвестны.
Pseudotaphoxenus angusticollis (Fischer von Waldheim, 1823).
Юг европейской части России, Западный Казахстан. Указан и для Украины (Casale,
2017), но нами пока не обнаружен, т. е. его распространение требует подтверждений.
Особенности биологии неизвестны.
211
Это же относится и к Ps. horvathi (Jedlička, 1952), малоизвестному виду с неточным
указанием распространения - «Rossia mer.» (Casale, 2017), что, возможно, означает
юг Русской равнины (Kryzhanovskij et al., 1995). В представленную ниже таблицу
включены все 3 вида, но их морфологические характеристики все же требуют уточне-
ний.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ И ПОДВИДОВ РОДА PSEUDOTAPHOXENUS
ФАУНЫ ВОСТОЧНОЙ ЕВРОПЫ
1(2). Членики лапок дорсально почти гладкие (на 2-5-м члениках средних и задних ла-
пок у основания есть слабые бороздки). Переднеспинка явственно поперечная;
ее боковые края и задние углы округлены, а наибольшая ширина - в передней
трети. Надкрылья овальные, с широким прямым основанием; промежутки упло-
щенные, бороздки слабо выражены. 19-21 мм
1. Ps. horvathi (Jedl.).
2(1). Членики лапок с дорсальными (иногда слабовыраженными) бороздками у ос-
нования. Переднеспинка слабо поперечная. Надкрылья удлиненно-овальные,
их бороздки тонкие, но более или менее явственные.
3(4). Переднеспинка почти квадратная, слабо сужена к основанию, которое не ýже
переднего края. Бока переднеспинки слабо округлены; задние углы слегка от-
тянуты и заострены на вершине (рис. 16). Наибольшая ширина переднеспинки
в передней трети или почти сразу за серединой; вдавления у задних углов неглу-
бокие, почти гладкие. Надкрылья у основания с небольшим вдавлением около
щитка; бороздки слабо намечены по всей длине. 20-22 мм
2. Ps. rufitarsis (F.-W.).
4(3). Переднеспинка явственно выемчатая перед задними углами; ее основание ýже пе-
реднего края; наибольшая ширина перед серединой; бока слегка сходятся к поч-
ти прямым задним углам. Надкрылья с очень тонкими бороздками. 15-19 мм
........................................................................................... 3. Ps. angusticollis (F.-W.).
Род SPHODRUS Clairville, 1806
В Палеарктике отмечено 2 вида. В Европе единственный вид - Sphodrus leucophthalmus
(Linnaeus, 1758).
Sphodrus leucophthalmus (Linnaeus, 1758).
Северная Африка, Европа (кроме Северной), Малая Азия, Ближний Восток, Афгани-
стан, Йемен, Кашмир. В Восточной Европе почти повсеместно в лесной и на севере
лесостепной зон. В Молдавии указан для Нижнего Приднестровья (Nekuliseanu,
Matalin, 2000).
Редкий вид, находки которого датируются первой половиной XX в. (данные этике-
ток). Несмотря на широкий ареал, за последние десятилетия сведения о его находках
практически отсутствуют. В Северной и Центральной Европе указан как синантроп.
Отмечен в погребах, подвалах, и даже картофельных кагатах (Lindroth, 1945; Bercio,
Folwaczny, 1979; Федоренко, 1988; Александрович, 2014). Указан для городов Европы
(Клаузницер, 1990). На юге Европы встречается за пределами зданий (Assmann, 2004).
По-видимому, мезофил. Активность жуков сумеречная и ночная. Имаго зарегистриро-
ваны в конце мая, а отдельные ювенильные особи отмечены в середине июля (Рiзун,
212
2003а). Размножение осеннее, зимуют личинки (Assmann, 2004). Длиннокрылый вид,
отмечен в полете (Lindroth, 1949).
Род TAPHOXENUS Motschulsky, 1850
В Европе 2, а в Палеарктике - 11 видов. В Восточной Европе зарегистрирован один
вид - Taphoxenus gigas (Fischer von Waldheim, 1823).
Taphoxenus (s. str.) gigas (Fischer von Waldheim, 1823).
Юг Восточной Европы, Предкавказье, Казахстан, юг Сибири (до Алтая и Саян),
Монголия, Западный Китай. Нечасто отмечен в южных регионах Лесостепи Украины
(особенно на востоке); в сопредельной лесостепи России (Гусева, 1984). Встречается
по всей степной зоне Восточной Европы: от Украины и Молдавии (Nekuliseanu,
Matalin, 2000) до Среднего (Утробина, 1964) и Нижнего Поволжья (Калюжная и др.,
2000).
Степной вид. Предпочитает относительно сухие травянистые участки, норы грызу-
нов, обычно попадается под камнями, в подах балок, иногда на солончаках. Изредка
встречается и в пойменных биотопах юга Степи (изреженные древесно-кустарниковые
сообщества). Местами обычен на посевах зерновых, многолетних трав и в садах,
но только в степной зоне (Сумароков, 2009) и в Крыму.
Отмечен с начала мая до конца сентября, но обычно в июне-июле. Имаго активны
преимущественно ночью. Зимуют жуки и, по-видимому, личинки старших возрастов.
Мезоксерофил, стратоботробионт. Зарегистрированы случаи нападения на новоро-
жденных мышевидных грызунов (устное сообщение А. А. Петрусенко).
Подтриба S Y N U C H I N A Lindroth, 1956
В Палеарктике известно 4 рода (крупнейший - Synuchus) и более 100 видов.
Род SYNUCHUS Gyllenhal, 1810
В Палеарктике более 80 видов, встречающихся главным образом на Дальнем Восто-
ке. В Европе (в том числе Восточной) распространен S. (s. str.) vivalis vivalis (Illiger,
1798), встречающийся почти повсеместно, кроме сухих степей.
Подрод S y n u c h u s Gyllenhal, 1810
Synuchus vivalis vivalis (Illiger, 1798).
Полизональный лесо-кустарниковый мезофильный вид. В Украине тяготеет к ку-
старниковым насаждениям и рощам, обычен и в изреженных широколиственных лесах
(Полесье, Лесостепь). В Карпатах встречается в древесных формациях разных типов,
где его доля составляла 0.21-0.76 % всех жужелиц в хвойных (особенно пихтовых)
и 0.10-2.67 % - в широколиственных (чаще буковых) лесах (Різун, 2003а). Изредка
отмечен в садах и парках, единично также в отдельных загущенных лесополосах (Пуч-
ков, 2018). Зарегистрирован на лугах и полях от Соловецких островов (Болотов и др.,
2011) до Нижнего Поволжья (Калюжная, 2000) и от Белоруссии (Александрович, 2014)
до Среднего Поволжья (Алейникова, Утробина, 1969).
213
Имаго встречаются с начала апреля до конца октября. Молодые жуки отмечены с на-
чала мая, чаще со второй половины июля до начала августа (Різун, 2003а), что, возмож-
но, свидетельствует о зимовке преимущественно личинок. Характерен крыловой ди-
морфизм, длиннокрылые особи не летают (Lindroth, 1945).
ОБСУЖДЕНИЕ
Из 24 видов фауны Восточной Европы наибольшее число приходится на роды
Calathus (10) и Laemostenus (6). Pseudotaphoxenus представлен 3, а Dolichus, Platyderus,
Sphodrus, Synuchus и Taphoxenus - одним видом каждый (см. таблицу).
Шесть видов распространены почти на всей исследуемой территории, а некоторые
представители трибы характерны только для отдельных регионов Восточной Европы
(см. таблицу). Так, исключительно в Горном Крыму отмечены 3 вида (Laemostenus
cimmerius, L. venustus и эндемичный L. jailensis), по одному в Карпатах (Calathus
metallicus) и подзоне широколиственных лесов (L. tichyi - эндемик Западной Украины).
Только в степной зоне встречаются Calathus fuscipes punctipennis и Laemostenus
terricola punctatus, а также все виды рода Pseudotaphoxenus. На крайнем юге Восточ-
ной Европы чаще встречаются Calathus distinguendus, C. syriacus и Laemostenus sericeus
tauricus. Taphoxenus gigas характерен преимущественно для степной зоны, а Calathus
cinctus встречается на крайнем западе лесной зоны и в Закарпатье (хотя возможны на-
ходки на западном макросклоне Карпат, на Волыни и в южных регионах).
Некоторые различия наблюдаются и при сравнении видового состава (фаунистиче-
ского сходства) трибы в отдельных регионах (см. таблицу). При сравнении фаунисти-
ческого сходства (с использованием коэффициента Жаккара) выявлены существенные
различия, особенно в направлении с севера на юг. Отмечено низкое фаунистическое
сходство северной части Русской равнины с другими регионами (от 0.30 с южными
регионами и до 0.60 - для лесной зоны), что обусловлено крайней обедненностью
в этом регионе видового состава трибы (рис. 29). Фауны Sphodrini подзон смешанных
и широколиственных лесов, а также Карпат оказались более близкими (0.77-0.93)
(рис. 29). Наибольшее фаунистическое своеобразие свойственно степной зоне и Горно-
му Крыму, уровень сходства фаун которых с фаунами других регионов достигал 0.29-
0.47. Фауна Лесостепи занимала промежуточное положение, коэффициенты ее сход-
ства составляли от 0.50-0.71 (при сравнении с лесной зоной и Карпатами) до 0.42-0.47
(для Степи) (рис. 29).
Пять видов, отмеченные нами на юге Украины и в Горном Крыму (Calathus mollis,
Laemostenus cimmerius, L. terricola, L. venustus, Pseudotaphoxenus rufitarsis), и два вида
в Белоруссии (Laemostenus terricola и Sphodrus leucophthalmus) - в каталоге жужелиц
Палеарктики (Hovorka, 2017a; Casale, 2017) для этих территорий не указаны, хотя све-
дения о находках этих видов в отмеченных регионах (кроме P. rufitarsis, действительно
нового для фауны Украины) имеются в ряде работ, в том числе и XX в. (Якобсон, 1905;
Эйдельберг и др., 1988; Александрович, 1991; Різун, 2003а, 2003б; Коваль, 2009; Пуч-
ков, 2012).
Большинство видов рода Calathus, а также Dolichus halensis и Synuchus vivalis, были
обычными во многих ксеро- (на севере и в центре) и мезофитных (на юге и востоке)
травянистых биотопах (чаще открытых) и даже в древесно-кустарниковых стациях.
Представители других родов биотопически тяготели как к разнообразным древесным
(Laemostenus cimmerius, L. terricola, L. venustus, Platyderus rufus), так и к остепненным
214
участкам (Laemostenus sericeus, Taphoxenus gigas). К очень редким видам можно отне-
сти Laemostenus jailensis (яйлы Крыма), Pseudotaphoxenus rufitarsis (степные биотопы),
Laemostenus tichyi и Sphodrus leucophthalmus (отмечены в подвалах).
Все представители трибы - типичные полизоофаги, а представители подтриб
Dolichina и Sphodrina - облигатные хищники. Их имаго и личинки питаются мелкими
червями и их яйцевыми коконами, личинками и куколками насекомых, связанных
с почвой (пластинчатоусые, щелкуны и долгоносики из жесткокрылых, а также чешу-
екрылые и двукрылые), небольшими наземными моллюсками и пауками сем. Lycosidae
(Пучков, 2018). Другие виды трибы в отдельных случаях могут поедать и раститель-
ную пищу. Для некоторых видов рода Calathus отмечено питание недозревшими семе-
нами крестоцветных и злаков, одуванчика, плодовой падалицы и ягодами земляники,
а также мякотью грибов из родов Russula, Boletus и Luteus. В скоплениях некоторых
массовых видов рода Calathus наблюдался каннибализм (Петрусенко, 1973).
Биологические особенности видов трибы исследованы с разной полнотой, а для не-
которых - неизвестны вообще. Лучше изучена биономия большинства политопных
видов - Calathus ambiguus, C. erratus, C. fuscipes, C. melanocephalus, C. micropterus,
C. cinctus, Dolichus halensis (Gilbert, 1956; Kůrka, 1972; van Dijk, 1973, 1994, 1996; Ве-
рещагина, 1984; Gergely, Lovei, 1987; Aukema, 1995; Sharova, Denisova, 1995; Різун,
2003а; Филиппов, 2006; Александрович, 2014; Пучков, 2018). Слабее изученными,
по отдельным из вышеприведенных работ (Різун, 2003а; Александрович, 2014; Пуч-
ков, 2018), остаются более редкие и стенотопные представители трибы (Calathus
metallicus, Laemostenus terricola, Synuchus vivalis). Сведения об эколого-биологических
особенностях других видов Sphodrini, известных по единичным экземплярам, отсут-
ствуют.
Сезонная активность, период размножения и особенности развития большинства ви-
дов трибы обычно поливариантны и зависят от наличия кормовой базы, погодных
и микроклиматических условий, характера фотопериодической реакции (Маталин,
2007).
БЛАГОДАРНОСТИ
Авторы выражают глубокую благодарность за помощь при создании этой работы
(предоставление коллекционного материала, обсуждение ряда вопросов) Б. М. Катае-
ву, И. И. Кабаку (Санкт-Петербург, Россия), В. Б. Ризуну (Львов, Украина), В. В. Мар-
тынову (Донецк, Украина) и А. В. Мартынову (Киев, Украина).
СПИСОК ЛИТЕРАТУРЫ
Абдурахманов Г. М., Нахибашева Г. М., Клычева С. М., Эльдерханова З. М. 2011. Сравнительный анализ
видовых составов жужелиц прибрежных и островных экосистем западного Каспия. Юг России: эко-
логия, развитие 4: 51-71.
Алейникова М. М., Утробина Н. М. 1969. Животное население почв в агроценозах Среднего Поволжья.
В кн.: Алейникова М. М. (ред.). Животное население почв агроценозов и его изменение под влиянием
сельскохозяйственного производства. Казань: КГУ: 3-62.
Александрович О.Р. 1991. Жуки жужелицы (Coleoptera, Carabidae) фауны Белоруссии. В кн: И. К. Лопатин,
Э. И. Хотько (ред.). Фауна и экология жесткокрылых Белоруссии. Минск: Навука i тэхнiка: 37-78.
Александрович О. Р. 2014. Жужелицы (Coleoptera, Carabidae) запада лесной зоны Русской Равнины (фауна,
зоогеография, экология, фауногенез). Saarbrücken: Lambert Academic Publishing, 456 с.
215
Александрович O. Р., Козулько Н. Г. 2017. Семейство Carabidae Latreille, 1802. В кн.: B. A. Цинкевич (ред.).
Каталог насекомых (Insecta) Национального парка «Беловежская пуща». Минск: Белорусский Дом
печати: 53-69.
Болотов И. Н., Зубрий Н. А., Цыварева Е. П., Христофорова Н. С. 2011. Видовой состав жужелиц (Coleoptera,
Carabidae) Соловецких островов. Вестник Северного (Арктического) федерального университета.
Серия: Естественные науки 2: 45-52.
Верещагина Т. Н. 1984. Жужелицы рода Calathus Bon. (Coleoptera, Carabidae) фауны СССР. Энтомологиче-
ское обозрение 63 (4): 751-767.
Гусева Н. А. 1984. Жужелицы (Carabidae, Coleoptera) Центрально-Черноземного заповедника. В кн.:
Д. А. Криволуцкий (ред.). Эколого-фаунистические исследования центральной лесостепи Европей-
ской части СССР. Сборник научных трудов ЦНИЛ Главохоты РСФСР. М.: 12-17.
Гусева О.Г., Коваль А.Г. 2008. Особенности комплексов жужелиц (Coleoptera, Carabidae) в агроценозах Ле-
нинградской области с различными почвенными условиями. Вестник защиты растений 4: 3-11.
Ильина Е. В., Алиев М. А. 2015. Географическое распространение жужелиц рода Calathus Bonelli, 1810
(Coleoptera: Carabidae) в Республике Дагестан. Вестник Дагестанского Научного Центра. Биологиче-
ские науки 58: 5-11.
Калюжная Н. С., Комаров Е. В., Черезова Л. Б. 2000. Жесткокрылые насекомые (Insecta, Coleoptera) Нижнего
Поволжья. Волгоград. 204 с.
Кириченко М. Б., Бабко Р. В. 2007. Підсумки досліджень фауни жуків-скакунів і турунів (Coleoptera,
Cicindelidae, Carabidae) Полісся та Лісостепу Лівобережної України. Известия Харьковского энтомо-
логического общества 14 (1-2): 28-36.
Клаузницер Б. 1990. Экология городской фауны. М.: Мир, 248 с.
Коваль А. Г. 2009. Жужелицы (Coleoptera, Carabidae) агроценоза картофеля европейской части России и со-
предельных территорий. Чтения памяти Н. А. Холодковского. СПб 61 (2), 111 с.
Крыжановский О. Л. 1965. Семейство Carabidae - Жужелицы. В кн.: Г. Я. Бей-Биенко (ред.). Определитель
насекомых европейской части СССР в 6 томах. Т. 2. Жесткокрылые и веерокрылые. М.; Л.: Наука:
29-77.
Крыжановский О. Л. 1983. Жуки подотряда Adephaga: семейства Rhysodidae, Trachypachidae; семейство
Carabidae (вводная часть и обзор фауны СССР). Л.: Наука, 342 с. (Фауна СССР. Нов. сер., № 128.
Жесткокрылые; т. 1, вып. 2).
Маринич О. М., Ланько А. І., Щербань М. І., Тищенко П. Г. 1982. Фізична географія Української РСР. Київ:
Вища школа, 208 с.
Маталин А. В. 2007. Типология жизненных циклов жужелиц (Coleoptera, Carabidae) Западной Палеарктики.
Зоологический журнал 86 (10): 1196-1220.
Петрусенко А. А. 1973. Жужелицы - Carabidae . В кн.: В. П. Васильев (ред.). Вредители сельскохозяйствен-
ных культур и лесных насаждений в 3 томах. Т. 1. К.: Урожай: 363-387.
Петрусенко О. А., Петрусенко С. В. 1971. До вивчення фауни жужелиць (Coleoptera, Carabidae) лучних сте-
пів України. Збірник праць Зоологічного музею АН УРСР 34: 60-64.
Петрусенко А. А., Никитенко Г. Н., Пучков А. В. 1999. Жужелицы (Coleoptera, Carabidae) плодово-ягодных
культур Украины. Вестник зоологии. Отдельный выпуск 10: 61-91.
Пономарчук В. И. 1963. О вертикальном распределении жужелиц (Coleoptera, Carabidae) Закарпатской обла-
сти. Зоологический журнал 42 (10): 1485-1493.
Пучков А. В. 1990. Жесткокрылые (Coleoptera) пшеничного поля юго-запада степной зоны европейской ча-
сти СССР. Энтомологическое обозрение 3: 538-549.
Пучков А. В. 2001. Новые находки жужелицы Laemostenus tauricus sericeus (Coleoptera, Carabidae) в Украи-
не. Вестник зоологии 35 (1): 94.
Пучков А. В. 2012. Фаунистический обзор карабоидных жуков (Coleoptera, Сaraboidea) Украины. Українсь-
кий ентомологічний журнал 2 (5): 3-44.
Пучков А. В. 2018. Жуки-жужелицы (Coleoptera, Carabidae) трансформированных ценозов Украины. Киев:
Институт зоологи им. И.И. Шмальгаузена НАН Украины, 448 с.
Різун В. Б. 2003а. Туруни Українських Карпат. Львів: Державний природознавчий музей НАН України,
210 с.
Ризун В. Б. 2003б. Жесткокрылые Западного Подолья. В кн.: Чернобай Ю. Н. (ред.). Экология и фауна поч-
венных беспозвоночных Западного Волыно-Подолья. Киев: Наукова думка: 173-270.
216
Різун В. Б. 2009. До фауни жуків-турунів (Coleoptera, Carabidae) Українського Полісся. В кн.: М. Д. Будз
(ред.). Збереження та відтворення біорізноманіття заповідних територій: матеріали міжнародної на-
уково-практичної конференції, присвячені 10-річчю Рівненського природного заповідника. Рівне:
535-543.
Сигида С. И. 1993. Ландшафтно-биотопическое распределение и экологическая характеристика жужелиц
(Coleoptera, Carabidae) Предкавказья и северных склонов центрального Кавказа. Энтомологическое
обозрение 72 (1): 11-38.
Сумароков А. М. 2009. Восстановление биотического потенциала биогеоценозов при уменьшении пести-
цидных нагрузок. Донецк: «Вебер», 194 с.
Утробина Н. И. 1964. Обзор жужелиц Среднего Поволжья. В кн.: Алейникова М. М. (ред.). Почвенная фауна
Среднего Поволжья. Казань: Наука: 93-119.
Федоренко Д. М. 1988. Фауна жужелиц (Coleoptera, Carabidae) Московской области. Насекомые Московской
области. Проблемы кадастра и охраны. М.: Наука: 20-46.
Феоктистов В. Ф., Душенков В. М. 1982. Сезонная динамика активности жужелиц (Coleoptera, Carabidae)
в различных типах леса у южной границы тайги. Зоологический журнал 61 (2): 227-232.
Филиппов Б. Ю.
2006. Сезонные аспекты жизненных циклов жужелиц Calathus melanocephalus
и С. micropterus (Coleoptera, Carabidae) в северной тайге. Зоологический журнал
85
(10):
1196-1204.
Физико-географическое районирование СССР. Характеристика региональных единиц. 1968. Под ред.
Н. А. Гвоздецкого. М.: Издательство Московского университета, 578 с.
Хоменко В. Н., Вакаренко Е. Г. 1993. Карабидофауна (Coleoptera, Carabidae) заповедника Аскания-Нова:
структура и тенденции изменения. Вестник зоологии 5: 26-35.
Шарова И. X., Соболева-Докучаева И. И. 1984. Эколого-фаунистическая характеристика полевых жужелиц
(Coleoptera, Carabidae) в зоне смешанных лесов Московской области. В кн.: Н. М. Чернова (ред.).
Фауна и экология беспозвоночных животных. Межвузовский сборник научных трудов. М.: МГПИ
им. В. И. Ленина: 117-124.
Эйдельберг М. М. 1989. Жужелицы (Coleoptera, Carabidae) плодовых садов Крыма (фауна, экология, биоло-
гия). Автореферат диссертации на соискание ученой степени кандидата биологических наук. Киев;
Ялта, 19 с.
Эйдельберг М. М., Мальцев И. В., Перваков В. П. 1988. Видовой состав жужелиц (Coleoptera, Carabidae)
Крыма. В кн.: Г. Л. Топчий (ред.). Экология и таксономия насекомых Украины. Киев: Наукова думка:
61-68.
Якобсон Г. Г. Триба Platynina (Anchomenina). 1905. В кн.: Жуки Россiи и Западной Европы. СПб.: Издатель-
ство Девриена: 317-333.
Aleksandrowicz O., Stachowiak M., Putchkov A. 2016. Additions, corrections and comments to the Carabidae part
of: I. Löbl & A. Smetana 2003. Catalogue of Palaearctic Coleoptera. Vol. 1, Archostemata-Myxophaga-
Adephaga for Belarus, Ukraine and Poland. Słupskie Prace Biologiczne 13: 5-18.
Assmann T. 2004. Sphodrini. In.: H. Freude, K.-W. Harde, G. A. Lorenz (еds). Die Käfer Mitteleuropas. Band 2.
Adephaga 1. Carabidae. Munchen: Akademische Verlag, p. 284-298.
Aukema B. 1995. Flying for Life: Wing Dimorphism in Closely Related Species of the Genus Calathus (Coleoptera:
Carabidae). Wageningen: Ponsen & Looijen, 168 p.
Bacal S., Munteanu N., Toderaș I. 2013. Checklist of Beetles (Insecta: Coleoptera) of the Republic of Moldova.
Brukenthal. Acta Musei 8 (3): 415-450.
Bercio H., Folwaczny B. 1979. Verzeichnis der Käfer Preußens. Gedruckt im Rahmen der Veröffentlichungen des
Vereins für Naturkunde in Osthessen. Fulda: Verlag Parzeller & Co., p. 288-315.
Burakowski B., Mroczkowski M., Stefańska J. 1974. Chrząszcze - Coleoptera. Biegaczowate - Carabidae 2.
Katalog Fauny Polski. Cz. 23, t. 3. Warszawa: PWN, 430 p.
Casale A. 1988. Revisione degli Sphodrina (Coleoptera, Carabidae, Sphodrini). Monographie V. Museo Regionale
di Scienze Naturali Torino, 1024 p.
Casale A. 2003. Subtribe Synuchina. In: I. Löbl, A. Smetana (eds). Catalogue of Palaearctic Coleoptera. Vol. 1.
Archostemata-Myxophaga-Adephaga. Stenstrup: Apollo Books, p. 548-577.
Casale A. 2017. Subtribe Sphodrina. In: J. Löbl, D. Löbl (eds). Catalogue of Palaearctic Coleoptera. Vol. 1.
Archostemata-Myxophaga-Adephaga. Leiden; Boston: Brill, p. 773-790.
Catalogue of Palaearctic Coleoptera. 2017. Vol. 1. Archostemata-Myxophaga-Adephaga. J. Löbl, D. Löbl. (eds).
Leiden; Boston: Brill, 1444 p.
217
Dijk Th. S. van. 1973. The age-composition of populations of Calathus melanocephalus L. Analysed by studying
marked individuals kept within fenced sites. Oecologia (Berlin) 12: 213-240.
Dijk Th. S. van. 1978. Calathus mollis erythroderus Gemminger & Harold in Drenthe (Coleoptera, Carabidae).
Enthomologische Berichten 38 (10): 151-153.
Dijk Th. S. van. 1994. On the relationship between food, reproduction and survival of two carabid beetles: Calathus
melanocephalus and Pterostichus versicolor. Ecological Entomology 19 (3): 263-270
Dijk Th. S. van. 1996. The influence of environmental factors and food on life cycle, ageing and survival of some
carabid beetles [melanocephalus]. In: Booij K., Nijs L.-den (eds.). Acta Jutlandica, Aarhus (Denmark).
Natural Science Series. Proceedings of the second EU workshop: Arthropod natural enemies in arable land,
2: Survival, reproduction and enhancement. Aarhus (Denmark). Acta Jutlandica 71: 11-24
Ferenca R., Tamutis V., Inokaitis V., Martinaitis K. 2016. Data on beetle (Coleoptera) species new to Lithuanian
fauna. Naujos ir retos Lietuvos vabzdžių rūšys 28: 21-31.
Gergely G., Lovei G. L. 1987. Phenology and reproduction of the ground beetle Dolichus halensis in maize fields:
a preliminary report. Acta Phytopathologica et Entomologica Hungarica 22 (1-4): 357-361.
Gilbert O. 1956. The natural histories of four species of Calathus (Coleoptera, Carabidae) living on sand dunes in
Anglesey, North Wales. Oikos 7 (1): 22-47.
Hammer Ø., Harper D. A. T., Ryan P. D. 2001. PAST: Paleontological Statistics software package for education and
data analysis. Palaeontologia Electronica 4 (1): 9 p.
Honek A., Martinkova Z., Saska P., Pekar S. 2007. Size and taxonomic constraints determine the seed preferences
of Carabidae (Coleoptera). Basic and Applied Ecology 8 (4): 343-353.
Hovorka O. 2017a. Subtribes Atranopsina, Calathina, Dolichina. In: I. Löbl, D. Löbl (eds). Catalogue of Palearctic
Coleoptera. Vol. 1. Archostemata-Myxophaga-Adephaga. Leiden; Boston: Brill, p. 755-772.
Hovorka O. 2017b. Subtribe Synuchina. In: I. Löbl, D. Löbl (eds). Catalogue of Palearctic Coleoptera. Vol. 1.
Archostemata-Myxophaga-Adephaga. Leiden; Boston: Brill, p. 790-794.
Kryzhanovskij O. L., Belousov I. A., Kabak I. I., Kataev B. M., Makarov K. V., Shilenkov V. G. 1995. A Checklist of
the Ground-Beetles of Russia and Adjacent Lands (Insecta, Coleoptera, Carabidae). Sofia; Moscow: Pensoft
Publishers, 271 p.
Kůrka A. 1972. Bionomy of the Czechoslovak species of the genus Calathus Bon. Coleoptera, Carabidae), with
notes on their rearing. Vestnik Československe Společnosti Zoologicke 36 (2): 101-114.
Lindroth C.H. 1945. Die fennoskandischen Carabidae. Eine tiergeographisce Studie. 1. Spezieller Teil. Goteborgs
Kungliga Vetenskaps och Vitter Hets-Samhalles Handlingar Sjatte Foljden. Ser. B 4 (1): 1-709.
Nekuliseanu Z. Z., Matalin A. V. 2000. A Сatalogue of the Ground-Beetles of the Republic of Moldova (Insecta,
Coleoptera: Carabidae). Sofia; Moscow: Pensoft, 164 p.
Sharova I. Kh., Denisova M. I. 1995. Life cycles and reproduction rhythms of Calathus erratus C. R. Sahlberg and
C. micropterus Duftschmid (Coleoptera, Carabidae) in the forests of Russian forest-steppe zone. Russian
Entomological Journal 4 (1-4): 159-165.
Šustek Z. 1999. Light attraction of carabid beetles and their survival in the city centre. Biologia, Bratislava 54 (5):
539-551.
Tamutis V., Ziogas A., Saluchaite A., Kazlauskaite S., Amsiejus A. 2007. Epigeic beetle (Coleoptera) communities
in summer barley agrocenoses. Baltic Journal of Coleopterology 7 (1): 83-98.
Telnov D., Bukejs A., Gailis J., Kalniņš M. 2008. Contributions to the knowledge of Latvian Coleoptera. 7. Latvijas
Entomologs 46: 47-58.
GROUND-BEETLES OF THE TRIBE SPHODRINI (COLEOPTERA, CARABIDAE)
OF EAST EUROPE
A. V. Putchkov, O. R. Aleksandrowicz
Key words: Carabidae, Sphodrini, East Europe, species, distribution, ecology, key.
218
SUMMARY
In East Europe 24 species of the 8 Sphodrini genera are recorded: Calathus (10), Laemostenus (6),
Pseudotaphoxenus (3), Dolichus, Sphodrus, Platyderus, Synuchus and Taphoxenus (one species in
each). Five species which occur in Ukraine (Calathus cinctus, Laemostenus cimmerius, L. terricola,
L. venustus, Pseudotaphoxenus rufitarsis) and two in Belarus (Laemostenus terricola, Sphodrus
leucophthalmus) were not recorded for these countries in the Catalogue of Palaearctic Coleoptera
(2017). Six species are distributed almost throughout East Europe, while some species occur in separate
regions only. Three species (Laemostenus cimmerius, L. venustus and L. jailensis) are known only in
the Crimean Mountains. One species (Calathus metallicus) occurs only in the Carpathians, and one
(Laemostenus tichyi, as endemic to Western Ukraine) is known only in the subzone of broadleaf forests.
All species of the genus Pseudotaphoxenus, Calathus fuscipes punctipennis and C. terricola punctatus
are found only in the steppe zone. Calathus distinguendus, C. mollis, C. syriacus and Laemostenus
sericeus occur in the very extreme south of East Europe and Crimea. Taphoxenus gigas is distributed
mainly in the steppe zone, and Calathus cinctus, only in the western part of the forest zone and in
the Carpathians. Most species of the genera Calathus, Dolichus and Synuchus are common in open
mesophytic biotopes. Species of other genera are met sporadically and not often in the woods, bushes
(Laemostenus cimmerius, L. terricola, L. venustus and Platyderus rufus) and steppe biotops (Laemostenus
sericeus, Taphoxenus gigas). Laemostenus jailensis (Crimean mountains), Pseudotaphoxenus rufitarsis
(southern steppe), L. tichyi, Sphodrus leucophthalmus (found in basements) and Calathus cinctus
(western part of the forest zone) are very rare in East Europe. The species essays and keys to 24 species
(27 species-group taxa including subspecies) of 8 genera are given.
219