ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 99, 2, 2020
УДК 632.936.2
СРАВНИТЕЛЬНАЯ ОЦЕНКА РЕАКЦИЙ ЗАПАДНОГО
ЦВЕТОЧНОГО ТРИПСА FRANKLINIELLA OCCIDENTALIS PERG.
(THYSANOPTERA, THRIPIDAE) И ХИЩНОГО КЛОПА
ORIUS LAEVIGATUS FIEBER (HETEROPTERA, MIRIDAE)
НА МАСЛО PONGAMIA PINNATA (L.) PIERRE
© 2020 г. Е. А. Степанычева, 1* И. М. Пазюк, 1** Т. Д. Черменская, 1***
М. О. Петрова, 1**** Р. Павела 2*****
1 Всероссийский научно-исследовательский институт защиты растений
шоссе Подбельского, 3, С.-Петербург-Пушкин, 196608 Россия
*e-mail: stepanycheva@yandex.ru, **e-mail: ipazyuk@gmail.com, *** e-mail: tchermenskaya@
yandex.ru, ****e-mail: mar34915696@yandex.ru
2 Crop Research Institute (CRI)
Drnovska 507, Praha 6 - Ruzyně, 161 06 Czech Republic
*****e-mail: pavela@vurv.cz
Поступила в редакцию 21.01.2020 г.
После доработки 3.04.2020 г.
Принята к публикации 3.04.2020 г.
В статье представлены результаты сравнительной оценки реакции западного цветочного
трипса Frankliniella occidentalis и хищного клопа Orius laevigatus на масло из семян Pongamia
pinnata L. Применение масла P. pinnata оказывало репеллентное и токсическое действие на
имаго и личинок трипса, а также снижало количество отложенных яиц. Orius laevigatus был
высоко чувствителен к маслу P. pinnata (даже в минимальной эффективной для трипса концен-
трации 0.75 %) при разных способах воздействия. Выявленное нами побочное действие масла на
ориуса показывает необходимость разработки тактики совместного применения масла и данного
энтомофага или замены ориуса другим видом хищника, не восприимчивым к маслу.
Ключевые слова: Pongamia pinnata, масло, семена, Frankliniella occidentalis, Orius laevigatus,
фитофаг, энтомофаг, поведение, размножение, токсичность.
DOI: 10.31857/S0367144520020057
Западный цветочный трипс, Frankliniella occidentalis Pergande, - широкий полифаг,
наносящий как прямой (при питании), так и косвенный (в качестве переносчика вирус-
ной инфекции) вред культивируемым растениям (Schneweis et al., 2017). Обладая высо-
кой репродуктивной способностью и коротким периодом развития генерации, этот
фитофаг в условиях закрытого грунта способен развиваться непрерывно, давая от
10 до 12 поколений в год. Стремление снизить численность вредителя дополнительны-
ми обработками инсектицидами оборачивается быстрым развитием у него резистент-
ности (Gao et al., 2012). Это стимулирует поиск новых веществ, способных снижать
численность фитофага и при этом сохранять благоприятную экологическую обста-
новку.
298
Одно из активно развивающихся направлений в защите растений связано с поиском
биологически активных веществ растительного происхождения. За последнее десяти-
летие опубликовано несколько обзорных статей, посвященных обсуждению различ-
ных механизмов воздействия эфирных масел на вредных членистоногих, что
подтверждает перспективность таких исследований (Regnault-Roger et al., 2012; Mossa,
2016; Pavela, 2016; Ikbal, Pavela, 2019). Есть достаточно свидетельств относительной
безопасности масел для многих энтомофагов (Ribeiro et al., 2016; Born et al., 2018;
Castilhos et al., 2018; Papadimitriou et al., 2019; Soares et al., 2019), что не исключает
необходимость такой оценки для вновь создаваемых препаратов.
Высокая эффективность вторичных метаболитов широкого круга высших растений
как средства контроля различных вредных членистоногих показана неоднократно
(Pino et al., 2013). Значительный интерес представляет бобовое Pongamia pinnata L.
(= Millettia pinnata L., Derris indica (Lamk.) Bennet из монотипического рода (Kumar,
Kalindhar, 2003). Pongamia pinnata - богатый источник флавоноидов, характеризую-
щихся высокой биологической активностью. Масло P. pinnata обладает антифидант-
ной активностью, оказывает репеллентное действие на членистоногих и вызывает
у них нарушение яйцекладки (Negi et al., 1997; Kumar, Kalindhar, 2003).
Цель нашей работы - изучить инсектицидные и модулирующие поведение свойства
масла растения Pongamia pinnata L. в отношении западного цветочного трипса и его
хищника - клопа Orius laevigatus Fieber.
МАТЕРИАЛ И МЕТОДИКА
Лабораторную популяцию клопа O. laevigatus (около 12 лет без обновления) разводили
в пластиковых контейнерах (объем 500 мл) на яйцах зерновой моли Sitotroga cerealella Oliv. и
обыкновенной злаковой тле Schizaphis graminum Rondani при температуре (23 ± 3) °C и продол-
жительности светового периода 16 ч. Субстратом для откладки яиц служили растения фасоли
Phaseolus vulgaris L.
Культуру западного цветочного трипса F. occidentalis содержали на растениях фасоли в поме-
щении с температурой 23 ± 2 °C и продолжительностью светового дня 16 ч.
Изучаемый образец - эмульгированное модифицированное масло из семян P. pinnata (со-
держание масла 86.8 %) было получено из Исследовательского института растениеводства
в Чешской Республике (Crop Research Institute, Czech Republic). Для испытаний готовили 3.0, 1.5,
0.75 и 0.187%-ные концентрации.
Влияние масла на выбор имаго трипса и клопа растений для питания
и откладки яиц
Растения фасоли, выращенные индивидуально в пластиковых стаканчиках (200 мл),
обрабатывали растворами масла разной концентрации, контрольные - водой.
В опыте с трипсом обработанные и контрольные растения размещали в боксы с сам-
ками для свободного заселения. Через сутки учитывали количество имаго на каждом
растении, после чего насекомых удаляли. В связи с тем, что самки откладывают яйца
в паренхиму листа, потенциал размножения оценивали по количеству отродившихся
личинок дочернего поколения. Опыт повторяли 10 раз.
В опыте с клопом по 2 контрольных и 2 опытных растения помещали в садки
40 × 40 × 60 см, обтянутые мельничным газом. В качестве корма для энтомофага
на листья в одинаковом количестве добавляли яйца зерновой моли S. cerealella, после
чего в садки выпускали по 40 взрослых особей клопов без разделения по полу.
299
Через двое суток учитывали распределение имаго клопа на растениях и подсчитыва-
ли число отложенных яиц. Повторностью служило одно растение. Всего использовали
5 садков.
Для определения продолжительности действия масла на клопа использовали такую
же методику, но ориусов выпускали на растения через 3, 5 и 7 дней после обработки.
Влияние масла на поведение обоих видов оценивали по индексу предпочтения (ИП),
рассчитываемому по следующей формуле (Pascual-Villalobos, Robledo, 1998):
ИП = [(Число особей на обработанном растении - число особей на контрольном
растении) / (число особей на контрольном растении + число особей на обработанном
растении) × 100
При положительном значении показателя ИП регистрируется аттрактантное дей-
ствие, при отрицательном - репеллентное.
Снижение численности потомства (личинок дочернего поколения) в опыте по срав-
нению с контролем (СЧП) рассчитывали по следующей формуле (Abbott, 1925):
СЧП, % = [(число личинок в контроле - число личинок в опыте) / число личинок
в контроле] × 100
В опыте с хищным клопом в этой формуле использовали количество отложенных
яиц.
Влияние масла на самок трипса Frankliniella occidentalis, хищного клопа
и их репродуктивную активность
Обработанные путем погружения на 2-3 сек в растворы препарата листья фасоли
размещали в чашки Петри на фильтровальную бумагу. На каждый лист выпускали по
10 самок трипса. Чашки закрывали пленкой с отверстиями для вентиляции. Через 2 су-
ток самок удаляли, подсчитывая количество живых и погибших особей. Через 5-7 дней
проводили учет отродившихся личинок. В эксперименте было 10 повторностей.
На дно пластиковых контейнеров объемом 212 мл помещали фильтровальную бума-
гу, яйца зерновой моли Sitotroga cerealella (для питания) и стебли фасоли (для отклад-
ки яиц). Раствором масла опрыскивали внутреннюю поверхность садка и корм. Расход
рабочей жидкости на 1 контейнер составлял 1 мл. После испарения излишней влаги в
контейнеры выпускали по 5 взрослых особей клопов и закрывали контейнеры крышка-
ми с вентиляционным отверстием, затянутым мельничным газом. Через сутки учиты-
вали живых и погибших особей, их пол, а также количество отложенных яиц. Оба
варианта опыта (включая контроль) повторяли десять раз.
Снижение численности потомства (%) рассчитывали по следующей формуле (Abbott,
1925):
[(к - о)/к] × 100,
[(численность потомства в контроле
- численность потомства в опыте)/
численность потомства в контроле] × 100
где к - численность потомства в контроле, о - численность потомства в опыте.
Снижение плодовитости самок (%) определяли по количеству личинок дочернего
поколения:
[(число личинок на самку в контроле - число личинок на самку в опыте) / число
личинок на самку в контроле] × 100
300
Ларвицидная активность масла
В опыте с трипсом для получения выравненных по возрасту личинок на растения
фасоли выпускали самок вредителя, которых затем удаляли через сутки, а растения
помещали в изолированный бокс для предотвращения повторного заселения. Через
5-7 суток отродившихся личинок использовали в экспериментах.
Листья фасоли погружали в растворы препарата на 2-3 сек. После испарения влаги
листья помещали в чашки Петри на влажную фильтровальную бумагу и выпускали на
них по 15-20 личинок. Чашки закрывали пленкой с отверстиями для вентиляции. Учет
выживших и погибших особей проводили через 1 и 3 дня. Опыты проводили в 10 по-
вторностях.
В опыте с клопом использовали личинок 5-го возраста. В пластиковые контейнеры
(V = 212 мл) помещали на дно фильтровальную бумагу, яйца зерновой моли S. cerealella
(для питания) и стебли фасоли (для поддержания влажности). Внутреннюю поверх-
ность контейнера и корм обрабатывали 0.75%-ным раствором масла P. pinnata. Расход
рабочей жидкости на 1 контейнер - 1 мл. После испарения излишней влаги в контейне-
ры выпускали по 5 личинок клопа и закрывали их крышками с вентиляционным отвер-
стием, затянутым мельничным газом. Через сутки учитывали живых и погибших
особей. В опыте и контроле было по 10 повторностей. Активность препарата с учетом
контроля (смертность с учетом контроля, %) рассчитывали по следующей формуле
(Abbott, 1925):
[(гибель в опыте - гибель в контроле) / (100 - гибель в контроле)] × 100
РЕЗУЛЬТАТЫ
Влияние масла на выбор имаго трипса и клопа растений для питания
и откладки яиц
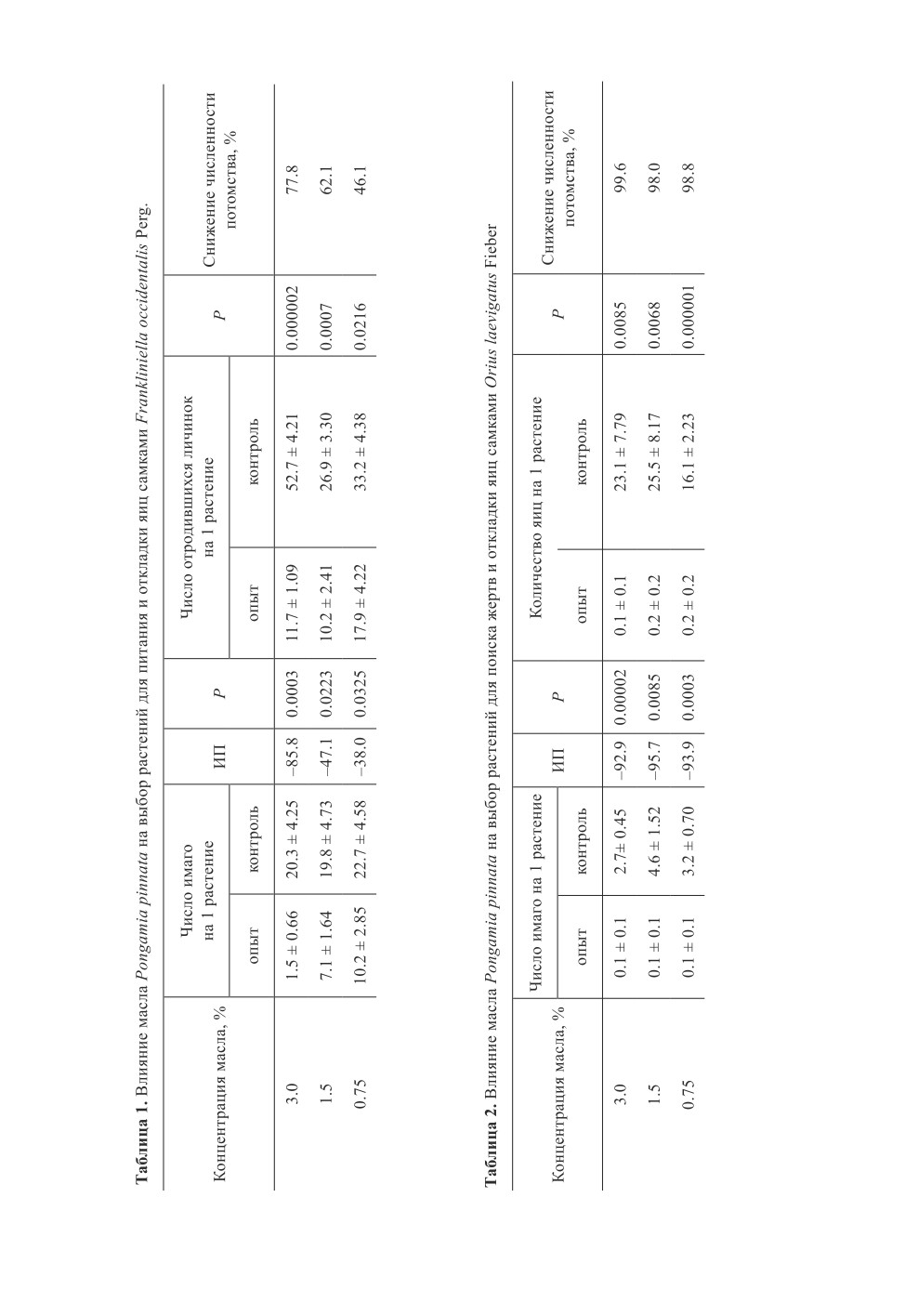
При превентивной (до заселения фитофагом) обработке растений в вариантах с при-
менением 3.0 и 1.5%-ного растворов самки предпочитали контрольные растения (ИП
составлял -85.8 и -47.1 соответственно). В дальнейшем этот выбор сказался на чис-
ленности потомства. Снижение числа личинок дочернего поколения на растениях при
обработке 3%-ным раствором составило 77.8 % по сравнению с контролем, а при ис-
пользовании 1.5%-ного раствора препарата - 62.1 % (табл. 1).
В аналогичном эксперименте с использованием O. laevigatus обработка растений
маслом P. pinnata резко снижала их привлекательность для клопа. На опытных расте-
ниях находились единичные взрослые особи и яйца. Все три концентрации масла по-
казали практически одинаковый результат. Наблюдения за продолжительностью
сохранения такого эффекта (изменение периода ожидания от обработки выпуска кло-
пов) продемонстрировали негативное влияние масла даже через 7 суток (табл. 2).
Влияние масла на самок трипса F. occidentalis, хищного клопа
и их репродуктивную активность
Принудительное содержание самок трипса на листьях фасоли, обработанных раство-
рами 3.0 и 1.5%-ной концентрациями, привело к гибели более 90 % взрослых особей
фитофага и к снижению числа отродившихся личинок также более чем на 90 %. Следу-
ет отметить, что в варианте с 3.0%-ной концентрацией все отродившиеся личинки поч-
ти сразу погибли (табл. 3). Из испытанных концентраций 0.75%-ная оказалась
минимальной, обеспечивающей эффективность более 50 %.
301
Таблица 3. Влияние масла Pongamia pinnata на численность самок Frankliniella occidentalis
Perg. и их потомства
Число
Снижение
Концентрация
Исходное
Число погибших
Смертность
отродившихся
численности
масла,
число
самок
самок с учетом
личинок
потомства
%
самок
через 2 суток, %
контроля, %
на 1 самку
на 1 самку, %
3.0
98
96.1 ± 2.19
95.8
0.03 ± 0.02
98.5
1.5
97
97.1 ± 0.02
96.9
0.07 ± 0.03
96.5
0.75
100
62.2 ± 3.81
59.6
0.30 ± 0.04
85.0
0.375
102
25.3 ± 3.16
20.1
1.40 ± 0.12
30.0
Контроль
197
6.5 ± 1.82
-
2.00 ± 0.11
-
Таблица 4. Ларвицидная активность масла Pongamia pinnata для трипса Frankliniella
occidentalis Perg.
Время до гибели личинок, сут
Смертность
Концентрация
Суммарная
по отношению
масла, %
гибель, %
1
3
к контролю, %
3.0
96.3 ± 1.35
3.7 ± 1.35
100 ± 0.0
100.0
1.5
89.1 ± 2.69
5.0 ± 1.80
94.1 ± 1.89
92.3
0.75
74.1 ± 1.26
7.0 ± 1.99
81.1 ± 2.81
75.3
0.375
33.5 ± 2.10
13.7 ± 1.30
47.2 ± 2.35
31.1
0.187
29.8 ± 2.75
9.0 ± 1.56
38.8 ± 2.49
20.1
Контроль
14.3 ± 1.26
9.1 ± 1.14
23.4 ± 1.53
-
В опыте с хищным клопом полученные результаты продемонстрировали высокую
токсичность 0.75%-ного раствора масла для имаго энтомофага в условиях закрытого
пространства (пластиковый садок-контейнер). Через сутки в опыте погибли все про-
тестированные особи. Количество отложенных яиц составило 0.05 ± 0.03 из расчета
на 1 самку. В контроле гибель взрослых особей составила 7.3 %, а количество яиц -
2.4 на самку.
Ларвицидная активность масла
На стадии личиночного развития трипс оказался достаточно чувствительным к маслу
P. pinnata. Образец 0.75%-ной концентрации вызывал гибель 75.4 % тестируемых
особей. Разведение рабочего раствора в 2 раза привело к резкому снижению ларвицид-
ной активности до 31.5 % (табл. 4).
Содержание личинок 5-го возраста O. laevigatus на обработанных 0.75%-ным
раствором масла корме и поверхности вызвало их 100%-ную гибель через сутки.
За этот же период в контроле погибло всего 4 % особей.
ОБСУЖДЕНИЕ
Полученные нами результаты показывают перспективность применения масла
Pongamia pinnata для снижения численности западного цветочного трипса за счет ре-
303
пеллентного и токсического действия на имаго и личинок, а также уменьшения коли-
чества откладываемых трипсом яиц. Наши результаты о различных формах воздействия
P. pinnata на F. оccidentalis согласуются с данными, полученными ранее для этого и
некоторых других вредителей. Например, содержание личинок F. occidentalis на ли-
стьях, обработанных 0.2%-ным раствором масла понгамии, привело к гибели более
50 % особей через 5 суток (Uçak et al., 2014). Токсическое действие P. pinnata установ-
лено также на совку Spodoptera litura (Tran et al., 2017), тлей Melanaphis sacchari
(Balikai, Lingappa, 2005) и Lipaphis pseudobrassicae (Tran et al., 2016), вредителей запа-
сов жуков Trogoderma granarium и Tribolium castaneum (Kumar et al., 2006; Mondal,
Khalequzzaman, 2006; Tayoub et al., 2012). Снижение плодовитости на 97.6 % при дей-
ствии экстракта P. pinnata показано на примере комара Aedes aegypti (Swathi et al.,
2010) и тли Lipaphis pseudobrassicae (Tran et al., 2016). Выявлено репеллентное дей-
ствие масла на белокрылку Trialeurodes vaporariorum (Pavela, Herda, 2007), комаров
(Lale, Kulkarni, 2010) и жука Oryzaephilus surinamensis (Shah et al., 2008). Проявление
антифидантной активности P. pinnata установлено на гусеницах хлопковой совки
Helicoverpa armigera (Packiam et al., 2015).
Общемировая тенденция при выращивании культур в закрытом грунте - расширение
применения против вредителей их естественных врагов с использованием как моно-
фагов, так и полифагов. К последним относится хищный клоп Orius laevigatus, способ-
ный питаться на целом комплексе фитофагов, поэтому чрезвычайно важна оценка
возможного побочного действия любого пестицида независимо от его происхождения
(результат химического синтеза или природные источники). Эфирные масла растений
нельзя считать полностью безопасными для полезных членистоногих. Масла из
Foeniculum vulgare и Citrus limon с высокой биологической активностью для паутинно-
го клеща Tetranychus turkestani были токсичны и для хищного клопа Orius albidipennis,
хотя и в меньшей степени, чем для жертвы (Faraji et al., 2016). Orius strigicollis был
менее восприимчив к 13 образцам эфирных масел по сравнению с фитофагом Thrips
palmi (Yi et al., 2006). Масла из Artemisia sieberi, Pelargonium roseum и Ferula gummosa
при фумигационном воздействии вызывали гибель как хищника Orius albidipennis, так
и его жертвы Bemisia tabaci (Zandi-Sohani et al., 2018). Показано, что на участках, где
использовались растительные вещества, включая 1%-ный раствор масла понгамии,
было значительно больше энтомофагов (насекомых из семейств Coccinelidae и Syr-
phidae и отряда Hymenoptera), чем на участках, где применялись традиционные
инсектициды (Onkara Naik et al., 2019).
По нашим данным, представленным в этой статье, хищный клоп O. laevigatus обла-
дает высокой чувствительностью к маслу P. pinnata при различных способах воздей-
ствия (даже в минимальной эффективной для трипса концентрации 0.75 %). Увеличение
периода ожидания (до 7 дней от обработки до выпуска клопов) не позволило избежать
негативного действия масла на энтомофага.
Полученные нами данные позволяют считать масло P. pinnata, обладающее различ-
ными механизмами действия, эффективным средством снижения численности запад-
ного цветочного трипса. Существенным аргументом в пользу применения данного
масла в защищенном грунте могут быть сведения об его активности для Trialeurodes
vaporariorum и Myzus persicae, которые часто присутствуют в теплицах на растениях
вместе с трипсом (Kumar, Kalindhar, 2003; Pavela, Herda, 2007; Stepanycheva et al.,
2014).
304
Выявленное нами негативное действие масла P. pinnata на O. laevigatus обусловлива-
ет необходимость поиска особой тактики выпуска хищного клопа, сопряженного
с применением этого масла, или использования другого, менее восприимчивого к нему,
биологического объекта.
ФИНАНСИРОВАНИЕ
Доктор Р. Павела выражает благодарность Министерству сельского хозяйства Чеш-
ской Республики за финансовую поддержку исследований по пестицидам раститель-
ного происхождения.
СПИСОК ЛИТЕРАТУРЫ
Abbott W. S. 1925. A method of computing the effectiveness of an insecticide. Journal of Economic Entomology
18: 265-267.
Balikai R. A., Lingappa S. 2005. Management of aphid, Melanaphis sacchari (Zehntner) on rabi sorghum through
botanicals. In: S. Ignacimuthu, S. Jayaraj (eds). Sustainable Insect Pest Management. New Delhi: Narosa
Publishing House, p. 204-209.
Born F. S., da Camara C. A. G., de Melo J. P. R., de Moraes M. M. 2018. Acaricidal property of the essential oil from
Lippia gracilis against Tetranychus urticae and a natural enemy, Neoseiulus californicus, under greenhouse
conditions. Experimental and Applied Acarology 75 (4): 491-502.
Castilhos R. V., Grutzmacher A. D., Coats J. R. 2018. Acute toxicity and sublethal effects of terpenoids and essential
oils on the predator Chrysoperla externa (Neuroptera: Chrysopidae). Neotropical Entomology 47: 311-317.
Faraji N., Seraj A. A., Yarahmadi F., Rajabpour A. 2016. Contact and fumigant toxicity of Foeniculum vulgare and
Citrus limon essential oils against Tetranychus turkestani and its predator Orius albidipennis. Journal of Crop
Protection 5 (2): 283-292.
Gao Y., Lei Z., Reitz S. R. 2012. Western flower thrips resistance to insecticides: detection mechanisms and
management strategies. Pest Management Science 68 (8): 1111-1121.
Ikbal C., Pavela R. 2019. Essential oils as active ingredients of botanical insecticides against aphids. Journal of Pest
Science 92 (3): 971-986.
Kumar S. M. B., Kalidhar S. B. 2003. A review of the chemistry and biological activity of Pongamia pinnata.
Journal of Medicinal and Aromatical Plant Sciences 25: 441-465.
Kumar V., Chandrashekar K., Sidhu O. P. 2006. Efficacy of karanjin and different extracts of Pongamia pinnata
against selected insect pests. Journal of Entomological Research 30: 103-108.
Lale A., Kulkarni D. K. 2010. Mosquito repellent Karanj Kunapa from Pongamia pinnata. Asian Agri-History 14
(2): 207-211.
Mondal M., Khalequzzaman M. 2006. Toxicity of essential oils against Red Flour Beetle, Tribolium castaneum
(Herbst) (Coleoptera: Tenebrionidae). Journal of Bio-Science 14: 43-48.
Mossa A.-T. H. 2016. Green Pesticides: Essential oils as biopesticides in insect-pest management. Journal of
Environmental Science and Technology 9: 354-378.
Negi R. S., Srivastava M., Saxena M. M. 1997. Egg laying and adult emergence of Callosobruchus chinensis on
green gram (Vigna radiata) treated with pongam oil. Indian Journal of Entomology 59 (2): 170-172.
Onkara Naik S., Kannan G. S., Chakravarthy A. K. 2019. Impact of integrated pest management modules on natural
enemies of whiteflies, Bemisia tabaci (Genn.) in bitter gourd ecosystem. Journal of Biological Control 33
(1): 63-69.
Packiam S. M., Emmanuel C., Baskar K., Ignacimuthu S. 2015. Feeding deterrent and genotoxicity analysis of
a novel phytopesticides by using comet assay against Helicoverpa armigera (Hubner) (Lepidoptera:
Noctuidae). Brazilian Archives of Biology and Technology 58 (4): 487-493.
Papadimitriou D. M., Petrakis E. A., Arvaniti K. A., Kimbaris A. C., Polissiou M. G., Perdikis D. Ch. 2019.
Comparative bioactivity of essential oils from two Mentha pulegium (Lamiaceae) chemotypes against
Aphis gossypii, Aphis spiraecola, Tetranychus urticae and the generalist predator Nesidiocoris tenuis.
Phytoparasitica 47 (5): 683-692.
Pascual-Villalobos M. J., Robledo A. 1998. Screening for anti-insect activity in Mediterranean plants. Industrial
Crop and Products 8 (3): 183-194.
Pavela R. 2016. History, presence and perspective of using plant extracts as commercial botanical insecticides and
farm products for protection against insects - a review. Plant Protection Science 52: 229-241.
Pavela R., Herda G. 2007. Repellent effects of pongam oil on settlement and oviposition of the common greenhouse
whitefly Trialeurodes vaporariorum on chrysanthemum. Insect Science 14: 219-224.
305
Pino O., Sánchez Y., Rojas M. M. 2013. Plant secondary metabolites as an alternative in pest management.
I: Background, research approaches and trends. Revista de Protección Vegetal 28 (2): 81-94.
Regnault-Roger C., Vincent C., Arnason J. T. 2012. Essential oils in insect control: low-risk products in a high-stakes
world. Annual Review of Entomology 57: 405-424.
Ribeiro N., Camara C., Ramos C. 2016. Toxicity of essential oils of Piper marginatum Jacq. against Tetranychus
urticae Koch and Neoseiulus californicus (McGregor). Chilean Journal of Agricultural Research 76: 71-76.
Schneweis D. J., Whitfield A. E., Rotenberg D. 2017. Thrips developmental stage-specific transcriptome response
to tomato spotted wilt virus during the virus infection cycle in Frankliniella occidentalis, the primary vector.
Virology 500: 226-237.
Shah M. M. R., Prodhan M. D. H., Siddquie M. N. A., Mamum M. A. A., Shahjahan M. 2008. Repellent effect
of some indigenous plant extracts against saw-toothed grain beetle, Oryzaephilus surinamensis (L.).
International Journal of Sustainable Crop Production 3 (5): 51-54.
Soares M. A., Campos M. R., Passos L. C., Carvalho G. A., Haro M. M., Lavoir A.-V., Biondi A., Zappalà L.,
Desneux N. 2019. Botanical insecticide and natural enemies: a potential combination for pest management
against Tuta absoluta. Journal of Pest Science 92 (4): 1433-1443.
Stepanycheva E. A., Petrova M. O., Chermenskaya T. D., Roman P. 2014. Prospects for the use of Pongamia pinnata
oil-based products against the green peach aphid Myzus persicae (Sulzer) (Hemiptera: Aphididae). Psyche
2014: 5.
Swathi S., Murugananthan G., Ghosh S. K. 2010. Oviposition deterrent activity from the ethanolic extract of
Pongamia pinnata, Coleus forskohlii, and Datura stramonium leaves against Aedes aegypti and Culex
quinquefasciatus. Pharmacognosy Magazine 6 (24): 320-322.
Tayoub G., Abu Alnaser A., Ghanem I. 2012. Fumigant activity of leaf essential oil from Myrtus communis L. against
the khapra beetle. International Journal of Medicinal and Aromatic Plants 2 (1): 207-213.
Tran D. H., Le K. P., Tran H. D. T., Ueno T. 2016. Control efficacy of pongam (Pongamia pinnata L.) leaf extract
against the turnip aphid Lipaphis pseudobrassicae (Davis) (Hemiptera: Aphididae). Journal of the Faculty of
Agriculture, Kyushu University 61 (1): 141-145.
Tran D. H., Takagi M., Ueno T. 2017. Efficacy of the extract from pongam leaves (Pongamia pinnata L.) against
Spodoptera exigua (Hübner) and Spodoptera litura Fabricius (Lepidoptera: Noctuidae). Journal of the
Faculty of Agriculture, Kyushu University 62 (2): 439-443.
Uçak H., Karaca İ., Güven Ö. 2014. The effects of some biopesticides on Frankliniella occidentalis (Pergade)
(Thripidae: Thysanoptera). Türkiye Biyolojik Mücadele Dergisi 5 (2): 137-148.
Yi C. G., Choi B. R., Park H. M., Park C. G., Ahn Y. J. 2006. Fumigant toxicity of plant essential oils to Thrips
palmi (Thysanoptera: Thripidae) and Orius strigicollis (Heteroptera: Anthocoridae). Journal of Economic
Entomology 99 (5): 1733-1738.
Zandi-Sohani N., Rajabpour A., Yarahmadi F., Ramezani L. 2018. Sensitivity of Bemisia tabaci (Hemiptera:
Aleyrodidae) and the generalist predator Orius albidipennis (Hemiptera: Anthocoridae) to vapors of essential
oils. Journal of Entomological Science 53 (4): 493-502.
COMPARATIVE EVALUATION OF REACTIONS OF THE WESTERN FLOWER
THRIPS FRANKLINIELLA OCCIDENTALIS PERG. (THYSANOPTERA,
THRIPIDAE) AND PREDATORY BUG ORIUS LAEVIGATUS FIEBER
(HETEROPTERA, MIRIDAE) TO PONGAMIA PINNATA (L.) PIERRE OIL
E. A. Stepanycheva, I. M. Pazyuk, T. D. Chermenskaya, M. O. Petrova, R. Pavela
Keywords: Pongamia pinnata, oil, seeds, Frankliniella occidentalis, Orius laevigatus, pest,
predator, behaviour, reproduction, toxicity.
SUMMARY
The results of a comparative assessment of the reaction of the western flower thrips Frankliniella
occidentalis and the predatory bug Orius laevigatus to oil from seeds of Pongamia pinnata (L.) Pierre
are presented. The use of P. pinnata oil had a repellent and toxic effect on the thrips adults and larvae,
and also reduced the number of laid eggs. Orius laevigatus was highly sensitive to P. pinnata oil (even
at the minimum effective concentration for thrips 0.75%) under various methods of exposure. The side
effect of the oil on O. laevigatus revealed by us necessitates the development of tactics for the combined
use of the oil and this entomophage or replacing Orius with another predator that is not susceptible to
oil.
306