ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 100, 2, 2021
УДК 595.754 (571.651)
СОСТАВ ФАУНЫ И ОСОБЕННОСТИ БИОТОПИЧЕСКОГО
РАСПРЕДЕЛЕНИЯ ПОЛУЖЕСТКОКРЫЛЫХ (HETEROPTERA)
В ОКРЕСТНОСТЯХ ПЕВЕКА (ЧУКОТСКИЙ АО)
© 2021 г. О. А. Хрулева, 1* Н. Н. Винокуров 2**
1 Институт проблем экологии и эволюции имени А. Н. Северцова РАН
Ленинский пр., 33, Москва, 119071 Россия
*e-mail: oa-khruleva@mail.ru
2 Институт биологических проблем криолитозоны СО РАН
Якутск, 677980 Россия
**e-mail: vinok@ibpc.ysn.ru
Поступила в редакцию 31.01.2021 г.
После доработки 04.02.2021 г.
Принята к публикации 04.02.2021 г.
Полужесткокрылые - отряд насекомых, у которого видовое богатство фаун резко сокраща-
ется севернее границы леса. В статье рассмотрены результаты изучения комплексов полужест-
кокрылых на севере Чаунского района (Западная Чукотка, подзона типичных тундр). Сборы
проводились в 2011 г. в трех ландшафтных районах. Выявленную фауну отличает необычно
высокое для севера тундровой зоны видовое богатство: 32 вида из 8 семейств. Более полови-
ны ее состава - широкоареальные полизональные и бореальные виды клопов. Среди наиболее
обычных преобладали сухолюбивые виды, в том числе общие с криофитными степями Северо-
Востока Азии. За исключением криоксерофильного Nysius ericae groenlandicus, криофилы (в том
числе арктические Chiloxanthus arcticus, Ch. stellatus stellatus, Calacanthia trybomi, Orthotylus
artemisiae) лишь в отдельных биотопах имели высокое обилие. Группировки с наиболее устой-
чивым видовым составом приурочены к местообитаниям с высоким обилием полыней (зоо-
генные луговины, колеи, обочины дорог, где постоянно встречались Europiella artemisiae и
N. e. groenlandicus, локально - также O. artemisiae) и склонам сопок южной экспозиции (наибо-
лее обычны Chlamydatus pullus, N. e. groenlandicus, Coriomerus scabricornis, Antheminia eurynota
remota). В прочих типах местообитаний население клопов не имело постоянного состава. Эко-
логический облик комплексов полужесткокрылых в районах с разными ландшафтно-климати-
ческими условиями различался. Тундровые виды составляли основу населения лишь в сырых
и околоводных местообитаниях равнинного района. В северной (наиболее сухой) части горной
гряды в населении абсолютно преобладали различные ксерофильные элементы (при практи-
чески полном отсутствии гигрофилов и, соответственно, крайней бедности населения сырых
местообитаний), что сближало их с комплексами полужесткокрылых, населяющими высокого-
рья южного Верхоянья. В южной (более обводненной) части горной гряды сокращение участия
арктических и увеличение числа широкоареальных видов определило сходство фауны клопов
этого района с лесотундровыми фаунами. Предполагается, что высокое видовое богатство и
своеобразие комплексов полужесткокрылых изученной территории связаны с их преемствен-
ным развитием в условиях высокого ландшафтно-климатического разнообразия. Полученные
298
данные существенно дополнили список полужесткокрылых тундровых ландшафтов Азии, но не
изменили общего представления об ограниченной возможности освоения этим отрядом тундро-
вой зоны.
Ключевые слова: клопы, Северо-Восток Азии, Чукотка, зональная и горная тундры, аркти-
ческие виды, фауна, хорология, биотопическое распределение.
DOI: 10.31857/S0367144521020040
Как и другие насекомые с неполным превращением, клопы обладают весьма низким
адаптивным потенциалом к освоению тундровой зоны, что связано с особенностями
их биологии. По сравнению с другими отрядами насекомых у Heteroptera в целом
более высокий средний температурный порог развития (около 12 оС) и большая сумма
необходимых для развития эффективных температур (Саулич, Мусолин, 2007). По-
скольку южная граница тундровой зоны проходит по изотерме среднеиюльской темпе-
ратуры примерно в 12 оС, эти особенности биологии хорошо объясняют столь суще-
ственное снижение видового богатства полужесткокрылых севернее границы леса
(Чернов, 1978; Danks, 1981). Число видов, оптимум распределения которых находится
в тундровых равнинных ландшафтах, невелико и составляет всего около десяти (Ки-
риченко, 1960). Почти все они относятся к семействам Saldidae (Calacanthia trybomi
(J. Sahlberg, 1878) и два вида рода Chiloxanthus Reuter, 1891) и Miridae (в основном
виды из родов Orthotylus Fieber, 1858 и Chlamydatus Curtis, 1833). Именно эти таксоны
составляют основу фаун клопов азиатских (Кириченко, 1960; Чернов, 1978; Матис,
1986; Хрулева, Винокуров, 2007) и американских (Danks, 1981; Scudder, 1997) тундр.
Комплексы полужесткокрылых Чукотского автономного округа до сих пор недоста-
точно изучены. Наиболее подробные сведения о характере распределения клопов по
территории этого региона представлены в монографии Э. Г. Матиса (1986), еще ряд
публикаций содержит данные о местонахождении и биотопических связях отдельных
видов (Берман, 1986, 2001; Кержнер, 1988; Марусик, 1993; Берман и др., 2002). Лишь
совсем недавно сведения о фауне Heteroptera безлесных ландшафтов Чукотки были
обобщены в отдельной публикации (Vinokurov, Khruleva, 2021). В настоящем сооб-
щении рассматриваются специфика таксономического и экологического облика ком-
плексов полужесткокрылых, представленных на северо-восточном побережье Чаун-
ской губы в условиях высокого ландшафтного разнообразия среды.
РАЙОН ИССЛЕДОВАНИЯ, МАТЕРИАЛ И МЕТОДИКА
Сборы полужесткокрылых проводились в окрестностях Певека (рис. 1). Район расположен в
поясе арктического климата. Средняя температура самого холодного месяца (февраль) составля-
ет -27.5 оС. Положительные температуры воздуха наблюдаются с июня по сентябрь; среднеме-
сячные температуры в эти месяцы составляют соответственно 5.4, 8.4, 7.3 и 3 оС (данные с сайта
pogodaiklimat.ru/climate/25051.htm). Климат континентальный; годовая сумма осадков составля-
ет около 220 мм, из них примерно 100 мм приходится на безморозный период. Изученный район
находится в Западночукотском флористическом округе (Юрцев и др., 2010), в подзоне типичных
(средних гипоарктических) тундр, или подзоне D по международной классификации (Walker
et al., 2005). В растительном покрове содоминируют гипоарктические (Ledum decumbens,
Vaccinium uliginosum ssp. microphyllum, V. vitis-idaea ssp. minus, Empetrum subholarcticum)
и арктоальпийские (Dryas punctata, Cassiope tetragona, Salix phlebophylla, S. polaris) кустарнич-
ки. Кустарниковые ивняки вне поймы практически отсутствуют, а высота зарослей Betula exilis
не превышает 20-30 см.
299
Рис. 1. Места проведения работ в окрестностях г. Певек.
1 - нижнее течение р. Апапельгын; 2 - окрестности сопки Янрапаакэнай; 3 - северная часть сопки
Пээкинэй; 4 - южная часть сопки Пээкинэй; 5 - окрестности бывшего пос. Валькумей,
южная часть сопки Певек.
Низменные равнинные ландшафты (нижнее течение р. Апапельгын, примерно в 20 км СВ
Певека, 69°48ʹ N, 170°39ʹ E; рис. 2) включали долину реки с обширными заболоченными участ-
ками в низинах, окруженные полосой травяно-моховой ерниково-ивняковой тундры (средняя
высота ив 0.4-0.6 м, отдельные кусты до 1 м), а также пологие увалы с полидоминантной мохо-
во-кустарничковой тундрой. На сухих буграх в долине реки представлены сусликовины
с разнотравно-полынным покровом, на склонах речных террас юго-западной экспозиции -
остепненные разреженные группировки с осочково-разнотравно-полынным покровом.
В горной части (изолированный горный массив, образующий небольшой полуостров протя-
женностью около 15 км) работы велись в четырех точках, расположенных на расстоянии в
5-6 км одна от другой. Наиболее подробно была изучена северная часть горного массива (бли-
жайшие окрестности г. Певек; рис. 3): склоны и предгорные шлейфы сопок Янрапаакэнай
(1 км С Певека, 69о42ʹ N, 170о21ʹ E) и Пээкинэй (примерно 1 км Ю Певека, 69о40ʹ N, 170о16ʹ E).
Часто повторяющиеся феновые ветра (так называемые южаки), отсутствие крупных водотоков и
очень незначительная площадь сырых участков определяют формирование ландшафта с преоб-
ладанием местообитаний, которые становятся сухими уже к концу первого летнего месяца.
На пологих предгорных шлейфах различной экспозиции распространены полидоминантные
злаково-кустарничковые моховые тундры, на щебнистых склонах преобладают разнотравно-
кустарничково-дриадовые пятнистые тундры, в том числе на крутых склонах южных румбов -
остепненные разнотравно-осочково-дриадовые группировки. Сырые местообитания занимают
небольшие площади в полосах стока на склонах и нагорных террасах под снежниками. Ивняки
высотой до 1-1.5 м представлены только по долинам ручьев. Наиболее сухо в окрестностях
сопки Янрапаакэнай, что, видимо, связано с ее изолированным положением, небольшой высотой
и открытостью для ветров всех направлений.
Мезоклимат южной части горной гряды не столь суров, о чем свидетельствуют увеличение
участия в растительном покрове гипоарктических кустарничков, а также высота и площадь
300
Рис. 2. Местообитания равнинного района (нижнее течение р. Апапельгын).
1 - общий вид долины реки, 2 - пойма реки, 3 - зональная полидоминантная кустарничковая тундра,
4 - сырые участки в долине реки, 5 - сухой бугор с разнотравно-полынным покровом,
6 - сусликовина с разнотравно-полынным покровом.
кустарниковых зарослей ивы и березы, появление (очень локально) зарослей ольховника Alnaster
fruticosa (рис. 4). Площадь умеренно увлажненных и сырых стаций выше, чем на севере горной
части, а ксерофитные сообщества ограничены преимущественно сухими склонам сопок южной
экспозиции. Здесь были обследованы южная оконечность сопки Пээкинэй (7 км Ю Певека,
69о38ʹ N, 170о15ʹ E) и окрестности бывшего пос. Валькумей (18 км Ю Певека, сопка Певек,
69о36ʹ N, 170о13ʹ E).
301
Рис. 3. Местообитания северного горного района (окрестности сопки Янрапаакэнай
и северной части сопки Пээкинэй).
1 - кустарничково-травяно-моховая сырая тундра на нагорной террасе под снежником, 2 - зональная
полидоминантная кустарничковая тундра на предгорном шлейфе, 3 - сухая травяно-кустарничковая
тундра на щебнистом предгорном шлейфе, 4 - щебнистый склон южной экспозиции с пятнистым
разнотравно-дриадовым покровом, 5 - остепненная разнотравно-дриадово-осочковая
группировка на крутом склоне южной экспозиции, 6 - старая вездеходная колея
на склоне сопки с разнотравно-полынным покровом.
302
Рис. 4. Местообитания южного горного района (южные оконечности сопок Пээкинэй и Певек).
1 - заросли ивняков в долине реки, 2 - кустарничковая тундра в нижней части склона, 3 - остепненная
разнотравно-осочковая группировка на склоне южной экспозиции, 4 - сырая травяно-моховая тундра
на предгорном шлейфе, 5 - зональная полидоминантная кустарничковая тундра,
6 - заросли ольхи в долине ручья.
Исследования проводились с 25 мая по 1 августа 2011 г. В течение большей части этого
периода (с начала июня по середину июля) стояла очень теплая и сухая погода; среднемесячные
температуры июня составили 8.9, а июля - 10.3 оС, что соответственно на 3.5 и 1.9 оС выше сред-
немноголетних значений. Большинство клопов было собрано кошением энтомологическим
сачком, которое проводилось во всех районах в дневное время (обычно 50-100 взмахов за один
укос в каждом биотопе). Небольшая часть клопов собрана просеиванием подстилки и ручным
сбором. В окрестностях Певека и в нижнем течении р. Апапельгын использовались также поч-
303
венные ловушки (линии из пластиковых стаканов объемом 200 мл, на треть заполненных водой
с добавлением поваренной соли). Видовой состав, общее число собранных экземпляров, а также
объем количественных учетов в каждой из изученных точек приведены в табл. 1.
Ареалогия видов дана по каталогу полужесткокрылых азиатской части России (Винокуров и
др., 2010); при обобщении данных по экологии и трофическим связям использованы следующие
литературные источники: Böcher, 1971; Винокуров, 1979, 2003, 2005, 2008; Danks, 1981; Матис,
1986; Берман, 1986, 2001; Кержнер, 1988; Scudder, 1997; Степанов, 2003; Зиновьева, 2006, 2013;
Макарова, Макаров, 2006; Хрулева, 2007, 2014; Хрулева, Винокуров, 2007, 2009; Böcher,
Nachman, 2011. Анализ широтного распределения видов проводился с учетом зоны их экологи-
ческого оптимума (определяемой на основе регулярности встреч, численности, особенностей
внутриландшафтного распределения видов в районах с различными природными условиями),
а не общей широтной составляющей их ареалов. К арктическим отнесены виды, оптимум рас-
пределения которых в равнинных ландшафтах лежит севернее границы леса (Чернов, Матвеева,
2002). Большинство из них (клопы-прибрежники Chiloxanthus arcticus, Ch. stellatus stellatus,
Calacanthia trybomi и слепняки Orthotylus artemisiae, O. bermani, Chlamydatus wilkinsoni) входит
в число наиболее характерных компонентов тундровых фаун полужесткокрылых различных ре-
гионов, а за пределами равнинных тундр встречается спорадично (в основном в субарктических
высокогорьях). Гораздо шире в горных ландшафтах распространен только Nysius ericae
groenlandicus (Lygaeidae), аркто-монтанный подвид степного Nysius ericae ericae (Schilling,
1829). Помимо арктических в группу криофилов включены виды, имеющие высокую актив-
ность в гипоарктических ландшафтах, расположенных южнее границы тундровой зоны (северо-
таежных редколесьях и лесотундре). К ним отнесены несколько видов с гипоаркто-борео-
монтанными ареалами (прибрежник Salda littoralis, слепняки Teratocoris saundersi, T. viridis,
Psallus aethiops и Ps. betuleti), а также кружевница Acalypta elegans и слепняк Leptopterna
ferrugata, имеющие преимущественно гипоарктическое распространение. В группу южных
видов включены клопы с полизональными и температными (Городков, 1984) ареалами, оптимум
распределения которых лежит в бореальных (иногда также суббореальных) ландшафтах. Клас-
сификационные процедуры выполнены в программе PAST.
РЕЗУЛЬТАТЫ
Общая характеристика фауны окрестностей Певека
На изученной территории собрано 32 вида клопов, относящихся к 8 семействам
(табл. 1). Основу фауны составили Saldidae (5 видов) и Miridae (16 видов); остальные
семейства представлены единичными (1-3) видами. Особенность состава фауны по
типу долготного простирания ареалов - преобладание видов с голарктическими ареа-
лами (25 видов, 78 % фауны), из которых два имеют сибирско-американское (клопы
рода Orthotylus) и один (Antheminia eurynota remota) - центральноазиатско-восточно-
сибирско-американское распространение, у большинства остальных видов транс-
палеарктический или трансевразиатский тип ареала, исключение составляет лишь
восточносибирско-дальневосточный Leptopterna kerzhneri. Состав фауны по набору
широтных групп весьма разнороден. Значительную часть (18 видов, около 56 %
фауны) составили южные виды, оптимум распределения которых находится преиму-
щественно в бореальных ландшафтах. Остальные виды относятся к криофилам, среди
которых клопы с арктическим и преимущественно гипоарктическим распростране-
нием представлены равным числом (по 7 видов, 22 % фауны).
Отличительная особенность экологического облика изученной фауны - высокое раз-
нообразие ксеро- и мезоксерофильных видов (11 видов из 10 родов и 6 семейств, всего
34
% фауны). В их составе присутствуют широкоареальные мезоксерофилы
304
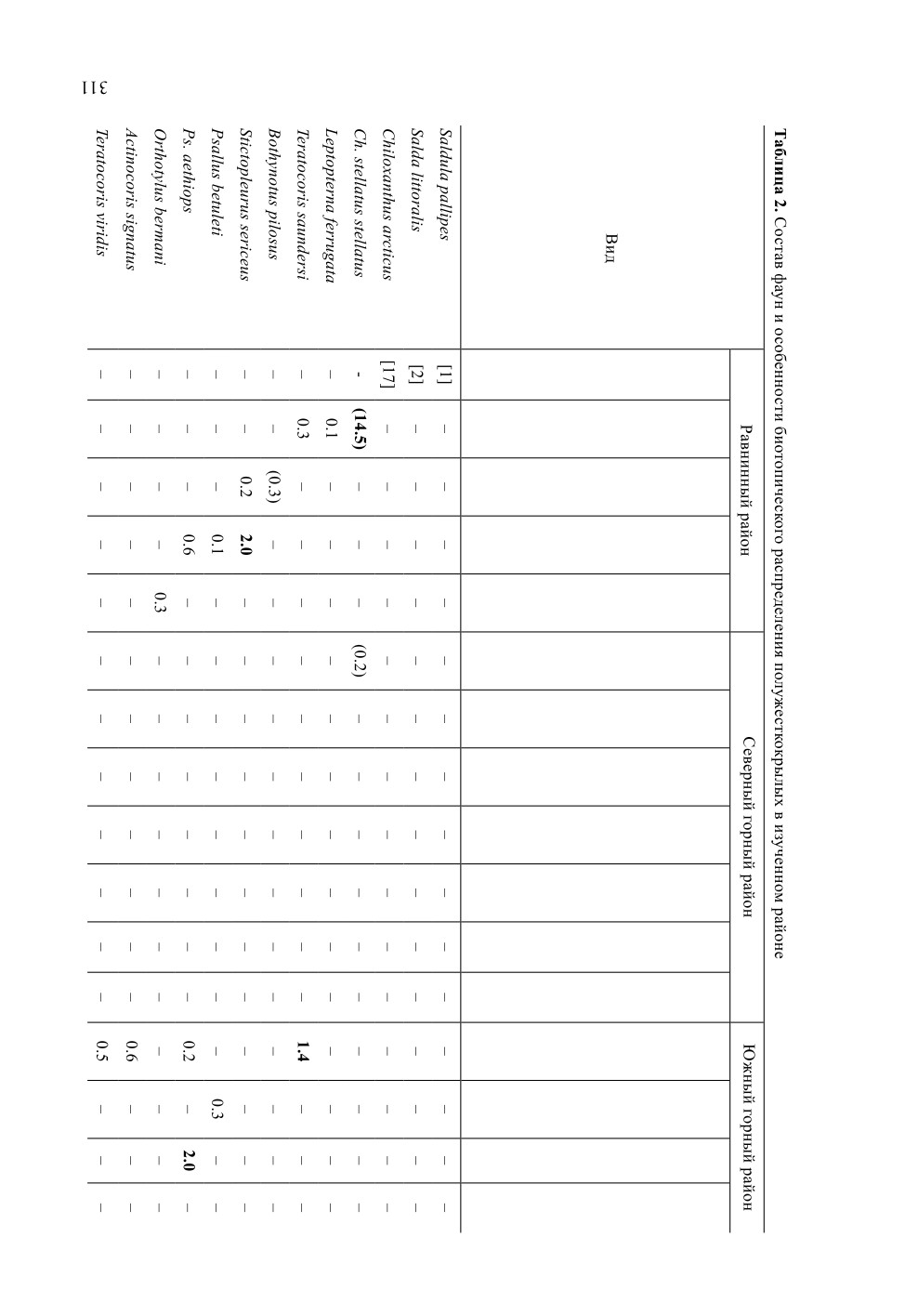
Таблица 1. Состав фаун наземных полужесткокрылых в разных точках в окрестностях Певека
Точка сборов
Таксон
Апап
Янр
СПэк
ЮПэк
Вальк
Сем. Saldidae
Chiloxanthus arcticus (J. Sahlberg,
Г
А
Г
17
-
-
-
-
1878)
Ch. stellatus stellatus (Curtis, 1835)
Г
А
Г
68
-
1
-
-
Calacanthia trybomi (J. Sahlberg,
Г
А
Кт
-
22
6
-
-
1878)
Saldula pallipes (Fabricius, 1794)
Г
П
Г
1
-
-
-
-
Salda littoralis (Linnaeus, 1758)
Г
ГБМ
Г
2
-
-
-
-
Сем. Miridae
Bothynotus pilosus (Boheman, 1852)
Г
Б
Э
1
-
-
-
-
Apolygus lucorum (Meyer-Dür, 1843)
П
П
Э
-
-
-
-
1
Lygus rugulipennis Poppius, 1911
Г
П
Э
10
-
1
4
7
Actinocoris signatus Reuter, 1878
Г
Б
Г
-
-
-
4
1
Leptopterna ferrugata (Linnaeus,
Г
Г
Г
1
-
-
-
-
1758)
L. kerzhneri Vinokurov, 1982
ВП
ГБМ
Э
-
3
-
-
-
Teratocoris saundersi Douglas
Г
ГБМ
Г
3
-
-
3
10
et Scott, 1869
T. viridis Douglas et Scott, 1867
Г
ГБМ
Г
-
-
-
3
1
Orthotylus artemisiae (J. Sahlberg,
СА
А
Кт
-
39
24
-
4
1878)
O. bermani Kerzhner, 1988
CА
А
Г
1
-
-
-
-
Chlamydatus pullus Kerzhner, 1988
Г
П
Кш
1
96
20
15
1
Ch. wilkinsoni (Douglas et Scott,
Г
А
Э
-
-
1
-
-
1866)
Europiella artemisiae (Becker, 1864)
Г
Б
Кш
36+
15
25
37
22+
Psallus betuleti (Fallén, 1826)
Г
ГБМ
Э
1
-
-
-
3
Ps. aethiops (Zetterstedt, 1838)
Г
ГБМ
Э
8
-
-
10
Ps. anticus (Reuter, 1876)
П
Б
Кс
-
3
-
-
2
Сем. Tingidae
Acalypta elegans Horváth, 1906
Г
Г
Э
-
3
-
-
-
A. nigrina (Fallén, 1807)
П
П
Э
-
1
1
-
-
305
Таблица 1 (продолжение)
Точка сборов
Таксон
Апап
Янр
СПэк
ЮПэк
Вальк
Сем. Aradidae
Aradus lugubris Fallén, 1807
Г
Б
Э
-
1
-
-
-
Сем. Lygaeidae
Nysius ericae groenlandicus
Г
А
Кт
92+
73+
21
6
57+
(Zetterstedt, 1838)
Trapezonotus desertus Seidenstücker,
Г
Б
Кш
-
-
-
2
-
1951
Сем. Coreidae
Coriomerus scabricornis (Panzer,
П
П
Кш
-
15
2
3
-
1805)
Сем. Rhopalidae
Stictopleurus crassicornis (Linnaeus,
П
П
Э
-
-
-
-
2
1758)
S. sericeus (Horváth, 1896)
П
П
Кс
19
-
-
-
-
Сем. Pentatomidae
Antheminia eurynota remota (Horváth,
СА
М
Кс
-
14
4
1
1
1907)
Sciocoris microphthalmus Flor, 1860
Г
П
Э
-
-
-
-
1
Rhacognathus punctatus (Linnaeus,
П
П
Э
-
-
-
-
1
1758)
Всего видов
15
12
11
11
15
Всего экземпляров
263
282
105
88
111
Число ловушко-суток
874
3341
1507
90
-
Число укосов
2750
4275
1800
900
1250
П р и м е ч а н и е. «-» - вид не отмечен; «+» - массовый вид, собрана лишь часть встреченных особей.
Долготное распространение: ВП - восточнопалеарктическое, Г - голарктическое, П - палеарктическое,
СА - сибирско-американское. Широтное распределение: А - преимущественно арктическое, Б - преиму-
щественно бореальное, Г - преимущественно гипоарктическое, ГБМ - преимущественно гипоаркто-борео-
монтанное, М - монтанное, П - полизональное. Экологические группы: Г - гигрофилы, К - ксерофилы
и мезоксерофилы (Кс - степные, Кт - тундровые, Кш - широкие), О - околоводные, Э - экологически
пластичные виды. Обозначение точек сбора: Апап - нижнее течение р. Апапельгын; Вальк - окрестности
бывшего пос. Валькумей, южная часть сопки Певек; СПэк - северная часть сопки Пээкинэй; ЮПэк - южная
часть сопки Пээкинэй; Янр - окрестности сопки Янрапаакэнай.
306
(Chlamydatus pullus, Europiella artemisiae, Trapezonotus desertus, Coriomerus
scabricornis), степные (Psallus anticus, Stictopleurus sericeus, Antheminia eurynota
remota), а также тундровые виды (Calacanthia trybomi, Orthotylus artemisiae,
O. bermani, Nysius ericae groenlandicus). Гигрофильные (включая околоводные)
виды представлены беднее (8 видов,
25
% фауны). В отличие от ксерофилов,
они относятся лишь к двум таксонам: сем. Saldidae и трибе Stenodemini
сем. Miridae; среди них преобладают криофильные виды (см. табл. 1): Chiloxanthus
spp., Salda littoralis, Leptopterna ferrugata, Teratocoris spp. Все прочие виды относятся
к экологически пластичным. Многие из них (Bothynotus pilosus, Acalypta nigrina,
Aradus lugubris, Rhacognathus punctatus, Sciocoris microphthalmus) - обитатели пре-
имущественно лесных биомов (Матис, 1986; Винокуров и др., 1988), хотя некоторые
(например Lygus rugulipennis) регулярно встречаются и севернее границы леса (Матис,
1986). Криофилы в этой группе представлены Chlamydatus wilkinsoni, Psallus aethiops
и Ps. betuleti.
По типу пищевой специализации в составе фауны преобладают фитофаги. Хищники
(около 22 % фауны) представлены клопами-прибрежниками (пять видов), щитником
Rhacognathus punctatus и слепняком Bothynotus pilosus. За исключением трех видов
(мицетофага Aradus lugubris и питающихся на мхах кружевниц рода Acalypta), фито-
фаги трофически связаны с высшими растениями. Значительную долю их составляют
широкие фитофаги, встречающиеся обычно в смежных ярусах: в травостое и кустар-
никовом ярусе (как Lygus rugulipennis и Stictopleurus crassicornis), на почве и в траво-
стое (Chlamydatus wilkinsoni, Nysius ericae groenlandicus), реже - только в одном ярусе
(Chlamydatus pullus в травостое, Trapezonotus desertus в подстилке). Среди видов,
ограниченных в питании одним семейством (олигофаги) или несколькими близкими
семействами (узкие полифаги), преобладают хортобионты. Шесть видов трофически
связаны с однодольными растениями: щитник Sciocoris microphthalmus и пять видов
слепняков трибы Stenodemini (роды Actinocoris, Leptopterna и Teratocoris). Среди
олигофагов на двудольных преобладают питающиеся на полынях (Artemisia,
сем. Asteraceae) Apolygus lucorum, Orthotylus artemisiae, Europiella artemisiae и
Stictopleurus sericeus. С бобовыми (Fabaceae) связан Coriomerus scabricornis; еще у од-
ного вида, Orthotylus bermani, в Северной Америке отмечено питание на сложно-
цветных рода Saussurea (Scudder, 1997), а в Азии - на бобовых рода Oxytropis
(Кержнер, 1988; Хрулева, 2007). Строгие тамнобионты представлены в фауне только
видами рода Psallus - зоофитофагами, связанными преимущественно с растениями из
родов Salix и Betula, а Ps. anticus - со Spiraea.
Состав локальных фаун
В пяти обследованных точках (см. рис. 1) собрано от 11 до 15 видов (см. табл. 1);
в каждой из них были специфические (отмеченные только в одной точке) виды, число
которых варьировало от одного до семи. Если не брать в расчет три вида сем. Saldidae,
собранных в долине р. Апапельгын (где были обследованы пойменные местооби-
тания), такие виды особенно разнообразно представлены на южной оконечности
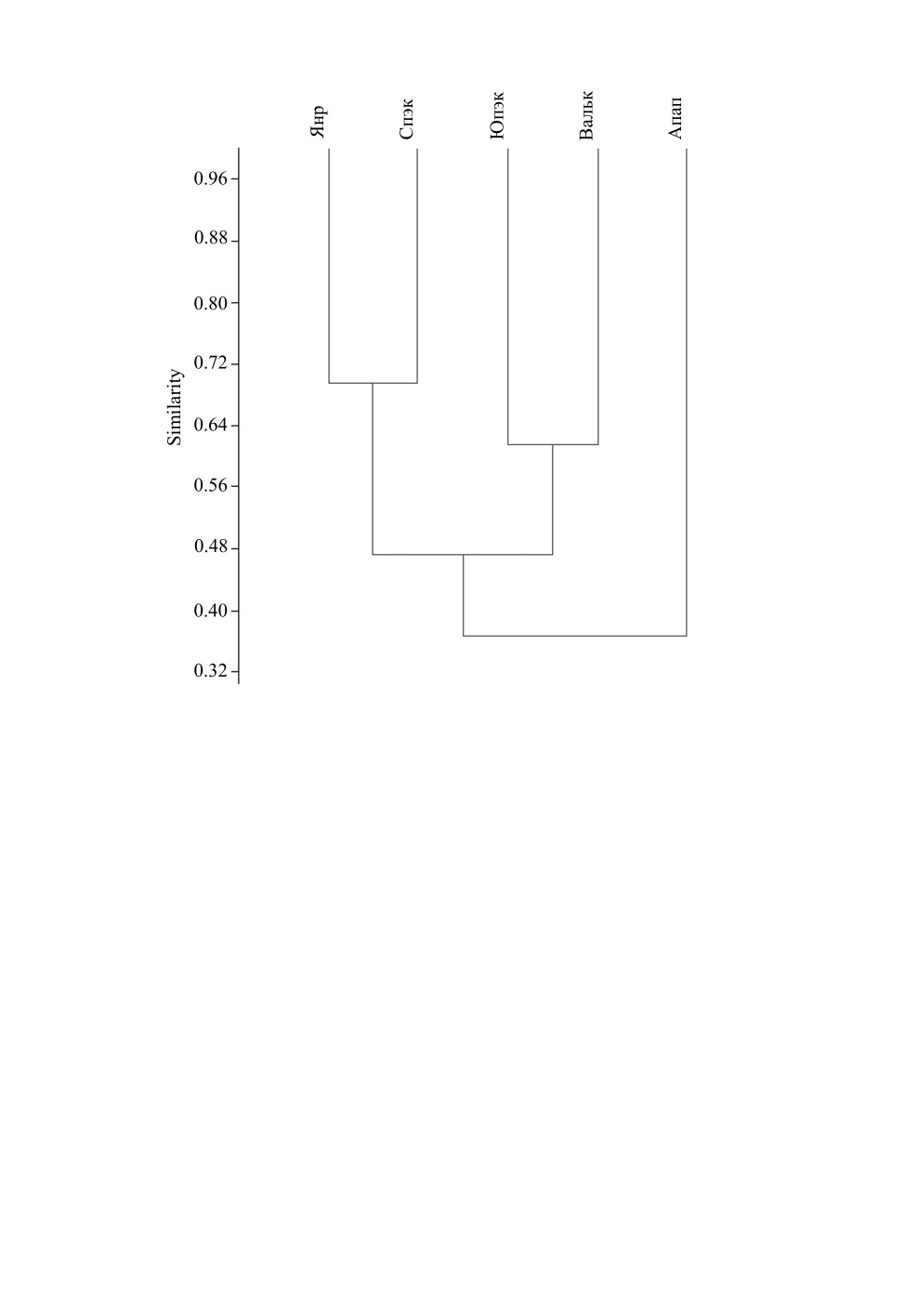
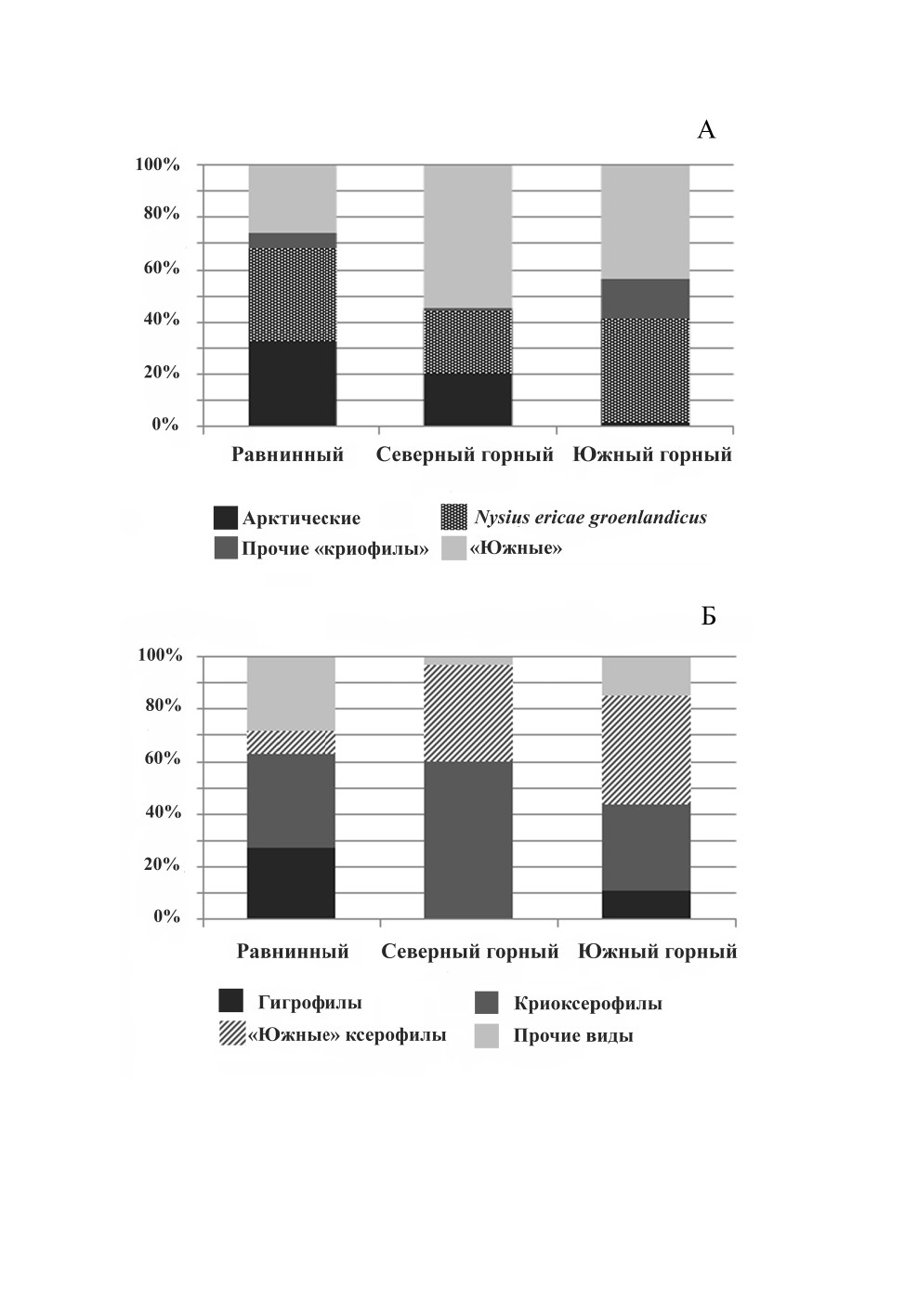
горной гряды. На дендрограмме (рис. 5) фауны изученных точек объединились в соот-
ветствии с их ландшафтным положением. Наиболее обособленные позиции заняла
фауна равнинного района (15 видов), еще две пары образовали фауны северного и юж-
ного горных ландшафтных районов. Характерно, что суммарное видовое богатство
307
Рис. 5. Дендрограмма сходства видового состава локальных фаун клопов в окрестностях
Певека (PAST program, Dice index, paired group).
Обозначение районов: Апап - нижнее течение р. Апапельгын; Вальк - окр. пос. Валькумей, южная часть
сопки Певек; СПэк - северная часть сопки Пээкинэй; ЮПэк - южная часть сопки Пээкинэй;
Янр - окрестности сопки Янрапаакэнай.
северного горного района, несмотря на его наилучшую изученность (см. табл. 1),
оказалось несколько ниже, чем южного (15 и 18 видов соответственно). Можно
ожидать, что действительное видовое богатство последнего района еще выше.
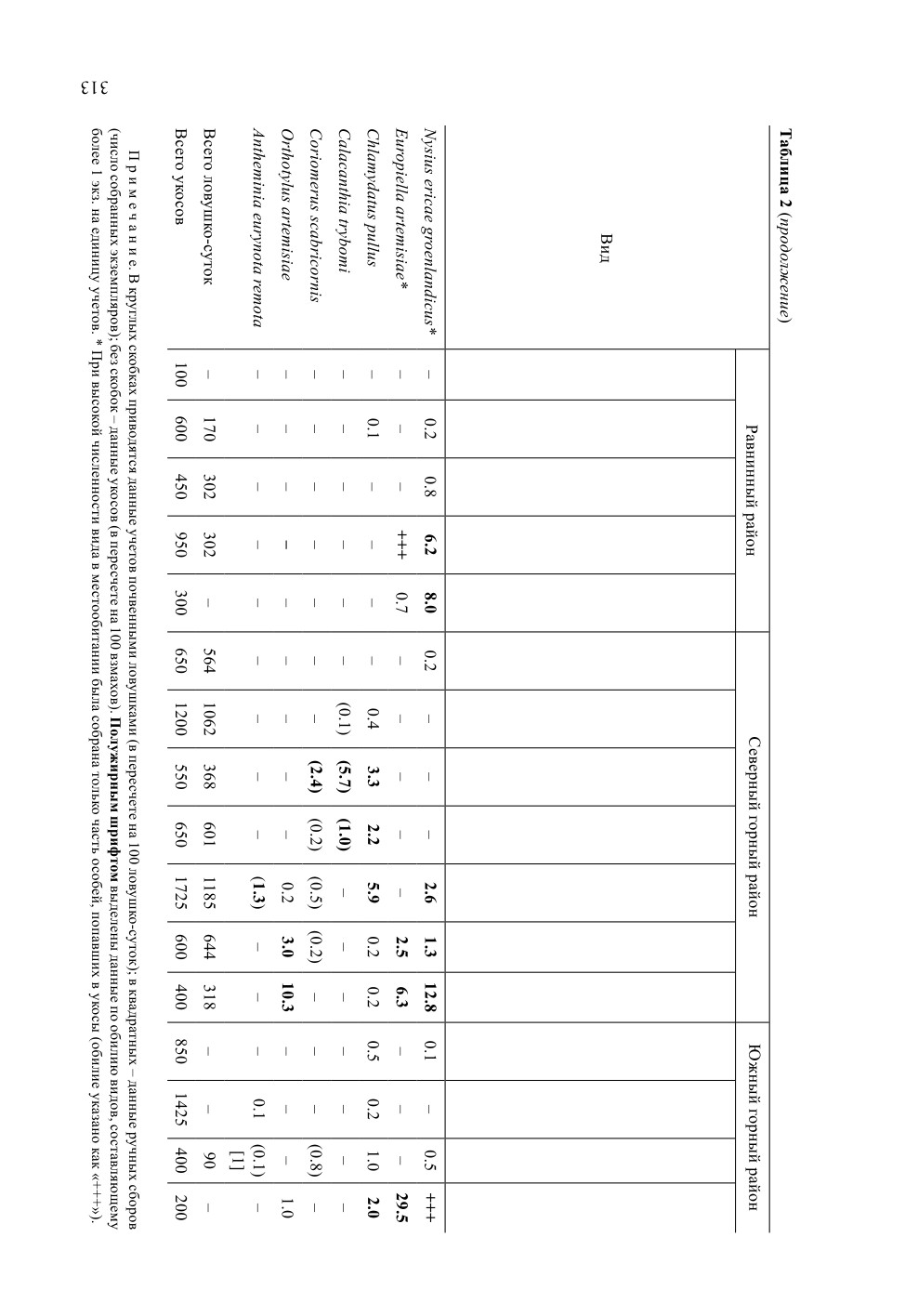
Сравнение широтного состава локальных фаун (рис. 6, А) показывает, что сум-
марная доля криофильных видов наиболее высока в равнинном районе (более поло-
вины от числа собранных). В горной части доля криофилов в локальных фаунах со-
ставляет около трети, а их состав заметно различается от района к району: на севере
горной гряды встречались в основном арктические виды, а на юге - криофилы с гипо-
арктическим распространением.
Сравнение экологического состава локальных фаун показывает, что ксерофильные
виды гораздо более равномерно заселяют изученную территорию, чем представители
прочих экологических групп. Особенно существенно от точки к точке варьировало
число гигрофилов, которые почти полностью отсутствовали в фаунах северного
308
Рис. 6. Особенности локальных фаун клопов в окрестностях Певека по широтному (А)
и экологическому составу (Б).
Обозначения районов как на рис. 5.
309
горного района (рис. 6, Б). С этими особенностями тесно связан и состав локальных
фаун по типам трофической специализации. Например, на севере горной гряды почти
не представлены олигофаги на однодольных растениях (большинство из которых от-
носится к группе гигрофилов), тогда как олигофаги на полынях (преимущественно
ксерофилы) распространены повсеместно.
Особенности биотопической приуроченности видов
Население клопов аналогичных типов местообитаний существенно варьировало от
района к району (табл. 2). Отличительная особенность равнинного района - высокое
обилие арктических видов клопов-прибрежников рода Chiloxanthus в местообитаниях
гидросерии: на галечниковой пойме - Ch. arcticus, а в переувлажненной депрессии -
Ch. stellatus stellatus. Во всех обследованных сухих биотопах (на зоогенных суслико-
винах с доминированием полыни Artemisia tilesii, сухих бровках с разнотравьем вдоль
грунтовых дорог) высокое обилие имел криоксерофил Nysius ericae groenlandicus.
На сухом бугре, где в массе росла петрофильная полынь Artemisia kruhsiana, помимо
N. e. groenlandicus высоким было обилие широкого ксерофила Europiella artemisiae и
степного Stictopleurus sericeus. Находка здесь последнего вида особенно интересна,
поскольку он не был найден ни в одном из биотопов горной части. Самое низкое
обилие клопов отмечено в зональной полидоминантной кустарничковой тундре, где
собраны единичные экземпляры нескольких видов.
На севере горной гряды в плакоротипных местообитаниях на умеренно увлаж-
ненных предгорных шлейфах клопы также были малочисленны. В других местооби-
таниях ландшафтного профиля их распределение резко отличалось от предыдущего
района: клопы практически полностью отсутствовали в сырых биотопах, тогда как в
сухих набор их был заметно разнообразнее. Здесь повсеместно встречались
Chlamydatus pullus и Coriomerus scabricornis, на щебнистых предгорных шлейфах и
склонах неюжных румбов к ним добавлялся арктический Calacanthia trybomi,
а на южных склонах - горностепной Antheminia eurynota remota. На зоогенных луго-
винах и участках с нарушенным покровом (в растительном покрове присутствовала
Artemisia tilesii) высокое обилие имели Eu. artemisiae, Orthotylus artemisiae и Nysius e.
groenlandicus. Последние два вида в этом районе отмечались также на склонах южной
экспозиции, но в гораздо меньшем обилии. Таким образом, основное своеобразие про-
странственной организации группировок клопов северной части горной гряды - засе-
ление ими преимущественно сухой части ландшафтного спектра, где совместно оби-
тали различные ксерофильные виды (в основном криоксерофилы и широкие
мезоксерофилы).
Отсутствие в южной горной части материалов из почвенных ловушек не позволило
получить данные по распределению видов, обитающих в наземном ярусе. Тем не
менее, некоторые из них в этом районе были собраны с помощью других методов (на-
пример, Trapezonotus desertus, A. eu. remota), но арктических видов клопов-прибреж-
ников среди них не было. Учитывая, что эти клопы обычно хорошо заметны благодаря
их передвижению прыжками, можно предположить, что если они и присутствуют в
данном районе, то в очень низком обилии. Укосов в этом районе также было прове-
дено меньше, чем в других (см. табл. 1), однако число видов, собранное с их помощью,
оказалось больше (16 и 9-10 видов соответственно). В группировках сырых местооби-
310
Галечниковая пойма
Переувлажненные участки в
долине реки
Умеренно увлажненный склон
увала
Сухой бугор и прилегающие
участки в долине реки
Участки с нарушенным
покровом
Переувлажненные участки на
склонах сопок
Умеренно увлажненные участки
предгорного шлейфа
Сухой предгорный шлейф
Склоны сопки северо-западной
экспозиции
Склоны сопок южной
экспозиции
Зоогенные луговины
Участки с нарушенным
покровом
Переувлажненные участки на
предгорном шлейфе
Умеренно увлажненные участки
на предгорном шлейфе
Склоны сопок южной
экспозиции
Участки с нарушенным
покровом
Галечниковая пойма
Переувлажненные участки в долине
реки
Умеренно увлажненный склон
увала
Сухой бугор и прилегающие
участки в долине реки
Участки с нарушенным покровом
Переувлажненные участки на
склонах сопок
Умеренно увлажненные участки
предгорного шлейфа
Сухой предгорный шлейф
Склоны сопки северо-западной
экспозиции
Склоны сопок южной экспозиции
Зоогенные луговины
Участки с нарушенным покровом
Переувлажненные участки на
предгорном шлейфе
Умеренно увлажненные участки на
предгорном шлейфе
Склоны сопок южной экспозиции
Участки с нарушенным покровом
Галечниковая пойма
Переувлажненные участки в долине
реки
Умеренно увлажненный склон
увала
Сухой бугор и прилегающие
участки в долине реки
Участки с нарушенным покровом
Переувлажненные участки на
склонах сопок
Умеренно увлажненные участки
предгорного шлейфа
Сухой предгорный шлейф
Склоны сопки северо-западной
экспозиции
Склоны сопок южной экспозиции
Зоогенные луговины
Участки с нарушенным покровом
Переувлажненные участки на
предгорном шлейфе
Умеренно увлажненные участки на
предгорном шлейфе
Склоны сопок южной экспозиции
Участки с нарушенным покровом
таний численно преобладали гигрофильные представители трибы Stenodemini -
Actinocoris signatus, Teratocoris saundersi и T. viridis. На умеренно увлажненных
участках и склонах южной экспозиции все виды встречались в небольшом обилии.
Криофилы были представлены двумя видами рода Psallus и Nysius ericae groenlandicus,
тогда как набор южных элементов отличался разнообразием: помимо видов, из-
вестных из других ландшафтных районов (Lygus rugulipennis, Chlamydatus pullus,
Psallus anticus, Coriomerus scabricornis, Antheminia eu. remota), здесь были собраны
Apolygus lucorum, Trapezonotus desertus, Stictopleurus crassicornis и Rhacognathus
punctatus. Самая высокая численность клопов в укосах отмечена на нарушенных
участках вдоль дорог с доминированием полыней. Только в этих биотопах наряду
с южными видами (Lygus rugulipennis, Europiella artemisiae, Chlamydatus pullus) в
массе отмечен криоксерофильный N. e. groenlandicus.
Основные черты пространственного распределения клопов
на изученной территории
Почти три четверти видов встречались в районе исследования локально и преиму-
щественно с низкой численностью. Среди достаточно обычных (найденных в двух и
более точках, в общей сложности не менее 15 экз.) преобладали ксерофилы -
Calacanthia trybomi, Orthotylus artemisiae, Nysius ericae groenlandicus, Europiella
artemisiae, Chlamydatus pullus, Coriomerus scabricornis, Antheminia eurynota remota.
Прочие экологические группы представлены гигрофильными Chiloxanthus stellatus
stellatus, Teratocoris saundersi и политопным Lygus rugulipennis.
Среди всех этих видов наиболее широкое распространение и высокое обилие имели
широкие полифаги (Ch. pullus, N. e. groenlandicus) и олигофаги на полынях
(O. artemisiae, Eu. artemisiae), в общей сложности на долю этих четырех видов при-
шлось около 44 % от числа всех собранных экземпляров клопов. Все они в различных
сочетаниях встречались в сухих местообитаниях, в основном (кроме Ch. pullus) до-
стигая максимального обилия на участках с нарушенным покровом и обилием по-
лыней (сусликовины, обочины дорог, колеи на склонах сопок). Помимо местооби-
таний с нарушенным покровом достаточно устойчивое по видовому составу население
клопов (состоящее из Ch. pullus, N. e. groenlandicus, Coriomerus scabricornis,
Antheminia eu. remota) в горной части было обнаружено на щебнистых склонах южной
экспозиции с пятнистым осочково-разнотравно-дриадовым покровом.
Напротив, видовой состав и обилие клопов в сырых местообитаниях существенно
различались от района к району. На севере горной гряды, где сырые участки встреча-
ются небольшими пятнами, собраны лишь единичные экземпляры клопов. В районах
с широким распространением переувлажненных стаций группировки клопов были до-
статочно богаты и состояли преимущественно из гигрофилов: в равнинном абсолютно
доминировал Chiloxanthus s. stellatus, а в южном горном - виды трибы Stenodemini.
Во всех зональных и близких к ним местообитаниях (полидоминантные кустарнич-
ковые тундры на умеренно увлажненных склонах увалов и предгорных шлейфов) на-
селение клопов было повсеместно малочисленно и не имело в своем составе ста-
бильных элементов. Тем не менее, именно в местообитаниях этих типов были собраны
многие крайне редкие для тундровой зоны виды - Bothynotus pilosus, Apolygus lucorum,
314
Psallus anticus, Aradus lugubris, Stictopleurus crassicornis, Rhacognathus punctatus. Их
концентрация была особенно высока в кустарничковых тундрах южного горного
района.
Обобщение всех имеющихся данных свидетельствует о наличии на изученной тер-
ритории трех комплексов полужесткокрылых, представленных в соответствующих
ландшафтных районах: равнинном, северном и южном горных. В каждом из них отме-
чены специфические виды (6-7 в каждом районе), причем некоторые - в значительном
обилии. В равнинном районе это Stictopleurus sericeus, в северном горном - Calacanthia
trybomi, а в южном горном - Actinocoris signatus и Teratocoris viridis. Комплексы полу-
жесткокрылых этих районов также различались по набору фоновых видов, особенно-
стям заселения ландшафтного профиля, суммарному участию в населении представи-
телей различных широтных (рис.7, А) и экологических (рис. 7, Б) групп. Так, от
равнинного к южному горному району в населении клопов отмечено заметное уси-
ление южных черт за счет сокращения доли арктических видов. Экологический облик
населения равнинного района определяет высокая доля в нем гигрофильных эле-
ментов, что, несомненно, в значительной степени связано с ландшафтными особенно-
стями этого района. Комплексы полужесткокрылых горной части, напротив, состоят
преимущественно из ксерофильных видов; на севере горной гряды на их долю прихо-
дится почти 97 % от числа всех собранных экземпляров.
Характер распределения большинства видов позволяет предположить, что для
многих из них существенным лимитирующим фактором на изученной территории вы-
ступает не только теплообеспеченность, но и сухость почвы. Это относится не только
к таким явным гигрофилам, как Actinocoris signatus, Leptopterna ferrugata, Teratocoris
saundersi, T. viridis, но и к большинству видов с высокой экологической пластично-
стью
- Bothynotus pilosus, Apolygus lucorum, Rhacognathus punctatus, Sciocoris
microphthalmus, Psallus aethiops, Ps. betuleti. Все они не были отмечены (или, как
L. rugulipennis, крайне малочисленны) в наиболее сухом северном горном районе, не-
смотря на его хорошую изученность, зато к нему явно тяготели (или были им ограни-
чены) такие ксерофильные элементы, как Chlamydatus pullus, Calacanthia trybomi и
Orthotylus artemisiae.
ОБСУЖДЕНИЕ
Особенности таксономического состава фауны
Ранее c изученной территории были известны шесть видов клопов (Матис, 1986):
Chiloxanthus arcticus, Saldula pallipes, Teloleuca pellicens (Fabricius, 1779), Orthotylus
artemisiae, Psallus betuleti и Nysius ericae groenlandicus. Из них в наших сборах отсут-
ствовал лишь T. pellicens, который и в материалах Э. Г. Матиса (1986) с Чукотки был
представлен существенно беднее, чем остальные пять видов. В результате проведен-
ного исследования к списку фауны добавлено 27 видов и их общее число достигло 33.
В настоящее время это самое большое число видов, собранных в тундровой зоне на
относительно небольшой территории (с наибольшей протяженностью около 40 км).
Ранее самая богатая фауна наземных полужесткокрылых (24 вида) была известна из
одного из районов Большеземельской тундры (несколько точек в верхнем течении
р. Шапкина, подзона южных тундр) (Зиновьева, 2006).
315
Рис. 7. Процентная доля разных широтных (А) и экологических (Б) групп
в населении клопов разных ландшафтных районов
окрестностей г. Певек.
316
Поскольку часть обнаруженных в этом районе видов широко распространена в
гипоарктических равнинных и (или) горных ландшафтах Северо-Восточной Азии
(Винокуров, 1979; Берман и др., 1984, 2002; Матис, 1986; Марусик, 1993; Бухкало,
1997; Винокуров, Степанов, 2003; Хрулева, Винокуров, 2007, 2009), находки практи-
чески каждого из них на севере Чукотки вполне ожидаемы. Тем не менее, столь вы-
сокая насыщенность ими обследованной территории весьма необычна. Еще более по-
казательно присутствие в фауне видов не только неизвестных из других районов
чукотской тундры, но и в целом на Северо-Востоке Азии достаточно редких (Вино-
куров, 1979, 2003, 2008; Матис, 1986; Степанов, 2003; Vinokurov, Khruleva, 2021). К их
числу относятся Bothynotus pilosus, Apolygus lucorum, Leptopterna ferrugata, Psallus
anticus, Acalypta elegans, Aradus lugubris, Stictopleurus crassicornis, Sciocoris
microphthalmus, Rhacognathus punctatus, составляющие около 28 % состава изученной
фауны.
Не исключено, что полученные результаты частично связаны с проведением иссле-
дований в период существенного потепления климата Арктики (Алексеев и др., 2015).
Согласно данным метеостанции в окрестностях Певека (сайт pogodaiklimat.ru), на-
чиная с 2002 г. суммы среднемесячных температур месяцев с положительными темпе-
ратурами (июнь-сентябрь) были постоянно выше, чем средние за период 1961-
1990 гг.; в 2005, 2007 и 2010 гг. эти различия достигали 10 оС и более. Очевидно, это
могло привести к росту численности ряда южных видов, ранее не попадавших в сборы
из-за редкости. Очень благоприятными были погодные условия и в 2011 г., что, несо-
мненно, способствовало успешному завершению развития видов клопов, находящихся
за пределами своего климатического оптимума.
Как правило, в подзоне типичных тундр локальные фауны наземных клопов со-
держат от 4 до 6 видов (Чернов, 1978; Scudder, 1997; Макарова, Макаров, 2006; Зино-
вьева, Долгин, 2006). Лишь на центральной Чукотке (среднее течение р. Амгуэма)
было собрано 9 видов (Марусик, 1993), что сравнимо с числом видов (11-15) в изу-
ченных локальных фаунах. По-видимому, более высокое богатство фаун Чукотки свя-
зано с ландшафтным разнообразием и преемственным развитием энтомокомплексов
крайнего Северо-Востока Азии (Киселев, 1981). В пользу этого предположения ясно
свидетельствует необычно высокая насыщенность окрестностей Певека, находящихся
в арктическом климатическом поясе, южными (в том числе достаточно ограниченно
проникающими в Субарктику) видами клопов.
В настоящее время фауны клопов лишь нескольких тундровых и лесотундровых
районов Азии подробно изучены. Имеющиеся данные свидетельствуют о стабильно
повторяющихся на зональном градиенте разнонаправленных преобразованиях их так-
сономического состава (табл. 3): выпадении арктических видов из родов Calacanthia и
Chiloxanthus (Saldidae) и, напротив, резком увеличении таксономического разноо-
бразия Miridae. Регулярными компонентами лесотундровых фаун клопов становятся
представители некоторых триб слепняков, почти не проникающих в тундровую зону:
Mirini (роды Apolygum, Capsus, Lygus, Polymerus), Stenodemini (Actinocoris, Leptopterna,
Stenodema, Trigonotylus), Halticini (Labops), а также роды Europiella и Plagiognathus из
Phylini. Эти черты в различной степени проявляются и в фаунах изученных ланд-
шафтных районов; очевидно, что наиболее близка к лесотундровым фауна южного
горного района. Подобный облик характерен и для фауны клопов Телекайской рощи
317
Р. Шапкина (Больше-
земельская тундра, ют)1
Озера Харбейской
системы (там же, ют)2
О. Долгий (Печорское
море, тт)3
Р. Нижняя Агапа
(Таймыр, ют)4
Ур. Ары-Мас
(там же, ют)**
Пос. Кремянка
(Чукотский АО, ют)5
Нижнее течение р.
Апапельгын (там же, тт)*
Север горной гряды в окр.
Певека (там же, тт)*
Юг горной гряды в окр.
Певека (там же, тт)*
Среднее течение р.
Амгуэма (там же, тт)6
Р. Большая Осиновая
(там же, анклав лт)6
Пос. Походск (Якутия,
лт)7
Пос. Черский (там же,
лт)7
О. Карагинский
(Камчатский край, лт)8
Хр. Сунтар-Хаята
(Якутия, пгп)4
Хр. Сунтар-Хаята
(там же, гтп)4
Хр. Улахан-Чистай
(там же, гтп)4
Р. Шапкина (Больше-
земельская тундра, ют)1
Озера Харбейской системы
(там же, ют)2
О. Долгий (Печорское море,
тт)3
Р. Нижняя Агапа
(Таймыр, ют)4
Ур. Ары-Мас
(там же, ют)**
Пос. Кремянка
(Чукотский АО, ют)5
Нижнее течение р.
Апапельгын (там же, тт)*
Север горной гряды в окр.
Певека (там же, тт)*
Юг горной гряды в окр.
Певека (там же, тт)*
Среднее течение р. Амгуэма
(там же, тт)6
Р. Большая Осиновая
(там же, анклав лт)6
Пос. Походск (Якутия, лт)7
Пос. Черский (там же, лт)7
О. Карагинский
(Камчатский край, лт)8
Хр. Сунтар-Хаята
(Якутия, пгп)4
Хр. Сунтар-Хаята
(там же, гтп)4
Хр. Улахан-Чистай
(там же, гтп)4
(Марусик, 1993) - самого северного изолированного анклава с пойменным лесом из
Chosenia arbutifolia, расположенного на р. Левый Телекай (или Большая Осиновая).
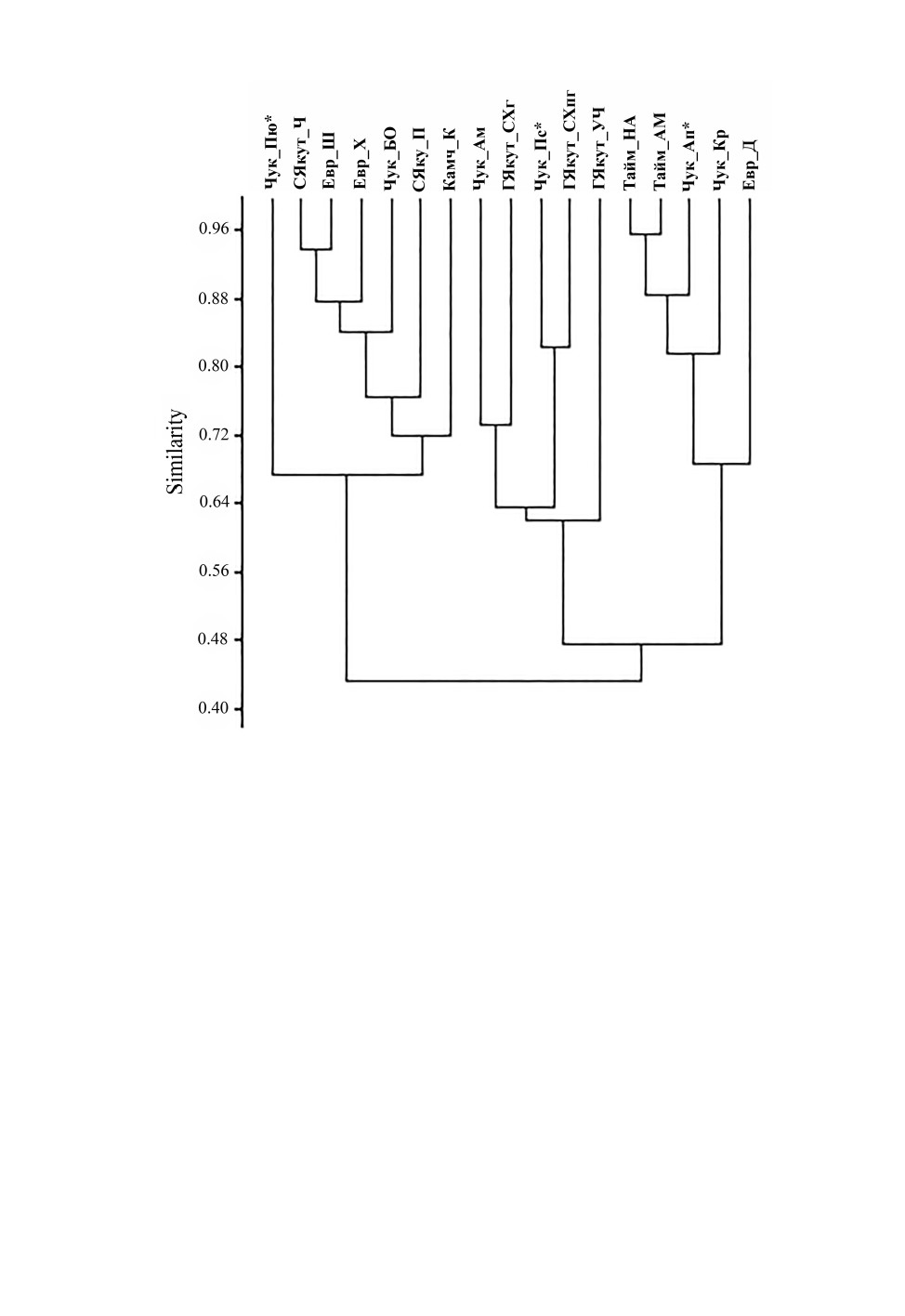
Дендрограмма (рис. 8), построенная на основе данных табл. 3, подтверждает рас-
хождение большинтсва тундровых и лесотундровых фаун по различным кластерам.
Но часть тундровых фаун, которые отличает высокое таксономическое разнообразие
слепняков и уменьшение числа арктических видов прибрежников (табл.
3),
объединилась с лесотундровыми. В их числе оказались фауны Большеземельской
тундры, а также фауна южной части горной гряды в окрестностях Певека. Из числа
изученных в тундровый кластер вошла только фауна клопов нижнего течения р. Апа-
пельгын, тогда как фауна севера горной части объединилась вместе с высокогорными
фаунами клопов Якутии. Подобное расхождение связано с различиями в их таксономи-
ческом составе. В то время как в равнинном районе он вполне традиционен для тун-
дровых фаун клопов (см. табл. 3), в горной части разнообразие семейств заметно выше
и сравнимо с таковым высокогорных фаун Якутии. Вхождение в тот же кластер еще
одной фауны, расположенной в горах центральной Чукотки, предполагает, что ком-
плексы полужесткокрылых с подобным таксономическим составом могут быть доста-
точно широко распространены на территории этого региона.
Особенности комплексов полужесткокрылых изученной территории
в сравнении с другими субарктическими и высокогорными
районами Северной Азии
Как уже отмечено, несмотря на чрезвычайно высокое видовое богатство фауны,
набор видов, достигающих на изученной территории хотя бы локально высокого
обилия, невелик. Экологический облик изученных комплексов полужесткокрылых
в значительной степени определяется достаточно высокой активностью клопов, вхо-
дящих в число характерных обитателей различных ксерофитных сообществ (в том
числе и высокогорных вариантов криофитных степей) Верхояно-Колымской горной
страны (Берман, 2001; Хрулева, Винокуров, 2007): Chlamydatus pullus, Europiella
artemisiae (Miridae), Coriomerus scabricornis (Coreidae), Stictopleurus sericeus
(Rhopalidae), Antheminia eurynota remota (Pentatomidae).
На изученной территории особенно высокую активность имеют два широких фито-
фага, Ch. pullus и Nysius ericae groenlandicus. На их долю в целом приходится около
30 % от числа всех собранных экземпляров, а в наиболее сухом (северном горном)
районе они абсолютно лидируют по числу собранных экземпляров (79 %). Интересно
отметить, что лишь эти два вида относятся к числу массовых в крайне бедной фауне
Гренландии. Очевидно, их процветание в криоаридных ландшафтах Чукотки и Грен-
ландии с совершенно разной плейстоценовой историей связано не только с ксеро-
фильностью, но и с высокой эколого-физиологической пластичностью этих видов,
проявляющейся в политрофности, способности сохранять активность в широком тем-
пературном диапазоне и формировать партеногенетические популяции (Böcher, 1971;
Böcher, Nachman, 2011).
Другая особенность трофического состава изученных комплексов - существенно
более высокая активность олигофагов на полынях по сравнению с видами, связан-
ными с другими группами растений. Показательно, что и самые устойчивые по
обилию и составу доминантов группировки клопов приурочены именно к участкам с
320
Рис. 8. Дендрограмма сходства таксономического состава локальных фаун клопов различных
районов субарктической Евразии, построенная на основе данных табл. 3 (PAST program,
Correlation index (paired group).
ГЯкут_СХг - горы Якутии, хр. Сунтар-Хаята, гольцово-тундровый пояс; ГЯкут_СХпг - там же,
подгольцовый пояс; ГЯкут_УЧ - там же, хр. Улахан-Чистай; Евр_Д - о. Долгий (Печорское море);
Евр_Х - Большеземельская тундра, озера Хабрейской системы; Евр_Ш - там же, р. Шапкина; Кам_К -
Камчатский край, о. Карагинский; СЯкут_П - северная Якутия, пос. Походск; СЯкут_Ч - там же, пос.
Черский; Тайм_АМ - Таймыр, ур. Ары-Мас; Тайм_НА - там же, р. Нижняя Агапа; Чук_Ам - Чукотский
АО, р. Амгуэма; Чук_Ап* - там же, р. Апапельгын; Чук_БО - там же, р. Большая Осиновая; Чук_Кр - там
же, Кремянка; Чук_Пс* - там же, северная часть горной гряды в окрестностях Певека; Чук_Пю* - там же,
южная часть горной гряды в окрестностях Певека. Звездочкой обозначены фауны изученных
ландшафтных районов в окрестностях Певека.
массовым произрастанием полыней (особенно Artemisia tilesii). Похожие данные были
получены и для еще одной группы Hemipteroidea: самый многочисленный в окрестно-
стях Певека вид тлей, Pleotrichophorus knowltoni Corpuz-Raros et Cook,
1974
(Aphididae), также трофически связан с полынями (Stekolshchikov, Khruleva, 2015).
Важно отметить и локальность обитания на изученной территории большинства
арктических видов (исключение составляет лишь криоксерофильный Nysius ericae
321
groenlandicus). Об их достаточно низкой активности косвенно свидетельствуют и ма-
териалы Э. Г. Матиса (1986) из окрестностей Певека: в отличие от других районов Чу-
котки, в них отсутствуют Chiloxantus stellatus, Calacanthia trybomi и Chlamydatus
wilkinsoni. Поскольку севернее границы леса эти виды встречаются в широком диапа-
зоне природных условий, включающем все подзоны тундровой зоны (табл. 4), фак-
тором, ограничивающим их распределение в окрестностях Певека, вряд ли выступает
теплообеспеченность. Нельзя, однако, исключить возможной лимитирующей роли вы-
Таблица 4. Относительное обилие некоторых наиболее обычных видов клопов с разными
типами распространения в окрестностях Певека* и других тундровых и высокогорных районах
Северной Азии
Окрестности
Певека, тт
Вид
Арктические
Chiloxanthus arcticus
++
-
++
+++
+++
-
-
-
-
Ch. stellatus stellatus
++
-
++
+++
+++
+++
+++
-
+
Calacanthia trybomi
-
++
++
+++
+++
+++
++
+
+
Chlamydatus wilkinsoni
-
+
++
+++
+
++
+++
++
+
Orthotylus artemisiae
-
++
++
++
+++
-
-
+
-
O. bermani
+
-
-
-
-
++
-
-
-
Nysius groenlandicus
+++
+++
-
-
+
+
++
-
-
Прочие
+
Europiella artemisiae
+++
+++
-
-
-
-
-
-
+++
Lygus rugulipennis
++
+
-
-
-
-
-
-
+
Chlamydatus pullus
+
+++
-
++
+++
-
++
++
+++
Coriomeris scabricornis
-
++
-
-
-
-
-
+
+
Antheminia eurynota remota
-
++
-
-
-
-
-
-
+
П р и м е ч а н и е. * приводятся данные по двум наиболее полно изученным районам. «-» - вид не
найден; «+» - единичные находки; «++» - вид локально обычен; «+++» - многочислен. Использованная
литература: 1 - Чернов, 1978; 2 - Хрулева, Винокуров, 2007; 3 - Матис, 1986; 4 - Хрулева, 1987, 2014. Выделы
зональной и поясной классификаций: ат - подзона арктических тундр, гтп - гольцово-тундровый высотный
пояс, лт - ландшафты лесотундрового облика, пгп - подгольцовый высотный пояс, тт - подзона типичных
тундр, ют - подзона южных тундр.
322
сокой континентальности климата (в сочетании с особенностями местной орографии),
а также истории развития ландшафтов этой территории в прошлом.
Почти полное отсутствие тундровых гигрофилов в переувлажненных местообита-
ниях самого сухого (северного горного) района, а также содоминирование в сухих ста-
циях тундровых и широких ксерофилов - черты, сближающие комплексы клопов
окрестностей Певека и гольцово-тундрового пояса южного Верхоянья (см. табл. 4).
Очевидно, что подобный характер распределения клопов можно связать с общей сухо-
стью и низкой обводненностью обоих районов. Пока недостаточно данных для заклю-
чения об их близости к плейстоценовым тундростепным комплексам полужестко-
крылых, однако своеобразие населения клопов северного горного района в сравнении
с таковыми других тундровых территорий косвенно свидетельствует о его реликтовом
статусе.
По сравнению с северным горным в двух других ландшафтных районах заметно
выше разнообразие гигрофильных и экологически пластичных видов; последние
представлены преимущественно южными элементами. В равнинном районе арктиче-
ские виды, тем не менее, составляют значительную долю населения (за счет домини-
рования в местообитаниях гидросерии). Напротив, в южном горном районе их уча-
стие как в фауне, так и в населении резко сокращается, что сближает комплекс
полужесткокрылых этого района с лесотундровыми, для которых характерно незначи-
тельное участие в их составе арктических видов (Матис, 1986; Ольшванг, 1992; Хру-
лева, Винокуров, 2009).
Таким образом, в зависимости от конкретных местных (мезоклиматических, орогра-
фических) условий комплексы клопов изученной территории имеют аналоги в ланд-
шафтах, занимающих различные позиции на зонально-поясном градиенте субарктиче-
ской Азии. Существенные различия в их видовом составе указывают, что большинство
видов находится здесь за пределами климатического оптимума. Очевидно, что вы-
сокое видовое богатство полужесткокрылых связано с разнообразием природной
среды этой равнинно-горной территории.
ЗАКЛЮЧЕНИЕ
Изученные комплексы полужесткокрылых отличаются от известных в других рай-
онах тундровой зоны как более низкой активностью арктических, так и присутствием
значительного числа бореальных и полизональных видов. Эти черты определяют их
более южный (относительно зонального положения территории) облик. Тем не менее,
в целом эти комплексы хорошо вписываются в общую картину освоения полужестко-
крылыми Северо-Востока Азии, имея ряд общих черт с фаунами как лесотундровых,
так и высокогорных ландшафтов этого региона. Поскольку большинство южных
видов клопов на изученной территории малочисленны, полученные результаты не
противоречат представлению о достаточно низком адаптивном потенциале к засе-
лению этим отрядом тундровой зоны. Вместе с данными по другим районам северной
Чукотки они подтверждают наличие в этом горном регионе длительно существующих
рефугиумов, способствующих сохранению видов, которые в современных тундровых
ландшафтах находятся за пределами своего климатического оптимума.
323
БЛАГОДАРНОСТИ
Мы благодарны директору заповедника «Остров Врангеля» А. Р. Груздеву за по-
мощь в проведении полевых исследований в окрестностях Певека.
ФИНАНСИРОВАНИЕ
Работа О. А. Хрулёвой выполнена при поддержке Российского фонда фундамен-
тальных исследований (грант № 20-04-00165); исследования Н. Н. Винокурова под-
держаны базовым проектом СО РАН № ААААА-А17-117020110058-4 (2017-2020) и
грантом Российского фонда фундаментальных исследований № 18-04-00464 (2018-
2021).
СПИСОК ЛИТЕРАТУРЫ
Алексеев Г. В., Родионов В. Ф., Александров В. И., Иванов Н. Е., Харланенкова Н. Е. 2015. Изменение кли-
мата Арктики при глобальном потеплении. Проблемы Арктики и Антарктики 1 (103): 32-42.
Берман Д. И. 1986. Фауна и население членистоногих в тундростепях о. Врангеля. В кн.: Ф. Б. Чернявский,
И. А. Черешнев (ред.). Биогеография Берингийского сектора Субарктики. Владивосток: ДВНЦ АН
СССР, с. 146-161.
Берман Д. И. 2001. Беспозвоночные животные. В кн.: Д. И. Берман (ред.). Холодные степи Северо-Востока
Азии. Магадан: ИБПС СВНЦ ДВО РАН, с. 96-161.
Берман Д. И., Алфимов А. В., Коротяев Б. А. 2002. Ксерофильные членистоногие в тундростепях урочища
Утесики (Чукотка). Зоологический журнал 81 (4): 444-450. [Berman D. I., Alfimov A. V., Korotyaev
B. A. 2002. Xerophilic arthropods in the tundra-steppe of the Utyosiki Locality (Chukchi Peninsula).
Entomological Review 82 (1): 94-100].
Беpман Д. И., Будаpин А. М., Бухкало С. П. 1984. Фауна и население беспозвоночных животных гоpных
тундp хpебта Большой Анначаг. В кн.: Д. И. Берман (ред.). Почвенный яpус экосистемы гоpных тундp
хpебта Большой Анначаг (веpховье Колымы). Владивосток: ДВНЦ АН СССР, с. 98-134.
Бухкало С. П. 1997. Население наземных беспозвоночных стационара «Контакт». Ч. 3. Видовой состав и
биотопическое распределение беспозвоночных. Препринт. Магадан: СВНЦ ДВО РАН, 56 с.
Винокуров Н. Н. 1979. Насекомые полужесткокрылые (Heteroptera) Якутии. Л.: Наука, 232 с.
Винокуров Н. Н. 2003. Полужесткокрылые (Heteroptera) бассейна р. Мома. В кн.: А. П. Исаев (ред.). Энтомо-
логические исследования в Якутии. Якутск: ИБПК СО РАН, с. 54-58.
Винокуров Н. Н. 2005. Обзор полужесткокрылых рода Chiloxanthus Reut. (Heteroptera, Saldidae) фауны Рос-
сии и сопредельных стран. Энтомологическое обозрение 84 (1): 46-61. [Vinokurov N. N. 2005. A review
of the shore-bug genus Chiloxanthus Reut. (Heteroptera, Saldidae) in the fauna of Russia and adjacent
countries. Entomological Review 85 (2): 118-130].
Винокуров Н. Н. 2008. Состав фауны полужесткокрылых (Heteroptera) Верхоянской горной страны. В кн.:
Н. С. Данилова (ред.). Исследования членистоногих животных в Якутии. Якутск: ИБПК СО РАН,
с. 97-120.
Винокуров Н. Н., Голуб В. Б., Канюкова Е. В., Кержнер И. М., Чернова Г. П. 1988. Отряд Heteroptera
(Hemiptera) - Полужесткокрылые, или клопы. В кн.: П. А. Лер (ред.). Определитель насекомых Даль-
него Востока СССР. Том 2. Равнокрылые и полужесткокрылые. Л.: Наука, с. 727-930.
Винокуров Н. Н., Канюкова Е. В., Голуб В. Б. 2010. Каталог полужесткокрылых насекомых (Heteroptera) ази-
атской части России. Новосибирск: Наука, 317 с.
Винокуров Н. Н., Степанов А. Д. 2003. Полужесткокрылые (Heteroptera) лесотундры и низменных редколе-
сий Северо-Восточной Якутии. Зоологический журнал 82 (6): 744-747.
Городков К. Б. 1984. Типы ареалов насекомых тундры и лесных зон европейской части СССР. В кн.: К. Б. Го-
родков (ред.). Ареалы насекомых европейской части СССР. Л.: Наука, с. 3-20.
Зиновьева А. Н. 2006. К фауне полужесткокрылых (Heteroptera) Большеземельской тундры. Russian
Entomological Journal 15 (2): 127-130.
Зиновьева А. Н. 2013. Фауна полужесткокрылых (Heteroptera) окрестностей озер Харбейской системы (Боль-
шеземельская тундра). Известия Коми научного центра УрО РАН 1 (13): 43-49.
Зиновьева А. Н., Долгин М. М. 2006. Зонально-ландшафтное распределение полужесткокрылых (Heteroptera)
на Европейском Северо-Востоке России. Вестник Северного (Арктического) федерального универси-
тета. Серия Естественные науки 2: 47-52.
324
Кеpжнеp И. М. 1988. Hовые и малоизвестные полужесткокpылые насекомые (Heteroptera) с Дальнего Восто-
ка СССР. Владивосток: Биолого-почвенный институт, 83 с.
Киpиченко А. H. 1960. Hастоящие полужесткоpылые (Heteroptera) восточного сектоpа аpктической Евpазии.
Энтомологическое обозpение 39 (3): 617-628.
Киселев С. В. 1981. Позднекайнозойские жесткокpылые Севеpо-Востока Сибиpи. М.: Hаука, 116 с.
Макарова О. Л., Макаров К. В. 2006. Полужесткокрылые насекомые (Heteroptera) арктического острова
Долгий (Баренцево море). Зоологический журнал 85 (6): 702-711. [Makarova O. L., Makarov K. V. 2006.
Bugs (Heteroptera) of Arctic island Dolgiy, Barents Sea. Entomological Review 86 (4): 423-432.
doi: 10.1134/S0013873806040063].
Марусик Ю. М. 1993. Наземные членистоногие. В кн.: Д. И. Берман (ред.). Экология бассейна р. Амгуэмы.
№ 1. Владивосток: ДВО АН СССР, с. 164-186.
Матис Э. Г. 1986. Hасекомые Азиатской Беpингии: пpинципы и опыт эколого-геосистемного изучения. М.:
Hаука, 312 с.
Ольшванг В. H. 1992. Стpуктуpа и динамика населения насекомых Южного Ямала. Екатеpинбуpг: Hаука,
Уpальское отделение, 104 с.
Саулич А. Х., Мусолин Д. Л. 2007. Времена года: разнообразие сезонных адаптаций и экологических меха-
низмов контроля сезонного развития полужесткокрылых (Heteroptera) в умеренном климате. В кн.:
Стекольников А. А. (ред.). Стратегии адаптаций наземных членистоногих к неблагоприятным услови-
ям среды. Сборник памяти профессора Виктора Петровича Тыщенко (к семидесятилетию со дня
рождения). Труды БиНИИ СПбГУ, 53: 25-106.
Степанов А. Д. 2008. Фаунистический состав полужесткокрылых (Heteroptera) Западной Якутии. В кн.:
А. П. Исаев (ред.). Энтомологические исследования в Якутии. Якутск: ИБПК СО РАН, с. 32-54.
Чернов Ю. И. 1978. Структура животного населения Субарктики. М.: Наука, 166 с.
Чернов Ю. И., Матвеева Н. В. 2002. Ландшафтно-зональное распределение видов в Арктике. Успехи совре-
менной биологии 122 (1): 26-45.
Хpулева О. А. 1987. Беспозвоночные животные. В сб.: В. Е. Соколов, Т. М. Корнеева (ред.). Фауна заповед-
ника «Остpов Вpангеля» (Аннотиpованные списки видов). М.: ИЭМЭЖ, с. 6-36.
Хрулева О. А. 2007. Своеобразие фауны наземных членистоногих острова Врангеля как отражение особен-
ностей его природной среды. В кн.: А. Р. Груздев (ред.). Природа острова Врангеля: современные ис-
следования. СПб.: Астерион, с. 136-181.
Хрулева О. А. 2014. Комплексы наземных членистоногих острова Врангеля: изменения, отмеченные в усло-
виях потепления климата. В кн.: Г. Г. Матишов, Г. А. Тарасов (ред.). Комплексные исследования при-
роды Шпицбергена и прилегающего шельфа: Материалы Международной научной конференции
(Мурманск, 6-8 ноября 2014). Выпуск 12. М.: ГЕОС, с. 316-322.
Хрулева О. А., Винокуров Н. Н. 2007. Наземные полужесткокрылые (Heteroptera) высокогорий хр. Сун-
тар-Хаята (Восточная Якутия). Зоологический журнал 86 (9): 1057-1072. [Khruleva O. A., Vinokurov
N. N. 2007. Terrestrial bugs (Heteroptera) in high mountains of the Suntar-Khayata Range (Eastern Yakutia).
Entomological Review 87 (9): 1126-1141. doi: 10.1134/S0013873807090047]
Хрулева О. А., Винокуров Н. Н. 2009. Наземные полужесткокрылые (Heteroptera) западного побережья
острова Карагинского (Берингово море). Материалы Х международной научной конференции, посвя-
щенной 300-летию со дня рождения Г. В. Стеллера «Сохранение биоразнообразия Камчатки и приле-
гающих морей». Петропавловск-Камчатский: Камчатпресс, с. 138-141.
Юрцев Б. А., Королева Т. М., Петровский В. В., Полозова Т. Г., Жукова П. Г., Катенин Ф. Е. 2010. Конспект
флоры Чукотской тундры. СПб.: ВВМ, 628 с.
Böcher J. 1971. Preliminary studies on the biology and ecology of Chlamydatus pullus (Reuter) (Heteroptera:
Miridae) in Greenland. Meddelelser om Grønland 191 (3): 1-29.
Böcher J., Nachman G. 2011. Coexistence of bisexual and unisexual populations of Nysius groenlandicus in the
Zackenberg Valley, Northeast Greenland. Entomologia Experimentalis et Applicata 140: 196-206.
doi: 10.1111/j.1570-7458.2011.01153.x
Danks Н. V. 1981. Arctic Arthropods. A review of Systematics and Ecology with Particular Reference to the North
American Fauna. Ottawa: Entomological Society of Canada, 605 р.
Scudder G. G. E. 1997. True bugs (Heteroptera) of the Yukon. In: H. V. Danks, J. A. Downes (eds). Insects of the
Yukon. Ottawa: Biological Survey of Canada (Terrestrial arthropods), p. 241-336.
Stekolshchikov A. V., Khruleva O. A. 2015. Contributions to the aphid fauna (Hemiptera, Aphidoidea) of Chukotka
Autonomous Okrug with descriptions of five new species. Zootaxa 4044 (1): 1-44. doi: 10.11646/
zootaxa.4044.1.1
Vinokurov N. N., Khruleva O. A. 2021. Bugs (Heteroptera) of treeless areas of Chukotka (Russia). Ecologica
Montenegrina 41: 15-34. doi: 10.37828/em.2021.41.4
Walker D. A., Raynolds M. K., Daniëls F. J. A., Einarsson E., Elvebakk A., Gould W. A., Katenin A. E., Kholod S. S.,
Markon C. J., Melnikov E. S., Moskalenko N. G., Talbot S. S., Yurtsev B. A. 2005. The Circumpolar Arctic
vegetation map. Journal of Vegetation Science 16: 267-282. doi: 10.1111/j.1654-1103.2005.tb02365.x
325
COMPOSITION OF THE FAUNA AND PATTERN OF BIOTOPIC DISTRIBUTION
OF BUGS (HETEROPTERA) IN THE VICINITY OF PEVEK
(CHUKOTKA AUTONOMOUS OKRUG)
O. A. Khruleva, N. N. Vinokurov
Key words: bugs, Northeastern Asia, Chukotka, zonal and mountain tundra, Arctic species,
fauna, сhorology, biotopic distribution.
SUMMARY
Heteroptera is an insect order with a sharp decline in species richness north of the forest boundary.
The article presents the results of a study of this group in the north of Chaunskiy District (Western
Chukotka, the subzone of typical tundra). The collection was carried out in 2011 in three landscape
areas. A total of 32 species of bugs from 8 families were collected. More than half of the entire fauna
is made up of widespread multizonal and boreal species of bugs. Among the most widespread species,
xerophilic bugs prevailed, including those dominant in the cryophytic steppes of Northeast Asia. With
the exception of the cryoxerophilic Nysius ericae groenlandicus, cryophiles (including the Arctic
species Chiloxanthus arcticus, Ch. stellatus stellatus, Calacanthia trybomi, Orthotylus artemisiae)
had a high abundance only in some biotopes. The composition of dominants was most constant in
habitats with high percentage of wormwoods (zoogenic meadows, ruts, roadsides), where Europiella
artemisiae and N. e. groenlandicus always were present. A stable composition of bug complexes (with
Chlamydatus pullus, N. e. groenlandicus, Сoriomerus scabricornis, Antheminia eurynota remota) was
also found on the southern hill slopes. In other types of habitats, the species composition of bugs was
varying. In areas with different landscape and climatic conditions, the Heteroptera complexes differed
in their taxonomic, latitudinal, and ecological composition. The greatest participation of Arctic bug
species was found in humid and coastal habitats of the lowland part of the investigated area. The
absolute predominance of xerophilic elements (including tundra and steppe species) with an almost
complete absence of hygrophilic (and, accordingly, the extreme poverty of the bug assemblages of
humid habitats) determined the similarity of the bug complex of the northern (driest) part of the ridge to
those in the highlands of the southern Verkhoyansk Area. In the southern (more humid with more water
bodies) part of the ridge, the decrease in the participation of Arctic species and the increase of that of
the southern ones determined the similarity of the fauna of this area to those of the forest-tundra. It is
assumed that the high species richness and uniqueness of the Heteroptera assemblages in the study area
is associated with their successive development in the conditions of the mountainous landscape and
climatic diversity. The data obtained significantly supplemented the list of bugs inhabiting the Asian
tundra, but did not change the general concept of the limited possibilities for the development of this
insect group in the tundra zone.
326