ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 99, 4, 2020
УДК 595.728
СЕМЕЙСТВА STENOPELMATIDAE И ANOSTOSTOMATIDAE
(ORTHOPTERA). 1. НАДРОДОВАЯ КЛАССИФИКАЦИЯ,
НОВЫЕ И МАЛОИЗВЕСТНЫЕ ТАКСОНЫ
© 2020 г. А. В. Горохов
Зоологический институт РАН
Университетская наб., 1, Санкт-Петербург, 199034 Россия
e-mail: orthopt@zin.ru
Поступила 28.08.2020 г.
После доработки 2.09.2020 г.
Принята к публикации 2.09.2020 г.
Семейство Stenopelmatidae подразделено лишь на 3 подсемейства: Stenopelmatinae с 4 или
5 трибами (Stenopelmatini, Siini stat. resurr., Maxentiini trib. n., Oryctopterini trib. resurr. и, воз-
можно, Oryctopini), Schizodactylinae (= Comicinae Ander, 1939, syn. n.), не разделенное на трибы,
и Gryllacridinae (= Hyperbaeninae Cadena-Castañeda, 2019, syn. n.) с проблематичным составом
триб. Описаны следующие новые таксоны: Sia bugajus sp. n.; Amaxentius subgen. n. и Eumaxentius
subgen. n. в составе рода Maxentius Stål, 1876, stat. resurr.; Comicus orangensis sp. n.; C. myburghi
sp. n. и C. namibicus sp. n. Роды Oryctopterus Karny, 1937, gen. resurr. и Brauckmannia Martins-
Neto, 2007, gen. resurr. восстановлены из синонимов Oryctopus Brunner-Wattenwyl, 1888 и
Schizodactylus Brullé, 1835 соответственно. Семейство Anostostomatidae включает только 2 под-
семейства: Anostostomatinae с 9 трибами (Anostostomatini, Anabropsini, Lutosini; Cratomelini stat.
n., Brachyporini, Deinacridini stat. n., Cooloolini stat. n., Leiomelini stat. n., Glaphyrosomatini) и,
возможно, Lezininae, не разделенное на трибы. Род Anabropsis Rehn, 1901 считается состоящим
из 5 подродов: Anabropsis s. str.; Paterdecolyus Griffini, 1913, stat. n.; Pteranabropsis Gorochov,
1988, stat. n. (= Brevipenna Shi et Bian, 2016, syn. n.); Apteranabropsis Gorochov, 1988, stat. n.;
Carnabropsis subgen. n. Предложены новые комбинации для 20 видов этого рода и описаны сле-
дующие новые виды и подвиды: Apteranabropsis (Pteranabropsis) intermedia sp. n., A. (P.) falcata
pallescens subsp. n., A. (Apteranabropsis) ailaoshanica sp. n., A. (A.) maculata sp. n., A. (A.) tarasovi
sp. n., A. (A.) abramovi sp. n., A. (A.) abramovi namlikensis subsp. n. Описаны ранее неизвестные
самец Anabropsis (Аpteranabropsis) minuta (Gorochov, 2001) и самка A. (A.) sinica Bey-Bienko,
1962. Описаны также Proctovitettix subgen. n. из рода Glaphyrosoma Brunner-Wattenwyl, 1888
и Spinolezina gen. n. для S. obscura (Burr, 1900), comb. n. из Lezininae; род Gryllacropsis Brunner-
Wattenwyl, 1888 условно включен в последнее подсемейство. Приведены определительные
таблицы для ряда таксонов и новые данные по распространению некоторых видов.
Ключевые слова: прямокрылые, систематика, новые таксоны, номенклатурные изменения,
распространение.
DOI: 10.31857/S0367144520040073
905
Данное сообщение посвящено вопросам надродовой классификации семейств
Stenopelmatidae и Anostostomatidae, а также следующим родам из этих семейств: Sia
Giebel, 1861; Maxentius Stål, 1876; Comicus Brunner-Wattenwyl, 1888; Anabropsis Rehn,
1901; Glaphyrosoma Brunner-Wattenwyl, 1888; Gryllacropsis Brunner-Wattenwyl, 1888;
Spinolezina gen. n. Материал, рассмотренный в данной статье, хранится главным об-
разом в Зоологическом институте РАН, Санкт-Петербург [ЗИН], но некоторые экзем-
пляры - в следующих учреждениях: Музей и Институт зоологии Польской Академии
наук (Muzeum i Instytut Zoologii PAN), Варшава [МИЗ]; Институт зоологии Китайской
Академии наук (Institute of Zoology, CAS), Пекин [ИЗК].
В течение длительного времени вышеназванные семейства вместе с сем. Rha-
phidophoridae обычно рассматривались как родственные и иногда объединялись в
одно семейство или надсемейство (Brunner-Wattenwyl, 1888; Karny, 1937; Gorochov,
1984), но мнения об их филогенетических отношениях были весьма различны - они то
помещались в основание ствола подотряда Ensifera (Zeuner, 1939) или инфраотряда
Tettigoniidea (Ander, 1939), то сближались с надсем. Hagloidea и инфраотрядом
Gryllidea (Шаров, 1968). В работах Горохова (1988а, 1988б, 1995а), ставших в неко-
тором роде завершающим этапом построения филогении группы на основе нефор-
мальных методов морфологической филогенетики, эти три семейства были объеди-
нены в голофилетическое надсем. Stenopelmatoidea, которое считалось родственным
голофилетическому надсем. Tettigonioidea, и оба этих надсемейства выводились из
одной и той же группы парафилетического надсем. Hagloidea (все эти три надсемей-
ства включались им в особый инфраотряд Tettigoniidea). Эта классификация хорошо
согласовывалась с палеонтологическим материалом, что давало основание считать ее
более или менее обоснованной, в отличие от классификации более низких таксонов
(подсемейств, триб и др.; Gorochov, 2001а).
Однако одновременно с этими работами начали появляться публикации на основе
формальных методов морфологической (Gwynne, 1995; Desutter-Grandcolas, 2003) и
молекулярной (Flook et al., 1999; Jost, Shaw, 2006) филогенетики, в кладограммах ко-
торых обнаружились существенные разногласия. В двух первых из них Steno-
pelmatoidea оказались как минимум парафилетическими и попали в основание инфра-
отряда Tettigonioidea, причем одно из подсемейств Stenopelmatidae (Schizodactylinae)
то сближалось с инфраотрядом Gryllidea, то оказывалось между Stenopelmatidae и
Rhaphidophoridae. Причиной таких разногласий могло бы быть стремление авторов
набрать побольше морфологических признаков для кладистического анализа, но фи-
логенетически обоснованных синапоморфий для давно обособившихся групп не-
много, и авторам пришлось вовлечь в анализ множество слабых признаков, которые
легко могли затушевать немногочисленные истинные синапоморфии анализируемых
таксонов. В отношении молекулярных признаков надежд было больше, поскольку
ожидалось, что гомоплазии среди таких признаков не должны быть многочисленны.
Однако установление полярности для молекулярных признаков еще сложнее, чем для
морфологических, поскольку палеонтологический тест и эволюционно-логические
критерии здесь пока практически неприменимы, в связи с чем синапоморфии могут
быть легко спутаны с симплезиоморфиями. Возможно, это и было причиной сбли-
жения Rhaphidophoridae, Schizodactylinae и Gryllidea, лишенных надежных общих
морфопризнаков, и помещения Tettigonioidea в основание общего ствола этих групп,
т. е. между ними и остальными таксонами Stenopelmatoidea (Jost, Shaw, 2006). Тем не
менее, в еще одной из перечисленных публикаций (Flook et al., 1999) кладограмма
906
Ensifera оказалась похожей на филогенетическую схему Горохова, разработанную по
данным морфологии (стоит отметить, что молекулярная кладограмма для подотряда
Caelifera из той же статьи также оказалась похожей на филогенетическую схему этой
группы, предложенную на основе неформальных методов морфологической фило-
генетики; Горохов, 1995б).
В последнее десятилетие количество опубликованных молекулярных кладограмм
заметно возросло, но, вероятно, по рассмотренным выше причинам разногласия
между ними уменьшились незначительно. В работах некоторых авторов (Sheffield
et al., 2010; Zhang et al., 2013; Zhou et al., 2014; Song et al., 2015) Rhaphidophoridae, а в
последней работе и другие группы Stenopelmatoidea (включая Schizodactylinae), сбли-
жаются не с Gryllidea, а с Tettigoniidea (как у Горохова), но Rhaphidophoridae распола-
гаются между Schizodactylinae и остальными Stenopelmatoidea (причем ближе к по-
следним). У других же авторов Rhaphidophoridae и Schizodactylinae оказываются
синопоморфными относительно других групп Ensifera и сближаются с Gryllidea
(Vandergast et al., 2017), либо два близких рода Schizodactylinae помещаются в разные
инфраотряды (в Tettigoniidea среди Rhaphidophoridae и в Gryllidea среди Myrmeco-
philidae; Chintauan-Marquier et al., 2016). Последнее предположение несомненно оши-
бочно (Gorochov, 2019), и близость этих родов будет продемонстрирована ниже.
Таким образом, следует признать, что молекулярная филогенетика медленно и му-
чительно подтверждает основные выводы по филогении Ensifera, полученные много
лет назад на основе неформального анализа морфологии. В ряде случаев, например, в
отношении Schizodactylinae и Rhaphidophoridae, путаница продолжается, и ее причи-
нами могут оказаться 1) утрата очень старой и обособленной группой (Schizodactylinae)
большей части молекулярных синапоморфий с ближайшими родственниками и 2) не-
точное определение филогенетического положения для полностью бескрылого (т. е.
утратившего целый набор важных морфологических признаков) сем. Rhaphidophoridae.
Поэтому я считаю необходимым обратить внимание коллег на то, что наружные по-
ловые органы (включая гениталии самца) и сперматофор в надсем. Stenopelmatoidea
очень недостаточно изучены, и что подробное исследование их строения может про-
лить дополнительный свет как на процесс эволюции этих органов, так и на филогене-
тическое положение главных групп Stenopelmatoidea.
Некоторые интересные в этом отношении особенности строения таких органов рас-
сматриваются здесь. Гениталии самцов инфраотряда Tettigoniidea обычно полностью
мембранозные и довольно простые по строению, состоят из дорсальной и вентральной
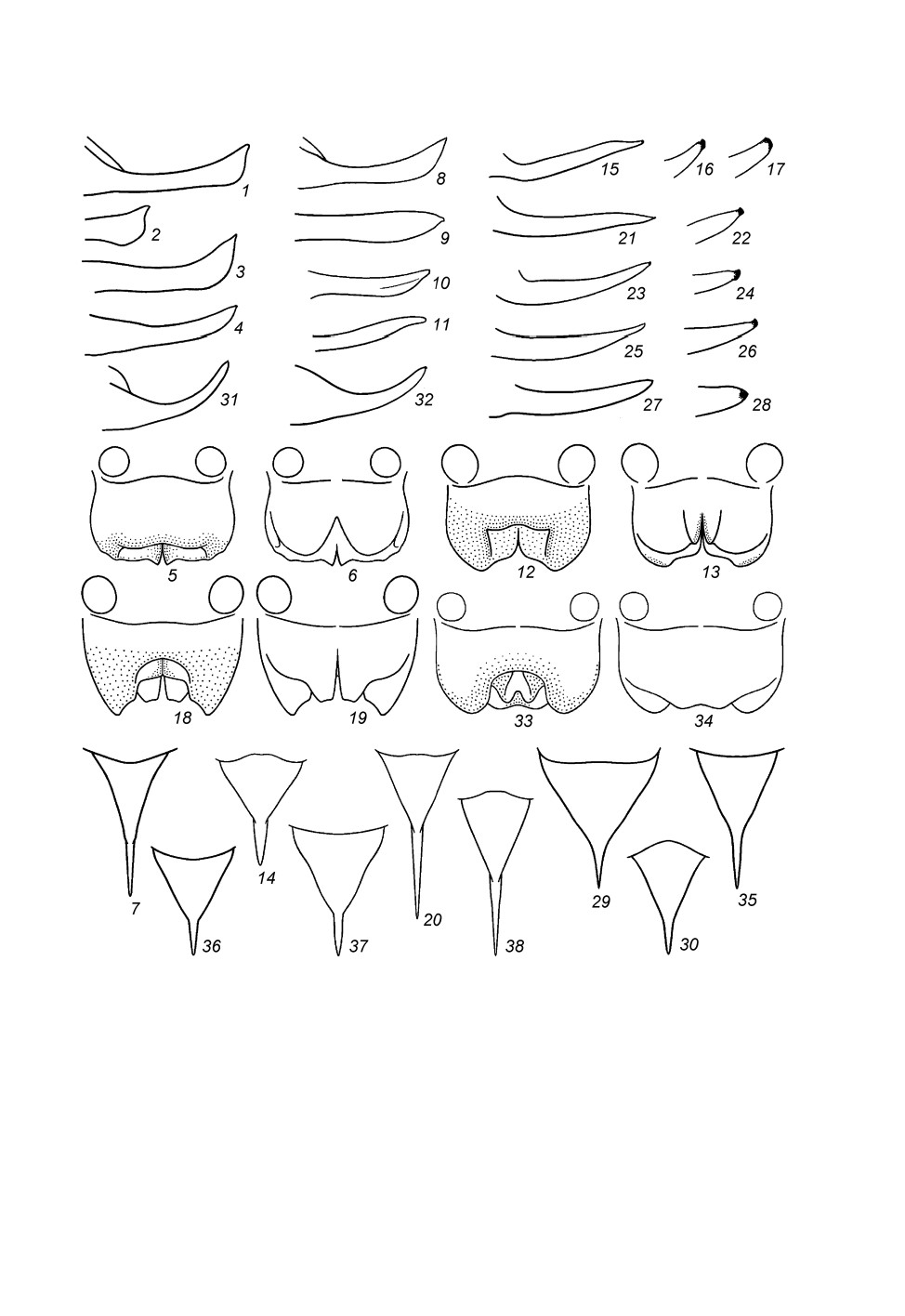
складок, часто подразделенных на несколько лопастей (рис. 1, 1-12, 14, 15, 21, 22),
причем если дорсальная складка с лопастями, то их обыкновенно 3 (срединная и
1 пара боковых; рис. 1, 1-8, 10-12, 21, 22). По литературным данным, сперматофор
Tettigonioidea, Stenopelmatidae и Anostostomatidae, а также современных представи-
телей Hagloidea отличается от такового у Rhaphidophoridae, Schizodactylidae и Gryl-
loidea наличием пары содержащих сперму капсул (Gwynne, 1995), т. е. ампула такого
сперматофора двойная, состоит из 1 пары флаконов, частично или почти полностью
слитых один с другим (Болдырев, 1914; Weissman, 2001: fig. 19.10). Но для некоторых
Anostostomatidae (Gwynne, 1995; Stringer, 2001) и Rhaphidophoridae (Болдырев, 1914)
отмечено наличие одиночной ампулы c остатком перегородки, частично отделяющим
одну капсулу со спермой от другой и указывающим на то, что у их предков ампула
была или могла быть двойной. Следовательно, отсутствие следов этой перегородки у
907
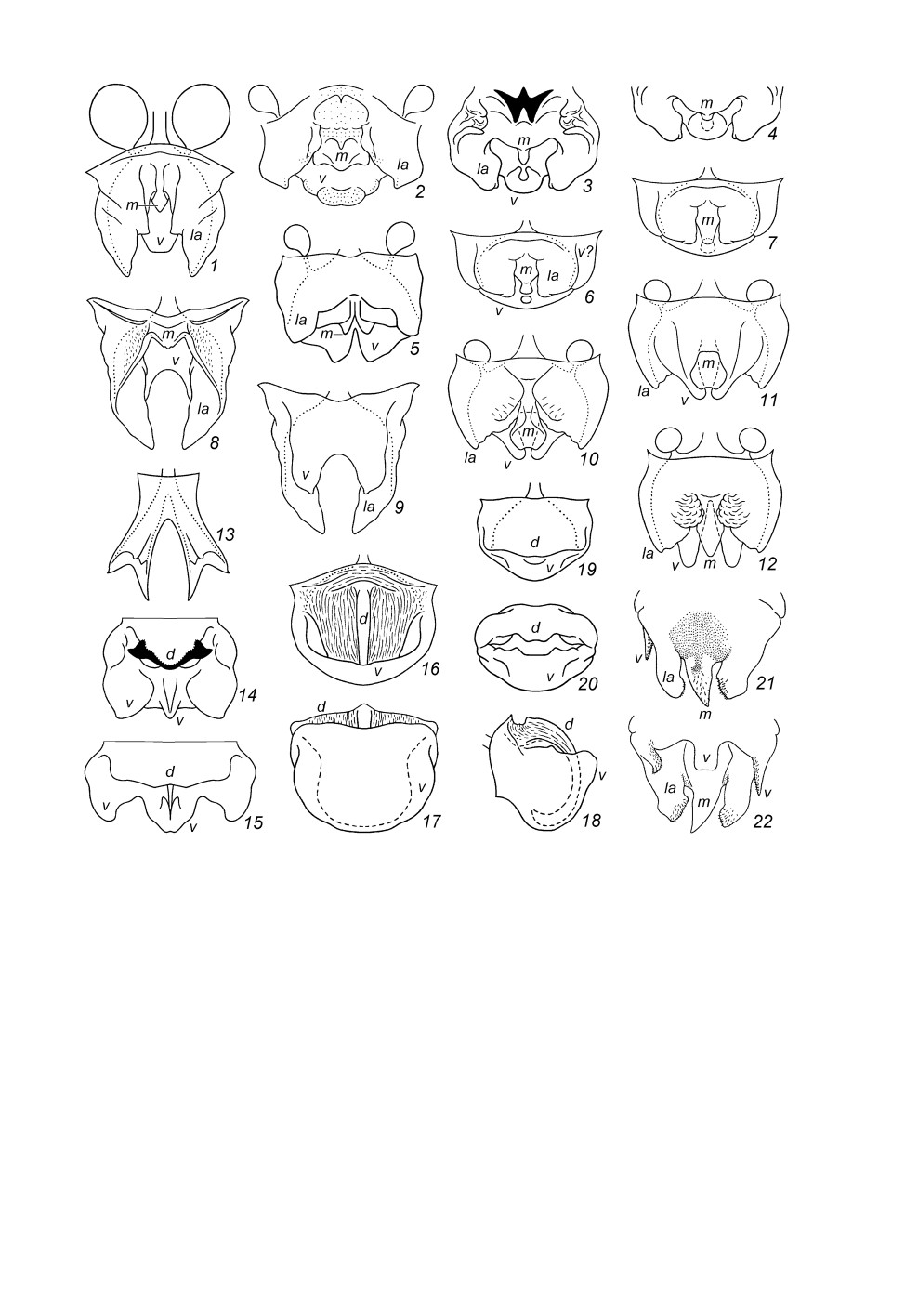
Рис. 1. Гениталии самцов инфраотряда Tettigoniidea, вид сверху (1-8, 10, 12-16, 19, 21), снизу
(9, 11, 22), сзади (17, 20) и сбоку (18), но с парой округлых придаточных желез (1, 5, 10-12)
или без них (2-4, 6-9, 13-22), либо без проксимальной части гениталий (4) (схематично).
1 - Prophalangopsidae (Cyphoderris Uhler); 2-4 - Tettigoniidae (2 - Nesoecia Scudd.; 3, 4 - Clonia Stål); 5-7 -
Anostostomatidae (5 - Anabropsis carli Griff.; 6, 7 - Lezina Walk.); 8-20 - Stenopelmatidae
(8, 9 - Stenopelmatopterus Gor.; 10, 11 - Maxentius pinguis Walk.; 12 - M. pallidus Walk.; 13 - Sia Giebel;
14 - Hyperbaenus Br.-W.; 15 - Capnogryllacris Karny; 16-18 - Schizodactylus Brullé;
19, 20 - Comicus orangensis sp. n.); 21, 22 - Rhaphidophoridae (Tamdaotettix semipullus Gor.).
5, 21, 22 - по: Горохов, 1998, с изменениями.
d - дорсальная складка; la, m - ее боковые (la) и срединная (m) лопасти; v - вентральная складка
или ее лопасти. Линиями из точек условно отмечены боковые края полости гениталий, расположенной
между d и v; пунктиром - края m, v или d, видимые сквозь мембранозные части.
других представителей Rhaphidophoridae (Болдырев, 1912) и у Schizodactylinae
(Gwynne, 1995) может быть результатом полного слияния не только флаконов спер-
матофорной ампулы, но и капсул со спермой внутри образованной таким образом
одиночной ампулы.
908
Формирование двойной ампулы сперматофора происходит, возможно, при замы-
кании генитальной полости срединной лопастью дорсальной складки, которая может
быть короткой, но погруженной внутрь генитальной полости и иногда раздвоенной и
складчатой (см. рис. 1, 1, 2, 5, 8); сходный тип гениталий самца развит у современных
Hagloidea, некоторых Tettigoniidae, Stenopelmatidae и Anostostomatidae. Однако
у других представителей трех последних семейств срединная лопасть дорсальной
складки может быть удлиненной и снабженной на вершине утолщением, которое
вставляется в специальное углубление срединной части вентральной складки
(рис. 1, 6, 7), в выемку между лопастинками этой части (рис. 1, 3, 4) или между лопа-
стями этой складки (рис. 1, 10, 11); у ряда родов Gryllacridinae имеется тонкий мем-
бранозный срединный вырост в основании нижней части дорсальной складки, вер-
шина которого фиксируется между лопастинками срединной лопасти вентральной
складки (рис. 1, 15).
В некоторых группах Stenopelmatidae такой план строения гениталий самца значи-
тельно изменен и будет рассмотрен отдельно (рис. 1, 13, 16-20), а у Rhaphidophoridae
срединный вырост дорсальной складки сильно вырьирует по длине, но может быть не
погружен внутрь генитальной полости. Если к этому добавить маловероятную воз-
можность независимого (от остальных Stenopelmatoidea) частичного разделения
капсулы со спермой у некоторых Rhaphidophoridae на две части и учесть иную трак-
товку последовательности изменений в характере сцепления створок в яйцекладе
(Клюге, 2016), то полностью бескрылые Rhaphidophoridae могут оказаться не близ-
кими родственниками Stenopelmatidae, а ответвлением от основания общего ствола
Stenopelmatoidea или даже от еще более ранней группы. Кстати, во второй половине
мезозоя нам известны две хорошо обособленные (но загадочные) группы крылатых
Ensifera (Vitimiidae, ранний мел; Phasmomimidae, поздняя юра), у последней из ко-
торых известны тела, несколько напоминающие таковые Rhaphidophoridae (Горохов,
2000; рис. 2, 2).
Сем. STENOPELMATIDAE - лжекузнечики
Это семейство было разбито на 5 подсемейств (Горохов, 1988б): Stenopelmatinae
Burmeister, 1838; Oryctopinae Kevan, 1986; Siinae Gorochov, 1988; Schizodactylinae
Blanchard, 1845 и Gryllacridinae Blanchard, 1845. Три первых подсемейства - близко-
родственны и в цитированной работе были объединены в особую группу
«Stenopelmatidae», причем морфологические различия между ними не превышают та-
ковых внутри крупного подсем. Phaneropterinae (Tettigoniidae), ранг которого не вызы-
вает ныне больших разногласий среди авторов. По этой причине я считаю нецелесо-
образным сохранять ранг подсемейства для Oryctopinae и Siinae и предлагаю
рассматривать их лишь как трибы подсем. Stenopelmatinae. Таким образом, в насто-
ящем я подразделяю сем. Stenopelmatidae лишь на три подсемейства: Stenopelmatinae,
Schizodactylinae и Gryllacridinae. Диагностические признаки этих подсемейств даны
в определительной таблице ниже.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ СЕМ. STENOPELMATIDAE
1. Глаза расположены примерно в средней (но не в верхней) части эпикраниума; про-
странство между усиковыми впадинами широкое и почти без рострума, т. е.
практически без какого-либо выступа, направленного вперед и вверх (рис. 2, 3,
4). Среднеспинка и заднеспинка в различной степени уменьшенные - короче и
909
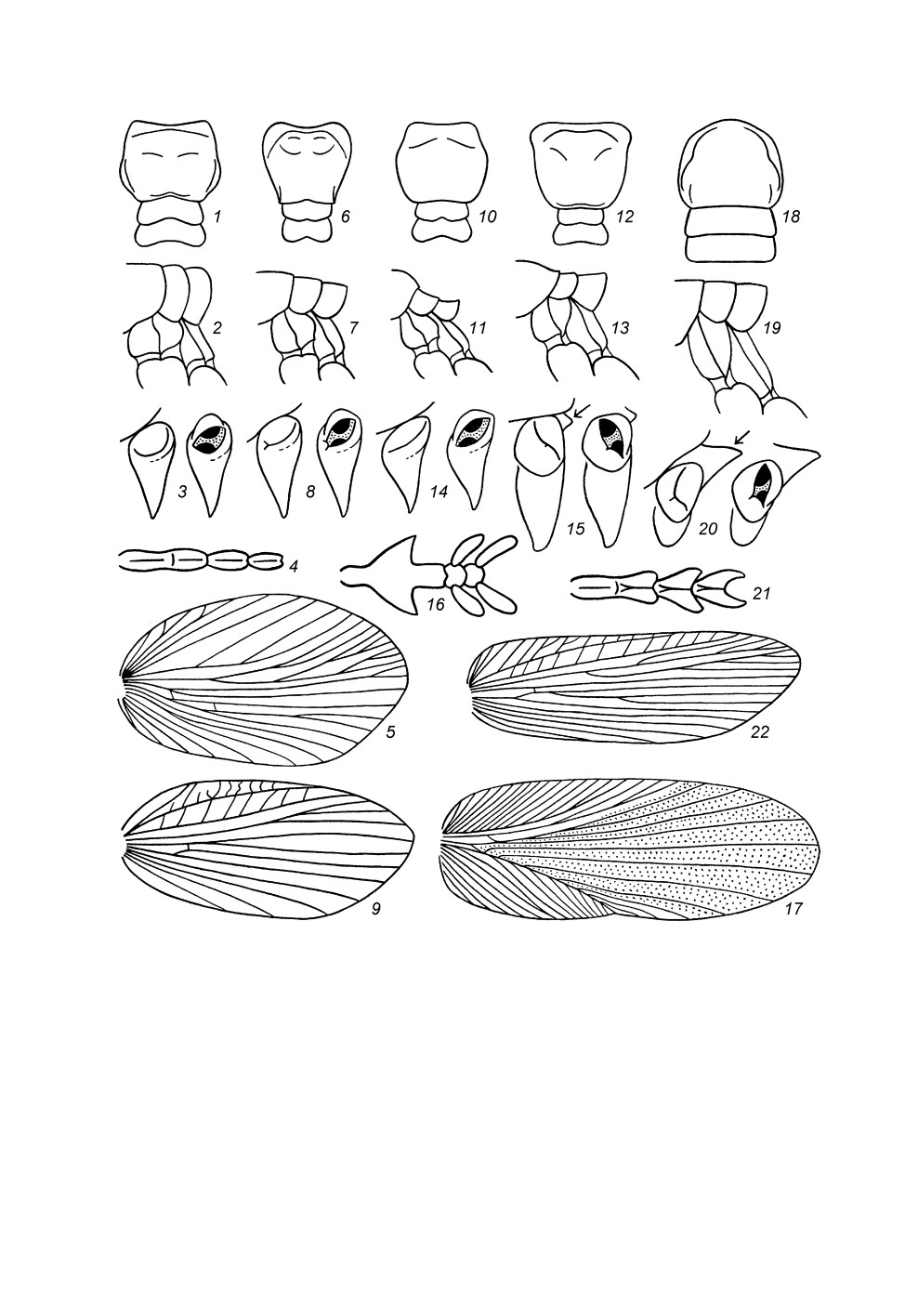
Рис. 2. Stenopelmatidae и Phasmomimidae.
1 - самец Schizodactylus inexspectatus (Wern.) в природной обстановке, Турция; 2 - отпечаток самца
неопределенного представителя сем. Phasmomimidae из верхней юры Казахстана; 3-5 - Sia
bugajus sp. n., самец (голотип) в позе угрозы с поднятыми передними ногами (3) и в более
спокойном состоянии (4), самка (паратип) в позе обороны (тело перевернуто на спину) (5).
1 - фотография А. В. Горохова; 2 - по: Горохов, 2000; 3-5 - фотографии М. В. Березина.
уже переднеспинки; плейриты средне- и заднегруди укороченные (рис. 3, 1, 2, 6,
7, 10-13); переднее грудное дыхальце с косо-горизонтальной щелью на верти-
кальной выпуклости, лишенной заднего выступа (рис. 3, 3, 8, 14). Лапки узкие,
т. е. без заметных боковых лопастевидных выростов, но с 4 парами маленьких
подошвенных подушечек (2 из них на базальном членике и иногда значительно
редуцированы; рис. 3, 4). Продольное жилкование надкрылий, если они разви-
ты, почти полностью параллелизованное (типичное для надсем. Stenopel-
910
Рис. 3. Stenopelmatidae. Грудь бескрылых экземпляров сверху (1, 6, 10, 12, 18) и их птероторакс
с плейритами сбоку (2, 7, 11, 13, 19); переднее грудное дыхальце и выпуклость, на которой оно
расположено, с сомкнутыми и раздвинутыми лопастинками дыхальца (3, 8, 14, 15, 20; стрелка
указывает на задний выступ выпуклости); 3 первых членика задней лапки снизу (4, 16, 21);
надкрылье (5, 9, 17, 22; точками покрыта веерообразно складывающаяся часть надкрылья,
дистальный участок которой закручивается в колечко в покое) (схематично).
1-3 - Maxentius pinguis (Walk.); 4, 5 - Sia Giebel; 6-9 - Oryctopterus lagenipes (Karny), самка (6, 7) и самец
(8, 9); 10, 11 - Oryctopus prodigiosus Bol., самка; 12-14 - Stenopelmatus talpa Burm.; 15-17 - Schizodactylus
hesperus B.-Bien.; 18, 19 - Nippancistroger Griff.; 21 - Capnogryllacris Karny; 22 - Hadrogryllacris Karny.
По: Горохов, 1995, с изменениями.
911
matoidea; рис. 3, 5, 9); дистальные части обеих пар крыльев у способных к
полету форм не закручиваются в колечки в состоянии покоя (рис. 2, 4). Нижние
части передних тергитов брюшка у форм с развитым феморо-абдоминальным
стридуляционным аппаратом снабжены мелкими и беспорядочно разбросанны-
ми шипиковидными щетинками, а также одним косо-вертикальным рядом из ча-
стых стридуляционных зубчиков на 2-м или 2-м и 3-м тергитах (такой
стридуляционный аппарат развит как у имаго, так и у нимф старших возрастов;
рис. 4, 1, 2, 9); церки с характерными мелкими грушевидными сенсиллами
в проксимально-медиальной части (рис. 4, 3); гениталии самца мембранозные, с
дорсальной и вентральной складками обычно разделенными на 3 и 2 лопасти
соответственно (причем срединная лопасть первой складки несколько погруже-
на внутрь генитальной полости; рис. 1, 8-12), но иногда преобразованными в
2 боковые трубчатые структуры (рис. 1, 13); яйцеклад значительно укорочен
(рис. 4, 7, 8, 12, 13, 20, 21) или редуцирован до нескольких мелких пальцевид-
ных лопастинок, немного выступаюших позади генитальной пластинки
........................................................................................................ Stenopelmatinae.
- Глаза расположены в верхней части эпикраниума; пространство между усиковыми
впадинами узкое или с коротким и округлым в профиль ростральным выступом
(рис. 2, 1; 15, 4). Среднеспинка и заднеспинка обычно не уже, а заднеспинка ча-
сто не короче переднеспинки; плейриты средне- и заднегруди неукороченные
(рис. 3, 18, 19); переднее грудное дыхальце с косо-вертикальной щелью на
разнообразной по высоте выпуклости, обычно несущей характерный задний вы-
ступ в верхней части (рис. 3, 15, 20). Лапки с заметными боковыми лопасте-
видными выростами, а их базальный членик с 1 парой маленьких подошвенных
подушечек или без них (рис. 3, 16, 21). Строение крыльев также более разно-
образно, чем у Stenopelmatinae (рис. 3, 17, 22). Беспорядочно разбросанные ши-
пиковидные щетинки феморо-абдоминального стридуляционного аппарата,
если он развит, отсутствуют или незаметны, но косо-вертикальный ряд или 2 та-
ких ряда из частых стридуляционных зубчиков на каждой нижней части 2-го и
3-го или 2-4-го тергитов брюшка часто хорошо развиты (в том числе и у нимф
старших возрастов; рис. 4, 14, 15); церки без грушевидных сенсилл; гениталии
самца (рис. 1, 14-20) с дорсальной складкой обычно почти не подразделенной
на лопасти и нередко частично склеротизованной, и с вентральной складкой,
полностью мембранозной и довольно разнообразной по строению (но иногда
дорсальная складка преобразована в удлиненную структуру, вдающуюся внутрь
генитальной полости; рис. 1, 16-18); яйцеклад от хорошо развитого и длинного
до практически совсем редуцированного (рис. 4, 25, 26; 6, 12, 19, 20)
2.
2. Пространство между усиковыми впадинами очень узкое и без рострального высту-
па (рис. 2, 1). Переднее грудное дыхальце расположено на высокой вертикаль-
ной выпуклости, снабженной небольшим задним выступом сверху (иногда этот
выступ может быть редуцирован; рис. 3, 15). Лапки с 1 парой довольно тонких
боковых лопастевидных выростов на 2-м и 3-м члениках; 1-й членик задней лап-
ки с 1 парой похожих или расширенных выростов и практически без подошвен-
ных подушечек (рис. 3, 16). Продольное жилкование надкрылий, если они
хорошо развиты, частично не параллелизовано, поскольку вдоль бокового края
дорсальной плоскости надкрылья расположена продольная жилка, от которой
последовательно и косо отходят многочисленные ветви, направленные немного
912
в разные стороны (рис. 3, 17); дистальные части обеих пар крыльев у способных
к полету форм закручиваются в колечки в состоянии покоя (столь значительные
изменения в строении крыльев связаны, наиболее вероятно, с адаптацией к рою-
щему образу жизни, сопровождающейся компактизацией сложенных крыльев и
коробчатым, почти как у сверчков, складыванием надкрылий). Феморо-абдоми-
нальный стридуляционный аппарат, если развит, состоит из 1 ряда стридуляци-
онных шипиков на каждой нижней части 2-го, 2-го и 3-го или 2-4-го тергитов
брюшка (рис. 4, 14); гениталии самца с вентральной складкой не подразделен-
ной на лопасти и с дорсальной складкой сильно удлиненной, полусклеротизо-
ванной (полумембранозной) и глубоко вдающейся внутрь генитальной полости
(рис. 1, 16-18), но иногда сильно редуцированной, возможно, в связи с ларва-
лизацией имаго (рис. 1, 19, 20); яйцеклад практически совсем редуцирован
(рис. 4, 25, 26; 6, 12, 19, 20)
Schizodactylinae.
– Пространство между усиковыми впадинами сравнительно широкое, с коротким,
округлым в профиль ростральным выступом (рис. 15, 4). Переднее грудное ды-
хальце расположено на сравнительно низкой выпуклости, снабженной крупным
задним выступом сверху (рис. 3, 20). Лапки с 1 парой расширенных боковых ло-
пастевидных выростов на 1-м, 2-м и 3-м члениках; 1-й членик задней лапки так-
же с 1 парой маленьких подошвенных подушечек в проксимальной половине
(рис. 3, 21). Крылья, если хорошо развиты, приблизительно как у Stenopelmatinae
(рис. 3, 22), т. е. без вышеперечисленных специализаций, характерных для
Schizodactylinae. Феморо-абдоминальный стридуляционный аппарат, если раз-
вит, обычно состоит из 2 рядов стридуляционных зубчиков на каждой нижней
части 2-го и 3-го тергитов брюшка (рис. 4, 15); гениталии самца полностью мем-
бранозные или с частично склеротизованной дорсальной складкой, причем по-
следняя складка довольно короткая, а вентральная складка обычно подразделена
на 3 мембранозные лопасти (рис. 1, 14, 15); яйцеклад развит (от длинного до за-
метно укороченного)
Gryllacridinae.
Подсем. STENOPELMATINAE - настоящие лжекузнечики
Это подсемейство здесь разделено как минимум на 4 трибы: Stenopelmatini
Burmeister, 1838; Siini Gorochov, 1988, stat. resurr.; Maxentiini trib. n.; Oryctopterini
Gorochov, 1988, trib. resurr. Первая триба известна лишь из Центральной и Северной
Америки и включает следующие роды: Stenopelmatus Burmeister,
1838; Steno-
pelmatopterus Gorochov, 1988; Ammopelmatus Tinkham, 1965 и Viscainopelmatus Tink-
ham, 1970, близкие к Stenopelmatus или похожие на него, но c недостаточно понят-
ными отличиями от этого рода.
Вторая триба содержит один род с Малайского архипелага (Sia Giebel, 1861) и один
ископаемый род из эоценового балтийского янтаря (Electrosia Gorochov, 2010),
а третья - только род Maxentius Stål, 1876, stat. resurr. из Африки. Следует отметить,
что Д. Отте (Otte, 2000) впервые придал Siini статус трибы и сделал Maxentius под-
родом рода Sia. Позднее эта триба снова стала рассматриваться как подсемейство
(Cigliano et al., 2020), но Maxentius продолжал считаться подродом (Weissman, Bazelet,
2013; Cigliano et al., 2020). Такое объединение родов Maxentius и Sia явно ошибочно,
поскольку по строению копулятивного аппарата Maxentius более сходен с родами
913
Stenopelmatini, чем с настоящими представителями рода Sia, и заслуживает не только
родового статуса, но и обособления в составе отдельной (новой) трибы.
Четвертая триба (Oryctopterini) включает один род из Шри-Ланки (Oryctopterus
Karny, 1937, gen. resurr.), но может быть синонимом или подтрибой трибы Oryctopini
Kevan, 1986, содержащей один род из Индии (Oryctopus Brunner-Wattenwyl, 1888). Ко-
пулятивные структуры рода Oryctopus практически не изучены, а передние голени у
его представителей значительно короче и толще, чем у Oryctopterus, и снабжены креп-
кими копательными шипами в дистальной части, тогда как у Oryctopterus передние
голени тонкие и довольно длинные, снабжены маленькими шипами лишь на вершине
и не участвуют в рытье почвы или участвуют в этом особым образом, либо могут
использоваться как колющие орудия защиты или нападения. Таким образом,
синонимизация этого рода с Oryctopus (Johns, 1997) ошибочна, а Oryctopterini с
Oryctopini (Cigliano et al., 2020) - преждевременна. Диагностические признаки триб
подсем. Stenopelmatinae приведены в определительной таблице ниже.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ТРИБ ПОДСЕМ. STENOPELMATINAE
1. Передние голени умеренно или слабо утолщенные, не тонкие на вершине (рис. 4, 5,
18); 1-й стернит брюшка разнообразный по строению (рис. 4, 4, 16, 17); 10-й
тергит брюшка самца с 1 парой широко расставленных копулятивных крючков
(рис. 4, 6, 19; 5, 1); генитальная пластинка самца без грифельков (рис. 4, 6, 19;
5, 1); гениталии самца короткие, полностью мембранозные и не втяжные, т. е. их
проксимальная часть не удлиненно-трубчатая, а более дистальная часть не спо-
собна частично втягиваться внутрь трубчатой части; дорсальная складка генита-
лий подразделена на 3, а вентральная - на 2 лопасти (рис. 1, 8-12); яйцеклад
сильно укорочен (рис. 4, 7, 8, 20, 21)
2.
– Передние голени примерно такие же, но могут быть сильно утолщенными или тон-
кими на вершине (рис. 4, 11, 22, 24). 1-й стернит брюшка довольно узкий (его
ширина не более чем в 1.5 раза больше длины) и хорошо обособлен от стернита
заднегруди (рис. 4, 10); 10-й тергит брюшка самца без копулятивных крючков
(рис. 4, 23; 5, 2); генитальная пластинка самца с грифельками или без
них (рис. 4, 23; 5, 3-5, 9, 12); гениталии самца также более разнообразные
(рис. 1, 13; 4, 23; 5, 7, 8, 10, 11); яйцеклад заметно укорочен (рис. 4, 12, 13) или
редуцирован до нескольких мелких пальцевидных выростов, незначительно вы-
ступающих позади генитальной пластинки
3.
2. 1-й стернит брюшка довольно широкий (его ширина более чем в 1.5 раза больше
длины) и плотно примыкает к стерниту заднегруди (рис. 4, 16, 17); феморо-
абдоминальный стридуляционный аппарат отсутствует (полностью редуциро-
ван?); крючки 10-го тергита брюшка самца расположены на особых выступах
тергита, направленных назад и вниз (рис. 4, 19)
Stenopelmatini.
– 1-й стернит брюшка довольно узкий (его ширина не более чем в 1.5 раза больше
длины) и хорошо обособлен от стернита заднегруди (рис. 4, 4); феморо-абдоми-
нальный стридуляционный аппарат развит, состоит из множества беспорядочно
разбросанных мелких бугорков или палочковидных щетинок на нижних полови-
нах 1-3-го тергитов брюшка, а также 1 почти вертикального ряда из частых
стридуляционных зубчиков на каждой нижней части 3-го или 2-го и 3-го терги-
тов брюшка (рис. 4, 1, 2); крючки 10-го тергита брюшка самца расположены на
914
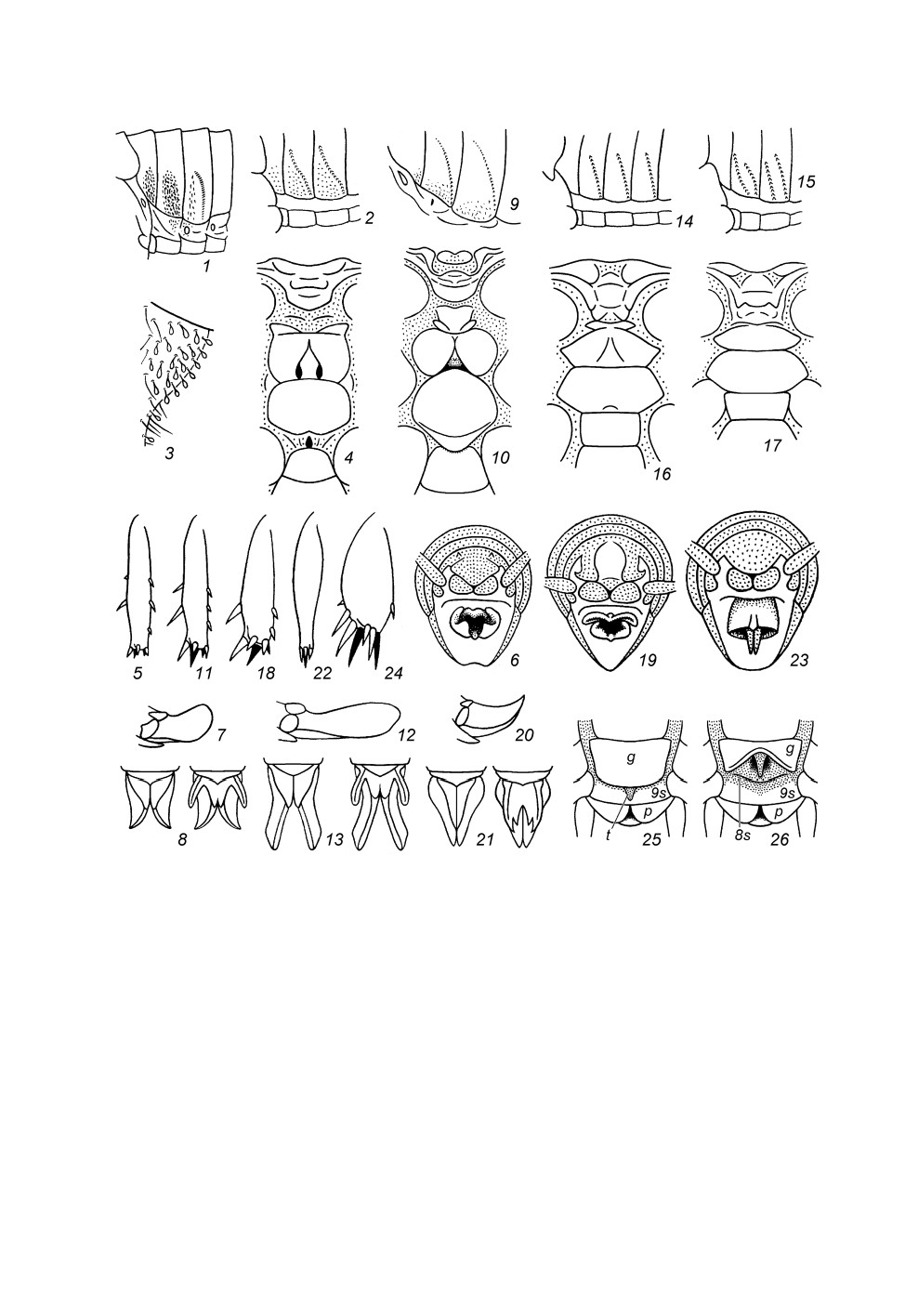
Рис. 4. Stenopelmatidae. Феморо-абдоминальный стридуляционный аппарат на передних
тергитах брюшка (1, 2, 9, 14, 15); медиальная часть основания церка сверху (3); стерниты
груди снизу (4, 10, 16, 17); передняя голень сбоку (5, 11, 18, 22, 24); вершина брюшка самца
сзади (6, 19, 23); яйцеклад сбоку (7, 12, 20); яйцеклад снизу, но с сомнутыми и раздвинутыми
нижними створками (8, 13, 21); генитальная пластинка самки и область полностью
редуцированного яйцеклада снизу, нормальное положение (25) и с отогнутой
вниз дистальной частью этой пластинки (26) (схематично).
1 - Maxentius pallidus (Walk.); 2-8 - M. pinguis (Walk.); 9, 10 - Sia bugajus sp. n.; 11-13 - Sia sp.;
14 - Schizodactylus sp., протонимфа; 15 - Diaphanogryllacris Karny; 16 - Stenopelmatopterus Gor.;
17-21 - Stenopelmatus talpa Burm.; 22, 23 - Oryctopterus lagenipes (Karny); 24 - Oryctopus
prodigiosus Bol.; 25, 26 - Schizodactylus hesperus B.-Bien. 2, 3, 5-8, 11-15, 18-24 - по: Горохов,
1995, с изменениями.
8s, 9s - 8-й и 9-й стерниты брюшка; g - генитальная пластинка; p - парапрокт; t - срединный язычок
генитальной пластинки.
915
незначительно увеличенных участках этого тергита (рис. 4, 6; 5, 1)
....................................................................................... Maxentiini Gorochov, trib. n.
[Типовой и единственный род трибы - Maxentius Stål, 1876 (Африка), название его -
существительное мужского рода.]
3. Передние голени слабо утолщенные и не тонкие на вершине (рис. 4, 11). Фемо-
ро-абдоминальный стридуляционный аппарат развит и похож по строению на
таковой Maxentiini, но беспорядочно разбросанные бугорки менее многочислен-
ные, а ряд из стридуляционных зубчиков расположен на каждой нижней части
2-го и 3-го тергитов брюшка (рис. 4, 9); гениталии самца полностью мембраноз-
ные или лишь с очень мелкими склеротизованными участками, удлиненные и
втяжные, т. е. их проксимальная часть удлиненно-трубчатая, а более дистальная
часть способна частично втягиваться внутрь трубчатой части; боковые части ге-
нитальной полости самца еще более обособлены одна от другой, чем у
Stenopelmatini и Maxentiini, поскольку дистальная половина гениталий преобра-
зована в пару трубчатых структур, открывающихся в единую полость лишь в
проксимальной половине гениталий (рис. 1, 13; 5, 7, 8, 10, 11); яйцеклад заметно
укорочен (рис. 4, 12, 13)
Siini.
– Передние голени слабо утолщенные и тонкие на вершине (рис. 4, 22), но если
Oryctopini относятся к этой же трибе, то эти голени могут быть также сильно
утолщенными и не тонкими на вершине (рис. 4, 24). Феморо-абдоминальный
стридуляционный аппарат отсутствует; гениталии самца напоминают таковые
Stenopelmatini и Maxentiini, но с дорсальной складкой, лишенной лопастей (кро-
ме очень короткой срединной лопастинки) и, возможно, со значительно более
длинными лопастями вентральной складки (рис. 4, 23); яйцеклад редуцирован
до нескольких мелких пальцевидных выростов, незначительно выступающих
позади генитальной пластинки
Oryctopterini.
Род SIA Giebel, 1861
Этот род был первоначально описан для одного яванского вида (Sia ferox Giebel,
1861). Вскоре тоже с Явы был описан Anostostoma couloni Saussure, 1862, который был
ошибочно включен в австралийский род, принадлежащий к другому семейству
(Anostostomatidae). Для последнего вида дважды и под разными названиями (Licola
Walker, 1869 и Bugajus Brunner-Wattenwyl, 1888) устанавливался самостоятельный
род. Но затем все эти виды и роды были синонимизированы (Kirby, 1906; Caudell,
1909), что, вероятно, правильно, но проверить уже невозможно, поскольку сохранился
лишь один типовой экземпляр S. ferox (самка) (Cigliano et al., 2020). Позднее был обна-
ружен еще один вид (S. incisa Karny, 1926 с Малайского полуострова); сначала он был
описан как подвид S. ferox, а затем его ранг был поднят до видового (Karny, 1937).
В последней работе S. ferox также был указан для Суматры, Борнео и о. Оби около
Новой Гвинеи, а S. incisa был дополнительно, но под вопросом, указан для Борнео.
Такое широкое распространение этих очень крупных, плохо летающих и не приспосо-
бленных к жизни на древесных стволах видов вызывает большое сомнение. Более
того, я имел возможность изучить экземпляры этого рода с Явы, Суматры и Борнео,
а также сравнить их с фотографиями типовых экземпляров S. ferox и S. incisa
(см. Cigliano et al., 2020), что позволило выявить три морфологически различимых и
географически обособленных вида: один из них (S. incisa) распространен на Малай-
916
ском полуострове и Суматре, которые фаунистически очень близки (за исключением
самого юга Суматры, по видовому составу более сходного с Явой); второй вид
(S. ferox) известен с Явы; третий вид обнаружен в северной части Борнео и описыва-
ется ниже как новый для науки.
Sia bugajus Gorochov, sp. n. (рис. 2, 3-5; 4, 9, 10; 5, 2-8).
М ате риа л. Малайзия, о. Борнео, штат Сабах («Sabah»): национальный парк «Crocker
Range» недалеко от г. «Keningau», 1000-1300 м, на ветвях кустов у лесной тропы, ночью,
2-6.V.2013 (A. Горохов, В. Горохова), 2 ♂ - голотип и паратип, 1 ♀ - паратип [ЗИН]; гора «Trus
Madi», ~1000 м, в лесу, на свет, 13-25.V.2007 (A. Горохов), 1 ♂ - паратип [ЗИН]; «Nord Borneo»,
«Sia ferox Giebel det. Ander», 1 ♂ - паратип [МИЗ].
С а ме ц (голотип). Тело очень крупное, типичного для рода облика (рис. 2, 3, 4). Окраска од-
нотонно светло-коричневая, но с коричневыми мандибулами, желтоватыми нижней половиной
клипеуса и верхней частью лабрума, желтовато-полупрозрачными мембранами вдоль косталь-
ного и анального краев надкрылья, почти прозрачными мембранами большей части заднего
крыла (кроме костальной лопасти, желтоватой в дистальной части и светло-коричневой прокси-
мальнее) и затемненными дистальными частями шипов и шпор на ногах. Голова крупная, высо-
кая, но довольно короткая (слегка уплощенная спереди); ее ширина приблизительно равна
ширине и длине переднеспинки; глаза вертикально удлиненные, овально-почковидные, но
суженные в нижней части, расположены в средней (по высоте) части эпикраниума и практиче-
ски не выступающие по бокам; рострум редуцирован, а на его месте кверху от середины усико-
вых впадин расположена слабая вогнутость с едва выпуклой срединной частью, которая
расширяется книзу и кверху (причем нижняя порция этой части существенно длиннее верхней),
а также снабжена низкими и узкими (но отчетливыми) килями по бокам; расстояние между уси-
ковыми впадинами примерно вдвое больше ширины скапуса; клипеальный шов стерт в средин-
ной трети и округло загнут кверху по бокам; ротовые части нормального строения, с мощными
мандибулами и тонкими и умеренно длинными щупиками (максиллярный щупик с предпослед-
ним члеником самым длинным, с 3-м члеником почти в 1.3 раза более коротким и с последним -
промежуточным между ними по длине). Переднеспинка слегка расширяющаяся кпереди,
с довольно глубокой бороздой вдоль переднего края, тянущейся от одной боковой лопасти до
другой, и со слабой поперечной вогнутостью на диске вдоль его заднего края; передний и задний
края диска слабовогнутые; задняя лопасть переднеспинки очень короткая и отделена от боковых
лопастей очень пологими плечевыми выемками; каждая боковая лопасть с округло-угловидным
передневерхним выступом и волнистым нижним краем. Крылья длинные; вершины надкрылий
и задних крыльев достигают вершины генитальной пластинки в состоянии покоя; ширина над-
крылья приблизительно в 2.2 раза меньше длины, а его дистальная часть заметно сужающаяся
к закругленной вершине; продольное жилкование, типичное для рода, параллелизованное в над-
крыльях и веерообразное в заднем крыле (поперечные жилки в обеих парах крыльев хорошо
развиты и умеренно многочисленные). Ноги также типичного для рода строения, сильные и
умеренно длинные; переднее и среднее бедра несколько утолщенные, с 4-6 небольшими неот-
члененными шипами только на нижнем внутреннем киле среднего бедра; заднее бедро значи-
тельно более длинное и заметно (но не сильно) более утолщенное в проксимальной половине
(т. е. адаптировано к сравнительно слабым прыжкам), с 4 небольшими неотчлененными шипами
лишь на нижнем наружном киле; передняя голень с 2 внутренними и 1 наружным верхними
шипами, 2 наружными и 1 внутренним нижними шипами, а также с 2 наружными и 3 внутрен-
ними вершинными шпорами, причем только проксимальный верхний шип почти неотчленен-
ный и расположен в средней части бедра, а все остальные шипы и шпоры отчлененные и
расположены дистальнее (по величине все шипы и шпоры немного крупнее таковых на бедрах,
но дистальный внутренний шип и 2 верхние внутренние вершинные шпоры значительно более
длинные); вооружение средней голени похоже на таковое передней, но верхних внутренних ши-
пов 3, нижних шипов 1 пара (внутренний из этих шипов приближен к шпорам), а вершина голе-
917
ни с 2 внутренними и 3 наружными шпорами, причем все шипы и шпоры отчлененные и
некрупные, а проксимальный внутренний верхний шип расположен в основании голени; задняя
голень с 5 внутренними и 4 наружными верхними неотчлененными и некрупными шипами, без
нижних шипов и с 3 парами вершинных шпор, из которых 2 внутренние и 2 наружные верхние
шпоры заметно более длинные. Брюшко сравнительно длинное; феморо-абдоминальный стри-
дуляционный аппарат развит, включает 1 почти вертикальный и слегка дуговидный ряд стриду-
ляционных зубчиков на каждой нижней части 2-го и 3-го тергитов брюшка, а также
беспорядочно разбросанные и немногочисленные короткие шипиковидные бугорки в нижних
частях 1-3-го тергитов брюшка (рис. 4, 9); предпоследний тергит брюшка с очень короткой (едва
выпуклой) верхнезадней лопастью, прикрывающей сверху лишь переднюю часть последнего
тергита; последний тергит снабжен округло-прямоугольной поперечной верхнезадней лопа-
стью, которая заметно (но не далеко) выступает позади основания церок и имеет едва выемча-
тую (почти обрубленную) вершину; какие-либо крючки или зубчики на последнем тергите
отсутствуют (рис. 5, 2); эпипрокт и парапрокты небольшие, простые, с широкоокругленными
видимыми частями; церки также простые, умеренно длинные и тонкие, без каких-либо иных
специализаций кроме тех, что свойственны Stenopelmatinae; генитальная пластинка заметно уд-
линенная, с расширенным основанием и суженной более дистальной частью, которая по бокам
выпуклая (т. е. эта пластинка с S-образными боковыми краями), а на вершине снабжена очень
узкой и умеренно глубокой срединной выемкой, а также 1 парой умеренно коротких и почти
пальцевидных грифельков (рис. 5, 3); гениталии практически полностью мембранозные, с ка-
ждой из дистальных ветвей несущей сравнительно короткую шиповидную вентромедиальную
лопасть и немного более длинную дорсолатеральную лопасть (последняя лопасть снабжена дву-
мя округлыми и почти полумембранозными лопастинками на вершине; рис. 5, 7, 8).
Ва р иа ции. Другие самцы (паратипы) отличаются от голотипа лишь немного
меньшим или незначительно большим числом шипов на среднем и заднем бедрах, а
также слегка более глубокой (рис. 5, 4) или едва более широкой (рис. 5, 5) вершинной
выемкой генитальной пластинки; у самца с горы «Trus Madi» верхняя половина кли-
пеуса интенсивно коричневая, а мандибулы - темно-коричневые.
С ам ка. Тело похоже на таковое самцов, но еще крупнее; окраска похожа на таковую самца с
горы «Trus Madi», но нижняя часть эпикраниума почти вся интенсивно коричневая, а верхняя
половина клипеуса почти темно-коричневая; голова очень крупная, немного шире длины перед-
неспинки; рострум почти без килей в верхней части (выше наиболее узкого участка между киля-
ми); расстояние между усиковыми впадинами примерно в 2.3 раза шире скапуса; переднеспинка
немного сильнее расширяется кпереди; феморо-абдоминальный стридуляционный аппарат по-
хож на таковой самца, но 1-й тергит брюшка практически без шипиковидных бугорков; предпо-
следний тергит без задней лопасти; последний тергит с немного более маленькой (но сходной по
форме) задней лопастью; генитальная пластинка треугольная, с отчетливо угловидной и едва
оттянутой кзади вершиной, но без какой-либо вершинной выемки (рис. 5, 6); яйцеклад практи-
чески такой же, как на рис. 4, 12, 13.
Длина (в мм). Тело: ♂ - 55-65, ♀ - 63; тело с крыльями: ♂ - 61-65, ♀ - 68; переднеспинка:
♂ - 11-12, ♀ - 13; надкрылье: ♂ - 44-47, ♀ - 49; переднее бедро: ♂ - 14.5-15, ♀ - 16; заднее
бедро: ♂ - 24-25.5, ♀ - 27; задняя голень: ♂ - 24.5-26, ♀ - 28; задняя лапка: ♂ - 17-18, ♀ - 19;
яйцеклад - 6.2.
Сравнение. Новый вид отличается от S. ferox менее постепенно сужающейся к
вершине генитальной пластинкой самца, менее глубокой и менее широкой вершинной
выемкой этой пластинки, значительно более короткими грифельками (ср. рис. 5, 3-5 и
5, 9), более короткими вентромедиальными лопастями в гениталиях самца, не склеро-
тизованными и не крючковидными вершинными лопастинками их дорсомедиальных
лопастей (рис. 5, 7, 8 и 5, 10, 11), а также угловидной (а не закругленной) вершиной
918
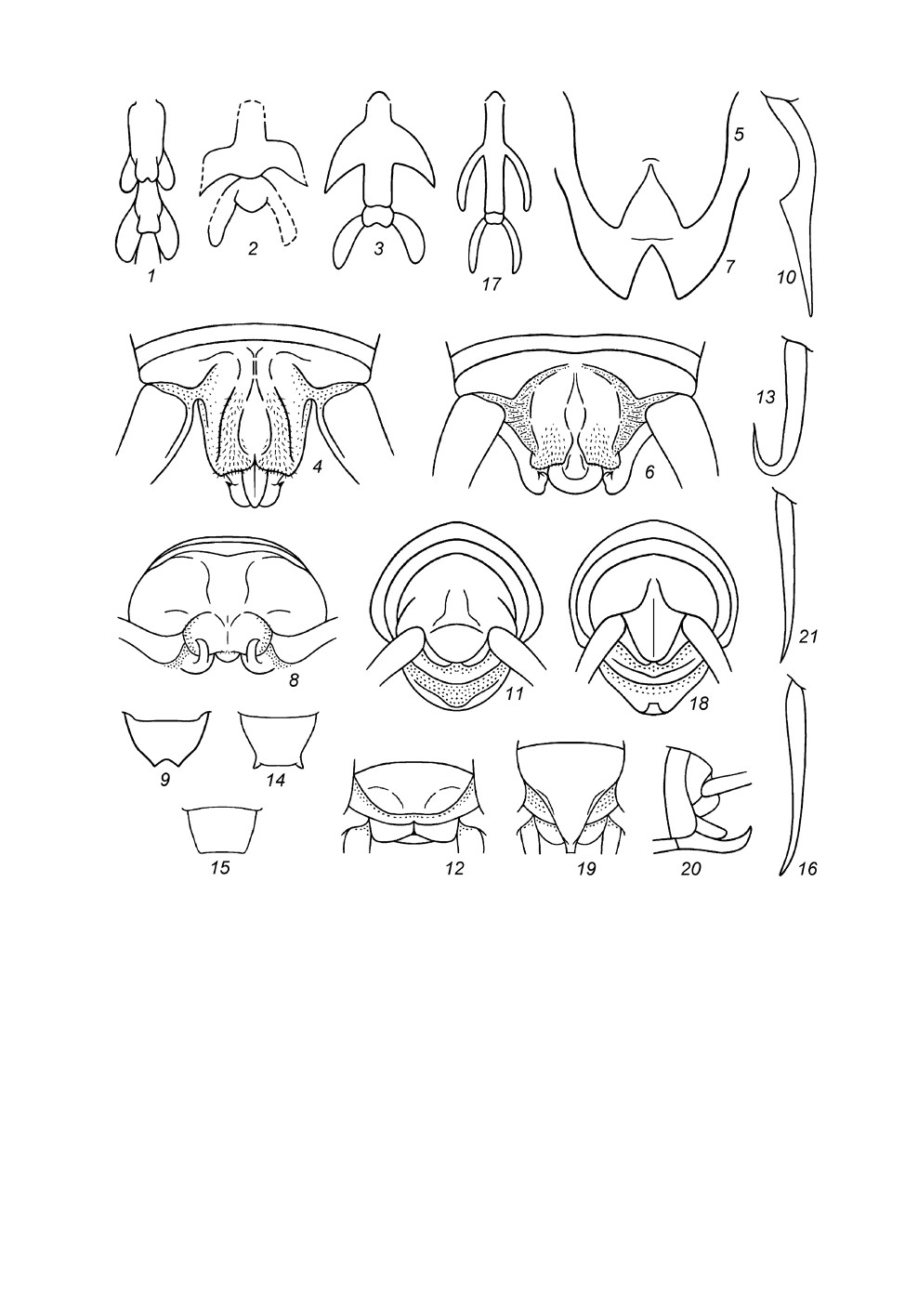
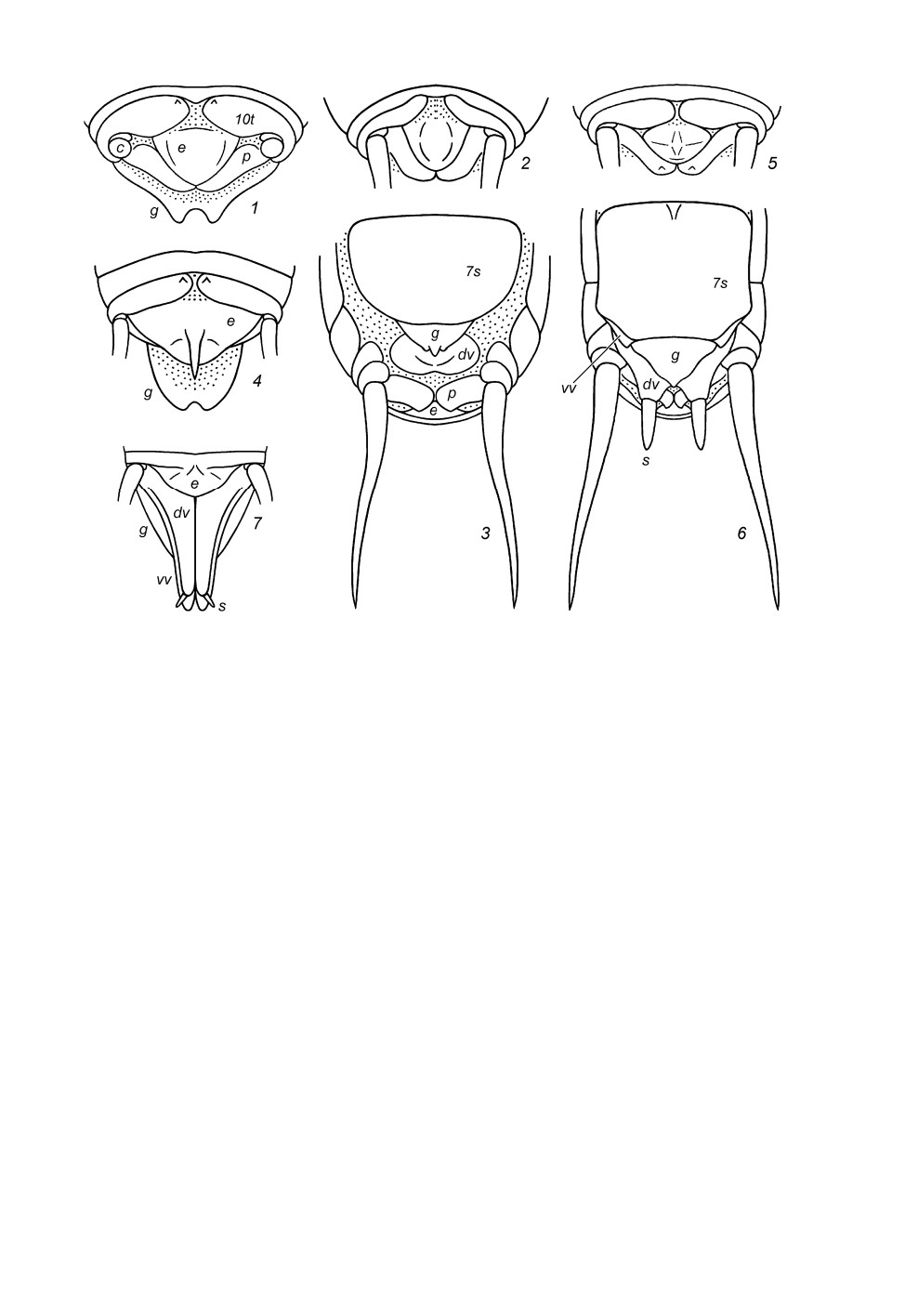
Рис. 5. Stenopelmatinae. Вершина брюшка самца сверху и слегка сзади (1, 2); генитальная
пластинка самца (3-5, 9, 12) и самки (6) снизу; гениталии самца с дистальной половиной,
втянутой в проксимальную (7, 8), и с обоими половинами, не втянутыми одна в другую
(рис. 10, 11), сверху (7, 10) и снизу (8, 11) (схематично).
1 - Maxentius pallidus (Walk.); 2-8 - Sia bugajus sp. n.; 9-11 - S. ferox Giebel; 12 - S. incisa Ander.
генитальной пластинки самки (рис. 5, 6). От S. incisa новый вид отличается выпу-
клыми боковыми краями дистальной половины генитальной пластинки самца, более
узкой (угловидной) передней частью вершинной выемки этой пластинки, более широ-
кими несущими грифельки лопастями по бокам от этой выемки (рис. 5, 3-5 и 5, 12),
а также отсутствием какой-либо выемки на вершине генитальной пластинки самки
(рис. 5, 6).
Этимология. Видовое название - существительное мужского рода, дано по
родовому названию Bugajus (от тюркского корня, означающего быка) в связи с круп-
ными размерами и крепким телосложением.
919
Sia ferox Giebel, 1861 (рис. 5, 9-11).
Мат ер иа л. Индонезия, о. Ява, «Halimun Salak N. P., WP 677», 20.III.2012 (Н. Юнаков), 1 ♂
[ZIN].
Этот самец внешне очень похож на самцов S. bugajus sp. n., но участки клипеуса
вдоль остатков клипеального шва и мандибулы коричневые, остальные части кли-
пеуса и весь лабрум светло-коричневые, верхнезадняя лопасть последнего тергита не-
сколько более вогнутая сзади, генитальная пластинка с почти прямыми боковыми
краями (если смотреть снизу) и более крупной вершинной выемкой, грифельки этой
пластинки значительно более длинные (рис. 5, 9), а гениталии с вентромедиальными
шиповидными лопастями более длинными, чем дорсолатеральные лопасти (причем
дистальные лопастинки на каждой из последних лопастей очень короткие: одна из них
с маленьким склеротизованным и крючковатым шипиком на вершине, а другая - пол-
ностью мембранозная и округлена на вершине; рис. 5, 10, 11). Самка этого яванского
вида, судя по фотографии его голотипа (Cigliano et al., 2020), отличается от самки
S. bugajus sp. n. округленной (а не угловидной) вершиной генитальной пластинки.
Sia incisa Karny, 1926 (рис. 5, 12).
Мат ер иа л. Индонезия, о. Суматра, «Bugajus couloni Sss», «Sia incisa Karny det. Ander», 1 ♂
[МИЗ].
Этот самец отличается от самцов S. bugajus sp. n. признаками, перечисленными в
разделе о сравнении последнего вида с другими видами рода (ср. рис. 5, 3-5 и 5, 12).
От S. ferox он отличается более узкой в дистальной части генитальной пластинкой
самца с более маленькой вершинной вырезкой и значительно более короткими гри-
фельками (рис. 5, 9 и 5, 12), а также гениталиями самца, похожими на таковые
S. bugajus sp. n., но, возможно, имеющими почти обрубленные дистальные части дор-
солатеральных лопастей. Самка S. incisa, судя по фотографиям голотипа этого вида c
Малайского полуострова (Cigliano et al., 2020) и неправильно определенного экзем-
пляра с Суматры (Desutter-Grandcolas, 2020a: S. ferox), отличается от обоих перечис-
ленных выше видов наличием маленькой выемки на вершине генитальной пластинки.
Род MAXENTIUS Stål, 1876
Этот род включает 4 вида из Южной Африки (Cigliano et al., 2020): M. pinguis
(Walker, 1869); M. pallidus (Walker, 1869); M. kuhlgatzi Karny, 1910 и M. canus Péringuey,
1916. Кроме того, в этом роде были описаны еще 4 вида, названия которых были све-
дены в синонимы к M. pinguis и M. pallidus в те времена, когда на тонкие признаки
стридуляционных аппаратов и копулятивных устройств еще почти не обращали вни-
мания (Kirby, 1906); таким образом, получилось, что эти бескрылые виды имеют
весьма широкое распространение (от Центральноафриканской Республики до юга
Южной Африки), тогда как другие бескрылые роды того же семейства в Африке
(Comicus) и Центральной Америке (Stenopelmatus) представлены многочисленными
видами со значительно более узкими ареалами. Возможно, эти синонимизированные
названия или некоторые из них принадлежат хорошим видам, но для выяснения этой
проблемы требуется дополнительное исследование. В данный момент можно лишь
констатировать, что виды этого рода распадаются на 3 группы, различия между кото-
рыми предполагают подродовой уровень дивергенции. Эти различия перечислены
ниже в определительной таблице подродов и признанных видов рода Maxentius.
920
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ И ВИДОВ РОДА MAXENTIUS
1. Передние ноги умеренно короткие (длина их голени приблизительно в 1.3-1.5 раза
меньше высоты головы) и с 2 крупными дорсальными шипами на наружном
крае голени
3.
– Передние ноги довольно длинные (длина их голени примерно равна высоте головы)
и с 3 крупными дорсальными шипами на наружном крае голени. Подрод
Amaxentius Gorochov, subgen. n
2.
[Состав: типовой вид - Maxentius kuhlgatzi Karny, 1910; M. canus Péringuey, 1916. Эти-
мология: название образовано от родового названия Maxentius с латинизирован-
ной греческой приставкой «a-», означающей «не» или «против»).]
2. Окраска контрастная - с поперечными темными полосами на светлом фоне
........................................................................................................ M. (A.) kuhlgatzi.
- Окраска почти однотонно светлая - лишь с затемненными вершинами шипов и
шпор на ногах
M. (A.) canus.
3. Феморо-абдоминальный стридуляционный аппарат с 1 рядом стридуляционных
зубчиков на каждой нижней части 2-го и 3-го тергитов брюшка, а также с корот-
кими и почти зубчиковидными беспорядочно разбросанными бугорками около
этого ряда (рис. 4, 2; 7, 8, 9); последний тергит брюшка самца слит с эпипроктом
в срединной части (рис. 4, 6); гениталии самца с заметно расширенной верши-
ной срединной лопасти дорсальной складки, т. е. с запирательным механизмом
(рис. 1, 10, 11)
подрод Maxentius s. str.
[Состав: типовой вид - Maxentius repens Stål, 1876; его возможный синоним -
Stenopelmatus pinguis Walker, 1869.]
- Феморо-абдоминальный стридуляционный аппарат с 1 рядом стридуляционных
зубчиков на каждой нижней половине только 3-го тергита брюшка, а также с
тонкими и умеренно длинными палочковидными, беспорядочно разбросанными
щетинками (вместо коротких бугорков) на нижних частях 1-3-го тергитов
брюшка (рис. 4, 1; 7, 7); последний тергит брюшка самца не слит с эпипроктом
(рис. 5, 1); гениталии самца с узкой (не расширенной) вершиной срединной ло-
пасти дорсальной складки, т. е. без запирательного механизма (рис. 1, 12)
................................................................ подрод Eumaxentius Gorochov, subgen. n.
[Состав: типовой вид - Stenopelmatus pallidus Walker, 1869; его возможные синони-
мы: S. bicolor Walker, 1869, S. zonatus Walker, 1869 и Maxentius fuscofasciatus Stål, 1876.
Этимология: название образовано от родового названия Maxentius c латинизирован-
ной греческой приставкой «eu-», означающей «хороший» или «настоящий».]
Подсем. SHIZODACTYLINAE - монстровидные лжекузнечики
Это подсемейство включает 2 близких современных рода: Shizodactylus Brullé, 1835
(= Dactylocomicus Karny, 1931), распространенный от южной Турции до Юго-
Восточной Азии, и Comicus Brunner-Wattenwyl, 1888, встречающийся только в Аф-
рике. Сравнительно недавно были также описаны ископаемый род Brauckmannia
Martins-Neto, 2007, gen. resurr. из верхней половины нижнего мела Бразилии и сем.
Brauckmanniidae Martins-Neto, 2007 для этого рода. Вскоре они были синонимизиро-
921
ваны с Schizodactylus и Schizodactylinae (Heads, Leuzinger, 2011), что правильно для
надродовых таксонов, но очень сомнительно для родовых; здесь я восстанавливаю род
Brauckmannia, поскольку, судя по приведенной фотографии в вышеупомянутой статье,
боковые лопастевидные выросты 1-го членика его задней лапки расположены заметно
дистальнее (т. е. более примитивно), чем у Schizodactylus и Comicus, и, таким образом,
2 последних рода явно синапоморфны по этому признаку в сравнении с Brauckmannia
и Gryllacridinae (ср. рис. 6, 1-3, 17).
Следует также отметить, что для Comicus было описано отдельное подсемейство
(Comicinae Ander, 1939, syn. n.), которое позднее было понижено в ранге до трибы
(Irish, 1986), а здесь синонимизируется с Shizodactylinae. Эта синонимизация вызвана
тем, что Comicus отличается от Schizodactylus в основном лишь редукцией некоторых
структур (исчезновением крыльев и заднего выступа выпуклости переднего грудного
дыхальца, но также утончением боковых лопастевидных выростов на лапках и, воз-
можно, ларвализацией гениталий самца; рис. 1, 16-20; 6, 3, 17) и обладает сходным
строением некоторых других копулятивных структур: последний тергит самца с до-
вольно крупной задней срединной лопастью (более длинной у Schizodactylus и ко-
роткой у Comicus), которая снабжена парой низких килей сверху-сзади, усаженных
мелкими щетинковидными шипиками, но у Schizodactylus эти кили широкие, упло-
щенные и расположены почти параллельно и близко один от другого (рис. 6, 4, 6),
тогда как у Comicus кили узкие, расположены в виде пары дуговидных складочек
у основания церок и далеко один от другого (рис. 6, 8); парапрокты самца с крючками,
направленными вверх, но у Schizodactylus эти парапрокты несколько удлиненные
и с мелким крючком на каждом (рис. 6, 4, 6), а у Comicus они короче и со сравнительно
более крупными крючками (рис. 6, 8); генитальная пластинка самца без грифельков,
но у Comicus довольно разнообразна по форме (рис. 6, 5, 7, 9, 14); у самок яйцеклад
полностью редуцирован - от него остаются лишь один видимый снизу стернит
брюшка (9-й) в форме поперечной частично склеротизованной складки, располо-
женной между парапроктами и генитальной пластинкой, и мембранозный рудимент
8-го стернита брюшка, полностью прикрытый снизу генитальной пластинкой; у Schi-
zodactylus эта пластинка снабжена срединным мягким язычком, выдающимся сзади за
ее задний край (рис. 4, 25, 26), а у Comicus она без такого язычка, но также довольно
разнообразна по форме (рис. 6, 11, 12, 15, 18-20).
Comicus orangensis Gorochov, sp. n. (рис. 1, 19, 20; 6, 8-13; 7, 1, 2; 8, 1).
Мат ер иа л. Южная Африка, штат «Northern Cape», ~80 км к западо-юго-западу от
г. «Upington», национальный парк «Augrabies», 300-400 м, полупустыня недалеко от р. Оранже-
вая, ночью на песке, 5-6.I.2008 (A. Горохов, A. Сочивко), 3 ♂ - голотип и паратипы, 1 ♀ - пара-
тип [ЗИН].
С ам ец (голотип). Тело маленькое для рода. Окраска желтоватая со следующими отметина-
ми (рис. 7, 1, 2; 8, 1): голова с почти темно-коричневыми глазами, коричневым полем между
глазами сверху и чуть позади середины глаз, 1 парой светло-коричневых вертикальных полосок
на эпикраниуме под глазами, темно-коричневыми вершинными частями мандибул и коричнева-
тыми вершинными частями максиллярных лациний; переднеспинка с коричневой узкой поло-
ской вдоль заднего края диска; другие тергиты со следами светло-коричневых полосок в их
верхней половине вдоль заднего края, но 3 последних тергита почти без затемнений; вершинные
части шипов и шпор на ногах светло-коричневые; крючки парапроктов коричневые. Голова
очень крупная, немного шире переднеспинки, примерно в 1.4 раза более высокая, чем широкая,
но очень короткая (уплощенная спереди) и без рострума (на его месте присутствует заметная
922
Рис. 6. Schizodactylinae. Два проксимальных членика задней лапки (1, 3, 17) и их отпечаток
(2) сверху; вершина брюшка самца без генитальной пластинки (4, 6, 8) и самки с генитальной
пластинкой (11, 18, 20) сверху и слегка сзади (4, 6), сзади и слегка сверху (8, 11, 18),
и сбоку (20); генитальная пластинка самца (5, 7, 9, 14) и самки (15) снизу; церк самца
(10, 16) и самки (13, 21) более или менее сбоку; генитальная пластинка, 9-й стернит
брюшка и парапрокты самки снизу и слегка сзади (12, 19) (схематично).
1 - Capnogryllacris sp.; 2 - Brauckmannia groeningae Martins-Neto; 3-5 - Schizodactylus
inexspectatus (Wern.); 6, 7 - Sch. hesperus B.-Bien.; 8-13 - Comicus orangensis sp. n.;
14-16 - C. myburghi sp. n.; 17-21 - C. namibicus sp. n. 2 - по фотографии: Heads, Leuzinger, 2011.
вогнутость над серединой усиковых впадин); минимальное расстояние между усиковыми впади-
нами почти вдвое меньше ширины скапуса; глаза крупные (высокие) и почти почковидные; глаз-
ков нет. Переднеспинка очень короткая, приблизительно в 1.3 раза короче головы; передняя
голень с 1 нижним внутренним, 1 нижним наружным и 1 верхним наружным небольшими пред-
вершинными шипами, а также с 5 вершинными шпорами, из которых 2 наружные и 1 нижняя
923
Рис. 7. Stenopelmatidae. Голова самца (1, 3) и самки (5) спереди; тело самца (2, 4) и самки (6)
сбоку; специализированные щетинки на нижнебоковой поверхности 2-го тергита брюшка
взрослой самки (7) и дейтонимфы самца (8, 9).
1, 2 - Comicus orangensis sp. n.; 3, 4 - C. myburghi sp. n.; 5, 6 - C. namibicus sp. n.; 7 - Maxentius
(Eumaxentius) pallidus (Walk.); 8 - M. (Maxentius) pinguis Walk. с нормальной (темной) окраской тела;
9 - M. (Maxentius) sp. со светлой окраской тела.
924
внутренняя незначительно крупнее шипов, а средняя и верхняя внутренние еще более длинные
(причем средняя из них - самая длинная); средняя голень со сходным вооружением, но нижний
внутренний и верхний наружный предвершинный шипы отсутствуют, а все вершинные шпоры
(3 наружные и 2 внутренние) более или менее сходны по длине с более короткими шпорами пе-
редней голени; задняя голень лишь с 4 небольшими верхними внутренними шипами, дисталь-
ный из которых расположен примерно на одинаковом расстоянии от ближайшего шипа и от
ближайшей шпоры, а также с 3 парами длинных вершинных шпор (эти шпоры длиннее всех
шпор передней голени, а самая длинная из них - средняя внутренняя - почти достигает вершины
базитарсуса; между этой и верхней внутренней шпорами развит короткий и очень тонкий допол-
нительный шипик); боковые лопасти на 1-3-м сегментах задней лапки приблизительно вдвое
короче таковых, изображенных на рис. 6, 17; все лапки с 1 парой длинных и тонких коготков, но
на задних лапках коготки примерно в 1.7 раза длиннее, чем на передних и средних. Феморо-аб-
доминальный стридуляционный аппарат редуцирован - от него остаются лишь следы в виде
редких и очень мелких бугорков вдоль задних краев 2-4-го тергитов брюшка; 10-й тергит брюш-
ка увеличен, с парой вздутий в верхнебоковых частях и довольно крупной (но неглубокой) во-
гнутостью между их медиальными краями, с очень короткой задней срединной лопастью,
которая преобразована в 1 пару почти пластинчатых округлых образований, направленных вниз
и в стороны и охваченных по бокам и сверху узкими валиками (килями), покрытыми мелкими
шипиковидными щетинками (рис. 6, 8); церк довольно длинный, с очень тонкой вершинной ча-
стью и заметным угловидным выступом около середины (немного ближе к основанию церка;
рис. 6, 10); эпипрокт очень маленький, частично покрыт сзади вышеупомянутой лопастью 10-го
тергита брюшка, с округлой вершиной; парапрокты округлые, но с довольно крупным и направ-
ленным вверх склеротизованным крючком на каждом (рис. 6, 8); генитальная пластинка овально
удлиненная, сужается к слабовыемчатой вершине и снабжена 1 парой почти зубчиковидных вы-
ступов (рис. 6, 9); гениталии мембранозные, но с полумембранозной и очень короткой средин-
ной лопастью на нижней стороне дорсальной складки (рис. 1, 19, 20).
В ар и аци и. Светло-коричневые полоски на тергитах бывают более заметны, плей-
риты заднегруди обычно частично затемнены, а угловидный выступ на церках иногда
расположен практически посередине церка.
С а мка. Похожа на более темного самца, но со следующими особенностями: расстояние меж-
ду усиковыми впадинами примерно в 1.7 раза меньше ширины скапуса; одна из передних голе-
ней без нижнего внутреннего предвершинного шипа; последний тергит брюшка несколько
уже и менее специализированный, чем у самца, но со следами рельефа, отдаленно напоминаю-
щими кили самца (рис. 6, 11); эпипрокт более крупный и округлый в дистальной части, слабо
обособлен от последнего тергита (частично слит с ним); парапрокты без крючков (рис. 6, 11, 12);
церки длинные, с очень тонкой вершинной частью, но без каких-либо специализаций
(рис. 6, 13); генитальная пластинка короткая и округленно-обрубленная на вершине, с 1 парой
почти овальных и блестящих (более склеротизованных) участков в заднебоковых частях
(рис. 6, 12); 9-й стернит брюшка в форме поперечной складки между генитальной пластинкой
и парапроктами (рис. 6, 12).
Длина (в мм). Тело: ♂ - 10.5-11, ♀ - 12.5; переднеспинка: ♂ - 1.6-1.7, ♀ - 1.8; переднее
бедро: ♂ - 4.6-4.8, ♀ - 5; заднее бедро: ♂ - 8.5-8.7, ♀ - 9; задняя голень: ♂ - 5.9-6.1, ♀ - 6.3;
задняя лапка: ♂ - 2.8-2.9, ♀ - 3.
С р ав не ние. Новый вид напоминает С. campestris Irish, 1986 по форме церка
самца, но отличается от него расположением угловидного выступа в середине или
перед серединой этого церка, а не позади его середины, а также отчетливо двузубой
вершиной генитальной пластинки самца и значительно более широкой генитальной
пластинкой самки. Самцы C. cavillorus Irish, 1986, C. calaharicus Irish, 1986, C. calcaris
Irish, 1986 и C. capensis Brunner-Wattenwyl, 1888 имеют сходный выступ на церке, но
он расположен еще более дистально, чем у обоих предыдущих видов, а генитальные
925
пластинки их самцов и самок также совсем другие. У C. cabonegrus Irish, 1986 и
C. arenarius Ramme, 1931 генитальная пластинка самки несколько похожа на таковую
нового вида, но c более заметной вершинной выемкой, а генитальные пластинки их
самцов совсем иные по форме; кроме того, задняя голень у C. cabonegrus без шипов,
а у С. arenarius эти шипы отделены от вершинных шпор значительно более длинным
промежутком. От C. carnalli Irish, 1995, самец которого неизвестен, новый вид легко
отличается генитальной пластинкой самки без глубокой задней срединной вырезки.
Эт им ол о гия. Видовое название образовано от названия р. Оранжевая (Orange
River).
Comicus myburghi Gorochov, sp. n. (рис. 6, 14-16; 7, 3, 4; 8, 2).
Мат ер иа л. Южная Африка, штат «Northern Cape», окрестности г. «Springbok», ферма
«Mr. Maritz Myburgh», 100-200 м, полупустыня (пастбище) с остатками воды в сайре и отдель-
ными деревьями рядом, 9-11.I.2008 (A. Горохов, A. Сочивко), 2 ♂ - голотип и паратип, 1 ♀ -
паратип [ЗИН].
С ам ец (голотип). Тело маленькое для рода. Окраска и строение тела похожи на таковые
C. orangensis sp. n., но со следующими отличиями (рис. 7, 3, 4; 8, 2): глаза, поле над глазами,
1 пара вертикальных полосок на эпикраниуме под глазами, пятна в срединной части диска пе-
реднеспинки, верхние половины боковых лопастей переднеспинки, задние части 1-6-го терги-
тов брюшка и крючки парапроктов светло-коричневые (затемненные участки переднеспинки
связаны чуть более темной полоской вдоль заднего края), а остальные части тела желтоватые;
минимальное расстояние между усиковыми впадинами приблизительно в 1.5 раза меньше ши-
рины скапуса; средняя голень с 2 парами небольших и примерно одинаковых по длине вершин-
ных шпор и лишь с 1 (наружным нижним) предвершинным шипом; задняя голень с 4 небольшими
верхними внутренними неотчлененными шипами (дистальный из них маленький и удален от
ближайшей шпоры на расстояние примерно вдвое большее, чем между этим и ближайшим ши-
пами, а самый проксимальный шип очень маленький); боковые лопасти на 1-3-м сегментах зад-
ней лапки примерно в 1.5 раза короче, чем таковые, изображенные на рис. 6, 17; коготки задней
лапки почти вдвое длиннее, чем таковые передней и средней лапок; последний тергит почти без
вздутий в верхнебоковых частях; церк без какого-либо выступа в средней или дистальной частях
(рис. 6, 16); генитальная пластинка (рис. 6, 14) сравнительно короткая, сужается к широкообру-
бленной вершине, несущей 1 пару коротких и зубчиковидных заднебоковых лопастинок (у сухо-
го экземпляра эта пластинка деформирована - продольно согнута таким образом, что эти
лопастинки более или менее сближены одна с другой).
Ва р иа ции. У другого самца затемнения над глазами и в средней части диска пе-
реднеспинки почти не выражены, затемнения на тергитах короче (в виде более узких
поперечных полосок вдоль задних краев), средняя голень с 3 небольшими наружными
вершинными шпорами, а задняя голень с дистальным шипом, примерно равным по
величине двум ближайшим к нему шипам.
Са м ка. Окраска и строение тела - примерно как у самцов, но затемненные полоски
под глазами расширены в нижней части (почти в форме треугольников), затемнения
над глазами в виде 4 коротких продольных полосок, окраска диска переднеспинки -
почти как у самца-паратипа, окраска тергитов - как у голотипа, вооружение средней и
задней голеней - как у самца-паратипа (но на одной из задних голеней 2 прокси-
мальных шипа маленькие), а расстояние между дистальным шипом задней голени и
ближайшей шпорой примерно в 1.5 раза больше, чем между этим и ближайшим ши-
пами. Вершина брюшка похожа на таковую самки C. orangensis sp. n., но генитальная
пластинка с немного более угловидными заднебоковыми участками (рис. 6, 15).
926
Рис. 8. Stenopelmatidae и Anostostomatidae, тело самца (1, 2) и самки (3) сверху; голова
и переднеспинка самца (4) и самки (6) сверху; тело самца (5) и самки (7) сбоку,
но без задней части и ног.
1 - Comicus orangensis sp. n.; 2 - C. myburghi sp. n.; 3 - C. namibicus sp. n.; 4, 5 - Anabropsis (Pteranabropsis)
intermedia sp. n.; 6, 7 - A. (P.) falcata pallescens subsp. n.
927
Длина (в мм). Тело: ♂ - 10.2-13.5, ♀ - 14.2; переднеспинка: ♂ - 1.7-1.8, ♀ - 1.9;
переднее бедро: ♂ - 4.2-4.4, ♀ - 4.5; заднее бедро: ♂ - 8-8.5, ♀ - 8.7; задняя
голень: ♂ - 6-6.5, ♀ - 6.2; задняя лапка: ♂ - 2.6-2.9.
Сравнение. Новый вид напоминает С. campestris Irish, 1986 по форме гени-
тальной пластинки самца, но эта пластинка менее короткая, с более развитыми задне-
боковыми лопастинками и с практически обрубленным (не волнистым) задним краем
между ними, а также церк самца без какого-либо выступа в средней части, и гени-
тальная пластинка самки значительно более широкая. От C. calcaris Irish, 1986, слегка
похожего на новый вид по форме генитальной пластинки самки, последний отлича-
ется совсем иной формой генитальной пластинки самца и отсутствием выступа в дис-
тальной части церка самца. От других видов рода новый вид легко отличается формой
генитальных пластинок самца и самки, отсутствием выступа на церке самца или нали-
чием шипов на задней голени.
Эт им ол о гия. Вид назван в честь хозяина фермы, мистера Маритца Майбурга
(Mr. Maritz Myburgh), любезно предоставившего нам возможность для изучения насе-
комых на своей территории.
Comicus namibicus Gorochov, sp. n. (рис. 6, 17-21; 7, 5, 6; 8, 3).
Мат ер иа л. Namibia, окр. заповедника «Gondwana Canyon» около национального парка
«Fish River Canyon», 200-300 м, пустыня у высохшего ручья, ночью на песке, 6-7.I.2008 (A. Го-
рохов, A. Сочивко), 1 ♀ - голотип [ЗИН].
С ам ка (голотип). Тело довольно крупное для рода. Окраска похожа на окраску C. orangensis
sp. n. - желтоватая с немногочисленными затемнениями (рис. 7, 5, 6; 8, 3): голова с темно-корич-
невыми глазами, вершинными частями мандибул и максиллярных лациний, а также с коричне-
вым небольшим вершинным участком лабрума и светло-коричневым полем на верхней части
эпикраниума между глаз (но позади их передних участков); переднеспинка со светло-коричне-
вым крупным пятном в верхней половине каждой боковой лопасти и 1 парой мелких попереч-
ных пятнышек на диске позади его середины; вершинные шпоры передних и средних голеней
светло-коричневые со слегка затемненными вершинами; 1-4-й тергиты брюшка с широкой
поперечной полосой вдоль заднего края каждый; дистальная половина эпипрокта коричневая
сверху. Строение тела (кроме вершины брюшка) также похоже на таковое этого вида, но со сле-
дующими отличиями: темя над (между) глазами с 1 парой слабых (сглаженных) продольных
бороздок; ростральная вогнутость менее развита (почти незаметна); минимальное расстояние
между усиковыми впадинами приблизительно в 1.4 раза больше ширины скапуса; длина перед-
неспинки примерно равна длине головы; передняя голень с 1 парой нижних предвершинных
шипов и 2 парами вершинных шпор, а вооружение средней голени отличается от вышеуказанно-
го лишь отсутствием 1 нижнего внутреннего предвершинного шипа (по величине шипы и шпо-
ры этих голеней близки к таковым C. orangensis sp. n.); задние голени с 4 небольшими
внутренними верхними шипами, расположенными довольно далеко от шпор (расстояние между
дистальным шипом и ближайшей шпорой приблизительно вдвое больше, чем между дисталь-
ным и ближайшим к нему шипами; рис. 7, 6), а также с 1 очень мелким верхним наружным ши-
пиком почти напротив вышеупомянутого дистального шипа на одной из голеней (на этой голени
2 проксимальных верхних внутренних шипа также очень мелкие) и 6 вершинными шпорами,
похожими на таковые C. orangensis sp. n.; все лапки с неукороченными боковыми лопастями
(рис. 6, 17) и с парными тонкими коготками, но на задней лапке коготки почти вдвое длиннее,
чем на передней и средней лапках. Вершина брюшка (рис. 6, 18-21) отличается от таковой
C. orangensis sp. n. главным образом несколько более длинным и узким эпипроктом со средин-
ной бороздкой на верхней поверхности, а также половыми признаками: генитальная пластинка
с сильно удлиненной и постепенно сужающейся к вершинной части срединной лопастью, при-
928
чем эта вершинная часть весьма узкая, загнута кверху и обрублена на вершине (рис. 6, 18-20);
9-й стернит брюшка в форме поперечной лопасти, которая слабо и округло выступает назад,
слегка утолщена по краям и расположена между генитальной пластинкой и парапроктами
(рис. 6, 19, 20).
Са ме ц неизвестен.
Длина (в мм). Тело - 14; переднеспинка - 2.4; переднее бедро - 5.6; заднее бедро - 10; задняя
голень - 7; задняя лапка - 4.
С р ав не ние. Новый вид по строению генитальной пластинки самки несколько на-
поминает С. capensis, но суженная задняя срединная часть этой пластинки заметно
уже и длиннее, окраска более однотонно светлая, а передняя голень с 2 (а не 3) пред-
вершинными шипами (не учитывая 2 пары вершинных шпор). От остальных видов
рода C. namibicus sp. n. легко отличается сильно удлиненной суженной срединной ча-
стью генитальной пластинки самки, которая в вершинной порции очень узкая и
загнута кверху, тогда как у других видов эта пластинка на вершине выемчатая, широ-
кообрубленная или заметно менее узкая и длинная.
Э ти моло ги я. Видовое название образовано от названия государства Намибия.
Подсем. GRYLLACRIDINAE - древесные лжекузнечики
Hyperbaeninae Cadena-Castañeda, 2019, syn. n.
Это подсемейство, иногда трактуемое как самостоятельное семейство (Cigliano
et al., 2020), недавно было подразделено на 2 подсемейства (номинативное и Hyper-
baeninae Cadena-Castañeda, 2019) и несколько триб (Cadena-Castañeda, 2019). Однако
такой дробительский подход находится в резком противоречии с классификацией
остальных групп длинноусых прямокрылых, поскольку морфологически подсем.
Gryllacridinae s. l. весьма близко к другим подсемействам Stenopelmatidae и довольно
однообразно (т. е. строение его тела менее разнообразно, чем у таких крупных подсе-
мейств Ensifera, как Podoscirtinae, Phaneropterinae и Сonocephalinae, и примерно соот-
ветствует по разнообразию Gryllinae, Trigonidiinae и Pleminiinae). Таким образом, даже
целесообразность деления Gryllacridinae на трибы сомнительна, но эта проблема тре-
бует отдельного исследования. Подобного мнения, очевидно, придерживается и
другой исследователь, также опубликовавший недавно крупную работу по таксономии
этого подсемейства (Ingrisch, 2018), в которой Gryllacridinae не подразделены на
какие-либо надродовые таксоны. Таким образом, Hyperbaeninae syn. n. здесь синони-
мизируются с Gryllacridinae.
Cем. ANOSTOSTOMATIDAE - лжесверчки
В настоящей работе это семейство понимается как родственное сем. Stenopelmatidae
и условно разбито лишь на 2 подсемейства: Anostostomatinae Saussure, 1859 и Lezininae
Karny, 1932. Ранее это семейство подразделялось на большее число подсемейств (Го-
рохов, 1988а, 1995а; Gorochov, 2001a), но, как и в случае с сем. Stenopelmatidae, ранг
многих из них кажется ныне несколько завышенным, поскольку морфологическое раз-
нообразие внутри этих таксонов и морфологические отличия между ними в целом не
превышают таковые для триб из семейств Tettigoniidae и Gryllidae. Условность де-
ления Anostostomatidae на 2 подсемейства обусловлена в основном лишь тем, что со-
гласно молекулярным данным филогенетическое положение Lezininae противоречиво:
929
одни авторы помещают эту группу в стороне от прочих Anostostomatidae (Jost, Shaw,
2006), видимо, считая ее более близкой к Stenopelmatidae; другие - сближают Lezininae
с некоторыми группами Anostostomatinae (Vandergast et al., 2017). Однако следует от-
метить, что морфологически Lezininae ничем особенно не выделяются среди прочих
групп этого семейства и вполне могут считаться лишь одной из триб Anostostomatinae
(различия между этими двумя условными подсемействами даны в разделе о Lezininae
ниже).
Подсем. ANOSTOSTOMATINAE - настоящие лжесверчки
Здесь это подсемейство предварительно разделено на 9 триб: Anostostomatini
Saussure, 1859; Cratomelini Brunner-Wattenwyl, 1888, stat. n.; Deinacridini Karny, 1932,
stat. n.; Glaphyrosomatini Rentz et Weissman, 1973; Anabropsini Rentz et Weissman, 1973;
Cooloolini Rentz, 1980, stat. n.; Lutosini Gorochov, 1988; Brachyporini Gorochov, 2001;
Leiomelini Gorochov, 2001, stat. n. Однако многие роды этого подсемейства недоста-
точно изучены и принадлежность их к какой-либо трибе неясна. Кроме того, специа-
лизированная роющая триба Cooloolini часто понимается даже как самостоятельное
семейство (Cigliano et al., 2020), хотя морфологические отличия этой трибы от других
триб не больше, чем различия между родами Maxentius и Oryctopus из разных триб
подсем. Stenopelmatinae, а по молекулярным данным она ближе к Anostostomatini и
Deinacridini, чем к Glaphyrosomatini (Song et al., 2015; Vandergast et al., 2017). Ниже
дана упрощенная таблица для определения триб Anostostomatinae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ТРИБ ПОДСЕМ. ANOSTOSTOMATINAE
1. Задние бедра с хорошо развитым перистым рельефом на наружной стороне
(рис. 8, 5, 7; 10, 4-6; 11, 1, 3, 5; 15, 1-3)
2.
– Задние бедра без такого рельефа или только с его следами (рис. 15, 5)
6.
2. Предпоследний тергит самца с 1 парой небольших (но заметных) или очень ма-
леньких (слабозаметных) задних лопастинок, разделенных довольно широким
промежутком; последний тергит самца не разделен на 2 половины мембраноз-
ным участком и несет 1 пару маленьких крючков, также отделенных один от
другого довольно широким промежутком (рис. 12, 2, 3, 6, 14, 19, 28)
................................................................................................................. Anabropsini.
- Предпоследний тергит самца разнообразный; последний тергит самца разделен на
2 половины мембранозным участком и несет 1 пару крючков довольно разной
величины и расположения
3.
3. Предпоследний тергит самца без крупных специализаций; медиальные части по-
следнего тергита самца с мелкими сближенными крючками и не отделены каки-
ми-либо полусклеротизованными участками от его латеральных частей; задние
крылья не превращены в стридуляционный орган
4.
– Предпоследний тергит самца разнообразный; последний тергит самца c крупными
крючками, а задние крылья у обоих полов превращены в стридуляционный
орган, либо медиальные части этого тергита (несущие крючки) отделены очень
узкими полусклеротизованными участками от его латеральных частей
5.
930
4. Тело в профиль более или менее дуговидное (переднеспинка в профиль с отчетливо
выпуклым верхним краем, а остальные тергиты груди с высокими боковыми ча-
стями, расположенными примерно как в сем. Rhaphidophoridae); мандибулы без
половых специализаций - одинаковые у самца и самки; крылья отсутствуют;
тимпанумы и феморо-абдоминальный стридуляционный аппарат обычно разви-
ты
Lutosini.
– Тело в профиль не дуговидное (переднеспинка в профиль с почти прямым верхним
краем, а остальные тергиты груди с низкими боковыми частями по крайней мере
у бескрылых форм и расположены таким образом, что их верхние края в про-
филь образуют прямую или слегка вогнутую линию); мандибулы самца обычно
с половой специализацией (удлиненные и дуговидно изогнутые или с дополни-
тельными выростами) - не похожие на таковые самки; крылья, тимпанумы и
феморо-абдоминальный стридуляционный аппарат развиты или отсутствуют
.......................................................................................................... Anostostomatini.
5. Предпоследний тергит самца разнообразный; задние крылья у обоих полов превра-
щены в стридуляционный орган; медиальные части последнего тергита самца
никак не обособлены от его латеральных частей и с крупными крючками
................................................................................................................. Cratomelini.
- Предпоследний тергит самца с 1 парой задних лопастинок или короткой непарной
задней лопастью; крылья отсутствуют; медиальные части последнего тергита
самца с небольшими или мелкими крючками и отделены очень узкими полу-
склеротизованными участками от его латеральных частей
Brachyporini.
6. Ноги типичного для подсемейства строения (не преобразованы в мощные копа-
тельные органы), т. е. с более или менее тонкими голенями, снабженными сла-
быми (тонкими или маленькими) вершинными шпорами, и с нормальным
вооружением задних голеней (их верхние шипы развиты как на внутренней, так
и на наружной сторонах); крылья отсутствуют; тимпанумы и феморо-абдоми-
нальный стридуляционный аппарат обычно развиты
7.
– Ноги в той или иной степени специализированы: преобразованы в мощные копа-
тельные органы с утолщенными голенями, снабженными лишь крепкими вер-
шинными шпорами, или тонкие, со слабыми шпорами и с мелкими шипиками
на месте верхних шипов лишь на внутренней стороне задней голени; крылья
развиты хотя бы у самца; тимпанумы и феморо-абдоминальный стридуляцион-
ный аппарат отсутствуют
8.
7. Задние бедра прыгательные - заметно утолщенные в проксимальной половине;
предпоследний тергит самца с 1 парой коротких задних лопастей либо с 1 такой
лопастью, часто снабженной выемкой посередине; крючки последнего тергита
самца разнообразные по величине
Glaphyrosomatini.
– Задние бедра практически не прыгательные - слабо утолщенные в проксимальной
половине; предпоследний тергит самца без каких-либо задних лопастей либо с
маленькой и очень короткой непарной задней лопастинкой; крючки последнего
тергита самца маленькие
Deinacridini.
931
8. Ноги преобразованы в мощные копательные органы (см. антитезу 6); задняя голень
без верхних шипов (только с вершинными шпорами); самец со значительно уко-
роченными крыльями, самка бескрылая
Cooloolini.
– Ноги тонкие, со слабыми шпорами; задняя голень с мелкими шипиками на месте
верхних шипов лишь на внутренней стороне (но вершинные шпоры присутству-
ют); крылья развиты, неукороченные
Leiomelini.
Род ANABROPSIS Rehn, 1901
Этот род, относящийся к трибе Anabropsini, отличается от Melanabropsis Wang
et Liu, 2020 (другого рода, возможно, принадлежащего к данной трибе) следующими
признаками: стерниты передне- и среднегруди с парными шиповидными выростами;
стернит заднегруди крупнее стернита среднегруди и с 1 парой крупных выступов или
выростов (рис. 9, 1, 2, 6, 8, 10, 12, 13, 15, 17, 21, 22, 24, 25, 27, 30, 31); предпоследний
тергит самца с 1 парой широко расставленных задних лопастинок в верхней части;
генитальная пластинка самца со сравнительно маленькими грифельками, которые не
сближены один с другим (рис. 9, 3-5, 7, 9, 11, 16, 18-20, 23, 26, 28, 29, 32-36). От
Exogryllacris Willemse, 1963 (еще одного возможного рода этой трибы из Австралии)
длиннокрылые виды рода Anabropsis отличаются почти не расширенными в дис-
тальной половине надкрыльями, но другие различия между этими родами неясны.
Ранее род Anabropsis был разбит на 5 родов (Горохов, 1988а; Griffini, 1913; Shi, Bian,
2016): Anabropsis; Paterdecolyus Griffini,
1913; Pteranabropsis Gorochov,
1988;
Apteranabropsis Gorochov, 1988; Brevipenna Shi et Bian, 2016. Первый из них объединял
американские виды, а остальные - азиатские, что вполне логично, поскольку, если не
учитывать связанные с редукцией крыльев признаки, азиатские виды несколько более
разнообразны по морфологии, чем американские. Более того, короткокрылые и бес-
крылые американские виды, вероятно, произошли от общего длиннокрылого предка,
который смог достичь Америки уже после того, как началась диверсификация этой
группы в Старом Свете.
Род Anabropsis, скорее всего, относительно молодой, так как морфологические раз-
личия между всеми его видами по копулятивным признакам незначительны, да и по
другим особенностям внешнего строения внутриродовая дифференциация была свя-
зана главным образом с различными редукциями крыльев и тимпанумов, с частичной
или полной редукцией плечевых выемок переднеспинки у нелетающих форм и с изме-
нениями в размерах и окраске тела. Остальные различия также невелики, и наиболее
важные из них, как мне теперь кажется, характеризуют лишь подроды. Эти различия
даны ниже в определительной таблице подродов рода Anabropsis, и следует отметить,
что в этой таблице бывший род Apteranabropsis, недавно снова объединенный с
Paterdecolyus (Wang, Liu, 2020), рассматривается как подрод, 1 родовое название сво-
дится в синонимы и еще 1 подрод описывается как новый для науки.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ РОДА ANABROPSIS
1. Стернит заднегруди с 1 парой длинных пальцевидных выступов (рис. 9, 1, 2, 6,
8, 10); генитальная пластинка самца с почти прямым задним краем между гри-
фельками, либо этот край слабо вогнут или с неглубокой срединной выемкой
(рис. 9, 3-5, 7, 9, 11)
2.
932
Рис. 9. Anabropsis spp, стернит заднегруди снизу (1, 2, 6, 8, 10, 12, 13, 15, 17, 21, 22, 24, 25, 27,
30, 31), генитальная пластинка самца (3, 7, 9, 11, 14, 16, 20, 23, 26, 28, 29, 32, 35)
и ее задняя часть (4, 5, 18, 19, 33, 34, 36) снизу.
1-5 - A. (Pteranabropsis) carli Griff.; 6, 7 - A. (P.) intermedia sp. n.; 8, 9 - A. (P.) falcata pallescens subsp. n.;
10, 11 - A. (Paterdecolyus) panteli (Griff.); 12 - A. (Anabropsis) longipenna Gor. et Cad.-Cast.;
13, 14 - A. (A.) mexicana Sauss.; 15, 16 - A. (Carnabropsis) pusilla (Ingr.); 17-20 - A. (C.) carnarius (Gor.);
21 - A. (Apteranabropsis) cervicornis Karny; 22, 23 - A. (A.) tonkinensis Rehn; 24 - A. (A.) miser B.-Bien.;
25, 26 - A. (A.) ailaoshanica sp. n.; 27, 28 - A. (A.) tarasovi sp. n.; 29 - A. (A.) abramovi sp. n.;
30 - A. (A.) costulata (Gor.); 31-34 - A. (A.) minuta (Gor.); 35, 36 - A. (A.) sinica B.-Bien. 1, 3, 10, 13, 17,
20-24, 30, 31 - по: Горохов, 1998; Gorochov, 2001b, с изменениями.
933
- Стернит заднегруди с 1 парой коротких или умеренно удлиненных угловидных вы-
ступов сзади (рис. 9, 12, 13, 15, 17, 21, 22, 24, 25, 27, 30, 31); генитальная пла-
стинка - как в предыдущей тезе либо с более или менее глубокой срединной
выемкой между грифельками (рис. 9, 14, 16, 18-20, 23, 26, 28, 29, 32-36)
3.
2. Тело от средних до крупных размеров (длина переднеспинки 6.8-11 мм); передняя
голень с 2 хорошо развитыми тимпанумами; крылья длинные или умеренно
укороченные (обычно не короче или незначительно короче переднеспинки;
рис. 8, 5, 7)
подрод Pteranabropsis stat. n. (= Brevipenna syn. n.).
[Cостав: типовой вид Pteranabropsis - Anabropsis carli Griffini, 1911; типовой вид
Brevipenna - B. falcata Shi et Bian, 2016; Pteranabropsis angusta Ingrisch, 2019; P. bavi
Ingrisch, 2019; P. copia Ingrisch, 2019; P. cuspis Ingrisch, 2019; A. (P.) intermedia sp. n.]
- Тело от мелких до средних размеров (длина переднеспинки 5.3-7.5 мм); передняя
голень лишь с внутренним тимпанумом; крылья сильно укороченные (намного
короче переднеспинки) или отсутствуют
подрод Paterdecolyus stat. n.
[Cостав: типовой вид - Paterdecolyus panteli Griffini, 1913; Anabropsis griffini Karny,
1930, синонимизированный с типовым видом (Johns, 1997); P. magnimaculatus Bian
et Shi, 2019; возможно, P. dubius Würmli, 1973.]
3. Стернит заднегруди с короткими (почти угловидными) задними выступами
(рис. 9, 12, 13, 21, 22, 24, 25, 27, 30, 31); генитальная пластинка самца с прямым
задним краем между грифельками (рис. 9, 26, 28, 34), либо этот край слегка во-
гнут (рис. 9, 23, 36) или со срединной выемкой, но в последнем случае эта выем-
ка от малозаметной (очень маленькой и округленной или угловидной; рис. 9, 32,
33, 35) до умеренно глубокой и довольно широкой (рис. 9, 14, 29)
4.
– Стернит заднегруди со слабо или умеренно удлиненными задними выступами
(рис. 9, 15, 17); генитальная пластинка самца с умеренно глубокой (но узкой)
или глубокой задней срединной выемкой (рис. 9, 16, 18-20)
................................................................ подрод Carnabropsis Gorochov, subgen. n.
[Состав: типовой вид - Pteranabropsis carnarius Gorochov, 1998; P. tenchongensis
Wang, Liu et Li, 2015; P. infuscatus Wang, Liu et Li, 2015; P. karnyi Wang, Liu et Li, 2015;
P. parallelus Wang, Liu et Li, 2015; P. crenatis Song, Bian et Shi, 2016; P. incisa Song, Bian
et Shi, 2016; P. guadun Ingrisch, 2019; P. pusilla Ingrisch, 2019. Этимология: название
подрода - существительное женского рода, образованное как сочетание частей родо-
вого и видового названий типового вида.]
4. Передняя голень с 2 хорошо развитыми тимпанумами, но иногда лишь с 1 таким
тимпанумом (внутренним), а наружный тимпанум недостаточно отчетливый,
значительно уменьшенный или отсутствует; крылья сильно укорочены (значи-
тельно короче переднеспинки), рудиментарны (рис. 12, 1, 13) или практически
отсутствуют (рис. 12, 29); генитальная пластинка заметно удлиненная и значи-
тельно суженная в дистальной части (рис. 9, 23, 26, 28, 32, 35), но у одного вида
слабо удлиненная и слабо суженная в дистальной части, причем с довольно
широкой и неглубокой вершинной выемкой и срединным бугорком на ее дне
(рис. 9, 29)
подрод Apteranabropsis stat. n.
934
[Состав: типовой вид - Anabropsis miser Bey-Bienko, 1968; A. tonkinensis Rehn, 1906;
A. cervicornis Karny, 1930; A. sinica Bey-Bienko, 1962; Apteranabropsis costulata
Gorochov, 2001; A. minuta Gorochov, 2001; A. paracostulata Gorochov, 2010; A. guang-
xiensis Bian et Shi, 2015; 4 новые вида, описываемые ниже.]
- Передняя голень с 2 хорошо развитыми тимпанумами; крылья разнообразны (от
длинных - полностью развитых - до рудиментарных), но иногда, возможно от-
сутствуют; генитальная пластинка самца короткая или слабо удлиненная, со сла-
бо суженной дистальной частью (вершинная выемка этой пластинки, если
развита, то округлая либо более или менее угловидная; рис. 9, 14)
..................................................................................................... подрод Anabropsis.
[Состав: типовой вид - Schoenobates mexicanus Saussure, 1859; все другие американ-
ские виды рода. Отличия Anabropsis s. str. от Apteranabropsis недостаточно отчетливы,
но сходство между этими подродами, вероятно, обусловлено лишь симплезиомор-
фиями - американские виды, скорее всего, дивергировали независимо от азиатских,
что может быть подтверждено или опровергнуто молекулярными исследованиями.]
Подродовая принадлежность некоторых других видов неясна. Anabropsis tibetensis
(Wang, Liu et Li, 2015), comb. n., cудя по фотографии (Wang et al., 2015: fig. E,
Pteranabropsis), внешне очень похож на виды подрода Carnabropsis subgen. n., но
генитальная пластинка его самца - как у других подродов рода Anabropsis; кроме
того, в подписи к этой фотографии написано «Male», но изображена самка (т. е. экзем-
пляры могли быть перепутаны). Для A. femorata (Pictet et Saussure, 1893) и A. frater
(Brunner-Wattenwyl, 1888), описанных из «Les Indes orientalis» (Pictet, Saussure,
1893: Onosandrus) и «Hinterindien» (Brunner-Wattenwyl, 1888: Schoenobates) соответ-
ственно, известны только такие признаки, которые позволяют включить их лишь
в Anabropsis s. l. Единственный вид из Африки (A. rehni Griffini, 1909) также не может
быть отнесен к какому-либо из подродов без дополнительного изучения. Голотип
A. chopardi Karny, 1932 (см. Desutter-Grandcolas, 2020b), происходящий якобы из
Мадагаскара, чрезвычайно похож на А. tonkinensis из Вьетнама, т. е. самостоятель-
ность и типовая местность A. chopardi проблематичны.
Anabropsis (Pteranabropsis) intermedia Gorochov, sp. n. (рис. 8, 4, 5; 9, 6, 7; 10, 1;
12, 2; 13, 3).
М ат е риа л. Вьетнам, провинция «Lao Cai», гора «Phan Si Pan», лес в окрестностях пос. «Sa
Pa», ~1400 м, V.2003 (Н. Орлов, С. Рябов), 1 ♂ - голотип [ЗИН].
С ам е ц (голотип). Тело небольшое для рода. Окраска темно-серо-коричневая со следующим
рисунком (рис. 8, 4, 5): голова с почти черными глазами, светло-коричневыми боковыми глазка-
ми, 1 парой розовых узких вертикальных полосок вдоль боковых килей рострального бугорка, с
желтоватыми пятном под этим бугорком и точками на остальной части эпикраниума и клипеусе
(на клипеусе эти точки редкие, а на эпикраниуме - многочисленные), со светло-коричневыми в
целом максиллярными пальпами (их вершинный членик почти желтоватый, а предвершинный -
серо-коричневый) и с мелкими и редкими желтоватыми и светло-коричневыми пятнышками на
усиках (но не на скапусах); переднеспинка коричневая с рыжеватым оттенком и умеренно мно-
гочисленными желтоватыми точками; надкрылья серо-коричневые с полупрозрачными косталь-
ными полями и редкими беловатыми пятнышками на остальных частях (рис. 10, 1); ноги похожи
по окраске на переднеспинку, но тимпанальные мембраны темно-коричневые, а дистальная по-
ловина заднего бедра более однотонная и с желтоватым верхним пятном на вершинном участке;
брюшко темно-коричневое с многочисленными светло-коричневыми и желтоватыми пятнышка-
935
Рис. 10. Anabropsis spp., правое надкрылье (1, 2; стрелки указывают на интермедиальное и
интеркубитальное поля); яйцеклад сбоку (3); тело самки (4) и самца (5, 6) сверху.
1 - A. (Pteranabropsis) intermedia sp. n.; 2, 3 - A. (P.) falcata pallescens subsp. n.; 4 - A. (Apteranabropsis)
maculata sp. n.; 5 - A. (A.) minuta (Gor.); 6 - A. (A.) ailaoshanica sp. n.
ми, но 2 последних тергита брюшка, церки, эпипрокт, парапрокты и генитальная пластинка
однотонные, а задние лопастинки предпоследнего тергита черноватые. Строение тела очень по-
хоже на таковое A. (P.) carli, но с некоторыми отличиями: переднеспинка с очень слабо выражен-
ными плечевыми выемками (рис. 8, 5); стернит переднегруди с заостренными шипами, которые
существенно короче, чем шипы стернита среднегруди, а последние шипы лишь незначительно
короче выростов стернита заднегруди (рис. 9, 6) и так же закруглены на вершине (у A. carli все
шипы и выросты стернитов груди не заостренные, а разница в длине между шипами стернита
среднегруди и шипами и выростами других грудных стернитов примерно одинаковая); крылья
укорочены и достигают лишь 8-го тергита брюшка (рис. 8, 5), причем костальное поле надкры-
лья почти вдвое уже, чем у этого вида, а поперечные жилки в остальной части надкрылья более
регулярные и более частые (рис. 10, 1); передние бедра с 3 или 4 нижними внутренними неотчле-
ненным шипиками, а средние - с 1 нижним наружным шипиком, сходным по строению с преды-
936
дущими, или без него; задние бедра с 4 или 5 наружными и 3-6 внутренними нижними
неотчлененными шипиками; передняя голень с 2 хорошо развитыми тимпанумами практически
одинакового размера (у A. carli тимпанумы обычно тоже одинакового размера, но иногда наруж-
ный чуть меньше внутреннего); задняя голень с 10 наружными и 10 внутренними верхними не-
отчлененными шипами, а также с 2 неотчлененными нижними шипиками; лопастинки
предпоследнего и крючки последнего тергитов очень похожи на таковые A. carli, но слегка коро-
че (рис. 12, 2); эпипрокт также незначительно короче (в форме почти равностороннего треуголь-
ника); парапроктальные выросты незначительно толще и с немного более загнутыми вверх
вершинными частями (основания этих частей с менее выпуклыми нижнезадними расширения-
ми; см. рис. 13, 1, 2 и 13, 3); генитальная пластинка со слегка суженной вершинной (но не пред-
вершинной) порцией, а расстояние между грифельками примерно равно длине грифелька
(у A. carli это расстояние приблизительно равно длине грифелька или немного меньше;
рис. 9, 3-5 и 9, 7); гениталии практически неотличимы от таковых A. (P.) carli (рис. 1, 5).
С а мка неизвестна.
Длина (в мм). Тело - 28; переднеспинка - 8.1; надкрылье - 15.5; переднее бедро - 11.5; заднее
бедро - 26.5; задняя голень - 25; задняя лапка - 7.
С р ав не ние. Новый вид наиболее похож на A. (P.) falcata из Юньнани, но отлича-
ется от него чуть более развитыми плечевыми выемками переднеспинки, немного
более узкими выростами стернита заднегруди, заметно более длинными крыльями
(у A. falcata они незначительно короче переднеспинки) и суженной вершинной пор-
цией генитальной пластинки самца (у нового вида расстояние между основаниями
грифельков примерно равно длине грифелька, но у A. falcata вершинная порция гени-
тальной пластинки не сужена, и длина грифелька отчетливо меньше расстояния между
основаниями грифельков). От остальных представителей подрода новый вид отлича-
ется главным образом более мелкими размерами, слаборазвитой плечевой выемкой и
значительно более короткими крыльями, а также некоторыми другими перечислен-
ными в описании признаками, отличающими новый вид от A. (P.) carli.
Этимология. Видовое название «intermedia» («промежуточная» на латыни) дано
в связи с промежуточной длиной крыльев между короткокрылым A. (P.) falcatа и
остальными известными представителями подрода с более или менее нормально раз-
витыми крыльями.
Anabropsis (Pteranabropsis) falcata (Shi et Bian, 2016), comb. n. (рис. 13, 36).
М ате риа л. Китай, провинция Юньнань: хребет
«Wuliangshan»,
24º45ʹ02ʹʹ с. ш.,
100º30ʹ24ʹʹ в. д., 2270 м, 12.VI. 2011 (И. Белоусов, И. Кабак, А. Королев), 1 ♂, 6 ♀ [ЗИН];
к востоко-северо-востоку от г. «Lincang», 23º54ʹ58ʹʹ с. ш., 100º18ʹ33ʹʹ в. д., 2190 м, 25.VI.2011
(И. Белоусов, И. Кабак, А. Королев), 1 ♂, 4 ♀ [ЗИН]; к юго-юго-востоку от г. «Shuangjiang»,
23º23ʹ19ʹʹ с. ш., 99º55ʹ28ʹʹ в. д., 2255 м, 21.VI.2011 (И. Белоусов, И. Кабак, А. Королев), 6 ♀ [ЗИН];
к юго-западу от г. «Dali», 20.V.2019 (И. Белоусов, И. Кабак, Г. Давидьян), 2 ♀ [ЗИН].
Эти экземпляры вполне соответствуют первоописанию этого вида, который был
описан в составе монотипического рода Brevipenna (Shi, Bian, 2016).
Anabropsis (Pteranabropsis) falcata pallescens Gorochov, subsp. n. (рис. 8, 6, 7; 9, 8,
9; 10, 2, 3; 12, 3-5; 13, 4-7).
М ате риа л. Китай, провинция Юньнань, хребет «Ailaoshan» к западу от г. «Shuitangzhen»,
24º07ʹ18ʹʹ с. ш., 101º27ʹ44ʹʹ в. д., 1965-2300 м, 1.VI.2011 (И. Белоусов, И. Кабак, А. Королев),
2 ♂ - голотип и паратип, 1 ♀ - паратип [ЗИН]; тот же хребет, но к северо-западу от
937
г. «Shuitangzhen», 24º09ʹ51ʹʹ с. ш., 101º25ʹ29ʹʹ в. д., 2005 м, 7.VI.2011 (И. Белоусов, И. Кабак,
А. Королев), 1 ♀ - паратип [ЗИН].
С ам ец (голотип). Тело немного меньше, чем у A. (P.) intermedia sp. n. Окраска светлее, чем у
этого вида (рис. 8, 6, 7): голова серо-коричневая с серовато-желтоватыми передней поверхно-
стью рострального бугорка, участком над этим бугорком (эти светлые части практически не со-
прикасаются одна с другой) и нижней частью эпикраниума (ниже глаз и усиковых впадин, но
с вертикальной срединной серо-коричневой полосой), желтоватыми точками на серо-коричне-
вых участках, беловатыми глазками, светло-коричневыми глазами, ротовыми частями (но вер-
шины мандибул затемненные), скапусом и педицелом (скапус и педицел, а также жгутик усиков
с мелкими желтоватыми отметинами, более многочисленными в проксимальной части усиков и
более редкими - в средней и дистальной частях); переднеспинка светло-коричневая с серо-
коричневой средней частью диска, а также с многочисленными светло-коричневыми точками на
этой более темной части; надкрылья по окраске - как у A. (P.) intermedia sp. n., но беловатые
пятнышки немного более редкие (менее заметные; рис. 10, 2); ноги также светло-коричневые
с малозаметными желтоватыми пятнышками, а заднее бедро с коричневыми продольной средин-
ной линией на наружной поверхности и боковыми участками в вершинной порции; грудь и
брюшко более или менее светло-коричневые, но стерниты груди почти желтоватые, а яйцеклад
коричневый со светло-коричневым основанием. Строение тела отличается от такового A. (P.) in-
termedia sp. n. следующими признаками: шипы и выросты всех стернитов груди - примерно как
у А. (P.) carli (но выросты стернита заднегруди немного более сближены один с другим, чем у
A. intermedia sp. n. и некоторых самцов A. carli; ср. рис. 9, 2, 6 и 9, 8); надкрылья более короткие
и узкие, достигают лишь 4-го тергита брюшка (рис. 8, 7), с интеркубитальным полем почти
вдвое более узким, чем интеррадиальное поле (у A. intermedia sp. n. эти поля примерно равны по
максимальной ширине; рис. 10, 1 и 10, 2); передние бедра с 2 или 3 нижними внутренними ши-
пиками; средние бедра без шипов и шипиков; задние бедра с 4 наружными и 6 или 7 внутрен-
ними нижними неотчлененными шипами; задние голени с 3-5 нижними шипиками, 9 или
10 верхними внутренними и 10 верхними наружными шипами; предпоследний тергит с хорошо
развитыми, но немного менее широкими лопастинками (рис. 12, 3), крючки последнего тергита
незначительно мельче, чем у A. (P.) intermedia sp. n., но очень похожи по форме (рис. 12, 4, 5);
эпипрокт и парапроктальные выросты - как на рис. 12, 3 и 13, 4; генитальная пластинка почти
неотличима от таковой номинативного подвида (рис. 9, 9); гениталии отличаются от таковых
A. (P.) carli менее глубокой задней срединной выемкой дорсальной складки и немного более ко-
роткими лопастями вентральной складки, которые отделены одна от другой более глубокой вы-
емкой (ср. рис. 1, 5 и 13, 5, 6).
Ва р иа ции. Второй самец со слегка более темной (серо-коричневой) нижней ча-
стью эпикраниума, с надкрыльями достигающими лишь середины 4-го тергита
брюшка, и с незначительно отличающимся числом шипиков и шипов на ногах.
Са м ка. Тело похоже на таковое самцов; окраска - как у самца-паратипа, причем у одной из
самок переднеспинка более светлая (желтоватая) и с 1 парой очень крупных и отчетливых почти
темно-коричневых пятен (эти пятна снабжены желтоватыми точками и отделены одно от другого
узкой срединной полоской); у другой самки передние бедра без шипиков. Вершина брюшка ти-
пичная для самок этого рода (тергиты и парапрокты не специализированы - без лопастинок,
крючков и выростов; эпипрокт - примерно как у самца A. intermedia sp. n.), но генитальная пла-
стинка узкая и длинная, с шиповидной дистальной частью, которая лишь незначительно короче
остальной части (рис. 13, 7), а яйцеклад длинный, тонкий, слабодуговидный и с узкозакруглен-
ной вершиной (рис. 10, 3).
Длина (в мм). Тело: ♂ - 21.8-24, ♀ - 24.5-27.5; переднеспинка: ♂ - 5.5-6, ♀ - 6.2-6.8; над-
крылье: ♂ - 10-11, ♀ - 9-11; переднее бедро: ♂ - 9-10, ♀ - 9-10.4; заднее бедро: ♂ - 20.5-23,
♀ - 20.7-23.5; задняя голень: ♂ - 19-22.5, ♀ - 20.5-23; задняя лапка: ♂ - 8.3-9, ♀ - 8; яйцеклад -
16-18.
938
Сравнение. От A. (P.) f. falcata новый подвид отличается заметно более круп-
ными (длинными) крыльями (у нового подвида надкрылья отчетливо длиннее перед-
неспинки, а у A. f. falcata они примерно равны ей по длине) и более узкой и длинной
генитальной пластинкой самки (ср. рис. 13, 7 и 13, 36). От A. (P.) intermedia sp. n.
новый подвид отличается признаками, перечисленными в его описании, включая явно
более светлую окраску нижней части тела; от других представителей подрода новый
подвид отличается меньшими размерами тела, редукцией плечевых выемок передне-
спинки, значительно укороченными крыльями и менее расширенными у вершины па-
рапроктальными выростами самца.
Этимология. Название нового подвида «pallescens» («светлеющая» на латыни)
дано в связи со светловатой окраской нижней части тела.
Anabropsis (Pteranabropsis) carli Griffini, 1911 (рис. 1, 5; 9, 1-5; 13, 1, 2).
В коллекции ЗИН хранится довольно представительный материал по этому виду -
около 60 экз. обоих полов из разных точек китайской провинции Юньнань и 3 се-
верных вьетнамских провинций: «Vinh Phu», «Lao Cai» и «Cao Bang». Эти экземпляры
несколько варьируют по величине, окраске, расстоянию между вершинами выростов
стернитов заднегруди (рис. 9, 1, 2), форме вершинной части парапроктальных выро-
стов самца (рис. 13, 1, 2) и генитальной пластинки самца (рис. 9, 3-5), а также, но
менее значительно, варьируют по форме генитальной пластинки самки и длине яй-
цеклада. Эти вариации иногда достигают различий, указанных С. Ингришем (Ingrisch,
2019) для видов, описанных им из северной части Вьетнама: A. (P.) copia comb. n.,
A. (P.) angusta comb. n., A. (P.) bavi comb. n., A. (P.) cuspis comb. n. Однако по длине
крыльев экземпляры в коллекции ЗИН довольно сходны - вершины крыльев у всех
заметно заходят за вершины задних бедер, а у A. angusta, A. bavi и A. cuspis крылья
достигают или немного не достигают вершин этих бедер. В связи с этим я допускаю,
что последние виды могут быть синонимами или подвидами особого (близкого
к A. carli) вида, а A. copia - лишь подвидом A. carli, но для решения этой проблемы
необходимо изучение дополнительного материала.
В мае-июне в окрестностях с. «Tam Dao» (вьетнамская провинция «Vinh Phu»)
первоначально наблюдалось множество нимф старших возрастов двух видов
A. (P.) carli и А. (C.) carnarius, сидящих на ветвях в подлеске. Затем эти нимфы начали
линять на имаго, причем имаго A. carnarius продолжали сидеть на тех же местах,
а имаго А. carli куда-то исчезали, хотя в садках линька обоих видов проходила одно-
временно. Возможно, имаго А. carli после линьки сразу улетали, что менее вероятно,
или перемещались в более высокий ярус леса. Было бы интересно проверить эти
данные для других видов подродов Pteranabropsis и Carnabropsis subgen. n.
Anabropsis (Carnabropsis) carnarius Gorochov, 1998 (рис. 9, 17-20; 13, 8).
Этот вид также хорошо представлен в коллекции ЗИН - около 50 экз. из следующих
северных провинций Вьетнама: «Ba Vi», «Lao Cai», «Vinh Phu» и «Cao Bang». Самцы
и самки из этих мест почти столь же вариабельны, как и A. (P.) carli (рис. 9, 18-20).
Однако в последние годы было описано множество новых видов из Южного Китая и
северного Вьетнама (Wang et al., 2015; Song et al., 2016; Ingrisch, 2019), явно близких к
A. (C.) carnarius и с отличиями, которые укладываются в рамки изменчивости
A. carnarius или лишь незначительно превышают ее: A. (C.) tenchongensis comb. n.,
939
A. (C.) infuscatus comb. n., A. (C.) karnyi comb. n., A. (C.) parallelus comb. n.,
A. (C.) crenatis comb. n., A. (C.) incisa comb. n., A. (C.) guadun comb. n., A. (C.) pusilla
comb. n. Для A. pusilla видовая самостоятельность относительно A. carnarius не вызы-
вает сомнений (см. ниже), но остальные виды нуждаются в проверке их вариабель-
ности и возможной синонимии с двумя последними видами.
Anabropsis (Carnabropsis) pusilla Ingrisch, 2019 (рис. 9, 15, 16; 13, 9).
Мат ер иа л. Вьетнам: провинция «Lao Cai», 6 км к З от пос. «Sa Pa», северный склон горы
«Phan Si Pan», национальный парк «Hoang Lien», станция «Tram Ton», 22º21ʹ с. ш., 103º46ʹ в. д.,
1930-2000 м, V.2008 (А. Абрамов), 1 ♂ [ЗИН]. Провинция «Cao Bang», гора «Phia Oac», 1600 м,
16.VII.2009 (М. Багатуров), 1 ♀ [ЗИН]; те же провинция и гора, но национальный парк «Phia
Oac - Phia Den» на восточном склоне (40 км к западу от г. «Cao Bang»), 22º36ʹ27ʹʹ с. ш., 105º52ʹ00ʹʹ
в. д., 1600-1800 м, 22.V-6.VI.2018 (Л. Анисюткин, А. Абрамов), 1 ♀ [ЗИН]; те же данные, но
22º37ʹ41.8ʹʹ c. ш., 105º54ʹ41.5ʹʹ в. д., 900 м, 3-15.VI.2019 (Л. Анисюткин), 1 ♀ [ЗИН].
Этот вид отличается от A. (C.) carnarius более мелкими размерами (длина передне-
спинки у A. pusilla - 6.8-8.1 мм, а у A. carnarius - 9.5-12 мм), генитальной пластинкой
самца с более крупной (глубокой и широкой) задней срединной выемкой и с выступа-
ющими назад (позади основания грифельков) округлыми медиальными выступами
между этой выемкой и грифельками (ср. рис. 9, 16 и 9, 18-20), почти не изогнутыми
вверх выростами парапроктов самца (рис. 13, 8 и 13, 9), а также более короткими кры-
льями у самки (у нее крылья незначительно заходят за вершины задних бедер или
только достигают их, тогда как у самца этого вида и у обоих полов A. carnarius крылья
значительно заходят за эти вершины).
Anabropsis (Apteranabropsis) ailaoshanica Gorochov, sp. n. (рис. 9, 25, 26; 10, 6;
11, 1, 2; 12, 6-10; 13, 10-14).
Мат ер иа л. Китай, провинция Юньнань, хр. «Ailaoshan» к северу от г. «Shuitangzhen»,
24º09ʹ51ʹʹ с. ш., 101º25ʹ29ʹʹ в. д., 2005 м, 7.VI.2011 (И. Белоусов, И. Кабак, А. Королев), 2 ♂ -
голотип и паратип, 1 ♀ - паратип [ЗИН].
Са м ец (голотип). Тело сравнительно маленькое для подрода. Окраска умеренно пестрая
(рис. 10, 6; 11, 1): эпикраниум серо-коричневый с желтоватыми глазами, глазками и пятном меж-
ду усиковыми впадинами, с темно-коричневой верхней частью (включая остальные части боко-
вых поверхностей рострального бугорка) и с многочисленными светло-коричневыми точками
(частично сливающимися одна с другой) на щеках и ниже глаз и усиковых впадин; усики свет-
ло-коричневые с несколькими коричневыми отметинами на скапусах и педицелах, а также с до-
вольно частыми умеренно затемненными и мелкими пятнышками на жгутиках; ротовые части
также серо-коричневые, но с немного более светлой верхней частью клипеуса и немного более
темной остальной его частью, со светло-коричневыми максиллами и лабиумом (включая их щу-
пики), а также с рыжевато-светло-коричневым лабрумом и темными вершинами мандибул; пе-
реднеспинка светло-коричневая с 1 парой крупных темно-коричневых пятен на боковых
лопастях, заходящих на диск и почти соприкасающихся одно с другим вдоль срединной линии
диска, а также с темной полоской вдоль заднего края диска, частично прерванной 6 мелкими
желтоватыми пятнышками; остальные тергиты серо-коричневые с многочисленными светло-
серо-коричневыми точками и с темной полоской вдоль каждого заднего края (эта полоска также
прервана несколькими желтоватыми пятнышками, но 2 последних тергита почти однотонно
серо-коричневые с 1 парой темных лопастинок 9-го тергита брюшка и 1 парой темных крючков
10-го тергита брюшка); плейриты средне- и заднегруди темно-коричневые; ноги светло-коричне-
вые с коричневыми пятнышками на коксах, бедрах и голенях, сливающимися в более крупные
пятна в верхней части передней коксы, дистальной половине среднего бедра и на вершине задне-
940
Рис. 11. Anabropsus (Apteranabropsis) spp., тело самца (1, 5) и самки (3) сбоку; яйцеклад сбоку
(2, 4, 6, 7).
1, 2 - A. ailaoshanica sp. n.; 3, 4 - A. maculata sp. n.; 5-7 - A. minuta (Gor.).
го бедра (вершина шипа передней коксы светлая, а дистальная половины заднего бедра перед
вершинным затемнением почти однотонно рыжеватая); стерниты груди светло-коричневые с
легким затемнением в основании стернита заднегруди, а стерниты брюшка и генитальная пла-
стинка слегка затемнены, но с желтоватыми как точками на стернитах, так и боковыми частями
задней половины генитальной пластинки (включая грифельки; рис. 9, 26); эпипрокт серо-корич-
невый; парапрокты коричневые с желтоватыми выростами; церки светло-коричневые. Строение
941
тела типичное для подрода, но со следуюшими особенностями: бугорок рострума с узкой верти-
кальной вмятиной спереди; темя со слабозаметным срединным швом (рис. 10, 6); боковые глазки
довольно крупные (рис. 11, 1); расстояние между усиковыми впадинами приблизительно в
2.2 раза меньше ширины скапуса; переднеспинка почти одинаковая по длине и ширине, ее пе-
редний и задний края почти прямые, а боковые лопасти без каких-либо плечевых выемок
(рис. 11, 1); средне- и заднеспинка с крохотными рудиментами крыльев в нимфальном положе-
нии (эти рудименты в покое полностью прикрыты задними частями впередилежащих тергитов и
очень похожи на таковые A. cervicornis; рис. 12, 1); стернит заднегруди - как на рис. 9, 25; перед-
няя голень с хорошо развитым внутренним тимпанумом и с маленьким (почти втрое более мел-
ким) наружным; заднее бедро с 9 или 10 мелкими внутренними нижними шипиками; задняя
голень с 9 парами немного более крупных нижних шипов; предпоследний тергит с 1 парой хоро-
шо заметных верхнебоковых лопастинок сзади; крючки последнего тергита хорошо развиты, но
при рассматривании сверху с округло-обрубленными вершинами (рис. 12, 7, 8); эпипрокт почти
удлиненно-овальный (рис. 12, 6); парапроктальные выросты более или менее прямые и направ-
лены вверх и назад от своих оснований (их форма - как на рис. 13, 10); генитальная пластинка
удлиненная, несколько сужена в дистальной половине, с обрубленной вершиной и с грифелька-
ми, длина которых незначительно больше расстояния между их основаниями (рис. 9, 26); гени-
талии - как на рис. 13, 12, 13.
Ва р иа ции. Второй самец с более светлой нижней частью клипеуса, несколькими
темноватыми отметинами на лабруме, очень незначительными отличиями в числе ши-
пиков и шипов задних ног, незначительно иной формой парапроктальных выростов
(рис. 13, 11), широкоокруглыми при рассматривании сверху вершинами крючков по-
следнего тергита и немного более короткой генитальной пластинкой.
С ам ка. Внешне очень похожа на самца (паратип), но крупные темные пятна на переднеспин-
ке немного мельче, 2 последних тергита почти темно-коричневые, церки с затемненными верши-
нами, наружный тимпанум передней голени значительно меньше на одной ноге и отсутствует на
другой ноге, последние тергиты без лопастинок и крючков, эпипрокт лишь более маленький
(рис. 12, 9), a парапрокты с очень короткими и угловидными задними выступами, чуть загнуты-
ми кверху (рис. 12, 10); генитальная пластинка и яйцеклад - как на рис. 11, 2 и 13, 14 (эта
пластинка светло-коричневая, а яйцеклад - интенсивно коричневый).
Длина (в мм). Тело: ♂ - 21.5-23, ♀ - 23.5; переднеспинка: ♂ - 5.6-5.8, ♀ - 6; переднее
бедро: ♂ - 8.8-9.3, ♀ - 9.1; заднее бедро: ♂ - 18-19.5, ♀ - 19.8; задняя голень: ♂ - 18-20, ♀ - 20;
задняя лапка: ♂ - 8-8.5, ♀ - 7.9; яйцеклад - 11.
Сравнение. Новый вид похож на A. (Apteranabropsis) sinica по отсутствию за-
метных килевидных ребрышек на тергитах, по очень сильной редукции крыльев
и по форме генитальной пластинки самца, но окраска его тела ясно более пятнистая
(особенно на переднеспинке и задних бедрах), лопастинки 9-го тергита брюшка самца
хорошо развиты (у A. sinica они почти незаметны - угадываются по характерным мор-
щинистостям, развитым на этом тергите около крючков 10-го тергита брюшка), пара-
проктальные выросты самца несколько иной формы (ср. рис. 13, 10, 11 и 13, 31, 32),
а грифельки генитальной пластинки самца более длинные (у нового вида они длиннее,
чем расстояние между их основаниями, а у A. sinica - короче). От A. (A.) paracostulata,
A. (A.) costulata, A. (A.) cervicornis и A. (A.) tonkinensis новый вид отличается отсут-
ствием заметных килевидных ребрышек на тергитах; кроме того, от первого вида он
отличен значительно более длинными грифельками генитальной пластинки самца, от
остальных - меньшими размерами тела, и дополнительно от двух последних видов -
не двулопастными вершинами парапроктальных выростов самца или рудиментар-
ными крыльями (у A. tonkinensis надкрылья очень сильно уменьшены, но все же
942
Рис. 12. Anabropsis spp. Нижнебоковые части тергитов птероторакса с мелкими (1) или очень
мелкими (13) рудиментами крыльев, либо только со следами этих рудиментов (29); вершина
брюшка самца сверху и немного сзади (2, 3, 6, 14, 19, 28); крючок последнего тергита самца
сбоку (4, 7, 15, 20, 22, 24, 26, 30, 32) и сверху (5, 8, 16, 21, 23, 25, 27, 31, 33); эпипрокт самки
сверху (9, 11, 17, 34); парапрокт самки сбоку (10, 12, 18, 35).
1 - A. (Apteranabropsis) cervicornis Karny; 2 - A. (Pteranabropsis) intermedia sp. n.; 3-5 - A. (P.) falcata
pallescens subsp. n.; 6-10 - A. (Apteranabropsis) ailaoshanica sp. n.; 11, 12 - A. (A.) maculata sp. n.;
13-18 - A. (A.) tarasovi sp. n.; 19-21 - A. (A.) abramovi sp. n.; 22-29 - A. (A.) minuta (Gor.);
30-35 - A. (A.) sinica B.-Bien. 1 - по: Горохов, 1998, с изменениями.
значительно крупнее и не рудиментарны, т. е. в обычном для крылатых имаго не-
перевернутом положении). От A. frater и A. femorata с неясной подродовой принад-
лежностью и от A. (A.) guangxiensis новый вид также отличается меньшими размерами
тела, причем первый из них значительно крупнее нового вида (длина переднеспинки и
заднего бедра у самца A. frater - соответственно 9 и 31 мм), а остальные - с иными
943
Рис. 13. Anabropsis spp., парапроктальный вырост самца (1, 3, 4, 8-11, 15, 21, 23, 25, 27, 31, 32)
и его вершинная часть (2, 16, 17, 22, 24, 26, 28) сбоку; гениталии самца сверху (5, 12, 18, 33)
и снизу (6, 13, 19, 34); генитальная пластинка самки снизу (7, 14, 20, 29, 30, 35-38).
1, 2 - A. (Pteranabropsis) carli Griff.; 3 - A. (P.) intermedia sp. n.; 4-7 - A. (P.) falcata pallescens subsp. n.;
8 - A. (Carnabropsis) carnarius (Gor.); 9 - A. (C.) pusilla (Ingr.); 10-14 - A. (Apteranabropsis)
ailaoshanica sp. n.; 15-20 - A. (A.) tarasovi sp. n.; 21, 22 - A. (A.) abramovi sp. n.; 23-30 - A. (A.) minuta
(Gor.); 31-35 - A. (A.) sinica B.-Bien.; 36 - A. (Pteranabropsis) falcata falcata (Shi et Bian);
37 - A. (Apteranabropsis) maculata sp. n.; 38 - A. (A.) abramovi namlikensis subsp. n.
31 - по: Gorochov, 2010.
944
пропорциями частей тела (отношения длины заднего бедра к длине переднеспинки
и яйцеклада у нового вида - 3.2-3.4 и 1.8, а у A. femorata - соответственно 2.6 и 1.4)
или с другой формой вершин парапроктальных выростов самца (в отличие от нового
вида у A. guangxiensis эти выросты слегка двулопастные). Anabropsis (A.) minuta и
A. (A.) miser, наоборот, мельче нового вида и лишены даже рудиментов крыльев
(рис. 12, 29).
Эт им оло ги я. Название нового вида образовано от названия хребта «Ailaoshan».
Anabropsis (Apteranabropsis) maculata Goroсhov, sp. n. (рис. 10, 4; 11, 3, 4; 12, 11,
12; 13, 37).
М ате риа л. Вьетнам: провинция «Lao Cai», 6 км к З от пос. «Sa Pa», северный склон горы
«Phan Si Pan», национальный парк «Hoang Lien», станция «Tram Ton», 22º21ʹ с. ш., 103º46ʹ в. д.,
1930-2000 м, V.2008 (А. Абрамов), 1 ♀ - голотип [ЗИН].
С а мка (голотип). Внешним обликом напоминает A. (A.) ailaoshanica sp. n., но тело немного
крупнее, а окраска светлее и более пятнистая (рис. 10, 4; 11, 3): эпикраниум желтоватый с мно-
гочисленными светло-коричневыми точками и пятнышками, а также со светло-коричневой верх-
ней частью, снабженной немного менее многочисленными желтоватыми точками, и с почти
беловатыми глазами и глазками; усики также желтоватые, но со светло-коричневыми прокси-
мальными члениками жгутиков, несущими редкие и мелкие желтоватые колечки, и с нескольки-
ми светло-коричневыми пятнышками на скапусах и педицелах; ротовые части в основном
однотонно желтоватые, но со светло-коричневыми точками на клипеусе, светло-коричневой
дистальной частью лабрума и затемненными вершинами мандибул; переднеспинка интенсивно
коричневая с многочисленными желтоватыми и светло-коричневыми точками и мелкими пят-
нышками; остальные тергиты похожи по окраске на переднеспинку, но тергиты птероторакса в
передней части почти однотонно желтоватые, а в нижних (боковых) частях эти тергиты и терги-
ты брюшка серовато-светло-коричневые (последние 2 тергита почти полностью серовато-свет-
ло-коричневые); ноги желтоватые со светло-коричневыми многочисленными пятнышками на
передних и средних бедрах и голенях (передние и средние коксы со светло-коричневыми верх-
ними пятнами и полностью желтоватыми шипами), а также с почти коричневой наружной по-
верхностью заднего бедра, снабженной многочисленными беловатыми пятнышками, и со слабо
выраженной пятнистостью задней голени; плейриты средне- и заднегруди светло-коричневые;
все стерниты, эпипрокт, парапрокты и генитальная пластинка светло-серо-коричневые; церки
желтоватые; яйцеклад коричневый со светло-коричневой базальной частью. Строение тела так-
же похоже на таковое самки A. (A.) ailaoshanica sp. n., включая состояние рудиментов крыльев,
но наружный тимпанум примерно в 1.5 раза меньше внутреннего, задние бедра с 3 или 4 нижни-
ми внутренними шипиками, задние голени с 10 нижними шипами на каждом крае, эпипрокт
почти в виде равностороннего треугольника (рис. 12, 11), парапрокты овально-удлиненные
и практически без выступов сзади (рис. 12, 12), генитальная пластинка с вершинным шипом
более коротким относительно остальной части (рис. 13, 37), а яйцеклад - как на рис. 11, 4.
С а мец неизвестен.
Длина (в мм). Тело - 21; переднеспинка - 7.1; переднее бедро - 9.3; заднее бедро - 22; задняя
голень - 21.5; задняя лапка - 8.4; яйцеклад - 11.
С р ав не ние. Новый вид наиболее похож на A. (A.) ailaoshanica sp. n., но отлича-
ется от него перечисленными в описании признаками. От A. (A.) sinica и A. (A.) minuta
новый вид отличается ясно более пятнистой окраской тела, а также дополнительно от
первого вида - более или менее одинаковыми шипами задней голени (у A. sinica на
этой голени более мелкие шипы перемежаются более крупными) и от второго - более
крупными размерами тела. От A. (A.) paracostulata, A. (A.) costulata, A. (A.) cervicornis
945
и A. (A.) tonkinensis новый вид отличен отсутствием заметных килевидных ребрышек
на тергитах и дополнительно от последнего вида - рудиментарными (не просто сильно
уменьшенными) крыльями; от A. (A.) guangxiensis новый вид отличается наличием за-
метных рудиментов крыльев (как у A. cervicornis - рис. 12, 1) и несколько меньшими
размерами (длина заднего бедра и яйцеклада у самки A. guangxiensis - соответственно
27.8-29 мм и 14.5-16 мм), от A. frater - также меньшими размерами (длина заднего
бедра у самца A. frater - 31 мм), от A. (A.) miser - заметно более крупной величиной
тела (длина заднего бедра у самца A. miser - 14 мм), а от A. femorata - иными про-
порциями частей тела (отношения длины заднего бедра к длине переднеспинки
и яйцеклада у нового вида - 3 и 2, a у A. femorata - соответственно 2.6 и 1.4).
Эт им оло г ия. Название нового вида «maculata» («пятнистая» на латыни) дано в
связи с характерной окраской тела.
Anabropsis (Apteranabropsis) tarasovi Gorochov, sp. n. (рис. 9, 27, 28; 12, 13-18;
13, 15-20; 14, 1, 2; 15, 1).
Мат ер иа л. Лаос, провинция «Champasak», плато «Bolaven», 14 км к юго-востоку от «Muang
Paxong», «Ban Houayteuay», 1200 м, 15º4.655ʹ с. ш., 106º16.848ʹ в. д., нарушенный горный лес,
почвенные ловушки, V-VI.2008 (С. Тарасов), 12 ♂ - голотип и паратипы, 11 ♀ - паратипы [ЗИН].
С ам ец (голотип). Тело по величине примерно как у A. (A.) maculata sp. n., но окраска менее
пятнистая (рис. 15, 1): эпикраниум интенсивно коричневый с темно-коричневыми боковыми
частями рострального бугорка (не считая желтоватых боковых глазков) и крупным полем под
глазами и усиковыми впадинами, в котором расположены довольно крупное желтоватое пятно
со срединным глазком и несколько желтоватых точек, а также с желтовато-сероватыми участка-
ми в нижней части щек, сероватой передневерхней поверхностью рострального бугорка и свет-
ло-коричневыми глазами (рис. 14, 1); усики беловатые с рыжевато-светло-коричневыми
проксимальными сегментами жгутиков и педицелами, а также с коричневыми скапусами, снаб-
женными желтоватыми пятном сверху и крупным полем снизу; ротовые части почти желтые с
коричневой верхней частью клипеуса, желтоватыми мандибулами, максиллами и лабиумом
(включая их щупики), а также с затемненными вершинами мандибул; переднеспинка однотонно
темно-коричневая (почти черная); остальные тергиты также темно-коричневые, но с редкими
желтоватыми точками на 1-3-м тергитах брюшка, мелкими пятнышками на 4-7-м тергитах
брюшка, более крупными пятнами на 8-м и 9-м брюшных тергитах и серовато-желтоватыми
участками на боковых частях тергитов брюшка (на передних тергитах эти участки небольшие,
а на более задних - постепенно увеличиваются к почти полностью серовато-желтоватому по-
следнему тергиту); плейриты груди, ноги и стерниты желтоватые, но передние и средние бедра
с рыжевато-коричневатыми дистальными половинами, заднее бедро почти полностью рыжева-
то-коричневатое со светло-коричневым основанием снаружи и желтоватой проксимальной тре-
тью изнутри и снизу, а все голени с рыжевато-коричневатыми проксимальными двумя третями;
генитальная пластинка едва затемненная в проксимальной половине (рис. 9, 28); крючки послед-
него тергита коричневые, а церки, эпипрокт и парапрокты желтоватые. Строение тела - как
у предыдущих видов подрода, описанных здесь, но со следующими особенностями: ростраль-
ный бугорок с более заметной срединной бороздкой; расстояние между усиковыми впадинами
примерно в 1.4 раза меньше ширины скапуса; задняя часть тергитов груди и 1-6-го тергитов
брюшка с несколькими продольными килевидными ребрышками, слабо развитыми на 5-м и 6-м
тергитах брюшка и хорошо заметными на предшествующих тергитах; рудименты крыльев едва
заметны, более маленькие, чем у A. (A.) ailaoshanica sp. n. и A. (A.) maculata sp. n. (рис. 12, 13);
стернит заднегруди - как на рис. 9, 27; передняя голень с 2 хорошо развитыми (но небольшими)
тимпанумами, причем наружный из них немного короче и почти в 1.5 раза уже внутреннего;
заднее бедро без шипиков, а задняя голень с 9 парами нижних шипов, из которых самая дисталь-
ная пара (перед парой предвершинных шпор) заметно мельче остальных нижних шипов; пред-
946
Рис. 14. Anabropsis (Apteranabropsis) spp., голова спереди (1, 3, 5), яйцеклад сбоку (2, 7, 8),
участок задней голени в области верхних проксимальных шипов сбоку и немного сверху (4, 6).
1, 2 - A. (A.) tarasovi sp. n.; 3, 4 - A. (A.) abramovi sp. n.; 5-7 - A. (A.) abramovi namlikensis subsp. n.;
8 - A. (A.) sinica B.-Bien.
последний тергит с 1 парой очень пологих и еле заметных лопастинок (рис. 12, 14); последний
тергит с крючками характерной формы (конусовидная вершинная часть крючка отходит вперед
от более широкого и высокого округлого бугорка, т. е. при рассматривании сбоку верхний край
крючка с вогнутостью; рис. 12, 15, 16); генитальная пластинка с очень широкой проксимальной
половиной и значительно суженной дистальной частью, обрубленной сзади и несущей довольно
короткие грифельки (длина каждого из грифельков чуть меньше расстояния между их основани-
947
ями; рис. 9, 28); эпипрокт, выросты парапроктов, церки и гениталии - как на рис. 12, 14 и 13, 15,
16, 18, 19.
Ва р иа ции. Некоторые самцы с наиболее светлыми участками тела от желтова-
то-сероватых до светло-коричневых, но лабрум всегда желтый; наружный тимпанум
иногда лишь незначительно уже внутреннего; обнаружены также незначительные ва-
риации в форме парапроктальных выростов (рис. 13, 17) и числе шипов задних го-
леней; ширина вершины генитальной пластинки также слегка варьирует.
С ам ка. Очень похожа на самцов, но затемненные части бедер нередко интенсивно коричне-
вые в вершинных участках и иногда с редкими светловатыми точками, рудименты крыльев
иногда полностью исчезают на одной стороне и/или на одном из тергитов, 2 последних тергита
брюшка без специализаций, а эпипрокт, парапрокты, генитальная пластинка и яйцеклад - как на
рис. 13, 20 и 14, 2 (последний рыжевато-коричневый с желтоватой проксимальной четвертью).
Длина (в мм). Тело: ♂ - 22-27, ♀ - 25-29; переднеспинка: ♂ - 6.5-7.5, ♀ - 7-8; переднее
бедро: ♂ - 8.8-10, ♀ - 8.9-10.5; заднее бедро: ♂ - 23-26, ♀ - 25-27; задняя голень: ♂ - 22-25, ♀ -
23-26; задняя лапка: ♂ - 8-8.5, ♀ - 8.5-9; яйцеклад - 14.5-16.5.
Сравнение. Новый вид похож на A. (Apteranabropsis) sinica и A. (A.) minuta по
более или менее однотонной и частично темной окраске, но отличается от первого
вида более расширенной проксимальной и более узкой дистальной частями гени-
тальной пластинки самца, а от второго - более крупными размерами тела и наличием
заметных продольных ребрышек на тергитах. От A. (A.) costulata, A. (A.) cervicornis,
A. (A.) tonkinensis и A. (A.) paracostulata, также снабженных продольными ребрыш-
ками на тергитах, новый вид отличается более мелкими (почти полностью утерян-
ными) рудиментами крыльев (от 3 первых видов) или более однотонной окраской (от
A. paracostulata); от A. (A.) ailaoshanica sp. n. и A. (A.) maculata sp. n. он отличается
теми же признаками и наличием продольных ребрышек на тергитах, от A. (A.) guang-
xiensis - совсем не двулопастными вершинами парапроктальных выростов самца, от
A. (A.) miser - значительно более крупными размерами тела (длина переднеспинки и
заднего бедра у самца A. miser - соответственно 4.5 и 14 мм), от A. frater - менее круп-
ными размерами (длина переднеспинки и заднего бедра у самца A. frater - соответ-
ственно 9 и 31 мм), а от A. femorata - иными промерами тела (отношения длины зад-
него бедра к длине переднеспинки и яйцеклада у нового вида - 3.4-3.6 и 1.6-1.7,
а у A. femorata - соответственно 2.6 и 1.4).
Э т имоло ги я. Вид назван в честь его сборщика.
Anabropsis (Apteranabropsis) abramovi Gorochov, sp. n. (рис. 9, 29; 12, 19-21; 13, 21,
22; 14, 3, 4; 15, 2).
Мате р иа л. Вьетнам, провинция «Dien Bien», уезд «Muong Nhe», коммуна «Sin
Thau», 22º21ʹ54ʹʹ с. ш., 102º14ʹ15ʹʹ в. д., 800 м, VI.2014 (А. Абрамов), 1 ♂ - голотип
[ЗИН].
С ам ец (голотип). Величина, окраска и строение тела очень похожи на таковые A. (A.) tarasovi
sp. n. (ср. рис. 15, 1 и 2), но со следующими отличиями: эпикраниум светло-коричневый с 1 ма-
леньким темно-коричневым пятнышком на верхней части рострального бугорка (над боковыми
глазками), коричневыми пятнышками позади глаз и по бокам рострального бугорка (впереди
боковых глазков), 4 темноватыми пятнышками между усиковыми впадинами (1 сразу под
ростральным бугорком и 1 срединным немного ниже) и у верхнебоковых углов клипеуса, а так-
же желтоватыми боковыми глазками и сероватыми глазами (рис. 14, 3); усики беловатые со свет-
948
Рис. 15. Anostostomatidae и Stenopelmatidae, тело самца (1, 2) и самки (3) сбоку, голова сверху
(4) и заднее бедро сбоку (5).
1 - Anabropsis (Apteranabropsis) tarasovi sp. n., 2 - A. (A.) abramovi sp. n., 3 - A. (A.) abramovi namlikensis
subsp. n., 4 - Diaphanogryllacris annamita (Griff.), 5 - Glaphyrosoma (Proctovitettix) dentatum Gor.
ло-коричневыми 2 проксимальными члениками жгутиков, имеющими также желтоватые
участки, и желтоватыми скапусами и педицелами, снабженными коричневыми (почти темно-
коричневыми) крупным участком на каждом педицеле и полосками и пятнами на скапусе; рото-
вые части с серовато-светло-коричневыми клипеусом, лабрумом и мандибулами (последние с
затемненными вершинами), а также с желтоватыми остальными структурами, имеющими едва
949
затемненные участки на максиллах и лабиуме (но не на их щупиках); тергиты груди и передних
сегментов брюшка - как у A. (A.) tarasovi sp. n. по окраске и строению, но задние тергиты брюш-
ка светлее (светло-коричневые с многочисленными коричневыми пятнышками), а 3 последних
тергита и эпипрокт почти однотонно серовато-желтоватые с затемненными лопастинками и
крючками на предпоследнем и последнем тергитах соответственно; ноги по окраске также похо-
жи на таковые этого вида, но немного светлее - светло-коричневые с затемненными мелкими
участками в области сочленения каждого бедра и голени и с беловато-желтоватыми лапками;
стерниты груди и первого сегмента брюшка сероватые, а остальные стерниты, генитальная пла-
стинка, парапрокты и церки желтоватые; продольная (вертикальная) вмятина на ростральном
бугорке спереди менее выражена, чем у A. (A.) tarasovi sp. n.; продольный киль на верхней части
эпикраниума сильнее выражен, чем у этого вида (ср. рис. 14, 1 и 14, 3); наружный тимпанум
передней голени лишь незначительно меньше внутреннего; остальные структуры тела (включая
гениталии) также очень близки по строению к таковым этого вида, но крючки последнего терги-
та обычной формы (рис. 12, 20, 21), парапроктальные выросты - как на рис. 12, 19 и 13, 21, 22,
а генитальная пластинка совсем иной формы (слабо сужена в дистальной части и с характерной
фигурной вырезкой между грифельками; рис. 9, 29).
С ам ка неизвестна.
Длина (в мм). Тело - 28; переднеспинка - 8; переднее бедро - 10.7; заднее бедро - 26; задняя
голень - 26; задняя лапка - 8.8.
Сравнение. От остальных видов подрода с известными самцами новый вид
легко отличается сравнительно короткой генитальной пластинкой самца, которая
слабо сужена в дистальной части и с характерной (широкой, неглубокой и слегка раз-
двоенной вследствие наличия срединного бугорка; рис. 9, 29) вырезкой между гри-
фельками. От A. (A.) costulata и A. femorata c неизвестными самцами новый вид от-
личен значительно более редуцированными рудиментами крыльев (у дейтонимфы
A. costulata эти рудименты примерно как на рис. 12, 1) или иными пропорциями
частей тела (отношение длины заднего бедра к длине переднеспинки у нового вида -
3.25, а у A. femorata - 2.6), а от недостаточно подробно описанного А. frater - менее
крупными размерами (длина переднеспинки и заднего бедра у самца нового вида -
8 мм и 26 мм, а у самца A. frater - 9 мм и 31 мм соответственно).
Эт им ол о ги я. Вид назван в честь его сборщика.
Anabropsis (Apteranabropsis) abramovi namlikensis Gorochov, subsp. n. (рис. 13, 38;
14, 5-7; 15, 3).
Мат ер иа л. Лаос, провинция «Vientiane», ~70 км северо-северо-западнее г. Вьентьян, «Nam
Lik Eco Village» на реке «Nam Lik», 18.61469º с. ш., 102.40847º в. д., ~200 м, среди подстилки
вторичного леса ночью, 10-30.VI.2017 (А. Горохов, М. Омелько), 1 ♀ - голотип [ЗИН].
С ам ка (голотип). Окраска и строение тела очень похожи на таковые номинативного подвида
(рис. 15, 3), но со следующими отличиями: размеры тела заметно меньше; окраска головы не-
много светлее - почти желтоватая, но с серо-коричневыми ростральным бугорком (кроме бело-
ватых боковых глазков по бокам и пятна на вершине) и вертикальными отметинами на
эпикраниуме под глазами (рис. 14, 5), а также с очень тонкими продольными светло-коричневы-
ми линиями на верхней части эпикраниума позади глаз; переднеспинка темно-коричневая с ко-
ричневыми и светло-коричневыми отметинами (сливающимися одна с другой) в средней части
груди; остальные тергиты груди и 1-4-й тергиты брюшка светло-коричневые в передней и тем-
но-коричневые в задней половинах; ноги однотонно светло-коричневые с темной мелкой отме-
тиной на вершинной части заднего бедра, неотчетливым беловато-желтоватым пятном на
верхней поверхности этой части и слегка затемненными небольшими проксимальными участка-
950
ми на голенях и на их шипах и шпорах; предпоследний и последний тергиты брюшка светло-
серовато-коричневые, но с желтоватыми боковыми частями; стерниты, генитальная пластинка,
эпипрокт, парапрокты и церки желтоватые, а яйцеклад коричневый со светло-коричневым ос-
нованием; срединный киль на эпикраниуме и продольные килевидные ребрышки на тергитах
менее выражены, чем у A. (A.) abramovi abramovi (ср. рис. 15, 2 и 15, 3); передние голени с более
короткими, почти округленно-квадратными (не овальными) тимпанумами; задние голени
с 11-15 наружными и 11 или 12 внутренними шипами и редкими шипиками между ними,
а боковые края голени между некоторыми из этих шипов очень мелко и округленно зазубрены
(у A. a. abramovi не зазубрены; см. рис. 14, 4 и 14, 6); строение вершины брюшка очень похоже
на таковое самки A. (A.) tarasovi sp. n., но дистальный шип генитальной пластинки и яйцеклад
заметно более короткие (см. рис. 13, 20, 38; 14, 2, 7).
С а мец неизвестен.
Длина (в мм). Тело - 26; переднеспинка - 6.7; переднее бедро - 8.8; заднее бедро - 21.5; задняя
голень - 20.5; задняя лапка - 7; яйцеклад - 11.
С р ав не ние. Отличия нового подвида от номинативного даны выше (в его опи-
сании). От A. (A.) costulata, A. (A.) cervicornis, A. (A.) tarasovi и A. (A.) paracostulata,
сходных с новым таксоном по наличию продольных килевидных ребрышек на тер-
гитах и по невидимым (прикрытым тергитами) в состоянии покоя рудиментам
крыльев, а также по отсутствию шипиков на бедрах, этот подвид отличается следую-
щими признаками: от первых двух видов - явно сильнее редуцированными рудимен-
тами крыльев (у A. costulata и A. cervicornis они как на рис. 12, 1, а у нового подвида -
как на рис. 12, 13); от третьего вида - относительно более короткими дистальным
шипом генитальной пластинки самки и яйцекладом (ср. рис. 13, 20, 38; 14, 2, 7);
от четвертого вида - более узким промежутком между усиковыми впадинами (этот
промежуток примерно в 1.4 раза уже скапуса, но у A. paracostulata он почти равен
скапусу по ширине), более короткими (округленно-квадратными, не овальными)
тимпанумами, более светлыми (не затемненными) верхней частью эпикраниума, пара-
проктами и церками, а также менее пятнистой окраской ног.
З а м еча ни я. Новый подвид отнесен к данному виду лишь условно (в основном
из-за более или менее сходной окраски головы), поскольку он известен только по
самке и может оказаться как отдельным видом, так и подвидом A. (A.) tarasovi.
Anabropsis (Apteranabropsis) minuta (Gorochov, 2001), comb. n. (рис. 9, 31-34;
10, 5; 11, 5-7; 12, 22-29; 13, 23-30).
М ате риа л. Вьетнам, провинция «Lao Cai»: 6 км к западу от с. «Sa Pa», северный склон
горы «Fan Si Pan», национальный парк «Hoang Lien», около станции «Tram Ton», 22º21ʹ с. ш.,
103º46ʹ в. д., 1930-2000 м, V.2008 (А. Абрамов), 1 ♂, 7 ♀ [ЗИН]; тот же национальный парк,
но 2000 м, 16-30.VII.2007 (С. Головач), 2 ♂, 6 ♀ [ЗИН].
С а ме ц (nov.). Размеры тела довольно маленькие для подрода. Окраска сравнительно одно-
тонная (рис. 10, 5; 11, 5): эпикраниум от коричневого до темно-коричневого с желтоватыми глаз-
ками и иногда светло-коричневым пятном позади нижней половины каждого глаза, а также со
светловатым пятнышком на вершине рострального бугорка; усики светло-коричневые или жел-
товатые с немного более темными пятнами на скапусе, почти полностью затемненным педице-
лом и иногда желтовато-беловатыми мелкими отметинами на проксимальной части жгутика;
ротовые части светло-коричневые с коричневой верхней или желтоватой нижней частями клипе-
уса, с желтоватыми щупиками, часто слегка затемненными на вершине, и иногда с коричневыми
пятнами на наружной поверхности проксимальной части мандибул; переднеспинка однотонно
коричневая или темно-коричневая, а остальные тергиты от коричневых до светло-коричневых и
951
с многочисленными узкими продольными беловатыми штрихами на 3-8-м тергитах брюшка или
почти без них; ноги светло-коричневые с рыжеватым оттенком или почти желтоватые, но обыч-
но с коричневыми предвершинной и вершинной частями бедер и с маленьким или довольно
крупным верхним беловатым пятном на вершинной части заднего бедра (иногда этого пятна
нет), а также часто со слегка затемненными проксимальными половинами голеней; стерниты от
серовато-желтоватых до светло-коричневых с чуть более темными генитальной пластинкой или
ее предвершинной частью; эпипрокт и парапрокты коричневые или светло-коричневые с желто-
ватыми вершиной эпипрокта и дистальной половиной парапроктального выроста; церки также
желтоватые. Голова без заметного срединного киля сверху, с расстоянием между усиковыми впа-
динами в 2-2.3 раза меньшим, чем ширина скапуса; тергиты без заметных продольных килевид-
ных ребрышек, а переднеспинка по форме - как на рис. 10, 5 и 11, 5; крылья отсутствуют, т. е.
почти без следов крыловых рудиментов на тергитах птероторакса (рис. 12, 29); выступы стерни-
та заднегруди - как на рис. 9, 31; наружный тимпанум немного или значительно меньше вну-
треннего; все бедра без нижних шипиков; передняя голень с 4 парами нижних шипов и 1 верхним
внутренним шипом; средняя голень с такими же нижними, 3 верхними внутренними и 3 верхни-
ми наружными шипами; задняя голень с 8 или 9 внутренними и 9 или 10 наружными верхними
шипами, а также с 2 или 3 нижними шипиками; предпоследний тергит лишь со следами задних
лопастинок (рис. 12, 28); эпипрокт, генитальная пластинка, крючки последнего тергита и пара-
проктальные выросты - как на рис. 9, 32-34 и 12, 22-28, а также 13, 23-28 (форма этих структур,
кроме эпипрокта, у разных самцов не идентична); гениталии почти неотличимы от таковых,
изображенных на рис. 13, 33, 34.
C ам ка. Тело похоже на таковое самцов, но стерниты, 2 последних тергита, эпипрокт, пара-
прокты и генитальная пластинка однотонно желтоватые (генитальная пластинка иногда слегка
затемненная), узкие продольные беловатые штрихи расположены часто и на 2 передних тергитах
брюшка, а вариации в величине наружного тимпанума менее выражены; эпипрокт и парапрокты
очень похожи на таковые, изображенные на рис. 12, 34, 35; яйцеклад и генитальная пластинка -
как на рис. 11, 6, 7 и 13, 29, 30 (эти структуры у разных самок несколько различаются: первый -
по длине, а вторая - по форме).
Длина (в мм, в скобках - для голотипа). Тело: ♂ - 14-18, ♀ - 16.5-21 (21); переднеспинка:
♂ - 5-5.9, ♀ - 5.8-6.2 (6.4); переднее бедро: ♂ - 6.2-7, ♀ - 6.3-6.9; заднее бедро: ♂ - 15.6-
17.7, ♀ - 16-17.3 (18.5); задняя голень: ♂ - 15.8-17, ♀ - 16-17 (18); задняя лапка: ♂ - 5.5-6.5,
♀ - 5-6; яйцеклад - 7.8-9.6 (10).
За меча ния. Этот вид был описан по единственной самке с той же горы (Gorochov,
2001b). Большинство ее признаков вполне соответствуют охарактеризованным выше
экземплярам, но по относительной длине яйцеклада и по форме генитальной пла-
стинки голотип занимает промежуточное положение между крайними вариантами
у этих экземпляров (приблизительные отношения длины заднего бедра и яйцеклада
у голотипа - 1.9, а у крайних вариантов - 1.8 и 2; ср. также рис. 11, 6, 7 и 13, 29, 30).
Различия между самцами в форме генитальной пластинки (рис. 9, 32-34), крючков
последнего тергита (рис. 12, 22-27) и парапроктальных выростов (рис. 13, 23-28)
также несколько превышают различия между некоторыми ранее описанными видами
или подвидами этого рода.
Anabropsis (Apteranabropsis) sinica Bey-Bienko, 1962 (рис. 9, 35, 36; 12, 30-35;
13, 31-35; 14, 8).
Мат ер иа л. Китай, провинция Юньнань, горы Давейшань, окр. Бинбяня, 350 м, 22.VI.1956
(Д. Панфилов), 1 ♂ - голотип [ИЗК]. Вьетнам, провинция «Cao Bang»: 40 км к западу от
г. «Cao Bang», национальный парк «Phia Oac - Phia Den», восточный склон горы «Phia Oac»,
22º36ʹ27ʹʹ с. ш., 105º52ʹ00ʹʹ в. д., 1600-1800 м, 22.V-6.VI.2018 (А. Анисюткин, А. Абрамов), 2 ♂,
952
3 ♀ [ЗИН]; уезд «Ha Lang», коммуна «Duc Quang», дер. «Ban Pooc», 800 м, 13-20.V.2019 (Н. Ор-
лов, Л. Иогансен, А. Острошабов), 1 ♀ [ЗИН].
Са мка (nov.). Внешний облик близок к таковому A. (A.) minuta, но размеры тела крупнее,
а окраска со следующими особенностями: эпикраниум темно-коричневый (почти черный) с се-
рыми или частично серыми глазами, беловатыми глазками и узким кольцом вокруг каждого из
них, светло-серо-коричневатыми нижними частями щек и иногда малозаметными пятнами
у клипеального шва; усики серо-коричневые со светло-серовато-коричневыми пятнами на ска-
пусе и малозаметными мелкими пятнышками на проксимальной части жгутика; ротовые части с
серо-коричневыми лабрумом и клипеусом (но верхняя часть последнего почти темно-коричне-
вая), со светло-серовато-коричневыми в целом остальными частями, снабженными мелкими и
редкими темноватыми пятнышками на максиллярных щупиках, и с темными вершинными ча-
стями мандибул; тергиты груди однотонно черные; ноги, плейриты груди и все стерниты свет-
ло-серо-коричневые с серо-коричневыми шипом на коксе передней ноги и дистальными
половинами бедер (иногда задние бедра затемнены в дистальных двух третях или лишь в дис-
тальной трети), а также пятнами на всех голенях (только дистальные части голеней почти пол-
ностью светлые); 1-3-й или 1-4-й тергиты брюшка темно-коричневые со светлыми боковыми
участками, а остальные тергиты мелкопятнистые (кроме однотонно светлого последнего терги-
та); стерниты светло-серо-коричневые; церки и генитальная пластинка серо-коричневые; эпи-
прокт, парапрокты и яйцеклад почти темно-коричневые, но последний с серо-коричневым
основанием. Строение тела также похоже на таковое A. (A.) minuta: срединный киль на верхней
части головы и продольные килевидные ребрышки на тергитах незаметны; скапус почти вдвое
шире расстояния между усиковыми впадинами; рудименты крыльев практически отсутствуют
(примерно как на рис. 12, 29); тимпанумы овальные, наружный немного меньше внутреннего;
задняя голень с мелкими шипиками (между верхними шипами) очень редкими, и какая-либо
зазубренность по краям голени между этими шипами отсутствует; эпипрокт, парапрокты и гени-
тальная пластинка - как на рис. 12, 34, 35 и 13, 35, а яйцеклад - как на рис. 14, 8.
С а ме ц. Голотип достаточно подробно охарактеризован в первоописании (Бей-Биенко, 1962),
но следует добавить, что парапроктальные выросты самца очень своеобразные - дугообразно
изогнутые кверху (рис. 13, 31). Перечисленные выше самцы из Вьетнама очень похожи на выше-
описанных самок по окраске и признакам внешнего строения, а также почти ничем не отличают-
ся от голотипа этого вида (ср. рис. 13, 31 и 13, 32), за исключением того, что мелкие шипики
между верхними шипами задней голени более редкие (как у самок). Однако некоторые признаки
требуют дополнительного уточнения: генитальная пластинка незначительно варьирует по степе-
ни вогнутости ее заднего края между грифельками (рис. 9, 35, 36); крючки последнего тергита
отличны от таковых A. (A.) minuta лишь меньшей вариабельностью и большей направленностью
вверх (у A. minuta они направлены вверх и вперед; см. рис. 12, 22-27 и 12, 30-33); гениталии,
совсем не изученные у голотипа, с более глубокой выемкой между боковыми лопастями
дорсальной складки, чем у A. (P.) falcata pallescens, c менее глубоко раздвоенной срединной
лопастью этой складки, чем у A. (A.) ailaoshanica и A. (A.) tarasovi, и с почти не разделенными
вентральными лопастями (в отличие от всех этих видов; см. рис. 13, 5, 6, 12, 13, 18, 19
и 13, 33, 34).
Длина (в мм, в скобках - для голотипа). Тело: ♂ - 18.5-21 (22), ♀ - 19-22; переднеспинка: ♂ -
6.8-7 (7.7), ♀ - 6.2-7; переднее бедро: ♂ - 6.7-7, ♀ - 6.8-7.4; заднее бедро: ♂ - 18-18.7
(23), ♀ -18.5-20; задняя голень: ♂ - 17-17.5, ♀ - 16.7-19; задняя лапка: ♂ - 5.9-6.6, ♀ - 6-6.8;
яйцеклад - 10.5-12.
З а м еча ни я. Дейтонимфа самки, описанная Г. Я. Бей-Биенко как паратип этого
вида, хорошо отличается от рассмотренного голотипа более крупными размерами и
менее однотонной окраской, а также заметно более крупными рудиментами крыльев;
она относится к другому виду и была обозначена как голотип A. (A.) costulata, поэтому
самка A. (A.) sinica впервые описана лишь здесь. Голотип из Китая немного крупнее
953
вьетнамских экземпляров, что может быть объяснено вариабельностью или принад-
лежностью к разным подвидам.
Род GLAPHYROSOMA Brunner-Wattenwyl, 1888
Этот центральноамериканский род, принадлежащий к трибе Glaphyrosomatini, не-
давно стал объектом особого внимания со стороны систематиков - в нем было опи-
сано множество новых видов, а сам он был разбит на 4 группы видов, отличающиеся
одна от другой главным образом наличием или отсутствием тимпанумов, гладким или
зазубренным яйцекладом и формой парапроктальных выростов самца (Gorochov,
Cadena-Castañeda, 2016; Cadena-Castañeda, Weissman, 2020). Среди этих признаков
лишь строение яйцеклада может отражать какие-либо существенные эволюционные
различия, способные свидетельствовать о родовом статусе такой группы, а все
остальные связаны либо с малозначимыми для систематики редукциями, могущими
проходить очень быстро и без существенных изменений генотипа, либо с незначи-
тельными изменениями формы копулятивных структур, которые также могут легко
меняться от вида к виду, поскольку они менее подвержены стабилизирующему
давлению внешней среды, чем многие другие морфологические признаки. Таким об-
разом, я предлагаю делить этот теперь уже довольно богатый видами род как минимум
на 2 подрода, которые описываются в подродовом ключе для рода Glaphyrosoma ниже,
но, возможно, заслуживают родового ранга.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ РОДА GLAPHYROSOMA
1. Окраска тела более или менее однотонно светлая; парапроктальные выросты самца
небольшие и почти пальцевидные, т. е. дистально не разделены на более или
менее шиповидную латеральную часть и мягкую медиальную лопасть; яйцеклад
отчетливо зазубрен в дистальной трети верхнего края (см. Gorochov, Cadena-
Castañeda, 2016: figs 213, 214, 218, 219, 242, 243, 267)
................................................................ подрод Proctovitettix Gorochov, subgen. n.
[Состав: типовой вид - Glaphyrosoma dentatum Gorochov, 2016; G. dilutum Gorochov,
2016. Этимология: название нового подрода - существительное мужского рода, ана-
грамма из частей названия рода Cnemotettix и морфологических терминов
«paraproctum» и «ovipositor» с латинизированными греческими и латинскими корнями
соответственно.]
- Окраска тела контрастная: поперечнополосатая или светлая с темной верхней
частью; парапроктальные выросты самца от небольших до крупных, но
дистально или на вершине разделены на более или менее шиповидную лате-
ральную часть и мягкую медиальную лопасть (иногда шиповидная часть или
медиальная лопасть сильно редуцированы, но от них остаются более или менее
заметные следы); яйцеклад не зазубрен, но нередко со слабой выемкой в верх-
ней части около вершины, которая иногда окружена несколькими крохотными
зубчиками (см. Gorochov, Cadena-Castañeda, 2016: figs 185, 186, 191, 192, 197,
198, 202, 203, 207, 224, 225, 230, 235-241, 248, 257, 259, 268-275, 292, 296, 298)
....................................................................................... подрод Glaphyrosoma s. str.
[Состав: типовой вид - Daihinia mexicana Saussure, 1859; остальные виды рода,
включенные в «Mexicanum Group», «Gracile Group» и, вероятно, «Magnoproctalis
954
Group», а также, возможно, G. karnyi Cadena-Castañeda et Gorochov, 2016 из «Karnyi
Group» (Cadena-Castañeda, Weissman, 2020).]
Подсем. LEZININAE - пустынные лжесверчки
Это подсемейство до настоящего момента считалось содержащим только один бес-
крылый род Lezina Walker, 1869, представители которого связаны главным образом с
норами в аридных зонах Юго-Западной Азии и Северной Африки. По своим морфоло-
гическим признакам, включая строение копулятивных органов самца (см. рис. 1, 6, 7;
16, 1), род Lezina ничем особо не выделяется среди прочих триб Anostostomatinae, и
подсем. Lezininae вполне могло бы рассматриваться как еще одна триба Anosto-
stomatinae. Но остатки яйцеклада у самки этого рода совсем не похожи на рудименты
яйцеклада тех групп Anostostomatinae, у которых он редуцирован: у Lezina от яйце-
клада сохраняется лишь непарная слабо склеротизованная выпуклость, располо-
женная над вершиной генитальной пластинки (рис. 16, 3); тогда как у Hemiandrus
Ander, 1938 и Cooloola Rentz, 1980 (Anostostomatinae) от яйцеклада остаются 6 ко-
ротких или умеренно коротких пальцевидных выростов, соответствующих 6 яйце-
кладным створкам, причем у Hemiandrus верхние и нижние створки более или менее
равные по длине и заметно длиннее внутренних, а верхние створки, кроме того, несут
на вершине маленькие либо очень маленькие грифельки (рис. 16, 7). Это един-
ственный четкий признак, позволяющий отделить Lezina от Anostostomatinae; второй,
но намного менее четкий признак, - обычно более длинные и тонкие (иногда почти
нитевидные) церки (рис. 16, 3).
Однако при исследовании длиннокрылого рода Gryllacropsis Brunner-Wattenwyl,
1888 из Индии нами установлено, что этот род, как и Lezina, лишен заметного пери-
стого рельефа на задних бедрах, его церки похожи на таковые последнего рода, а стро-
ение вершины брюшка самки у него необычное и отчасти также напоминает таковое
самки Lezina: сверху эта вершина, как и у самки Lezina, похожа на таковую самца, но
без крючков на 10-м тергите (рис. 16, 2, 5); 7-й стернит брюшка очень крупный,
а генитальная пластинка маленькая (намного меньше этого стернита), треугольная
и с округло-угловидной вершинной лопастинкой (рис. 16, 3, 6). Кроме того, и это
наиболее интересно, редуцированный яйцеклад Gryllacropsis совсем не похож на ре-
дуцированный яйцеклад Anostostomatinae, но очень напоминает рудимент яйцеклада у
реликтового рода Tarragoilus Gorochov, 2001 (Hagloidea: Prophalangopsidae), так как
его верхние створки (dv) короткие, толстые, сросшиеся одна с другой в проксимальной
половине, с расставленными вершинами и со сравнительно крупными грифельками
(s), его внутренние створки неразвиты, а от нижних створок (vv) сохранились лишь
маленькие боковые лопастинки (у Hemiandrus, например, верхние створки более
длинные, но тонкие, отделенные одна от другой до самого основания, со сближен-
ными вершинами и с намного более мелкими грифельками, а внутренние и нижние
створки хорошо развиты; ср. рис. 16, 6 и 16, 7). Происхождение характерного непар-
ного остатка яйцеклада Lezina от сросшихся верхних створок рудиментарного яйце-
клада, близкого к таковому Gryllacropsis и Tarragoilus, кажется наиболее логичным
вариантом его происхождения, а, следовательно, принадлежность Gryllacropsis к Le-
zininae также очень вероятна. Здесь я условно включаю последний род в подсем.
Lezininae.
955
Рис. 16. Anostostomatidae, вершина брюшка самца (1, 4) и самки (2, 5, 7) сверху и немного
сзади (без церок или лишь с основаниями церок) и дистальная часть брюшка самки
снизу (3, 6) (схематично).
1-3 - Lezina mutica (Br.-W.); 4 - Spinolezina obscura (Burr); 5, 6 - Gryllacropsis magniceps (Walk.);
7 - Hemiandrus pallitarsis (Walk.).
7s - 7-й стернит брюшка; 10t - 10-й тергит брюшка; с - место удаленного церка; dv - верхняя створка
рудимента яйцеклада; e - эпипрокт; g - генитальная пластинка самца и самки; p - парапрокт; s - грифелек
на вершине верхней створки рудимента яйцеклада; vv - нижняя створка рудимента яйцеклада.
Следует также отметить, что один вид, обычно рассматриваемый в составе рода
Lezina (Popov, 1984; Cigliano et al., 2020), весьма отличен от прочих его видов, включая
типовой, и заслуживает обособления в составе отдельного рода, который описывается
ниже в родовом ключе к подсем. Lezininae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ ПОДСЕМ. LEZININAE
1. Крылья отсутствуют у обоих полов; рудимент яйцеклада в виде единой слабо скле-
ротизованной выпуклости, расположенной над вершиной генитальной пластин-
ки и лишенной грифельков и дополнительных лопастинок (рис. 16, 3)
2.
– Крылья полностью развиты; рудимент яйцеклада состоит из отчетливо раздвоенной
верхней умеренно склеротизованной лопасти, вершины которой выступают
позади генитальной пластинки и снабжены крупными грифельками, а также
из 1 пары маленьких нижнебоковых лопастинок у основания этой лопасти
(рис. 16, 6)
Gryllacropsis.
956
[Состав: типовой вид - Gryllacropsis perturbans Brunner-Wattenwyl, 1888; Gryllacris
magniceps Walker, 1870, синонимизированный с типовым видом (Kirby, 1906).]
2. Тело маленькое, со сравнительно короткими церками (длина переднеспинки и цер-
ка - 5.1 и 4.8 мм соответственно); эпипрокт самца широкий (шире его гениталь-
ной пластинки) и с довольно крупным непарным шипом, направленным назад и
заметно выступающим позади остальных частей эпипрокта (рис. 16, 4)
...................................................................................... Spinolezina Gorochov, gen. n.
[Состав: типовой и единственный вид - Magrettia obscura Burr, 1900 (= Spinolezina
obscura comb. n.). Этимология: навание рода образовано от родового названия Lezina и
латинского морфологического термина «spina» (шип) в связи с наличием шипа на эпи-
прокте самца.]
- Тело крупнее и с более длинными церками (длины переднеспинки и церка - 5.3-
8.5 и 6-15.3 мм соответственно); эпипрокт самца менее широкий (значительно
уже генитальной пластинки) и без какого-либо шипа или иных заметных копу-
лятивных специализаций (рис. 16, 1)
......................................................... Lezina (= Magrettia Brunner-Wattenwyl, 1888).
[Состав: типовой вид Lezina - Lezina concolor Walker, 1869; типовой вид Magrettia -
Magrettia abominata Brunner-Wattenwyl, 1888, синонимизированный с L. concolor
(Uvarov, 1924); 11 остальных видов, включенных в этот род в каталоге Международ-
ного общества ортоптерологов (Cigliano et al., 2020).]
БЛАГОДАРНОСТИ
Автор благодарен коллегам, передавшим в коллекцию ЗИН свои сборы из Вьетнама,
Китая и Лаоса, а также всем зарубежным ортоптерологам, приславшим необходимую
для работы литературу, и М. В. Березину (Московский зоопарк), предоставившему
мне для опубликования сделанные им в природе фотографии.
ФИНАНСИРОВАНИЕ
Работа выполнена в рамках государственного исследовательского проекта Россий-
ской Федерации № АААА-А19-119082990107-3.
СПИСОК ЛИТЕРАТУРЫ
Бей-Биенко Г. Я. 1962. Результаты Китайско-Советских зоолого-ботанических экспедиций в юго-западный
Китай 1955-1957 гг. Новые или малоизвестные кузнечиковые (Orthoptera) из Сычуани и Юньнани.
Труды Зоологического института АН СССР, т. 30: 110-138.
Болдырев В. Ф. 1912. Способ оплодотворения и сперматофора у Tachycines asynamorus Аdеl. (Orthoptera,
Stenopelmatidae). Русское энтомологическое обозрение 12: 552-570.
Болдырев В. Ф. 1914. Материалы к познанию строения сперматофор и особенностей спаривания у Locustodea
и Gryllodea. Труды Русского энтомологического общества 41 (6): 1-244.
Горохов А. В. 1988а. Система и филогения современных прямокрылых надсемейств Hagloidea и
Stenopelmatoidea (Orthoptera) с описанием новых таксонов. Сообщение 1. Зоологический журнал 67
(3): 353-366.
Горохов А. В. 1988б. Система и филогения современных прямокрылых надсемейств Hagloidea и
Stenopelmatoidea (Orthoptera) с описанием новых таксонов. Сообщение 2. Зоологический журнал 67
(4): 518-529.
Горохов А. В. 1995а. Система и эволюция прямокрылых подотряда Ensifera (Orthoptera). Части I и II. Труды
Зоологического института РАН, т. 260, с. 1-224 + 1-212.
957
Горохов А. В. 1995б. О системе и эволюции отряда прямокрылых (Orthoptera). Зоологический журнал 74
(10): 39-45.
Горохов А. В. 1998. Материалы по фауне и систематике лжекузнечиковых (Orthoptera, Stenopelmatoidea) Ин-
докитая и некоторых других территорий. I. Энтомологическое Обозрение 77 (1): 73-105.
Горохов А. В. 2000. Phasmomimidae: прямокрылые (Orthoptera) или привиденьевые (Phasmatoptera). Палеон-
тологический журнал 3: 67-72.
Клюге Н. Ю. 2016. Строение яйцекладов и кладоэндезис прыгающих прямокрылых (Saltatoria, или
Orchesopia). Энтомологическое обозрение 95 (4): 765-800.
Шаров А. Г. 1968. Филогения ортоптероидных насекомых. Труды Палеонтологического института АН
СССР, т. 118. 216 с.
Ander K. 1939. Vergleichend-anatomische und phylogenetische Studien über die Ensifera (Saltatoria). Opuscula
Entomologica, suppl. 2, 306 p.
Brunner-Wattenwyl C. 1888. Monographie der Stenopelmatiden und Gryllacriden. Verhandlungen der Kaiserlich-
Königlichen Zoologisch-Botanischen Gesellschaft in Wien 38: 247-394, pls. 5-9.
Cadena-Castañeda O. J. 2019. A proposal towards classification of the raspy crickets (Orthoptera: Stenopelmatoidea:
Gryllacrididae) with zoogeographical comments: an initial contribution to the higher classification of the
Cadena-Castañeda O. J., Weissman D. B. 2020. Review of Glaphyrosoma (Orthoptera: Stenopelmatoidea:
Anostostomatidae), including new species and biological information. Zootaxa 4779 (1): 1-37. https://doi.
org/10.11646/zootaxa.4779.1.1
Caudell A. N. 1909. Miscellaneous notes on Orthoptera. Proceedings of the Entomological Society of Washington
11: 111-114.
Chintauan-Marquier I. C., Legendre F., Hugel S., Robillard T., Grandcolas Ph., Nel A., Zuccon D., Desutter-
Grandcolas L. 2016. Laying the foundations of evolutionary and systematic studies in crickets (Insecta,
Cigliano M. M., Braun H., Eades D. C., Otte D. 2020. Orthoptera Species File Online (Version 5.0/5.0). Visited 28
Desutter-Grandcolas L. 2003. Phylogeny and the evolution of acoustic communication in extant Ensifera (Insecta,
Desutter-Grandcolas L. 2020a. Sia ferox Giebel, 1861. Museum Nacional d’Histoire Naturelle. Available from:
Desutter-Grandcolas L. 2020b. Anabropsis chopardi Karny, 1932. Museum Nacional d’Histoire Naturelle. Available
Flook P. K., Klee S., Rowell C. H. F. 1999. Combined molecular phylogenetic analysis of the Orthoptera (Arthropoda,
Insecta) and implications for their higher systematics. Systematic Biology 48 (2): 233-253. https://doi.
org/10.1080/106351599260274
Gorochov A. V. 1984. On the higher classification of recent Ensifera (Orthoptera). In: Z. Kaszab (ed.). Verhandlungen
des Zehnten Internationalen Symposiums über Entomofaunistik Mitteleuropas (SIEEC X), 15-20 August
1983, Budapest. Budapest: Múzsák Közművelődési Kiadó, p. 187-189.
Gorochov A. V. 2001a. The higher classification, phylogeny and evolution of the superfamily Stenopelmatoidea.
In: L. H. Field (ed.). The Biology of Wetas, King Crickets and Their Allies. Wallingford: CABI Publishing,
p. 3-33.
Gorochov A. V. 2001b. New taxa of Anostostomatidae and Prophalangopsidae (Orthoptera). Zoosystematica Rossica
9 (2) (for 2000): 299-315.
Gorochov A. V. 2010. New species of the families Anostostomatidae and Rhaphidophoridae (Orthoptera:
Stenopelmatoidea) from China. Far Eastern Entomologist
206:
FEE/00000295.pdf
Gorochov A. V. 2019. The cricket subfamily Phalangopsinae (Orthoptera: Gryllidae) in Peru. Zoosystematica
Gorochov A. V., Cadena-Castañeda O. J. 2016. New and little known Stenopelmatoidea (Orthoptera: Ensifera) from
America. Zoosystematica Rossica
25
(1):
zr_2016_25_1_Gorochov_2.pdf
Griffini A. 1913. Sopra alcuni Grillacridi e Stenopelmatidi della collezione Pantel. Atti della Società italiana di
scienze naturali e del Museo civico di storia naturale in Milano 52: 61-104.
Gwynne D. T. 1995. Phylogeny of the Ensifera (Orthoptera): a hypothesis supporting multiple origins of acoustical
signalling, complex spermatophores and maternal care in crickets, katydids, and weta. Journal of Orthoptera
Research 4: 203-218. doi:10.2307/3503478
Heads S. W., Leuzinger L. 2011. On the placement of the Cretaceous orthopteran Brauckmannia groeningae from
Brazil, with notes on the relationships of Schizodactylidae (Orthoptera, Ensifera). ZooKeys 77: 17-30.
958
Ingrisch S. 2018. New taxa and records of Gryllacrididae (Orthoptera, Stenopelmatoidea) from South East Asia and
Ingrisch S. 2019. Review of the genus Pteranabropsis (Anostostomatidae: Anabropsinae) with description of six
Irish J. 1986. The species of Comicus Brunner v. Wattenwyl (Orthoptera: Schizodactylidae) with a discussion of
their origin. Navorsinge Nationale Museum Bloemfontein 5 (10): 253-276.
Irish J. 1995. A new species of Comicus Brunner v. Wattenwyl, with additional biogeographical data on the genus
(Orthoptera: Schizodactylidae). Cimbebasia 14: 37-41.
Johns P. M. 1997. The Gondwanaland weta: family Anostostomatidae (formerly in Stenopelmatidae, Henicidae or
Mimnermidae): nomenclatural problems, World checklist, new genera and species. Journal of Orthoptera
Jost M. C., Shaw K. L. 2006. Phylogeny of Ensifera (Hexapoda: Orthoptera) using three ribosomal loci, with
implications for the evolution of acoustic communication. Molecular Phylogenetics and Evolution 38 (2):
Karny H. H. 1937. Orthoptera fam. Gryllacrididae, subfamiliae omnes. In: P. Wytsman (ed.). Genera Insectorum,
fasc. 206, p. 1-317.
Kirby W. F. 1906. A Synonymic Catalogue of Orthoptera. Vol. 3. Orthoptera Saltatoria. Part I. (Achetidae
et Phasgonuridae). London: British Museum, 562 p.
Pictet A., Saussure H. 1893. De quelques orthoptères nouveaux. Mitteilungen der Schweizerischen Entomologischen
Gesellschaft 8 (8) (for 1891): 293-318.
Popov G. B. 1984. Insects of Saudi Arabia Orthoptera: Fam. Stenopelmatidae and Gryllacrididae. In: W. Büttiker,
F. Krupp (eds). Fauna of Saudi Arabia, vol. 6, p. 175-202.
Sheffield N. C., Hiatt K. D., Valentine M. C., Song H., Whiting M. F. 2010. Mitochondrial genomics in Orthoptera
Shi F., Bian X. 2016. A new addition to the subfamily Anabropsinae (Orthoptera: Anostostomatidae) from China.
Song H., Amedegnato Ch., Cigliano M. M., Desutter-Grandcolas L., Heads S. W., Huang Yu., Otte D., Whiting
M. F. 2015. 300 million years of diversification: elucidating the patterns of orthopteran evolution based on
Song Q., Guo H., Bian X., Shi F. 2016. Review of the genus Pteranabropsis Gorochov, 1988 (Orthoptera:
Anostostomatidae: Anabropsinae) with description of two new species from China. Zootaxa 4121 (4): 473-
Stringer I. A. N. 2001. The reproductive biology and the eggs of New Zealand Anostostomatidae. In: L. H. Field
(ed.). The Biology of Wetas, King Crickets and Their Allies. Wallingford: CABI Publishing, p. 379-397.
Uvarov B. P. 1924. Some new and interesting Orthoptera in the collection of the Ministry of Agriculture, Cairo.
Bulletin of the Ministry of Agriculture, Egypt, Technical Scientific Services 41: 1-41.
Vandergast A. G., Weissman D. B., Wood D. A., Rentz D. C., Bazelet C. S., Ueshima N. 2017. Tackling an intractable
problem: Can greater taxon sampling help resolve relationships within the Stenopelmatoidea (Orthoptera:
Wang H.-Q., Liu X.-W. 2020. A new genus and a new species of Anabropsinae (Orthoptera: Anostostomatidae) from
Wang M., Liu X., Li K. 2015. Review of the genus Pteranabropsis Gorochov (Orthoptera: Stenopelmatoidea:
zootaxa.3905.4.6
Weissman D. B. 2001. Communication and reproductive behavior in North American Jerusalem crickets
(Stenopelmatus) (Orthoptera: Stenopelmatidae). In: L. H. Field (ed.). The Biology of Wetas, King Crickets
and Their Allies. CABI Publishing, p. 351-373.
Weissman D. B., Bazelet C. S. 2013. Notes on southern Africa Jerusalem crickets (Orthoptera: Stenopelmatidae:
Zeuner F. E. 1939. Fossil Orthoptera Ensifera. London: British Museum, 321 p.
Zhang H., Huang Yu., Lin L., Wang X., Zheng Zh. 2013. The phylogeny of the Orthoptera (Insecta) as deduced from
Zhou Zh., Shi F., Zhao L. 2014. The first mitochondrial genome for the superfamily Hagloidea and implications for
journal.pone.0086027
959
THE FAMILIES STENOPELMATIDAE AND ANOSTOSTOMATIDAE
(ORTHOPTERA). 1. HIGHER CLASSIFICATION,
NEW AND LITTLE KNOWN TAXA
A. V. Gorochov
Key words: Orthoptera, systematics, new taxa, nomenclatural changes, distribution.
SUMMARY
The family Stenopelmatidae is here divided into 3 subfamilies only: Stenopelmatinae with
4 or 5 tribes (Stenopelmatini; Siini stat. resurr.; Maxentiini trib. n.; Oryctopterini trib. resurr. and
possibly Oryctopini); Schizodactylinae (= Comicinae Ander, 1939, syn. n.) undivided into tribes, and
Gryllacridinae (= Hyperbaeninae Cadena-Castañeda, 2019, syn. n.) with problematic tribal composition.
The following new taxa are described: Sia bugajus sp. n.; Amaxentius subgen. n. and Eumaxentius
subgen. n. in the genus Maxentius Stål, 1876, stat. resurr.; Comicus orangensis sp. n.; C. myburghi
sp. n.; C. namibicus sp. n. The genera Oryctopterus Karny, 1937, gen. resurr. and Brauckmannia
Martins-Neto, 2007, gen. resurr. are restored from synonymy with Oryctopus Brunner-Wattenwyl,
1888 and Schizodactylus Brullé, 1835, respectively. The family Anostostomatidae includes only
2 subfamilies: Anostostomatinae with 9 tribes (Anostostomatini; Anabropsini; Lutosini; Cratomelini
stat. n.; Brachyporini; Deinacridini stat. n.; Cooloolini stat. n.; Leiomelini stat. n.; Glaphyrosomatini)
and possibly Lezininae undivided into tribes. The genus Anabropsis Rehn, 1901 is considered as
consisting of 5 subgenera: Anabropsis s. str.; Paterdecolyus Griffini, 1913, stat. n.; Pteranabropsis
Gorochov, 1988, stat. n. (= Brevipenna Shi et Bian, 2016, syn. n.); Apteranabropsis Gorochov,
1988, stat. n.; Carnabropsis subgen. n. New combinations for 20 species of this genus are formed,
and the following new species and subspecies are described: A. (Pteranabropsis) intermedia sp. n.;
A. (P.) falcata pallescens subsp. n.; A. (Apteranabropsis) ailaoshanica sp. n.; A. (A.) macu-
lata sp. n.; A. (A.) tarasovi sp. n.; A. (A.) abramovi sp. n.; A. (A.) abramovi namlikensis subsp. n.
Previously unknown male for Anabropsis (Аpteranabropsis) minuta (Gorochov, 2001) and female for
A. (A.) sinica Bey-Bienko, 1962 are described. Proctovitettix subgen. n. in the genus Glaphyrosoma
Brunner-Wattenwyl, 1888 as well as Spinolezina gen. n. for S. obscura (Burr, 1900), comb. n. from
Lezininae are also described; the genus Gryllacropsis Brunner-Wattenwyl, 1888 is tentatively included
in the latter subfamily. Keys to several taxa and new data on distribution of some species are given.
960