ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 99, 2, 2020
УДК 595.773.4
КОМПЬЮТЕРНАЯ МИКРОТОМОГРАФИЯ ЛИЧИНКИ
3-ГО ВОЗРАСТА ВОСТОЧНОЙ ФРУКТОВОЙ МУХИ
BACTROCERA DORSALIS (HENDEL, 1912) (DIPTERA,
TEPHRITIDAE) С НОВЫМИ ДАННЫМИ
О МОРФОЛОГИЧЕСКИХ ПРИЗНАКАХ ЛИЧИНОК
МУХ-ПЕСТРОКРЫЛОК ДЛЯ ВИДОВОЙ ДИАГНОСТИКИ
© 2020 г. Т. В. Галинская, 1, 2* О. Г. Овчинникова, 3** И. О. Камаев,
2*** М. Ю. Арапова, 1, 2**** Л. Ю. Крючкова 4*****
1 Кафедра энтомологии Биологического факультета Московского государственного
университета им. М. В. Ломоносова
Ленинские горы, 1-12, Москва, 119234 Россия
*e-mail: nuha1313@gmail.com, ****e-mail: maria.yurevna@bk.ru
2 Научно-методический отдел энтомологии Всероссийского центра карантина растений
***e-mail: ilyakamayev@yandex.ru
ул. Пограничная, 32, пос. Быково, Раменский р-н, Московская обл., 140150 Россия
3 Зоологический институт РАН
Университетская наб., 1, С.-Петербург, 199034 Россия
**e-mail: brach@zin.ru
4 Санкт-Петербургский государственный университет
Университетская наб., 7/9, С.-Петербург, 199034 Россия
*****l.kruchkova@spbu.ru
Поступила в редакцию 28.02.2020 г.
После доработки 9.03.2020 г.
Принята к публикации 9.03.2020 г.
Приведены результаты изучения методом рентгеновской компьютерной микротомографии
(micro-CT) мышечной системы личинки восточной фруктовой мухи Bactrocera dorsalis (Hendel,
1912) (Diptera, Tephritidae), карантинного вредителя многих плодовых культур. Проведено
сравнение мускулатуры у Bactrocera dorsalis и Drosophila melanogaster Meigen, 1830 (Diptera,
Drosophilidae), личинка которого изучена наиболее подробно среди круглошовных двукрылых
(Cyclorrhapha). Несмотря на принадлежность к разным и не близко родственным семействам,
у этих видов обнаружен сходный набор мышц, которые отличаются по местам прикрепления и
степени развития. Более сильное развитие мышц фарингеального склерита и ротовых крючьев
у B. dorsalis по сравнению с D. melanogaster может быть связано с различиями в плотности
субстрата, в котором прокладывают ходы их личинки (у B. dorsalis - ткани плодов и почва (при
зарывании перед окукливанием), а у D. melanogaster - разлагающиеся субстраты).
Ключевые слова: двукрылые, томография, диагностика, карантинные виды.
DOI: 10.31857/S0367144520020136
399
Восточная фруктовая муха Bactrocera dorsalis (Hendel, 1912) - представитель семей-
ства мух-пестрокрылок (Tephritidae) с чрезвычайно высокой расселительной способ-
ностью, происходящий из Юго-Восточной Азии и в последние десятилетия широко
распространившийся на африканском континенте. Этот вид повреждает плоды более
чем 200 видов растений, в том числе сахарное яблоко (Annona squamosa L.), яблоня
(Malus pumila Mill.), банан (Musa paradisiaca L.), перец рода Capsicum, гуава (Psidium
guajava L.), манго (Mangifera indica L.), апельсин (Citrus sinensis (L.) Pers.) и другие
виды рода Citrus, папайя (Carica papaya L.), персик (Persica vulgaris Mill.), слива
(Prunus domestica L.), груши Pyrus spp. и др. Высокая инвазионная активность и спо-
собность осваивать новые виды растений-хозяев, а также находки личинок этого вре-
дителя в импортируемых плодах привели к тому, что он стал объектом фитосанитарного
контроля во многих странах мира, в том числе членах Евразийского экономического
союза (Воробьева, Камаев, 2017).
Личинок мух-пестрокрылок идентифицируют по внутренним и внешним морфоло-
гическим признакам (Кандыбина, 1977; White, Elson-Harris, 1992; Carroll et al., 2004;
Камаев, 2017 и др.). Идентификация их крайне сложна, так как набор диагностических
признаков ограничен, а видам свойственна широкая изменчивость. До сих пор описа-
ны личинки лишь небольшого числа видов рода Bactrocera.
В связи с этим необходимы тщательные исследования строения склеритов, покровов
и мускулатуры личинок мух рода Bactrocera для использования их особенностей
в диагностике карантинных объектов.
Ранее мускулатура личинок двукрылых была изучена методом классического препа-
рирования у пестрокрылки Rhagoletis pomonella (Walsh, 1867) (Snodgrass, 1923);
у Tipulidae (Snodgrass, 1935; Birket-Smith, 1984; Neugart et al., 2009); Nymphomyiidae
(Schneeberg et al., 2012); Exechia (Mycetophilidae) и Bibio (Bibionidae) (Bauernfeind
et al., 2015) и Piophilidae (Martin-Vega, Niederegger, 2015).
Нидереггер с соавт. (Niederegger et al., 2017) показали, что места прикрепления мышц
специфичны для некоторых видов семейств Calliphoridae и Sarcophagidae, используе-
мых в криминологии, однако для видов сем. Piophilidae этот признак оказался не при-
меним.
Мускулатура личинок была изучена с помощью компьютерной томографии Випфле-
ром с соавт. (Wipfler et al., 2012a) у Protanyderus (Tanyderidae), а наиболее подробно
среди круглошовных мух (Diptera, Cyclorrhapha) теми же авторами (Wipfler et al.,
2012b) - у Drosophila melanogaster Meigen, 1830 (Drosophilidae).
МАТЕРИАЛ И МЕТОДИКА
Два экземпляра личинок третьего возраста Bactrocera dorsalis, полученные из образцов для
лабораторных исследований плодов из Таиланда, были умерщвлены кипятком и зафиксированы
в 70%-ном этаноле. У одного экземпляра была отсечена передняя часть тела, у второго была
отсечена задняя часть тела для наилучшей фиксации в 96%-ном этаноле и контрастирования
тканей. Фрагменты тела были подготовлены для компьютерной микротомографии (micro-CT)
путем контрастирования йодом по методике, описанной в работе Жиньяка и Клея (Gignac, Kley,
2014) с последующим высушиванием в критической точке. Экземпляры были наклеены на уз-
кую вертикальную деревянную подставку, один - вверх передним концом тела, второй - задним;
оба экземпляра были зафиксированы вертикально, настолько близко к оси вращения томографа,
насколько возможно.
400
Исследования проводились на микротомографе Bruker SkyScan-1172 (Bruker micro-CT,
Belgium) в ресурсном центре «Рентгенодифракционные методы исследования» Санкт-Петер-
бургского государственного университета (Россия). Режимы микротомографического сканиро-
вания: излучение медного анода, ускоряющее напряжение 25 кВ, сила тока 139 mА, разрешение
0.9 мкм, угол поворота образца 0.2°, число сканирований в одном положении 6, экспозиция
2.7 сек.
Для реконструкции массива теневых изображений использовалась программа NRecon (Bruker
Micro-CT), позволяющая нивелировать приборные артефакты и задавать диапазон градаций се-
рого цвета, соответствующих величине рентгеновского поглощения и, соответственно, рентге-
новской плотности. Для анализа полученных микротомографических данных использовались
программы DataViewer и CTVox (Bruker Micro-CT).
При описании строения личинки использована терминология М. Н. Кандыбиной (1977);
в Таблице 1 приведены соответствующие названия в наиболее употребляемых системах англо-
язычных авторов (Wipfler et al., 2012b; Balmès, Mouttet, 2017). Номера мышц даны по Випфлеру
с соавт. (Wipfler et al., 2012b)..
РЕЗУЛЬТАТЫ
Общее строение личинки
Личинка 3-го возраста конической формы; средние и задние сегменты цилиндриче-
ские, около 4-5 мм в длину и около 1 мм в ширину; тело наиболее широкое в средней
области и сужается к псевдоцефалону. Кутикула тонкая, очень слабо склеротизованная,
прозрачная. Сильно склеротизованы только части ротоглоточного аппарата (= цефало-
фарингеального скелета, рис. 1), которые видны через наружную кутикулу. Ротоглоточ-
ный аппарат включает следующие части: парные мандибулярные склериты (ротовые
Таблица 1. Используемые в статье названия склеритов ротоглоточного аппарата по тер-
минологиям разных авторов (Кандыбина, 1977; Wipfler et al., 2012b; Balmès, Mouttet, 2017)
Кандыбина, 1977
Wipfler et al., 2012b
Balmès, Mouttet, 2017
Мандибулярные склериты
Mouth hook
-
(ротовые крючья)
Постеродорсальные отростки
Posterodorsal process of the
-
мандибулярных склеритов
mouth hook
-
Posterior process of the mouth
Ventral apodeme of mandible
hook
(в русскоязычной
литературе закрепился
термин «вентральные
аподемы мандибуляр-
ных склеритов»)
Зубные склериты
Mouth angle piece
-
Гипостомальные склериты
H-piece, sclerite between mouth
-
hooks and cephaloskeleton
Фарингеальные склериты
Cephaloskeleton
-
Задние дорсальные отростки
Dorsal posterior process (arms)
-
фарингеальных склеритов
of cephaloskeleton
Вентральные отростки фаринге-
Ventral posterior process (arms)
-
альных склеритов
of cephaloskeleton
401
Рис. 1. Bactrocera dorsalis (Hendel), ротоглоточный аппарат личинки 3-го возраста.
Обозначения структур по: Кандыбина, 1977; в скобках - по Випфлеру с соавт.
(Wipfler et al., 2012б).
1 - мандибулярный склерит (the mouth hook), 2 - вентральная аподема мандибулярного склерита (posterior
process of the mouth hook), 3 - постеродорсальный отросток мандибулярного склерита (posterodorsal process
of the mouth hook), 4 - зубной склерит (mouth angle piece), 5 - гипостомальный склерит, 6 - фарингеальный
склерит (cephaloskeleton), 7 - задние дорсальные отростки фарингеального склерита
(dorsal posterior process (arms) of cephaloskeleton), 8 - вентральные отростки
фарингеального склерита (ventral posterior process (arms) of cephaloskeleton).
крючья), имеющие постеродорсальные отростки и вентральные аподемы (термин от-
сутствует в монографии Кандыбиной (1977), но теперь принят (Balmès, Mouttet, 2017);
зубные склериты, гипостомальные склериты, фарингеальные склериты с задними дор-
сальными отростками и вентральными отростками.
Тело подразделяется на отчетливо уменьшенный псевдоцефалон (= ложноголова),
три грудных сегмента и удлиненное брюшко. Псевдоцефалон втягивается в передне-
грудь, которая несет пару дорсолатеральных дыхалец в виде пластин с небольшими
лопастями. Другие грудные и брюшные сегменты I-VII лишены дыхалец и очень сход-
ны по форме. Последний абдоминальный «сегмент» точнее называть анальным отде-
лом, поскольку он состоит из нескольких брюшных сегментов. В его верхней части
есть задняя пара дыхалец, а в нижней располагаются анальные бугорки. Грудные и
брюшные, или ложные, ноги отсутствуют.
Наиболее подробное описание габитуса личинки приведено в монографии М. Н. Кан-
дыбиной (1977) и в интернет-ресурсе (Carroll et al., 2006).
Мускулатура
Мышечная система личинки образует плотный трехмерный каркас под стенкой тела,
включает только продольные (прямые и косые) и дорсовентральные мышцы, тогда как
кольцевые мышцы стенки тела отсутствуют (в отличие от имаго насекомых). Продоль-
ные межсегментные мышцы обычно соединяют два последовательно расположенных
402
сегмента, но некоторые из них связывают не соседние сегменты, а проходят насквозь
через несколько сегментов. Продольные внутрисегментные мышцы прикрепляется
обоими концами внутри одного сегмента. Мышцы ротоглоточного аппарата хорошо
развиты, простираются между фарингеальным склеритом, мандибулярными склерита-
ми и стенкой тела.
Все описанные ниже мышцы парные, если не указано иное.
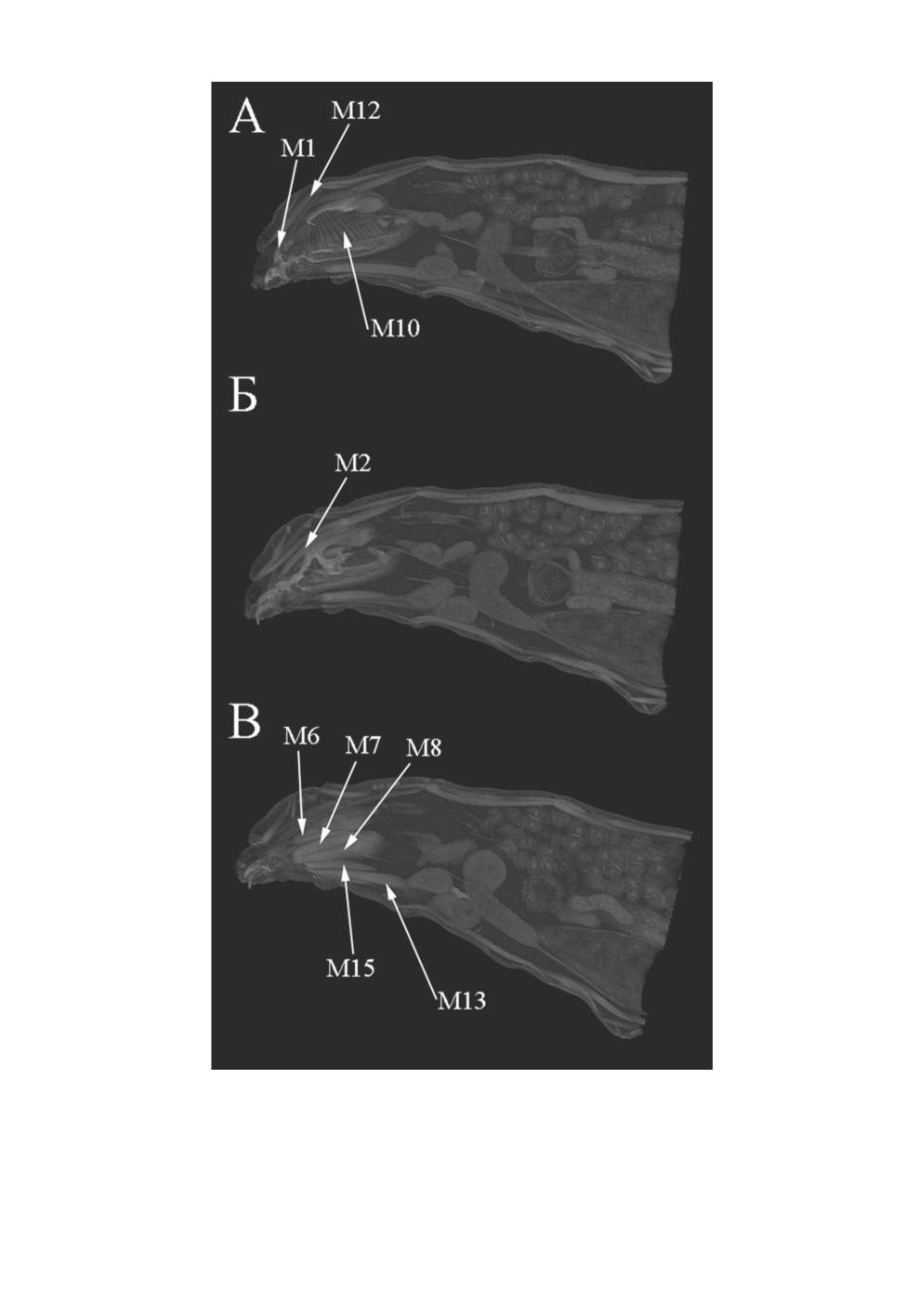
Ротоглоточный аппарат (рис. 1-3). Мышцы M1 связывают дорсальную сторону
мандибулярного склерита с задней границей дорсолатеральной стенки переднегруди.
Мышцы М2 прикрепляются к гипостомальному склериту и к средней части дорсола-
теральной стенки среднегруди.
Мышцы М3 тянутся от задних дорсальных отростков фарингеального склерита
к передней части дорсальной стенки переднегруди.
Мышцы M4 соединяют задние дорсальные отростки фарингеального склерита
со средней частью дорсальной стенки переднегруди дорсальнее M3.
Широкие мышцы М5 связывают задние дорсальные отростки фарингеального скле-
рита с задней границей вентролатеральной стенки переднегруди.
Мышцы М6 прикрепляются к мембране у постеродорсального отростка мандибуляр-
ного склерита и к месту отхождения вентральных отростков фарингеального склерита.
Мышцы М7 тянутся от дорсальной поверхности вентральных отростков фаринге-
ального склерита к мембране задней поверхности вентральной аподемы мандибуляр-
ного склерита и к зубному склериту, который примыкает к указанной аподеме.
Мышцы M8 связывают дорсальную поверхность вентральных отростков фаринге-
ального склерита поствентральнее мышц M7 с мембраной задней поверхности вент-
ральной аподемы мандибулярного склерита и зубным склеритом, который примыкает
к указанной аподеме. Мышцы М8 очень близко примыкают к М7, образуя общий
пучок.
Мышцы M9 соединяют вентральные отростки фарингеального склерита (ближе
к центральной оси тела, расположены под M8) с вентральной аподемой мандибулярно-
го склерита и зубным склеритом, который примыкает к указанной аподеме.
Широкие мышцы M10 прикрепляются к задним дорсальным и вентральным отрост-
кам фарингеального склерита, охватывая дорсальную сторону кишки.
Мышцы М11 тянутся от дорсальной стенки слюнного протока к внешней поверхно-
сти вентральных отростков фарингеального склерита кпереди от мышц М7.
11 пар глоточных расширителей простираются между задними дорсальными отрост-
ками фарингеального склерита и дорсальной стороной пищевода.
Псевдоцефалон и грудные отделы. В п е р е д н е г р у д н о м отделе имеются 3 па-
ры внутрисегментных продольных мышц дорсолатерально, латерально и вентролате-
рально, а также 1 пара дорсовентральных мышц (рис. 2).
Мышцы М12 (рис. 3) связывают вентральную стенку псевдоцефалона медиальнее
мандибулярных склеритов со средней частью дорсальной стенки переднегруди лате-
ральнее мышц М3 и М4.
403
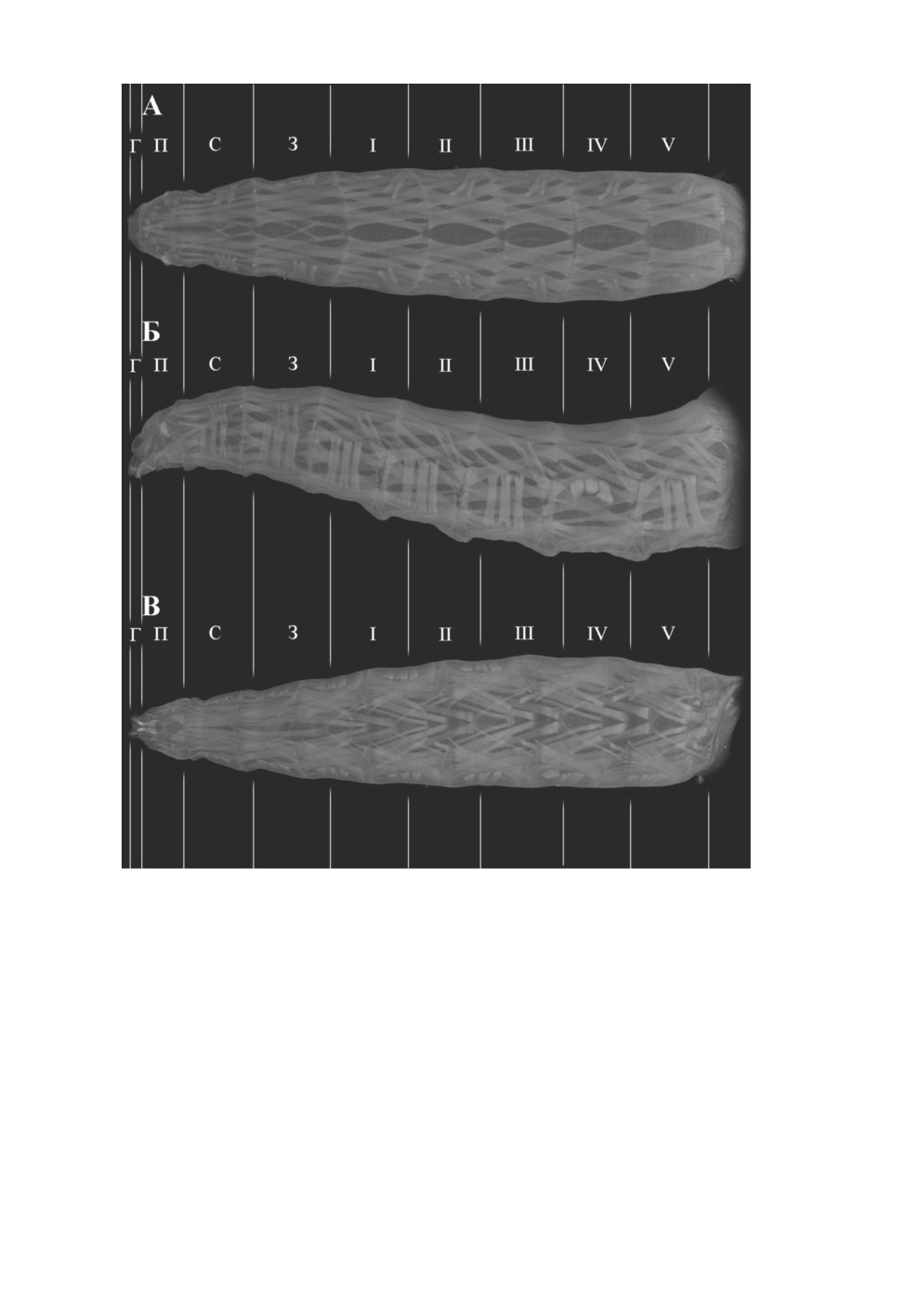
Рис. 2. Bactrocera dorsalis (Hendel), общий вид передней части тела личинки сверху (А),
сбоку (Б) и снизу (В).
Г - псевдоцефалон; П, С, З - передне-, средне- и заднегрудь; I-V - брюшные сегменты.
Найдены 6 пар межсегментных мышц (М13-М18).
Мышцы М13 тянутся от вентральной части заднего края псевдоцефалона к задневен-
тральной границе брюшных сегментов I и II.
Мышцы M14 связывают переднюю вентральную границу переднегруди с задней
латеровентральной границей заднегруди. Оба места прикрепления расположены
дорсолатеральнее мышц М13.
Мышцы M15 соединяют переднюю вентральную границу переднегруди с задней
латеровентральной границей заднегруди. Оба места прикрепления расположены
дорсолатеральнее мышц М14.
404
Рис. 3. Bactrocera dorsalis (Hendel), внутреннее строение передней части тела личинки.
405
Мышцы M16 идут от передней части вентральной стенки переднегруди к задней
части латеровентральной стенки среднегруди. Оба места прикрепления расположены
вентролатеральнее М13.
Мышцы M17 прикрепляются к латерофронтальной стенке псевдоцефалона и к зад-
ней части дорсальной стенки среднегруди.
Мышцы M18 тянутся от латеральной стенки псевдоцефалона позади мандибулярных
склеритов к задней части медиодорсальной стенки среднегруди.
В с р е д н е г р у д н о м отделе найдено 11 пар внутрисегментных продольных мышц
и 5 пар дорсовентральных мышц. Продольные мышцы, ближайшие к латеральной
стенке тела, очень широкие.
В з а д н е г р у д н о м отделе имеется 11 пар внутрисегментных продольных мышц
и 5 пар дорсовентральных мышц.
Найдены следующие 3 пары межсегментных мышц (М19-М21).
Мышцы M19 тянутся от передней части медиодорсальной стенки заднегруди к зад-
ней части медиодорсальной стенки брюшного сегмента II.
Мышцы M20 прикрепляются к передней части вентральной стенки заднегруди и к
вентральной стенке брюшного сегмента I.
Мышцы M21 связывают переднюю часть вентральной стенки заднегруди с задней
частью вентральной стенки брюшного сегмента I. Оба места прикрепления располо-
жены вентральнее М20.
Брюшной сегмент I. В брюшном сегменте I найдено 14 пар внутрисегментных про-
дольных мышц и 5 пар дорсовентральных мышц.
Брюшные сегменты II-VI. В сегментах II и III найдены одинаковые внутри-
сегментные продольные и дорсовентральные мышцы.
В каждом сегменте есть по 16 пар продольных и 5 пар дорсовентральных мышц.
Мышцы брюшных сегментов IV-VI очень сходны по структуре и местам прикрепле-
ния с описанными выше мышцами сегментов II и III, поэтому мы не приводим их
описания.
Найдены 4 пары межсегментных мышц (M22-M25).
Мышцы M22 идут от передней части вентральной стенки брюшного сегмента II
к задней части латеровентральной стенки сегмента VII.
Мышцы M23 тянутся от передней части вентральной стенки брюшного сегмента II
к задней части латеровентральной стенки сегмента VII, оба места прикрепления рас-
положены дорсальнее мышц М22.
Мышцы M24 связывают переднюю часть вентральной стенки брюшного сегмента II
со средней частью вентральной стенки сегмента III.
Мышцы M25 соединяют переднюю часть вентральной стенки брюшного сегмента II
со средней частью вентральной стенки сегмента III, оба места прикрепления располо-
жены вентральнее мышц M24.
406
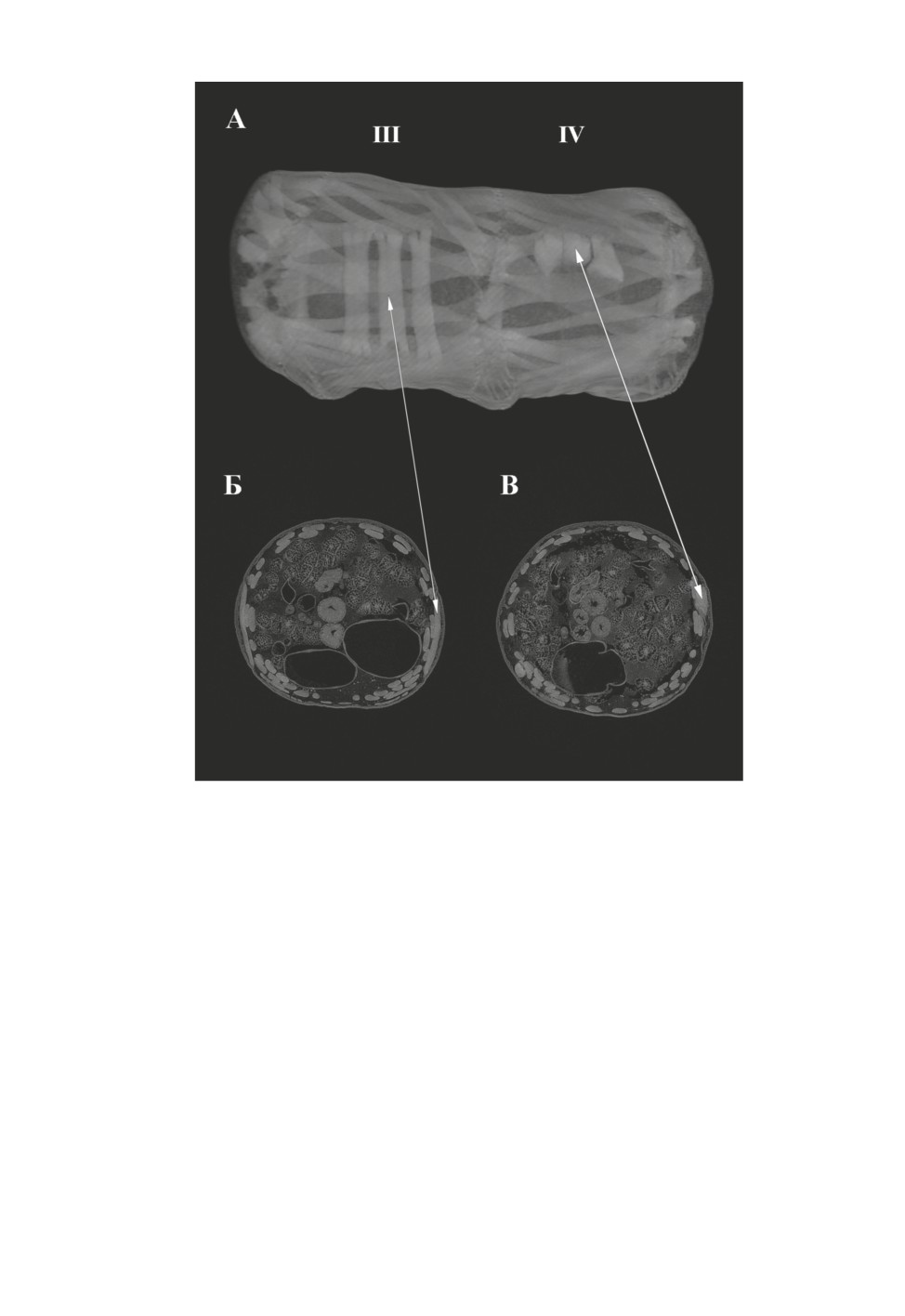
Рис. 4. Bactrocera dorsalis (Hendel), брюшные сегменты личинки. Сегмент III включает
не сокращенные дорсовентральные мышцы, сегмент IV - сокращенные дорсовентральные
мышцы (А). Эти же мышцы на поперечных срезах (Б и В).
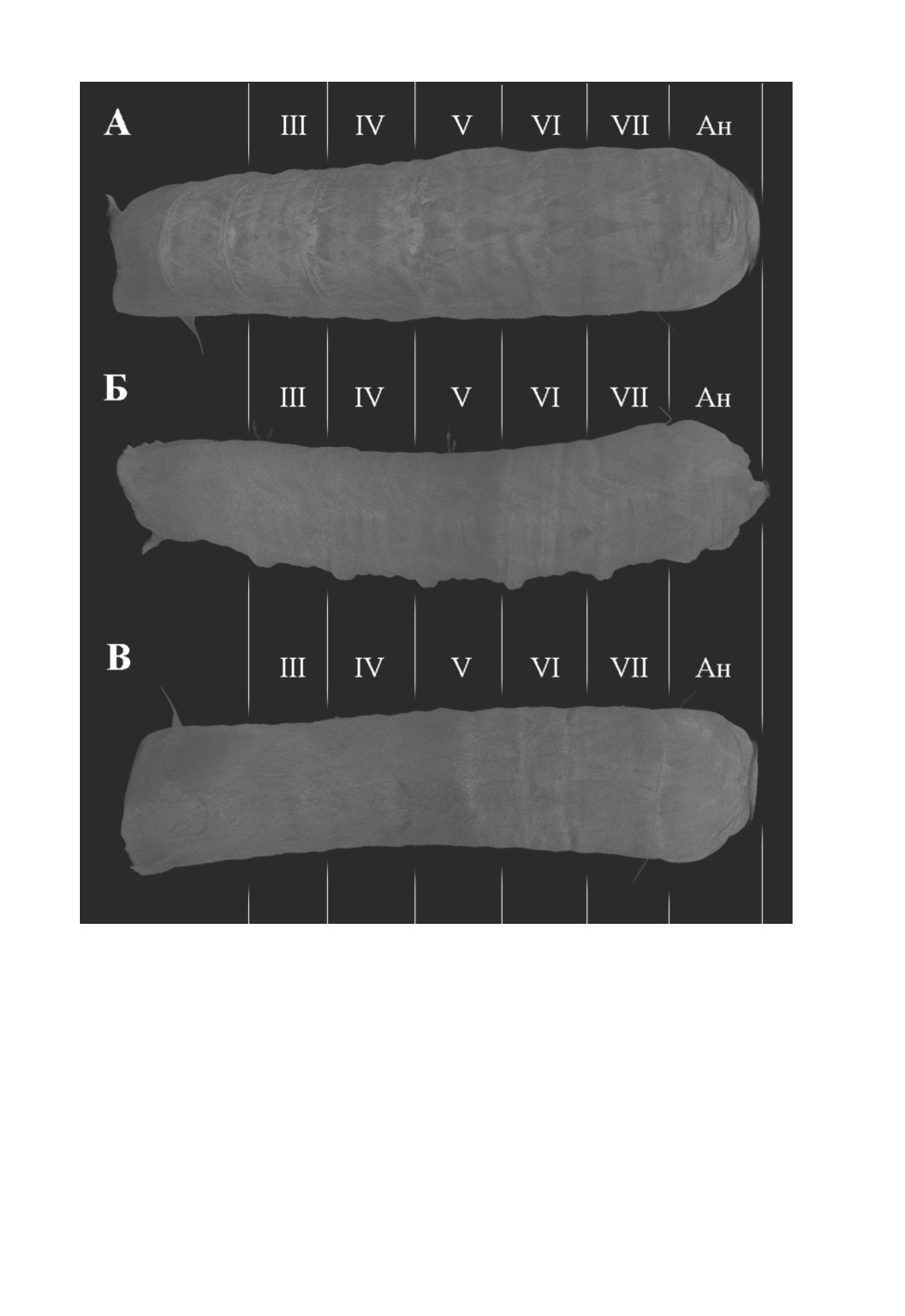
Брюшной сегмент VII и анальный отдел. Мускулатура брюшного сегмента VII
очень сходна с мускулатурой предшествующего сегмента VI и включает главным обра-
зом продольные и дорсовентральные мышцы (рис. 4, 5), разница лишь в большем ко-
личестве пучков мышц в сегменте VII. Анальный отдел характеризуется явно
уменьшенным набором мышц (рис. 6): содержит только 5 пучков как продольных, так
и дорсовентральных мышц и 4 мышцы, связанные с дыхальцем.
Мышцы M26, 4 пучка, тянутся парамедиально от переднего дорсального края брюш-
ного сегмента VII к переднему дорсальному краю анального отдела; функционируют
как ретракторы анального отдела.
Мышцы M27, 3 пучка, связывают передний край сегмента VII латеральнее мышц
M26 с передним краем анального отдела вентральнее мышц M26.
407
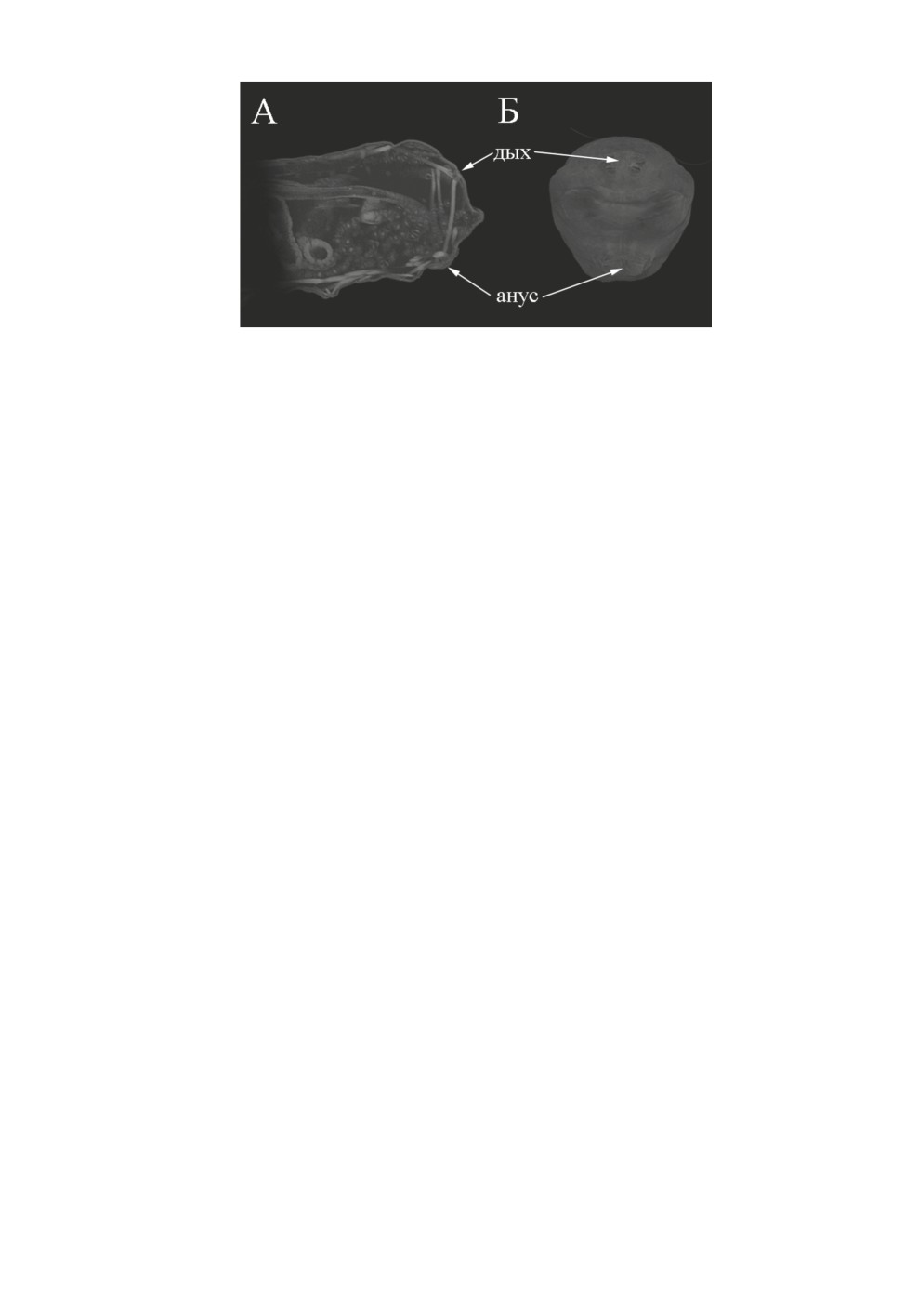
Рис. 5. Bactrocera dorsalis (Hendel), общий вид задней части тела личинки сверху (А), сбоку (Б)
и снизу (В).
Ан - анальный сегмент, III-VII - брюшные сегменты.
Мышца M28, 1 непарный пучок, идет от передней латеральной части сегмента VII
к переднему краю анального отдела вентральнее мышц M26.
Мышца M29, 1 непарный пучок, тянется от передней части боковой стенки сегмента
VII вентральнее мышц M28 к передней части боковой стенки анального отдела.
Мышца M30, 1 непарный пучок, идет от передней части боковой стенки сегмента VII
вентральнее мышц M26 и латеральнее мышц M36 и M37 к передней части боковой
стенки анального отдела.
408
Рис. 6. Bactrocera dorsalis (Hendel), внутреннее строение анальной части тела личинки.
Дых - дыхальце.
Мышца M31, 1 непарный пучок, связывает переднюю часть боковой стенки сегмента
VII вентральнее мышцы M30 с передней частью боковой стенки анального отдела
(передняя граница сегмента) вентральнее мышцы M29.
Мышца М32, 5 пучков, идет от передней части вентральной стенки сегмента VII
к передней части вентральной стенки анального отдела.
Мышца M33, 5 пучков, соединяет переднюю часть брюшной стенки сегмента VII
(граница сегмента) латеральнее мышцы M32 с передней частью вентролатеральной
стенки анального отдела латеральнее мышц М34.
Мышца M34, 5 пучков, тянется от передней части боковой стенки сегмента VII дор-
сальнее М31 к передней части боковой стенки анального отдела ближе к центральной
оси тела, чем мышцы M33.
Мышца M35, 1 непарный пучок, соединяет переднюю часть боковой стенки сегмен-
та VII латеральнее мышцы M34 с передней частью вентролатеральной стенки анально-
го отдела латеральнее мышцы M33.
Дорсовентральная мышца M36, 1 непарный пучок, прикрепляется латерально к пе-
редней границе сегмента VII ближе к оси тела, чем мышца M30.
Дорсовентральная мышца M37, 3 пучка, прикрепляется латерально к средней обла-
сти сегмента VII позади M36 и латеральнее M28, ближе к оси тела, чем мышца M30.
Дорсовентральная мышца М38, 1 непарный пучок, прикрепляется латерально к зад-
нему краю сегмента VII позади М37 латеральнее М27 и М28.
Мышца М39, 1 непарный пучок, вентральная поперечная мышца, расположена близ-
ко к заднему краю сегмента VII.
Мышца M40, 1 непарный пучок, связывает переднюю часть дорсальной стенки
анального отдела с дорсальной частью терминальных дыхалец.
Мышца M41, 1 непарный пучок, идет паралатерально от вентральной части анально-
го отдела к вентральной части терминальных дыхалец.
Мышца M42, 1 непарный пучок, тянется парамедиально от вентральной части аналь-
ного отдела к вентральной части терминальных дыхалец.
409
Кольцевая мышца M43 охватывает терминальные дыхальца.
Мышца M44, 1 непарный пучок, связывает переднюю часть дорсолатеральной стен-
ки анального отдела с ее задней частью.
Мышцы М45, 3 пучка, идут от передней части вентролатеральной стенки анального
отдела вентральнее М41 к его задневентральной стенке латеральнее М42.
Мышца M46, 1 непарный пучок, дорсовентральная мышца, идет от передней дор-
сальной части анального отдела к его вентролатеральной стенке; функционирует как
депрессор анального отдела.
Мышца М47, 1 непарный дорсовентральный пучок, тянется от передней дорсолате-
ральной части анального отдела позади М46 к вентральной стенке отдела.
Мышца M48, 1 непарный дорсальный поперечный пучок, прикрепляется к средней
области анального отдела и к его вентральной стенке.
Мышца M49, 1 непарный поперечный пучок, наклонно простирающийся через
анальный отдел от задней части его дорсолатеральной стенки к вентролатеральной
стенке.
Мышца M50, 1 непарный пучок, связывает вентролатеральную стенку анального от-
дела с прямой кишкой латеральнее мышцы M41.
Мышца М51, 1 непарный пучок, связывают боковую стенку анального отдела лате-
ральнее мышцы М46 с противоположной боковой стенкой латеральнее мышцы М49.
ОБСУЖДЕНИЕ
Мы впервые полностью изучили мышечную систему личинки представителя сем.
Tephritidae, которая оказалась сходной с той частью мускулатуры Rhagoletis pomonella,
вида этого же семейства, которая была изучена Снодграссом (Snodgrass,
1923).
У Bactrocera dorsalis также найдены «брюшные тяжи» (в данной статье мышцы
М13-М16) и диагональные мышцы по всему телу, образующие каркас (в данной статье
дорсовентральные мышцы сегментов груди и брюшка).
Проведено сравнение мускулатуры Bactrocera dorsalis с мускулатурой Drosophila
melanogaster - вида с наиболее подробно изученной личинкой среди круглошовных
двукрылых (Diptera Cyclorrhapha) (Wipfler et al., 2012b). Несмотря на то, что B. dorsalis
и D. melanogaster - представители не только разных семейств (Tephritidae и Droso-
philidae), но и не близко родственных надсемейств Acalyptratae (Tephritoidea и Ephydro-
idea), набор мышц у них очень сходный. Вероятно, сходство личиночной мускулатуры
у высших акалиптратных мух (Diptera, Cyclorrhapha, Acalyptratae) объясняется специ-
фическим образом жизни.
Места прикрепления и степень развития мышц несколько отличаются: так, у D. me-
lanogaster мышцы М7 и М8 тянутся от заднего конца вентральных отростков фаринге-
ального склерита к вентральной аподеме мандибулярного склерита, в то время как
у B. dorsalis эти мышцы идут от дорсальной поверхности вентральных отростков фа-
рингеального склерита к мембране задней поверхности вентральной аподемы манди-
булярного склерита и зубному склериту, который примыкает к указанной аподеме.
Таким образом, у личинок B. dorsalis плечо рычага, образованного мышцами M7
и M8 и фарингеальным склеритом, становится больше, поэтому при одинаковом мы-
410
шечном усилии производимое действие будет эффективнее, чем у дрозофилы. Кроме
этого, мышцы M10 у B. dorsalis развиты сильнее, чем у D. melanogaster, что дополни-
тельно усиливает фарингеальный склерит.
Более сильное развитие мышц фарингеального склерита и ротовых крючьев
у B. dorsalis по сравнению с D. melanogaster может быть связано с прокладыванием
личинками ходов в разных субстратах (у B. dorsalis - ткани плодов и почва (при зары-
вании перед окукливанием), у D. melanogaster - разлагающиеся субстраты). Усиление
мускулатуры фарингеального склерита, возможно, нужно также для защиты склеритов
во время ударов о субстрат при так называемых «прыжках» личинок - специфической
манере передвижения личинок B. dorsalis.
Мышцы в IV сегменте брюшка B. dorsalis на рис. 4, возможно, сокращены из-за пе-
ристальтики. На нашем материале видно, однако, что сокращение мышц в тех случаях,
когда они прикрепляется не к склеритам, может визуально изменять расположение
мест их прикрепления относительно других мышц, вероятно, за счет стягивания
стенки тела. Тем не менее, мы предполагаем, что сравнительный анализ мышц рото-
глоточного аппарата может быть полезным для определения видов на личиночной ста-
дии - степень развития мышц и места их прикрепления могут различаться в связи
с особенностями строения склеритов у разных видов Tephritidae.
БЛАГОДАРНОСТИ
Авторы выражают благодарность специалистам ФГБУ «Всероссийский центр каран-
тина растений» за помощь в сборе материала при выполнении данной работы.
ФИНАНСИРОВАНИЕ
Работа Т. В. Галинской выполнена при финансовой поддержке Российского научного
фонда (грант № 18-74-00035). Работа О. Г. Овчинниковой выполнена на базе Зоологи-
ческого института РАН (гостема АААА-А19-119020690082-8) и при финансовой под-
держке Российского фонда фундаментальных исследований (грант № 18-04-00354-а).
СПИСОК ЛИТЕРАТУРЫ
Воробьева Л. В., Камаев И. О. 2017. Таксономический статус и инвазионная активность восточной фрукто-
вой мухи Bactrocera dorsalis (Diptera: Tephritidae). Карантин растений. Наука и практика 20 (2): 20-55.
Камаев И. О. 2017. Идентификация личинок карантинных для Российской Федерации видов мух-пестрокры-
лок (Diptera: Tephritidae). Карантин растений. Наука и практика 22 (4): 52-64.
Кандыбина М. Н. 1977. Личинки плодовых мух-пестрокрылок (Diptera, Tephritidae). Л.: Наука, 211 с.
Balmès V., Mouttet R. 2017. Development and validation of a simplified morphological identification key for larvae
of tephritid species most commonly intercepted at import in Europe. EPPO Bulletin 47 (1): 91-99.
Bauernfeind R., Schneeberg K., Beutel R. G. 2015. The larval head of Exechia (Mycetophilidae) and Bibio
(Bibionidae) (Diptera). Arthropod Structure and Development 44: 326-345.
Birket-Smith S. J. R. 1984. Prolegs, legs and wings of insects. Entomonograph 5: 1-128.
Carroll L. E., Norrbom A. L., Dallwitz M. J., Thompson F. C. 2006. Pest fruit flies of the world - larvae. Version:
Gignac P. M., Kley N. J. 2014. Iodine-enhanced micro-CT imaging: Methodological refinements for the study of the
soft-tissue anatomy of post-embryonic vertebrates. Journal of Experimental Zoology Part B: Molecular and
Developmental Evolution 322 (3): 166-176.
Martín-Vega D., Niederegger S. 2015. Larval muscle attachment site (MAS) patterns are a conserved character
among Piophilini flies (Diptera, Piophilidae). Deutsche Entomologische Zeitschrift 62 (2): 239-245.
Neugart C., Schneeberg K., Beutel R. G. 2009. The morphology of the larval head of Tipulidae (Diptera, Insecta) -
The dipteran ground plan and evolutionary trends. Zoologischer Anzeiger 248: 213-235.
411
Niederegger S., Döge K. P., Peter M., Eickhölter T., Mall G. 2017. Connecting the dots: From an easy method to
computerized species determination. Insects 8 (2): 52.
Schneeberg K., Friedrich F., Courtney G. W., Wipfler B., Beutel R. G. 2012. The larvae of Nymphomyiidae (Diptera,
Insecta) - Ancestral and highly derived? Arthropod Structure and Development 41: 293-301.
Snodgrass R. E. 1935. Principles of Insect Morphology. New York, London: McGraw-Hill Book Co., Inc., 667 p.
Snodgrass R. E. 1923. The anatomy and metamorphosis of the apple maggot (Rhagoletis pomonella Walsh.). Journal
of the Washington Academy of Sciences 13: 260-261.
White I. M., Elson-Harris M. M. 1992. Fruit Flies of Economic Significance: Their Identification and Bionomics.
Wallingford: CAB International, 601 p.
Wipfler B., Courtney G. W., Craig D. A., Beutel R. G. 2012a. First μ-CT-based 3D reconstruction of a dipteran
larva - The head morphology of Protanyderus (Tanyderidae) and its phylogenetic implications. Journal of
Morphology 273: 968-980.
Wipfler B., Schneeberg K., Löffler A., Hünefeld F., Meier R., Beutel R. G. 2012b. The skeletomuscular system of the
larva of Drosophila melanogaster (Drosophilidae, Diptera) - A contribution to the morphology of a model
organism. Arthropod Structure and Development 42: 47-68.
COMPUTER MICROTOMOGRAPHY OF THE 3RD INSTAR LARVA
OF THE EASTERN FRUIT FLY BACTROCERA DORSALIS (HENDEL, 1912)
(DIPTERA, TEPHRITIDAE), WITH NEW DATA ON MORPHOLOGICAL
CHARACTERS OF FRUIT FLIES’ LARVAE USEFUL
FOR SPECIES DIAGNOSTICS
T. V. Galinskaya, O. G. Ovtshinnikova, I. O. Kamaev, M. Yu. Arapova, L. Yu. Kryuchkova
Key words: Diptera, tomography, diagnostics, quarantine species.
SUMMARY
The results of a study of the muscular system of the larva of the eastern fruit fly Bactrocera dorsalis
(Hendel, 1912) (Diptera, Tephritidae), a quarantine pest of many fruit crops, by X-ray computer
microtomography (micro-CT) are presented. The muscles of Bactrocera dorsalis were compared with
those of Drosophila melanogaster Meigen, 1830 (Diptera, Drosophilidae) as the most thoroughly
studied larva of the Diptera Cyclorrhapha. Despite belonging to different unrelated families, a similar
set of muscles was found in these species, the differences being present in the places of their attachment
and degree of development.
The stronger muscle development of pharyngeal scleritis and oral hooks in Bactrocera dorsalis
compared to Drosophila melanogaster can be associated with the larval habitation in different substrates
(Bactrocera dorsalis - fruit tissues and soil (when buried before pupation), Drosophila melanogaster -
decaying substrates).
412