ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 99, 4, 2020
УДК 595.721
MONGOLABIS BOCHKOVI SP. N., НОВЫЙ ВИД УХОВЕРТОК
(DERMAPTERA, ANISOLABIDIDAE: ANISOLABIDINAE)
ИЗ СЕВЕРНОГО ВЬЕТНАМА
© 2020 г. Л. Н. Анисюткин
Зоологический институт РАН
Университетская наб., 1, С.-Петербург, 199034, Россия;
Совместный Российско-Вьетнамский Тропический научно-исследовательский
и технологический центр
Нгуен Ван Хуен, Кау Зяй, Ханой, Вьетнам
e-mail: leonid.dictyoptera@gmail.com, Leonid.Anisyutkin@zin.ru
Поступила в редакцию 3.03.2020 г.
После доработки 29.07.2020 г.
Принята к публикации 27.04.2020 г.
Описан новый вид рода Mongolabis, M. bochkovi sp. n. из северного Вьетнама. Приведены под-
робное морфологическое описание и краткие сведения об образе жизни нового вида. Уточнен
диагноз рода Mongolabis, в него из рода Anisolabis перенесены Mongolabis interjacens (Anisyutkin,
1988), comb. n., M. incrassatus (Anisyutkin, 1988), comb. n., M. incurvatus (Anisyutkin, 1988),
comb. n., M. fusoideus (Anisyutkin, 1988), comb. n. и M. dealbatus (Anisyutkin, 1988), comb. n.
Ключевые слова: уховертки, морфология, новый вид, Anisolabidinae.
DOI: 10.31857/S0367144520040085
Подсем. Anisolabidinae включает многочисленных, преимущественно бескрылых,
геофильных уховерток (Sakai, 1987; Srivastava, 1999). В настоящей работе приведено
описание нового вида рода Mongolabis Zacher, 1911, M. bochkovi sp. n., из северного
Вьетнама с замечаниями о его образе жизни, а также уточнены диагноз и состав рода
Mongolabis.
МАТЕРИАЛ И МЕТОДИКА
Весь материал, включая голотип, был собран в 70%-ный этанол и хранится в нем в настоящее
время. Экземпляры изучались в чашках Петри под бинокулярными микроскопами Leica MZ16
и МБС-10. Рисунки выполнены на рисовальном аппарате микроскопа Leica MZ16. Гениталии
самцов обрабатывались 10-15%-ным раствором КОН по стандартной методике (12-24 ч) для
удаления мягких тканей.
Все экземпляры, включая голотип, хранятся в коллекции Зоологического института РАН
в Санкт-Петербурге.
961
РЕЗУЛЬТАТЫ
Сем. ANISOLABIDIDAE Verhoeff, 1902
Подсем. ANISOLABIDINAE Verhoeff, 1902
Род MONGOLABIS Zacher, 1911
Ти по во й в ид: Gonolabis woodwardi Burr, 1906, по последующему обозначению
(Burr, 1911).
В настоящее время нет общепринятой родовой системы уховерток подсем.
Anisolabidinae. В этой работе я принимаю трактовку рода Mongolabis по Г. К. Срива-
ставе (Srivastava, 1999), который выделил в него уховерток с метапарамерами (параме-
рами у Сриваставы) немного (примерно в 1.5 раза) длиннее своей ширины, с угло-
видно выступающим наружным краем и оттянутой вершиной.
Следуя такому определению рода Mongolabis, я переношу в него ряд видов, перво-
начально описанных в роде Anisolabis Fieber, 1853 в широком смысле (Анисюткин,
1998): M. interjacens (Anisyutkin, 1998), comb. n., M. incrassatus (Anisyutkin, 1998),
comb. n., M. incurvatus (Anisyutkin, 1998), comb. n., M. fusoideus (Anisyutkin, 1998),
comb. n. и M. dealbatus (Anisyutkin, 1998), comb. n.
Автор не во всем согласен с Г. К. Сриваставой (Srivastava, 1999). Его родовая клас-
сификация Anisolabidinae основана исключительно на признаках строения метапара-
меров, что приводит к ошибкам при определении систематической принадлежности
некоторых видов. Например, если принять вышеприведенный диагноз, Euborellia lata
Anisyutkin, 1998 должен быть перенесен в род Mongolabis, так как сходен с его пред-
ставителями по строению метапарамеров (Анисюткин, 1998). Такому решению проти-
воречит очень своеобразное строение генитальных лопастей у Eu. lata, свойственное
всем видам рода Euborellia Burr, 1910 (склеротизованные поля генитальной лопасти
образуют пару овальных пузырьков; вероятно, это апоморфия рода - Анисюткин,
1998) и отличное от такового у Mongolabis. Очевидно, набор признаков, на основании
которых строится родовая система, должен быть расширен.
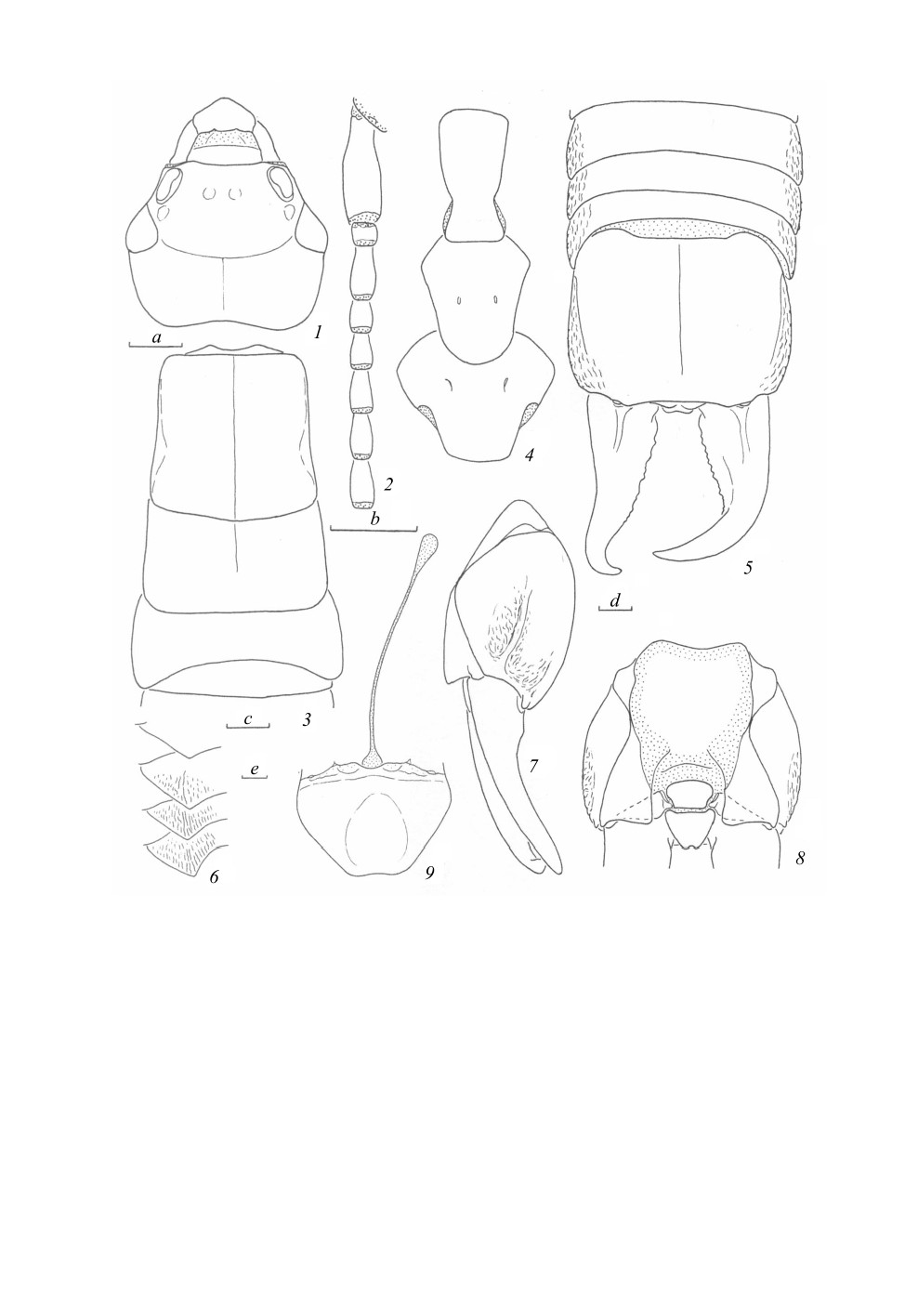
Mongolabis bochkovi Anisyutkin, sp. n. (рис. 1, 2).
Мат ер иа л. Вьетнам. Провинция Каобанг (Cao Bang), национальный парк Фиа-Оак -
Фиа-Ден (Phia Oac - Phia Den), восточный склон горы Фиа-Оак (Phia Oac), 40 км В г. Каобанг
(Cao Bang), 22º36′27′′ N, 105º52′00′′ E, h = 1600-1800 м, 22.V-VI.2018 (Л. Н. Анисюткин,
А. В. Абрамов), 5 ♂, голотип и паратипы; 1 ♀, 1 личинка, паратипы; те же данные, но
3-15.VI.2019 (Л. Н. Анисюткин), 2 ♀, паратипы.
С ам ец (голотип). Голова, грудь сверху и брюшко темно-коричневые, почти черные; брюшко
темнее по направлению к вершине, 10-й тергит черный; церки черные, с красноватым отливом;
генитальная пластинка красновато-коричневая. Глаза черные. Антеклипеус светло-желтый,
мембранозный. Ротовые органы, антенны, за исключением белесых 16-го и 17-го члеников,
стерниты груди, бедра и голени коричневые. Лапки более светлые, грязно-желтые.
Покровы гладкие и блестящие. Брюшко с мелкой густой пунктировкой, немного более
крупной и глубокой на 10-м тергите, где она образует слабо заметные продольные полосы, и на
генитальной пластинке; боковые края 7-10-го тергитов брюшка с продольной морщинистостью
(рис. 1, 5-8). Опушение по большей части слабо выраженное, короткое и редкое, присутствует
на ротовых органах, антеннах и ногах, плотное опушение развито только на голенях и лапках.
962
Рис. 1. Mongolabis bochkovi sp. n., самец, голотип.
1 - голова сверху; 2 - базальная часть левой антенны; 3 - грудь и 1-й сегмент брюшка сверху; 4 - грудные
стерниты снизу; 5 - вершина брюшка сверху; 6 - боковые углы 6-9-го тергитов брюшка слева;
7, 8 - 10-й тергит брюшка и церки (гипандрий удален) слева (7) и снизу, показаны только
основания церок (8); 9 - гипандрий снизу. Пунктировкой показаны мембранозные участки.
Масштабная линейка - 1 мм: a - к 1; b - к 2; c - к 3, 4; d - к 5, 7, 8; e - к 6, 9.
Голова длиннее ширины, выпуклая (рис. 1, 1); эпикраниальные швы в виде тонких линий;
глаза небольшие, по длине примерно равны заглазничному пространству; глазки отсутствуют;
задний край головы плавно вырезан. Левая и правая антенны 21- и 22-члениковые (рис. 1, 2),
скапус крупнее других члеников, немного короче расстояния между антеннальными впадинами;
педицеллус примерно равной длины и ширины, отношение длин 10 базальных члеников (с 1-го
по 10-й) примерно 5 : 1.0 : 2.2 : 1.4 : 1.6 : 2 : 2 : 2.2 : 2.4 : 2.4; далее членики очень постепенно
удлиняются, но вершинные снова немного укорачиваются.
963
Пронотум длиннее ширины, слабо расширяется к заднему краю (рис. 1, 3); его передний
и боковые края почти прямые, задний - слабо оттянут назад; про- и метазона не разделены; сре-
динная борозда отчетливая.
Надкрылья и крылья отсутствуют. Средне- и заднегрудь без боковых килей. Среднегрудь
с полустертой срединной бороздой и прямым задним краем (рис. 1, 3). Заднегрудь короче и
шире (рис. 1, 3) среднегруди, срединная борозда практически неразличима. Грудные стерниты
(рис. 1, 4): простернум удлиненный, передний край слабо закругленный, боковые сходятся при-
мерно к задней трети стернита, задний - прямой, фуркальные ямки не различимы; мезостернум
удлиненный, по заднему краю округленный, выступающий, фуркальные ямки слабо выражены;
метастернум шире своей длины, по заднему краю слабо выступающий, фуркальные ямки хо-
рошо заметны.
Ноги относительно короткие; передние бедра утолщены; лапки с наиболее длинным 1-м чле-
ником, 2-й членик короткий, немного длиннее своей ширины, 3-й - немного короче 1-го; 1-й
и 2-й членики снизу густо покрыты короткими щетинками; коготки простые, симметричные;
аролиум отсутствует.
Брюшко слабо уплощенное, постепенно расширяется к вершине; 2-й и 3-й тергиты со слабо
выраженными боковыми бугорками; боковые, загнутые вниз края 7-9-го тергитов оттянуты
назад, заострены и латерально морщинистые (рис. 1, 5, 6). 10-й тергит поперечный, слабо
вздутый (рис. 1, 5, 7, 8), его боковые края латерально морщинистые и несут боковые кили, кау-
дально с небольшими выростами над основаниями церок; задний край между выростами слабо
выемчатый; срединная борозда слабо выражена. Церки относительно короткие, немного
длиннее 10-го тергита, асимметричные, на виде сверху сильно (рис. 1, 5), в профиль - слабо
равномерно изогнутые (рис. 1, 7); левая церка почти прямая, изогнута у вершины, правая -
резко изогнута примерно у середины, обе с редкими тупыми зубцами по внутреннему краю.
Пигидий и парапрокты - см. рис. 1, 5, 8. Генитальная пластинка трапециевидная, со слабо выем-
чатым каудальным краем и нечетким округлым вдавлением; манубриум длинный и узкий
(рис. 1, 9).
Гениталии самца склеротизованные, симметричные (рис. 2, 1-5), проксимальный стебель
длинный, более чем в 10 раз длиннее мета- и пропарамеров вместе взятых. Метапарамеры отно-
сительно короткие и широкие, немного длиннее своей ширины, с угловидно выступающим на-
ружным краем и немного оттянутой вершиной, в профиль прямые (рис. 2, 1-4). Пропарамеры
сильнее склеротизованы латерально, разделены примерно на 1/3 своей длины, со склеритом,
расположенным вдоль линии сочленения (рис. 2, 1). Генитальные лопасти короткие, с парой
склеритов у основания, вместе с дистально выворачивающимися мембранозными мешками
примерно вдвое длиннее метапарамеров (рис. 2, 1, 3, 4); мембранозные мешки вентрально
с более склеротизованными участками, покрытыми папиллами (рис. 2, 4). Вирга не склеротизо-
вана, трудно различима; какие-либо дополнительные склеротизованные структуры, связанные
с виргой, в генитальных лопастях отсутствуют. В основании проксимального стебля располо-
жена пара удлиненных базальных пузырьков (рис. 2, 5).
Изм ен чиво ст ь. Антенны 20-24-члениковые, белесым может быть только один,
18-й или 19-й членик.
Са м ка. Сходна с самцом, отличается следующими признаками. Стерниты груди
более светлые, частично коричнево-желтые; у одного экземпляра бедра примерно
в дистальной четверти, голени и лапки грязно-желтые. Глаза по длине равны
примерно 0.6-0.8 заглазничного пространства. Антенны 22- или 23-члениковые, беле-
сыми могут быть 2-4 членика (в последнем случае 1-й и последний членики белесые
лишь частично) с 14-го по 18-й. 7-9-й тергиты брюшка с боковыми краями очень
слабо оттянутыми назад и более слабой, чем у самца, морщинистостью. 10-й тер-
964
Рис. 2. Mongolabis bochkovi sp. n., самец, голотип (1-5), и самка, паратип (6, 7).
1 - гениталии, общий вид сверху, генитальные лопасти вывернуты, показано только основание
проксимального стебля; 2 - метапарамеры и левая вывернутая генитальная лопасть снизу (правая
генитальная лопасть не показана); 3, 4 - правые метапарамер и генитальная лопасть сверху (3)
и справа (4); 5 - базальный пузырек; 6 - 10-й тергит брюшка и церки сверху; 7 - 10-й тергит
брюшка снизу. Пунктировкой показаны мембранозные участки.
Масштабная линейка - 1 мм: a - к 1, 3, 4; b - к 2; c - к 5; d - к 6, 7.
гит каудально слабо сужается (рис. 2, 6). Церки удлиненные, почти симметричные
(рис. 2, 6). Генитальная пластинка каудально выступающая (рис. 2, 7).
Л и чин ка самки последних возрастов. Сходна с имаго, но меньших размеров, антенны без
белесых члеников (осталось 17, остальные обломаны), дистальные участки бедер, голени
и лапки грязно-желтые.
965
Р азме ры (в мм; в скобках - для голотипа). Длина головы: самец - 3.6-4.0 (4.0), самка -
3.8-4.1; ширина головы: самец - 3.2-3.4 (3.4), самка - 3.0-3.5; длина пронотума: самец - 3.4-4.1
(4.1), самка - 3.6-4.0; ширина пронотума: самец - 3.5-3.9 (3.7), самка - 3.3-3.6; ширина мезоно-
тума: самец - 3.9-4.2 (4.2), самка - 3.7-4.2; ширина метанотума: самец - 4.3-4.7 (4.7), самка -
4.2-4.8; длина передних / средних / задних бедер: самец - 3.0-3.3 / 3.5-3.8 / 4.5-4.9 (3.2 / 3.8 /
4.9), самка - 3.0-3.3 / 3.4-3.8 / 4.2-5.0; ширина 10-го тергита брюшка: самец - 6.0-6.7 (6.5),
самка - 5.1-6.1; длина церок: самец - 5.2-5.8 (5.8), самка - 5.2-5.6. Ширина головы измерялась
в наиболее широком месте, сразу за глазами.
Ср а в нит ел ьны е з ам еча ни я. Новый вид крупными размерами и темной,
почти черной окраской сходен с Mongolabis magna (Bey-Bienko, 1959), описанным из
Юньнани (Бей-Биенко, 1959), но отличается относительно более короткими и резко
изогнутыми церками (сравнить рис. 1, 5 и рис. 22 в: Бей-Биенко, 1959) и значительно
менее выступающими наружными углами метапарамеров (сравнить рис. 2, 1-3
и рис. 26 в: Бей-Биенко, 1959).
Mongolabis bochkovi sp. n. по строению метапарамеров, широких, с сильно выступа-
ющим наружным углом и оттянутой вершиной, сходен с M. incurvatus (Anisyutkin,
1998) и M. fusoideus (Anisyutkin, 1998), описанными соответственно из Южного Китая
(Юньнань, окр. Фохая) и северного Вьетнама (провинция Виньфу, Тамдао)
(Анисюткин, 1998). Форма метапарамеров нового вида промежуточная между этими
видами: массивность метапарамера и его наружный угол более выражены у M. in-
curvatus, но менее у M. fusoideus. От обоих видов M. bochkovi sp. n. хорошо отличается
большими размерами, более темной, почти черной окраской и одноцветно темными
бедрами.
Эт им оло г ия. Вид назван в честь систематика и паразитолога А. В. Бочкова
(1968-2018), друга, однокурсника и коллеги.
За меча ния. Все экземпляры имаго M. bochkovi sp. n. были собраны в крупных
кусках разлагающейся древесины, лежащих на лесной подстилке под пологом первич-
ного леса и пронизанных многочисленными ходами личинок жуков-ксилофагов.
В лесной подстилке была собрана только одна личинка уховертки. Уховертки обитали
в ходах вместе с личинками жуков.
Будучи в основном всеядными насекомыми, уховертки, включая представителей
рода Anisolabis (см.: Marucci, 1955), могут хищничать, особенно активно поедая ли-
чинок насекомых с мягкими покровами (Marucci, 1955; Sueldo et al., 2010; Zhong et al.,
2016). Можно предположить, что и M. bochkovi sp. n. играет заметную роль в процессе
разложения мертвой древесины, регулируя численность насекомых-ксилофагов.
БЛАГОДАРНОСТИ
Автор искренне благодарен всем друзьям и коллегам, участникам экспедиции
Совместного Российско-Вьетнамского Тропического научно-исследовательского и
технологического центра, а также сотрудникам национального парка Фиа-Оак -
Фиа-Ден за помощь в полевых исследованиях.
ФИНАНСИРОВАНИЕ
Работа выполнена в рамках гостемы AAAA-A19-119020690101-6.
966
СПИСОК ЛИТЕРАТУРЫ
Анисюткин Л. Н. 1998. К познанию уховерток подсем. Anisolabidinae (Dermaptera, Anisolabididae) Юго-Вос-
Бей-Биенко Г. Я. 1959. Результаты китайско-советских зоолого-ботанических экспедиций 1955-1957 гг.
в Юго-Западный Китай. Энтомологическое обозрение 38 (3): 590-627.
Burr M. 1911. Dermaptera. In: P. Wytsman (ed.). Genera Insectorum, vol. 122, p. 1-112.
Marucci P. E. 1955. Notes on the predatory habits and life cycle of two Hawaiian earwigs. Proceedings of Hawaiian
Entomological Society 15 (3): 565-569.
Sakai S. 1987. Dermapterorum catalogus XIX-XX: Iconographia IV-V. Chelisochidae and Anisolabididae. Tokyo:
Daito Bunka University, 1567 p.
Srivastava G. K. 1999. On the higher classification of Anisolabididae (Insecta: Dermaptera) with a check-list of
genera and species. Records of the Zoological Survey of India 97 (Part 1): 73-100.
Sueldo M. R., Bruzzone O. A., Virla E. G. 2010. Characterization of the earwig, Doru lineare, as a predator of larvae
of the fall armyworm, Spodoptera frugiperda: A functional response study. Journal of Insect Science 10: 38.
Zhong B., Lv C., Qin W. 2016. Preliminary study on biology and feeding capacity of Chelisoches morio (Fabricius)
(Dermaptera: Chelisochidae) on Tirathaba rufivena (Walker). SpringerPlus 5 (1): 1944. doi: 10.1186/s40064-
016-3628-9
MONGOLABIS BOCHKOVI SP. N., A NEW EARWIG (DERMAPTERA,
ANISOLABIDIDAE: ANISOLABIDINAE) FROM NORTHERN VIETNAM
L. N. Anisyutkin
Key words: earwigs, morphology, new species, Anisolabidinae.
SUMMARY
A new species of the genus Mongolabis, M. bochkovi sp. n., is described from northern Vietnam.
Detailed morphological description and notes on the biology of the new species are given. The
diagnosis of the genus Mongolabis is emended. Mongolabis interjacens (Anisyutkin, 1988), comb. n.,
M. incrassatus (Anisyutkin, 1988), comb. n., M. incurvatus (Anisyutkin, 1988), comb. n., M. fusoideus
(Anisyutkin, 1988), comb. n. and M. dealbatus (Anisyutkin, 1988), comb. n., originally described in
the genus Anisolabis, are transferred to Mongolabis.
967