ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 99, 3, 2020
УДК 595.754
ДИАГНОСТИКА ГРУППЫ ВИДОВ РОДА POLYMERUS HAHN
(HETEROPTERA, MIRIDAE), ВКЛЮЧАЮЩЕЙ ВРЕДИТЕЛЕЙ
АГРОКУЛЬТУР В ВОСТОЧНОЕВРОПЕЙСКОЙ ЛЕСОСТЕПИ,
ПО МОРФОЛОГИЧЕСКИМ И МОЛЕКУЛЯРНЫМ ПРИЗНАКАМ
В. Б. Голуб, 1* М. Ю. Сыромятников, 1, 2** А. В. Кокина, 1***
В. А. Соболева, 1**** Е. В. Нестерова, 1***** В. Н. Попов, 1, 2******
1 Воронежский государственный университет
Университетская пл., 1, Воронеж, 394018 Россия
*e-mail: v.golub@inbox.ru, **e-mail: syromyatnikov@bio.vsu.ru,
***e-mail: nastenka.kokina@mail.ru, ****e-mail: soboleva@bio.vsu.ru,
*****e-mail: katya.nesterova.1997@mail.ru, ******e-mail: pvn@bio.vsu.ru
2 Воронежский государственный университет инженерных технологий
проспект революции, 19, Воронеж, 394036 Россия
Поступила в редакцию 7.05.2020 г.
После доработки 1.08.2020 г.
Принята к публикации 1.08.2020 г.
В статье представлены усовершенствованная таблица для определения совместно обитающих
видов рода Polymerus Hahn, 1831 (Heteroptera, Miridae) на основе морфологических признаков и
полученных авторами данных о различиях между видами в нуклеотидной последовательности
ДНК гена COI. Комплекс этих показателей с учетом хранящихся в GenBank данных обеспечи-
вает возможность достоверной и оперативной идентификации Polymerus unciniger Gapon, 2014,
P unifasciatus (Fabricius, 1794), P. cognatus (Fieber, 1858), P. vulneratus (Panzer, 1806), и P. brevicornis
(Reuter, 1879). Подроды Poeciloscytus Fieber, 1858 и Pachycentrum Gapon, 2014 характеризуются
значительной обособленностью по морфологическим и молекулярно-генетическим признакам.
Разработан ПЦР-ПДРФ-метод быстрой идентификации вредного вида P. vulneratus. Выявлена
высокая внутрипопуляционная изменчивость морфологических признаков P. unifasciatus и ну-
клеотидной последовательности гена COI у форм, депонированных в GenBank как P. unifasciatus
и P. microphthalmus (Wagner, 1951). Выдвинуто предположение о том, что P. microphthalmus пред-
ставляет собой форму P. unifasciatus с укороченными надкрыльями и более мелкими глазами по
сравнению с полнокрылой формой того же вида, особи которой имеют более крупные глаза.
Ключевые слова: Polymerus, виды, вредители, определительная таблица, баркодинг ДНК,
ПЦР-ПДРФ анализ, определение.
DOI: 10.31857/S0367144520030120
Род Polymerus Hahn, 1831 включает 20 палеарктических видов, относящихся по
классификации Д. А. Гапона (Gapon, 2014) к трем подродам - Polymerus s. str. (4 вида),
Poeciloscytus Fieber, 1858 (14 видов) и Pachycentrum Gapon, 2014 (3 вида) (Kerzhner,
Josifov, 1999; Gapon, 2014). В европейской части России к настоящему времени из
666
подрода Polymerus известны 2 вида: P. holosericeus Hahn, 1831 (типовой вид) и P. rus-
satus Gapon, 2014; из подрода Poeciloscytus указаны 6 видов: P. asperulae (Fieber, 1861),
P. brevicornis (Reuter, 1879), P. cognatus (Fieber, 1858), P. palustris (Reuter, 1907),
P. unifasciatus (Fabricius, 1794) (типовой вид) и P. vulneratus (Panzer, 1806); подрод
Pachycentrum представлен в европейской части России всеми тремя видами (Gapon,
2014): P. nigrita (Fallén, 1807), P. carpathicus Horváth, 1882 (типовой вид) и P. unciniger
Gapon, 2014.
Таксономии палеарктических видов рода Polymerus и выяснению различий между
близкими видами, включая вредителей агрокультур, посвящено много публикаций
(Wagner, 1951, 1961, 1971; Carvalho, Leston, 1952; Кержнер, Ячевский, 1964; Пучков,
1966, 1972; Голуб, 1983; Rinne, 1989; Herczek, 1991; Gapon, 2014, и др.). В статье
М. Шварца с соавт. (Schwartz et al., 1991) впервые указаны из Северной Америки
P. cognatus и P. vulneratus, а также уточнены особенности внешнего строения и гени-
талий самцов, отличающие их от другого голарктического вида, P. unifasciatus, и неко-
торых похожих неарктических видов.
Polymerus cognatus и P. vulneratus (свекловичные клопы), относятся к числу вреди-
телей свеклы и бобовых культур (люцерны, эспарцета, клевера, гороха, фасоли и др.)
в Европе, особенно в ее восточной части (Пучков, 1966, 1972; Голуб, 1983). Они оби-
тают в природных ландшафтах и агроценозах совместно с другими видами этого рода
(Пучков, 1966), что затрудняет их быструю идентификацию и прогнозирование чис-
ленности. Перекрывание внутривидовой изменчивости морфологических признаков,
обычно использующихся для идентификации палеарктических видов, ограничивает
их пригодность для оперативного выявления вредителей. Наиболее надежно опреде-
ление с использованием признаков выдутого эдеагуса по методике Д. А. Гапона
(Gapon, 2001), но овладение ею требует длительной работы. Личинки ранних возрас-
тов, особенно видов подрода Poeciloscytus, достоверно не различаются, и таблиц для
их определения нет, но именно идентификация личинок ранних возрастов необходима
для своевременного выявления и прогнозирования численности вредных видов.
Первая попытка создания способа оперативной и точной идентификации вредного
вида клопов на основе комплекса морфологических и молекулярных признаков был
сделана нами для вредной черепашки Eurygaster integriceps Puton, 1881 и других
видов этого рода (Syromyatnikov et al., 2017). В настоящей статье представлены резуль-
таты попытки использования комплекса морфологических и молекулярных особен-
ностей для идентификации 5 видов рода Polymerus: 4 из подрода Poeciloscytus -
P. (P.) unifasciatus, P. (P.) brevicornis, P. (P.) vulneratus, P. (P.) cognatus - и P. (Pa-
chycentrum) unciniger. Кроме того, нами рассмотрена внутрипопуляционная изменчи-
вость морфологических признаков и нуклеотидных последовательностей гена COI
массового вида P. unifasciatus.
МАТЕРИАЛ И МЕТОДИКА
Материал для морфологических и молекулярных исследований был собран в 20 км СВ г. Во-
ронеж: Усманский бор, окр. биологического учебно-научного центра Воронежского государ-
ственного университета «Веневитиново», 51°49′ с. ш., 39°23′ в. д.; высота - 97-102 м над ур. м.
Всего исследовано 280 экз. Изученный материал хранится в коллекции кафедры экологии и си-
стематики беспозвоночных животных Воронежского государственного университета.
667
Морфологические признаки были исследованы у P. unifasciatus, P. brevicornis, P. vulneratus,
P. cognatus и P. unciniger, встречающихся обычно совместно в природных ландшафтах и в агро-
ценозах (в последних P. unciniger чаще отсутствует). Идентификация видов проводилась с помо-
щью определителей (Wagner, 1951, 1957; Кириченко, 1951; Кержнер, Ячевский, 1964; Rinne,
1989; Gapon, 2014) и проверена по коллекции Зоологического института РАН (С.-Петербург).
Морфологическая терминология принята по определителю И. М. Кержнера и Т. Л. Ячевского
(Кержнер, Ячевский, 1964), для структур эдеагуса - по работе Д. А. Гапона (Gapon, 2014).
Для проведения сравнительно-морфологического анализа были использованы традиционные
методы сбора и хранения энтомологического материала (Голуб и др., 2012). Снимки имаго кло-
пов сделаны камерой Canon методом послойного фотографирования на разной глубине резкости
под стереоскопическим микроскопом МБС-10; снимки эдеагусов сделаны под микроскопом
с использованием камеры DCM 800; все первичные фотографии скомбинированы с использова-
нием программы CombineZP, находящейся в свободном доступе, и отретушированы в програм-
ме Adobe Photoshop CS5. Рисунки парамеров сделаны с использованием рисовального аппарата
РА-6. Парамеры и эдеагус предварительно подвергались обработке в 4%-ном растворе KOH
(Кержнер, Ячевский, 1964; Голуб и др., 2012). При проведении морфометрического анализа
P. unifasciatus были использованы по 20 самцов и самок из указанного выше пункта (20 км СВ
Воронежа).
Для баркодинга ДНК были использованы те же виды, что и для проведения морфологического
анализа (P. unifasciatus, P. brevicornis, P. vulneratus, P. unciniger; рис. 1), кроме P. cognatus; чис-
ленность последнего вида в пунктах отбора проб при проведении исследований оказалась недо-
статочной для получения достоверных результатов. Собранные насекомые помещались
индивидуально в пробирки с 96%-ным этиловым спиртом и этикетировались. До проведения
анализов образцы хранились при температуре -20 °C для замедления деградации ДНК.
ДНК выделяли из ног и головы насекомых с помощью набора ZR Tissue & Insect DNA
MicroPrep (Zymo Research, США). ДНК из образцов исследованных видов использовали для
амплификации гена COI. Фрагменты гена длиной в 455 п. н. амплифицировали с помощью прай-
меров mHemF1/LepR1 и затем секвенировали. Полимеразную цепную реакцию проводили на
Eppendorf MasterCycler Personal Cycler (Eppendorf, Германия). Реакционная смесь для ПЦР со-
держала 5 мкл 5X ScreenMix-HS (Евроген, Россия), 1 мкл 10 мкМ прямого праймера, 1 мкл
10 мкМ обратного праймера, 2 мкл ДНК и 16 мкл деионизированной воды. Режим ПЦР включал
начальную денатурацию при 94 °C в течение 3 мин; 35 циклов: денатурация при 94 °C в течение
30 сек, отжиг праймеров при 51 °C в течение 30 сек, элонгация при 72 °C в течение 45 сек; ко-
нечная элонгация при 72 °C в течение 10 мин. При проведении ПЦР использовались следующие
праймеры: прямой mHemF1 (5ʹ-GCATTYCCACGAATAAATAAYATAAG) и обратный LepR1
(5ʹ-TAAACTTCTGGATGTCCAAAAAATCA) (Hebert et al., 2004; Wilson, 2012). Продукты ПЦР
разделяли электрофорезом в 2.5%-ном агарозном геле, окрашивали бромидом этидия и визуали-
зировали с помощью трансиллюминатора TCP-20LM. Размер продуктов ПЦР определяли с ис-
пользованием стандартов длин ДНК (Евроген, Россия).
Продукты ПЦР очищали от агарозного геля с помощью набора Cleanup Standard (Евроген,
Россия) и секвенировали с помощью генетического анализатора Applied Biosystems 3500 с ис-
пользованием BigDye Terminator v3.1 Kit. Для секвенировании использовали те же праймеры,
что и для баркодинга ДНК - mHemF1 и LepR1. Полученные нуклеотидные последовательности
для проверки на отсутствие стоп-кодонов и «пробелов» транслировали в аминокислотные по-
следовательности с помощью EMBOSS Transeq на сайте European Molecular Biology Laboratory’s
При проведении ПЦР-ПДРФ-анализа для видовой дифференциации были выбраны эндону-
клеазы рестрикции с использованием теоретических диаграмм расщепления ДНК, доступных
на сайте научно-производственного объединения
products/restrictases). ПЦР проводили с использованием праймеров LepF1 и mHemR1 (см. выше).
668
Рис. 1. Polymerus Hahn.
1 - P. unciniger Gapon, самец; 2, 3 - P. vulneratus (Panzer), самец (2 - вид сверху, 3 - вид сбоку);
4, 5 - P. unifasciatus (F.) (4 - самец, forma macroptera; 5 - самка, forma submacroptera); 6 - P. brevicornis
(Reuter), самец (Россия, 20 км СВ Воронежа, Усманский бор).
Шкала: 1 мм. (Ориг.).
Продукты ПЦР (10 мкл) расщепляли в реакционной смеси, содержащей 1.5 мкл 10Х реакцион-
ного буфера (СибЭнзим, Россия) и 10 ед. эндонуклеазы рестрикции (СибЭнзим, Россия) в об-
щем объеме 15 мкл (при необходимости объем до нужного значения доводили стерильной
водой). Смесь инкубировали в течение 2 ч при температуре +37 °C. Фермент инактивировали
при температуре +75 °C в течение 15 мин. После электрофореза продукты рестрикции визуали-
зировали окрашиванием бромидом этидия в 2.5%-ном агарозном геле.
Анализ степени филогенетической близости видов по молекулярным показателям проводился
с использованием программного обеспечения MEGA6 (Center for Evolutionary Medicine and
Informatics, США). Анализируемые последовательности ДНК были усечены до 412 пар нуклео-
669
тидов (п. н.). Парные генетические расстояния между образцами были рассчитаны с использова-
нием модели Kimura-2-Parameter (K2P) (Kimura, 1980). Реконструкция генетического дерева
была осуществлена с использованием метода ближайших соседей (Saitou, Nei, 1987). Процент
дублирующих деревьев, в которых таксоны сгруппированы в «bootstrap» тесте (500 повторов),
показаны рядом с ветвями (Felsenstein, 1985). Было проанализировано 27 нуклеотидных после-
довательностей. Реконструкция генетических деревьев была проведена в программе Molecular
Evolutionary Genetics Analysis 6 (MEGA6) (Tamura et al., 2013). В качестве внешней группы был
выбран Stenotus binotatus (Heteroptera, Miridae).
Эволюционное расхождение между видами было оценено как число нуклеотидных замен на
сайт, рассчитанное путем усреднения всех пар последовательностей между анализируемыми
группами. Анализ проводился с использованием модели K2P (Saitou, Nei, 1987).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Морфологические особенности имаго Polymerus (Poeciloscytus) и P. (Pachycentrum),
обитающих совместно в природных ландшафтах и агроэкосистемах восточноевропей-
ской лесостепи, приведены в определительной таблице. Длина члеников усиков, их
соотношение и другие пропорции тела, часто использующиеся для идентификации
видов, в ней не используются из-за перекрывания границ их внутривидовой изменчи-
вости; кроме того, у большинства коллекционных экземпляров 3-й и 4-й членики уси-
ков обломаны. Особенности окраски тела и его опушения пригодны для идентификации
только полностью окрепших и окрашенных имаго.
ТАБЛИЦА ДЛЯ РАЗЛИЧЕНИЯ POLYMERUS UNCINIGER, P. VULNERATUS, P. COGNATUS,
P. UNIFASCIATUS И P. BREVICORNIS
(по признакам, использованным в работах: Кержнер, Ячевский, 1964; Schwartz et al.,
1991; Gapon, 2014, с изменениями)
1(2). Переднеспинка, щиток и надкрылья одноцветно-черные, только кунеус часто
окаймлен узкой желтой полосой; задний край переднеспинки по бокам иногда
также с узкой желтой полосой. Левый (крупный, г-образно изогнутый) парамер
с явственно заостренной вершиной (рис. 2, 1). Медиальная спикула везики эдеа-
гуса головчато утолщенная вблизи вершины (рис. 3, 1). 4.6-5.0 мм. Рис. 1, 1
(подрод Pachycentrum)
P. unciniger.
2(1). Щиток и надкрылья не одноцветно-черные, а с крупными желтыми и черными
пятнами; щиток обычно почти весь желтый, иногда (у P. vulneratus) желтый цвет
преобладает и в окраске надкрылий. Левый парамер со слегка притупленной
вершиной (рис. 2, 2-5). Медиальная спикула везики постепенно утончается
к вершине, вблизи нее без головчатого утолщения (рис. 3, 2-5)
................................................................................................ (Подрод Poeciloscytus).
3(6). У полностью окрашенных особей с окрепшими после линьки на имаго покрова-
ми боковой край надкрылий по всей длине или на большей его части с очень
тонкой черной или буровато-черной линией (см. сбоку); у молодых особей тем-
ная линия часто заметна только в базальной половине надкрылья. Тело сверху
преимущественно в серебристых волосках.
4(5). Большая часть верхней стороны тела темная; большая часть надкрылий обычно
черная (у молодых, недоокрашенных особей черная окраска может быть слабо
развита). Передние углы переднеспинки у полностью окрашенных особей с чер-
ным, нередко слабо блестящим пятном. Кунеус нередко с буро-красным и чер-
670
Рис. 2. Polymerus Hahn, левый парамер.
1 - P. unciniger Gapon, 2 - P. cognatus (Fieber), 3 - P. vulneratus (Panzer), 4 - P. unifasciatus (F.),
5 - P. brevicornis (Reuter) (Россия, 20 км СВ Воронежа, Усманский бор).
Шкала: 0.2 мм. (Ориг.).
Рис. 3. Polymerus Hahn, везика эдеагуса.
1 - P. unciniger Gapon, 2 - P. cognatus (Fieber), 3 - P. vulneratus (Panzer), 4 - P. unifasciatus (F.),
5 - P. brevicornis. MS - медиальная спикула (Россия, 20 км СВ Воронежа).
Шкала: 0.2 мм. (Ориг.).
671
ным пятном. Левый парамер и везика - см. рис. 2, 2; 3, 2. 3.8-4.7 мм
................................................................................................................... P. cognatus.
5(4). Большая часть верхней стороны тела светлая, желтая или зеленовато-желтая;
надкрылья обычно почти целиком светлые, лишь с отдельными черными про-
дольными пятнами. Передние углы переднеспинки без черного пятна. Кунеус с
большим красным пятном и обычно с черной полоской по наружному краю. Ле-
вый парамер и везика - см. рис. 2, 3; 3, 3. 4.3-4.9 мм. Рис. 1, 2, 3
................................................................................................................ P. vulneratus.
6(3). Боковой край надкрылий без черной линии или она есть только в основании над-
крылий. Тело сверху преимущественно в желтоватых волосках.
7(8). Кунеус с большим или маленьким (нередко очень узким, в виде полоски) чер-
ным пятном, прилегающим к его наружному краю и не доходящим до внутрен-
него края, и с красным пятном, прилегающим к его внутреннему краю, часто не
полностью окружающим черное пятно; иногда кунеус почти полностью черный.
Левый парамер и везика эдеагуса - см. рис. 2, 4; 3, 4. 4.3-4.9 мм. Рис. 1, 4, 5
............................................................................................................. P. unifasciatus.
8(7). Кунеус с черным пятном, доходящим до его внутреннего края, и с желтым или
красноватым окаймлением черного пятна. Левый парамер и везика эдеагуса -
см. рис. 2, 5; 3, 5. 4.2-4.8 мм. Рис. 1, 6
P. brevicornis.
При разработке метода ПЦР-ПДРФ для быстрой идентификации вредного вида
P. vulneratus установлено, что специфичные для этого вида фрагменты ДНК образуют-
ся под действием семи рестриктаз (AcsI, BstF5I, FokI, PpsI, RsaI, RsaNI и SspMI). Для
дальнейших лабораторных процедур была выбрана рестриктаза RsaI, поскольку она
имеет сайт рестрикции у всех исследуемых видов, но только у P. vulneratus образуется
фрагмент размером 131 п. н., который хорошо регистрируется в 2.5%-ном агарозном
геле при проведении электрофореза. Электрофореграмма продуктов ПЦР с праймера-
ми mHemF и LepR1, обработанных рестриктазой RsaI, представлена на рис. 4.
Рис. 4. Электрофореграмма продуктов рестрикции RsaI.
K - продукт ПЦР до рестрикции, R - после рестрикции.
672
Таким образом, обработка рестриктазой RsaI продукта ПЦР, полученного с помо-
щью праймеров mHemF1/LepR1, обеспечивает достоверную идентификацию
P. vulneratus.
Нуклеотидные последовательности, полученные нами секвенированием гена COI
у исследованных видов, были зарегистрированы в системе GenBank под следующими
номерами: P. unciniger - MH752746-MH752750; P. vulneratus - MH607428, MH607436;
P. unifasciatus - MH607420-MH607427, MH607440, MH607441; P. brevicornis -
MH607415-MH607419. Баркодинг ДНК P. brevicornis и P. unciniger был выполнен
впервые.
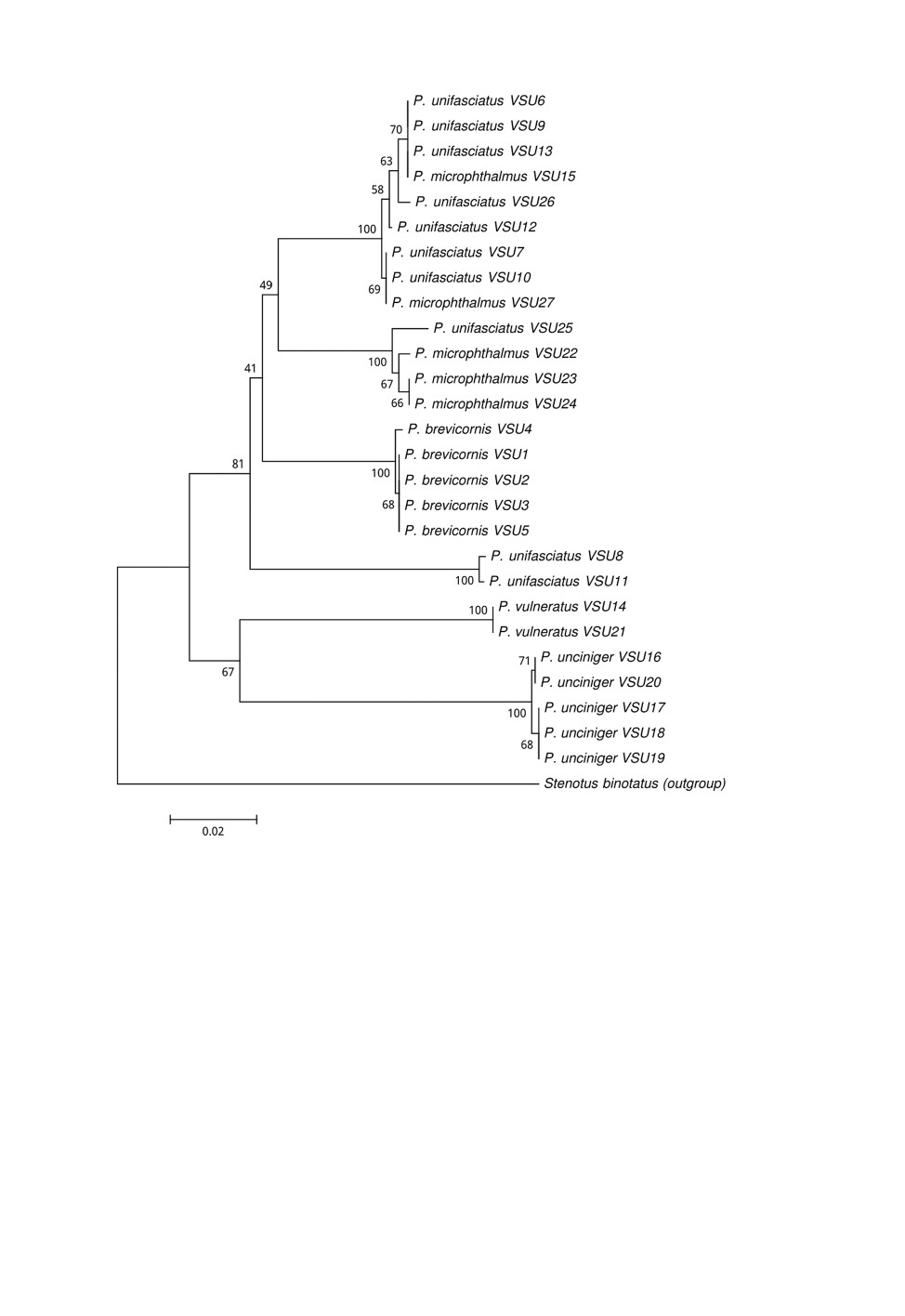
Генетические расстояния между изученными видами Polymerus на основе получен-
ных нами данных о последовательности генов COI (VSU («Voronezh State University»);
пункт отбора проб: Россия, 20 км СВ Воронежа) и с использованием данных в системе
NCBI GenBank представлены соответственно на рис. 5 и 6.
Результаты секвенирования гена COI показали значительную дистанцированность
кластера, включающего два вида подрода Pachycentrum (P. (P.) nigrita и P. (P.) un-
ciniger), от всех видов подрода Poeciloscytus (рис. 5, 6). Морфологические различия
между двумя подродами по окраске тела и признакам гениталий (Gapon, 2014; см. та-
блицу для определения видов и рис. 1, 1-5; 2, 1-5) также более значительны, чем раз-
личия между видами подрода Poeciloscytus. Таким образом, комплекс морфологических
и молекулярных различий обеспечивает возможность точной идентификации двух
подродов и, соответственно, видов подрода Pachycentrum на стадиях имаго и личинок.
Морфологические различия между видами подрода Poeciloscytus не всегда обеспе-
чивают точную идентификацию видов. На основе материала из восточноевропейской
лесостепи (Воронежская обл.) и c других территорий России и сопредельных стран
(материал ЗИН) установлено, что наличие тонкой темной линии на боковом крае над-
крылий P. vulneratus и P. cognatus проявляется только у полностью окрепших и окра-
шенных особей. У недавно линявших на имаго особей темная линия обычно слабо
выражена и присутствует только в базальной части надкрылий. Напротив, боковой
край надкрылий у особей P. unifasciatus в базальной половине часто с темной линией,
как у P. cognatus и P. vulneratus. Сравнительный анализ особенностей везики позволя-
ет констатировать, что ее медиальные спикулы у P. cognatus, P. vulneratus и P. brevi-
cornis (рис. 3, 2, 5), различаются почти исключительно степенью изогнутости. Однако
результаты секвенирования гена COI показали отчетливость кластера P. vulneratus
и P. cognatus (рис. 5, 6): его генетическая дистанцированность от других изученных
нами видов Polymerus составила не менее 0.123, что обеспечивает надежную иденти-
фикацию этой пары видов среди встречающихся совместно с ними.
Медиальная спикула P. unifasciatus длиннее, чем у других изученных видов подрода
Poeciloscytus (рис. 3, 4), форма ее слабо изменчива. По результатам секвенирования
гена COI этот вид, с учетом внутривидовой вариабельности, примерно равноудален от
других видов рода (рис. 5, 6). Таким образом, комплекс признаков P. unifasciatus,
включающих особенности строения эдеагуса и сиквенса гена COI, также обеспечива-
ет его надежную идентификацию.
Некоторые нуклеотидные последовательности, полученные нами (MH607429,
MH607437, MH607438, MH607439, MH607442), соответствуют таковым P. micro-
phthalmus (Wagner, 1951), зарегистрированным прежде в базе данных GenBank. Одна-
673
Рис. 5. Кладограмма нуклеотидных последовательностей гена COI видов рода Polymerus Hahn,
полученных авторами.
ко различия между нуклеотидными последовательностями гена COI P. unifasciatus
и P. microphthalmus настолько малы, что эти таксоны не могут быть достоверно диф-
ференцированы классическим методом баркодинга ДНК (см. рис. 5, 6). Очень высокая
вариабельность нуклеотидных последовательностей гена COI у P. unifasciatus (0.0519)
и P. microphthalmus (0.0341) (рис. 5, 6) не позволяет различить их по этому признаку.
При этом нуклеотидная вариабельность P. unciniger не превышала 0.001, а у P. brevi-
cornis и P. vulneratus она вообще отсутствовала (рис. 5, 6).
В связи с тем, что P. microphthalmus прежде не указывался из европейской части
России, мы провели морфометрический анализ внутрипопуляционной изменчивости
признаков P. unifasciatus, которые наиболее часто использовались ранее для диффе-
ренциации его от P. microphthalmus (табл. 1).
674
Рис. 6. Кладограмма нуклеотидных последовательностей гена COI видов рода Polymerus Hahn,
полученных авторами и депонированных в системе NCBI GenBank.
675
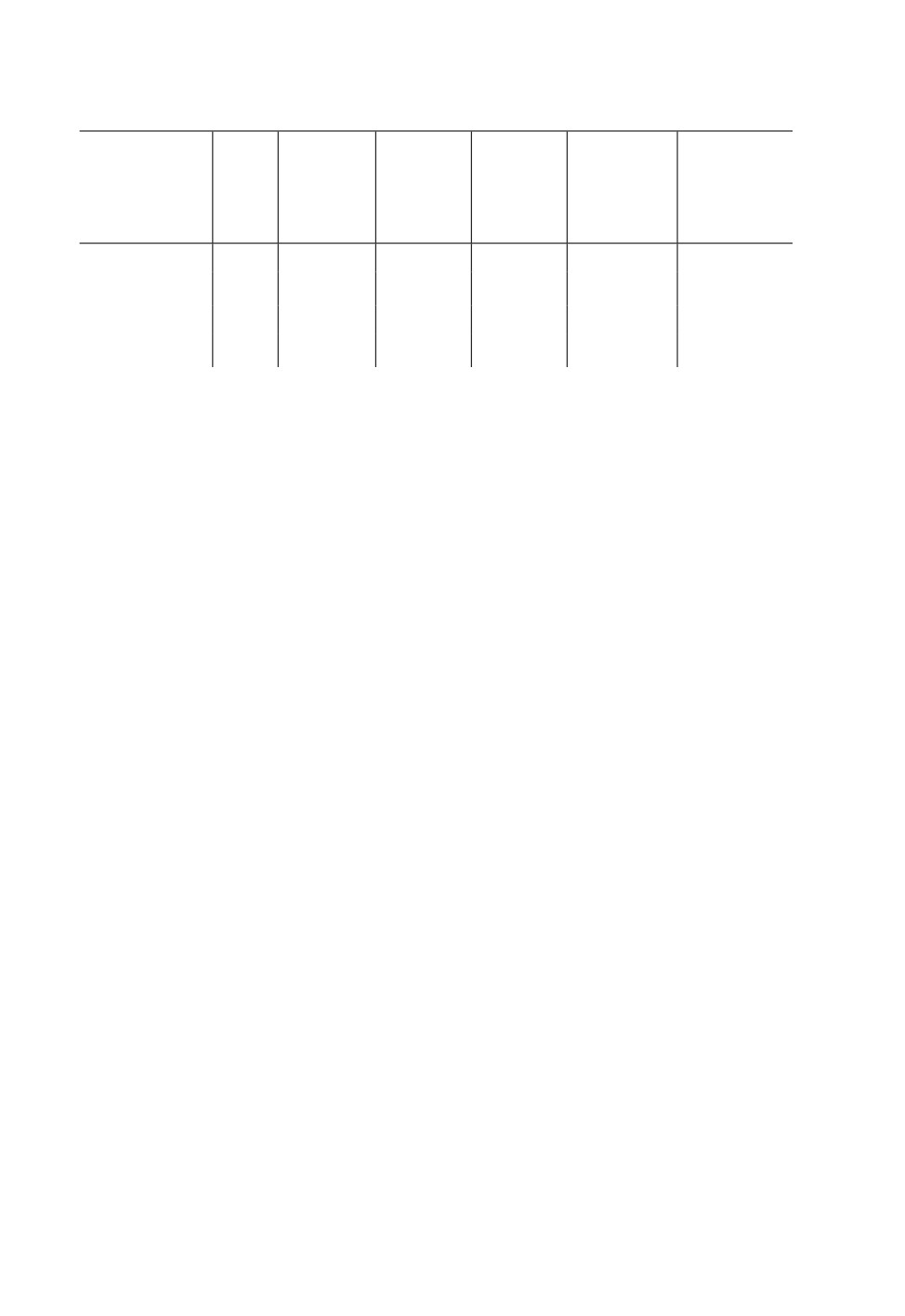
Таблица 1. Морфометрические показатели Polymerus unifasciatus (F.) (20 км СВ Воронежа,
Усманский бор, 2016-2018 гг.)
Число
Отношение
Отношение
изучен-
ширины
Длина тела,
Ширина
Ширина
ширины
Пол, форма
ных
темени
мм
темени, мм
глаза, мм
головы
экземп-
к ширине
к ее высоте
ляров
глаза
Самцы,
21
5.50-7.60
0.40-0.50
0.26-0.35
1.14-1.93
1.05-1.30
f. macroptera
5.9 ± 0.123
0.43 ± 0.006
0.30 ± 0.005
1.43 ± 0.046
1.17 ± 0.015
Самки,
20
4.50-6.10
0.47-0.57
0.25-0.33
1.58-2.12
1.10-1.28
f. submacroptera
5.23 ± 0.096
0.51 ± 0.006
0.28 ± 0.005
1.86 ± 0.032
1.22 ± 0.011
Примечание. Над чертой - минимальное и максимальное значения признака, под чертой - среднее
значение и ошибка.
Кроме того, изучено отношение длин III и IV члеников усиков у особей с полностью
сохранившимися усиками (8 самцов и 8 самок), которое составило у самцов 0.96-1.0,
у самок - 0.9-1.0.
Установлено, что значения всех включенных в табл. 1 показателей полнокрылых
самцов (forma macroptera) полностью соответствуют таковым P. unifasciatus, приве-
денным Э. Вагнером при описании P. microphthalmus и в более поздних публикациях
(Wagner, 1951, 1961, 1971), а также дополненным последующими авторами (Rinne,
1989; Herczek, 1991). В отличие от самцов, у исследованных самок перепоночка над-
крылий слегка или заметно укорочена (forma submacroptera). При этом теменная часть
головы у них шире, а глаза меньше, чем у самцов (табл. 1), что соответствует описан-
ным Э. Вагнером признакам P. microphthalmus (Wagner, 1951, 1961, 1971). Особенно-
сти парамеров и спикул эдеагуса (включая медиальную), которые указывали бы на
присутствие в изученной выборке двух видов, не выявлены.
Полученные данные об изменчивости морфологических и молекулярно-генетиче-
ских показателей свидетельствуют о том, что мы, вероятнее всего, имеем дело с одним
видом, P. unifasciatus, с довольно широкой изменчивостью размеров тела и пропорций
его частей. Эти различия усиливаются половым диморфизмом с проявлением у самок
укороченности надкрылий и, несомненно, коррелирующими с ней меньшими размера-
ми глаз и, соответственно, большей шириной темени. Высокая морфологическая из-
менчивость (табл. 1) и половой диморфизм P. unifasciatus, очевидно, обусловлены
значительной генотипической изменчивостью, затрагивающей и ген COI (рис. 5, 6).
Окончательно вопрос о самостоятельности P. microphthalmus может быть решен на ос-
новании результатов изучения его межпопуляционных различий и детального изуче-
ния эдеагуса в его различных проекциях.
ЗАКЛЮЧЕНИЕ
1. Совместно обитающие в природных ландшафтах и агроценозах восточноевропей-
ской лесостепи Polymerus unifasciatus, P. cognatus, P. vulneratus, P. brevicornis и P. un-
ciniger могут достоверно идентифицироваться по разработанному нами комплексу
особенностей внешнего строения, гениталий и результатам баркодинга ДНК.
676
2. Почти полностью черная окраска тела (см. рис. 1, 1), острая вершина левого пара-
мера (рис. 2, 1) и головчато утолщенная близ вершины медиальная спикула везики
(рис. 3, 1) P. (Pachycentrum) unciniger в сочетании с обособленностью кластера подро-
да Pachycentrum в кладограмме сиквенсов гена COI изученных таксонов (рис. 6)
обеспечивают возможность точной идентификации двух подродов Polymerus.
3. Разработанный нами ПЦР-ПДРФ-метод быстрой и надежной идентификации вре-
дителя сельскохозяйственных культур P. vulneratus заключается в разрезании ампли-
фицированных фрагментов ДНК эндонуклеазой RsaI, в результате чего формируется
уникальная картина рестрикции (см. рис. 4).
4. Polymerus (Poeciloscytus) unifasciatus характеризуется наиболее длинной медиаль-
ной спикулой везики среди изученных видов этого подрода (рис. 3, 4), что в сочетании
с данными баркодинга ДНК (рис. 5, 6) позволяет достоверно определять этот мас-
совый вид.
5. Внутрипопуляционная изменчивость признаков строения головы P. unifasciatus
связана с проявлением полового диморфизма, при котором у особей (чаще - самок)
с укороченными надкрыльями глаза немного меньше, а ширина темени, соответствен-
но, больше, чем у полнокрылых особей, обычно самцов. Высокая внутрипопуляцион-
ная изменчивость внешних морфологических признаков P. unifasciatus сопутствует
очень высокой изменчивости нуклеотидной последовательности гена COI у форм,
представленных в базе данных GenBank под названиями P. unifasciatus и P. micro-
phthalmus. Не исключено поэтому существование всего одного вида, P. unifasciatus,
крайние проявления генетической и морфологической изменчивости которого отно-
сятся авторами к разным видам - P. unifasciatus и P. microphthalmus.
6. Впервые выполненный баркодинг ДНК P. brevicornis и P. unciniger расширяет
представления о межвидовых различиях в нуклеотидной последовательности гена
COI в роде Polymerus. Полученные авторами данные о нуклеотидных последователь-
ностях COI всех исследованных видов Polymerus зарегистрированы в базе данных
GenBank.
ФИНАНСИРОВАНИЕ
Работа выполнена при финансовой поддержке Российского фонда фундаменталь-
ных исследований (грант № 18-04-00464-а) и гранта президента для ведущих научных
школ (соглашение НШ-2535.2020.11).
СПИСОК ЛИТЕРАТУРЫ
Голуб В. Б. 1983. Отряд Полужесткокрылые, или Клопы, - Hemiptera. В кн.: Л. М. Копанева (со-
ставитель). Определитель вредных и полезных насекомых и клещей однолетних и много-
летних трав и зернобобовых культур в СССР. Л.: Колос, Ленинградское отделение,
с. 73-84.
Голуб В. Б., Цуриков М. Н., Прокин А. А. 2012. Коллекции насекомых: сбор, обработка и хране-
ние материала. М.: Товарищество научных изданий КМК, 334 с.
Кержнер И. М., Ячевский Т. Л. 1964. Отряд Hemiptera (Heteroptera) - полужесткокрылые, или
клопы. В кн.: Г. Я. Бей-Биенко (ред.). Определитель насекомых европейской части СССР
в 5 томах. Т. 1. М.; Л.: Наука, с. 655-845.
Кириченко А. Н. 1951. Настоящие полужесткокрылые европейской части СССР (Hemiptera).
Определитель и библиография. М.; Л.: Издательство АН СССР, 423 с.
677
Пучков В. Г. 1966. Главнейшие клопы-слепняки - вредители сельскохозяйственных культур.
Киев: Наукова думка, 172 с.
Пучков В. Г. 1972. Отряд Hemiptera (Heteroptera) - полужесткокрылые. В кн.: О. Л. Крыжанов-
ский, Е. М. Данциг (ред.). Насекомые и клещи - вредители сельскохозяйственных культур.
Т. 3. Л.: Наука, с. 222-262.
Carvalho J. C. M., Leston D. 1952. The classification of the British Miridae (Hem.), with keys to the
genera. Entomologist’s Monthly Magazine 88: 231-251.
Felsenstein J. 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39
(4): 783-791.
Gapon D. A. 2001. Inflation of heteropteran aedeagi using microcapillaries (Heteroptera: Pentatomidae).
Zoosystematica Rossica 9 (1): 157-160.
Gapon D. A. 2014. Revision of the genus Polymerus (Heteroptera: Miridae) in the Eastern Hemisphere.
Part 1: Subgenera Polymerus, Pachycentrum subgen. nov. and new genus Dichelocentrum gen.
nov. Zootaxa 3787 (1): 1-87.
Hebert P. D., Penton E. H., Burns J. M., Janzen D. H., Hallwachs W. 2004. Ten species in one: DNA
barcoding reveals cryptic diversity in the Neotropical skipper butterfly Astraptes fulgerator.
Proceedings of the National Academy of Sciences of the United States of America 101 (41):
14812-14817.
Herczek A. 1991. Notes on three species of Polymerus Westwood (Heteroptera, Miridae). Polskie
Pismo Entomologiczne 61: 23-29.
Kerzhner I. M., Josifov M. 1999. Family Miridae Hahn, 1833. In: B. Aukema, Ch. Rieger (eds).
Catalogue of the Heteroptera of the Palaearctic Region. Vol. 3. Cimicomorpha II. Amsterdam,
The Netherlands: The Netherlands Entomological Society, 577 p.
Kimura M. 1980. A simple method for estimating evolutionary rate of base substitutions through
comparative studies of nucleotide sequences. Journal of Molecular Evolution 16: 111-120.
Rinne V. 1989. Review of the European Polymerus subgenus Poeciloscytus (Heteroptera, Miridae),
with two new species and special reference to the Finnish fauna. Annales Entomologici Fennici
55: 89-101.
Saitou N., Nei M. 1987. The neighbor-joining method: A new method for reconstructing phylogenetic
trees. Molecular Biology and Evolution 4: 406-425.
Schwartz M. D., Scudder G. E., Henry T. J. 1991.The first Nearctic records of two Holarctic species of
Polymerus Hahn, with remarks on a monophyletic species-group (Heteroptera: Miridae: Mirinae).
Canadian Entomologist 123 (4): 721-743.
Syromyatnikov M. Y., Golub V. B., Kokina A. V., Soboleva V. A., Popov N. V. 2017. DNA barcoding
and morphological analysis for rapid identification of most economically important crop-
infesting Sunn pests belonging to Eurygaster Laporte, 1833 (Hemiptera, Scutelleridae). ZooKeys
706: 51-71.
Tamura K., Stecher G., Peterson D., Filipski A., Kumar S. 2013. MEGA6: Molecular Evolutionary
Genetics Analysis version 6.0. Molecular Biology and Evolution 30 (12): 2725-2729.
Wagner E. 1951. On the genus Poeciloscytus Fieb. (Hem. Het., Miridae). Acta Entomologica Musei
Nationalis Pragae 26 (351): 1-8.
Wagner E. 1961. Unterordnung: Ungleichflügler, Wanzen, Heteroptera (Hemiptera). In: Die Tierwelt
Mitteleuropas, Vol. 4, p. 1-173.
Wagner E. 1971. Die Miridae Hahn, 1831, des Mittelmeerraumes und der Makaronesischen Inseln
(Hemiptera, Heteroptera). Teil 1. Entomologische Abhandlungen Staatliches Museum für
Tierkunde Dresden 37, suppl., 484 p.
Wilson J. J. 2012. DNA barcodes for insects. In: W. J. Kress, D. L. Erickson (eds). Methods in Molecular
Biology (Methods and Protocols), vol. 858. Totowa, NJ: Humana Press, p. 17-46.
678
DIAGNOSTICS OF A GENUS POLYMERUS HAHN (HETEROPTERA, MIRIDAE)
SPECIES GROUP INCLUDING AGRICULTURAL PESTS IN THE EAST
EUROPEAN FOREST-STEPPE, BASED ON THE MORPHOLOGICAL
AND MOLECULAR CHARACTERS
V. B. Golub, M. Yu. Syromyatnikov, A. V. Kokina, V. A. Soboleva,
E. V. Nesterova, V. N. Popov
Key words: Polymerus, species, pests, key, DNA barcoding, PCR-RFLP method, identifi-
cation.
SUMMARY
The paper presents an improved key to the co-occurring species of Polymerus Hahn, 1831
(Heteroptera, Miridae) based on morphological characters and authors’ data on the differences in
the DNA sequences between the COI gene of the same species. The complex of these characters,
together with the data stored in the NCBI GenBank, enables accurate and rapid identification of
imagoes and larvae of Polymerus unciniger Gapon, 2014, P. unifasciatus (Fabricius, 1794), P. cognatus
(Fieber, 1858), P. vulneratus (Panzer, 1806) and P. brevicornis (Reuter, 1879). Correspondence of
the differences between the subgenera Poeciloscytus Fieber, 1858 and Pachycentrum Gapon, 2014
in the morphological and molecular genetic characteristics was established. A PCR-RFLP method for
rapid identification of the pests, P. vulneratus and P. cognatus, was developed. High intrapopulation
variability of morphological characters of P. unifasciatus and the DNA sequences of the COI gene in the
forms registered in GenBank as P. unifasciatus and P. microphthalmus (Wagner, 1951) was revealed. It
has been suggested that P. microphthalmus is a submacropterous form of P. unifasciatus with shortened
hemelytra and smaller eyes, in comparison with the macropterous form of the same species, whose
individuals have larger eyes.
679