ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 100, 4, 2021
УДК 595.798 (591.151)
ФЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ОСЫ POLISTES
ALBELLUS GIORDANI SOIKA, 1976 (HYMENOPTERA, VESPIDAE)
© 2021 г. A. Ю. Косякова,1* А. В. Гилев,2** P. П. Ноймейер,3***
Л. Ю. Русина4****
1 Национальный парк «Мещера»
ул. Интернациональная, 111, Гусь-Хрустальный, Владимирская обл., 601501 Россия
*e-mail: ainsel@list.ru
2 Институт экологии растений и животных УрО РАН
ул. 8 Марта, 202, Екатеринбург, 620144 Россия
**e-mail: gilev@ipae.uran.ru
3 Цюрихский университет
Люгисландштрассе, 56, Цюрих, 8051 Швейцария
***e-mail: neumeyer.funk@icloud.com
4 Московский зоопарк
ул. Большая Грузинская, 1, Москва, 123242 Россия
****e-mail: lirusina@yandex.ru
Поступила в редакцию 2.07.2021 г.
После доработки 14.11.2021 г.
Принята к публикации 14.11.2021 г.
Работа посвящена описанию изменчивости окраски и меланинового рисунка социальной осы
Polistes albellus Giordani Soika, 1976 (Hymenoptera, Vespidae), распространенной от Западной
Европы (Франция) до Монголии, Китая и Приморского края России. Изучены 590 самок и 101 са-
мец. Фенотипическая изменчивость у самок P. albellus прослежена на мезоскутуме, 1-м и 2-м
тергитах и 4-6-м стернитах метасомы, а у самцов помимо этих структур на клипеусе и тазиках.
Анализ связи элементов рисунка с пространственно-временными особенностями гнездования
был проведен в двух локальных поселениях ос в Центральной Мещере (Рязанская обл., Россия).
Самки-основательницы разных морфотипов различаются по срокам начала гнездования, высоте
прикрепления гнезда и по склонности к образованию скоплений. Выборки самок-основательниц
из разных поселений фенотипически отличаются как от выборок рабочих, так и одна от другой.
В каждом локальном поселении выборки рабочих и будущих самок-основательниц фенотипи-
чески сходны.
Варианты рисунка мезоскутума связаны с влажностью местообитания: в сухих биотопах
у рабочих выше доля темных вариантов. Неоднородность условий среды может способствовать
увеличению изменчивости меланинового рисунка ос.
Ключевые слова: социальные осы, Vespidae, Polistes albellus, фенотипическая изменчивость,
меланиновый рисунок.
DOI: 10.31857/S0367144521040043
728
Осы-полисты широко распространены по всему миру и относятся к наиболее интен-
сивно изучаемым группам ресоциальных насекомых (Reeve, 1991; Röseler, 1991).
Черно-желтый рисунок ос-полистов чрезвычайно изменчив и дает простые и на-
дежные признаки в популяционно-экологических и поведенческих исследованиях
(Tibbets, Dale, 2004; Русина, 2009).
Одна из основных особенностей ос-полистов умеренных широт как объектов таких
исследований - наличие в их жизненном цикле трех периодов: 1) спаривания самцов и
будущих основательниц (конец лета - начало осени), 2) зимовки оплодотворенных
основательниц и 3) основания семей (последний длится весной немногим более ме-
сяца). Представляется интересным изучение такого цикла методами популяционной
морфологии (Яблоков, 1976; Яблоков, Ларина, 1985; Сергиевский, 1985, 1987 и др.):
поиск таких признаков меланинового рисунка (фенов), которые могут маркировать
в популяции ос-полистов функционально различающихся особей.
У ряда видов ос-полистов описана изменчивость меланинового рисунка и на этой
основе детально изучены фенотипическая структура локальных поселений ос и ее се-
зонная и многолетняя динамика, а также выбор брачного партнера будущими основа-
тельницами (Русина и др., 2004а, 2004б, 2006, 2007а, 2007б, 2007в, 2009; Русина, Ло-
патин, 2009; Русина, Орлова, 2009; Фирман, Русина, 2009). Выявлены циклические
изменения фенотипической структуры в меняющихся погодных условиях сезона (Ру-
сина и др., 2004а, 2007а; Rusina et al., 2011). Существенное влияние на фенотипиче-
ский состав популяции оказывает степень зараженности паразитами и паразитоидами
(Русина, 2009; Русина, Орлова, 2011). У самок-основательниц разных морфотипов
были выявлены отчетливые предпочтения способов основания гнезда и различия
в агрессивности (Русина и др., 2007б, 2007в, 2009). Так, самки Polistes dominula
(Christ, 1791) (Hymenoptera, Vespidae) с темными вариантами рисунка клипеуса пред-
почитали гнездиться в одиночку (гаплометроз), а в случае основания семьи группой
(плеометроз) становились доминантами, тогда как осы со светлыми вариантами чаще
предпочитали гнездиться совместно (Русина и др., 2007б). Некоторые авторы отме-
чают, что рисунок клипеуса у этого вида выполняет социальную функцию, позволяя
особям распознать ранг друг друга (Tibbets, 2002; Tibbets, Dale, 2004).
Широко распространенные виды отличаются большим разнообразием условий оби-
тания и часто демонстрируют высокий уровень изменчивости и внутривидовой фено-
типической дифференциации (Русина и др., 2008). Одним из перспективных объектов
популяционно-экологических исследований является оса P. albellus Giordani Soika,
1976, которая распространена от Западной Европы (Франция) до Приморского края
России, Монголии и Китая (Neumeyer et al., 2014, 2015).
Целями нашей работы были описание фенотипической изменчивости ос P. albellus
из локальных поселений в Центральной Мещере и изучение связи меланинового ри-
сунка самок-основательниц с пространственно-временными особенностями их гнез-
дования.
МАТЕРИАЛ И МЕТОДИКА
Основная часть материала по изменчивости окраски и меланинового рисунка P. albellus была
собрана в июне-августе 2019 г. на территории Центральной Мещеры (Россия, Рязанская обл.): из
двух локальных поселений в окрестностях дер. Уткино (табл. 1, рис. 1) и дер. Снохино (табл. 1,
729
Таблица 1. Места сбора самок и самцов Polistes albellus Giordani Soika
Выборки
Населенный пункт
Координаты
Число, экз.
Россия
Рязанская обл.
1-е локальное
Клепиковский р-н,
55°08′ с. ш., 40°32′ в. д.
110 ♀ (живые осы)
поселение
дер. Уткино
2-е локальное
дер. Снохино
55°08′ с. ш., 40°36′ в. д.
302 ♀, 101 ♂
поселение
(живые осы)
Одиночные
с. Гришино
55°04′6″ с. ш., 40°21′18″ в. д.
158 ♀ (живые осы)
гнезда
дер. Савино
55°06′47″ с. ш., 40°33′33″ в. д.
55°06′35″ с. ш., 40°33′32″ в. д.
дер. Коренево
55°09′20″ с. ш., 40°34′52″ в. д.
пос. Тума
55°09′30″ с. ш., 40°32′47″ в. д.
дер. Снохино
55°08′40″ с. ш., 40°36′07″ в. д.
55°08′40″ с. ш., 40°36′12″ в. д.
55°08′40″ с. ш., 40°36′03″ в. д.
Башкирия
Ишимбайский р-н,
53°36ʹ57″ с. ш., 56°34ʹ14″ в. д.
3 ♀ (по фотографиям
с. Зигановка
В. А. Валуева)
Благоварский р-н,
54°31ʹ41″ с. ш., 54°57ʹ28″ в. д.
дер. Удрякбаш
Буздякский р-н,
54°29ʹ57″ с. ш., 54°37ʹ14″ в. д.
дер. Новокили-
мово
Забайкальский
Дульдургинский р-н,
50°41ʹ50″ с. ш., 113°27ʹ6″ в. д.
1 ♀ (по фотографиям
край
Национальный
Р. Ю. Абашеева;
парк «Алханай»
Осы России. [Интер-
нет-ресурс])
Оренбургская
Сакмарский р-н,
52°3ʹ28″ с. ш. 55°25ʹ12,6″ в. д.
1 ♀ (Neumeyer et al.,
обл.
дер. Донское
2015)
Швейцария
Баума
47°23′00″ с. ш., 08°50′49″ в. д
8 ♀ (по фотографиям
Р. П. Ноймейера,
К. Доблер Гросс)
Казахстан
г. Зайсан
47°14′ с. ш., 84°56′
1 ♀ (Neumeyer et al.,
2015)
рис. 2). Особи из одиночно расположенных гнезд были описаны по материалу, собранному близ
с. Гришино, пос. Тума и деревень Савино, Снохино и Коренево. Кроме того, меланиновый рису-
нок ос был проанализирован по фотографиям, сделанным Р. П. Ноймейром и К. Доблер Гросс в
окрестностях населенного пункта Баума (Швейцария), В. А. Валуевым в Башкирии, Р. Ю. Аба-
шеевым в Забайкальском крае (Осы России. [Интернет-ресурс]) и опубликованным в статье
(Neumeyer et al., 2015). Всего изучены 590 самок и 101 самец.
При описании изменчивости меланинового рисунка на разных частях тела P. albellus исполь-
зовали его схемы, предложенные для других видов, в частности P. dominula и P. nimpha (Christ,
730
Рис. 1. Влажный злаково-разнотравный луг, окр. дер. Уткино (55°08′ N, 40°32′ E, 01.VII.2019).
Фотография А. Ю. Косяковой.
1791) (Русина и др., 2004б, 2006; Русина, 2009), выделяя ряды изменений по усилению степени
меланизации (Филиппов, 1961).
Сезонные изменения фенотипической структуры локальных поселений были изучены на тер-
ритории Центральной Мещеры.
Локальные поселения ос были обнаружены на двух участках, которые различались структу-
рой и увлажненностью почвы: 1-й - дерново-подзолистая с перегнойно-глеевыми пятнами под
влажными злаково-разнотравными лугами (преимущественно с манником большим Glyceria
maxima (Harm.) Holmb); 2-й - хорошо дренированная, сложенная песками с отдельными просло-
ями суглинков, дерново-слабоподзолистая почва под разнотравно-вейниковой залежью, зараста-
ющей соснами Pinus sylvestris L. (Анненская и др., 1983).
Были описаны пространственные (высота прикрепления сота, вид и высота растения, рассто-
яние до ближайшего гнезда и скученность гнезд на площадке 10 × 10 м) и демографические
(число ячей, личинок IV и V возрастов, куколок, рабочих) параметры каждого гнезда и морфоло-
гические особенности (меланиновый рисунок) у отловленных в них и окольцованных 76 са-
мок-основательниц (рис. 3). Демографические замеры для всех гнезд были произведены в один
день для каждого из поселений в период до выхода рабочих. В 1-м поселении (04.VI.2019) были
описаны меланиновые рисунки 36 самок-основательниц из 35 гаплометротичных семей и одной
плеометротичной, во 2-м (08.VI.2019) описаны 40 самок из 40 гаплометротичных гнезд.
Для каждого поселения определяли коэффициент корреляции морфологических параметров
самок-основательниц и размеров их семей, а также пространственных характеристик гнезд. Раз-
731
Рис. 2. Залежь, зарастающая соснами (Pinus sylvestris L.), окр. дер. Снохино.
Фотография А. Ю. Косяковой (55°08′ N, 40°36′ E, 28.IX.2019).
Рис. 3. Polistes albellus Giordani Soika.
Справа: самка-основательница (обозначена красной стрелкой) и рабочие в Центральной Мещере (окр. дер.
Коренево, 55°08′44ʺ N, 40°36′00ʺ E, 26.VI.2019). Фотография А. Ю. Косяковой. Слева: будущие самки-
основательницы на гнезде в Швейцарии (ZH, Bauma: Joggelisböl, 47°23′00ʺ N, 08°50′49ʺ E, 07.IX.2012).
Фотография Р. П. Ноймейера
732
меры семей использовали для косвенной оценки их относительного возраста и, следовательно,
сроков начала гнездования: более крупные семьи считали основанными раньше. Дополнительно
рассчитывали корреляцию между вариантами меланинового рисунка самок-основательниц и
пространственными характеристиками их семей.
Всего в первом поселении за время исследований (май-август 2019) было обнаружено 54 гнез-
да, во втором - 168; 19 гнезд располагались поодиночке. Повторные осмотры гнезд проводили
1 или 2 раза в неделю. Регистрировали состав семьи и ее выживание (рис. 4); кольцевали моло-
дых, не меченных ранее насекомых (рабочих, самцов и будущих основательниц), описывали их
меланиновые рисунки, а также отмечали присутствие паразитов и паразитоидов. Клеща
Sphexicozela connivens Mahunka, 1970 (Acari, Astigmata: Winterschmidtiidae) и осиного веерокры-
ла Xenos vesparum Rossi, 1793 (Strepsiptera, Stylopidae) искали, осматривая личинок, поздних
рабочих, самцов и будущих основательниц.
При картировании гнезд фиксировали следы заражения расплода P. albellus паразитоидами.
Так, заражение наездником Latibulus argiolus (Rossi, 1790) (Hymenoptera: Ichneumonidae) опреде-
ляли по присутствию в ячейке гнезда овально скошенных остатков личиночной кутикулы свет-
ло-желтого или светло-оранжевого цвета и/или коконов (Makino, 1983), а Elasmus schmitti
Ruschka, 1920 (Hymenoptera, Eulophidae) - по наличию крышечки темно-серого цвета, которую
личинки этого грегарного паразитоида сообща формируют из мекониев перед окукливанием
(Gumovsky et al., 2007).
Для множественного сравнения независимых выборок параметров гнезд использовали тест
Крускала-Уоллиса (критерий H) и тест Данна (критерий Q). Попарное сравнение двух независи-
мых выборок проводили с помощью теста Манна-Уитни (критерий U) (Гланц, 1999).
Связь особенностей меланиновых рисунков самок-основательниц со спецификой простран-
ственных и/или временных характеристик их гнездования оценивали с помощью корреляции
Спирмена (rs).
Для оценки достоверности различий между выборками по частотам встречаемости вариантов
окраски использовали критерий Хи-квадрат (χ²). Сравнение производилось как между локаль-
ными поселениями на сходных стадиях развития, так и в ходе сезонного развития семей каждого
локального поселения. Однако универсальный критерий сравнения χ² недостаточно чувствите-
лен к редким вариантам (Животовский, 1991), поэтому для оценки фенотипических различий
локальных поселений были дополнительно использованы специальные меры расстояний (дис-
танций), такие как расстояния Кавалли-Сфорцы, которые учитывают редкие признаки (Cavalli-
Рис. 4. Самцы (обозначены синими стрелками) и рабочие Polistes albellus Giordani Soika
на разоренном птицами гнезде (окр. дер. Коренево, 55°08′43ʺ N, 40°36′07ʺ E, 28.VI.2019).
Фотографии А. Ю. Косяковой
733
Sforza, Edvards, 1967; Животовский, 1991; Вейр, 1995). Для наглядного отображения различий
между выборками матрицы расстояний были обработаны методами многомерного неметриче-
ского шкалирования.
Все расчеты выполнены в программах Microsoft Excel 2019, Statistica v.10.0 (StatSoft, Inc.
1984-2020), а также в программе Биостатистика (Primer Of Biostatistics) Biostatistica v. 4.03
(Гланц, 1999). Для анализа вида исследуемых параметров использовали критерий Шапиро-Уил-
ка. При нормальном распределении признаков выборку представляли в виде среднего
M ± среднеквадратическоe отклонение SD, а в случае ненормального распределения - в виде
медианы Me [25; 75] [min; max] - медианы, 1-го и 3-го квартилей, минимального и максимально-
го значений (Гланц, 1999).
РЕЗУЛЬТАТЫ
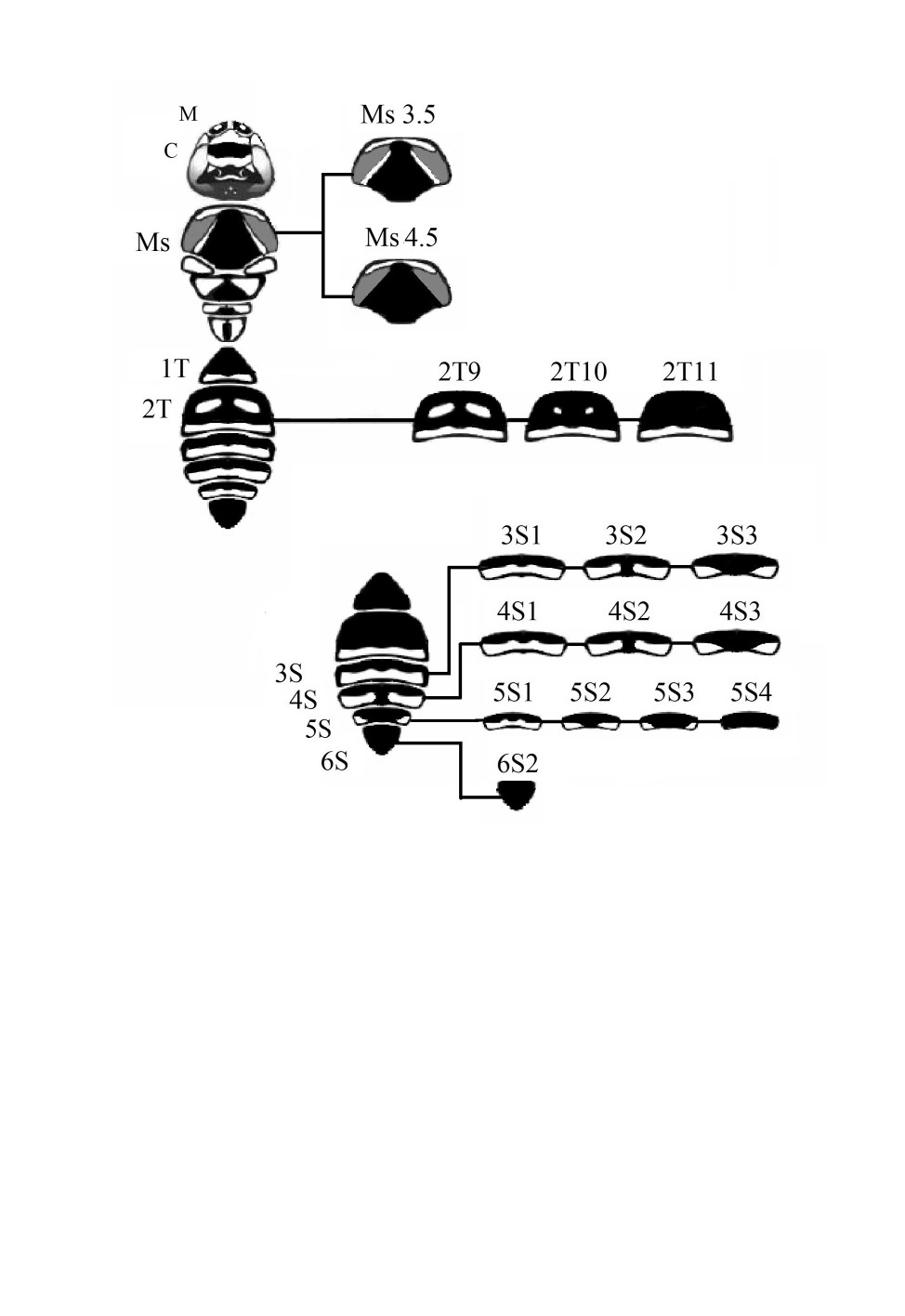
Меланиновый рисунок самок Polistes albellus
Общая картина изменчивости меланинового рисунка самок и самцов P. albellus
представлена на схемах (рис. 5, 6). Практически на всех частях тела обнаруженные
варианты рисунка представляли собой только часть спектра изменчивости, в целом
характерного для изученных в этом отношении ос-полистов (Русина, 2009). По резуль-
татам этих исследований для некоторых частей тела удалось построить схемы, на ко-
торых представлены полный спектр вариантов и направления развития рисунка
(рис. 7-13).
Изменчивость рисунка P. albellus сходна с таковой у P. dominula и P. nimphа, но есть
и ряд особенностей. У самок перечисленных видов изменчивость проявляется на
жвалах, клипеусе, груди, 1-м и 2-м тергитах и 4-6-м стернитах метасомы (см. рис. 5).
Рисунок на жвалах полистов характеризуется наличием (М1) или отсутствием
желтых пятен (М2). Жвалы P. albellus всегда были пигментированы лишь частично.
Варианты рисунка на клипеусе связаны с появлением пятен пигмента и их последу-
ющим слиянием (рис. 7). Клипеус P. albellus имел только один сильно меланизиро-
ванный вариант рисунка с широкой центральной поперечной полосой (С5).
На груди имеются парные непигментированные участки (просветы) вдоль промезо-
нотального шва и в центре мезоскутума. В связи с этим рисунок мезоскутума описы-
вается по комбинативному типу (двумя цифрами, разделенными точкой; рис. 8).
Начальная цифра отображает вариант рисунка первой области просветов, последняя -
второй. У ос-полистов для просветов вдоль шва выделены 4 варианта, которые отобра-
жают степени их развития: просветы могут достигать переднего края, передняя часть
пронотума остается немеланизированной (Мs1); просветы достигают (Ms2) или не до-
стигают (Ms3) края мезоскутума; просветы отсутствуют (Мs4).
Просветы в центре мезоскутума описываются пятью вариантами в порядке возрас-
тания степени меланизации: просветы в виде больших удлиненных запятых (Ms1-4.1),
небольших запятых (Ms1-4.2), треугольничков (Ms1-4.3), точек (Ms1-4.4), просветов
нет (Ms1-4.5). У самок P. albellus просветы в центре мезоскутума не обнаружены,
а вдоль промезонотального шва отмечены 2 варианта (Ms3.5 и Ms4.5; см. рис. 5).
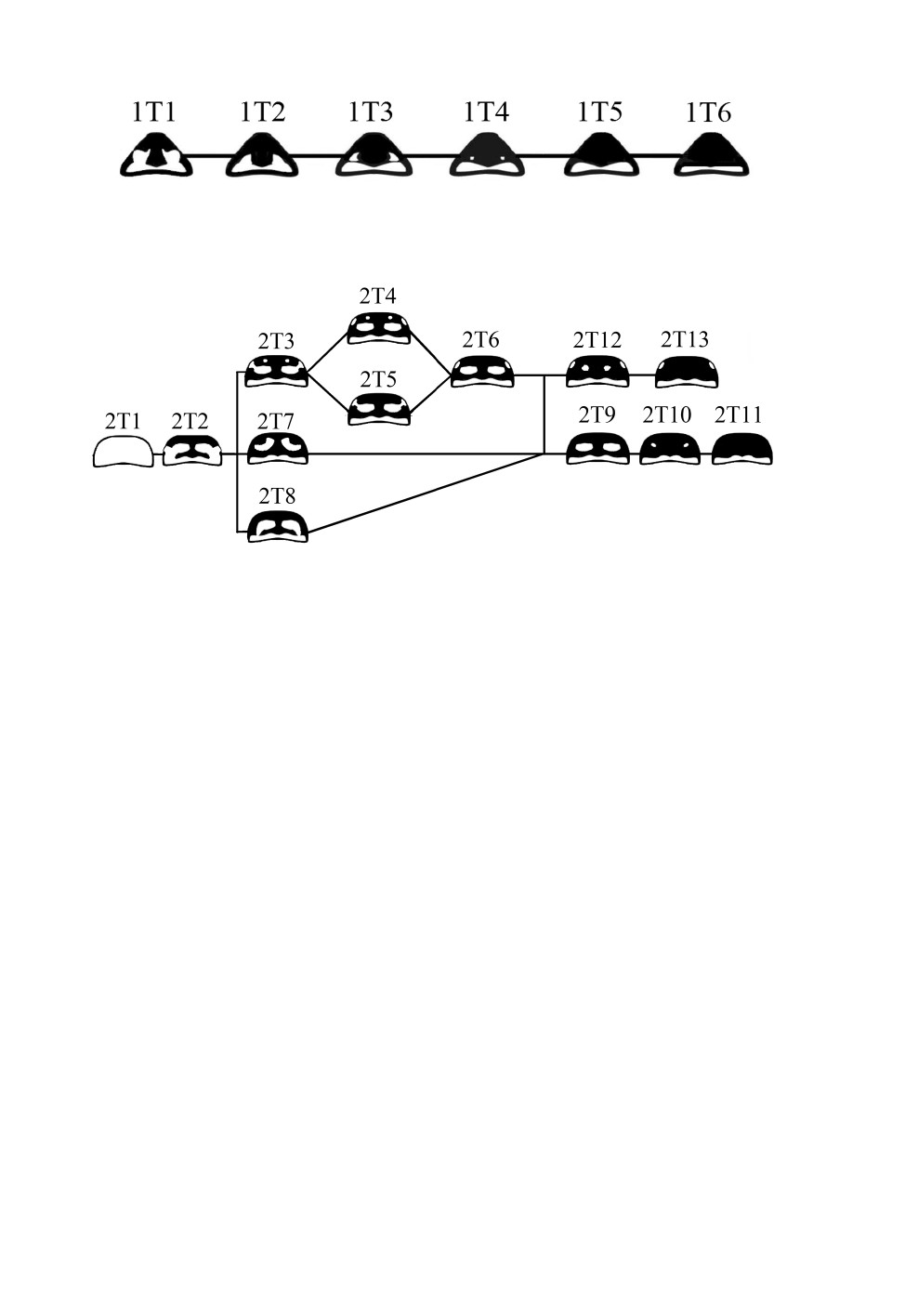
Варианты рисунка 1-го тергита метасомы у ос-полистов выделяют по степени мела-
низации его дорсальной части, а дистальный край всегда с желтой полосой (рис. 9).
У самых светлых вариантов просветы сложной формы, сливающиеся с дистальной
734
Рис. 5. Варианты рисунка самки Polistes albellus Giordani Soika.
M - жвалы, С - клипеус, Ms - мезоскутум, 1Т - 1-й тергит метасомы, 2Т - 2-й тергит метасомы,
3S-6S - 3-6-й стерниты метасомы.
желтой полосой. Дальнейшее развитие рисунка на этой части тела приводит к обособ-
лению просветов и их последующему уменьшению. Для P. albellus отмечен новый ва-
риант рисунка 1-го тергита, не встречающийся у других видов. Изменения рисунка
затрагивают только дистальную желтую полосу, которая становится уже, при этом
пятна отсутствуют (1Т6).
Варианты рисунка на 2-м тергите метасомы выделены в зависимости от наличия
парных просветов, их размеров и формы (рис. 10). У наиболее светлых вариантов про-
светы или сливаются с нижней желтой полосой, или не сливаются, но имеют при этом
большие размеры и сложную форму. У P. albellus на 2-м тергите обособленные цен-
тральные просветы отмечены в виде больших пятен (2Т9), точек (2Т10), или они от-
сутствовали (2Т11; см. рис. 5).
735
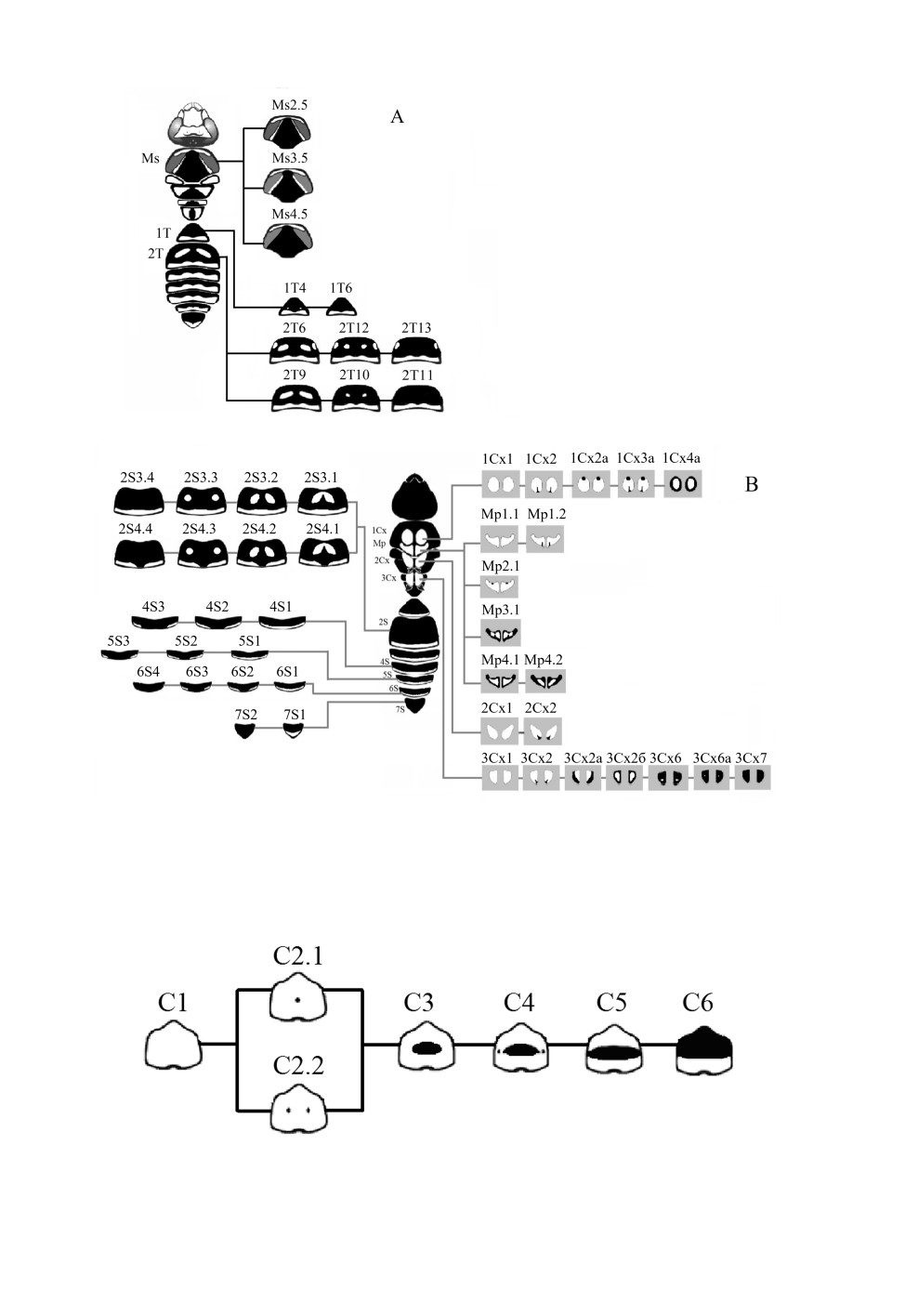
Рис. 6. Варианты рисунка самца Polistes albellus Giordani Soika. А - вид сверху, B - вид снизу.
2S - 2-й стернит метасомы; 1Cx-3Cx - передние, средние и задние тазики; Mp - мезоплевры.
Остальные обозначения как на рис. 5.
Рис. 7. Варианты рисунка клипеуса самок ос-полистов.
736
Рис. 8. Варианты рисунка мезоскутума ос-полистов.
На стернитах метасомы полистов подвержена изменчивости желтая поперечная пе-
ревязь (см. рис. 5). Она может быть сплошной или разделенной языком пигмента на
две части. У некоторых видов 6-й стернит может быть полностью черным или иметь
небольшой просвет. У P. albellus изменчивость рисунка отмечена на 3-5-м стернитах.
На 3-м и 4-м стернитах были выделены по 3 варианта рисунка: сплошная желтая по-
лоса (S1), а также перевязь, разделенная пигментом на 2 части, которая могла быть
более (3S2 и 4S2) или менее (3S3 и 4S3) выражена. На 5-м стерните помимо ранее пе-
речисленных вариантов (5S1, 5S2, 5S3) отмечен и вариант без желтой перевязи (5S4).
На 6-м стерните самок P. albellus выявлен только один вариант с полностью черной
окраской (6S2).
Меланиновый рисунок самцов Polistes albellus
Самцы P. albellus гораздо более изменчивы, чем самки, и в целом окрашены заметно
светлее. Изменчивость рисунка у них выявлена на клипеусе, мезоскутуме, тазиках
всех пар ног, 1-м и 2-м тергитах и на 2-м и 4-7-м стернитах метасомы (см. рис. 6).
Как и у других видов ос-полистов, у самцов P. albellus изменчивость рисунка груди
сходна с таковой у самок (рис. 8). У большинства описанных нами самцов встречались
те же варианты рисунка мезоскутума, что и у самок: с просветами вдоль промезоно-
тального шва (Ms3.5) или без них (Ms4.5). У единичных особей отмечен вариант ри-
сунка Ms2.5, при котором просветы вдоль шва доходят до переднего края мезоскутума,
но при этом отсутствуют парные центральные просветы.
737
Рис. 9. Варианты рисунка 1-го тергита ос-полистов.
Рис. 10. Варианты рисунка 2-го тергита ос-полистов.
Рисунок мезоплевр самцов полистов очень изменчив (рис. 11) и включает в себя
практически полный спектр от полностью желтого до самого темного варианта без
просветов. При этом пятна пигмента могут возникать в разных местах и иметь раз-
личную форму.
На мезоплеврах P. albellus отмечено шесть вариантов рисунка, три из них встреча-
ются и у других видов полистов: самый светлый вариант (Mp1.1), а также вариант
с очагом пигмента сзади (Mp1.2) и пигментированный вариант c центральным про-
светом (Мp4.2). Помимо этого, у P. albellus встречаются три не описанных ранее вари-
анта. В первом из них точка пигмента появляется у верхнего края (Mp2.1). Во втором -
пигментация захватывает весь внешний край мезоплевры (Mp4.1). Третий вариант
Mp3.3 сходен с предыдущим и отличается тем, что внутренняя часть полосы пигмента
имеет два выступа.
Рисунок на тергитах метасомы самцов в целом развивается по тому же плану, что и
у самок. У P. albellus 1-й тергит имеет два варианта рисунка. Наиболее часто встре-
чался вариант с желтой перевязью по краю стернита, отмеченный и у самок этого вида
(1Т6). Помимо этого, у самцов есть и вариант с парными просветами в виде точек
(1Т4).
Для 2-го тергита P. albellus отмечены 6 вариантов рисунка (см. рис. 10). Три из них
сходны с теми, что встречаются и у самок этого вида: с одной парой центральных
пятен (2Т9), которые затем уменьшаются до точек (2Т10) и впоследствии исчезают
полностью (2Т11). Кроме этого, отмечен один более светлый вариант с двумя парами
пятен (2Т6). Дальнейшее развитие этого рисунка у P. albellus идет по пути умень-
738
Рис. 11. Варианты рисунка мезоплевр самцов ос-полистов.
шения больших центральных пятен до точек (2Т12) и их полного исчезновения, при
этом боковые пятна сохраняются (2Т13). Последние 2 варианта рисунка не встреча-
ются у P. dominula и P. nimphа.
Основные преобразованием рисунка на 2-м стерните метасомы полистов связаны с
изменением центральных просветов (рис. 12). Так, у P. nimpha отмечены самый
светлый вариант и вариант с парными пигментированными пятнами, не встречаю-
щиеся у P. dominula. При этом число вариантов рисунка у P. dominula достигает шести,
в то время как у P. nimpha отмечены только два.
Значительная изменчивость 2-го стернита у самцов P. albellus связана с наличием
или отсутствием просветов, их числом, размерами и формой. Помимо этого, изменчи-
вость затрагивает желтую перевязь, идущую вдоль дистального края стернита,
739
Рис. 12. Варианты рисунка 2-го стернита ос-полистов.
которая может быть цельной или разделяться пигментом на 2 части. Многообразие
рисунков 2-го стернита можно разделить на две группы.
Первая группа рисунков имеет цельную желтую перевязь. Для самого светлого ва-
рианта характерно наличие двух крупных сливающихся пятен, расположенных
в центре (2S3.1), впоследствии они разделяются на 2 обособленных пятна (2S3.2),
которые затем уменьшаются в размерах (2S3.3) и полностью исчезают (2S3.4).
Рисунки второй группы повторяют в общих чертах варианты первой с той лишь раз-
ницей, что желтая перевязь разделена в центре пигментом на две части (2S4.1-2S4.4).
Варианты меланинового рисунка на 4-7-м стернитах метасомы P. albellus различа-
ются по особенностям желтой перевязи (S1-S2-S3), при этом 6-й стернит помимо
вышеперечисленных вариантов может быть полностью пигментирован (S4). По-
следний, 7-й стернит имеет небольшой желтый просвет (7S1) или не имеет его (7S2).
Как и у других видов ос-полистов, изменчивость рисунка всех пар тазиков самцов
P. albellus связана с уменьшением размеров просветов и усилением пигментации в
любом месте (рис. 13). На всех парах тазиков встречаются как полностью немеланизи-
рованные варианты (1Сх1, 2Сх1, 3Сх1), так и варианты с пигментированным участком
у нижнего края (1Сх2, 2Сх2, 3Сх2). При этом на передних и задних тазиках встреча-
ются варианты рисунков, не отмеченные ранее у самцов P. dominula и P. nimpha. Так,
на передних тазиках помимо рисунков 1Сх1 и 1Сх2 встречается вариант с пятном,
расположенным сверху (1Cх2а). В другом варианте помимо верхнего округлого пятна
пигмента возникает 2-й очаг пигмента, находящийся снизу (1Сх3а). Отмечен вариант
с полосами пигмента по внешним краям тазиков и непигментированными централь-
ными их частями (1Сх4а).
Для задних тазиков отмечены варианты 3Сх1, 3Сх2 и, кроме того, ранее не встречав-
шийся у других видов рисунок с полосой пигмента, идущей по внешнему краю
(3Сх2а). Обнаружен и более пигментированный вариант, у которого полоса захваты-
вает весь край тазика (3Сх2б): усиление меланизации в этом случае приводит к вари-
анту с небольшим центральным просветом (3Сх6). Немеланизированные просветы у
P. albellus сохраняются в виде маленьких точек (3Сх6а). У самого темного варианта
задние тазики полностью пигментированы (3Сх7).
740
Рис. 13. Варианты рисунка тазиков самцов ос-полистов.
Места гнездования и фенология развития семей Polistes albellus
в Центральной Мещере
Перезимовавшие самки-основательницы P. albellus в Центральной Мещере весной
сооружали гнезда на растениях (N = 222, табл. 2). Большая часть гнезд была обнару-
жена в двух поселениях, при этом часть их располагалась очень близко друг к другу,
а другие находились в небольших разреженных группах. Кроме того, были обнару-
жены одиночные гнезда за пределами поселений.
В первом поселении самки P. albellus чаще всего прикрепляли свои гнезда к сухим
прошлогодним стеблям трав (77.8 %, 42 из 54), реже - к стеблям невысоких вегетиру-
ющих молодых деревьев и кустарников.
Во втором поселении большинство гнезд (80.95 %, 136 из 168) было расположено на
подросте сосны обыкновенной Pinus sylvestris (в среднем высотой 0.62 ± 0.30 м);
реже - на лиственных кустарниках и деревьях и на стеблях травянистых растений.
Перезимовавшие самки в мае основывали свои гнезда поодиночке. В одном посе-
лении 2 июня наблюдали, как самка шершня Vespa crabro L. (Hymenoptera, Vespidae)
села на гнездо P. albellus, вытащила личинку и стала ее пережевывать; она прилетала
в гнездо в этот день несколько раз и сильно повредила его. Самка-хозяйка, имевшая
741
Таблица 2. Видовой состав растений, используемых для гнездования Polistes albellus Giordani
Soika (N - число гнезд)
Встречаемость, %
Вид растения
1
2
3
4
(N = 54)
(N = 168)
(N = 19)
(N = 241)
Сем. Pinaceae
Pinus sylvestris L.
-
79.17
42.11
58.5
Сем. Betulaceae
Betula pendula Roth
-
1.19
10.53
1.66
Сем. Compositae
Tanacetum vulgare L.
3.7
0.59
-
1.24
Сем. Elaeagnaceae
Hippoophaë rhamnoides L.
-
-
10.53
0.83
Сем. Fagaceae
Quercus robur L.
-
-
5.26
0.42
Сем. Gramineae
Calamagrostis epigeios (L.) Roth
18.52
0.59
-
4.56
Dactylis glomerata L.
-
1.79
-
1.24
Сем. Onagraceae
Epilobium sp.
-
-
5.26
0.42
Сем. Polygonaceae
Rumex confertus Willd.
16.67
2.38
-
5.39
Сем. Ranunculaceae
Ranunculus acris L.
1.85
-
-
0.42
Сем. Rhamnaceae
Frangula alnus Mill.
-
2.98
-
2.07
Сем. Rosaceae
Amelanchier spicata (Lam.) C. Koch
1.85
-
-
0.42
Malus domestica Borkh.
7.41
5.37
15.79
6.64
Rosa majalis Herrm.
1.85
-
5.26
0.83
Rubus idaeus L.
1.85
-
-
0.42
742
Таблица 2 (продолжение)
Встречаемость, %
Вид растения
1
2
3
4
(N = 54)
(N = 168)
(N = 19)
(N = 241)
Сем. Rubiaceae
Galium mollugo L.
-
0.59
-
0.42
Сем. Salicaceae
Populus alba L.
-
0.59
-
0.42
Salix cinerea L.
9.26
3.57
5.26
4.98
Сем. Sapindaceae
Acer negundo L.
1.85
1.19
-
1.24
Не определены
35.19
-
-
7.88
П р и м е ч а н и е. 1 - первое поселение, 2 - второе поселение, 3 - одиночные гнезда, 4 - всего.
индивидуальное кольцо-метку, переселилась на следующий день в соседнее гнездо,
расположенное в 8 м. Агрессивных взаимодействий между самками не было отме-
чено, и в дальнейшем они обитали в гнезде совместно.
Сравнительный анализ пространственно-временных характеристик гнезд выявил
статистически значимые различия в высоте прикрепления гнезда между поселениями.
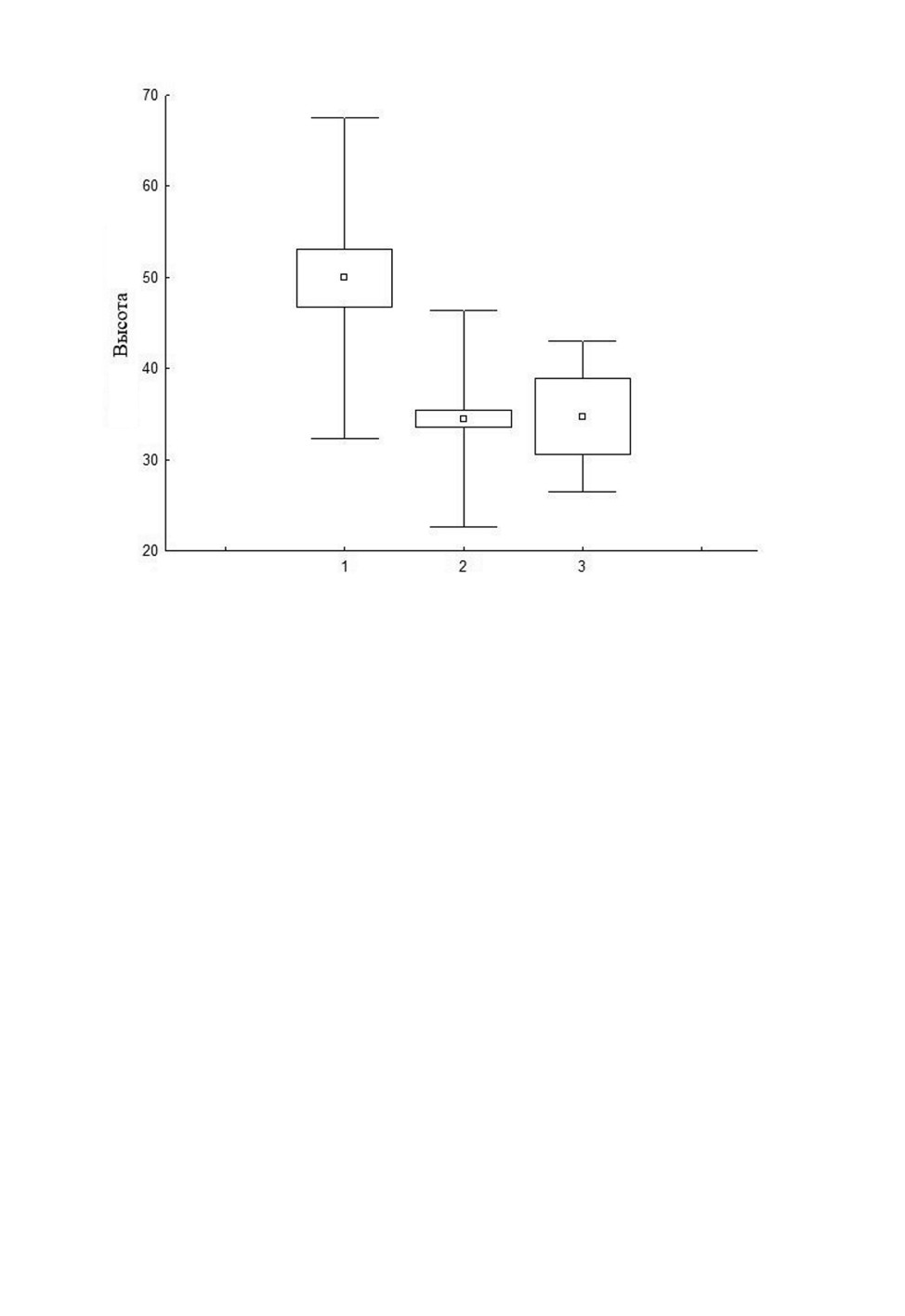
Так, самки-основательницы в первом поселении, расположенном в более увлаж-
ненном биотопе, прикрепляли гнезда выше, чем во втором поселении и в одиночных
гнездах (рис. 14).
Выход первых рабочих отмечен в первой декаде июня, в среднем 10 июня ± 3 дня
(N гнезд = 39). Самцы в отдельных гнездах были отмечены 15 июня, массовый выход
самцов, в среднем, 9 июля ± 6 дней. Будущие основательницы появлялись после вы-
хода самцов, в последней декаде июля. Они отмечены лишь для части гнезд второго
поселения и для одиночных гнезд, поскольку все семьи первого поселения погибли
из-за выпаса стада коров.
Различий между поселениями по срокам выхода рабочих, самцов и будущих основа-
тельниц в 2019 г. обнаружено не было.
Паразиты клещ Sphexicozela connivens и осиный веерокрыл Xenos vesparum на осмо-
тренных имаго (N = 671) не найдены. В гнездах также не были найдены крышечки
паразитоида E. schmitti.
В 10.81 % (24 из 222) гнезд обнаружены следы пребывания паразитоида Latibulus
argiolus.
743
Рис. 14. Первоначальная высота расположения гнезд (см) Polistes albellus Giordani Soika
в Центральной Мещере.
1 - первое локальное поселение, 2 - второе локальное поселение, 3 - одиночные гнезда.
Результаты общего теста Краскела-Уоллиса (H = 27.9; p < 0.001) и попарное сравнение по критерию Данна
(1 и 2: Q = 27.7; p < 0.001; 1 и 3: Q = 4.3; p < 0.05; 2 и 3: Q = 0.02; p > 0.05 соответственно).
В трех гнездах (из 54; 5.36 %) первого поселения обнаружена лишь 1-я генерация
паразитоида. Во втором поселении доля семей, зараженных 1-й генерацией парази-
тоида, составила 3.57 % (6 из 168 первичных гнезд, заложенных основательницами);
зараженных 2-й генерацией - 41.4 % (12 из 29 выживших). Две семьи были заражены
обеими генерациями L. argiolus. Наблюдалось заражение расплода вторичного (пере-
заложенного рабочими после разрушения первичного) гнезда небольших размеров
(38 ячей).
Среди 8 одиночных выживших гнезд было обнаружено одно со следами пребывания
1-й генерации L. argiolus, а в 3 гнездах - его 2-й генерации.
В каждом из поселений зараженные и не зараженные L. argiolus семьи были сходны
по размерам.
Образование локальных поселений самками-основательницами Polistes albellus
разных морфотипов в Центральной Мещере
Фенотипические различия между самками-основательницами двух поселений были
выявлены в частотах вариантов рисунка 3-го и 5-го стернитов метасомы: χ² = 15.85,
р < 0.001 и χ² = 13.52, р < 0.01 соответственно (табл. 3, 4). В обоих поселениях наблюда-
744
Таблица 3. Встречаемость вариантов рисунка у самок-основательниц, рабочих и будущих
основательниц Polistes albellus Giordani Soika в локальных поселениях Центральной Мещеры,
%
Вариант
1
2
3
4
5
6
7
Часть тела
рисунка
(N = 36)
(N = 74)
(N = 40)
(N = 248)
(N = 14)
(N = 106)
(N = 48)
Мезоскутум
Ms 3.5
80.56
72.97
70
68.14
78.57
80.19
89.58
Ms 4.5
19.44
27.03
30
31.86
21.43
19.81
10.42
2-й тергит
2Т9
66.67
36.49
55
33.47
0
24.53
14.58
2Т10
19.44
36.49
35
36.69
71.43
38.68
31.25
2Т11
13.89
27.02
10
29.84
28.57
36.79
54.17
3-й стернит
3S1
2.78
1.35
5
0
0
0
0
3S2
38.89
27.03
2.5
25
0
8.49
12.5
3S3
58.33
71.62
92.5
75
100
91.51
87.5
4-й стернит
4S1
2.78
1.35
0
0
0
0
0
4S2
16.67
0
2.5
0.81
0
0.94
2.08
4S3
80.55
98.65
97.5
99.19
100
99.06
97.92
5-й стернит
5S2
5.56
0
2.5
0
0
0
0
5S3
66.66
8.11
27.5
6.45
0
1.89
14.58
5S4
27.78
91.89
70
93.55
100
98.11
85.42
П р и м е ч а н и е. 1 - самки-основательницы первого поселения, 2 - рабочие первого поселения,
3 - самки-основательницы второго поселения, 4 - рабочие второго поселения, 5 - будущие самки-
основательницы второго поселения, 6 - рабочие из одиночных гнезд, 7 - будущие самки-основательницы
из одиночных гнезд. N - число самок.
Таблица 4. Различия во встречаемости разных вариантов рисунка у самок-основательниц и
рабочих Polistes albellus Giordani Soika в поселениях Центральной Мещеры по критерию χ2
Вариант
1 и 2
3 и 4
1 и 3
2 и 4
рисунка
Мs
0.75
0.06
1.13
0.62
2Т
8.87
9.41**
2.33
0.31
3S
2.01
21.73***
15.85***
3.53
4S
13.46**
-
5.84
3.95
5S
48.60***
24.59***
13.52**
-
П р и м е ч а н и е. 1 - самки-основательницы первого поселения, 2 - рабочие п ервого поселения, 3 -
самки-основательницы второго поселения, 4 - рабочие второго поселения. ** - p < 0.01, *** - p < 0.001.
745
лось заметное преобладание самок с фенотипом Ms3.5-1T9 (58.34 % и 35 % соответ-
ственно).
Вместе с тем, в каждом поселении обнаружен неслучайный выбор мест и сроков
начала гнездования самками-основательницами разных морфотипов. Размеры гнезд и
семей в день проведения измерений в среднем составили: в первом поселении по
числу ячей (здесь и далее приводятся Me [25; 75] [min; max]) - 56 [49; 61] [26; 72], по
числу куколок - 6 [4; 8] [0; 17], по числу личинок IV возраста - 2 [1; 3] [0; 5], по числу
личинок V возраста - 2 [0; 3] [0; 7]; во втором поселении по числу ячей - 47 [28; 58]
[4; 93], по числу куколок - 0 [0; 9] [0; 14], по числу личинок IV возраста - 0 [0; 2]
[0; 6], по числу личинок V возраста - 0 [0; 0] [0; 5].
Так, для самок-основательниц первого поселения было показано, что чем светлее
был 5-й стернит, тем больше ячей имели их гнезда: rs = ˗0.39, p < 0.05.
Кроме того, чем более меланизированным был 4-й стернит самок, тем выше они
прикрепляли свои гнезда: rs = 0.42; p < 0.05. Во втором поселении выявлена связь
между степенью меланизации 3-го стернита и скученностью гнезд (rs = ˗0.41;
p < 0.05): чем светлее был окрашен стернит самки-основательницы, тем более они
были склонны гнездиться скоплениями.
Связи между степенью меланизации покровов основательниц и зараженностью рас-
плода Latibulus аrgiolus не найдено ни в одном из локальных поселений.
Предпочтений в выборе растений и ориентации сота самками разных морфотипов
обнаружено не было.
Сезонная динамика фенооблика Polistes albellus в Центральной Мещере
Основательницы и рабочие обоих поселений имели единственные, одинаковые для
всех самок варианты рисунка жвал (M1), клипеуса (C5), 1-го тергита (1T6) и 6-го стер-
нита (6S2), но различались встречаемостью вариантов рисунка мезоскутума, 2-го тер-
гита и 3-5-го стернитов метасомы (табл. 3).
В первом поселении частота встречаемости вариантов рисунка 2T у основательниц
и рабочих была близкой. У обеих каст наиболее часто отмечались самые светлые вари-
анты рисунка. Кроме того, эти касты оказались сходными по частотам встречаемости
вариантов рисунка 3S, который у большинства самок был сильно пигментирован.
Во втором поселении по частоте встречаемости вариантов рисунка мезоскутума ос-
новательницы были сходны с рабочими (у обеих каст преобладал вариант с просве-
тами вдоль промезонотального шва - Ms3.5). Кроме того, сходство отмечено для 4-го
стернита, который чаще всего имел темные варианты рисунка.
Основательницы и рабочие 1-го поселения статистически значимо различались по
частоте встречаемости вариантов рисунка 4-го и 5-го стернитов метасомы (табл. 4): у
основательниц, в отличие от рабочих, был отмечен вариант 4S с выраженными про-
светами (4S2), при этом по частоте встречаемости в обеих группах преобладал самый
темный вариант (4S3); 5-й стернит основательниц чаще всего имел просветы, в то
время как у подавляющего большинства рабочих он был полностью пигментирован.
Помимо этого, у рабочих не было отмечено самого светлого варианта рисунка 5S2.
Мезоскутум обеих рассматриваемых групп самок чаще всего имел самый светлый ва-
746
риант Ms3.5, однако у рабочих его доля оказалась заметно ниже, чем у основательниц.
В целом по всем рассматриваемым признакам рабочие отличались от основательниц
преобладанием более меланизированных форм.
Основательницы второго поселения статистически значимо отличались от рабочих
по частоте встречаемости вариантов рисунка 2Т, 3S и 5S (см. табл. 4). У основательниц
преобладал самый светлый вариант 2-го тергита с крупными пятнами (2Т9), а у ра-
бочих - более темный с небольшими просветами (2Т10). Во втором поселении только
у рабочих встречался самый светлый вариант рисунка 3S со сплошной перевязью
(3S1). Кроме того, у основательниц чаще, чем у рабочих, отмечались варианты ри-
сунка 5-го стернита с просветами. У рабочих совсем не встречались самые светлые
варианты рисунков 4S и 5S (4S1, 5S2).
Обнаружены различия между двумя скоплениями гнезд в Центральной Мещере по
частотам встречаемости вариантов меланинового рисунка у самок-основательниц и
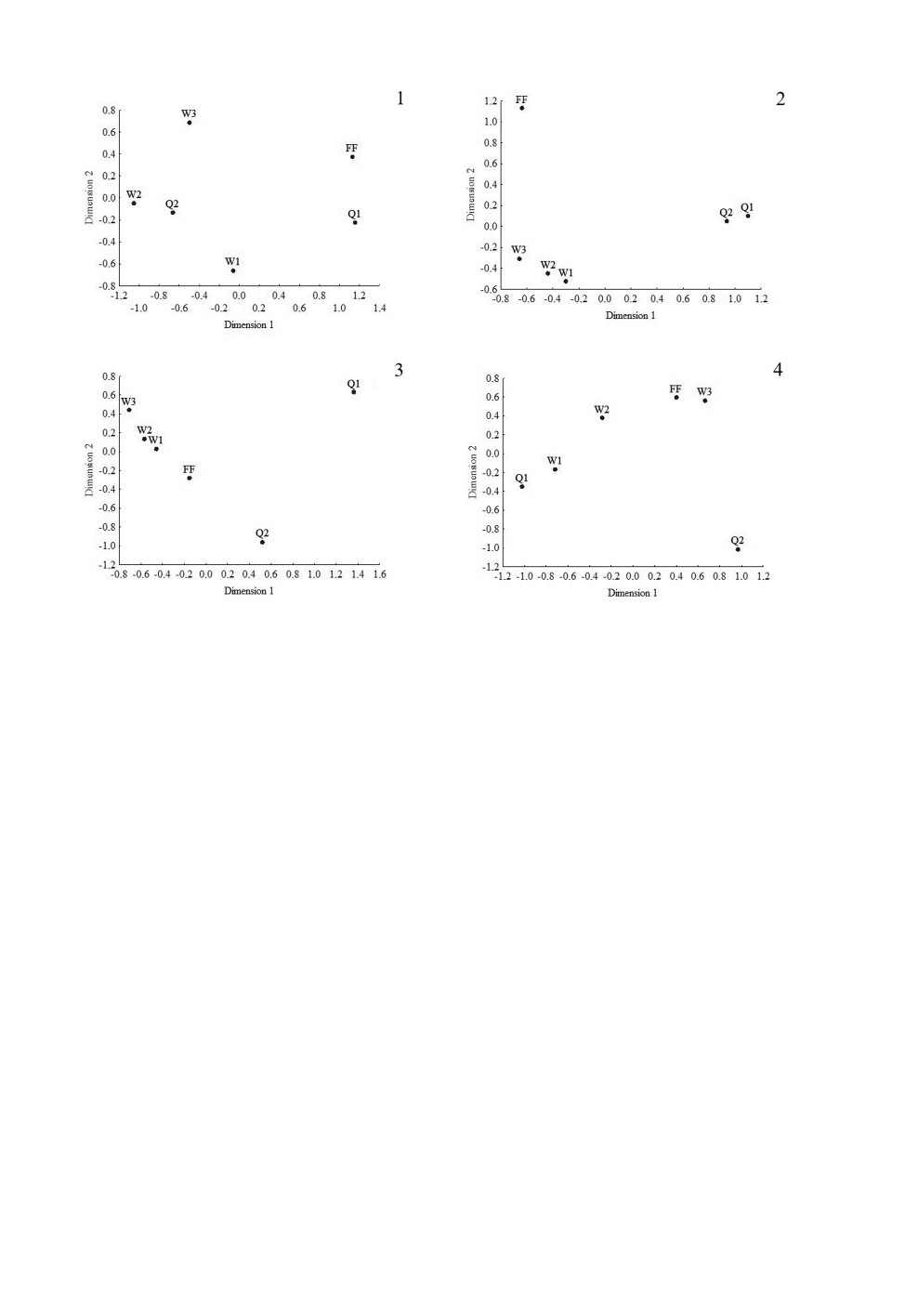
рабочих P. albellus (см. табл. 3, рис. 15, 1-4). У самок-основательниц из разных ско-
плений были сходны встречаемость вариантов рисунка 2-го тергита, мезоскутума и
4-го стернита, но существенно различались они по частоте 3-го и 5-го стернитов.
У основательниц второго поселения чаще встречались более меланизированные вари-
анты рисунков на этих частях тела. Соотношение вариантов различных рисунков на
всех частях тела у рабочих обоих поселений было сходным (см. табл. 4).
В целом выборки рабочих и будущих основательниц из разных поселений и оди-
ночных гнезд похожи, а выборки основательниц фенотипически отличались как от
них, так и одна от другой.
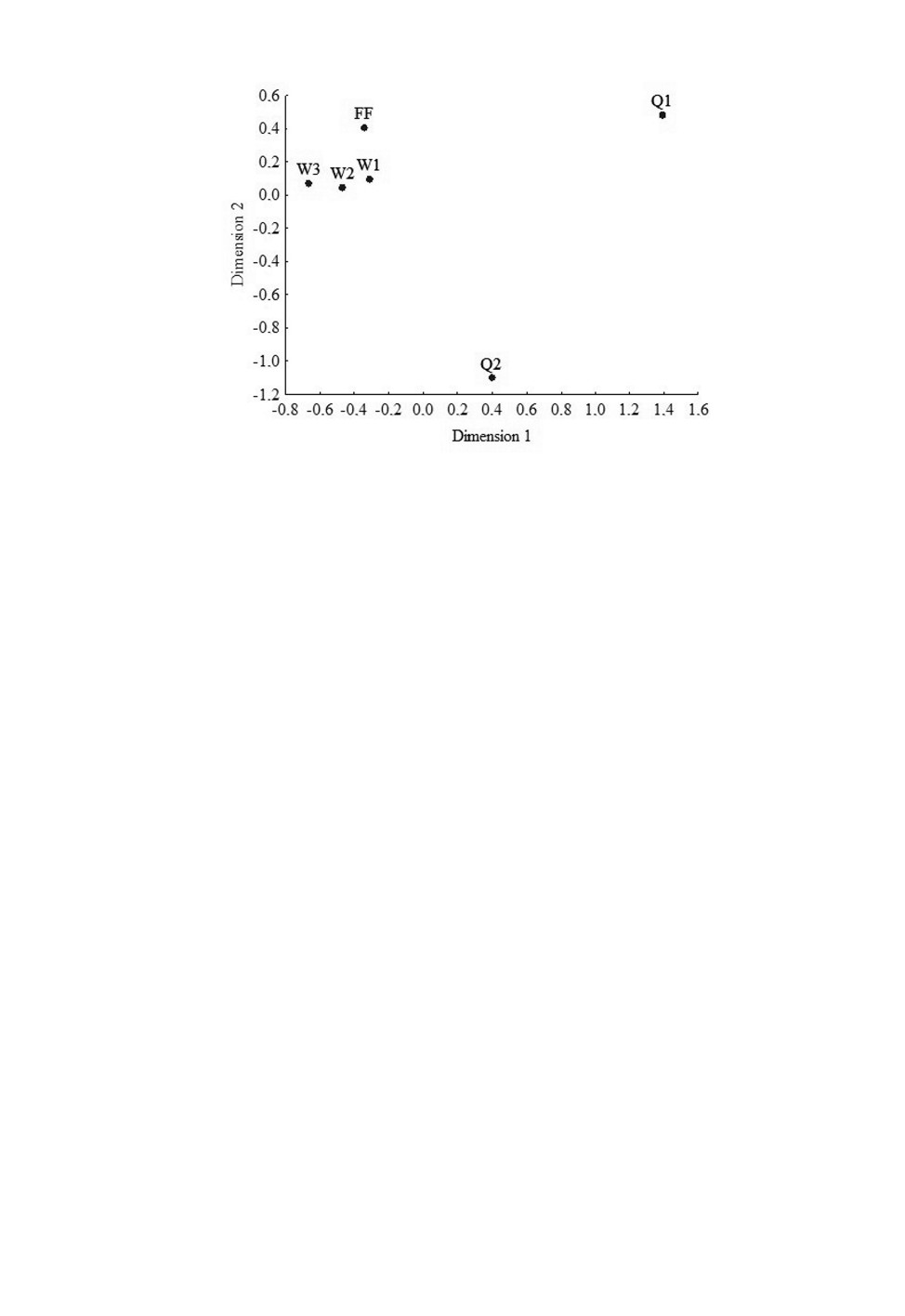
Обращает на себя внимание тот факт, что практически по всем признакам выборки
рабочих выстраиваются в последовательности W1-W2-W3 вдоль одной из осей
(рис. 15, 16). Иногда в той же последовательности выстраиваются и выборки са-
мок-основательниц. Эта последовательность смены фенооблика отражает градиент
изменений в местах гнездования от более влажных к более сухим местообитаниям.
Для анализа сочетаний вариантов рисунков были выбраны части тела самок, для ко-
торых была характерна наибольшая изменчивость (мезоскутум и 2-й тергит мета-
сомы) (табл. 5).
Таблица 5. Встречаемость разных фенотипов Polistes albellus Giordani Soika в поселениях
Центральной Мещеры, %
Фенотип
1 (N = 36)
2 (N = 74)
3 (N = 40)
4 (N = 248)
Ms3.5-2Т9
58.34
31.08
35
27.02
Ms3.5-2Т10
11.11
25.67
27.5
27.82
Ms3.5-2Т11
11.11
16.22
7.5
13.31
Ms4.5-2Т9
8.33
5.41
20
6.45
Ms4.5-2Т10
8.33
10.81
7.5
8.87
Ms4.5-2Т11
2.78
10.81
2.5
16.53
Всего
100
100
100
100
П р и м е ч а н и е. 1 - самки-основательницы первого поселения, 2 - рабочие первого поселения, 3 -
самки-основательницы второго поселения, 4 - рабочие второго поселения. N - число самок.
747
Рис. 15. Фенотипические дистанции по мезоскутуму (1), 2-му тергиту (2), 5-му стерниту
метасомы (3) и 3-му стерниту метасомы (4) между основательницами 1-го поселения - Q1,
рабочими 1-го поселения - W1, основательницами 2-го поселения - Q2, рабочими 2-го
поселения - W2, рабочими из одиночных гнезд - W3 и будущими самками-
основательницами - FF, взятыми в общей совокупности.
Dimension 1 - измерение 1, Dimension 2 - измерение 2.
У основательниц обоих поселений наиболее часто отмечались светлые варианты ри-
сунков Ms3.5-2Т9 и Ms3.5-2Т10. Различие между поселениями заключалось в том,
что у основательниц 2-го поселения часто встречался и более темный вариант
Ms4.5-2Т9.
У рабочих обоих поселений наиболее часто встречались варианты рисунка
Ms3.5-2Т9, Ms3.5-2Т10, Ms3.5-2Т11. Однако только в 1-м поселении также были от-
мечены особи Ms4.5-2Т10.
Отметим, что в мещерских выборках были представлены все фенотипы, которые
были нами описаны по фотографиям из других, значительно удаленных частей ареала
вида (см. табл. 1). Так, рабочие и будущие основательницы P. albellus из Швейцарии
имели фенотип Ms3.5-2T9 (N = 8). Из Башкирии описаны 3 самки с сочетаниями ва-
риантов Ms3.5-2T9 (2 случая) и Ms3.5-2T10. Самка из Оренбургской обл. имела фе-
нотип Ms3.5-2T10. В Забайкальском крае было отмечено сочетание Ms4.5-2T11 (Осы
России. [Интернет-ресурс]), в Казахстане - Ms3.5-2Т11.
748
Рис. 16. Суммарные фенотипические дистанции между основательницами 1-го поселения -
Q1, рабочими 1-го поселения - W1, основательницами 2-го поселения - Q2, рабочими 2-го
поселения - W2, рабочими одиночных гнезд - W3, будущими самками-основательницами - FF.
Dimension 1 - измерение 1, Dimension 2 - измерение 2.
ОБСУЖДЕНИЕ
Меланиновый рисунок насекомых изучается уже более 100 лет, преимущественно
на примере жесткокрылых. Вначале число известных аберраций было сравнительно
невелико и им даже присваивались латинские названия, не имеющие, впрочем, таксо-
номического статуса (см., напр., Плавильщиков, 1936; Захаров, 1997, и др.). Однако по
мере накопления материала число вариантов рисунка для отдельных видов перева-
лило за сотни и использование их в прежнем виде стало невозможным. Важный шаг
сделал Н. Н. Филиппов (1961), показавший, что многие аберрации выстраиваются
в ряды форм, связанных определенными преобразованиями. Дальнейшее изучение по-
казало, что рисунок насекомых состоит из определенных элементов, появление или
исчезновение которых связаны с уровнем пигментации (количеством пигмента в кути-
куле насекомого), и подчиняется видоспецифичным закономерностям (Креславский,
1975, 1977; Васильев, 1988, 2005, и др.).
Проведенный нами анализ показывает, что изменчивость меланинового рисунка
у P. dominula, P. nimpha и P. albellus сходна (рис. 7-13; Русина и др., 2006, 2007в),
но у P. nimpha она обнаружена на большем числе сегментов, однако у P. albellus спектр
вариантов в некоторых случаях оказывается шире, например, на мезоскутуме
(см. рис. 8), а также на мезоплеврах и тазиках самцов (см. рис. 11, 12).
Polistes albellus, в отличие от P. nimpha и P. dominula, гнездится открыто на расте-
ниях; перезимовавшие самки основывают семью гаплометротично. У видов, заселя-
ющих помимо растений разного рода укрытия естественного и антропогенного проис-
хождения, самки-основательницы P. nimpha с более светлыми вариантами рисунка
1-го тергита метасомы предпочитают гнездиться в укрытиях, а с более темными - на
растениях, тогда как у P. dominula - наоборот (Русина, 2014). Данная закономерность
749
у P. nimpha прослеживается на значительной части ареала от Крыма до Зауралья (Ру-
сина и др., 2008).
В ходе полевых исследований локальных поселений P. аlbellus в Центральной Ме-
щере установлено, что особенности меланинового рисунка отражают разные компо-
ненты изменчивости ос-полистов. (см. табл. 4). Выборки рабочих и будущих самок-ос-
новательниц
фенотипически сходны. Кроме того, проведенный анализ
фенотипических дистанций и результаты многомерного шкалирования показали, что
взаиморасположение выборок хорошо соответствует градиенту увлажнения микроме-
стообитаний. Выборки ос с влажного злаково-разнотравного луга (первое поселение)
имеют наиболее темную окраску, а осы из более сухих местообитаний (одиночные
гнезда) - наиболее светлую. Осы из поселения на залежи, зарастающей сосновым под-
ростом (как известно, сосна предпочитает сухие, хорошо дренируемые почвы), зани-
мают промежуточное положение. Можно предположить, что на рабочих ос и будущих
основательниц, развивающихся в разных условиях, фактор увлажнения может дей-
ствовать как непосредственно, так и опосредованно через различающиеся спектры их
питания на личиночных стадиях.
Фенотипические различия между самками-основательницами из разных местооби-
таний уже были отмечены для других видов полистов. В частности, исследования
P. nimpha в разных отделениях Луганского природного заповедника (ЛПЗ) показы-
вают, что различия между выборками проявляются в частотах вариантов рисунка 1-го
тергита метасомы (Русина и др., 2007а). Кроме того, по данному признаку были выяв-
лены различия между основательницами P. nimpha, гнездящимися в разных расти-
тельных сообществах Стрельцовского отделения ЛПЗ. Межгодовые различия между
основательницами P. nimpha из ЛПЗ и P. dominula из Черноморского биосферного за-
поведника (ЧБЗ) выражены в частоте вариантов рисунка мезоскутума (Русина и др.,
2007а; Русина, Орлова, 2009).
Остается не выясненным, какие факторы влияют на изменчивость меланинового ри-
сунка рабочих, самцов и будущих основательниц P. albellus. Можно предположить,
что, как и у P. dominula, размеры тела и характер меланинового рисунка особи зависят
от особенностей ее питания на личиночной стадии, обусловленных морфо-физиологи-
ческими чертами самки-основательницы, размерами и историей семьи (Русина,
Орлова, 2010; Фирман, Русина, 2013).
Так, чем темнее был рисунок клипеуса у самок-основательниц P. dominula из ЧБЗ,
тем темнее оказывался клипеус у будущих основательниц и тем светлее у них были
мезоскутум и 1-й тергит метасомы. Кроме того, чем больше были семьи, тем были
темнее клипеус и светлее мезоскутум и 1-й тергит метасомы у будущих основа-
тельниц. Семьи P. dominula существенно различаются не только по числу рабочих,
приходящихся на одну личинку IV и V возрастов, но также и по их фуражировочной
активности. В успешных семьях с активными самками-основательницами эти показа-
тели выше по сравнению с иными категориями семей: сиротскими, зараженными 1-й
генерацией паразитоидов или узурпированными (агрессивно захваченными чужими
самками) (Русина, 2009).
Экспериментальные исследования P. dominula показали, что изменение рациона пи-
тания личинок V возраста сказывалось в пределах одной семьи на меланизации 1-го
тергита метасомы, а в пределах поселения - на частотах встречаемости вариантов ри-
750
сунка мезоскутума (Русина, Орлова, 2010). Недостаточное питание личинок IV и V
возрастов в эксперименте (вследствие снижения в два раза числа фуражиров и кор-
милиц) усиливало меланизацию 1-го тергита метасомы, а клипеус будущих основа-
тельниц оказывался при этом почти непигментированным. Существенная нехватка
корма вела к измельчанию самок и к усилению пигментации мезоскутума.
В нашем исследовании была установлена зараженность некоторых семей P. albellus
паразитоидом Latibulus argiolus. Зараженные и незараженные семьи были сходны по
размерам; гнезд, в которых L. argiolus 1-й генерации заражал расплод до выхода ра-
бочих, не обнаружено. Вместе с тем, на территории ЛПЗ в отдельные годы доля семей
P. nimpha, зараженных1-й генерацией L. аrgiolus, достигала почти 30 % (Русина, 2009).
Самки паразитоида выбирали для заражения более крупные семьи до выхода рабочих,
которые были основаны самками с более темными вариантами рисунка. В семьях
P. albellus паразитоид Elasmus schmitti не найден, но он заражает куколок в семьях
P. nimpha, гнездящегося в Центральной Мещере не только в укрытиях (Косякова и др.,
2020), но также и на растениях (второе поселение, 2020 г.).
На самках и самцах P. albellus из Мещеры паразиты клещ Sphexicozela connivens и
осиный веерокрыл Xenos vesparum не были обнаружены, но в Швейцарии отмечены
случаи заражения изучаемого вида полистов X. vesparum (личная коллекция Р. П. Ной-
мейера).
Клещ S. connivens найден на самках и самцах P. nimpha, гнездящегося на растениях
неподалеку от P. albellus в тех же самых поселениях, а X. vesparum был отмечен на
самках P. dominula, гнездящихся в укрытиях в Рязани.
Показано, что зараженность паразитами также оказывает влияние на фенотипиче-
скую изменчивость самцов и будущих основательниц (Русина, Орлова, 2011). Так,
слабое и умеренное заражение личинок и куколок P. nimpha клещом S. connivens при-
водило к изменению частот встречаемости вариантов рисунка 4-го стернита, но не
сказывалось на размерах развивающихся из них будущих основательниц. При сильном
заражении личинок (более 5 клещей на особь) наблюдалось измельчание отродив-
шихся имаго и увеличение среди них доли особей со слабо меланизированным вари-
антом рисунка 4-го стернита метасомы (Русина, Орлова, 2011).
В целом процесс меланизации и формирования структуры рисунка P. albellus оста-
ется во многом не исследованным, поэтому приходиться ограничиваться приведенной
в статье схемой, видоизменяя и совершенствуя ее по мере накопления данных по онто-
генезу рисунка, с одной стороны, и фенооблику популяций в разных частях ареала
вида, с другой.
БЛАГОДАРНОСТИ
Авторы признательны А. П. Моргачеву, Е. А. Моргачевой, А. С. Лукиной, C. А. Лу-
кину, Д. A. Дробот, Д. A. Бовсуновскому за помощь в сборе материала, В. А. Валуеву,
А. В. Крутилину, К. Доблер Гросс (C. Dobler Gross) за разрешение использовать их
фотоматериалы. Мы глубоко благодарны также С. А. Белокобыльскому, С. Я. Резнику,
А. В. Лопатину, А. В. Фатерыге и Б. А. Коротяеву за критические замечания, выска-
занные при обсуждении рукописи.
751
СПИСОК ЛИТЕРАТУРЫ
Анненская Г. Н., Мамай И. И., Цесельчук Ю. Н. 1983. Ландшафты Рязанской Мещеры и возможности их
освоения. М.: Издательство Московского университета, 183 с.
Васильев А. Г. 1988. Эпигенетическая изменчивость: неметрические пороговые признаки, фены и их компо-
зиции. В кн.: А. В. Яблоков (ред.). Фенетика природных популяций. М.: Наука, с. 158-169.
Васильев А. Г. 2005. Эпигенетические основы фенетики: на пути к популяционной мерономии. Екатерин-
бург: Академкнига, 640 с.
Вейр Б. 1995. Анализ генетических данных. М.: Мир, 400 с.
Гланц С. 1999. Медико-биологическая статистика. М.: Практика, 459 с.
Животовский Л. А. 1991. Популяционная биометрия. М.: Наука, 271 с.
Захаров В. М. 1997. Асимметрия животных. М.: Наука, 216 с.
Косякова А. Ю., Лукина А. С., Русина Л. Ю. 2020. Функциональная реакция паразитоидов в локальных по-
селениях социальной осы Polistes nimpha (Christ, 1791) на территории национального парка «Мещер-
ский». В кн.: Р. Т. Багиров и др. (ред.). Концептуальные и прикладные аспекты научных исследований
и образования в области зоологии беспозвоночных: сборник статей V Международной конференции,
26-28 октября 2020 г., г. Томск, Россия. Томск: Издательство Томского государственного университе-
та, с. 84-87.
Креславский А. Г. 1975. Наследственный полиморфизм, наследственный мономорфизм и их роль в эволю-
ции окраски у листоедов (Coleoptera, Chrysomelidae). Журнал общей биологии 36 (6): 878-886.
Креславский А. Г. 1977. Некоторые закономерности изменчивости и эволюции рисунков на надкрыльях
у жуков-листоедов. Зоологический журнал 56 (7): 1043-1056.
Новоженов Ю. И. 1989. Хронографическая изменчивость популяций. Журнал общей биологии 33 (2):
171-183.
обращения: 11.05.2020).
Плавильщиков Н. Н. 1936. Фауна СССР. Насекомые жесткокрылые. Т. XXI. Жуки-дровосеки. Ч. 1. М.; Л.:
Издательство АН СССР, 612 с.
Русина Л. Ю. 2009. Структурно-функциональная организация популяций ос-полистов (Hymenoptera,
Vespidae). Труды Русского энтомологического общества 79: 1-217.
Русина Л. Ю. 2014. Структурно-функциональная организация популяций ос-полистин (Hymenoptera,
Vespidae: Polistinae). Диссертация на соискание ученой степени доктора биологических наук. СПб.:
Зоологический институт РАН, 497 с.
Русина Л. Ю., Богуцкий М. П., Гилев А. В., Орлова Е. С. 2007а. Фенотипическая структура поселений осы
Polistes nimphus (Christ) (Hymenoptera, Vespidae) в Луганском природном заповеднике. Известия Харь-
ковского энтомологического общества 15 (1-2): 121-130.
Русина Л. Ю., Гилев А. В. Скороход О. В., Филимонова Н. Б., Фирман Л. А. 2007б. Связь окраски осы Polistes
dominulus с пространственно-этологической структурой ее популяции в Нижнем Приднепровье.
Успехи современной биологии 127 (2): 157-165.
Русина Л. Ю., Гилев А. В., Фирман Л. А., Глотов С. В., Говорун А. В., Пеканова И. А., Русин И. Ю., Черствый
С. А., Фатерыга А. В. 2008. Изменчивость окраски осы Polistes nimphus (Christ) (Hymenoptera,
Vespidae) в Украине и южном Зауралье России. Природничий альманах. Біологічні науки 10: 158-173.
Русина Л. Ю., Лопатин А. В. 2009. Связь фенотипической изменчивости самок-основательниц Polistes
nimphus (Christ) (Hymenoptera, Vespidae) со способом основания семьи. В кн.: М. М. Марченко (ред.).
Науковий вісник Чернівецького університету: Збірник наукових праць. Вип. 455: Біологія. Чернівці:
«Рута», с. 120-125.
Русина Л. Ю., Орлова Е. С. 2009. Динамика фенооблика популяции Polistes dominulus в Черноморском био-
сферном заповеднике. Вісник Дніпропетровського унiверситету. Біологія. Екологія 17 (1): 205-211.
doi: 10.15421/010931
Русина Л. Ю., Орлова Е. С. 2010. Связь фенотипической изменчивости будущих основательниц осы Polistes
dominulа с режимом их питания в личиночном состоянии. Труды Русского энтомологического обще-
ства 81 (2): 165-171.
Русина Л. Ю., Орлова К. С. 2011. Связь фенотипической изменчивости будущих основательниц Polistes
nimpha (Hymenoptera, Vespidae, Polistinae) с характером их зараженности в личиночном состоянии
клещом Sphexicozela connivens (Acari, Astigmata, Winterschmidtiidae). Энтомологическое обозрение 90
Русина Л. Ю., Орлова Е. С., Богуцкий М. П. 2009. Агрессивность самок-основательниц Polistes dominulus
(Hymenoptera: Vespidae), гнездящихся в Черноморском биосферном заповеднике. Научные ведомости
752
Белгородского государственного университета. Серия естественные науки 3 (58): 68-74. https://www.
elibrary.ru/item.asp?id=15182991
Русина Л. Ю., Русин И. Ю., Старр Х. К., Фатерыга А. В., Фирман Л. А. 2007в. Способы основания семьи
самками различных морфотипов у бумажных ос-полистов (Hymenoptera, Vespidae, Polistes). Энтомо-
логическое обозрение 86 (4): 750-772.
Русина Л. Ю., Скороход О. В., Гилев А. В. 2004а. Дискретные вариации окраски осы Polistes dominulus
(Christ) (Hymenoptera: Vespidae) в Черноморском биосферном заповеднике. Труды Русского энтомоло-
Русина Л. Ю., Фирман Л. А., Скороход О. В., Гилев А. В. 2004б. Изменчивость окраски и основные направ-
ления развития рисунка ос рода Polistes (Hymenoptera: Vespidae) в Нижнем Приднепровье. Сообще-
ние 1. Polistes dominulus (Christ). Известия Харьковского энтомологического общества 12 (1-2):
195-207.
Русина Л. Ю., Фирман Л. А., Скороход О. В., Гилев А. В. 2006. Изменчивость окраски и основные направле-
ния развития рисунка ос рода Polistes (Hymenoptera: Vespidae) в Нижнем Приднепровье. Сообщение
3. Polistes nimphus (Christ). Известия Харьковского энтомологического общества 14 (1-2): 110-115.
Сергиевский С. О. 1985. Полифункциональность и пластичность генетического полиморфизма (на примере
популяционного меланизма двуточечной божьей коровки Adalia bipunctata (L.). Журнал общей биоло-
гии 46 (4): 491-502.
Сергиевский С. О. 1987. Полиморфизм как универсальная адаптивная стратегия популяций. Труды Зоологи-
ческого института АН СССР 160: 41-58.
Филиппов Н. Н. 1961. Закономерности аберративной изменчивости рисунка надкрылий жесткокрылых.
Зоологический журнал 40 (3): 372-385.
Фирман Л. А., Русина Л. Ю. 2009. Структура скрещиваний в популяции Polistes dominulus (Christ)
asp?id=12862330
Фирман Л. А., Русина Л. Ю. 2013. Продолжительность онтогенеза самцов в семьях осы Polistes dominulа
(Christ) (Hymenoptera, Vespidae: Polistinae) на юге Украины. Энтомологическое обозрение 92 (3):
Яблоков А. В. 1976. Популяционная морфология как новое направление эволюционно-морфологических и
популяционных исследований. Журнал общей биологии 38 (5): 649-659.
Яблоков А. В., Ларина Н. И. 1985. Введение в фенетику популяций. Новый подход к изучению природных
популяций. М.: Высшая школа, 159 с.
Cavalli-Sforza L. L., Edvards A. W. F.1967. Phylogenetic analysis models and estimation procedures. American
Journal of Human Genetics 19 (3): 233-257.
Gumovsky A., Rusina L., Firman L. 2007. Bionomics, morphological and molecular characterisation of Elasmus
schmitti and Baryscapus elasmi (Hymenoptera: Chalcidoidea, Eulophidae), parasitoids associated with
a paper wasp, Polistes dominulus (Vespoidea, Vespidae). Entomological Science
10
(1):
21-34.
Makino S. 1983. Biology of Latibulus argiolus (Hymenoptera: Ichneumonidae), a parasitoid of the paper wasp
Polistes biglumis (Hymenoptera: Vespidae). Kontyû 51 (3): 426-434.
Neumeyer R., Baur H., Guex G.-D., Praz C. 2014. A new species of the paper wasp genus Polistes (Hymenoptera,
Vespidae, Polistinae) in Europe revealed by morphometrics and molecular analyses. ZooKeys 400: 67-118.
Neumeyer R., Gereys B., Castro L. 2015. New data on the distribution of Polistes bischoffi Weyrauch, 1937 and
Polistes helveticus Neumeyer,
2014, a synonym of Polistes albellus Giordani Soika,
1976
N. STAT. (Hymenoptera: Vespidae). Boletín de la Sociedad Entomológica Aragonesa (S. E. A.) 57: 205-216.
doi: 10.5281/zenodo.47902
Reeve H. K. 1991. Polistes. In: K. G. Ross, R. W. Matthews (eds). The Social Biology of Wasps. New York: Cornell
University Press, p. 99-148.
Röseler P.-F. 1991. Reproductive competition during colony establishment. In: K. G. Ross, R. W. Matthews (eds).
The Social Biology of Wasps. New York: Cornell University Press, p. 309-335.
Rusina L. Y., Gilyov A. V., Firman L. A., Pekanova I. A., Orlova E. S. 2011. Dynamic stability of phenotypic
variability in Polistes wasps (Hymenoptera: Vespidae). Russian Entomological Journal 20 (3): 321-326.
Tibbets E. A. 2002. Visual signals of individual identity in the wasp Polistes fuscatus. Proceedings of the Royal
Society 269 (1499): 1423-1428. doi: 10.1098/rspb.2002.2031
Tibbets E. A., Dale J. 2004. A socially enforced signal of quality in a paper wasp Polistes dominulus. Nature 432
(7014): 218-222. doi: 10.1038/nature02949
753
PHENOTYPIC VARIABILITY OF POLISTES ALBELLUS GIORDANI SOIKA, 1976
(HYMENOPTERA: VESPIDAE)
A. Yu. Kosyakova, A. V. Gilev, R. P. Neumeyer, L. Yu. Rusina
Key words: social wasps, Vespidae, Polistes albellus, phenotypic variability, melanin
patterns.
SUMMARY
This study is dealing with the coloration and melanin patterns of the social wasp Polistes albellus
Giordani Soika, 1976 (Hymenoptera: Vespidae), which is distributed from Western Europe (France)
to the Pacific coast of Russia (Primorskiy Territory) and to Mongolia and China. The total number of
adult wasps examined was 590 females and 101 males. Phenotypic variability in females of P. albellus
is expressed on the mesoscutum, the first and second tergite, and the sternites 4 to 6 of the metasoma. In
males, however, the variability is additionally expressed on the clypeus and the coxae.
Analyses of the relationship between coloration elements on the one hand and spatial as well as
temporal nesting features on the other hand were done in three local populations of P. albellus of
Meshchera Lowlands (Ryazan prov., Russia). It was shown that overwintering foundresses of different
morphotypes differentiated by the timing of nest initiation, nest attachment height to plant, and tendency
to form clusters. Samples of queens of different settlements are phenotypically differentiated both from
workers and from each other. All samples of workers and future foundresses are phenotypically similar.
The mesoscutum pattern appears to be related to habitat moisture: in dry habitats, workers have a higher
proportion of dark pattern variants.
The heterogeneity of environmental conditions can be a factor in increasing the variability of the
melanin pattern of wasps.
754