ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 100, 2, 2021
УДК 595.763.1
ВОДОЛЮБЫ ПОДРОДА LUMETUS ZAITZEV (COLEOPTERA,
HYDROPHILIDAE: ENOCHRUS THOMSON) ФАУНЫ РОССИИ
И СОПРЕДЕЛЬНЫХ СТРАН
© 2021 г. С. В. Литовкин, 1* А. С. Сажнев, 2, 3** А. А. Прокин 2, 4***
1 Русское энтомологическое общество
Самара, 443016 Россия
*e-mail: sats.lit@gmail.com (автор, ответственный за переписку)
2 Институт биологии внутренних вод им. И. Д. Папанина РАН
пос. Борок, Ярославская обл., 152742 Россия;
3 Институт проблем экологии и эволюции им. А. Н. Северцова РАН
Ленинский просп., 33, Москва, 119071 Россия
**e-mail: sazh@list.ru
4 Воронежский государственный университет
Университетская пл., 1, Воронеж, 394036 Россия
***e-mail: prokina@mail.ru
Поступила в редакцию 3.02.2020 г.
После доработки 17.02.2021 г.
Принята к публикации 17.02.2021 г.
В фауне России и сопредельных стран зарегистрировано 11 видов подрода Lumetus
Zaitzev, 1908 рода Enochrus Thomson, 1859. Enochrus ater (Kuwert, 1888) впервые указан из
Украины и Киргизии; E. bicolor (Fabricius, 1792) - из Киргизии; E. fuscipennis (Thomson,
1884) - из Грузии; E. segmentinotatus (Kuwert, 1888) - из Украины. Обсуждаются строение
эдеагуса и методика изготовления препаратов гениталий. Приведены аннотированный список и
определительная таблица видов с оригинальными фотографиями и рисунками, описана измен-
чивость видов. Доказана принадлежность E. puetzi Hebauer, 1995 к подроду Lumetus. Установле-
на новая синонимия: Enochrus ochropterus (Marsham, 1802) = Philhydrus ferrugineus Motschulsky,
1849, syn. n., обозначены лектотип и паралектотип этого вида. Обсуждается таксономический
статус E. asiaticus (Kuwert, 1893), E. calabricus (Ferro, 1976) и E. sahlbergi (Fauvel, 1887).
Ключевые слова: Hydrophilidae, Enochrus, Россия, Палеарктика, фауна, жуки-водолюбы, опре-
делитель, новые находки.
DOI: 10.31857/S036714452102009X
Род Enochrus Thomson, 1859 относится к подсем. Enochrinae Short et Fikáček, 2013
сем. Hydrophilidae Latreille, 1802, распространен на всех материках, кроме Антарк-
тиды, и включает более 200 видов, которые на стадиях личинки и имаго обитают
в разнообразных водных объектах (Short, Fikáček, 2013). В Палеарктике зарегистри-
рован 61 вид из 5 подродов; 25 видов относятся к подроду Lumetus Zaitzev, 1908
(Fikáček et al., 2015; данные авторов). До сих пор не было проведено ревизии подрода
390
для фауны бывшего СССР, хотя последняя определительная таблица для имаго (Ки-
рейчук, Шатровский, 2001) включала все известные с территории страны виды, за ис-
ключением трех из группы bicolor, ревизованной Ш. Шёдлем (Schödl, 1998). Большая
изменчивость окраски и малое количество диагностических признаков в строении
эдеагуса не позволяли надежно различать многие виды, включая массовые и широко
распространенные.
МАТЕРИАЛ И МЕТОДИКА
Всего изучено более 1660 экз. имаго с территории бывшего СССР и около 100 экз. из других
стран: Швеции, Финляндии, Египта, Израиля, Турции, Ирана и Монголии, в том числе
E. salomonis (Sahlberg, 1900) из Израиля и Ирана и E. politus (Küster, 1849) из Испании и Италии.
В статье приводится весь изученный материал, включая ранее нами опубликованный, из
следующих коллекций.
ЗИН - Зоологический институт РАН, С.-Петербург, Россия;
ЗММУ - Зоологический музей Московского государственного университета им. М. В. Ломо-
носова, Москва, Россия;
ИБВВ - Институт биологии внутренних вод им. И. Д. Папанина РАН, пос. Борок, Ярославская
обл., Россия;
КСЛ - коллекция С. В. Литовкина, г. Самара, Россия;
КСР - коллекция С. К. Рындевича, г. Барановичи, Республика Беларусь.
В материале по наиболее интересным и новым для регионов видов приводятся подробные
данные этикеток, для более обычных видов указываются лишь административные регионы. Раз-
меры тела жуков приводятся по результатам собственных измерений, в скобках указываются
литературные данные. Длина жуков измерялась от переднего края головы до вершин надкрылий.
Основная часть фотографий получена с помощью камеры Nikon D3200, оснащенной фото-
объективом Helios-44-2 или микроскопными объективами ЛОМО ПЛАН 3.5 × 0.10 и ЛОМО
ПЛАН 9.0 × 0.20, а также камеры Canon PowerShot A640, соединенной с биологическим микро-
скопом ЛОМО Биолам Р-17. Фотографии эдеагуса в глицериновой среде сделаны в проходящем
свете. Обработка снимков проведена в программах Helicon Focus 6.3 и Adobe Photoshop CS3.
Рисунки эдеагуса выполнены на основе фотографий в программе Inkscape 0.92 с параллельным
просмотром препарата под микроскопом.
Рекомендации к определению представителей рода Enochrus и подрода Lumetus
Внешние морфологические признаки имаго изучаются на сухих экземплярах с воз-
можностью изменения направления и жесткости освещения. При монтировке хотя бы
у части жуков рекомендуется оставлять нижнюю сторону доступной для обзора. Для
определения по гениталиям самцов требуется изучение эдеагуса в жидкой среде без
обработки щелочью или молочной кислотой при увеличениях 40-60×. Применение
щелочи или молочной кислоты часто ведет к раздвиганию парамер и, как следствие,
к искажению их формы. Для быстрого просветления предварительно размоченный
в горячей воде или в спирте эдеагус помещают в каплю глицерина на предметное
стекло. Стекло нагревают при температуре около 70-100 ºC в течение 15 мин до заме-
щения исходной жидкости глицерином и вытеснения остатков воздуха. Остающиеся
иногда внутри небольшие пузырьки воздуха не мешают определению, но могут быть
391
удалены повторными нагреваниями или сами растворяются в глицерине в течение не-
скольких дней (если эдеагус остается в капле).
Самцы и самки видов подрода Lumetus узнаются благодаря хорошо выраженному
диморфизму в строении коготков и окраске головы. У самцов коготки резко изогнутые,
с увеличенным зубцом или лопастью в основании (рис. 1, 10); на передних лапках
один коготок может быть немного более толстым и немного более изогнутым, чем
другой; тарсомеры II-IV передних лапок удлиненные. У самок коготки серповидно
изогнутые, с небольшим зубчиком в основании, одинаковые по форме (рис. 1, 11, 12).
У самцов большинства видов верхняя губа светлая (желтая), у самок - темная (черная
или бурая). Темный рисунок на голове самок более развит, чем у самцов.
Если экземпляры с аберрантной окраской головы трудно с уверенностью опреде-
лить с помощью таблицы, необходимо сравнить их с видами, у которых голова более
темная. Нередко на покровах сухих экземпляров места с отслоившейся от кутикулы
гиподермой выглядят как темные пятна, которые можно ошибочно принять за особен-
ности окраски.
Строение и диагностические признаки эдеагуса видов подрода Lumetus
В публикациях по систематике рода Enochrus строение эдеагуса рассматривается
с разной степенью подробности. Общепринятых терминов для обозначения частей
пениса (средней доли эдеагуса) и парамер Enochrus нет. Названия «корона» (corona) и
«базальные апофизы» (basal apophyses) применяются в англоязычной литературе для
подобных (вероятно, гомологичных) структур пениса в других группах водолюбов.
Некоторые из названных далее структур пениса и парамер Lumetus ранее никем в ка-
честве самостоятельных не выделялись.
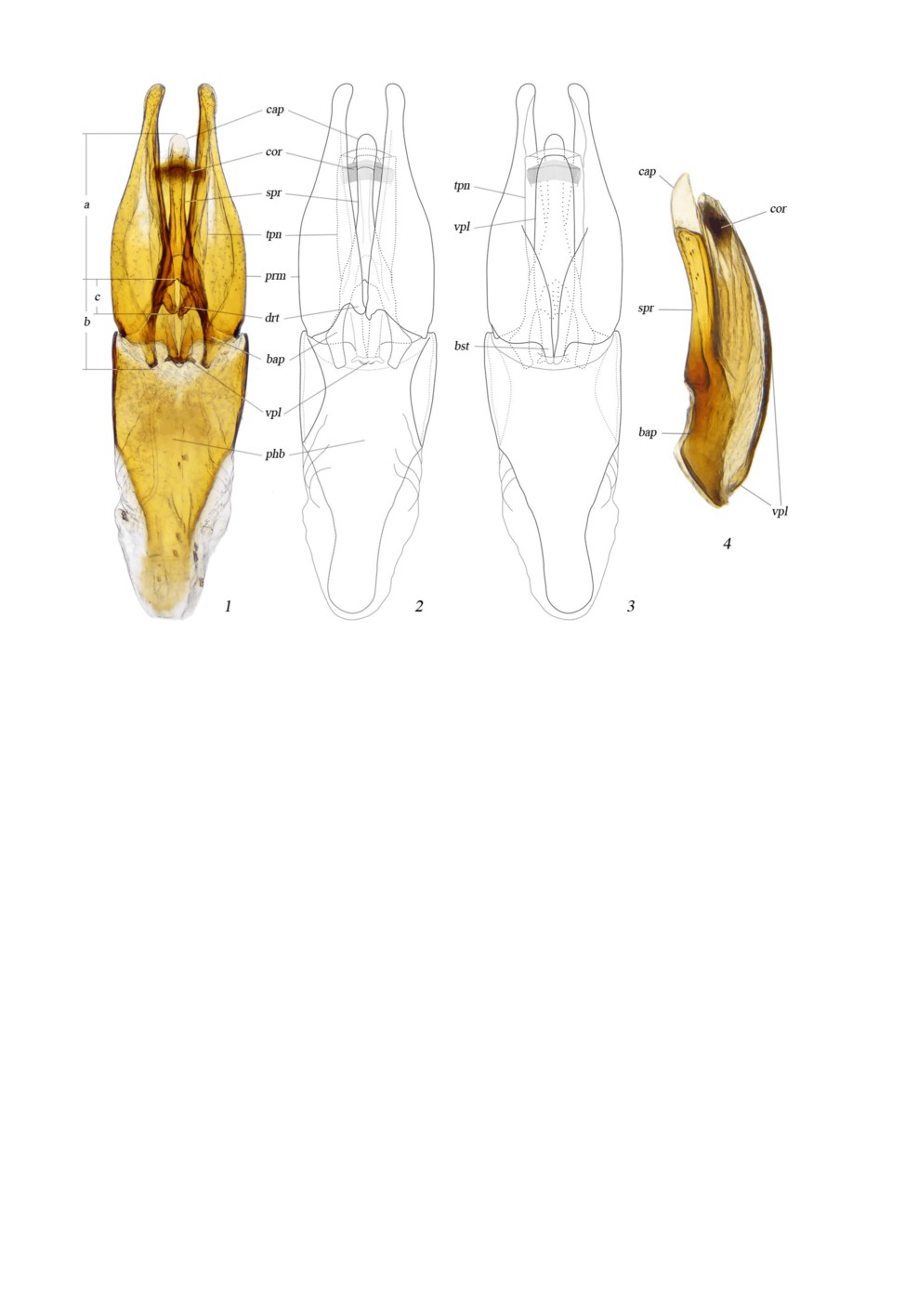
Расположение главных элементов эдеагуса показано на рис. 6, 1-4. Парамеры (prm)
посредством мембран и пары латеральных мыщелков подвижно сочленяются с фалло-
базой (phb) и удерживают пенис. Между собой парамеры удерживаются парой дор-
сальных зубцов (drt) и парой базальных зубцов (bst) вентрально. Дорсальные зубцы
обхватывают также базальные апофизы пениса и соединяются с ними посредством
связок. Базальные апофизы (bap) образуют арку в основании пениса, в которую входит
семявыносящий канал (ductus ejaculatorius). К ним крепятся некоторые мышцы па-
рамер. Шпора пениса (spr) отходит непосредственно от места слияния апофиз и почти
вплотную прилегает к трубке пениса, но не соединяется с ней. Вершина шпоры без
каких-либо наружных швов переходит в прозрачный наконечник (cap), отделенный от
нее резкой внутренней границей. Трубка пениса (tpn) сжата дорсовентрально, слива-
ется с базальными апофизами с одной стороны и укреплена вентральной пластинкой с
другой, половое отверстие открывается апикально. Склеротизация стенок трубки раз-
личима в световой микроскоп по крайней мере до основания шпоры пениса. Дор-
сальная стенка трубки с продольным медиальным желобком. Вентральная пластинка
(vpl) в форме узкого склерита тянется вдоль всего пениса, в основании подвижно со-
членяется с парой базальных зубцов парамер, а на вершине свободно оканчивается
дистальнее короны. Вершина вентральной пластинки частично пережимает половое
отверстие. Корона (cor) - внутренняя структура в субапикальной части трубки пениса.
Она окружает гонопор и образована множеством мелких, параллельно расположенных
склеритов, сжата дорсовентрально, как и вся расширенная дистальная часть семя-
выносящего канала.
392
Рис. 1. Enochrus spp.
1, 6 - E. melanocephalus (Olivier); 2 - E. affinis (Thunberg); 3 - E. quadripunctatus (Herbst);
4 - E. puetzi Hebauer; 5 - E. simulans (Sharp); 7, 10, 11 - E. fuscipennis (Thomson);
8 - E. ochropterus (Marsham); 9, 12 - E. turanicus Schödl.
1-4 - 5-й вентрит брюшка и его вершина (на врезке); 5-9 - пунктировка надкрылий (белыми стрелками
показаны видимые на фотографии точечные ряды, черными стрелками на врезках - точки
с трихоботриями,); 10-12 - коготки передней правой лапки, вид спереди (10 - самец;
11, 12 - самки). Каждая группа изображений представлена в едином масштабе.
Для диагностики могут использоваться форма парамер, соотношение длин трубки
и шпоры пениса с наконечником, соотношение длин дорсальных зубцов парамер
и базальных апофиз, а также форма вершины шпоры пениса. Длина трубки пениса
легко определяется по расположению короны относительно шпоры. При сравнимых
длинах обеих ветвей пениса учитывается взаимное расположение короны и границы,
393
разделяющей шпору и наконечник. Строение наконечника не используется в качестве
диагностического признака, так как эта часть нередко подвергается естественной де-
формации.
РЕЗУЛЬТАТЫ
В фауне России нами отмечено 10 видов из подрода Lumetus, еще один вид, Enochrus
turanicus Schödl, 1998, отмечен из Туркмении.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ РОДА ENOCHRUS РОССИИ
И СОПРЕДЕЛЬНЫХ СТРАН
1. 5-й (последний) вентрит брюшка с более или менее глубокой апикальной вырезкой,
несущей грубые щетинки (рис. 1, 1, 2)
2.
– 5-й вентрит брюшка цельнокрайний, без апикальной вырезки и грубых щетинок
(рис. 1, 3, 4)
3.
2. 2-й (1-й видимый дорсально) и 3-й максиллярные пальпомеры короткие, 4-й (по-
следний) пальпомер сравним по длине с 3-м (рис. 4, 1)
........................................................................................... Enochrus Thomson, 1859.
В Палеарктике один вид - E. melanocephalus (Olivier, 1793) (рис. 1, 1, 6; 2, 1; 4, 1;
7, 1): надкрылья, особенно в задней трети, со слабыми, но ясно различимыми точеч-
ными рядами; точки с трихоботриями на надкрыльях редкие и рассеянные; передне-
спинка без пятна; 4-й максиллярный пальпомер затемнен перед вершиной; ноги
темные; половой диморфизм в строении коготков слабо выражен, в окраске головы -
отсутствует; длина тела 4.5-5.5 мм.
- 2-й и 3-й максиллярные пальпомеры длинные, 4-й пальпомер заметно короче 3-го
(рис. 4, 2)
Methydrus Rey, 1885.
В Палеарктике 29 видов; бороздки на надкрыльях, кроме неполных пришовных, от-
сутствуют; E. affinis (Thunberg, 1794) (рис. 1, 2); E. nigritus (Sharp, 1873) (рис. 2, 2;
4, 2; 7, 2).
3. Надкрылья с углубленными в бороздки точечными рядами и грубыми точками
с трихоботриями в нечетных междурядьях (рис. 1, 5)
........................................................................... Holcophilydrus Kniž, 1912 (partim).
Enochrus simulans (Sharp, 1873) (рис. 1, 5; 2, 3; 4, 3; 7, 3) - наиболее широко распро-
страненный и известный вид из четырех, отмеченных в Палеарктике; половой димор-
физм в строении коготков и в окраске головы хорошо выражен; длина тела 4.5-5.2 мм.
- Надкрылья только с неполными пришовными бороздками, остальные точечные
ряды могут быть лишь едва заметными (их расположение нередко маркировано
рядами просвечивающих изнутри темных пятен), с более или менее раз-
личимыми рядами точек с трихоботриями на месте нечетных междурядий
(рис. 1, 7-9)
Lumetus Zaitzev, 1908.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПОДРОДА LUMETUS ФАУНЫ РОССИИ
И СОПРЕДЕЛЬНЫХ СТРАН
1. Боковые края надкрылий распластанные (рис. 1, 9). Пунктировка надкрылий и пе-
реднеспинки, за исключением точек с трихоботриями, поверхностная. Коготки
394
(у изученной нами самки) сильно удлиненные. Длина тела (4.7)-4.9-(5.2) мм
................................................................... E. turanicus Schödl, 1998 (рис. 1, 9, 12).
- Скаты надкрылий плавно скругленные (как на рис. 1, 6). Пунктировка надкрылий
и переднеспинки относительно грубая. Коготки относительно короткие
2.
2. Голова от частично бурой до полностью желтой (рис. 3, 1-4)
3.
– Как минимум лоб зачернен (рис. 3, 5-12)
6.
3. Голова и переднеспинка обычно с едва заметным буроватым рисунком или без
него. Максиллярные щупики полностью светлые. Шпора пениса с наконечником
выступают за корону более чем на 2/5 их общей длины. Вершины дорсальных
зубцов парамер достигают примерно 1/3 длины базальных апофиз
4.
– Лоб обычно бурый, наличник у самцов затемнен в развилке фронто-клипеального
шва, у самок - с размытым бурым пятном. 2-й и/или 4-й максиллярные пальпо-
меры иногда частично затемненные. Пятно на переднеспинке от едва заметного
до темного, у E. ater сильно размытое. Шпора пениса с наконечником высту-
пают за корону не более чем на 1/3 их общей длины
5.
4. Тело широкоовальное. Задние бедра с размытой продольной желтой полосой, реже
полностью черные. Наличник спереди обычно вогнут в форме широкой
трапеции. Внутренний край парамер без резких изгибов. Вершина шпоры
пениса равномерно отогнута дорсально. Длина тела 5.3-6.7 мм
......................... E. bicolor (Fabricius, 1792) (рис. 2, 5; 3, 1; 4, 4, 14, 21; 7, 4, 9, 11).
- Тело овальное или узкоовальное. Задние бедра с широкой контрастной продольной
желтой полосой. Наличник спереди обычно вогнут в форме узкой трапеции.
Внутренний край парамер с резким ступенчатым изгибом, различимым в дор-
сальном и дорсо-латеральном аспектах. Вершина шпоры пениса резко оттянута
дорсально. Длина тела 5.5-6.4 мм
.......... E. segmentinotatus (Kuwert, 1888) (рис. 2, 6; 3, 2; 4, 5, 15, 22; 7, 5, 10, 12).
5. Переднегрудь спереди с характерным крючковидным отростком. Бедра со светлой
полосой или полностью темно-бурые. Голени буровато-желтые, немного темнее
максиллярных щупиков. Вершины дорсальных зубцов парамер достигают почти
середины базальных апофиз. Наконечник шпоры отграничен немного дис-
тальнее короны. Мельче, длина тела 4.5-5.2 мм
.................................... E. hamifer (Ganglbauer, 1901) (рис. 2, 7; 3, 3; 4, 6, 20; 7, 6).
- Переднегрудь спереди без крючковидного отростка. Бедра, кроме красноватых
вершин, темно-бурые. Голени одного цвета с бедрами, отчетливо темнее максил-
лярных щупиков и лапок. Вершины дорсальных зубцов парамер достигают при-
мерно 1/4 длины базальных апофиз. Наконечник шпоры отграничен на уровне
короны. Крупнее, длина тела 4.5-6.2 мм
....................................................... E. ater (Kuwert, 1888) (рис. 2, 8; 3, 4; 4, 7; 7, 7).
6. Точки с трихоботриями не выделяются на фоне остальной пунктировки надкрылий.
Наличник у самцов с треугольным пятном, у самок - с широким пятном, его
фоновая окраска у обоих полов от светлой до темной. Пятно на переднеспинке
395
Рис. 2. Enochrus spp., общий вид имаго сверху.
1 - E. melanocephalus (Olivier), самка; 2 - E. nigritus (Sharp), самец; 3 - E. simulans (Sharp), самец;
4 - E. turanicus Schödl, самка; 5 - E. bicolor (F.), самец; 6 - E. segmentinotatus (Kuwert), самец;
7 - E. hamifer (Ganglbauer), самка; 8 - E. ater (Kuwert), самец; 9 - E. ochropterus (Marsham), самка;
10 - E. quadripunctatus (Herbst), самец; 11 - E. sahlbergi (Fauvel), самка (Ekerö); 12 - E. halophilus (Bedel),
самец; 13 - E. fuscipennis (Thomson), самец; 14 - E. testaceus (F.), самец; 15 - E. puetzi Hebauer, самец.
Масштабная линейка - 2 мм.
396
Рис. 2. (продолжение)
часто очень широкое. Тело широкоовальное. Шпора пениса с наконечником
примерно на половину их общей длины выступают за корону. Длина тела 4.7-
5.2 мм
E. ochropterus (Marsham, 1802) (рис. 1, 8; 2, 9; 3, 5; 7, 8).
– Ряды из точек с трихоботриями на месте нечетных междурядий хорошо различимы
(как на рис. 1, 7)
7.
7. 2-й максиллярный пальпомер светлый. Тело овальное. Шпора пениса с наконеч-
ником выступают за корону не более чем на 1/3 их общей длины, вершина
шпоры равномерно отогнута дорсально (как на рис. 7, 23). Вершины дорсальных
зубцов парамер достигают примерно 1/3 длины базальных апофиз
8.
– 2-й максиллярный пальпомер в основной половине и обычно 4-й перед вершиной
затемнены. Тело очень широко овальное. Шпора пениса с наконечником высту-
пают за корону не менее чем на 1/3 их общей длины. Вершина шпоры пениса
резко оттянута дорсально (как на рис. 7, 24), наконечник сжат с боков. Вершины
397
Рис. 3. Enochrus (Lumetus) spp., половой диморфизм окраски головы (cлева самец,
справа - самка).
1 - E. bicolor (F.), 2 - E. segmentinotatus (Kuwert), 3 - E. hamifer (Ganglbauer), 4 - E. ater (Kuwert),
5 - E. ochropterus (Marsham), 6 - E. quadripunctatus (Herbst), 7 - E. sahlbergi (Fauvel) (Ekerö),
8 - E. halophilus (Bedel), 9 - E. fuscipennis (Thomson), 10 - E. fuscipennis (самец аберрации
«calabricus» и самка, собранная вместе с ним), 11 - E. testaceus (F.), 12 - E. puetzi Hebauer.
Все изображения представлены в едином масштабе.
дорсальных зубцов парамер достигают 1/3 или 2/5 длины базальных апофиз
10.
8. Пятно переднеспинки расположено внутри четырехугольника из черных точек.
Наличник V-образно вогнут спереди, у самцов с треугольным пятном, значи-
тельно не доходящим до его переднего края, у самок - с широким резким
пятном. Верхняя губа у самцов светлая. 4-й максиллярный пальпомер отчетливо
398
Рис. 4. Enochrus spp.
1 - E. melanocephalus (Olivier); 2 - E. nigritus (Sharp); 3 - E. simulans (Sharp); 4, 14, 21 - E. bicolor (F.);
5, 15, 22 - E. segmentinotatus (Kuwert); 6, 20 - E. hamifer (Ganglbauer); 7 - E. ater (Kuwert);
8 - E. ochropterus (Marsham); 9, 16, 24 - E. quadripunctatus (Herbst); 10, 18, 25 - E. halophilus (Bedel);
11, 19, 23 - E. fuscipennis (Thomson); 12 - E. testaceus (F.); 13 - E. puetzi Hebauer; 17 - E. sahlbergi (Fauvel).
1-13 - максиллярные щупики дорсально; 14-19 - форма переднего края наличника (14, 15, 18 - самцы;
16, 17, 19 - самки); 20 - отросток переднегруди латеро-вентрально; 21, 22 - окраска
и гидрофобное опушение задних бедер самцов вентрально (стрелками показана граница опушения);
23-25 - отросток среднегруди справа. Каждая группа изображений представлена в едином масштабе.
затемнен перед вершиной. Передний край отростка среднегруди направлен
назад или вниз. Наконечник шпоры отграничен, как правило, на уровне короны,
редко дистальнее. Длина тела 4.0-5.5 мм
................... E. quadripunctatus (Herbst, 1797) (рис. 2, 10; 3, 6; 4, 9, 16, 24; 7, 13).
- Пятно переднеспинки покрывает прямоугольник из черных точек, у E. fuscipennis
нередко значительно шире его; пятно более или менее темное. Признаки и сино-
нимику E. sahlbergi см. в тексте
9.
399
9. Наличник V-образно вогнут спереди. Окраска наличника и верхней губы как у
E. quadripunctatus. Максиллярные щупики полностью светлые. Передний край
отростка среднегруди направлен назад. Наконечник шпоры отграничен значи-
тельно проксимальнее короны, едва выступает за вершину трубки пениса. Длина
тела 5.9-6.1 мм
....................... E. halophilus (Bedel, 1879) (рис. 2, 12; 3, 8; 4, 10, 18, 25; 7, 14, 20).
- Наличник дуговидно вогнут спереди, у самцов с широким треугольным пятном, до-
ходящим до его переднего края или широко с ним соприкасающимся, у самок -
с широким резким пятном. Верхняя губа у самцов темная до черной. 4-й мак-
силлярный пальпомер от отчетливо затемненного перед вершиной до почти
светлого. Передний край отростка среднегруди направлен вниз или вперед.
Наконечник шпоры отграничен на уровне короны или дистальнее. Длина тела
4.6-5.7 мм
................ E. fuscipennis (Thomson, 1884) (рис. 2, 13; 3, 9, 10; 4, 11, 19, 23; 7, 15).
10. Крупнее, длина тела 5.4-6.5 мм. Наличник у самцов затемнен в развилке фронто-
клипеального шва, у самок - с размытым пятном. Переднеспинка, как правило,
без отчетливого пятна. Вершины парамер шире, скругленные снаружи. Вер-
шины дорсальных зубцов парамер достигают примерно 2/5 длины базальных
апофиз
E. testaceus (Fabricius, 1801) (рис. 2, 14; 3, 11; 4, 12; 7, 16, 21, 24).
– Мельче, длина тела 5.2(-5.6) мм. Наличник у самцов с треугольным пятном,
у самок - с размытым пятном. Переднеспинка с размытым темным пятном, как
минимум, покрывающим прямоугольник из черных точек. Вершины парамер
суженные, слабо булавовидные. Вершины дорсальных зубцов парамер до-
стигают примерно 1/3 длины базальных апофиз
....................................... E. puetzi Hebauer, 1995 (рис. 2, 15; 3, 12; 4, 13; 7, 17, 22).
Аннотированный список изученных видов подрода Lumetus
Enochrus ater (Kuwert, 1888) (рис. 2, 8; 3, 4; 4, 7; 7, 7).
Мат ер иа л. Украина. Николаевская обл.: Кинбургская коса, тростниковое болото в степи,
1.V.2007 (В. Г. Дядичко), 1 ♂ (ИБВВ); там же, пресноводный водоем в степи, 30.VI.2007 (В. Г. Дя-
дичко), 1 ♀ (ИБВВ). Запорожская обл.: Бильмакский р-н, пос. Камыш-Заря, на свет, VII.2012
(В. В. Шапоринский), 2 ♂, 1 ♀ (КСЛ). Россия. Крым: Симферопольский р-н, окр. балки Тубай,
45°03ʹ N, 34°10ʹ E, на свет, 8-11.VII.2015, 28.VI.2016, 7.VII.2016 (В. В. Шапоринский), 3 ♂, 3 ♀
(КСЛ). Калмыкия: с. Приютное, пруд, 4.V.2011 (М. Я. и А. О. Беньковские), 1 ♀ (ЗММУ). Астра-
ханская обл.: Богдинско-Баскунчакский заповедник, 7 км СЗ горы Большое Богдо, 4.V.2006
(А. В. Ковалёв), 1 ♀ (ИБВВ); Красноярский р-н, с. Досанг, берег р. Ахтуба, на свет, 1-8.V.2011
(И. А. Забалуев), 1 ♀ (КСЛ). Ставропольский край: pg. Praskoveja [44°44ʹ N, 44°12ʹ E], Stavropol,
1.VIII.1904 (L. Banjkovski), 1 экз. (ЗИН). Дагестан: Петровск-порт [Махачкала], 10.V.1926,
1.VIII.1926 (Рябов), 1 ♂, 1 ♀ (ЗИН). Азербайджан. ?Масаллинский р-н: Кумбаши [р. Кумбаши =
Кумбашинка] (морцё [морцо - солоноватое озерко]), Ленкоран[ский] у[езд]., 23.IV.1909 (А. Н.
Кириченко), 2 экз. (подписаны Ф. А. Зайцевым как E. (Lumetus) kiritshenkoi sp. n.) (ЗИН). «Мони-
дыя, Ленкоранск[ий]. у[езд]», (без даты) (А. Н. Кириченко), 1 экз. (подписан Ф. А. Зайцевым как
E. (Lumetus) kiritshenkoi sp. n.) (ЗИН); «Гелядара, Ленк[оранский]. у[езд].», 10.VII.1909 (А. Н. Ки-
риченко), 3 экз. (подписаны Ф. А. Зайцевым как E. (Lumetus) kiritshenkoi sp. n.) (ЗИН); Baku,
Sumgait, 8.V.1927 (сборщик неизвестен), коллекция Ф. А. Зайцева, 1 экз. (ЗИН). Агдашский р-н:
Geok-Tapa, Caucasus (без даты) (A. Schelkownikow), 1 ♂, 2 ♀, 6 экз. (ЗИН); Aresch, Caucasus (без
даты) (A. Schelkownikow), 2 ♀, 1 экз. (ЗИН); Geok-Tapa, Aresch, VII.1908 (A. Schelkownikow), 1 ♀
400
(ЗИН). Сабирабадский р-н: Alexandrovka [~ 39°56ʹ30ʺ N, 48°42ʹ30ʺ E], Steppa Mugan, Mus. Caucas,
No. 101-13 (без даты) (N. Volčanetski), 4 экз. (ЗИН). Ленкоранский р-н: Гелякерань [Шихакаран],
Ленкоранск[ий]. у[езд]., 6.V.1909 (А. Н. Кириченко), 1 экз. (подписан Ф. А. Зайцевым как E. (Lu-
metus) kiritshenkoi sp. n.) (ЗИН); пос. Нариманабад, С Ленкорани, заболоченная низина, 24.V.1983
(А. Г. Шатровский), 8 экз. (ЗИН). Казахстан. Кызылординская обл.: Джулек, Оренбург-Ташкент-
ская железная дорога, Сырдарья, 14.VII.1910 (Кожанчиков), 1 экз. (ЗИН); 4.3 км ЮВ пос. Тар-
тогай, тугай близ р. Сырдарья,
44°24ʹ37ʺ N,
66°16ʹ30ʺ E,
145 м, на свет,
13-
14.VI.2015, 23-24.VII.2017 (С. В. Литовкин), 2 ♂, 5 ♀ (КСЛ). Туркестанская обл.: 50 km W of
Arys, Bayrkum, Syr-Darya River, 7.VII.1999 (M. L. Danilevskiy), 2 ♂ (КСР); 60 km NW of Zhuantobe,
4 km N of Chu River, stream, 25.V.2001 (E. Mongin), 2 ♂ (КСР); северный край оз. Кызылколь,
43°46ʹ34ʺ N, 69°30ʹ36ʺ E, 335 м, солончак, на свет, 5.V.2015, 4.VI.2015 (С. В. Литовкин), 2 ♂, 1 ♀
(КСЛ); там же, 43°45ʹ39ʺ N, 69°28ʹ47ʺ E, 327 м, в воде, 31.V-1.VI.2017 (С. В. Литовкин), 1 ♂
(КСЛ); 7.4 км СВ пос. Балтаколь, заводи р. Сырдарья, 43°09ʹ31ʺ N, 67°50ʹ51ʺ E, 178 м, 7.VI.2016
(С. В. Литовкин), 2 ♂ (КСЛ); там же, тугай, 43°09ʹ24ʺ N, 67°51ʹ00ʺ E, 180 м, на свет, 9-10.VI.2016
(С. В. Литовкин), 2 ♂ (КСЛ); 9.2 км СВ пос. Балтаколь, р. Сырдарья и ее заводи, 43°10ʹ19ʺ N,
67°51ʹ48ʺ E, 170 м, 10.VI.2016 (С. В. Литовкин), 1 ♂, 1 ♀ (КСЛ); 3 км З пос. Балтаколь, граница
пустыни Кызылкум, 43°07ʹ04ʺ N, 67°42ʹ49ʺ E, 180 м, на свет, 13-14.VI.2016 (С. В. Литовкин), 2 ♀
(КСЛ). Жамбылская обл.: Аксуек, на свет, 7-8.VI.1993 (А. С. Тилли), 1 ♂, 1 ♀ (КСЛ); 55 км ССВ
пос. Жайылма, пустыня Мойынкум, 44°17ʹ33ʺ N, 70°09ʹ54ʺ E, 315 м, на свет, 13-14.V.2014
(А. М. Шаповалов), 1 ♂ (КСЛ); там же, 31.V-1.VI.2015 (С. В. Литовкин), 2 ♂ (КСЛ); западный
край оз. Акколь, 43°24ʹ30ʺ N, 70°37ʹ30ʺ E, 405 м, тростниковый берег и на свет, 17-18.V.2015
(С. В. Литовкин), 2 ♂, 1 ♀ (КСЛ). Узбекистан. Бухарская обл.: Зап[адная] Бухара (без даты)
(Нилльберг), 1 ♀ (ЗИН); «Бухара, Зякеш у Кули-Суфиона», 19.V.1925 (И. Соколов), 1 экз. (ЗИН);
Бухара, болото Кули-Суфион [озеро-болото Кулисуфиён], 26.V.1925 (И. Соколов и П. Резвой),
1 экз. (ЗИН); Бухара, болото Углановское, 22.IV.1925 (И. Соколов), из коллекции Ф. А. Зайцева,
1 экз. (ЗИН). Ташкент и Ташкентская обл.: Бектемир (дата и сборщик неизвестны), из коллекции
Ф. А. Зайцева, 1 экз. (ЗИН); окр. Ташкента (дата и сборщик неизвестны), 2 ♂ (ЗИН). Сырдарьин-
ская обл.: ж.-д. ст. Голодная Степь [г. Гулистан], Среднеазиатская железная дорога, 31.III.1903,
8.IV.1903, 1.VI.1903 (Г. Г. Якобсон), 4 экз. (ЗИН). Ферганская обл.: окр. Скобелева [г. Фергана],
«Ак-тюбе», 21.IV.1919 (П. П. Архангельский), 1 экз. (ЗИН). Кашкадарьинская обл.: Камаши,
30.VI.1932 (В. Гуссаковский), 2 экз. (ЗИН). Туркмения. Балканский велаят: Transcaspien,
Sumbar, 1894 (Herz), из коллекции Г. Сиверса, 1 экз. (ЗИН); Моллакара, на свет электрического
фонаря, 7.VIII.1933 (Власов), 2 экз. (ЗИН); Копетдаг, 12 км ЮЗ Кизыл-Арвата [г. Сердар], на
свет, 4.VII.1953 (О. Л. Крыжановский), 1 экз. (ЗИН). Ахалский велаят: Душак, близ г. Теджен,
11-18.IV.1985 (Тарасенкова), 1 ♂ (КСЛ); окр. г. Теджен, 17-24.IV.1985 (А. Тилли и В. Никулин),
1 ♀ (КСЛ); Bakharden, at light, 19.VII.1990 (V. Pillipenko), 1 ♂ (ИБВВ). Киргизия. Джалал-Абад-
ская обл.: 5 км З г. Таш-Кумыр, 41°20ʹ08ʺ N, 72°07ʹ57ʺ E, 775 м, на свет 15-16.VI.2017,
16-17.VII.2017 (С. В. Литовкин), 4 ♂, 4 ♀ (КСЛ); 5 км В пос. Шекафтар, Ю пос. Карын-Кур,
41°13ʹ28ʺ N, 71°23ʹ25ʺ E, 990 м, на свет, 26-27.VI.2017 (С. В. Литовкин), 2 ♂, 4 ♀ (КСЛ); 10 км
СЗ г. Джалалабад, 40°56ʹ00ʺ N, 72°53ʹ20ʺ E, 917 м, холмы, на свет, 15-16.VII.2017 (С. В. Литов-
кин), 1 ♂ (КСЛ); 2 км З пос. Шекафтар, 41°13ʹ03ʺ N, 71°17ʹ38ʺ E, ~ 1140 м, на свет, 2-3.VI.2018
(С. В. Литовкин), 1 ♂ (КСЛ). Ошская обл.: берег Найманского водохранилища, 40°20ʹ31ʺ N,
72°22ʹ22ʺ E, 1200 м, 21-23.VI.2017 (С. В. Литовкин), 1 ♀ (КСЛ). Таджикистан. Хатлонская обл.:
«Бобо-сафид, р-н Пархара, р. Пяндж», 8, 29 и 31.VII.1934 (Е. П. Луппова), 8 экз. (ЗИН); «Ишан,
р-н Пархара, р. Пяндж», 12.VII.1934 (Е. П. Луппова), 1 экз. (ЗИН); «Пархар-Калай-Пучкак»,
19.VIII.1934 (Е. П. Луппова), 1 экз. (ЗИН); Пархар, на рисовом поле, 09.IX.1934 (Е. П. Луппова),
2 экз. (ЗИН); Курган-Тюбе [г. Бохтар], на свет, 9.VII.1939, 19.VII.1939 (О. Л. Крыжановский),
6 экз. (ЗИН). Иран. Iran (без даты) (Bashkevich), 1 ♂ (ЗИН). Провинция Гилян: «Persia l. m Caspii
[Каспийское море], Hassankiadeh [? Куйе-Хасан-Киаде]», 1915, 4.II.1915, 24.II.1915, 10.IV.1915
(B. Iljin), 5 экз. (ЗИН); «Persia l. m Caspii [Каспийское море] Tshemchala [? Чемхале]», 21.V.1915
(B. Iljin), 3 экз. (ЗИН). Провинция Систан и Белуджистан: «Гусейн-абад, Сеистан», 29.V.1896
(Н. А. Зарудный), 1 экз. (ЗИН); «Сеистан, Нэйзар», 21-30.V.1898 (Н. А. Зарудный), 6 экз. (ЗИН);
«Керман, стр.[ана] Касеркенд», 31.V.1898 (Н. А. Зарудный), 1 ♀ (ЗИН); «Керман, стр.[ана] Бам-
401
пур», 1-4.VI.1898 (Н. А. Зарудный), 11 ♂, 12 ♀ (ЗИН); «Сеистан», 8-9.VI.1898 (Н. А. Зарудный),
1 ♂ (ЗИН); «Базман-Тагаб, В Кирман», 8-10.VIII.1898 (Н. А. Зарудный), 1 экз. (ЗИН). Израиль.
Qesarya, 23.XII.2003 (V. Kravchenko, V. Chikatunov), 1 ♀ (КСР). Египет. «Aegypt» (дата и сбор-
щик неизвестны), из коллекции П. П. и А. П. Семеновых-Тян-Шанских, 1 ♀ (ЗИН).
И зм ен чиво с ть и сходн ы е в и ды. Цвет рисунка на голове варьирует от бу-
рого до буроватого, наличник у самцов иногда с коротким треугольным пятном. Ши-
рина пятна на наличнике у самок варьирует. Рисунок на лбу иногда редуцирован до
двух симметрично расположенных пятен. Нередко основная половина 2-го максилляр-
ного пальпомера затемненная, но менее интенсивно, чем обычно у E. testaceus. Среди
рассмотренных в статье видов E. ater выделяется темной окраской ног и обеих сторон
тела. Сходно окрашены E. politus и E. salomonis, известные с юга Западной Пале-
арктики, но отличаются от него поверхностной простой или двойной пунктировкой
переднеспинки, поверхностной пунктировкой надкрылий, строением эдеагуса и дру-
гими признаками (Ribera et al., 1997). По строению эдеагуса E. ater сходен с E. quadri-
punctatus и E. fuscipennis, но легко отличается самыми маленькими среди рассмо-
тренных видов дорсальными зубцами парамер, вершины которых достигают 1/4, а не
1/3 длины базальных апофиз. Длина тела самых мелких изученных экземпляров
E. ater из Ирана составляет 4.5 мм, что сопоставимо с размерами E. salomonis.
Р асп ро ст р анени е. Широко распространен на юге Палеарктики, на восток известен до
Синьцзян-Уйгурского автономного района Китая; Алжир, Египет (Fikáček et al., 2015); Азер-
байджан (Pallarés et al., 2017: Table S1). Здесь впервые указан из Украины и Киргизии. В России
ранее был известен лишь из Крыма (Greń, 2018).
П р им ечан ие. Несколько экземпляров E. ater из Азербайдажана (ЗИН) были
обозначены Ф. А. Зайцевым как «E. (Lumetus) kiritshenkoi sp. n.», но описание вида не
было опубликовано. См. также обсуждение E. asiaticus.
Enochrus bicolor (Fabricius, 1792) (рис. 2, 5; 3, 1; 4, 4, 14, 21; 7, 4, 9, 11, 18).
Мат ер иа л. Свыше 270 экз. из следующих регионов. Украина. Николаевская и Донецкая
области. Россия. Карелия, Вологодская, Тверская, Ярославская, Московская, Липецкая, Тамбов-
ская, Пензенская, Белгородская, Воронежская, Самарская, Саратовская и Астраханская области,
Крым, Краснодарский край, Башкирия, Оренбургская, Челябинская и Кемеровская области.
Казахстан. Кызылординская, Туркестанская, Жамбылская и Алматинская области. Киргизия.
Иссык-Кульская обл.: западное побережье оз. Иссык-Куль, 5 км Ю пос. Балыкчи, 42°23ʹ03ʺ N,
76°11ʹ10ʺ E, 1615 м, на свет, 8-9.VII.2018 (С. В. Литовкин), 11 ♂, 3 ♀ (КСЛ). Джалал-Абадская
обл.: 5 км З г. Таш-Кумыр, 41°20ʹ08ʺ N, 72°07ʹ57ʺ E, 775 м, на свет, 16-17.VII.2017 (С. В. Литов-
кин), 1 ♀ (КСЛ). Монголия. Баян-Улгий (оз. Хар-нуур, 9.VIII.2010 (А. А. Прокин), 1 ♂ (ИБВВ)),
Увс (Давст сомон, окр. оз. Увс-нуур, 50°34ʹ08ʺ N, 92°28ʹ49ʺ E, лужа, 30.VII.2010 (А. А. Прокин),
1 ♂, 3 ♀ (ИБВВ)), Булган (Тешиг сомон, оз. Олон-нуyр, 49°55ʹ12ʺ N, 102°37ʹ20ʺ E, заросшие
мелководья, 24.VII.2009 (А. А. Прокин), 1 ♂ (ИБВВ)) и Уверхангай (Улзийт сомон, озеро в окр.
оз. Сангийн-Далай-нуур, 46°40ʹ20ʺ N, 103°18ʹ06ʺ E, 31.VIII.2010 (А. А. Прокин), 1 ♂ (ИБВВ))
аймаки.
Изм ен чиво ст ь и сходн ы е в иды. У единичных самок есть хорошо за-
метные размытые буроватые пятна на голове и в центре переднеспинки. У многих
самок верхняя губа буроватая или даже желтая. Окраска задних бедер изредка сходна
с таковой у E. segmeninotatus. Форма тела у единичных экземпляров E. bicolor
овальная, и они могут быть приняты за E. segmentinotatus. Enochrus bicolor выделя-
ется среди рассмотренных видов самыми крупными размерами и почти полностью
402
светлой окраской верхней стороны тела, но очень сходен с E. segmentinotatus (см. при-
мечания к этому виду, а также ревизию Ш. Шёдля (Schödl, 1998). У одного самца
E. bicolor из Иссык-Кульской обл. Киргизии (см. Материал) обнаружена аберрация па-
рамер типа «calabricus» (рис. 7, 18) (cм. обсуждение E. calabricus).
Р а спр о ст р ан ен и е. Широко распространен по всей Палеарктике, более обычен на юге;
Северная Африка (Fikáček et al., 2015). Впервые указывается из Киргизии.
Примечание. Материал из Монголии приводится как дополнение к обзору
А. Прокина с соавторами (Prokin et al., 2020). См. также примечание к E. segmentino-
tatus.
Enochrus fuscipennis (Thomson, 1884) (рис. 1, 7, 10, 11; 2, 13; 3, 9, 10; 4, 11, 19, 23;
7, 15, 19).
М ат е риа л. Свыше 240 экз. из следующих регионов. Россия. Вологодская, Ярославская,
Московская, Курская, Нижегородская, Липецкая, Тамбовская, Пензенская области, Татарстан,
Самарская, Белгородская, Воронежская, Саратовская, Волгоградская, Астраханская области,
Крым, Краснодарский и Ставропольский края, Кабардино-Балкария (42°58ʹ29ʺ N, 43°20ʹ05ʺ E,
1995 м, болото Уштулу, 21.IX.2018 (А. А. Пржиборо, A. С. Сажнев), 12 экз. (ИБВВ)), Северная
Осетия, Оренбургская, Челябинская и Кемеровская области. Грузия. Край Мцхета-Мтианети:
Mleti Distr., Dušet [Душети], 17.VIII.1916 (сборшик неизвестен), 1 ♀ (ЗИН). Край Самцхе-Джа-
вахети: Baraleti Prov., Achalk, VI-VII.1916 (сборщик неизвестен), из коллекции Ф. А. Зайцева,
2 ♂, 3 ♀ (ЗИН). Тбилиси: «Tiflis», 17.VI.1880 (сборщик неизвестен), из коллекции Г. Сиверса, 1 ♂
(ЗИН); «Tiflis, Caucasus», 3.VII.1917 (сборщик неизвестен), из коллекции Ф. А. Зайцева, 1 ♀
(ЗИН). Армения. Котайкская и Гехаркуникская области. Азербайджан. Агдашский и Ленкоран-
ский районы. Казахстан. Актюбинская, Туркестанская и Жамбылская области. Туркмения.
Ахалский велаят. Киргизия. Иссык-Кульская, Джалал-Абадская и Ошская области. Таджики-
стан. Горно-Бадахшанская АО. Турция. Провинция Ардахан.
И зм е нчиво ст ь и сходн ые в и ды. Иногда передний край наличника вогнут
почти V-образно. Рисунок головы сходен у обоих полов. Ширина пятна на наличнике
изменчива, но обычно оно более развито у самок. Верхняя губа у самцов иногда
светлее, чем у самок, но не желтая. У единичных экземпляров основная половина 2-го
максиллярного пальпомера слабо затемненная. Помимо окраски (см. определи-
тельную таблицу), у максиллярных щупиков изменчива форма пальпомеров - от узких
и вытянутых до более утолщенных и коротких. Пятно переднеспинки варьирует от
ограниченного четырехугольником из черных точек до более широкого, оно может
быть отделено от переднего края диска (как на рис. 2, 13; 3, 9) или соприкасаться с
ним (как на рис. 3, 10). У единичных экземпляров передний край отростка средне-
груди отогнут назад. Полиморфизм E. fuscipennis требует более детального изучения,
в том числе молекулярно-генетическими методами.
Enochrus fuscipennis очень сходен с E. quadripunctatus, вместе с которым часто летит
на свет и может встречаться в одном водоеме. Как правило, E. fuscipennis отличается
от E. quadripunctatus более темной окраской верхней стороны тела: надкрылья у него
от буроватых до черно-бурых, темнее фона переденспинки и головы. Наиболее за-
метны различия в окраске головы самцов. Определение по окраске некоторых экзем-
пляров, особенно самок, затруднительно как в связи с естественной изменчивостью
окраски, так и с ее изменением при хранении коллекционного материала. Гениталии
самцов этих видов не имеют ясных отличий, хотя относительная длина трубки пениса
у E. fuscipennis обычно меньше, чем у E. quadripunctatus. У одного самца E. fuscipennis
403
из Кабардино-Балкарии (см. Материал) обнаружена аберрация парамер типа «cala-
bricus» (рис. 7, 19) (см. обсуждение E. calabricus). См. также обсуждение E. sahlbergi.
Р асп ро ст р анени е. Широко распространен в Палеарктике, на восток доходит до Восточ-
ной Сибири, Синьцзян-Уйгурского автономного района и провинции Хэбэй в Китае; Северная
Африка (Fikáček et al., 2015; Przewoźny, 2021); Туркмения (Зайцев, 1951; Старостин, 1992); Тад-
жикистан (Sazhnev, 2020). Впервые приводится для Грузии.
Enochrus halophilus (Bedel, 1879) (рис. 2, 12; 3, 8; 4, 10, 18, 25; 7, 14, 20).
Мат ер иа л. Украина. Николаевская обл.: Кинбургская коса, соленое озеро, 1.V.2007
(В. Г. Дядичко), 1 ♂ (ИБВВ); там же, тростниковое болото в степи, 29.VI.2007 (В. Г. Дядичко),
1 ♂, 1 ♀ (ИБВВ). Россия. Крым: Симферопольский р-н, окр. балки Тубай, 45°03ʹ N, 34°10ʹ E, на
свет, 28.VI.2016 (В. В. Шапоринский), 2 ♂, 1 ♀ (КСЛ); Керченский п-ов, г. Щёлкино, на свет,
6.VII.2009 (В. В. Шапоринский), 1 ♂ (КСЛ). Краснодарский край: «Tamań» [Тамань], 31.III.1913
(сборщик неизвестен), из коллекции А. Н. Кириченко, 1 ♂ (ЗИН).
Изм ен чиво ст ь и сходн ые в и ды. От сходных по окраске E. quadripunctatus
и E. fuscipennis изученные нами экземпляры отличаются заметно большими разме-
рами тела. Литературные данные о размерах тела E. halophilus существенно разнятся:
4.5-5.5 мм (Hebauer, Klausnitzer, 1998), 5.0-5.8 мм (Hansen, 1987) и 5.5-6.2 мм (Berge
Henegouwen et al., 1992). Мелкие экземпляры E. halophilus могут быть приняты за
E. quadripunctatus или E. fuscipennis.
Ра сп ро ст ра нен ие. Центральная и Южная Европа, Кипр, Турция, Иран (Fikáček et al.,
2015; Gentili et al., 2018).
Пр им еч ани е. Указание вида из Украины (Ryndevich, 2007) частью относится
к ошибочно определенным экземплярам E. bicolor: Донецкая обл.: «Novoazovsk Distr.,
3 km S. near Sedovo, estuary, 12.VII.2004 (S. K. Ryndevich)», 1 ♂, 2 ♀ (ИБВВ). Пре-
дыдущее указание из России (Воронежская обл.) (Prokin et al., 2008) также основано
на ошибочном определении E. bicolor (Прокин, 2018), а материал из Крыма (Ryndevich,
Fikáček, 2013) нуждается в проверке. Находки вида в Турции требуют подтверждения.
Перечисленные в одной из работ (Darilmaz, Kiyak, 2009) признаки не соответствуют
E. halophilus, а изображенный в ней эдеагус наиболее сходен с таковым E. hamifer.
Enochrus hamifer (Ganglbauer, 1901) (рис. 2, 7; 3, 3; 4, 6, 20; 7, 6).
Мат ер иа л. Украина. Херсонская обл.: Черноморский заповедник, постоянный степной во-
доем, 10.VII.1978 (А. Г. Шатровский), 1 ♂ (ИБВВ). Россия. Крым: Саки, Евпаторский уезд,
26.VI.1900 (W. Pliginski), 1 экз. (ЗИН). Самарская обл.: Кинельский р-н, окр. пос. Формальный,
болото Моховое-1, 53°07ʹ11ʺ N, 50°43ʹ33ʺ E, на свет, 30-31.VII.2020 (А. Е. Кузовенко), 72 ♂, 35 ♀
(КСЛ). Астраханская обл.: Богдинско-Баскунчакский заповедник, 7 км СЗ горы Большое Богдо,
4.V.2006 (А. В. Ковалёв), 1 ♀ (ИБВВ). Дагестан: Дербент, 10.VI.1904 (К. Сатунин), 2 экз. (ЗИН).
Оренбургская обл.: Соль-Илецкий р-н, окр. пос. Новоилецк, 5-6.V.2010 (А. М. Шаповалов), 1 ♀
(КСЛ). Иркутская обл.: «Иркутская губерния», VI.1907 (Д. А. Смирнов), 3 экз. (ЗИН). Азербайд-
жан. Агдашский р-н: «Geok-Tapa, Caucasus» (без даты) (A. Schelkownikow), 4 экз. (ЗИН); «Aresch,
Caucasus» (без даты) (A. Schelkownikow), 1 ♀ (ЗИН); «Geok-tapa distr., Areš», 28.IV.1913 (сбор-
щик неизвестен), 2 экз. (ЗИН). Астаринский р-н: «Ленкоранский округ, с. Мотал-Ятаг», в труто-
вике и гнилом дубе, 14.VII.[19]19 (П. Алексеев), 1 экз. (ЗИН). Армения. «Uluelralnu, prov.
Erivan», 13.V.1917 (Bening), 1 экз. (ЗИН). Казахстан. Актюбинская обл.: окр. Челкара [г. Шел-
кар], степь Большие Барсуки, 22.VI.1907 (Н. В. Андросов), 1 экз. (ЗИН); «Темирский уезд,
Уральская обл.», урочище Кок-Джида, 17.VI.1908 (Д. Бородин и В. Уваров), 1 экз. (ЗИН). Кара-
гандинская обл.: Бассага [с. Босага], 13.V.1957 (Грунин), 1 экз. (ЗИН). Кызылординская обл.:
404
4.3 км ЮВ пос. Тартогай, тугай близ р. Сырдарья, 44°24ʹ37ʺ N, 66°16ʹ30ʺ E, 145 м, на свет,
1.V.2014 (А. М. Шаповалов), 1 ♂, 1 ♀ (КСЛ). Туркестанская обл.: северный край оз. Кызылколь,
43°46ʹ34ʺ N, 69°30ʹ36ʺ E, 335 м, солончак, на свет, 5.V.2015, 4.VI.2015 (С. В. Литовкин), 9 ♂, 3 ♀
(КСЛ); там же, но рядом, 31.V-1.VI.2017 (С. В. Литовкин), 31 экз. (КСЛ); там же, 43°45ʹ39ʺ N,
69°28ʹ47ʺ E, 327 м, в воде, 31.V-1.VI.2017 (С. В. Литовкин), 1 ♂, 2 ♀ (КСЛ); 7.4 км СВ пос. Бал-
таколь, тугай близ р. Сырдарья, 43°09ʹ24ʺ N, 67°51ʹ00ʺ E, 180 м, на свет, 9-10.VI.2016 (С. В. Ли-
товкин), 1 ♂, 2 ♀ (КСЛ). Жамбылская обл.: «Туркестан, Аулие-Ата» [г. Тараз] (без даты)
(E. Willberg), 1 экз. (ЗИН); Аксуек, на свет, 7-11.VI.1993 (А. С. Тилли), 1 ♂ (КСЛ); 7 км СВ пос.
Кумкент, 43°48ʹ58ʺ N, 69°41ʹ46ʺ E, 300 м, солончак, на свет, 13-14.V.2015 (С. В. Литовкин), 4 ♂,
3 ♀ (КСЛ); западный край оз. Акколь, 43°24ʹ30ʺ N, 70°37ʹ30ʺ E, 405 м, на свет, 17-18.V.2015
(С. В. Литовкин), 35 экз. (КСЛ); берег соленого оз. Ащыколь, 43°32ʹ19ʺ N, 70°37ʹ48ʺ E, 380 м, на
свет, 18-20.V.2015 (С. В. Литовкин), 14 экз. (КСЛ). Алматинская обл.: берег р. Или, окр.
пос. Айдарлы, 43°58ʹ00ʺ N, 79°35ʹ55ʺ E, 3.VII.2010 (Д. В. Потанин), 1 ♀ (КСЛ); там же, обмелев-
шая протока р. Или, 43°58ʹ09ʺ N, 79°36ʹ03ʺ E, 500 м, 18.VI.2018 (С. В. Литовкин), 1 ♀ (КСЛ); окр.
г. Капчагай, берег р. Или, 43°55ʹ51.7ʺ N, 77°05ʹ30.8ʺ E, ~ 500 м, 22-23.IV.2014 (С. К. Корб), 1 ♂,
1 ♀ (КСЛ); 45.5 км ВСВ пос. Шелек, пустыня, 43°43ʹ59ʺ N, 78°49ʹ11ʺ E, 490 м, на свет,
11-12.VI.2018 (С. В. Литовкин), 2 ♂, 2 ♀ (КСЛ); 5.5 км ССЗ пос. Айдарлы, пустыня, 44°04ʹ48ʺ N,
79°28ʹ12ʺ E, 625 м, на свет, 15-16.VI.2018 (С. В. Литовкин), 3 ♂ (КСЛ). Узбекистан. Бухарская
обл.: Бухара, болото Углановское, 22.IV.1925 (И. Соколов), 3 экз. (ЗИН); Бухара, на оз. Кули-
Суфион [озеро-болото Кулисуфиён], 26.IV.1925 (И. Соколов), 1 экз. (ЗИН). Сырдарьинская обл.:
ж.-д. ст. Голодная Степь [г. Гулистан], Среднеазиатская железная дорога, 31.V.1903 (Г. Г. Якоб-
сон), 1 экз. (ЗИН). Киргизия. Джалал-Абадская обл.: 10 км ЗСЗ г. Таш-Кумыр, 41°23ʹ15ʺ N,
72°06ʹ05ʺ E, ~ 1010 м, на свет, 17-18.VI.2017 (С. В. Литовкин), 22 экз. (КСЛ); В пос. Кара-Алма,
~ 41°12ʹ50ʺ N, 73°21ʹ31ʺ E, ~ 1500 м, лесистый горный склон, на свет, 11-12.VII.2017 (С. В. Ли-
товкин), 3 ♀ (КСЛ). Туркмения. Балканский велаят: «Кизил-Арват [Сердар], Арман-Саад»,
VI.1896 (К. О. Ангер), 1 экз. (ЗИН). Ахалский велаят: «Karategen» [? гора Каратегелен],
4.VIII.1899 (сборщик неизвестен), 9 экз. (ЗИН); «Закаспийская обл., Душанг» [? Душак],
18.VI.1898 (К. О. Ангер), 1 экз. (ЗИН). Иран. «Хадери-Пефес, с.-в. Персия», 15.III.1916
(Б. Ильин), 1 экз. (ЗИН).
И зм е нчиво с ть и сходны е в и ды. Цвет рисунка на голове варьирует от бу-
рого до буроватого, редко рисунок почти исчезает. Ширина пятна на наличнике у
самок варьирует. Наличник самцов иногда с коротким размытым треугольным пятном
или с размытой медиальной полоской; редко верхняя губа самцов с буроватым пятном.
Рисунок на лбу иногда редуцирован до двух симметрично расположенных пятен. 2-й
максиллярный пальпомер в основной половине и 4-й перед вершиной могут быть
слабо затемнены. В зависимости от интенсивности окраски E. hamifer может быть
сходным с разными видами, но выделяется небольшими размерами и быстро распоз-
нается по характерному крючковидному отростку на переднегруди (рис. 4, 20).
Р а спр о ст р ан ен и е. Центральная и Южная Европа, Израиль, Средняя Азия, Иран (Fikáček
et al., 2015; Gentili et al., 2018); Армения, Азербайджан (Марджанян, 1988). Впервые приводится
из Сибири, ранее в России был известен на восток до Среднего Урала (Свердловская обл.)
(Кирейчук, Шатровский, 2001).
Enochrus ochropterus (Marsham, 1802) (рис. 1, 8; 2, 9; 3, 5; 4, 8; 5, 1-9; 7, 8).
Philhydrus ferrugineus Motschulsky, 1849, syn. n.
М ате риа л. 25 экз. из следующих регионов. Россия. Новгородская, Вологодская, Ярослав-
ская, Московская и Нижегородская области, Удмуртия, Тамбовская, Воронежская, Самарская,
Саратовская и Кемеровская области.
405
Рис. 5. Philhydrus ferrugineus Motschulsky, типовые экземпляры.
1, 8 - типовые экземпляры на пластинке; 2 - лектотип дорсально; 3 - паралектотип дорсально;
4, 5 - окраска головы, лектотип и паралектотип; 6 - пунктировка надкрылий, лектотип;
7 - максиллярный щупик, лектотип; 9 - этикетки типовых экземпляров.
Масштабная линейка для 1 - 10 мм.
Philhydrus ferrugineus был описан из Восточной Сибири («Daourie boréale») как
сходный с Ph. ochropterus вид с полностью темной головой, коричневыми («brun un
peu châtain») надкрыльями и немного более уплощенным телом (Motschulsky, 1849).
После описания статус таксона не проверялся, предполагалась его принадлежность
к подроду Methydrus (Hansen, 1999). В последнем издании Палеарктического каталога
вид был правильно помещен в подрод Lumetus (Fikáček et al., 2015).
Голотип в описании не обозначен и количество типовых экземпляров не указано.
Нами были изучены две самки-синтипа (помечены буквой «m.») плохой сохранности
из коллекции В. Мочульского в Зоомузее МГУ. В качестве лектотипа здесь обозначает-
ся самка с сохранившимися щупиками (рис. 5, 2) и этикеткой «fl Turra Sib. orient //
ferrugineus m.». Вторая самка обозначается как паралектотип (рис. 5, 3). Размеры и
форма тела, характер пунктировки надкрылий, окраска головы и переднеспинки, фор-
ма и окраска щупиков лектотипа (рис. 5, 1, 4, 6, 7) соответствуют признакам
E. ochropterus, на этом основании мы устанавливаем здесь новую синонимию.
Изм енч иво ст ь и сход ны е в и ды. 4-й максиллярный пальпомер пол-
ностью светлый или слабо затемнен перед вершиной. Размер темного пятна на налич-
нике у самцов сильно варьирует. Темное пятно на переднеспинке обычно более раз-
вито у самок, так, что светлыми остаются только ее края. Встречаются как
406
светлоокрышенные, так и темноокрашенные особи. Светлоокрашенные самки
E. ochropterus по рисунку головы, форме тела и размерам сходны с E. melanocephalus,
однако легко отличаются от него перечисленными в определительной таблице призна-
ками. По общей окраске тела темные экземпляры E. ochropterus могут быть сходны
с E. fuscipennis. Полиморфизм E. ochropterus по окраске тела требует более детального
изучения.
Р а спр о ст р ан ен и е. Широко распространен в Европе; Западная и Восточная Сибирь, Тур-
ция, Иран (Fikáček et al., 2015; Przewoźny, 2021).
Примечание. Экземпляр, указанный из Таджикистана (Sazhnev, 2020) как
E. ochropterus, в действительности относится к неизвестному нам виду подрода
Methydrus.
Enochrus puetzi Hebauer, 1995 (рис. 1, 4; 2, 15; 3, 12; 4, 13; 7, 17, 22).
М ате риа л. Россия. Приморский край: Лазовский заповедник, с. Лазо, на свет,
18-20.VIII.2005 (В. Шохрин, Ю. Сундуков), 5 экз. (ИБВВ); там же, 7-13.VIII.2006 (Ю. Сундуков,
В. Шохрин), 10 экз. (ИБВВ); там же, 23-29.VII.2007 (В. Шохрин), 2 экз. (ИБВВ); Уссурийский
р-н, окр. с. Кроуновка, 8-11.VII.2002 (А. С. Тилли), 1 ♂ (КСЛ); Октябрьский р-н, 7 км З с. По-
кровка, на свет, 11-12.VIII.2018 (С. Н. Иванов), 1 ♂, 2 ♀ (КСЛ).
Типо вой мат ер и а л. Изучены фотографии двух паратипов-самцов и эдеагуса (колл.
Andreas Pütz, Eisenhüttenstadt, Germany).
Enochrus puetzi был описан по серии экземпляров из Приморского края (Hebauer,
1995), где недавно был отмечен в Лазовском заповеднике (Прокин, 2009) (см. Мате-
риал). Изначально вид был помещен в подрод Lumetus, но впоследствии без пояснений
перенесен в подрод Methydrus (Hansen, 1999, 2004). Изучение нами фотографий ти-
повых экземпляров и коллекционного материала подтвердило принадлежность
E. puetzi к подроду Lumetus. В частности, вершина 5-го вентрита брюшка у него
цельная, без вырезки, и без грубых щетинок (рис. 1, 4). Enochrus puetzi очень близок
к E. testaceus как по внешним признакам, так и по строению эдеагуса; в частности,
вершина шпоры пениса у обоих видов характерно оттянута дорсально.
Р а спр о ст р ан ен и е. Юг Дальнего Востока России (Приморский край: Hebauer, 1995; Про-
кин, 2009; Fikáček et al., 2015).
Enochrus quadripunctatus (Herbst, 1797) (рис. 1, 3; 2, 10; 3, 6; 4, 9, 16, 24; 6, 1-4;
7, 13, 23).
М ате риа л. Свыше 430 экз. из следующих регионов. Украина. Николаевская обл. Россия.
Вологодская, Ярославская, Московская, Нижегородская, Брянская, Курская, Липецкая и Тамбов-
ская области, Чувашия, Удмуртия, Татарстан, Белгородская, Воронежская, Самарская, Саратов-
ская, Волгоградская и Астраханская области, Крым, Краснодарский и Ставропольский края,
Дагестан, Оренбургская и Кемеровская области, Алтайский край, Якутия (Якутск), Приморский
край. Грузия. Край Шида-Картли и Тбилиси. Казахстан. Актюбинская, Акмолинская, Кызылор-
динская, Туркестанская, Жамбылская и Алматинская области. Киргизия. Чуйская обл. (Суса-
мырская долина). Монголия. Увс, Хувсгел, Сэлэнгэ, Хэнтий и Баянхонгор аймаки.
И зм е нчиво ст ь и сходн ые в и ды. Иногда передний край наличника вогнут
почти дуговидно. Верхняя губа у самцов редко с маленьким темным пятном. У еди-
ничных экземпляров пятно переднеспинки может редуцироваться либо расширяться
407
Рис. 6. Enochrus quadripunctatus (Herbst), эдеагус.
1 - эдеагус дорсально; 2 - эдеагус дорсально, схема; 3 - эдеагус вентрально, схема; 4 - пенис латерально.
bap - базальные апофизы; bst - базальные зубцы парамер; cap - наконечник шпоры пениса; cor - корона;
drt - дорсальные зубцы парамер; phb - фаллобаза; prm - парамеры; spr - шпора пениса; tpn - трубка
пениса; vpl - вентральная пластинка пениса. a - длина шпоры пениса с наконечником; b - длина
базальных апофиз; c - длина дорсальных зубцов относительно базальных апофиз.
до границ четырехугольника из черных точек в виде темного гало. Вид, вероятно,
может образовывать меланистические формы (см. E. sahlbergi).
Enochrus quadripunctatus очень похож на E. fuscipennis (см. примечания к этому
виду). Как правило, экземпляры E. quadripunctatus отличаются от E. fuscipennis более
светлой окраской верхней стороны тела: надкрылья у него желтые или темно-желтые,
одного цвета с фоном переднеспинки и головы. Из-за изменчивости окраски обоих
видов определение некоторых экземпляров затруднительно, особенно самок.
Р асп ро ст р анени е. Широко распространен в Палеарктике, но не найден в Северной
Африке, Японии и на Курильских островах (Fikáček et al., 2015).
Enochrus segmentinotatus (Kuwert, 1888) (рис. 2, 6; 3, 2; 4, 5, 15, 22; 7, 5, 10, 12).
Мат ер иа л. Украина. Николаевская обл.: Кинбургская коса, соленое озеро, 1.V.2007
(В. Г. Дядичко), 2 ♂; там же, соленая лужа и озеро, 29-30.VI.2007 (В. Г. Дядичко), 1 ♂; там же,
соленое озеро, берег лимана, 29.VI.2007 (В. Г. Дядичко), 3 ♂, 2 ♀ (ИБВВ). Россия. Крым: Сим-
феропольский р-н, окр. балки Тубай, 45°03ʹ N, 34°10ʹ E, на свет, 28.VI.2016 (В. В. Шапорин-
ский),
2
♂ (КСЛ). Краснодарский край: Краснодар, ул. Вавилова, на свет,
5.VII.2018
408
(Е. Ю. Родионова) 1 ♂ (ИБВВ). Саратовская обл.: Ровенский р-н, 8 км ССВ с. Луговское,
50°42ʹ02ʺ N, 46°33ʹ21ʺ E, 7.V.2010 (Д. В. Потанин), 1 ♂ (КСЛ); Александрово-Гайский р-н, окр.
с. Ветелки, лиман, 27 м, на свет, 15.V.2019 (Е. Ю. Масолова), 4 ♂ (ИБВВ); Озинский р-н, 12 км С
п. г. т. Озинки, песчаный карьер, 142 м, песчаная степь, на свет, 27-28.V.2019 (В. В. Аникин), 2 ♂,
1 ♀ (ИБВВ). Астраханская обл.: Богдинско-Баскунчакский заповедник, 7 км СЗ горы Большое
Богдо, 4.V.2006 (А. В. Ковалёв), 3 ♂ (ИБВВ); Красноярский р-н, с. Досанг, берег р. Ахтуба, на
свет, 1-8.V.2011 (И. А. Забалуев), 1 ♂, 1 ♀ (КСЛ). Оренбургская обл.: Акбулакский р-н, окр. пос.
Акоба, 50°54ʹ N, 55°54ʹ E, на свет, 16.VI.2012 (А. М. Шаповалов), 1 ♂ (КСЛ). Казахстан. Юго-За-
падный Казахстан, 23-24.V.2007 (Д. Ф. Шовкун), 1 ♂ (КСЛ); Кызылординская обл.: 1 км Ю пос.
Акжарма, 44°50ʹ34ʺ N, 65°21ʹ09ʺ E, 120 м, на свет, 25-26.IV.2013 (С. В. Литовкин), 1 ♂ (КСЛ);
21 км СЗ г. Аральск, 46°57ʹ50ʺ N, 61°29ʹ07ʺ E, 80 м, пустыня, на свет, 7-8.V.2013 (С. В. Литов-
кин), 38 экз. (КСЛ). Туркестанская обл.: С оз. Кызылколь, 43°46ʹ34ʺ N, 69°30ʹ36ʺ E, 335 м, солон-
чак, на свет, 5.V.2015 (С. В. Литовкин), 1 ♂ (КСЛ); северный край оз. Кызылколь, 43°45ʹ39ʺ N,
69°28ʹ47ʺ E, 327 м, в воде, 31.V-1.VI.2017 (С. В. Литовкин), 3 ♂ (КСЛ); 7.4 км СВ пос. Балтаколь,
возле р. Сырдарья, 43°09ʹ24ʺ N, 67°51ʹ00ʺ E, 180 м, тугай, на свет, 9-10.VI.2016 (С. В. Литовкин),
1 ♂ (КСЛ). Жамбылская обл.: Южное Прибалхашье, Мынарал [~ 45°26ʹ N, 73° 39ʹ E], 6.VI.1993
(А. С. Тилли), 1 ♂ (КСЛ); окр. оз. Ащыколь, 43°29ʹ43ʺ N, 70°39ʹ37ʺ E, 375 м, остаточная соленая
лужа в русле небольшой реки, 3.VI.2015 (С. В. Литовкин), 1 ♂, 2 ♀ (КСЛ). Иран. Провинция
Систан и Белуджистан: «Керман, стр.[ана] Бампур», 10-11.VIII.1898 (Н. А. Зарудный), 1 ♂
(ЗИН); «Керман, стр.[ана] Саргад», 28.VIII-4.IX.1898 (Н. А. Зарудный), 1 ♂ (ЗИН). Монголия.
Увс, Булган и Баянхонгор аймаки (данные этикеток см. Prokin et al. (2020)).
И зм е нчиво ст ь и сход ны е в иды. У самок верхняя губа часто буроватая,
почти желтая. Вид по внешнему строению очень похож на E. bicolor, вместе с которым
часто летит на свет и может встречаться в одном водоеме, но отличается от него, как
правило, более узким телом и меньшими размерами. Достоверное определение воз-
можно только по строению эдеагуса. Самцы этих видов, кроме того, заметно различа-
ются по площади гидрофобного опушения задних бедер (ср. рис. 4, 21 и 4, 22). Опре-
деление самок затруднительно и часто возможно главным образом по объединению
с самцами.
Р а спр о ст р а не н ие. Широко распространен на юге Палеарктики, включая Северную
Африку (Fikáček et al., 2015).
Примечание. Экземпляр из Уверхангай аймака и часть экземпляров из Булган и
Увс аймаков Монголии, определенные как E. segmentinotatus (Prokin et al., 2020),
в действительности относятся к E. bicolor (см. материал по этому виду).
Enochrus testaceus (Fabricius, 1801) (рис. 2, 14; 3, 11; 4, 12; 7, 16, 21, 24).
М ат е риа л. Свыше 150 экз. из следующих регионов. Украина. Николаевская обл. Россия.
Тверская, Вологодская, Ярославская, Рязанская, Курская, Липецкая, Пензенская области, Мор-
довия, Удмуртия, Татарстан, Белгородская, Воронежская, Самарская, Саратовская и Астрахан-
ская области, Крым, Краснодарский и Ставропольский края, Оренбургская, Челябинская и
Кемеровская области, Якутия (Якутск), Курильские острова (о. Кунашир). Казахстан. Актюбин-
ская, Кызылординская, Туркестанская и Алматинская области. Киргизия. Джалал-Абадская
обл. Монголия. Баян-Улгий аймак.
И зм е нчиво ст ь и сходн ы е в и ды. 4-й максиллярный пальпомер почти
полностью светлый или в разной степени затемнен перед вершиной. У самцов верхняя
губа иногда с бурыми пятнами. Надкрылья иногда узкие, почти параллельносто-
ронние. Некоторые экземпляры с размытым бурым или буроватым рисунком головы
409
Рис. 7. Enochrus spp., эдеагус и детали его строения.
1 - E. melanocephalus (Olivier); 2 - E. nigritus (Sharp); 3 - E. simulans (Sharp); 4, 9, 11 - E. bicolor (F.),
типичная форма; 5, 10, 12 - E. segmentinotatus (Kuwert); 6 - E. hamifer (Ganglbauer); 7 - E. ater (Kuwert);
8 - E. ochropterus (Marsham); 13, 23 - E. quadripunctatus (Herbst); 14, 20 - E. halophilus (Bedel);
15 - E. fuscipennis (Thomson), типичная форма; 16, 21, 24 - E. testaceus (F.); 17, 22 - E. puetzi Hebauer;
18 - E. bicolor (F.), аберрация «calabricus»; 19 - E. fuscipennis (Thomson), аберрация «calabricus».
1-8, 13-19 - эдеагус дорсально; 9, 10 - форма внутреннего края парамер; 11, 12, 23, 24 - шпора пениса
с наконечником и трубка пениса латерально; 20 - диагностические признаки пениса, дорсально;
21, 22 - форма вершины парамер. Каждая группа изображений представлена в едином масштабе.
Масштабная линейка для 1-8, 13-19 - 0.5 мм.
410
сходны с E. ater, особенно жуки с более темными голенями. От E. ater такие экзем-
пляры отличаются более широким телом и/или большими размерами тела и/или зачер-
ненным (а не буроватым) в основной половине 2-м максиллярным пальпомером и/или
светлыми голенями и/или светлой окраской верхней стороны тела. Самки E. puetzi от-
личается от E. testaceus только заметно меньшими размерами тела и темным пятном
на переднеспинке. Недоокрашенные E. testaceus сходны с E. bicolor.
Р асп ро ст р анени е. Широко распространен в Европе и Сибири, отмечен на Дальнем Вос-
токе России, в Турции, Иране, Казахстане, Синьцзян-Уйгурском автономном районе Китая,
в Японии (Fikáček et al., 2015; Przewoźny, 2021), Туркмении (Зайцев, 1951; Старостин, 1992),
Киргизии (Овчинников, 1996).
Enochrus turanicus Schödl, 1998 (рис. 1, 9, 12; 2, 4).
Мат ер иа л. Туркмения. Ахалский велаят: окр. г. Теджен, 17-24.IV.1985 (А. Тилли и В. Ни-
кулин), 1 ♀ (КСЛ).
Описание вида и изображение его эдеагуса см. в работе Ш. Шёдля (Schödl, 1998).
И зм ен чиво с ть и сход ны е в и ды. Судя по литературным данным, вид
сходен с E. sinuatus dʹOrchymont, 1937 (Schödl, 1998), известным из Таджикистана
(Hebauer, 1991), Ирана, Кувейта и Пакистана (Fikáček et al., 2015; Przewoźny, 2021).
Р асп ро ст р анени е. Туркмения, Иран (Schödl, 1998; Fikáček et al., 2015).
ТАКСОНЫ С НЕЯСНЫМ СТАТУСОМ
Enochrus asiaticus (Kuwert, 1893).
Описан из Туркмении («Kissil Arrat» - Кизыл-Арват, сейчас Сердар) (Kuwert, 1893).
Впоследствии указан из Туркмении (Старостин, 1992), а также из Восточного Закав-
казья (Кирейчук, Шатровский, 2001), Ирана (Кирейчук, Шатровский, 2001; Gentili
et al., 2018), Казахстана (Ryndevich, 2011) и Северного Китая (Jia, Wang, 2010), однако
в изученном нами материале из Закавказья, Ирана, Южного Казахстана и Средней
Азии вид не обнаружен. Указания вида из Казахстана (Ryndevich, 2011) относятся
в действительности к E. ater (см. материал по этому виду). Определенные как
E. asiaticus экземпляры из Азербайджана и Туркмении в коллекции ЗИН также отно-
сятся к E. fuscipennis или E. ater.
По литературным данным, E. asiaticus сходен с E. ater, E. politus (Kuwert, 1893;
Hebauer, 1991) и E. fuscipennis (Jia, Wang, 2010). Его главными отличительными при-
знаками названы укороченный (или «вздутый») и затемненный на вершине 4-й мак-
силлярный пальпомер, более длинный (выступающий вперед в виде крупного зубца:
см. рис. 3d и 4d в: Hebauer, 1991), чем у E. ater, отросток среднегруди, а также
«длинная» шпора пениса с наконечником, отграниченным значительно дистальнее ко-
роны (см. Hebauer, 1991: рис. 4a; Jia, Wang, 2010: рис. 16). Статус E. asiaticus требует
уточнения; вероятно, он близок к E. fuscipennis или конспецифичен с ним.
Enochrus calabricus (Ferro, 1976).
Описан из Италии (Калабрия) по единственному самцу с необычным для Enochrus
строением вершин парамер (Ferro, 1976). Подродовая принадлежность в описании не
412
указана. Позже несколько самцов из Турции были отнесены к этому же виду, который
был помещен в подрод Lumetus (Polat et al., 2015). В сериях E. fuscipennis (см. мате-
риал из Кабардино-Балкарии) и E. bicolor (см. материал из Иссык-Кульской обл.
Киргизии) есть по одному самцу, сходному по строению парамер с E. calabricus
(рис. 7, 18, 19), а по внешним признакам самцы не отличаются от остальных экзем-
пляров серий (окраску головы самца E. fuscipennis см. на рис. 3, 10). Это говорит
о независимом происхождении аберрантного строения парамер, которое описано как
уникальный признак E. calabricus.
Судя по фотографиям турецких жуков (Polat et al., 2015), по крайней мере часть из
них может относиться к E. fuscipennis. В то же время, указанные в тексте признаки
(максимальная длина тела, форма тела, окраска максиллярных щупиков) не харак-
терны для E. fuscipennis. Приведенные в оригинальном описании (Ferro, 1976) при-
знаки (окраска головы, верхней губы, максиллярных щупиков и переднеспинки) соот-
ветствуют E. fuscipennis, однако на рисунке эдеагуса показаны относительно более
длинная в сравнении с E. fuscipennis трубка пениса и шпора пениса с отломленной
вершиной. Таким образом, без изучения типового экземпляра мы не можем досто-
верно судить о синонимии E. calabricus.
Enochrus sahlbergi (Fauvel, 1887) (рис. 2, 11; 3, 7; 4, 17).
Philydrus sahlbergi Fauvel, 1887.
М ат е риа л. Швеция. «Ekerö // 18.6.1922 // Håkan Lindb. // 203 // Philydrus sahlbergi Kuw.
K. Lindberg det.», 2 ♂ (один без головы) (ЗИН); «Ekerö // 18.6.1922 // Håkan Lindb. // Philydrus
sahlbergi Kuw. K. Lindberg det.», 3 ♀ (ЗИН). Финляндия. «H: fors // J. Sahlb. // Philydrus Sahlbergi
// 305 // к. Г. Сиверса», 1 ♂ (сохранился только эдеагус на картонной пластинке) (ЗИН); «H: fors
// J. Sahlb. // Philydrus Sahlbergi Fauv.», 1 ♀ (ЗИН); «221 // Finland // 51 Db // Philydrus. Sahlbergi
Kuw // колл. Семенова-Т.-Ш.», 1 ♀ (ЗИН); «Tvärminne. // Håkan Lindb. // Enochrus Sahlberg», 1 ♂,
1 ♀ (ЗИН); «Tvärminne. // Håkan Lindb.», 2 ♀ (ЗИН).
Enochrus sahlbergi (Fauvel, 1887) рассматривается современными авторами как
темная форма и синоним E. fuscipennis (Fikáček et al., 2015), что обсуждается М. Хан-
сеном (Hansen, 1987). Нами были изучены несколько экземпляров из Швеции и Фин-
ляндии, определенные как E. sahlbergi, в том числе из коллекции Дж. Сальберга
(J. Sahlberg). На некоторых определительных этикетках автором таксона, вероятно,
ошибочно назван Kuwert, так как Philydrus sahlbergi Kuwert, 1888 считается сино-
нимом E. bicolor (Fikáček et al., 2015). Признаки изученных экземпляров отчасти соот-
ветствуют признакам, упомянутым А. Оршимоном (Orchymont, 1938) и М. Хансеном
(Hansen, 1987). У самцов наличник с широким в основании треугольным пятном, до-
ходящим или почти доходящим до его переднего края; верхняя губа большей частью
или полностью светлая (рис. 3, 7). У самок голова полностью черная (3, 7) или с уз-
кими, более или менее отчетливыми светлыми пятнами по боковым краям наличника.
4-й максиллярный пальпомер отчетливо затемнен перед вершиной. Окраска передне-
спинки, надкрылий и ног как у E. fuscipennis (рис. 2, 11) или более светлая, переходная
к окраске E. quadripunctatus; очень темные экземпляры неизвестны. Отчетливо
V-образно вогнутый наличник (рис. 4, 17) и отклоненный назад передний край от-
ростка среднегруди характерны для E. quadripunctatus. Эдеагус не отличается от эдеа-
гуса E. quadripunctatus или E. fuscipennis, наконечник отграничен на уровне короны.
Длина тела 4.9-5.3 мм. Таким образом, до проверки цито- и молекулярно-генети-
413
ческими методами мы рассматриваем такие экземпляры E. sahlbergi как мелани-
стичную форму E. quadripunctatus с более обильным типичным паттерном окраски.
Только одна самка (Tvärminne; без определения; на одной булавке с темной самкой
«E. quadripunctatus»; см. материал) имеет сходные с E. fuscipennnis признаки (форма
переднего края наличника, форма отростка среднегруди), она выделяется более круп-
ными размерами тела (5.7 мм) и более темными надкрыльями, голова у нее полностью
черная. Не исключено, что под названием E. sahlbergi ранее подразумевались экзем-
пляры как E. fuscipennis, так и E. quadripunctatus, что оставляет вопрос о его сино-
нимии открытым. Он осложняется тем, что типовые экземпляры E. sahlbergi никем из
исследователей не упоминаются.
БЛАГОДАРНОСТИ
Авторы выражают благодарность А. А. Гусакову и П. Н. Петрову (Москва) за воз-
можность ознакомиться с типовыми экземплярами Philhydrus ferrugineus; А. Пютцу
(A. Pütz, Eisenhüttenstadt, Germany) за фотографии типовых экземпляров E. puetzi;
А. Г. Кирейчуку (С.-Петербург) за возможность работы с коллекцией ЗИН; В. В. Ани-
кину (Саратов), В. Г. Дядичко (Одесса, Украина), С. Н. Иванову (Владивосток),
А. В. Ковалёву (С.-Петербург), А. Е. Кузовенко (Самара), С. К. Рындевичу (Баранови-
чи, Беларусь), А. С. Тилли (Самара), И. С. Турбанову и Д. А. Филиппову (Борок),
В. В. Шапоринскому (Симферополь) за предоставленный материал.
ФИНАНСИРОВАНИЕ
Работа А. С. Сажнева выполнена при поддержке Российского научного фонда (грант
№ 21-74-20001), работа А. А. Прокина - в рамках госзадания 121051100109-1.
СПИСОК ЛИТЕРАТУРЫ
Зайцев Ф. А. 1951. Водные жуки Туркменистана. Труды Мургабской гидробиологической станиции 1: 53-76.
Кирейчук А. Г., Шатровский А. Г. 2001. Семейство Hydrophilidae (Водолюбы). В кн.: С. Я. Цалолихин (ред.).
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 5. Высшие насе-
комые. СПб.: Наука, с. 300-326.
Марджанян М. А. 1988. Ревизия рода Enochrus Thomson фауны Армянской ССР (Coleoptera, Hydrophilidae).
Биологический журнал Армении 41 (8): 650-660.
Овчинников С. В. 1996. Сем. Hydrophilidae - водолюбы. В кн.: Э. Д. Шукуров (ред.). Кадастр генетического
фонда Кыргызстана. Том 3. Надкласс Hexapoda - шестиногие (Entognatha и Insecta). Бишкек: Алейне,
с. 111-113.
Прокин А. А. 2009. Семейство Hydrophilidae - Водолюбы. В кн.: С. Ю. Стороженко (ред.). Насекомые Лазов-
ского заповедника. Владивосток: Дальнаука, с. 110-112.
Прокин А. А. 2018. Дополнения и исправления списка водных жесткокрылых (Coleoptera) Центрального
Черноземья. В кн.: В. С. Сарычев, О. П. Негробов, В. Б. Голуб, Л. В. Большаков (ред.). Изучение и
сохранение беспозвоночных Центрально-Черноземного региона России: сборник научных статей, по-
священный памяти М. Н. Цурикова. Воронеж: Научная книга, с. 101-103.
Старостин И. В. 1992. Фауна внутренних водоемов Туркменистана. Ашхабад: Ылым, 256 с.
Berge Henegouwen A. L. van, Cuppen J. G. M., Drost M. B. P., Huijbregts H. 1992. Hydrophilidae (Spinnende
waterorren). In: M. B. P. Drost, H. P. J. J. Cuppen, E. J. van Nieukerken, M. Schreijer (eds). De waterkevers
van Nederland. Utrecht: Koninklijke Nederlandse Natuurhistorische Vereniging, p. 181-224.
Darilmaz M. C., Kiyak S. 2009. The genus Enochrus Thomson (Coleoptera: Hydrophilidae) from Turkey, checklist
and new records. Archives of Biological Science, Belgrade 61 (4): 767-772. doi: 10.2298/ABS0904767D
Ferro G. 1976. Diagnosi preliminare di una nuova specie di Hydrobiini (Coleoptera, Hydrophilidae). Rivista di
Idrobiologia 15: 433-437.
Fikáček M., Angus R. B., Gentili E., Jia F., Minoshima Y. N., Prokin A., Przewoźny M., Ryndevich S. K. 2015.
Family Hydrophilidae Latreille, 1802. In: I. Löbl, D. Löbl (eds). Catalogue of Palaearctic Coleoptera. Vol. 2,
pt. 1. Revised and Updated Edition. Leiden-Boston: Brill, p. 37-76.
414
Gentili E., Ostovan H., Ghahari H., Komarek A. 2018. Annotated checklist of Iranian Hydrophilidae (Coleoptera:
Polyphaga: Hydrophiloidea). Aquatic Insects 39 (1): 55-88.
Greń C. 2018. Addition to Crimean fauna of water beetles (Coleoptera: Dytiscidae, Hydrophilidae). Annals of the
Нansen M. 1987. The Hydrophiloidea (Coleoptera) of Fennoscandia and Denmark. Fauna Entomologica
Scandinavica 18: 1-254.
Hansen M. 1999. World Catalogue of Insects 2: Hydrophiloidea (s. str.) (Coleoptera). Stenstrup: Apollo Books,
416 p.
Hansen M. 2004. Family Hydrophilidae Latreille, 1802. In: I. Löbl, A. Smetana (eds). Catalogue of Palaearctic
Coleoptera. Vol. 2. Hydrophiloidea - Histeroidea - Staphylinoidea. Stenstrup: Apollo Books, p. 44-68.
Hebauer F. 1991. Die Hydrophiliden-Ausbeuten H. Muches und D. W. Wrases aus Tadzhikistan. Ein Beitrag zur
Fauna der UdSSR (Coleoptera, Hydrophilidae). Entomologische Nachrichten und Berichte 35: 175-179.
Hebauer F. 1995. Bekannte und neue Hydrophiloidea aus Ostsibirien (Col.). Entomologische Nachrichten und
Berichte 39: 29-36.
Hebauer F., Klausnitzer B. 1998. Insecta: Coleoptera: Hydrophilidae (exkl. Helophorus). In: J. Schwoerbel, P. Zwick
(eds). Süsswasserfauna von Mitteleuropa. Band 20, parts 7-9, 10-1. Stuttgart; Jena: Lübek Ulm, 134 p.
Jia F., Wang Y. 2010. A revision of the species of Enochrus (Coleoptera: Hydrophilidae) from China. Oriental Insects
44: 361-385. doi: 10.1080/00305316.2010.10417622
Kuwert A. 1893. Neue Coleopteren. Societas Entomologica 8 (3): 17-18.
Motschulsky V. 1849. Coléoptères reçus dʹun voyage de M. Handschuh dans le midi de lʹEspagne enumeres et suivis
de notes. Bulletin de la Société Imperial des Naturalistes de Moscou 22 (3): 52-163.
Orchymont A. dʹ. 1938. Revision dʹune espèce dʹHydrophilide de Zetterstedt, suivie de la discussion dʹun “Philydrus”
de Thomson. Opuscula Entomologica 3: 7-10.
Pallarés S., Arribas P., Bilton D. T., Millán A., Velasco J., Ribera I. 2017. The chicken or the egg? Adaptation to
desiccation and salinity tolerance in a lineage of water beetles. Molecular Ecology 26 (20): 5614-5628.
Polat A., Taşar G. E., İncekara Ü. 2015. A new record of Enochrus Thomson, 1859 (Coleoptera: Hydrophilidae) for
the Turkish fauna. Turkish Journal of Science and Technology 10 (1): 9-12.
Prokin A. A., Chuluunbaatar G., Angus R. B., Jäch M. A., Petrov P. N., Ryndevich S. K., Byambanyam E., Sazhnev
A. S., Shaverdo H. 2020. New records of water beetles (Coleoptera: Gyrinidae, Haliplidae, Noteridae,
Dytiscidae, Helophoridae, Hydrophilidae, Hydraenidae) and shore beetles (Coleoptera: Heteroceridae) of
Mongolia. Aquatic Insects 41 (1): 1-44. doi: 10.1080/01650424.2019.1651870
Prokin A. A., Ryndevich S. K., Petrov P. N., Andrejeva T. R. 2008. New data on the distribution of Helophoridae,
Hydrochidae and Hydrophilidae (Coleoptera) in Russia and adjacent lands. Russian Entomological Journal
17 (2): 145-148.
Przewoźny M. 2021. Catalogue of Palearctic Hydrophiloidea (Coleoptera). Internet version 2021-01-01. URL:
Ribera I., Schödl S., Hernando C. 1997. Enochrus ater (Kuwert) and E. salomonis (Sahlberg) (Coleoptera:
Hydrophilidae), two widespread but overlooked species new to the European fauna. Hydrobiologia 354 (1):
183-188.
Ryndevich S. K. 2007. New records of Palaearctic water beetles (Coleoptera: Dytiscidae, Helophoridae, and
Hydrophilidae). In: V. B. Golub (ed.). Questions of Aquatic Entomology of Russia and Adjacent Lands:
Materials of the Third All-Russia Symposium on Amphibiotic and Aquatic Insects. Voronezh: Publishing
Polygraphic Centre of Voronezh State University, p. 284-287.
Ryndevich S. K. 2011. New data on Holarctic and Oriental Spercheidae and Hydrophilidae (Coleoptera:
Hydrophiloidea). Euroasian Entomological Journal 10 (3): 337-340.
Ryndevich S. K., Fikáček M. 2013. Faunistic and zoogeographic notes on hydrophiloid beetles from the Palaearctic
Sazhnev A. S. 2020. Additional data to the fauna of water beetles (Coleoptera) of Tajikistan. Entomological and
Schödl S. 1997 Taxonomic studies on the genus Enochrus (Coleoptera: Hydrophilidae). Entomological Problems
28 (1): 6166.
Schödl S. 1998. Taxonomic revision of Enochrus (Coleoptera: Hydrophilidae). I. The Enochrus bicolor species
complex. Entomological Problems 29: 111-127.
Short A. E. Z., Fikáček M. 2013. Molecular phylogeny, evolution and classification of Hydrophilidae (Coleoptera).
415
SPECIES OF THE SUBGENUS LUMETUS ZAITZEV (COLEOPTERA, HYDROPHILIDAE:
ENOCHRUS THOMSON) OF THE FAUNA OF RUSSIA AND ADJACENT COUNTRIES
S. V. Litovkin, A. S. Sazhnev, A. A. Prokin
Key words: Hydrophilidae, Enochrus, Russia, Palaearctic, fauna, key, new records.
SUMMARY
Eleven species of the subgenus Lumetus Zaitzev, 1908 of the genus Enochrus Thomson, 1859 are
recorded from Russia and adjacent countries. Enochrus ater (Kuwert, 1888) is recorded for the first
time from Ukraine and Kyrgyzstan; E. bicolor (Fabricius, 1792), from Kyrgyzstan; E. fuscipennis
(Thomson, 1884), from Georgia; E. segmentinotatus (Kuwert, 1888), from Ukraine. The structure of
the aedeagus and the method of male genitalia preparation are discussed. An annotated list and a key
to the species illustrated with original photographs and line drawings are given, the variability of the
species is characterised. Placement of E. puetzi Hebauer, 1995 in the subgenus Lumetus is confirmed.
A new synonymy is established: Enochrus ochropterus (Marsham, 1802) (= Philhydrus ferrugineus
Motschulsky, 1849, syn. n.). Lectotype and a paralectotype of Ph. ferrugineus are designated. The
taxonomic status of E. asiaticus (Kuwert, 1893), E. calabricus (Ferro, 1976), and E. sahlbergi (Fauvel,
1887) is discussed.
416