ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 100, 4, 2021
УДК 595.78
ПЯДЕНИЦЫ (LEPIDOPTERA, GEOMETRIDAE) БАЙКАЛЬСКОГО
РЕГИОНА: УТОЧНЕНИЕ ВИДОВОГО СОСТАВА И РЕЗУЛЬТАТЫ
МОЛЕКУЛЯРНОГО ДНК-БАРКОДИНГА
© 2021 г. И. А. Махов,* В. А. Лухтанов**
Зоологический институт РАН
Университетская наб., 1, С.-Петербург, 199034 Россия
*e-mail: maakhov@mail.ru, **e-mail: lukhtanov@mail.ru
Поступила в редакцию 30.09.2021 г.
После доработки 28.10.2021 г.
Принята к публикации 28.10.2021 г.
В работе приводятся дополнения к фаунистическому списку пядениц Байкальского региона.
Для семи видов уточнено распространение в его пределах; семь видов - Chlorissa obliterata
(Walker, 1863), Scotopteryx acutangulata (Inoue, 1941), Entephria byssata (Aurivillius, 1891), Minoa
lutea Schwingenschuss, 1954, Eupithecia sophia Butler, 1878, Idaea dimidiata (Hufnagel, 1767) и
Scopula flaccidaria (Zeller, 1852) - впервые приводятся для Байкальского региона, и один вид,
Charissa beljaevi Erlacher, Marrero et Erlacher, 2017 - впервые для России. Установлена новая
комбинация для Nebula korschunovi comb. n. Впервые создана библиотека ДНК-баркодов пяде-
ниц Байкальского региона, которая включает 631 нуклеотидную последовательность фрагмента
гена COI для 312 видов (86 % состава фауны). Анализ ДНК-баркодов и морфологии особей, для
которых баркоды были получены, приводит к следующим результатам: 1) добавляет один вид
(Chlorissa obliterata) к фауне региона; 2) выявляет 28 опубликованных в литературе и в публич-
ных базах данных случаев ошибочной видовой идентификации, ассоциированных с нуклеотид-
ными последовательностями; 3) позволяет однозначно делимитировать морфологически слабо
дифференцированные виды-двойники трех пар: пара Macaria alternata ([Denis et Schiffermüller],
1775) и M. shanghaisaria Walker, 1861; пара Isturgia murinaria ([Denis et Schiffermüller], 1775) и
I. kaszabi Vojnits, 1974, и пара Chlorissa viridata (Linnaeus, 1758) и Ch. obliterata; 4) показывает
глубокую (3 % и более) генетическую дифференциацию байкальских и/или восточноазиатских
популяций пяти видов, что свидетельствует о необходимости их таксономической ревизии.
Ключевые слова: Geometridae, пяденицы, фауна, Байкальский регион, Иркутская область,
Республика Бурятия, ДНК-баркодинг.
DOI: 10.31857/S0367144521040080
Пяденицы - одно из наиболее богатых видами семейств чешуекрылых, насчитыва-
ющее в мировой фауне около 24 000 описанных видов и распространенное всесветно,
за исключением полярных регионов (Yamamota, Sota, 2007; Van Nieukerken et al., 2011;
Murillo-Ramos et al., 2019).
836
Фауна пядениц России насчитывает около 1125 видов из 330 родов (Беляев, Ми-
ронов, 2019). Многие регионы России, однако, до сих пор изучены недостаточно, в
особенности это касается удаленных северных и северо-восточных территорий со
слабо развитой дорожной сетью. Данная работа посвящена одному из таких реги-
онов - Байкальскому, который включает территории двух административных субъ-
ектов, примыкающих к оз. Байкал: Иркутской области и Республики Бурятия.
В последние годы первый автор данной работы вместе с коллегами проводил инвен-
таризацию фауны сем. Geometridae Байкальского региона. Несмотря на то, что фауни-
стические списки региона неоднократно публиковались и обновлялись (Вийдалепп,
1987; Васильева, 1989; Белова, 2000; Берлов, Берлов, 2006; Гордеева, Гордеев, 2007),
за последние 10 лет нами был обнаружен ряд видов, не отмеченных на этой терри-
тории ранее, а также подтверждено обитание нескольких видов, указания которых
считались сомнительными. Кроме того, более 20 видов пядениц, известных из региона
по литературным источникам, были исключены из фаунистического списка после про-
верки коллекционного материала. Наиболее полный опубликованный список видов
региона, включающий 350 видов, можно найти в Каталоге чешуекрылых России (Бе-
ляев, Миронов, 2019). Всего с учетом находок последних лет (Makhov, Beljaev, 2019;
Махов, 2021; Makhov, 2021) для региона отмечено 353 вида.
В данной работе приводятся дополнения к фаунистическому списку пядениц Бай-
кальского региона, которые основаны на исследованиях первого автора, проведенных
в 2019-2021 гг. преимущественно в ранее не исследованных районах Иркутской обл. и
Бурятии, а также на анализе коллекций Зоологического института РАН и других на-
учных учреждений России.
Помимо этого, в статье публикуются результаты молекулярного ДНК-баркодинга
пядениц Байкальского региона. ДНК-баркодинг - метод молекулярной идентификации
видов, основанный на анализе и сравнении нуклеотидных последовательностей ко-
ротких, как правило, видоспецифичных участков генов (Hebert et al., 2003a). Многие
специалисты сходятся во мнении, что использование этого метода в сочетании с ана-
лизом данных по морфологии, биологии и экологии позволяет существенно повысить
надежность видовой идентификации (Silva-Brandão et al., 2009; Rajaei et al., 2013;
Rubio et al., 2013; Stadie et al., 2014; Feizpour et al., 2018; Wanke et al., 2019).
МАТЕРИАЛ И МЕТОДИКА
Основой работы стали сборы первого автора за период 2009-2021 гг., а также материалы лич-
ных коллекций Э. Я. Берлова, В. Г. Шиленкова, А. В. Косарева (Иркутск), А. В. Филиппова
(Улан-Удэ) и С. А. Князева (Омск). Помимо этого, были изучены коллекционные фонды следу-
ющих институтов и учреждений: Зоологический институт РАН (С.-Петербург), Институт систе-
матики и экологии животных СО РАН (Новосибирск), Бурятский научный центр СО РАН
(Улан-Удэ), Биолого-почвенный факультет Иркутского государственного университета
(Иркутск).
Сбор материала и его обработка. Бабочки собирались с конца марта по начало октября с
использованием световой ловушки, оборудованной ртутной лампой Sylvania HSL-BW 250W
E40, запитанной от бензинового электрогенератора FUBAG TI 1000, и переносным экраном из
белой ткани.
837
Фотографии гениталий получены с помощью стереомикроскопа Nikon SMZ25, фотокамеры
Nikon DS-Ri2 и программного обеспечения NIS-Elements BR на базе ресурсного центра «Так-
сон» Зоологического института РАН (С.-Петербург).
Составление списка новых для региона видов. При составлении аннотированного списка
видов была использована система типизации ареалов, предложенная К. Б. Городковым (1984,
1985, 1986, 1992). В списке после названия вида и характеристики типа его ареала приведено
распространение в Байкальском регионе с использованием условных обозначений: БР - Бай-
кальский регион (вид отмечен как в Иркутской обл., так и в Бурятии); ИО - вид отмечен только
в Иркутской обл. (Предбайкалье); РБ - вид известен только из Бурятии.
Виды, отмеченные впервые для всего Байкальского региона, помечены восклицательным зна-
ком; виды, впервые зарегистрированные в одном из входящих в него субъектов, отмечены звез-
дочкой (*). В разделе «Материал» приведены данные этикеток изученных экземпляров и место
их депонирования (в квадратных скобках): БНЦ - Бурятский научный центр Сибирского отделе-
ния РАН, Улан-Удэ; БПФ - Биолого-почвенный факультет Иркутского государственного универ-
ситета, Иркутск; ЗИН - Зоологический институт РАН, С.-Петербург; ИСЭЖ - Институт
систематики и экологии животных Сибирского отделения РАН, Новосибирск.
ДНК-баркодинг. Выделение ДНК проводилось с помощью колонок и набора реактивов
DNeasy Blood & Tissue Kit (QIAGEN, Germany) в соответствии с протоколом производителя.
Элюция ДНК проводилась с использованием 150-μl элюирующего буфера. Амплификация фраг-
мента гена субъединицы I цитохромоксидазы I (COI) длиной 658 пар нуклеотидов производи-
лась с использованием прямого праймера LCO1490 (5′-GGTCAACAAATCATAAAGATATT
GG-3′) и обратного праймера HCO2198 (5′-TAAACTTCAGGGTGACCAAAAAATCA-3′) (Folmer
et al., 1994) при следующих условиях: 95 °C - 3 мин, 95 °C - 30 сек, 50 °C - 45 сек, 72 °C - 1 мин
(с 34 повторами шагов 2-4), 72 °C - 10 мин.
Секвенирование полученных ПЦР-продуктов выполнено в Центре молекулярных и клеточ-
ных технологий Научного парка Санкт-Петербургского государственного университета и в ком-
пании Евроген (Москва). Для части образцов (181 проба) амплификация и секвенирование
фрагмента гена COI осуществлялись в Канадском центре ДНК-баркодинга (Canadian Centre for
DNA Barcoding, Biodiversity Institute of Ontario, University of Guelph) по стандартным методикам
(deWaard et al., 2008).
Анализ ДНК-баркодов. Расчет оптимальной модели эволюции и вычисления нескорректиро-
ванных генетических p-дистанций (отношения числа замен в двух сравниваемых сиквенсах к
общему числу нуклеотидов в изучаемой последовательности (Hebert et al., 2003)) были выполне-
ны в программе MEGA7 (Kumar et al., 2015). Нуклеотидные последовательности были выравне-
ны в программе BioEdit (Hall, 1999). Байесовский анализ был выполнен с использованием
программы MrBayes v.3.2.7 (Ronquist et al., 2012), для каждого набора последовательностей бы-
ло выполнено два прогона по 10 000 000 генераций с четырьмя цепями (одна холодная и три
горячих). Полученные филогенетические реконструкции были визуализированы с помощью
нии сравнений использовались не индивидуальные ДНК-баркоды, а гаплогруппы, под которыми
понимались совокупности сиквенсов, близких по нуклеотидному составу и образующих на
филогенетическом дереве отдельные клады.
838
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Дополнение и уточнение списка региональной фауны
Подсем. ENNOMINAE
*Gnophopsodos ravistriolaria (Wehrli, 1922). Южносибирский альпийский - БР.
Ранее не указывался для ИО.
М ате риа л. Иркутская обл.: Слюдянский р-н, 20 км Ю Слюдянки, пик Черского, 1850 м над
ур. м., 16.VII.1984, 1 ♂ (leg. С. Синёв) [ЗИН].
Charissa beljaevi Erlacher, Marrero et Erlacher, 2017. Южносибирский монтанный -
?БР. Гениталии ♂ - рис. 1, 1, 2).
М ате риа л. «Jrkut» [р. Иркут], 1 ♂ (сборщик неизвестен) [ЗИН].
З а м еча ни е. Вид впервые приводится для фауны России. Фото этого самца приве-
дено в предыдущей работе (Makhov, 2021: 82, fig. 10H), где он был ошибочно опре-
делен как Ch. (Pterygnophos) ochrofasciata.
Apocheima cinerarius (Erschoff, 1874). Центральноазиатский аридный - юг РБ.
М ате риа л. Бурятия: Тарбагатайский р-н, долина р. Селенга, 6 км С пос. Тарбагатай
(51°32ʹ37ʺ N, 107°21ʹ41ʺ E), 19.V.2021, 1 ♂ (leg. И. Махов) [ЗИН].
З а м еча ни е. Редкий, вероятно, залетный вид, впервые обнаруженный в регионе
Т. В. Гордеевой (БНЦ) в 2018 г. (личное сообщение).
Подсем. GEOMETRINAE
! Chlorissa obliterata (Walker, 1863). Сибирско-восточноазиатский суббореальный -
юг РБ.
М ате риа л. Бурятия: Джидинский р-н, долина р. Цагатуй, 15 км С улуса Гэгэтуй,
11.VII.2016, 1 ♂; кордон Малый Тасархай, 8.VII.2016, 3 ♂; там же, 7.VII.2016, 3 экз. Прибайкаль-
ский р-н, долина р. Селенга, о. Сенокосный, 26.VI.2016, 2 ♂; окр. пос. Мостовка, 28.VI.2015,
6 ♂; там же, 25.VI.2016, 1 экз. (И. Махов) [ЗИН]. Иволгинский р-н, г. Улан-Удэ, станция Комуш-
ка, разнотравный луг, 23.VI.1996, 1 ♀. Селенгинский р-н, долина р. Уда, 3-4 км выше пос. Гар-
топ, восточнее с. Курба, ильмовники, мезофитные луга, 26-27.VI.2016, 2 ♂ (GenBank: OK156233,
OK641942). Заиграевский р-н, окр. пос. Онохой, Добо-Енхор, ильмовники, 15.VII.2008, 2 ♂
(GenBank: OK641946); там же, 2-3.VI.2009, 2 ♂ (GenBank: OK641944, OK641945) (leg. Т. Горде-
ева) [БНЦ].
Подсем. LARENTIINAE
*Acasis appensata (Eversmann, 1842). Трансевразиатский температный - БР. Ранее
этот вид не приводился для ИО.
М ате риа л. Иркутская обл.: Иркутский р-н, Садоводческое некоммерческое товарищество
(СНТ) Лаврентьево (52°08ʹ41ʺ N, 104°18ʹ05ʺ E): 11, 13 и 16.VI.2012, 4 ♂; 15.VI.2013, 1 ♂; окр.
садоводства Родник (52°17ʹ59ʺ N, 104°29ʹ35ʺ E), 13.VI.2016, 1 ♀. Эхирит-Булагатский р-н, окр.
пос. Усть-Ордынский (52°44ʹ30ʺ N, 104°44ʹ19ʺ E), 8.VI.2016, 1 ♂, 1 ♀ (leg. И. Махов); «Иркутск»,
1 ♀ (сборщик неизвестен) [ЗИН]; окр. г. Иркутск, ст. Мельниково (52°15ʹ53ʺ N, 104°14ʹ44ʺ E),
6.VI.1941, 1 ♀; 9.VI.1941, 3 ♂; там же, 13 и 14.VI.1941, 2 ♂; там же, 24.VI.1940, 1 ♀ (leg. А. Бы-
ков) [БПФ].
839
Рис. 1. Geometridae, гениталии самцов.
1, 2 - Charissa beljaevi Erlacher, Marrero et Erlacher; 3, 4 - Scotopteryx acutangulata Inoue;
5-7 - Entephria byssata Aurivillius; 8-10 - Minoa lutea Schwingenschuss.
1, 3, 5, 8 - генитальный сегмент вентрально (1, 3, 5) и латерально (8); 2, 4, 7, 9, 10 - эдеагус дорсально (2)
и латерально (4, 7, 9, 10); 6 - отделенная вальва.
Масштабные линейки - 1 мм.
! Scotopteryx acutangulata (Inoue, 1941). Сибирско-восточноазиатский суббореаль-
ный - РБ.
Мат ер иа л. Бурятия: Селенгинский р-н, окр. оз. Гусиное (51°14ʹ07ʺ N, 106°31ʹ47ʺ E),
8.VII.2020, 2 ♀. Кяхтинский р-н, долина р. Чикой, 6 км ЗСЗ пос. Октябрьск (50°14ʹ05ʺ N,
106°58ʹ50ʺ E), 11.VIII.2019, 1 ♂ (leg. И. Махов) (гениталии - рис. 1, 3, 4) [ЗИН].
840
! Entephria byssata (Aurivillius, 1891). Трансевразиатский аркто-гольцовый - БР.
М ате риа л. Иркутская обл.: Слюдянский р-н, пик Черского, 2090 м над ур. м., 21.VII.1984,
1 ♂, 1 ♀ (leg. С. Ю. Синёв) (гениталии ♂ - рис. 1, 5-7); «Хамар Дабан 25.VII», 4 ♂, 4 ♀; «Jrkut»,
4 ♂, 2 ♀ (сборщик неизвестен) [ЗИН].
! Minoa lutea Schwingenschuss, 1954. Вид отмечен в Крыму, на Северном Кавказе,
в Закавказье и Турции, а также на Алтае - ИО.
М ате риа л. Красноярский край: «Саянскiя горы 24/V.1918 Кожанчиковъ, р. Казир-Сук
[р. Казырсуг]», 2 ♂; там же, 30.V.1918, 1 ♀[ЗИН]. Иркутская обл.: Иркутский р-н, пос. Большие
Коты (51°54ʹ12ʺ N, 105°04ʹ08ʺ E), 6.VII.2010, 1 ♂; СНТ «Лаврентьево» (52°08ʹ41ʺ N, 104°18ʹ05ʺ E),
13-16.VI.2018, 1 ♀ (leg. И. Махов); Глазунова падь (52°23ʹ11ʺ N, 104°15ʹ47ʺ E), 9.V.1914, 1 ♂ (leg.
С. Родионов); «Jrkutzk [Иркутск] 28/6.78. Niels.», 1 ♀; «Jrkutzk 28/5.80. Hns.», 1 ♂; «Jrkutzk
9/6.80. Hns.», 1 ♀; «Иркутская губер.», 1 ♀ [ЗИН].
З а м еча ни е. Недавно показано (Rajaei et al., 2021), что Minoa lutea заслуживает
видового статуса и вид был восстановлен из синонимии с Minoa murinata (Scopoli,
1763). Несмотря на отсутствие выраженной дифференциации по митохондриальным
ДНК-баркодам, M. lutea хорошо диагностируется по небольшому (по сравнению с
M. murinata) корнутусу и некоторым менее заметным признакам гениталий. В упомя-
нутой работе M. lutea впервые приводится для России (Республика Алтай и Северный
Кавказ). Изучение гениталий экземпляров M. lutea из Прибайкалья подтвердило их
определение (гениталии ♂ на рис. 1, 8-10).
*Asthena amurensis (Staudinger, 1897). Сибирско-дальневосточный суббореаль-
ный - БР. Обитание в ИО подтверждается недавней находкой.
М ате риа л. Иркутская обл.: Иркутский р-н, СНТ
«Лаврентьево»
(52°08ʹ41ʺ N,
104°18ʹ05ʺ E), 20.VI.2019, 1 ♀ (leg. И. Махов) [ЗИН].
З а м еча ни е. В базе данных BoLD зарегистрированы 2 экз. с Алтая (Акташ), пол-
ностью идентичные по COI прибайкальским экземплярам A. amurensis и ошибочно
определенные как A. albulata (Hufnagel, 1767). ДНК-баркоды A. amurensis и истинной
A. albulata различаются на 8 %.
*Eupithecia denotata (Hübner, 1813). Евро-кавказско-сибирский температный - БР.
Ранее не указывался для РБ.
М ате риа л. Бурятия: Прибайкальский р-н, долина р. Селенга, окр. пос. Ильинка, о. Сено-
косный (52°08′12″ N, 107°19′36″ E), 27.VI.2016, 1 ♀ (leg. И. Махов) [ЗИН].
*Eupithecia fennoscandica Knaben, 1949. Евро-сибирский аркто-альпийский - БР.
Ранее не указывался для РБ.
М ате риа л. Бурятия: Окинский р-н, Восточный Саян, верховья р. Архат (51°52ʹ23ʺ N,
101°33ʹ51ʺ E), 2000 м над ур. м., 20.VI.2018, 1 ♂ (leg. И. Махов) [ЗИН].
*Eupithecia groenblomi Urbahn, 1969. Трансевразиатский температный - БР. Ранее
не указывался для РБ.
М ате риа л. Бурятия: Кабанский р-н, пос. Речка Выдриная (51°28ʹ36ʺ N, 104°51ʹ32ʺ E),
18.VII.2014, 1 ♀ (leg. И. Махов) [ЗИН].
841
Eupithecia lanceata (Hübner, 1825). Субтрансевразиатский бореальный - БР.
Мат ер иа л. Бурятия: Заиграевский р-н, юго-восточный макросклон хр. Улан-Бургасы, падь
с ручьем Хара-Ацагат, 20 км СВ пос. Онохой, 1.V.2017, 1 ♀ (leg. Т. В. Гордеева) [БНЦ].
За меча ние. В Каталоге чешуекрылых России (Беляев, Миронов, 2019) вид не от-
мечен для РБ, хотя ранее приводился Васильевой (1989) для Тункинской долины.
! Eupithecia sophia Butler, 1878. Сибирско-дальневосточный суббореальный - ИО.
М ат ер иа л. Иркутская обл.: Ольхонский р-н, Байкало-Ленский заповедник, берег оз. Бай-
кал, мыс Большой Солонцовый, 31.VII.2005, 1 ♀ (leg. О. Берлов) [ЗИН].
За меча ние. Приведенный экземпляр ранее был упомянут как «Eupithecia sp. 1.
(sophia species group)» (Берлов, Берлов, 2006: 106).
*Horisme falcata (A. Bang-Haas, 1907). Южносибирский температный - БР. В ИО
ранее не отмечался.
Мат ер иа л. Иркутская обл.: Эхирит-Булагатский р-н, окр. пос. Усть-Ордынский
(52°44ʹ30ʺ N, 104°44ʹ19ʺ E), 22.VIII.2020, 3 ♂, 1 ♀. Ольхонский р-н, окр. пос. Сахюрта
(53°01ʹ52ʺ N, 106°51ʹ42ʺ E), 21.VIII.2020, 1 ♂ (leg. И. Махов) [ЗИН].
Подсем. STERRHINAE
! Idaea dimidiata (Hufnagel, 1767). Субциркумтемператный азиодизъюнктивный -
?ИО. Впервые приводится для БР.
Мат ер иа л. Иркутская обл.: «Лиственичное, Иркутская Область, Попова, 1953», 1 ♀ (leg.
Попова) [ЗИН].
За меча ние. Оригинальная этикетка приведенного экземпляра, очевидно, отно-
сится к пос. Листвянка, расположенному на берегу оз. Байкал. Возможно, этикетка
ошибочна и упомянутый экземпляр собран в другом месте, так как Idaea dimidiata ра-
нее никогда не приводился для исследуемой территории. Однако этот вид известен
в Забайкальском крае (Беляев, Миронов, 2019), поэтому его распространение в Бай-
кальском регионе исключить нельзя.
*Idaea sylvestraria (Hübner, 1799). Трансевразиатский температный - БР. Ранее не
указывался для ИО.
Мат ер иа л. Иркутская обл.: Ольхонский р-н, Тажеранская степь (52°57ʹ07ʺ N, 106°45ʹ01ʺ E),
23.VII.2019, 1 ♀ (leg. И. Махов) [ЗИН].
! Scopula flaccidaria (Zeller, 1852). Центральноазиатский аридный - ?ИО. Ранее не
приводился на БР.
Мат ер иа л. Иркутская обл.: «Лиственичное, Иркутская Область, Попова, 1953», 2 ♂
(leg. Попова); “Jrkut! [р. Иркут]”, 1 ♂ [ЗИН].
За меча ние. Изученные самцы соответствуют инфраподвидовому таксону Acid-
alia flaccidaria var. albidaria Staudinger, 1901, который одни авторы (Вийдалепп, 1988;
Viidalepp, 1996) считают подвидом S. flaccidaria, а другие - самостоятельным видом
(Hausmann, 2004).
842
*Scopula permutata (Staudinger, 1897). Сибирско-туранский суббореальный - БР. Ра-
нее не приводился для ИО.
Замечание. Вид был известен только с юга РБ, однако на сайте Lepiforum (http://
lepiforum.org/wiki/page/Scopula_permutata) представлена самка S. permutata из кол-
лекции Р. Мюллера (Rando Müller), собранная в окрестностях пос. Голоустное (берег
оз. Байкал). Пока это единственный известный экземпляр из ИО.
Результаты ДНК-баркодинга
Нами был обработан 631 образец прибайкальских пядениц, принадлежащих к
312 видам. Все полученные баркоды сибирских представителей сем. Geometridae де-
ступны по номерам: MW792274-MW792412, MZ148325-MZ148361, OK073093-
OK073273, OK156183-OK156397, OK205231-OK205252, OK314959-OK314981,
OK641942-OK641962.
Впервые ДНК-баркоды получены для следующих видов пядениц: Phigalia djakonovi
Molt., Aspitates curvaria Ev., Pseudobaptria corydalaria Graes., Xanthorhoe aridela Prt.,
Euphyia coangulata Prt., Solitanea defricata Pglr., Zola terranea Butl., Rheumaptera
neocervinalis Inoue, Horisme scotosiata Wil., Asthena corculina Butl., Eupithecia
insignioides Wehrli, Eu. nobilitata Stgr., Cleta jacutica Viid., Holarctias rufinularia Stgr.,
Scopula aequifasciata Chr., S. agutsaensis Vsl. и S. impersonata Wlk.
97 % изученных ДНК-баркодов оказались видоспецифичными, т. е. отличными по
нуклеотидному составу от баркодов близких видов. В большинстве случаев эти раз-
личия превышали уровень в 2-3 % (различия наблюдались более чем в 13-17 нуклео-
тидных позициях из 658 изученных).
Выявлено 10 пар видов, в которых баркоды не позволяют достоверно идентифици-
ровать вид, либо полностью (или почти полностью) совпадая у двух близких таксонов,
либо частично перекрываясь у части популяций (табл. 1). У пяти пар из этих десяти
такая ситуация была описана ранее для европейских популяций (Hausmann et al., 2011,
2013; Hausmann, Viidalepp, 2012; Mutanen et al., 2012; Skou, Sihvonen, 2015; Müller
et al., 2019). Четырнадцать видов из этих 10 пар распространены в Байкальском ре-
гионе. Тем не менее, с учетом данных по географическому распространению (табл. 1),
только 10 видов, населяющих Байкальский регион, невозможно делимитировать на
основании их ДНК-баркодов.
Показано, что байкальские популяции ряда видов (табл. 2) резко дифференцированы
по ДНК-баркодам от популяций из Европы, Турции, с Дальнего Востока и из Канады и
представлены специфическими гаплогруппами. При этом байкальские популяции
Yezognophos vittaria Thnb., Aspitates gilvaria Den. et Schiff., A. taylorae Butl., Perizoma
hydrata Tr. и Eupithecia pernotata Gn. демонстрируют значительную степень дивер-
генции митохондриальных ДНК-баркодов (генетическая дистанция с известными га-
плогруппами превышает 2.7-3.0 %). Это выше стандартных дистанций для баркодов в
2-3 %, которые используются в качестве критерия для разграничения аллопатричных
видов (Hebert et al., 2003b).
843
Таблица 1. Пары видов прибайкальских пядениц с идентичными ДНК-баркодами. Случаи,
выявленные в данном исследовании, помечены звездочкой
Видовая пара
Замечание
Chlorissa viridata (Linnaeus, 1758)
Ch. cloraria в Байкальском регионе не отмечен
Ch. cloraria (Hübner, 1813)
(Василенко, 2006), поэтому делимитация Ch. vi-
ridata в регионе возможна по ДНК-баркоду.
*Xanthorhoe sajanaria (Prout, 1914)
X. derzhavini в Байкальском регионе не отмечен,
X. derzhavini (Djakonov, 1931)
поэтому делимитация X. sajanaria в регионе
возможна по ДНК-баркоду.
Epirrhoe tartuensis Möls, 1965
E. rivata в Байкальском регионе не отмечен, поэто-
E. rivata (Hübner, 1813)
му делимитация E. tartuensis в регионе возможна
по ДНК-баркоду.
Thera obeliscata (Hübner, 1787)
Оба вида встречаются в Байкальском регионе,
Th. variata ([Denis & Schiffermüller],
поэтому не могут быть делимитированы по
1775)
баркодам.
Minoa murinata (Scopoli, 1763)
M. murinata в Байкальском регионе не отмечен
M. lutea Schwingenschuss, 1954
(смотреть раздел «Дополнение и уточнение
списка региональной фауны»), поэтому делими-
тация M. lutea в регионе возможна по ДНК-бар-
коду.
*Eupithecia gelidata Möschler, 1860
Eu. nanata в Байкальском регионе не отмечен,
Eu. nanata (Hübner, 1813)
поэтому делимитация Eu. gelidata в регионе
возможна по ДНК-баркоду.
*Eupithecia innotata (Hufnagel, 1767)
Оба вида встречаются в Байкальском регионе,
Eu. ochridata Schütze et Pinker, 1968
поэтому не могут быть делимитированы по
баркодам.
*Idaea dohlmanni (Hedemann, 1881)
I. serpentata может быть обнаружен на западе
I. serpentata (Hufnagel, 1767)
Иркутской области, поэтому баркоды не могут
быть использованы для делимитации этих видов
в Байкальском регионе.
Scopula ternata Schrank, 1802
Было высказано предположение, что идентичные
S. frigidaria (Möschler, 1860)
баркоды у этих видов могут быть следствием
спорадической межвидовой гибридизации и
последующей митохондриальной интрогрессии
(Hausmann et al., 2013). Оба вида встречаются
в Байкальском регионе, поэтому не могут быть
делимитированы по баркодам.
Морфологически сходные виды, делимитированные
на основании анализа ДНК-баркодов
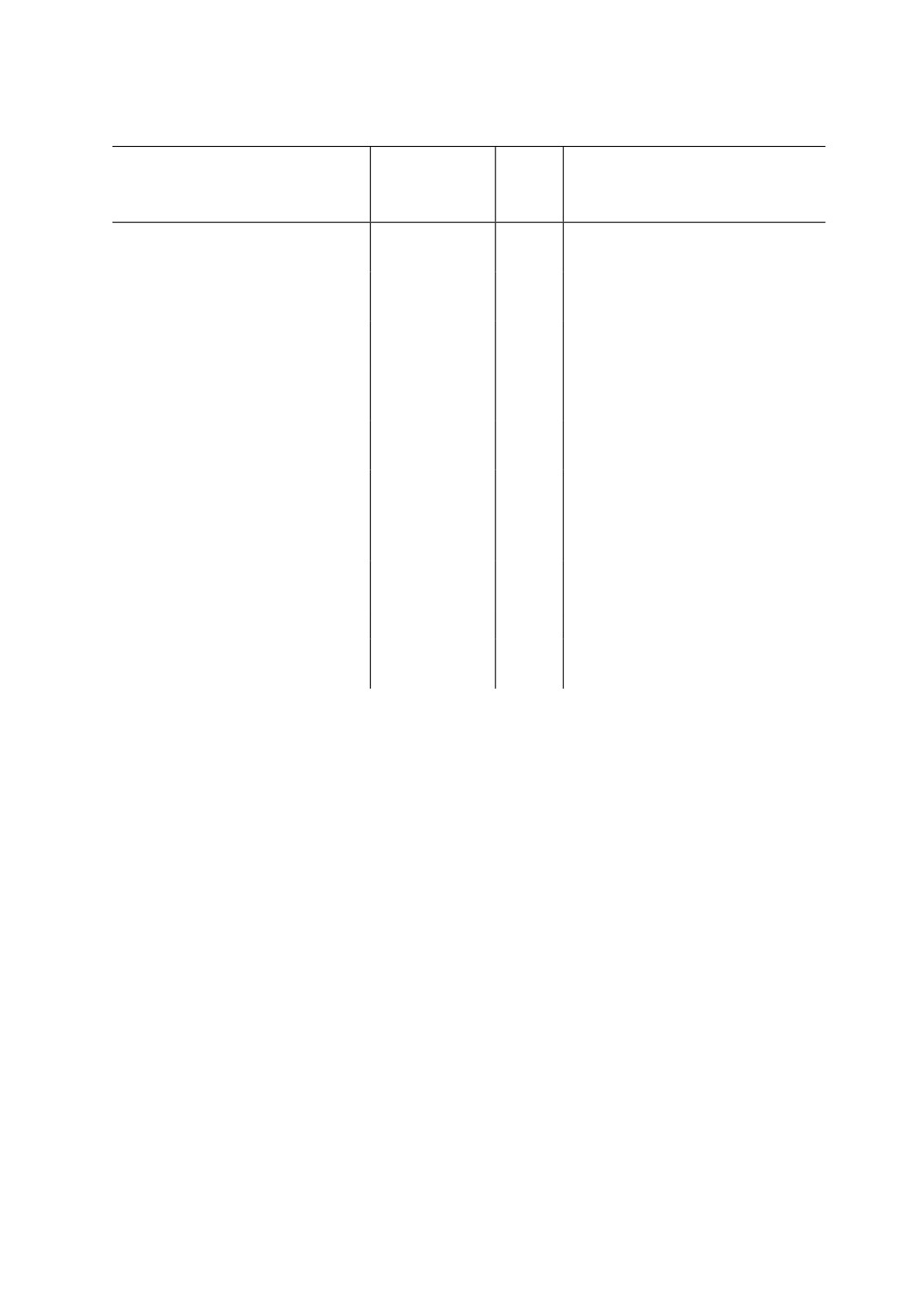
Macaria alternata ([Denis et Schiffermüller], 1775) - Macaria shanghaisaria Walker,
1861.
Виды внешне очень сходны как по строению гениталий, так и по окраске крыльев
(особенно облетанные экземпляры), из-за чего делимитация этой пары видов затруд-
нена, особенно в зоне их симпатрии. Полученные нами ДНК-баркоды M. alternata и
844
Таблица 2. Новые гаплогруппы пядениц, выявленные в ходе анализа ДНК-баркодов сибирских
представителей семейства. Min. d - минимальная генетическая дистанция до другой известной
гаплогруппы этого вида
ID в базе
Min. d,
Местонахождение ближайшей
Вид
данных
%
гаплогруппы
GenBank
Yezognophos vittaria (Thunberg,
OK156256,
3.0
Финляндия
1788)
OK156257
(Müller et al., 2019: 206)
Aspitates gilvaria
OK314973
3.2
Эстония
([Denis et Schiffermüller], 1775)
(Skou, Sihvonen, 2015: 410)
A. taylorae
OK156194
2.9
Канада (deWaard et al., 2011;
(Butler, 1893)
Hebert et al., 2016)
Ectropis crepuscularia
OK156241 -
1.2
Россия, Приморский край
([Denis et Schiffermüller], 1775)
OK156255
(Müller et al., 2019: 468)
Pseudentephria lamata (Staudinger,
OK073213
1.5
Россия, Алтай
1897)
(BoLD: GWORZ676-10)
Perizoma hydrata (Treitschke,
OK073166,
5.0
Грузия, Турция
1829)
OK073266,
(BIN BoLD:AAC3711)
OK073133,
OK073179
Eupithecia pernotata Guenée, 1858
OK073201,
3.2
Финляндия (Mutanen et al., 2016)
OK156296
Scopula immorata (Linnaeus, 1758)
MZ148352
1.2
Германия (BIN BoLD:AAC9960)
S. immutata (Linnaeus, 1758)
OK073115
1.2
Россия, Прибайкалье (данные
авторов)
M. shanghaisaria (GenBank: OK156338, OK156339, OK156340, OK073272) демонстри-
руют четкую молекулярную дифференциацию, различаясь 21 фиксированной нуклео-
тидной заменой (табл. 3) (минимальная p-дистанция - 3.2 %).
Isturgia murinaria ([Denis et Schiffermüller], 1775) - Isturgia kaszabi Vojnits, 1974.
I. murinaria и I. kaszabi демонстрируют слабые и неустойчивые отличительные при-
знаки во внешнем строении имаго. Также слабые, но устойчивые морфологические
различия в строении гениталиий обоих полов были показаны нами ранее (Makhov,
2021). Полученный нами ДНК-баркод (OK156332) для малоизвестного I. kaszabi пока-
зывает, что эти виды различаются 12 заменами (табл. 4) (p-дистанция - 1.8 %), хотя в
данном случае замены трудно интерпретировать как фиксированные, так как изучен
только 1 экз. I. kaszabi.
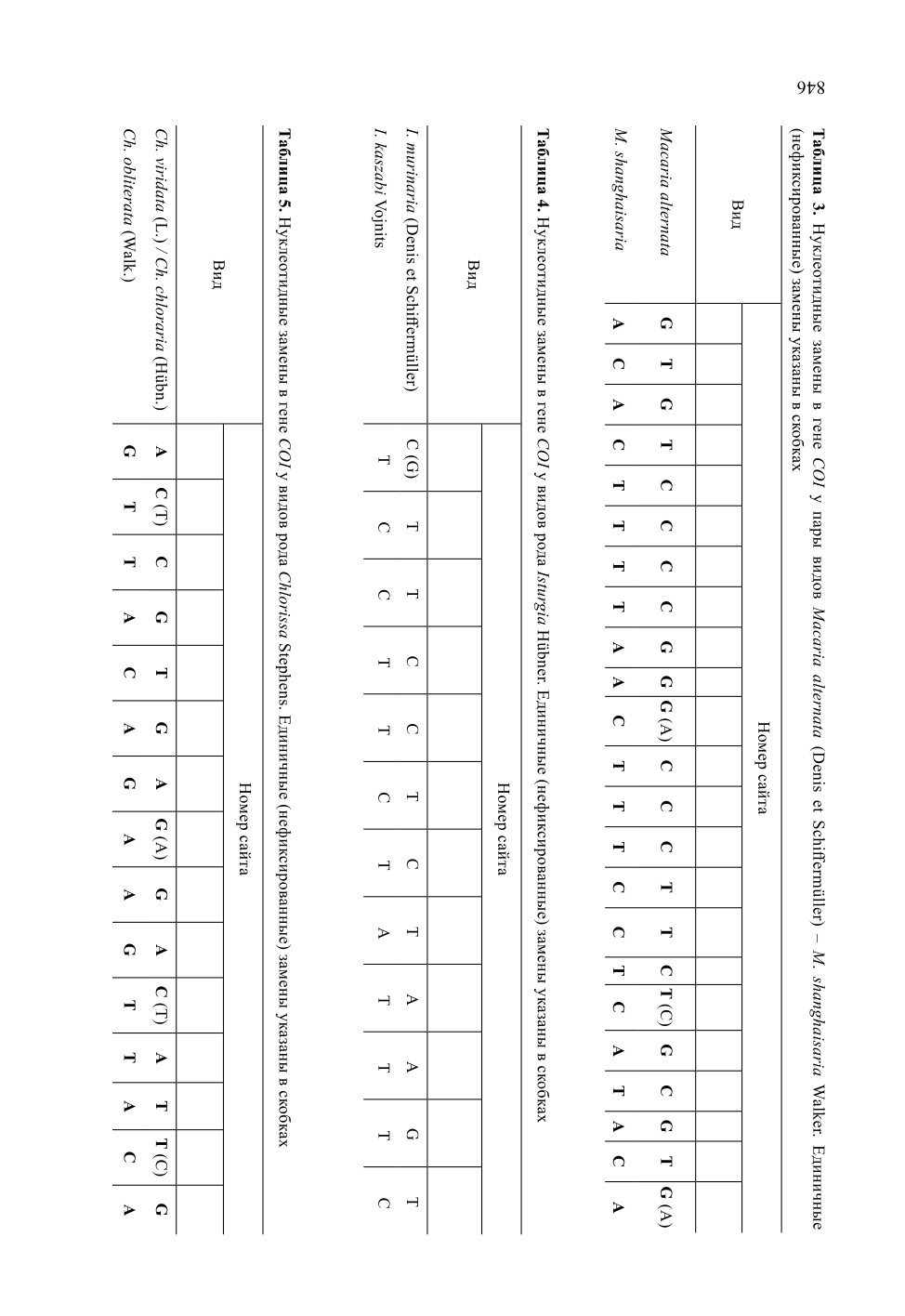
Chlorissa viridata (Linnaeus, 1758) - Chlorissa obliterata (Walker, 1863).
Ранг Ch. obliterata был понижен Леро (Leraut, 2009) до подвида Ch. viridata
(Linnaeus, 1758), однако впоследствии его видовой статус был восстановлен (Беляев,
2016; Müller et al., 2019). Ранее этот вид ошибочно указывался из Байкальского ре-
гиона под названием Ch. viridata (L.) (Гордеева, Гордеев, 2007: 129). Действительно,
он едва ли морфологически отличим от Ch. viridata / Ch. chloraria, но хорошо диффе-
ренцирован генетически (табл. 5). Виды различаются 11 фиксированными заменами
845
34
46
88
106
43
88
118
133
103
139
271
154
118
157
316
166
187
352
214
220
412
268
391
433
271
394
446
424
397
427
493
448
403
499
487
421
547
542
544
562
508
589
607
592
512
611
619
628
535
631
(p-дистанция - 1.7 %). Таким образом, достоверная регистрация Ch. obliterata в ре-
гионе стала возможной лишь благодаря использованию ДНК-баркодинга.
Примеры возможной синонимии и изменения таксономического статуса
Hemistola zimmermanni (Hedemann, 1879), H. intermedia Djakonov, 1926 и H. veneta
(Butler, 1879).
Дьяконов (1926) описал из окрестностей Минусинска H. intermedia, близкий к
H. zimmermanni (Hedemann, 1879), указав в качестве диагностического признака более
короткие гребни антенн. Позже Верли (Wehrli, 1929) предположил, что H. intermedia
может быть лишь формой H. zimmermanni либо не полностью дифференцированным
от H. chrysoprasaria молодым видом. Е. А. Беляев вместе с В. Г. Мироновым считают
(Беляев, 2016; Беляев, Миронов, 2019), что H. intermedia заслуживает видового ста-
туса. Молекулярно-генетический анализ мог бы внести ясность в этот вопрос. Среди
бабочек, собранных первым автором этой работы в Иркутской обл., Бурятии и Забай-
кальском крае, нет экземпляров с укороченными гребнями антенн. Анализ их бар-
кодов (n = 10, GenBank: MZ148328-MZ148335, OK073174, OK156317) показал, что
бабочки в этой выборке генетически гомогенны по COI (максимальная внутригруп-
повая нескорректированная p-дистанция - 1.7 %). На основе полученных нами бар-
кодов и последовательностей, доступных в базах данных GenBank и BoLD, было по-
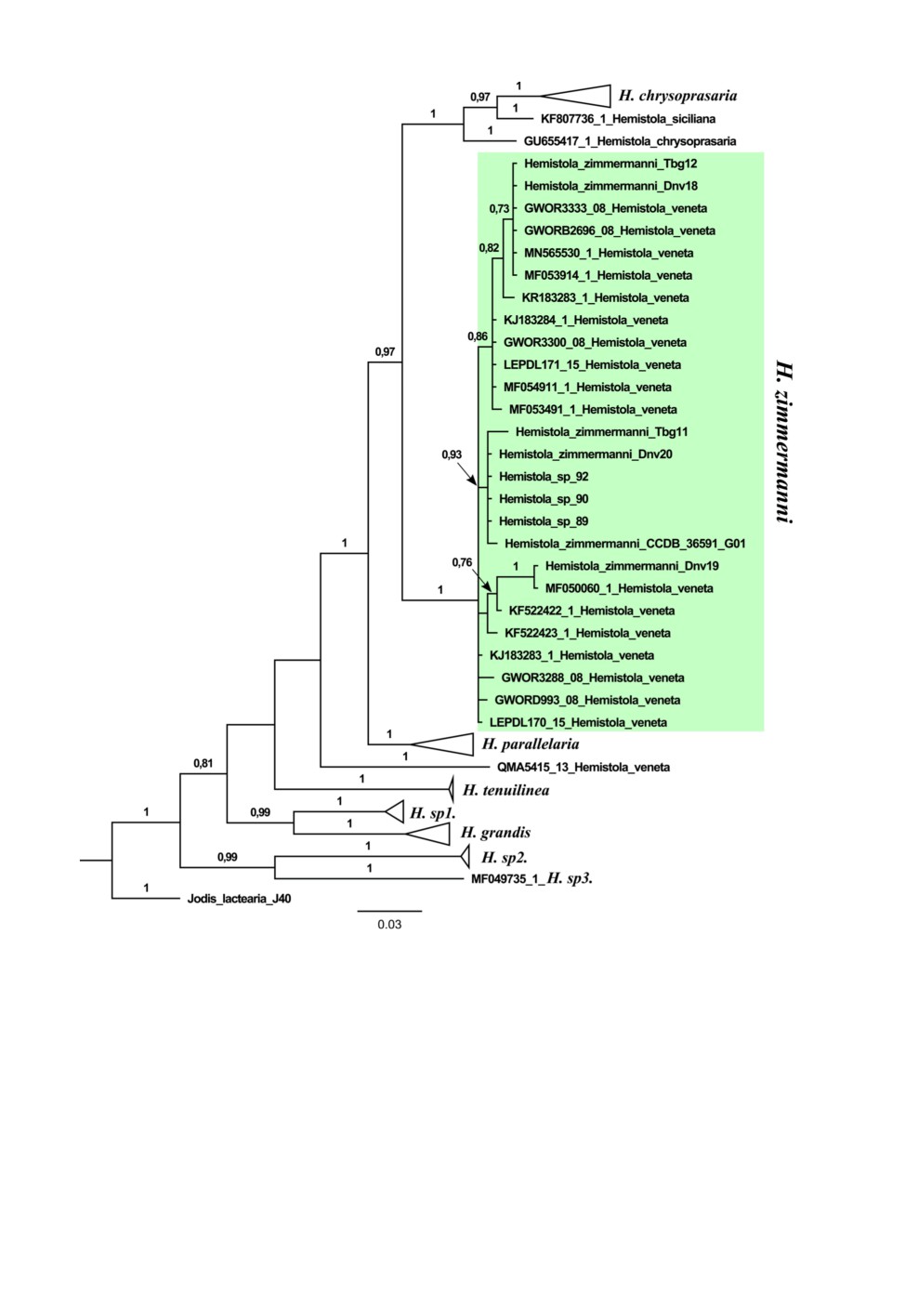
строено Байесово дерево (рис. 2). Интересующая нас клада (выделена зеленым)
объединяет все образцы H. zimmermanni из Сибири и ряд образцов из Китая, которые
определены как H. veneta (Butler, 1879). Мы изучили изображения этих пядениц, до-
ступные в базе BoLD, и пришли к выводу, что они определены неверно и в действи-
тельности относятся к H. zimmermanni (в трактовке Хана и Сю: Han, Xue, 2009). Клада,
сестринская к (H. chrysoprasaria + H. siciliana + H. zimmermanni + H. parallelaria),
также обозначена как H. veneta (единственный образец: QMA5415-13). Hemistola
zimmermanni, H. chrysoprasaria, H. siciliana и H. veneta довольно близки между собой,
и можно было ожидать, что настоящий H. veneta кластеризуется с тремя первыми
видами. Однако на полученном дереве H. veneta (QMA5415-13) ответвляется еще до
H. parallelaria, который определенно принадлежит к другой видовой группе. Бабочка
на фото H. veneta QMA5415-13 довольно потертая, поэтому трудно сказать, к какому
виду она относится. В связи с этим вопрос о синонимии H. zimmermanni и H. veneta
остается открытым и требует изучения типового материала. В выделенной зеленым
цветом кладе обнаруживаются лишь одна хорошо поддержанная (апостериорная веро-
ятность = 1) подклада: Dnv19+MF050060.1 и одна подклада со средним значением
поддержки (апостериорная вероятность = 0.93); поддержки остальных подклад
слабые. Образцы, формирующие два упомянутых значительно поддержанных кла-
стера, не соответствуют признакам предполагаемого H. intermedia, поэтому мы
склонны считать, что в Иркутской обл., Бурятии и Забайкальском крае распространен
только H. zimmermanni.
Nebula mongoliata (Staudinger, 1897) и Nebula korschunovi (Viidalepp, 1976),
comb. n.
Мы получили баркоды нескольких бабочек, отвечающих признакам рода Nebula
Bruand, 1846, собранных в разных местах в пределах Байкальского региона (см. «Ма-
териал»). Эти экземпляры различаются по размеру, форме крыльев и окраске; часть их
847
Рис. 2. Байесовское дерево Hemistola, основанное на анализе гена COI. Числа над ветвями
указывают Байесовскую апостериорную вероятность. Поддержки ниже 0.7 не показаны.
соответствует описанию Cidaria mongoliata Staudinger, 1897, а часть - Coenotephria
korschunovi Viidalepp, 1976. Анализ ДНК показал их генетическую гомогенность по
COI, а изучение гениталий не выявило значимых различий, которые можно было бы
трактовать как видовые. Это ставит вопрос о синонимизации Coenotephria korschunovi
c N. mongoliata, но для этого необходим генетический анализ особей C. korschunovi из
типовой местности (Хакасия). Кроме того, остается неясным статус Nebula egenata
848
(Prout, 1914), описанного из Сибири (р. Иркут) без приведения изображения, который
также может оказаться синонимом N. mongoliata.
М ате риа л. Красноярский край: Минусинский р-н, окр. Минусинска, 21.VI.1936, 1 ♀ (leg.
Кожанчиков) [ЗИН]. Иркутская обл.: Иркутский р-н, окр. пос. Б. Коты (51°54ʹ12ʺ N, 105°04ʹ08ʺ E),
30.VII.2016, 1 ♂; там же, 2.VIII.2016, 1 ♀ [ИСЭЖ]; там же, 3.VIII.2016, 1 ♂, 1 ♀ (GenBank:
OK156360) (leg. И. Махов) [ЗИН]. Бурятия: Тарбагатайский р-н, долина р. Селенга, 6 км С
пос. Тарбагатай (51°32ʹ37ʺ N, 107°21ʹ41ʺ E), 20.V.2021, 1 ♂ (GenBank: OK314970). Джидинский
р-н, долина р. Джида, 7 км ЮЗ пос. Петропавловка (50°32ʹ44ʺ N, 105°15ʹ38ʺ E), 9.VIII.2020, 1 ♂
(GenBank: OK073226) (leg. И. Махов) [ЗИН]. Иволгинский р-н, окр. Улан-Удэ, 17.V.1997, 1 ♂
(GenBank: OK073260). Заиграевский р-н, окр. пос. Онохой, Добо-Енхор, 14-15.VI.2012, 1 ♀
(GenBank: OK073188) (leg. Т. Гордеева) [БНЦ].
З а м еча ни е. Таксон korschunovi был описан как отдельный вид в составе рода
Coenotephria Prout 1914 (Вийдалепп, 1976). Позже он был приведен Я. Вийдалеппом
как Lampropteryx albigirata korschunovi (Viidalepp, 1996: 27). Беляев (2016: 623) спра-
ведливо отмечает, что морфологически он не соответствует C. albigirata. Однако впо-
следствии трактовка этого таксона в составе рода Lampropteryx Stephens, 1831 была
поддержана в первом и втором изданиях Каталога чешуекрылых России (Миронов и
др., 2008: 217; Беляев, Миронов, 2019: 266), где он вновь рассматривается в качестве
самостоятельного вида. Мы согласны, что отнесение C. korschunovi к роду
Coenotephria неверно: в гениталиях самца щетинки (hamuli) на лабидах нечленистые,
тогда как у Coenotephria щетинки двучлениковые. Более того, в описании C. kor-
schunovi и иллюстрации к нему отражены важные детали строения гениталий самца,
которые не характерны и для представителей рода Lampropteryx: эдеагус S-образной
формы, везика лишена корнутусов, в то время как у видов Lampropteryx эдеагус более
или менее прямой, а везика несет группу корнутусов. Окраска и рисунок крыльев
у изученного голотипа (этикетки: «Хакассия, 8.8.69. Бирикчуль. det. Viidalepp 197[?]»,
«preparaat K-342♂ Viidalepp», ИСиЭЖ СО РАН, Новосибирск) также не соответствуют
признакам Lampropteryx. По совокупности признаков C. korschunovi в большей сте-
пени соответствует роду Nebula. Таким образом, мы предлагаем рассматривать этот
вид как Nebula korschunovi comb. n.
Asthena anseraria (Herrich-Schäffer, 1855) и Asthena corculina Butler, 1878.
Некоторые авторы (Xue, Scoble, 2002; Choi, 2012; Kim et al., 2018) считают
A. corculina подвидом A. anseraria, другие рассматривают A. anseraria и A. corculina
как самостоятельные виды (Беляев, 2016; Беляев, Миронов, 2019). Впервые полу-
ченный нами баркод забайкальского экземпляра A. corculina (GenBank: OK073094) от-
личается от европейских A. anseraria на 1.0-1.34 %, что скорее свидетельствует в
пользу подвидового статуса A. corculina. Однако между этими таксонами существует
обширная дизъюнкция в Сибири от Южного Урала до Забайкалья, из области которой
их находки неизвестны. Указания A. corculina для Иркутской обл. (Берлов, Берлов,
2006: 105) ошибочны и относятся к A. amurensis (Staudinger, 1897) (проверены их изо-
возможно, что A. anseraria и A. corculina - аллопатричные виды с незначительной ге-
нетической дифференциацией по COI. Требуются дальнейшие исследования.
849
Признаки возможной митохондриальной интрогрессии
Cleta jacutica Viidalepp, 1976.
Полученный нами ДНК-баркод саянского экземпляра Cleta (GenBank: OK073236)
полностью совпал с баркодами экземпляров из западной Монголии и Крыма (BoLD:
GWOSU212-11; GBLAD323-14), определенными как C. perpusillaria. В то же время
изученный нами баркод якутского образца Cleta (GenBank: MZ148325) отличается от
баркодов этих бабочек на 4.2 %. По строению гениталий (форма вальвы и юксты:
рис. 3, 1-8, отмечены стрелками) экземпляры из Бурятии, с Алтая и из Якутии
(см. «Материал») однозначно соответствуют признакам C. jacutica. Таким образом,
можно предположить возможный генетический обмен между восточными популя-
циями C. perpusillaria и южными популяциями C. jacutica (путем межвидовой гибри-
дизации с последующей митохондриальной интрогрессией). Требуется дальнейшее
изучение вопроса на более обширном материале.
Мат ер иа л. Республика Алтай. Кош-Агачский р-н, хр. Чихачева: оз. Киндыктыкуль, 2550 м
над ур. м., 24.VII.2001, 1 ♂ (leg. В. Ковтунович) (гениталии - рис. 3, 7, 8); 13 км В пос. Кокоря
(49°57ʹ09ʺ N, 89°11ʹ02ʺ E), 11-12.VII.2017, 1 ♂ (leg. С. Князев). Бурятия: Окинский р-н, 6.5 км
ЮЮЗ пос. Самарта, 2000 м (52°02ʹ06ʺ N, 101°06ʹ26ʺ E), 18.VII.2019, 5 ♂ (leg. И. Махов) (генита-
лии - рис. 3, 1, 2). Тункинский р-н, окр. пос. Монды, 2000 м над ур. м., 14.VII.1965, 1 ♂
(leg. К. Б. Городков) (гениталии - рис. 3, 3, 4) [ЗИН]. Баунтовский р-н, 43 км С пос. Багдарин, окр.
оз. Талой, 18.VII.2007, 2 ♂(leg. Т. Гордеева) (гениталии - рис. 3, 5, 6) [БНЦ]. Якутия: Момский
р-н, хр. Черского, окр. с. Хонуу, тубдиспансер, 6.VII.2013, 1 ♂ (leg. С. Ноговицына) [ИСЭЖ].
Помимо этого вида, еще у 6 номинальных видов пядениц в Прибайкалье (Alcis
deversata Stgr., A. extinctaria Ev., Aspitates gilvaria Den. et Schiff., Ectropis crepuscularia
Den. et Schiff., Thalera chlorosaria Graes., Scopula immutata L.) в пределах одной попу-
ляции обнаружены два типа резко дифференцированных митохондриальных ДНК-бар-
кодов. Этот феномен может означать, что симпатрично встречающиеся дифференци-
рованные баркоды представляют нераспознанные криптические виды. Кроме того, это
может объясняться вторичным слиянием популяций, которые долгое время эволюцио-
нировали в условиях географической изоляции и накопили генетические различия,
или же интрогрессией от другого вида посредством межвидовой гибридизации. Те-
стирование этих альтернативных гипотез возможно с использованием дополни-
тельных ядерных генетических маркеров, что недавно было сделано для Alcis
deversata, A. extinctaria и Thalera chlorosaria (Makhov et al., 2021).
Выявление опубликованных в литературе и в публичных базах данных случаев
ошибочной видовой идентификации
Мы использовали функцию Identification Engine, реализованную на платформе
BoLD Systems (https://www. BoLDsystems.org/index.php/IDS_OpenIdEngine) и инстру-
cgi), для сравнения всех полученных нами сиквенсов с имеющимися в публичном до-
ступе. Во многих случаях наши определения не совпали с опубликованными, и мы
проверили исходные данные для образцов, баркоды которых в публикациях оказались
наиболее сходными с нашими. Были изучены изображения ваучерных экземпляров и
данные об их местонахождении. В результате было выявлено 28 ошибочных опреде-
лений, которые приведены в табл. 6.
850
Рис. 3. Cleta Duponchel, гениталии самцов.
1-8 - Cleta jacutica Viidalepp; 9-11 - C. perpusillaria Eversmann, 1847. 1, 3, 5, 7, 9, 11 - генитальный сегмент
вентрально; 2, 4, 6, 8, 10 - эдеагус, латерально. Стрелками указаны отличительные признаки C. jacutica
и C. perpusillaria (пояснения в тексте).
Масштабная линейка - 1 мм.
851
Таблица 6. Исправления ошибочных определений видов сем. Geometridae в базах данных
BoLD Systems и GenBank по результатам анализа ДНК-баркодов экземпляров из Прибайкалья.
В квадратных скобках указаны номера баркодов, размещенных нами в базе GenBank
Определение
Исправленное
Идентификационный номер
в базе данных
определение
«Eilicrinia parvula»
BIN BoLD:AAC9618
Eilicrinia unimacularia
Püngeler, 1914
[OK073217]
«Heterophleps fusca»
GenBank: MF054615, MF053586,
Astegania honesta
MF053511, MF051923, MF051469,
(Prout, 1908)
Опубликовано в: Hao et al.,
MF051212, MF050531, MF050364,
[OK156196]
2020
MF049553
BIN BoLD:AAC0759
«Jankowskia athleta»
GenBank: KJ183296.
Jankowskia
bituminaria
Опубликовано в: Jin et al.,
BoLD: GWOSU218-11, PEMOD896-20,
(Lederer, 1853)
2018
GWOR3243-08, GWOR3183-08,
[OK156333]
GWOR3241-08, GWOR3234-08,
GWOR3220-08, GWOR3382-08,
GWORK1468-15, GWOUG191-20
«Jankowskia fuscaria»
BoLD: GWORB2820-08,
GWORB2819-08, GWORB2770-08,
GWORD914-08
«Lomaspilis marginata»
ID в ограниченном доступе
Calcaritis pallida
Hedemann, 1881
«Dysgonia obscura»
GenBank: MF053212
[OK156204].
«Macaria signaria»
BoLD: GWOTL956-13
Macaria continuaria
(Eversmann,
1852) [OK156348,
OK156349]
«Macaria wauaria»
BoLD: GWORA2085-09, PEMOD953-20,
Macaria circumflexaria
PEMOD892-20
(Eversmann, 1848)
[OK073215]
«Macaria sp.»
BoLD: PEMOD911-20, PEMOD888-20,
Chiasmia saburraria
GWORL1150-16, GWORL1126-16,
(Eversmann, 1851)
GWOUG225-20, GWOSU217-11,
[OK073259].
GWOSU221-11, GWOSU262-11,
GWOSU229-11, GWOSR373-11
«Isturgia sp.»
BoLD: GWORB2823-08,
GWORB2880-08, GWORB2845-08
«Comibaena cassidara»
BoLD: GWOTL766-13
Comibaena amoenaria
(Oberthür, 1880)
«Comibaena quadrinotata»
GenBank: MF052099
[OK073119,
Опубликовано в: Hao et al.,
OK073237]
2020
852
Таблица 6 (продолжение)
Определение
Исправленное
Идентификационный номер
в базе данных
определение
«Hemistola veneta»
BoLD: GWOR3333-08, GWORB2696-08,
Hemistola
GWOR3300-08, LEPDL171-15,
zimmermanni
Опубликовано в: Jin et al.,
GWOR3288-08, GWORD993-08,
(Hedemann, 1879)
2018; Hao et al., 2020
LEPDL170-15
[MZ148328-
MZ148335]
GenBank: MN565530, MN565531,
MN565532, MN565533, MF053914,
KR183283, KJ183284, MF054911,
MF053491, MF050060, KF522422,
KF522423, KJ183283
«Pelurga comitata»
GenBank: MF050952
Pelurga
taczanowskiaria
Опубликовано в: Hao et al.,
(Oberthür, 1880)
2020
[OK073120]
«Paracolax tristalis»
GenBank: MF052802
Xenortholitha
propinguata
Опубликовано в: Hao et al.,
(Kollar, 1844)
2020
[OK156397]
«Pelurga comitata»
GenBank: MF050723, MF049687,
MF049691, MF049831
Опубликовано в: Hao et al.,
2020
«Lampropteryx sp.»
BoLD: GWOR3199-08, GWORD994-08,
GWOR3124-08, GWOR3132-08,
GWOR3147-08, GWOR3161-08,
PEMOD910-20
«Xenortholitha exacra»
BoLD: GWOTL950-13
«Asthena albulata»
BoLD: GWOSN889-11, GWOSN888-11
Asthena amurensis
(Staudinger, 1897)
[OK156197,
OK156198]
«Eupithecia irriguata»
BoLD: GWOR3313-08
Eupithecia
carpophilata
Staudinger, 1897
[OK073178,
OK156281].
BoLD: GWOR3434-08, GWOR3331-08
Eupithecia dissertata
(Püngeler, 1905)
[OK073110]
«Eupithecia extensaria»
BoLD: GWORD947-08, GWOR3253-08
Eupithecia impolita
Vojnits, 1980
[OK073230]
853
Таблица 6 (продолжение)
Определение
Исправленное
Идентификационный номер
в базе данных
определение
«Eupithecia pimpinellata»
BoLD: GWORB2698-08, GWOR3175-08
Eupithecia uliata
Staudinger, 1897
[OK073148,
OK156302]
«Horisme nigrovittata»
BoLD: LEALT331-16, GWORZ671-10,
Horisme falcata
LEALT757-16, GWORD1034-08
(A. Bang-Haas,
1907)
[OK073146,
OK073218]
«Idaea foedata»
BoLD: GWOR3245-08, GWORE2071-09,
Idaea falckii
GWORE2069-09, GWORM629-09
(Hedemann, 1879)
[MZ148339]
«Idaea effusaria»
GenBank: MF055524; MF054782;
Idaea nitidata
MF053001; MF051919; MF051844;
(Herrich-Schäffer,
Опубликовано в: Hao et al.,
MF051555; MF053413; MF049550,
1861)
2020; Xie, 2020
MN646772
[OK073137]
«Scopula subpunctaria»
BoLD: INSSD484-20, PEMOD887-20,
Scopula dignata
GWORB2781-08, GWORB2776-08,
(Guenée, 1858)
GWORB2772-08, GWOUG229-20,
[MZ148350]
GWOR3378-08, GWOR3284-08,
GWOR3259-08
«Scopula beckeraria»
BoLD: GWOR3323-08; GWORB2711-08;
Scopula permutata
GWOR3330-08
(Staudinger, 1897)
[MZ148354]
Проведенная нами проверка определений показывает, что часть ошибок обуслов-
лена плохим состоянием экземпляров, не позволяющим точно диагностировать вид по
внешним признакам (например, Eupithecia irriguata, Eu. pimpinellata, Scopula
subpunctaria, Idaea effusaria). Так, мы проанализировали полный митохондриальный
геном “I. effusaria”, полученный Си (Xie, 2020), и обнаружили, что его локус COI соот-
ветствует баркоду I. nitidata, т. е. весь опубликованный митогеном принадлежит не
I. effusaria, а I. nitidata. Часть ошибок, по-видимому, связана с тем, что авторы опреде-
лений имели дело лишь с изображениями видов (их ваучерных экземпляров), которые
представляли мало знакомую им фауну (Jin et al., 2018; Hao at al., 2020). В единичных
случаях можно предполагать операционные ошибки (путаница ваучеров и их но-
меров), как в случаях с неверным определением до уровня семейства: Dysgonia
obscura (Erebidae) - Calcaritis pallida (Geometridae), Paracolax tristalis (Erebidae) -
Xenortholitha propinguata (Geometridae) (Hao et al., 2020).
854
ЗАКЛЮЧЕНИЕ
С учетом представленных дополнений фауна пядениц Байкальского региона насчи-
тывает 361 вид из 146 родов.
Создана библиотека ДНК-баркодов пядениц Байкальского региона, которая вклю-
чает 631 нуклеотидную последовательность фрагмента гена COI для 312 видов. Для
17 видов ДНК-баркоды получены впервые; для 298 видов впервые получены ДНК-бар-
коды популяций, обитающих в Байкальском регионе. 97 % изученных ДНК-баркодов
оказались видоспецифичными, т. е. отличными от баркодов близких видов. В боль-
шинстве случаев эти различия превышали уровень в 2-3 % (т. е. наблюдались более
чем в 13-17 нуклеотидных позициях из 658 изученных). Показано, что надежная ви-
довая идентификация возможна для трех пар видов, Macaria alternata и M. shan-
ghaisaria, Isturgia murinaria и I. kaszabi, а также Chlorissa viridata и Ch. obliterata,
которые не имеют устойчивых диагностических признаков в окраске и рисунке кры-
льев и в строении гениталий обоих полов. Использование созданной библиотеки
ДНК-баркодов делает возможным идентификацию 302 видов региональной фауны,
причем эти определения можно производить не только по имаго, но и по любым преи-
магинальным стадиям, а также по фрагментам тела насекомого.
Выявлено 10 пар видов, в которых баркоды не позволяют достоверно идентифици-
ровать вид, полностью (или почти полностью) совпадая у двух близких таксонов либо
частично перекрываясь у некоторых популяций. Тем не менее, показано, что с учетом
данных по географическому распространению только 10 видов, населяющих Байкаль-
ский регион, невозможно делимитировать на основании ДНК-баркодов.
Показано, что у 6 номинальных видов пядениц в Прибайкалье (Alcis deversata Stgr.,
A. extinctaria Ev., Aspitates gilvaria Den. et Schiff., Ectropis crepuscularia Den. et Schiff.,
Thalera chlorosaria Graes., Scopula immutata L.) обнаруживается два типа дифференци-
рованных баркодов (= митохондриальных гаплогрупп) в одной популяции. Необхо-
димо дальнейшее изучение этих популяций с целью тестирования двух альтерна-
тивных гипотез: пары симпатричных гаплогрупп представляют 1) нераспознанные
криптические виды и 2) отражают внутривидовой полиморфизм.
В ходе анализа полученных ДНК-баркодов и их сравнения с доступными сиквен-
сами в базах данных сети Интернет было выявлено и исправлено 28 ошибочных опре-
делений видов по нуклеотидным последовательностям. Выявление таких ошибок
представляется критически важным, так как, попадая в базы генетических данных
ВoLD и GenBank, они существенно затрудняют работу с нуклеотидными последова-
тельностями, например, при их включении в филогенетический анализ или при прове-
дении молекулярной экспресс-идентификации видов, основанной на сравнении
ДНК-баркода неопределенного экземпляра с библиотеками ранее полученных бар-
кодов.
Девять видов региональной фауны значительно отличаются по ДНК-баркодам от по-
пуляций этих видов в других частях Голарктики. У 5 из 9 видов (Yezognophos vittaria,
Aspitates gilvaria, A. taylorae, Perizoma hydrata и Eupithecia pernotata) выявленные ми-
тохондриальные линии демонстрируют значительную генетическую дистанцию
(2-3 %) с гаплогруппами, которые были установлены для этих видов ранее. Эти
данные свидетельствуют о необходимости таксономических ревизий комплексов, в
855
которые входят виды с новыми гаплогруппами, с целью проверки гипотезы о том, что
носители этих гаплогрупп относятся к еще не описанным видам.
БЛАГОДАРНОСТИ
Авторы искренне благодарят Е. А. Беляева (Владивосток) и В. Г. Миронова (С.-Пе-
тербург) за помощь в определении ряда видов и ценные консультации. Авторы также
признательны О. А. Аненхонову (Улан-Удэ), М. К. Дементьевой, Е. В. Софроновой,
А. П. Софронову, В. В. Чепиноге (Иркутск), Д. В. Казакову (Тюмень) за плодотворное
сотрудничество и организацию полевых исследований; Э. Я. Берлову, В. Г. Шилен-
кову, А. В. Косареву (Иркутск), А. В. Филиппову (Улан-Удэ) и С. А. Князеву (Омск) за
предоставленные материалы по пяденицам.
Авторы также благодарны Т. В. Гордеевой за возможность работы с коллекционным
фондом БНЦ СО РАН (Улан-Удэ), С. В. Василенко за помощь в обработке материала
коллекции ИСиЭЖ СО РАН (Новосибирск) и Е. Ю. Городиловой (ресурсный центр
«Развитие молекулярных и клеточных технологий» Санкт-Петербургского государ-
ственного университета) за секвенирование образцов ДНК.
ФИНАНСИРОВАНИЕ
Сбор материала выполнен в рамках гостем № AAAA-A19-119020690101-6 и № АА-
АА-А19-119020790106-0. Молекулярные исследования и ДНК-баркодинг выполнены
при финансовой поддержке Российского научного фонда (грант № 19-14-00202
Зоологическому институту РАН).
Молекулярные исследования выполнены с использованием оборудования Центра
коллективного пользования оборудованием «Хромас» Ресурсного парка Санкт-Петер-
бургского государственного университета (препарирование образцов и секвениро-
вание ДНК) и Центра коллективного пользования оборудованием «Таксон» Зоологи-
ческого института РАН (фотографирование препаратов генитальных структур).
СПИСОК ЛИТЕРАТУРЫ
Белова Н. А. 2000. Высшие разноусые чешуекрылые Байкальского заповедника. Красноярск: Поликом,
144 с.
Беляев Е. А. 2016. Сем. Geometridae - Пяденицы. В кн.: А. С. Лелей (ред.). Аннотированный каталог насеко-
мых Дальнего Востока России. Т. 2. Чешуекрылые. Владивосток: Дальнаука, с. 518-666.
Беляев Е. А., Миронов В. Г. 2019. Geometridae. В кн.: С. Ю. Синёв (ред.). Каталог чешуекрылых (Lepidoptera)
России. Издание 2-е. СПб.: Зоологический институт РАН, с. 235-281.
Берлов О. Э., Берлов Э. Я. 2006. Материалы к фауне и экологии пядениц (Geometridae) Прибайкалья. Труды
государственного природного заповедника «Байкало-Ленский». Иркутск: РИО НЦ РВХ ВСНЦ СО
РАМН, выпуск 4, с. 102-110.
Василенко С. В. 2006. Пяденицы (Lepidoptera, Geometridae) лесостепной зоны Западно-Сибирской равнины.
Евразиатский энтомологический журнал 5 (3): 215-219.
Василенко С. В. 2014. Обзор видов Euphyia Hübner, 1816 (Lepidoptera: Geometridae, Larentiinae) азиатской
части России. Евразиатский энтомологический журнал 13 (2): 154-160.
Васильева Т. Г. 1989. К фауне пядениц (Lepidoptera, Geometridae) Южного Прибайкалья. В кн.: В. Г. Шилен-
ков, А. С. Плешанов, Е. П. Бессолицына, Л. Н. Дубешко, Н. А. Никулина (ред.). Насекомые и пауко-
образные Сибири. Иркутск: Иркутский государственный университет, c. 104-115.
Вийдалепп Я. Р. 1976. Список пядениц (Lepidoptera, Geometridae) фауны СССР. I. Энтомологическое обозре-
ние 55 (4): 842-852.
856
Вийдалепп Я. P. 1987. К фауне пядениц Эвенкийского автономного округа и зоны БАМ. В кн.: А. С. Рожков
(ред.). Насекомые зоны БАМ. Новосибирск: Наука, Сибирское отделение, с. 74-82.
Вийдалепп Я. Р. 1988. Фауна пядениц гор Средней Азии. М.: Наука, 240 с.
Гордеева Т. В., Гордеев С. Ю. 2007. Семейство Geometridae - пяденицы. В кн.: Л. Л. Убугунов, В. В. Дубато-
лов (ред.). Чешуекрылые Бурятии. Новосибирск: Сибирское отделение РАН, с. 95-148.
Городков К. Б. 1984. Типы ареалов насекомых тундры и лесных зон европейской части СССР. В кн.: К. Б. Го-
родков (ред.). Ареалы насекомых европейской части СССР, Атлас. Л.: Наука, с. 3-20.
Городков К. Б. 1985. Трехмерная климатическая модель потенциального ареала и некоторые ее свойства.
Ч. I. Энтомологическое обозрение 64 (2): 295-310.
Городков К. Б. 1986. Трехмерная климатическая модель потенциального ареала и некоторые ее свойства.
Ч. II. Энтомологическое обозрение 65 (1): 81-95.
Городков К. Б. 1992. Типы ареалов двукрылых (Diptera) Сибири. В кн.: Э. П. Нарчук (ред.), Систематика,
зоогеография и кариология двукрылых насекомых (Insecta: Diptera). СПб.: Зоологический институт
РАН, с. 45-55.
Дьяконов А. М. 1926. К познанию фауны Geometridae Минусинского края. Ежегодник Государственного му-
зея имени Н. М. Мартьянова 4 (1): 1-78.
Махов И. А. 2021. Дополнения ко второму изданию Каталога чешуекрылых России по трем восточносибир-
ским регионам. Часть
1. Энтомологическое обозрение
100
(2):
375-389. doi:
10.31857/
S0367144521020088
Миронов В. Г., Беляев Е. А., Василенко С. В. 2008. Geometridae. В кн.: С. Ю. Синёв (ред.). Каталог чешуе-
крылых (Lepidoptera) России. СПб.; М.: Товарищество научных изданий КМК, с. 190-226.
Choi S.-W. 2012. Taxonomic review of the genus Asthena Hübner (Lepidoptera: Geometridae) in Korea.
Entomological Research 42: 151-157. doi: 10.1111/j.1748-5967.2012.00450.x
deWaard J. R., Hebert P. D. N., Humble L. M. 2011. A comprehensive DNA barcode library for the looper moths
(Lepidoptera: Geometridae) of British Columbia, Canada. PLoS ONE 6 (3): e18290. doi:10.1371/journal.
pone.0018290
deWaard J. R., Ivanova N. V., Hajibabaei M., Hebert P. D. N. 2008. Assembling DNA barcodes: analytical protocols.
In: С. С. Martin (ed.). Environmental Genomics. (Methods in Molecular Biology 410). Totowa, New Jersey:
Humana Press, p. 275-283. doi: 10.1007/978-1-59745-548-0_15
Feizpour S., Fekrat L., Namaghi S., Stadie D., Rajaei H. 2018. Combination of morphological characters and DNA-
barcoding confirms Problepsis cinerea (Butler, 1886) (Geometridae: Sterrhinae: Scopulini) as a new genus
and species for the fauna of Iran. Integrative Systematics: Stuttgart Contributions to Natural History 1 (1):
47-57. doi: 10.18476/insy.v01.a6
Folmer O., Black M., Hoeh W., Lutz R., Vrijenhoek R. 1994. DNA primers for amplification of mitochondrial
cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and
Biotechnology 3: 294-299.
Hajibabaei M., Singer G. A. C., Hebert P. D. N., Hickey D. A. 2007. DNA barcoding: how it complements taxonomy,
molecular phylogenetics and population genetics. Trends in Genetics 23: 167-172. doi: 10.1016/j.
tig.2007.02.001
Hall T. A. 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows
95/98/NT. Nucleic Acids Symposium Series 41: 95-98.
Han H., Xue D. 2009. Taxonomic review of Hemistola Warren, 1893 from China, with descriptions of seven new
species (Lepidoptera: Geometridae, Geometrinae). Entomological Science
12:
382-410. doi:
10.1111/j.1479-8298.2009.00341.x
Hao M., Jin Q., Meng G., Yang C., Yang Sh., Shi Zh., Tang M., Liu Sh., Li Y., Li J., Zhang D., Su X., Shih Ch., Sun
Y., Wilson J.-J., Zhou X., Zhang A. 2020. Using full-length metabarcoding and DNA barcoding to infer
community assembly for speciose taxonomic groups: a case study. Evolutionary Ecology 34 (9). doi: 10.1007/
s10682-020-10072-y
Hausmann A. 2004. Sterrhinae. In: A. Hausmann (ed.). The Geometrid Moths of Europe. Vol. 2. Stenstrup: Apollo
Books, 600 p.
Hausmann A., Godfray H. C. J., Huemer P., Mutanen M., Rougerie R., van Nieukerken E. J., Ratnasingham S.,
Hebert P. D. N. 2013. Genetic patterns in European geometrid moths revealed by the Barcode Index Number
(BIN) System. PLoS ONE 8 (12): e84518.d. doi: 10.1371/journal.pone.0084518
Hausmann A., Haszprunar G., Hebert P. D. N. 2011. DNA Barcoding the Geometrid Fauna of Bavaria (Lepidoptera):
Successes, Surprises, and Questions. PLoS ONE 6 (2): e17134. doi: 10.1371/journal.pone.0017134
Hausmann A., Viidalepp J. 2012. Subfamily Larentiinae 1. In: A. Hausmann (ed.). The Geometrid Moths of Europe.
Vol. 3. Stenstrup: Apollo Books, 743 p.
Hebert P. D. N., Cywinska A., Ball S. L., de Waard J. R. 2003a. Biological identifications through DNA barcodes.
Proceedings of the Royal Society B: Biological Sciences 270: 313-321. doi: 10.1098/rspb.2002.2218
857
Hebert P. D. N., Ratnasingham S., deWaard J. R. 2003b. Barcoding animal life: cytochrome c oxidase subunit 1
divergences among closely related species. Proceedings of the Royal Society B: Biological Sciences 270:
S96-S99. doi: 10.1098/rsbl.2003.0025
Hebert P. D. N., Ratnasingham S., Zakharov E. V., Telfer A. C., Levesque-Beaudin V., Milton M. A., Pedersen S.,
Janetta P., deWaard J. R. 2016. Counting animal species with DNA barcodes: Canadian insects. Philosophical
Transactions of the Royal Society B: Biological Sciences, 371 (1702): 20150333. doi:10.1098/rstb.2015.0333
Jin Q., Hu X. M., Han H. L., Chen F., Cai W.-J., Ruan Q.-Q., Liu B., Luo G.-J., Wang H., Liu X., Ward R. D., Wu
Ch.-Sh., Wilson J.-J., Zhang A.-B. 2018. A two-step DNA barcoding approach for delimiting moth species:
moths of Dongling Mountain (Beijing, China) as a case study. Scientific Reports 8: 14256. doi: 10.1038/
s41598-018-32123-9
Kim N.-H., Choi S.-W., Kim S.-S. 2018. Additional Report of the Genus Asthena (Lepidoptera: Geometridae) from
Korea. Animal Systematics, Evolution and Diversity 34 (2): 92-95. doi: 10.5635/ASED.2018.34.2.016
Kumar S., Stecher G., Tamura K. 2015. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger
datasets. Molecular Biology and Evolution 33: 1870-1874. doi: 10.1093/molbev/msw054
Leraut P. 2009. Moths of Europe - Volume 2: Geometrid Moths. Paris: NAP Editions, 808 p.
Makhov I. A. 2021. Geometridae (Lepidoptera) of the Baikal region: keys to species and an annotated catalogue.
Part 1. Ennominae. Zootaxa 4962 (1): 1-125. doi: 10.11646/zootaxa.4962.1.1
Makhov I. A., Beljaev E. A. 2019. New data on Geometrid moths (Lepidoptera: Geometridae) of the Baikal region,
Russia. Far Eastern Entomologist 391: 1-23. doi: 10.25221/fee.391.1
Makhov I. A., Gorodilova Ye. Yu., Lukhtanov V. A. 2021. Sympatric occurrence of deeply diverged mtDNA lineages
in Siberian geometrid moths (Lepidoptera, Geometridae): cryptic speciation, mitochondrial introgression,
secondary admixture or effect of Wolbachia? Biological Journal of the Linnean Society 134 (2): 342-365.
doi: 10.1093/biolinnean/blab089/6314024
Müller B., Erlacher S., Hausmann A., Rajaei H., Sihvonen P., Skou P. 2019. Ennominae II. In: A. Hausmann,
P. Sihvonen, H. Rajaei, P. Skou (eds). Geometrid Moths of Europe. Vol. 6. Leiden: Brill, 906 p.
Murillo-Ramos L., Brehm G., Sihvonen P., Hausmann A., Holm S., Reza Ghanavi H., Õunap E., Truuverk A.,
Staude H., Friedrich E., Tammaru T., Wahlberg N. 2019. A comprehensive molecular phylogeny of
Geometridae (Lepidoptera) with a focus on enigmatic small subfamilies. PeerJ 7: e7386. doi: 10.7717/
peerj.7386
Mutanen M., Hausmann A., Hebert P. D. N., Landry J.-F., de Waard J. R., Huemer P. 2012. Allopatry as a Gordian
knot for taxonomists: Patterns of DNA barcode divergence in Arctic-Alpine Lepidoptera. PLoS ONE 7 (10):
e47214. doi:10.1371/journal.pone.0047214
Mutanen M., Kivelä S. M., Vos R. A., Doorenweerd C., Ratnasingham S., Hausmann A., Huemer P., Dincă V.,
van Nieukerken E. J., Lopez-Vaamonde C., Vila R., Aarvik L., Decaëns T., Efetov K. A., Hebert P. D. N.,
Johnsen A., Karsholt O., Pentinsaari M., Rougerie R., Segerer A., Tarmann G., Zahiri R., Godfray
H. C. J. 2016. Species-level para- and polyphyly in DNA barcode gene trees: Strong operational bias in
European Lepidoptera. Systematic Biology 65 (6): 1024-1040. doi: 10.1093/sysbio/syw044
Rajaei H., Gelbrecht J., Schulz N., Hausmann A. 2021. Minoa lutea Schwingenschuss, 1954 (Lepidoptera:
Geometridae: Larentiinae) recognized as bona species. Zootaxa 4903 (2): 255-264. doi: 10.11646/
zootaxa.4903.2.5
Rajaei H., Struwe J.-F., Raupach M., Ahrens D., Wägele W. 2013. Integration of cytochrome c oxidase I barcodes
and geometric morphometrics to delimit species in the genus Gnopharmia (Lepidoptera: Geometridae,
Ennominae). Zoological Journal of the Linnean Society 169: 70-83. doi: 10.1111/zoj.12053
Ronquist F., Teslenko M., Van Der Mark P., Ayres D. L., Darling A., Hohna S., Larget B., Liu L., Suchard M. A.,
Huelsenbeck J. P. 2012. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across
a large model space. Systematic Biology 61: 539-542. doi: 10.1093/sysbio/sys029
Rubio R. M., Guerrero J. J., Garre M., Ortiz A. S. 2013. DNA barcoding confirms the presence of Hydria cervinalis
(Scopoli, 1763) in the Iberian Peninsula (Lepidoptera: Geometridae: Larentiinae). Zootaxa 3702 (1): 97-99.
doi: 10.11646/zootaxa.3702.1.7
Silva-Brandão K. L., Lyra M. L., Freitas A. V. L. 2009. Barcoding lepidoptera: current situation and perspectives on
the usefulness of a contentious technique. Neotropical Entomology 38 (4): 441-451. doi:10.1590/s1519-
566x2009000400001
Skou P., Sihvonen P. 2015. Subfamily Ennominae 1. In: P. Skou, P. Sihvonen, A. Hausmann (eds). The Geometrid
Moths of Europe. Vol. 5. Stenstrup: Apollo Books, 657 p.
Stadie D., Hausmann A., Rajaei H. 2014. Cataclysme subtilisparsata Wehrli, 1932 (Lepidoptera, Geometridae,
Larentiinae) recognized as bona species - an integrative approach. Nota Lepidopterologica 37 (2): 141-150.
Van Nieukerken E. J., Kaila L., Kitching I. J., Kristensen N. P., Lees D. C., Minet J., Mitter C., Mutanen M.,
Regier J. C., Simonsen T. J., Wahlberg N., Yen S., Zahiri R., Adamski D., Baixeras J., Bartsch D., Bengtsson
B. Å., Brown J. W., Bucheli S. R., Davis D. R., Prins J. de, De Prins W., Epstein M. E., Gentili-Poole P.,
Gielis C., Hättenschwiler P., Hausmann A., Holloway J. D., Kallies A., Karsholt O., Kawahara A. Y.,
858
Koster J., Kozlov M., Lafontaine J. D., Lamas G., Landry J., Lee S., Nuss M., Park K., Penz C., Rota J.,
Schintlmeister A., Schmidt B. C., Sohn J., Solis M. A., Tarmann G. M., Warren A. D., Weller S.,
Yakovlev R. V., Zolotuhin V. V., Zwick A. 2011. Order Lepidoptera Linnaeus, 1758. In: Z.-Q. Zhang (ed.).
Animal biodiversity: an outline of higher-level classification and survey of taxonomic richness. Zootaxa
3148: 212-221. doi: 10.11646/zootaxa.3148.1.41
Viidalepp J. 1996. Checklist of the Geometridae (Lepidoptera) of the Former U.S.S.R. Stenstrup: Apollo Books,
111 p.
Wanke D., Hausmann A., Rajaei H. 2019. An integrative taxonomic revision of the genus Triphosa Stephens, 1829
(Geometridae: Larentiinae) in the Middle East and Central Asia, with description of two new species. Zootaxa
4603 (1): 39-65. doi: 10.11646/zootaxa.4603.1.2
Wehrli E. 1929. Beitrag zur Geometriden-Fauna von Minussinsk Sibirien, Gouv. Jenissej. Ежегодник государ-
ственного музея имени Н. М. Мартьянова 6 (1): 8-30.
Xie J.-L. 2020. Sequencing and characterization of mitochondrial genome of Idaea effusaria (Lepidoptera:
Geometridae). Mitochondrial DNA Part B 5 (1): 1001-1002. doi:10.1080/23802359.2020.1720542
Xue D., Scoble M. J. 2002. A review of the genera associated with the tribe Asthenini (Lepidoptera: Geometridae:
Larentiinae). Bulletin of the Natural History Museum: Entomology
71:
77-133. doi:
10.1017/
S0968045402000044
Yamamoto S., Sota T. 2007. Phylogeny of the Geometridae and the evolution of winter moths inferred from
a simultaneous analysis of mitochondrial and nuclear genes. Molecular Phylogenetics and Evolution 44:
711-723. doi:10.1016/j.ympev.2006.12.027
GEOMETRID MOTHS (LEPIDOPTERA, GEOMETRIDAE) OF THE BAIKAL
REGION: ADDITIONS TO THE SPECIES LIST AND RESULTS
OF DNA BARCODING
I. A. Makhov, V. A. Lukhtanov
Key words: Geometridae, fauna, Baikal Region, Irkutsk Province, Republic of Buryatia,
DNA barcoding, cytochrome c oxidase subunit I (COI).
SUMMARY
Additions to the species list of the Baikal region (Russia) are given in the present work. The
distribution of 7 geometrid species in the region is specified. Seven species are recorded in the study
area for the first time: Chlorissa obliterata (Walker, 1863), Scotopteryx acutangulata (Inoue, 1941),
Entephria byssata (Aurivillius, 1891), Minoa lutea Schwingenschuss, 1954, Eupithecia sophia Butler,
1878, Idaea dimidiata (Hufnagel, 1767), Scopula flaccidaria (Zeller, 1852), and one species, Charissa
beljaevi Erlacher, Marrero et Erlacher, 2017 is recorded for the first time in Russia. One species is newly
combined with genus Nebula Bruand, 1846: Nebula korschunovi comb. n. For the first time, a library
of DNA barcodes for Geometridae of the Baikal region is created. It includes 631 nucleotide sequences
of the COI gene fragment for 312 species (86% of the regional fauna). Analysis of these DNA barcodes
and the morphology of the individuals from which they were obtained leads to the following results:
(1) one new species (Chlorissa obliterata) for the fauna of the region is recorded; (2) 28 cases of
erroneous species identification published in the literature and in public databases are revealed;
(3) weakly morphologically differentiated sibling species Macaria alternata ([Denis et Schiffermüller],
1775) and M. shanghaisaria Walker, 1861, Isturgia murinaria ([Denis et Schiffermüller], 1775) and
I. kaszabi Vojnits, 1974, Chlorissa viridata (Linnaeus, 1758) and Ch. obliterata are unambiguously
discriminated using DNA barcodes; (4) a deep genetic differentiation of Baikal populations of 5
geometrid species (genetic divergence is about 3% or more, in comparison with European and / or
East Asian populations) is detected, which raises the question of the need for their taxonomic revision.
859