ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 101, 1, 2022
УДК 595.754
ФЕНОЛОГИЯ КЛОПА-КРАЕВИКА MOLIPTERYX FULIGINOSA
(UHLER) (HETEROPTERA, COREIDAE) НА ЮГЕ ДАЛЬНЕГО
ВОСТОКА РОССИИ
© 2022 г. Т. О. Маркова, 1* Е. В. Канюкова, 2** М. В. Маслов 1***
1 Федеральный научный центр биоразнообразия наземной биоты Восточной Азии ДВО РАН
пр. 100-летия Владивостока, 159, Владивосток, 690022 Россия
*e-mail: martania@mail.ru, ***nippon_mvm@mail.ru
2 Дальневосточный федеральный университет, Зоологический музей
Океанский пр., 37, Владивосток, 690091 Россия
**e-mail: evkany@mail.ru
Поступила в редакцию 13.06.2021 г.
После доработки 12.07.2021 г.
Принята к публикации 12.07.2021 г.
На основе шестилетнего мониторинга в естественных и приближенных к естественным усло-
виях изучено сезонное развитие Molipteryx fuliginosa (Uhler) в Приморском крае, включая сроки
откладки и развития яйца и личиночных стадий до выхода имаго. Жизненный цикл M. fuliginosa
относится к моновольтинному типу. Яйца откладываются беспорядочно на различные предме-
ты, отмечена откладка яиц на землю. Длительность эмбриональной стадии колеблется от 12 до
36 дней в зависимости от температуры воздуха. Развитие личинок после выхода из яйца до вы-
лета имаго продолжается от 55 до 83 дней. Продолжительность преимагинального развития
с момента откладки яйца до окрыления имаго составляет от 80 до 117 дней. Установлены опти-
мальные условия и сроки сезонного развития клопа в природе.
Ключевые слова: клопы, Molipteryx fuliginosa, сезонное развитие, фенология, яйцо, личинки,
зимовка, Приморский край, Heteroptera, Coreidae.
DOI: 10.31857/S0367144522010051
В фауне России род Molipteryx Kiritshenko, 1916 представлен единственным видом
M. fuliginosa (Uhler, 1860), 4 другие вида этого рода обитают на юго-востоке Палеар-
ктики и в Ориентальной области. До 90-х гг. XX века M. fuliginosa был известен только
из Северо-Восточного Китая, c Корейского п-ова и из Японии, но в 1987 и 1992 гг.
первые единичные особи были обнаружены на юге Дальнего Востока России
(Kerzhner, Kanyukova, 1998; Kanyukova, Vinokurov, 2009). Он стал единственным
видом рода Molipteryx, расширившим свой ареал на север до широты 49°26′26′′ N -
юга Хабаровского края - и сумевшим адаптироваться к новым климатическим усло-
виям. В последние годы клоп прижился на юге Дальнего Востока России, дает
вспышки численности и вредит малине (Канюкова, 2012), снижая ее урожайность.
Этот самый крупный представитель сем. Coreidae в нашей фауне стал удобным объ-
87
ектом для изучения процесса внедрения и адаптации еще одного чужеродного вида в
России (Маркова и др., 2021).
Нами были обобщены и проанализированы сведения о распространении
M. fuliginosa на российском Дальнем Востоке и первичные данные по его биотопиче-
скому распределению на юге Приморского края (Канюкова, 2012; Канюкова, Оста-
пенко, 2013; Markova et al., 2016; Маркова и др., 2016, 2021). В стационарных садках,
расположенных на приусадебном участке в условиях, приближенных к естественным,
изучена морфология преимагинальных фаз клопа, получены сведения о репродук-
тивном поведении самок и их плодовитости (Маркова и др., 2017а, 2017б).
Первые данные о цикле развития M. fuliginosa были получены в Южной Корее, где в
лабораторных условиях его наблюдали в течение одного летнего сезона (Park, 1996).
В местных климатических условиях установлены сроки появления имаго весной и
ухода на зимовку имаго следующего поколения. Собранных в природе имаго содер-
жали в инсектарии при постоянной температуре воздуха +25 оC, относительной влаж-
ности 65 % и фотопериоде 12 : 12. Установлены число откладываемых яиц, сроки раз-
вития личинок и кривая выживаемости M. fuliginosa; из отложенных 90 яиц получены
4 имаго, что привело автора к заключению о низкой выживаемости преимагинальных
стадий клопа. Прослежены сезонная динамика, поведение на малине при выборе объ-
екта питания. Сделан вывод, что клоп имеет одно поколение в году. Места зимовки
обнаружены не были.
В настоящей работе впервые представлены результаты многолетнего мониторинга
фенологии развития эмбриональной и личиночной стадий и лёта имаго M. fuliginosa
в климатических условиях юга российского Дальнего Востока.
В тексте приняты сокращения: n - число экземпляров, m - среднее значение, ЛI-
ЛV - личинки I-V возрастов.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводились с мая по октябрь 2014-2020 гг. на полевых стационарах и в поезд-
ках по Приморскому краю. Пункты стационарных исследований указаны в табл. 1.
Клопы содержались в полевых условиях в стационарных садках из сетчатой ткани, снабжен-
ной застежкой-молнией. Садками оборачивали куст живого растения, обеспечивающий доста-
точное питание и свободу передвижения насекомых (Маркова и др., 2017а, 2017б, 2018).
Проводились сравнительные наблюдения в садках и естественных ценозах с момента яйцеклад-
ки до выхода личинок I возраста, созревания и линьки последующих стадий вплоть до имаго.
Для выяснения сроков ухода в диапаузу и условий зимовки имаго нового поколения содержали
в садках в осенне-зимний период с сохранением влияния на них всех атмосферных условий
(рис. 1). Для обобщения многолетних данных по фенологии M. fuliginosa на Дальнем Востоке
привлечен также коллекционный и фотоматериал 2016-2020 гг.
Molipteryx fuliginosa (Uhler, 1860).
Мат ер иа л. Россия. Хабаровский край. Нанайский р-н: Анюйский национальный парк, бас-
сейн р. Пихца, кордон Пихур, 24.V.2019 (Н. М. Яворская), 2 экз. Хабаровский р-н: пос. Хехцир,
приусадебный участок, 21.VI.2020 (А. Ю. Олейников), 2 экз.; с. Осиновая Речка, садоводческое
некоммерческое товарищество «Лидер», 21.VII.2020 (Д. А. Тихонов), 2 экз. Приморский край.
Пожарский р-н: пгт. Лучегорск, приусадебный участок, 31.VIII.2020 (Н. А. Коляда), 1 экз. Крас-
ноармейский р-н: с. Рощино, приусадебный участок, травостой, 26.VI.2018 (А. Д. Гребенюк),
88
Таблица 1. Координаты пунктов стационарных исследований в Приморском крае (2014-
2020 гг.)
Пункт исследований
Координаты
Ханкайский р-н
с. Первомайское
044°00′05′′ N, 131°58′54′′ E
Спасский р-н
с. Спасское
44°36′52′′ N, 132°47′49′′ E
Чугуевский р-н
с. Чугуевка
44°10′00′′ N, 133°51′30′′ E
с. Новомихайловка
44°14′01′′ N, 133°51′59′′ E
Уссурийский городской округ
с. Каймановка
43°37′49′′ N, 132°13′49′′ E
Уссурийский заповедник
43°40′00′′ N, 132°30′00′′ E
4 экз. Тернейский р-н: Сихотэ-Алинский заповедник, окр. пос. Терней, пойма р. Серебрянка,
26.VI.2015 (М. Е. Сергеев), 1 экз. Дальнегорский городской округ: окр. г. Дальнегорск, проселоч-
ная дорога, 23-24.VI.2018 (Д. М. Черняк), 1 экз. Ханкайский р-н: с. Первомайское, приусадеб-
ный участок, травостой, VI-VIII.2017-2019 (А. В. Ховрина), 15 имаго, 14 личинок III-V
возрастов; с. Камень-Рыболов, приусадебный участок, 20.VI и 22.VIII.2018 (Ю. С. Заварзина),
4 экз.; с. Турий Рог, сорное придорожное разнотравье, 10.VI.2017 и 01.VII.2018 (К. А. Остапен-
ко), 2 экз.; долина р. Комиссаровка, с. Дворянка, придорожное крупнотравье, 01 и 15.VI.2017
(К. А. Остапенко), 4 экз. Спасский р-н: окр. с. Евсеевка, пойменный лес, травостой под пологом
леса, 12.V.2018 (А. К. Мрикот), 2 экз.; с. Чкаловское, приусадебный участок, 20.VII.2018
(Н. В. Репш), 1 экз.; с. Спасское, приусадебный участок, на земле, 22.V.2020 (А. К. Мрикот),
1 экз. Яковлевский р-н: с. Минеральное, кустарниковая растительность, 20.VII.2017 (С. В. Вери-
га), 2 экз.; с. Яковлевка, приусадебный участок, 16-25.VII.2020 (С. А. Макаревич), 7 имаго, 3 ли-
чинки III возраста. Черниговский р-н: с. Синий Гай, приусадебный участок, 22.VIII.2020
(Н. А. Коляда), 1 экз.; пос. Сибирцево, приусадебный участок, 26.V.2018 (Ю. Н. Глущенко),
1 экз. Анучинский р-н: г. Синегорка, травостой под пологом леса, 01.VI.2015 (С. В. Верига),
1 экз.; ключ Большой, пойма, подрост ильма долинного Ulmus propinqua, 10.VI.2017 (К. А. Оста-
пенко), 1 экз. Арсеньевский городской округ: г. Арсеньев, приусадебные участки, сорная придо-
рожная растительность, кустарниковые заросли, V-VIII.2014, 2016-2018 (К. А. Остапенко,
С. В. Верига), 10 экз.; 25.VII.2020 (А. Е. Ковалева), 4 имаго, 2 личинки III возраста. Октябрьский
р-н: пос. Чернятино, гора Синеловка, разнотравье на месте заброшенных сельскохозяйственных
угодий, 09.VII.2017, 03.VII.2018 (К. А. Остапенко), 3 экз. Чугуевский р-н: с. Кокшаровка, приу-
садебный участок, 08.VII.2020 (А. К. Мрикот), 4 экз.; с. Чугуевка, приусадебный участок, 30.V и
14.VI.2020 (А. К. Мрикот), 4 экз.; с. Новомихайловка, приусадебный участок, V-VIII.2018-2020
(З. А. Лимачко), 14 имаго, 14 личинок II-V возрастов. Уссурийский городской округ: с. Пушки-
но, травостой под пологом леса, 29.V.2018 (Ю. Н. Глущенко), 1 экз.; окраина г. Уссурийск, зверо-
совхоз (приусадебный участок), 13.VI и 20.VI.2018; 04-20.IX.2019 (Т. О. Маркова, Е. Н. Боловцов),
12 экз.; с. Горнотаежное, приусадебные участки, кустарники, разнотравье, сорная раститель-
ность, V-VIII.2016-2018 (К. А. Остапенко, М. М. Омелько), 12 имаго, 1 личинка IV возраста;
приусадебный участок, 16.IX.2019 (Н. А. Коляда), 1 экз.; питомник, 05.VIII.2020 (А. В. Куприн),
1 личинка IV возраста; с. Заречное, плодовый питомник, 23.VI.2020 (С. А. Макаревич), 1 экз.;
с. Каменушка, пойменный лес, обочина поля, 24.VIII.2016; 30.V-28.VIII.2017; 10.VI-25.
VIII.2018; 30.V-08.VIII.2019; 15.VIII и 30.IX.2020 (Т. О. Маркова, М. В. Маслов), 17 имаго,
13 личинок IV-V возрастов; с. Каймановка, опушка леса, обочина лесной дороги, приусадебный
89
Рис. 1. Стационарный садок в зимних условиях.
участок, 30.V-30.IX.2016-2020 (Т. О. Маркова, М.В. Маслов), 79 имаго, 46 личинок II-V возрас-
тов; Уссурийский заповедник, травостой под пологом леса, обочина лесной дороги, лесная поля-
на, V-IX.2016-2019 (М. В. Маслов, Л. А. Федина), 20 имаго, 2 личинки IV возраста, 3 личинки
V возраста. Шкотовский р-н: Уссурийский заповедник, придорожная растительность, кустарни-
ковые заросли, V-X.2016-2019 (М. В. Маслов, Л. А. Федина), 4 экз.; с. Анисимовка, ключ
Смольный, заросшие лесные дороги, высокотравье, 12.VII.2016 (К. А. Остапенко), 2 экз. Парти-
занский р-н: с. Сергеевка, приусадебный участок, травостой, 10.V.2016 (Е. А. Новая), 1 экз.; с.
Николаевка, приусадебный участок, 20.VIII.2019; 30.V.2020 (С. Е. Егоренчев), 5 экз.; дер. Васи-
льевка, 23-29.VIII.2020 (И. Д. Солодкий), 1 имаго, 4 личинки V возраста. Владивостокский го-
родской округ: окр. г. Владивосток, ст. Спутник, 11.V.2016 (К. А. Остапенко), 1 экз.; среднее
течение р. Богатая, под пологом долинного леса, приусадебные участки, VII.2017-2018 (С. В. Ве-
рига), 5 экз. Находкинский городской округ: г. Находка, садоводческое некоммерческое товари-
щество «Находка», 21.VII и 06.VIII.2020 (М. В. Аналеева), 2 экз. Хасанский р-н: о. Фуругельма,
луговое разнотравье, 17-22.VIII.2018 (М. Е. Сергеев), 4 личинки IV и V возрастов; п-ов Гамова,
бухта Витязь, луговое разнотравье, 12-14.VIII.2019 (М. Е Сергеев), 2 экз.; 13.IX.2019 (А. М. Ни-
90
колаева), 4 имаго, 6 личинок V возраста; 20-22.VII и 28.VII.2020 (Ю. А. Чистяков), 4 имаго, 1 ли-
чинка III возраста.
РЕЗУЛЬТАТЫ
Полный жизненный цикл клопов (период активной жизнедеятельности и диапаузы)
включает три стадии: эмбриональную, личиночную и имагинальную. Жизненный
цикл M. fuliginosa относится к моновольтинному типу (Саулич, Мусолин, 2007), он
укладывается в один год (с осени до следующего лета). По нашим многолетним на-
блюдениям, в Приморском крае имаго M. fuliginosa после зимней диапаузы встреча-
ются с начала мая (табл. 2). Первыми появляются самки (02-04.V.2017); с конца 1-й
декады мая в агроценозах наблюдаются массовые случаи спаривания, в этот период
клопы образуют скопления из 10 и большего числа имаго на излюбленных кормовых
растениях из сем. Rosaceae - малине и ежевике - в период бутонизации. Здесь собира-
ются особи обоих полов, копуляция прерывается для питания и вновь возобновляется,
продолжительность дополнительного питания перед откладкой яиц составляет от 2 ч.
до 3 сут.
Первые яйцекладки в стационарных садках получены нами в конце 1-й декады мая,
а в лесных стациях первые самки и их яйцекладки отмечены несколько позже, с сере-
дины мая, самцы - со 2-й декады июня. Копуляция и яйцекладка растянуты - их
можно наблюдать до конца 1-й декады августа.
Кладка происходит вразброс, яйца помещаются на субстрат беспорядочно. Среднее
число яиц, отложенных одной самкой в стационарных садках, составляет 28.4 (n = 25).
Ранее нами были приведены данные о локализации яиц M. fuliginosa на растениях в
естественных условиях и в стационарных садках (Маркова и др., 2017б). Наблюдения
2016-2020 гг. позволили привести новые данные о местах откладки яиц, в том числе
на поверхности почвы под растениями (табл. 3; рис. 2).
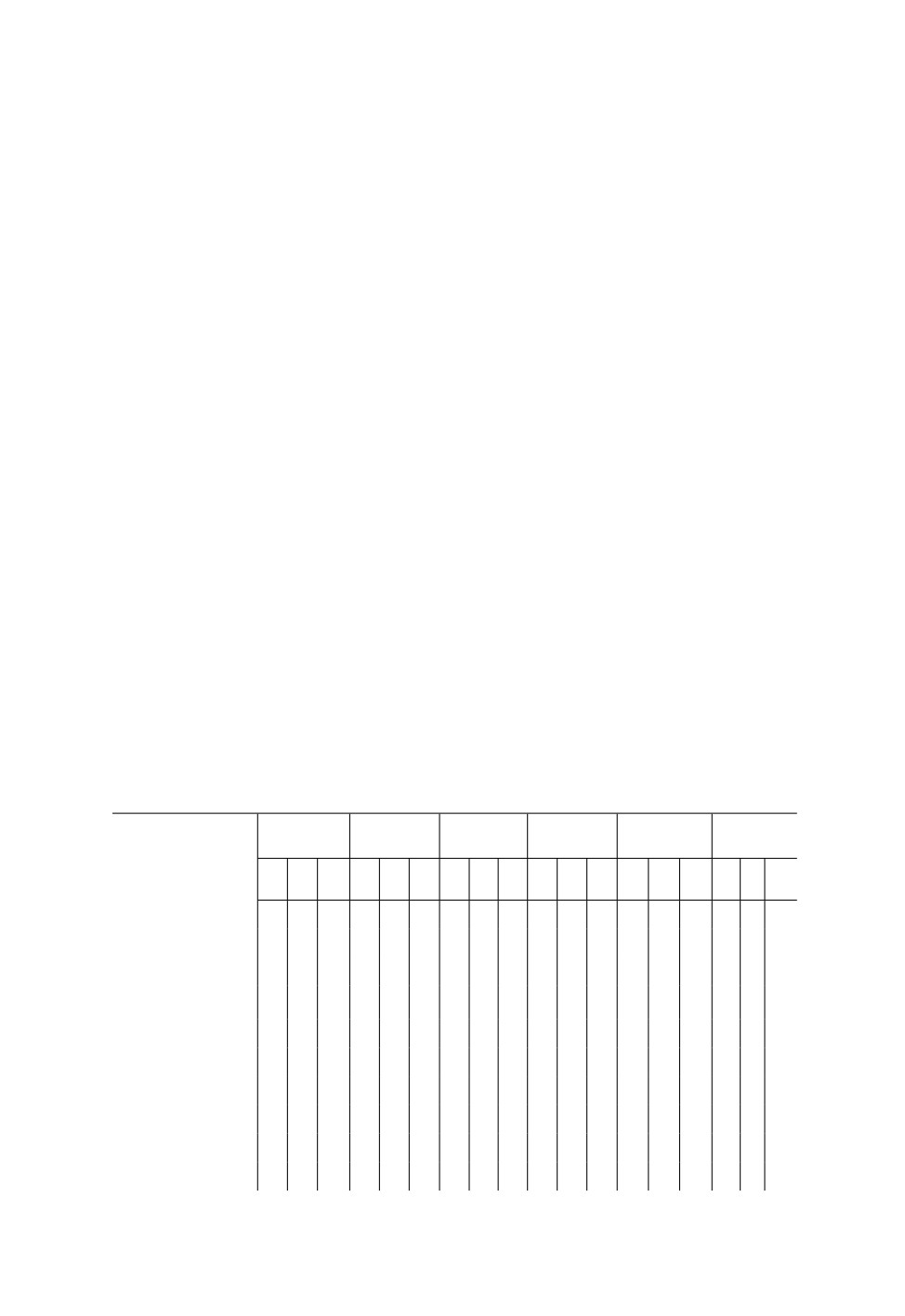
Таблица 2. Фенология Molipteryx fuliginosa (Uhler) (по декадам) в естественных условиях и
стационарных садках на территории Приморского края (2014-2020 гг.)
Стадия или
Май
Июнь
Июль
Август
Сентябрь
Октябрь
физиологический
период
1
2
3
1
2
3
1
2
3
1
2
3
1
2
3
1
2
3
Имаго
+
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
Копуляция
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
Яйцо
-
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
Личинка
I возраст
-
-
-
-
+
+
+
+
+
+
+
-
-
-
-
-
-
-
II возраст
-
-
-
-
-
+
+
+
+
+
+
+
+
-
-
-
-
-
III возраст
-
-
-
-
-
-
+
+
+
+
+
+
+
-
-
-
-
-
IV возраст
-
-
-
-
-
-
-
+
+
+
+
+
+
-
-
-
-
-
V возраст
-
-
-
-
-
-
-
-
+
+
+
+
+
+
+
-
-
-
Имаго
-
-
-
-
-
-
-
-
-
-
+
+
+
+
+
+
+
+
91
Таблица 3. Места откладки яиц Molipteryx fuliginosa (Uhler)
Биотоп
Локализация яиц
Число яиц
Приусадебный уча-
Lonicera edulis Turcz. ex Freyn
Верхняя сторона листа
2
сток в сельской
Trifolium repens L.
Нижняя сторона листа
5
местности
Стебель
2
Стационарный садок
Фрагменты коры на почве
Нижняя поверхность
2
Сухие листья и стебли злаков
Верхняя сторона
3
Поверхность почвы
2
Ambrosia artemisiifolia L.
Верхняя сторона листа
1
Нижняя сторона листа
1
Стебель
1
Solanum tuberosum L.
Верхняя сторона листа
8
Нижняя сторона листа
2
Стебель
1
Всего
30
Molipteryx fuliginosa - полифаг; из перечисленных выше растений жимолость съе-
добная Lonicera edulis (сем. Caprifoliaceae) и клевер ползучий Trifolium repens (сем.
Fabaceae) ранее в качестве кормовых для M. fuliginosa не были известны, но, очевидно,
используются самкой в качестве субстрата для размещения яиц. Кладка на стебле кар-
тофеля Solanum tuberosum (сем. Solanaceae) была обнаружена ранее в естественных
Рис. 2. Места откладки яиц Molipteryx fuliginosa (Uhler).
1 - фрагмент коры, 2 - стебель Ambrosia artemisiifolia.
92
Рис. 3. Личинки Molipteryx fuliginosa (Uhler) II и V возрастов.
условиях, а в садке яйца откладывались на листовые пластины. В естественных усло-
виях на амброзии Ambrosia artemisiifolia (сем. Asteraceae) личиночные стадии
M. fuliginosa до настоящего времени не обнаружены, однако в садке клопы размещали
яйца на этом растении (см. рис. 2).
В стационарных садках, расположенных в природных условиях, нами определена
длительность эмбрионального развития M. fuliginosa, которая колеблется от 12 до 36
(m = 23.6; n = 343) дней. Личинки M. fuliginosa до превращения в имаго проходят пять
возрастов (Маркова и др., 2017а) (рис. 3). Продолжительность развития каждого личи-
ночного возраста увеличивается в процессе онтогенеза и составляет соответственно
5-7 (m = 6.0) (I), 13-16 (m = 14.5) (II), 7-8 (m = 7.5) (III), 11-23 (m = 16.6) (IV) и 19-38
(m = 27.9) (V) дней (n = 68). Общая продолжительность развития от момента выхода
личинки из яйца до окрыления имаго - от 55 до 83 дней, от откладки яйца до окры-
ления имаго проходит от 80 до 117 дней.
Имаго перезимовавшего поколения отмирают в течение июля (самок со 2-й декады,
самцов - с конца месяца). В середине июля 2015 г. на почве в стационарном участке
с. Каймановка обнаружены погибшие самки, не закончившие яйцекладку. Отдельные
перезимовавшие особи M. fuliginosa доживают до середины августа.
По данным, полученным в стационарных садках, расположенных в природных ус-
ловиях, для полноценного развития клопа оптимальный период яйцекладки - с конца
1-й декады мая до конца 2-й декады июня. При этих сроках личинки I возраста появля-
ются с начала 2-й декады июня, II возраста - c конца 2-й - начала 3-й декады июня,
III возраста - c начала 1-й декады июля, IV - c конца 1-й - начала 2-й декады июля,
V возраста - с начала 3-й декады июля (см. табл. 2). Имаго нового поколения окрыля-
ются с середины августа до середины сентября, при этом с конца 1-й декады сентября
93
отмечено появление необычно мелких особей с ослабленной склеротизацией по-
кровов и с не полностью развитыми крыльями. Массовый вылет имаго нового поко-
ления в природе наблюдается с конца августа до начала сентября (по нашим данным,
30.VIII-02.IX.2016, 20-21.VIII.2017, 21.VIII-01.IX.2020). Активное преддиапаузное
питание и накопление клопами жирового тела для успешной зимовки продолжаются
до конца августа. Отдельные имаго нового поколения в хорошую погоду отмечались
на кормовых растениях до начала октября.
Для M. fuliginosa выявлена облигатная имагинальная диапауза (Маркова и др., 2017а,
2017б), которая формируется в конце сезона вегетации и сопровождается прекраще-
нием репродукции и проявлением миграционного синдрома - имаго покидают
прежние места обитания. Не успевшие окрылиться личинки продолжают питаться, но
с понижением ночных температур воздуха постепенно отмирают, не закончив раз-
витие. С конца августа наблюдается уменьшение численности клопов в агроценозах
и перемещение в другие (в том числе лесные) стации. Благодаря содержанию имаго в
стационарных садках с сохранением влияния на них всех атмосферных факторов
в осенне-зимний период выяснено, что M. fuliginosa зимуют близ поверхности и
в верхних слоях почвы под листовым опадом и сухими растительными остатками.
ЗАКЛЮЧЕНИЕ
Установлены особенности сезонного развития M. fuliginosa в климатических усло-
виях юга Дальнего Востока России. Для успешного завершения цикла развития
M. fuliginosa оптимальны следующие условия.
1. Яйцекладка в период с конца 1-й декады мая до конца 2-й декады июня общей
продолжительностью 40-43 дня. Среднее число отложенных самкой яиц составляет
28.4 (n = 25). Продолжительность эмбриональной стадии M. fuliginosa - от 12 до 36
(m = 23.6) дней.
2. Появление личинок I возраста наблюдается с начала 2-й декады июня, II возраста
- c конца 2-й - начала 3-й декады июня, III возраста - c начала 1-й декады июля, IV -
c конца 1-й - начала 2-й декады июля, V возраста - с начала 3-й декады июля. Продол-
жительность развития личинок M. fuliginosa с каждым возрастом увеличивается.
3. Общая продолжительность развития от яйцекладки до окрыления имаго состав-
ляет от 80 до 117 дней. Массовое окрыление имаго нового поколения наблюдается
с конца августа до начала сентября. Зимовка имаго происходит на поверхности почвы
под листовым опадом и сухими растительными остатками.
Продолжительность жизни имаго в природе после зимовки составляет в среднем
91 день, перезимовавшее поколение отмирает в течение июля.
БЛАГОДАРНОСТИ
Авторы выражают искреннюю благодарность Д. Л. Мусолину (Санкт-Петербург-
ский государственный лесотехнический университет им. С. М. Кирова),
Н. Н. Винокурову (Институт биологических проблем криолитозоны СО РАН, Якутск)
за ценные советы и замечания при подготовке рукописи, а также Л. А. Фединой (ФНЦ
Биоразнообразия ДВО РАН, Владивосток), А. К. Мрикот, А. В. Ховриной (ДВФУ,
Школа педагогики, Уссурийск), З. И. Лимачко (Средняя общеобразовательная школа
94
№ 7 с. Новомихайловка Чугуевского р-на) за помощь в проведении стационарных ис-
следований. Авторы глубоко благодарны также Н. М. Яворской («Заповедное Приа-
мурье», Хабаровск), А. Ю. Олейникову (Институт водных и экологических проблем
ДВО РАН, Хабаровск); Н. А. Коляде, А. В. Куприну, М. М. Омелько, М. Е. Сергееву,
Д. М. Черняк, Ю. А. Чистякову (ФНЦ Биоразнообразия ДВО РАН, Владивосток),
А. М. Николаевой (Окский государственный заповедник), К. А. Остапенко (ДВФУ,
Зоологический музей, Владивосток), Ю. Н. Глущенко, Н. В. Репш, Е. Н. Боловцову,
А. Д. Гребенюк, С. Е. Егоренчеву, Ю. С. Заварзиной, Е. А. Новой (ДВФУ, Школа педа-
гогики, Уссурийск), С. А. Макаревичу (личное подсобное хозяйство «Макаревич»,
плодовый питомник, с. Заречное), М. В. Аналеевой, С. В. Вериге, А. В. Ковалевой,
И. Д. Солодкому за предоставление сведений из Красноармейского, Тернейского, Хан-
кайского, Спасского, Яковлевского, Черниговского, Анучинского, Октябрьского, Чугу-
евского, Шкотовского, Партизанского, Хасанского районов; Дальнегорского, Арсень-
евского, Уссурийского, Владивостокского и Находкинского районов и городских
округов Приморского края.
СПИСОК ЛИТЕРАТУРЫ
Винокуров Н. Н., Канюкова Е. В., Остапенко К. А. 2016. Материалы по фауне цикадовых (Homoptera,
Cicadina) и полужесткокрылых (Heteroptera) насекомых Сихотэ-Алинского государственного природ-
ного биосферного заповедника. Амурский зоологический журнал 8 (4): 233-249.
Канюкова Е. В. 2012. Клоп-краевик Molipteryx fuliginosa (Heteroptera: Coreidae) - новый вредитель малины
на юге Дальнего Востока России. Амурский зоологический журнал 4 (4): 331-332.
Канюкова Е. В., Остапенко К. А. 2013. Новые и малоизвестные полужесткокрылые (Heteroptera) из Примор-
ского края. Амурский зоологический журнал 5 (4): 405-407.
Маркова Т. О., Канюкова Е. В., Маслов М. В. 2019. Полужесткокрылые насекомые (Heteroptera) с амброзии
(Ambrosia artemisiifolia L.) на юге Приморского края. Евразиатский энтомологический журнал 18 (1):
16-20.
Маркова Т. О., Канюкова Е. В., Маслов М. В. 2021. Динамика расселения клопа-краевика Molipteryx fuliginosa
(Uhler) (Heteroptera, Coreidae) на Дальнем Востоке России. Российский журнал биологических инва-
зий 14 (2): 70-80.
Маркова Т. О., Канюкова Е. В., Маслов М. В., Репш Н. В. 2017a. Преимагинальные фазы Molipteryx fuliginosa
(Uhler) (Heteroptera, Coreidae) с юга Дальнего Востока России. Энтомологическое обозрение 96 (3):
418-428.
[Markova T. O., Kanyukova E. V., Maslov M. V., Repsh N. V. 2017a. Immature stages of Molipteryx fuliginosa
(Uhler) (Heteroptera, Coreidae) in the South of the Russian Far East. Entomological Review 97 (6): 723-729.
Маркова Т. О., Маслов М. В. 2020. Амброзия полыннолистная (Ambrosia artemisiifolia L.) (Asteraceae) в пи-
тании клопов-краевиков (Heteroptera: Coreidae) на юге Приморского края. Чтения памяти Алексея
Ивановича Куренцова 31: 165-173.
Маркова Т. О., Маслов М. В., Канюкова Е. В., Репш Н. В. 2017б. Репродуктивное поведение клопов Molipteryx
fuliginosa (Uhler) (Heteroptera, Coreidae) на юге Дальнего Востока России. Энтомологическое обозре-
ние 96 (4): 725-736.
[Markova T. O., Maslov M. V., Kanyukova E. V., Repsh N. V. 2017b. Reproductive behaviour of the bug Molipteryx
fuliginosa Uhler (Heteroptera, Coreidae) in the South of the Russian Far East. Entomological Review
97 (9): 1227-1233.
95
Маркова Т. О., Маслов М. В., Репш Н. В. 2016. К экологии клопа-краевика Molipteryx fuliginosa (Heteroptera,
Coreidae) в Уссурийском районе Приморского края. Чтения памяти Алексея Ивановича Куренцова
27: 80-84.
Маркова Т. О., Маслов М. В., Репш Н. В. 2018. Модификации садков для исследования насекомых. Еврази-
атский энтомологический журнал 17 (5): 345-348.
Саулич А. Х., Мусолин Д. Л. 2007. Времена года: разнообразие сезонных адаптаций и экологических меха-
низмов контроля сезонного развития полужесткокрылых (Heteroptera) в умеренном климате. В кн.:
А. А. Стекольников (ред.). Стратегии адаптаций наземных членистоногих к неблагоприятным услови-
ям среды. Сборник статей, посвященный памяти профессора Виктора Петровича Тыщенко. CПБ.,
с. 25-106. (Труды Биологического НИИ СПбГУ, вып. 53).
Aistova E. V., Bezborodov V. G., Markova T. O., Maslov M. V., Fedina L. A. 2019. The formation of the consortia
relations of Molipteryx fuliginosa (Uhler, 1860) (Hemiptera, Coreidae) with Ambrosia artemisiifolia in the
Primorskii Krai of Russia. Ecologica Montenegrina 21: 90-99.
Kanyukova E. V., Vinokurov N. N. 2009. New data to the fauna of superfamilies Lygaeoidea, Pyrrhocoroidea and
Coreioidea (Heteroptera) of the Asian part of Russia. В кн.: А. И. Таскаев [и др.] (отв. ред.). Проблемы
изучения и охраны животного мира на севере: Материалы докладов Всероссийской научной конфе-
ренции с международным участием (Сыктывкар, Республика Коми, Россия, 16-20 ноября 2009 г.).
Сыктывкар: Коми научный центр УрО РАН, с. 57-59.
Kerzhner I. M., Kanyukova E. V. 1998. First record of Molipteryx fuliginosa Uhler from Russia (Heteroptera:
Coreidae). Zoosystematica Rossica 7 (1): 84.
Markova T. O., Maslov M. V., Repsh N. V., Ogorodnikov E. G. 2016. New data on distribution of Molipteryx
fuliginosa (Heteroptera: Coreidae) in Russia. Far Eastern Entomologist 316: 26-28.
Park S. O. 1996. Development of the leaf-footed bug, Molipteryx fuliginosa (Heteroptera: Coreidae). Korean Journal
of Ecology 19 (6): 575-582.
PHENOLOGY OF MOLIPTERYX FULIGINOSA (UHLER) (HETEROPTERA,
COREIDAE) IN THE SOUTH OF THE RUSSIAN FAR EAST
T. O. Markova, E. V. Kanyukova, M. V. Маslov
Key words: bugs, Molipteryx fuliginosa, seasonal development, phenology, egg, larvae,
wintering, Primorskiy Territory, Heteroptera, Coreidae.
SUMMARY
Seasonal development of Molipteryx fuliginosa (Uhler) in Primorskii Territory, including the time of
oviposition and development of all stages before the emergence of adults, was studied during a six-year
monitoring in natural and seminatural conditions. Molipteryx fuliginosa has a univoltine seasonal type.
Eggs are laid randomly on various substrates, including the ground. The duration of the embryonic
stage ranges from 12 to 36 days, depending on the air temperature. The development of larvae after
hatching until the emergence of adults lasts from 55 to 83 days. The duration of immature development
from the moment of oviposition to the emergence of adult is 80 to 117 days. The optimal conditions and
timing of the seasonal development of the bug in nature have been identified.
96