ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 101, 2, 2022
УДК 632.76:574.34:595.768.1
НОВАЯ ВСПЫШКА МАССОВОГО РАЗМНОЖЕНИЯ КОРОЕДА-
ТИПОГРАФА IPS TYPOGRAPHUS (L.) (COLEOPTERA,
CURCULIONIDAE) В ЛЕНИНГРАДСКОЙ ОБЛАСТИ
© 2022 г. А. В. Селиховкин,1* Н. А. Мамаев,1** М. Б. Мартирова,1***
С. А. Меркурьев,2,3**** Б. Г. Поповичев1*****
1 Санкт-Петербургский государственный лесотехнический университет им. С. М. Кирова
Институтский пер., 5, С.-Петербург, 194021 Россия
⁎e-mail: a.selikhovkin@mail.ru (автор, ответственный за переписку), ⁎⁎e-mail: mamaevld@bk.ru,
⁎⁎⁎e-mail: masha2340350@yandex.ru, ⁎⁎⁎⁎⁎e-mail: b.g.popovichev@yandex.ru
2 Санкт-Петербургский филиал Института земного магнетизма, ионосферы
и распространения радиоволн
Университетская наб., 5, С.-Петербург, 199034 Россия
3 Санкт-Петербургский государственный университет
Университетская наб., 7/9, С.-Петербург, 199034 Россия
⁎⁎⁎⁎e-mail: sam_hg@hotmail.com
Поступила в редакцию 01.02.2022 г.
После доработки 4.03.2022 г.
Принята к публикации 4.03.2022 г.
В Ленинградской обл. в 2021 г. началась новая и потенциально сильная вспышка массово-
го размножения короеда-типографа Ips typographus. Перманентное присутствие значительного
количества короеда-типографа в ельниках этого региона создало предпосылки для быстрого
увеличения численности этого вида. Спровоцировало развитие вспышки размножения резкое
повышение температуры в июне-июле 2021 г., благодаря которому успешно развились два се-
стринских и два основных поколения. Массовое размножение двух генераций короеда-типогра-
фа в ельниках Карельского перешейка отмечено впервые. По бивольтинному типу развивалась
часть популяции: примерно 56 % особей дали второе поколение, успешно завершившее разви-
тие, остальная часть молодых жуков ушла в диапаузу. Эта особенность развития обеспечивает
устойчивость популяций короеда-типографа к неблагоприятным погодным условиям. Часть вто-
рого поколения, начавшая развиваться раньше, имела высокую энергию размножения (отноше-
ние числа особей молодого поколения к числу особей родительского поколения) (4.7), близкую
к энергии размножения первого поколения (6.0). Следующую группу второго поколения, по-ви-
димому, в основном представляло потомство сестринского поколения. Показатели развития в
этой группе были существенно ниже (энергия размножения - 1.3) из-за того, что значительная
часть особей этого поколения не успела завершить развитие до начала осенних холодов и под-
готовиться к диапаузе.
Короедный запас (число особей на гектар), обеспечивающий быстрое нарастание численно-
сти короеда-типографа, в условиях Карельского перешейка можно оценить в 140-150 тыс. осо-
бей/га, или 10-12 заселенных типографом елей на гектар. В рассматриваемом случае короед-
ный запас был выше - 187 тыс. особей/га, что обеспечило появление очень большого потомства
(около 3 млн особей/га), способного обеспечить стремительное нарастание вспышки.
239
Быстрый и устойчивый тренд роста температуры, наличие большого объема кормовой базы
для короеда-типографа в Ленинградской обл. и в юго-западной части Карелии и сохранение ко-
роедного запаса позволяют предположить, что эта вспышка массового размножения будет стре-
мительно развиваться, а в ближайшем будущем можно ожидать ее повторения.
Ключевые слова: короед-типограф, Ips typographus, вспышка массового размножения, числен-
ность, генерация.
DOI: 10.31857/S0367144522020034, EDN: HJTBWL
Вспышки массового размножения короеда-типографа Ips typographus (Linnaeus,
1758) на территории Ленинградской обл. и в окрестностях С.-Петербурга отмечались
начиная с XIX в. (Семашко, 1864; Шевырёв, 1895), а затем многократно в XX
и XXI вв. (Катаев, 1948, 1952, 1956, 1983; Катаев и др., 1984; Осетров, Селиховкин,
1998; Селиховкин и др., 2016, 2017, 2018). В текущем столетии антропогенные
факторы в сочетании с климатическими изменениями привели к резкому обострению
ситуации. Различные виды рубок, в особенности санитарные рубки, строительство
скоростных магистралей, понижение уровня грунтовых вод, повышение средней тем-
пературы воздуха и другие факторы создали благоприятные условия для размножения
стволовых вредителей хвойных, в особенности короеда-типографа (Селиховкин, 2017;
Селиховкин и др., 2017; Selikhovkin et al., 2021). Последняя вспышка массового раз-
множения этого вредителя завершилась в 2016-2017 гг. (Селиховкин, 2017; Сели-
ховкин и др., 2017, 2018). При исследованиях видового состава вредителей и пато-
генов и их роли в изменении состояния хвойных древостоев Ленинградской обл.,
проводившихся нами в 2021 г., были обнаружены масштабные очаги короеда-типо-
графа в Ленинградской обл. на Карельском перешейке и в Гатчинском лесничестве,
а также в Республике Карелия в Сортавальском лесничестве. При этом короед-типо-
граф дал два поколения, что совершенно не характерно для его популяций в Ленин-
градской обл. и в особенности на Карельском перешейке (Селиховкин и др., 2017,
2018). В связи с этим была поставлена задача изучения особенностей размножения
короеда-типографа в обнаруженных очагах.
МАТЕРИАЛ И МЕТОДИКА
Основные исследования проводились в Ленинградской обл., в Рощинском и Северо-западном
лесничествах (Карельский перешеек) и в Учебно-опытном лесничестве Тосненского р-на (Ли-
синский лесхоз СПбГЛТУ), расположенном в 50 км к югу от С.-Петербурга.
По программе исследований в рамках проекта Российского научного фонда № 21-16-
00065 проводились маршрутные обследования, целью которых был подбор насаждений для за-
кладки пробных площадей по методике ICP Forests (Методика, 1995; Alekseev et al., 2018) и
оценки состояния окружающих древостоев.
В ходе маршрутных обследований на Карельском перешейке было выявлено несколько очагов
размножения короеда-типографа, шесть из которых занимали значительную площадь (табл. 1).
Предварительное обследование очагов было проведено в период с 16 по 25 августа 2021 г. В
процессе детального обследования очагов с 20 по 26 ноября 2021 г. были определены границы
очагов и выполнена их координатная привязка, установлена доля деревьев, заселенных короеда-
ми первого и второго поколений. Деревья, заселенные вторым поколением короедов, имели зе-
леную или бледно-зеленую крону и мало отличались от незаселенных деревьев в августе (рис. 1).
В ноябре хвоя приобрела желтую окраску, и эти отличия были хорошо видны. На деревьях, от-
работанных первым поколением, к этому времени хвои почти не осталось.
240
Таблица 1. Расположение и площадь очагов размножения короеда-типографа в Рощинском
лесничестве Ленинградской обл., сформировавшихся в 2021 г.
Координаты центра очага
№
Площадь, га
Широта, N
Долгота, E
1
60°17′1.36″
29°56′22.8″
9.5
2
60°17′47.6″
29°54′15.1″
1.7
3
60°17′44.1″
29°53′45.4″
0.5
4
60°26′1.64″
28°53′34.4″
2.3
5
60°26′55.3″
28°52′32.7″
1.0
6
60°20′29.6″
29°56′33.7″
2.0
Всего
-
-
17.0
Анализ популяционных характеристик проводился на основе классической методики учета
стволовых вредителей (Мозолевская и др., 1984). На каждом модельном дереве анализировалась
только одна палетка на высоте груди (1.3 м), так как валка деревьев не представлялась возмож-
ной из-за сложностей оформления разрешений на нее. Протяженность палетки составляла 50 см
по всей окружности ствола. На палетке подсчитывалось количество лётных отверстий, маточ-
ных ходов, брачных камер, живых жуков, куколок и личинок короеда-типографа. Кроме того,
фиксировались ходы и личинки других видов стволовых вредителей, а также места ухода в дре-
весину личинок жуков-усачей. Деревья отбирались случайным образом с исключением тех,
у которых нижняя часть ствола была затенена подростом. В общей сложности было проанализи-
ровано 21 дерево.
Рассчитывались следующие популяционные характеристики: плотность поселения (ПП) -
число жуков родительского поколения, приходящееся на единицу площади палетки (1 дм2); про-
дукция - число жуков молодого поколения, приходящееся на единицу площади палетки; энергия
размножения (ЭР) - отношение числа особей молодого поколения к числу особей родительского
поколения; короедный запас (КЗ) - число жуков родительского поколения на 1 гектар; коро-
едный прирост (КП) - число жуков молодого поколения на 1 гектар.
При анализе полученных данных по популяционным характеристикам использовалась про-
верка гипотезы о принадлежности выборок к одной генеральной совокупности, проведенная по
t-критерию Стьюдента для малых выборок.
Для выяснения характера изменения температуры в С.-Петербурге за последние 40 лет (1980-
2021 гг.) был применен регрессионный анализ средних температур. Отдельно исследовались
среднегодовая и средняя температуры за вегетационный сезон (май-сентябрь), а также сумма
эффективных температур (СЭТ) выше 10 °C на территории С.-Петербурга. Принята простая де-
терминированная линейная модель динамики температуры. При анализе изменения температу-
ры методом наименьших квадратов были рассчитаны параметры уравнения регрессии, а также
коэффициенты корреляции и доверительные интервалы. Использовались сведения Мирового
центра данных Всероссийского научно-исследовательского института гидрометеорологической
тельные, выверенные и корректные ряды значений метеорологических показателей. Эти данные
ru, поскольку база Мирового центра данных заканчивается 2020 годом.
Средние температуры воздуха за месяц или вегетационный сезон вычислялись из среднесу-
точной температуры путем осреднения дневных данных. После этого для исследуемого времен-
ного интервала строились графики их многолетней динамики и рассчитывались коэффициенты
241
Рис. 1. Ели, заселенные первым, сестринским и вторым поколениями короеда-типографа,
Рощинское лесничество, август 2021 г. (фото А. В. Селиховкина, 2017 г.)
уравнений линейной регрессионной модели, а также ее статистические параметры, включая ко-
эффициенты корреляции между фактическими данными и их линейными трендами, коэффици-
ент детерминации и среднеквадратическое отклонение (табл. 4). При расчете суммы эффективных
температур величина порога принималась равной 10 ºС.
РЕЗУЛЬТАТЫ
Обследования, проведенные в Учебно-опытном лесничестве (Лисинский учебно-
опытный лесхоз), не выявили очагов или куртин елей, заселенных в 2021 г. короедом-
242
типографом. Встречались только группы не более чем из трех деревьев или отдельные
ели, заселенные короедом-типографом. Следует отметить, что на территории лесниче-
ства более 20 лет практически не проводились никакие лесохозяйственные и, в том
числе, санитарно-защитные мероприятия в силу ряда нормативно-правовых обстоя-
тельств. Однако в Гатчинском лесничестве, к которому с востока примыкает Учеб-
но-опытное лесничество, нами были обнаружены 3 очага короеда-типографа пло-
щадью от 150 до 300 га. Эти очаги сформировались в 2021 г., располагались в
непосредственной близости от шоссейных дорог и имели следующие координаты:
1-й - 59°27′12.7″ с. ш., 030°06′48.1″ в. д.; 2-й - 59°20′15.3″ с. ш., 030°07′35.3″ в. д.;
3-й - 59°19′36.6″ с. ш., 030°07′31.5″ в. д.
В Рощинском лесничестве были обнаружены 6 крупных очагов, сформировавшихся
в 2021 г., которые послужили основными объектами исследований (см. табл. 1). Все
очаги находились в спелых и перестойных ельниках, чистых или с небольшой при-
месью сосны и относительно недалеко друг от друга. Преобладающий тип леса -
ельник-черничник. Насаждения имели нормальную полноту, только в одном очаге
(№ 6) в 2020 г. была проведена добровольно-выборочная рубка, в результате которой
полнота насаждения снизилась до 0.5.
В ходе обследования, проведенного в августе, было установлено, что первое поко-
ление короеда-типографа полностью завершило развитие, а второе заселило значи-
тельную часть деревьев. Деревья, заселенные вторым поколением короедов, имели
зеленую или бледно-зеленую крону и мало отличались от незаселенных деревьев
(см. рис. 1). Часть первого поколения закончила развитие быстро, в конце июня, и уже
в августе на этих деревьях отмечались участки с облетевшей корой. Личинки усачей
первого и второго возрастов на этих деревьях встречались, однако места ухода усачей
в древесину, как и на всех других деревьях, заселенных в 2021 г., отсутствовали, т. е.
деревья были заселены усачами в 2021 г. Позднее, при обследовании в ноябре, отмеча-
лись места ухода усачей Monochamus spp. и Tetropium spp. в древесину и куколочные
колыбельки Rhagium spp. под корой.
В августе на некоторых деревьях активно развивались одновременно сестринское
поколение и второе поколение короеда-типографа. Заселившиеся жуки хорошо разли-
чались: особи, откладывавшие яйца повторно, были темноокрашенными, почти чер-
ными, а заселившиеся жуки молодого поколения - светло-коричневыми. Заселение
деревьев шло тремя волнами.
1. Первое (основное) родительское поколение (РП1) заселяло деревья в начале мая.
2. Повторно первое родительское поколение (РП1с) заселяло деревья в конце июня
(его потомство составит сестринское поколение), и одновременно заселялись жуки
нового поколения (РП2) - потомства РП1. РП2, по-видимому, было сформировано жу-
ками основного родительского поколения и уже почти полностью закончило развитие
во второй половине августа. Большинство отродившихся жуков вылетело.
3. В июле и позднее заселялись только жуки нового поколения (РП2), которое было
представлено потомками преимущественно первого сестринского поколения. Значи-
тельная часть особей этого поколения не успела завершить развитие, и в ноябре под
корой встречались и жуки, и куколки, и личинки.
243
Соответственно, были выделены три группы заселенных деревьев в зависимости от
того, жуки какой генерации заселяли их.
1. Деревья, на которых развивалось первое поколение, т. е. заселенные родитель-
ским поколением РП1 (уже в начале августа на этих деревьях отмечались участки с
облетевшей корой в местах развития и пожелтевшая хвоя).
2. Деревья, на которых развивались первое и второе сестринские поколения, а также
второе поколение - потомки основного родительского поколения, т. е. деревья, засе-
ленные РП1с и РП2.
3. Деревья, на которых развивалось второе поколение - потомство сестринского по-
коления.
Во второй группе, по-видимому, были жуки, откладывающие яйца как во второй,
так и в третий раз и, соответственно, формирующие первое и второе сестринские
поколения. Долю особей, приступающих к повторной откладке (первое сестринское
поколение), нам не удалось установить. К откладке яиц второго сестринского поко-
ления приступало 12 % самок. В литературе приводятся весьма разные данные о соот-
ношении жуков, формирующих первое и второе сестринское поколения, но в целом
оно составляет примерно 1 : 3 (Маслов, 2010). Из этого соотношения мы исходили в
дальнейших расчетах.
При проведении повторного обследования в ноябре деревья, погибшие в результате
развития первого поколения, хорошо отличались: имели желтую, сильно изреженную
хвою, а на многих кора частично отвалилась. Деревья, на которых развивалось второе
поколение, имели довольно интенсивное охвоение, хвоя была желто- или бледно-
зеленой. На всех деревьях, заселенных в 2021 г. короедом-типографом, отсутствовали
летные отверстия усачей. Нередко отмечались места ухода на зимовку в древесину
личинок усачей Monochamus spp. и Tetropium spp., а также личинки усачей рода
Rhagium F. в куколочных колыбельках под корой. Паразитоиды и хищники отмечались
единично, в среднем менее одного случая на палетку.
Средний диаметр стволов заселенных деревьев и плотность поселений первого и
второго поколений существенно не различались (табл. 2). Нет значимых различий
между характеристиками развития короедов 1-й и 2-й групп, развивающихся соответ-
Таблица 2. Характеристики развития короеда-типографа в очагах размножения в Рощинском
лесничестве в 2021 г.
Диаметр ствола
Плотность
Продукция,
Энергия
Заселяющееся поколение
дерева на высоте
поселения,
особей/дм2
размножения
груди, см
особей/дм2
РП1 - первое
36.32 ± 7.95
1.67 ± 0.92
6.63 ± 3.40
6.04 ± 2.70
РП1с + РП2 - первое,
откладывающее яйца
31.00 ± 5.34
1.73 ± 0.25
8.15 ± 1.60
4.73 ± 0.90
повторно, и второе
РП2 - только второе
30.60 ± 8.8
2.03 ± 1.00
*2.57 ± 1.29
*1.31 ± 0.35
Примечание: * - различия с остальными значениями в данной колонке значимы при p < 0.05.
244
ственно весной и в первой половине лета; весьма высока энергия размножения, обес-
печивающая многочисленность молодого поколения, и благодаря этому очень велика
продукция. Однако второе поколение, начавшее заселять деревья позже (группа 3),
размножается гораздо хуже: энергия размножения снижается примерно в 4 раза при
некотором увеличении плотности поселения и закономерно низкой продукции.
Для расчета числа особей родительского поколения и молодого поколения жуков на
1 га (короедный запас и короедный прирост соответственно) было принято, что
средняя протяженность района поселения составляет 20 м, число деревьев основной
породы (ель) - 600 на гектар. Доля заселенных деревьев составила 80 %. Распреде-
ление деревьев по группам заселения в зависимости от генерации было довольно рав-
номерным: 1-я группа (РП1) - 23.1; 2-я группа (РП1с + РП2) - 25.7; 3 группа (РП2) -
51.2 %. На основании размера доли жуков, формирующих сестринское поколение
(РП1с), которая составляет 12 %, и показателей развития в разных группах заселения
(см. табл. 2) были рассчитаны короедный запас и короедный прирост для первого ро-
дительского поколения РП1 и РП1с и второго родительского поколения (РП2) (табл. 3).
Из 1711 тыс. жуков первого и сестринского поколений (РП1 + РПс) больше поло-
вины (949 тыс.) продолжили развитие и сформировали многочисленное второе поко-
ление. Остальные 762 тыс. особей и молодые жуки второго поколения (2131 тыс.)
ушли на зимовку. Таким образом, сформирован огромный короедный запас 2022 г.,
который с учетом возможной гибели жуков во время подготовки к диапаузе составит
примерно 2.5 млн особей.
ОБСУЖДЕНИЕ
Ухудшение состояние ельников Карельского перешейка в последние три десяти-
летия обеспечило наличие доступной кормовой базы для стволовых вредителей, в
особенности для короеда-типографа. После окончания вспышки размножения
I. typographus в 2010-2015 гг. небольшие группы или куртины заселенных им елей
(см. рис. 2) можно было легко обнаружить на Карельском перешейке в Рощинском,
Северо-западном и Приозерском лесничествах (Селиховкин и др., 1917, 1918; Сели-
ховкин, 2022). По утверждению А. Д. Маслова (2010), критическая численность коро-
еда-типографа составляет 90-110 тыс. особей на гектар, т. е. семи или восьми крупных
елей, которые заселил короедом-типограф и дал потомство, может оказаться вполне
достаточно для достижения численности, способной перерасти во вспышку массового
размножения. Вероятно, именно за счет таких небольших куртин и возникли очаги
Таблица 3. Плотность популяции короеда-типографа в очагах размножения в Рощинском лес-
ничестве в 2021 г., тыс. особей/га
Средний диаметр
Короедный
Короедный
Ушло
ствола дерева на
запас
прирост
на зимовку
высоте груди, см
Первое поколение
36.30
315
1711
762
(РП1 + РПс)
Второе поколение (РП2)
30.60
949
2131
2131
Итого
-
-
3842
2893
245
размножения 2021 г. Если из короедного запаса РП1 (см. табл. 3) исключить жуков
сестринского поколения, то число особей, заселивших деревья весной 2021 г., со-
ставит 187 тыс./га, т. е. будет выше критического значения, достаточного для возникно-
вения вспышки массового размножения вредителей при благоприятных условиях
(Маслов, 2010). Данные А. Д. Маслова получены в Центральной России, где для типо-
графа характерно бивольтинное развитие. В нашем случае также наблюдалось мас-
совое развитие второго поколения, не характерное для севера Ленинградской обл., од-
нако около половины родительского поколения развивалась только в одном поколении.
Соответственно, критическое значение численности типографа для севера Ленинград-
ской обл. и южной Карелии должно быть несколько выше, чем по оценке А. Д. Мас-
лова, предположительно 140-150 тыс. особей или 10-12 заселенных типографом елей
на гектар.
Следует отметить, что часть не вылетевших жуков второго поколения, не завер-
шивших развитие и оставшихся под корой на зиму, может выжить, дополнив коро-
едный запас следующего года (Štefková et al., 2017). В итоге на зимовку ушло более
2.5 млн жуков на один гектар, т. е. очень большое количество, которое с высокой веро-
ятностью приведет к гибели ельников на площади, примерно в 5-7 раз превышающей
площадь очагов 2021 г.
Хорошо известно, что каждое основное поколение короеда-типографа может давать
развитие двум сестринским поколениям (Jurc et al., 2006; Маслов, 2010; Öhrn, 2012).
В северотаежных лесах, в том числе в Норвегии, как правило, отмечаются одно
основное поколение типографа и одно или два сестринских (Маслов, 2010; Öhrn, 2012;
Schebeck et al., 2021). Однако исследования фенотипов популяции короеда-типографа
в Норвегии показали, что для нее характерно наличие как факультативной, так и обли-
гатной диапаузы. Это позволяет части популяции при благоприятных условиях в
июне-августе продолжить развитие, а второй части, имеющей облигатную диапаузу,
избежать рисков при резком ухудшении погодных условий и сохранить часть особей
(Schebeck et al., 2021). В нашем случае часть популяции (44 %) развивалась в одном
поколении, и весьма вероятно, что она, так же как и норвежская популяция, могла
иметь облигатную диапаузу. Эта особенность развития обеспечивает устойчивость по-
пуляций короеда-типографа к неблагоприятным погодным условиям. Важно отметить,
что часть второго поколения, начавшая развиваться раньше, имела высокую энергию
размножения, близкую к энергии размножения первого поколения. Следующую
группу второго поколения, по-видимому, в основном представляло потомство сес-
тринского поколения. Показатели развития в этой группе были существенно хуже, и
значительная часть особей не успела завершить развитие.
По мере потепления климата бивольтинная часть популяции начинает преобладать.
В Швеции по мере повышения эффективной температуры в последние 80 лет проис-
ходит отчетливо выраженный переход к бивольтинному развитию короеда-типографа,
начиная с южной части страны (Jönsson et al., 2007, 2009). В Норвегии с потеплением
климата частота бивольтинных моделей развития увеличивается медленнее, также на-
чиная с ельников, расположенных в южной части страны и прилегающих к морским
побережьям. Переход к выраженной бивольтинности резко увеличивает возможный
ущерб от вспышек массового размножения I. typographus (Lange et al., 2009). Появ-
ление двух генераций у короеда-типографа в Ленинградской обл., даже частичное, по-
зволило быстро реализовать накопленный потенциал как вспышку массового размно-
жения и привести к гибели жизнеспособных ельников на больших площадях.
246
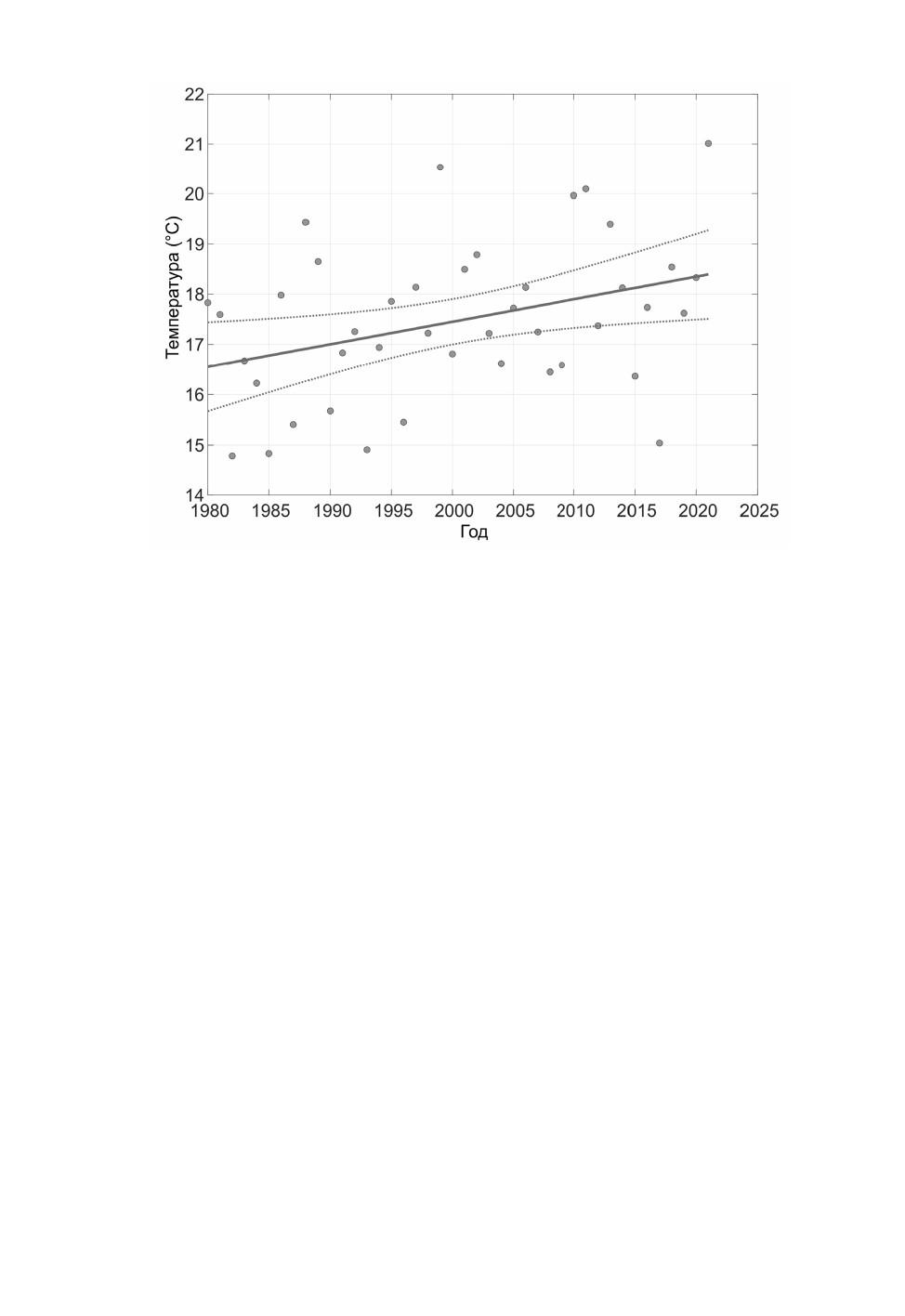
Рис. 3. Динамика средних температур за июнь-июль в С.-Петербурге с 1980 по 2021 г.
Точками обозначены расчетные значения; жирная линия - линия регрессии, полученная на основе
регрессионной модели (см. табл. 4); пунктирные линии - доверительный интервал при P ≤ 0.05.
Развитие короеда-типографа успешно протекает при температуре около 10 °C; оно
не останавливается и при более низких температурах (Wermelinger, 2004; Маслов,
2010; Štefková et al., 2017), однако откладка яиц происходит при среднесуточной тем-
пературе немного выше 10 °C (Wermelinger, 2004; Маслов, 2010).
При анализе изменения средней температуры воздуха - среднегодовой, средней за
вегетационный период и суммы эффективных температур (рис. 3-5, табл. 4) - уста-
новлены положительные достоверные линейные тренды изменения всех исследуемых
температур, при этом коэффициенты корреляции лежат в диапазоне 0.36-0.56.
В июне-июле 2021 г. в Ленинградской обл. произошел резкий скачок температуры
(рис. 3). Лето 2021 г. было весьма жарким - при летней норме 18.4 °C средняя темпе-
ратура за июнь-июль была 21 °C, что превышает предыдущие максимумы температур,
наблюдавшиеся в 1999, 2010 и 2011 гг. (20.5, 19.9 и 20.1 °C), и составляет абсолютный
максимум с 1980 г.
Резкое увеличение температуры обеспечило возможность быстрого развития первой
генерации, сестринского поколения и появления второго поколения, которое успешно
закончило развитие уже в августе. Потомство жуков, приступивших к откладке яиц
позднее (преимущественно сестринские поколения), развивалось не так успешно.
Температура в августе и сентябре была относительно низкой; многие особи к началу
холодов остались под корой на стадии куколки или имаго. Однако решающее зна-
чение, по-видимому, имела высокая температура в первой половине лета. Именно этот
жаркий период обеспечил стремительное нарастание численности. Интересно отме-
247
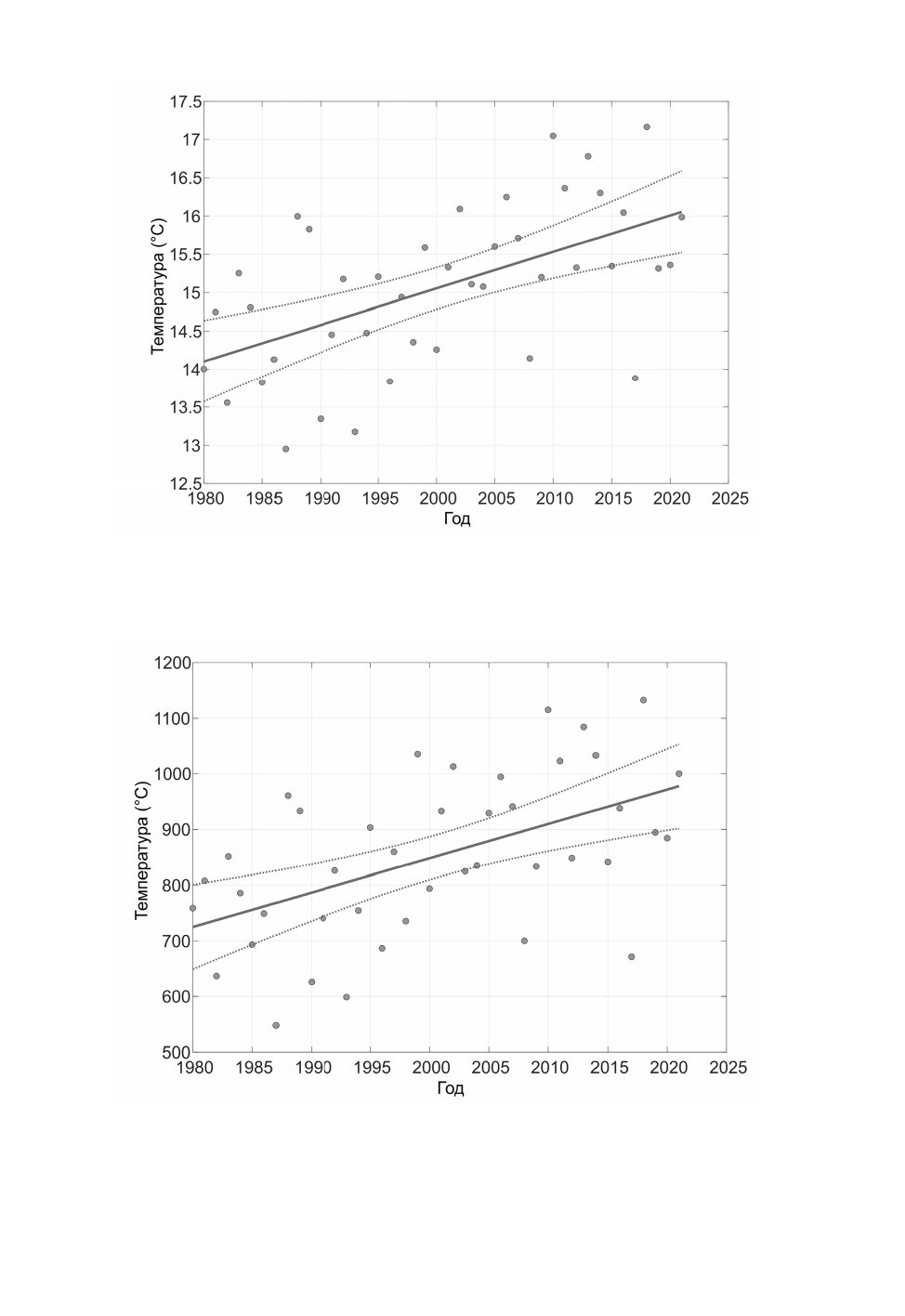
Рис. 4. Динамика средних температур за вегетационный сезон (май-сентябрь) 2021 г.;
Ленинградская обл.
Обозначения как на рис. 3.
Рис. 5. Динамика эффективных температур за вегетационный период (май-сентябрь) 2021 г.;
Ленинградская обл. Сумма эффективных температур рассчитывалась от порога 10 ºС.
Обозначения как на рис. 3, 4.
248
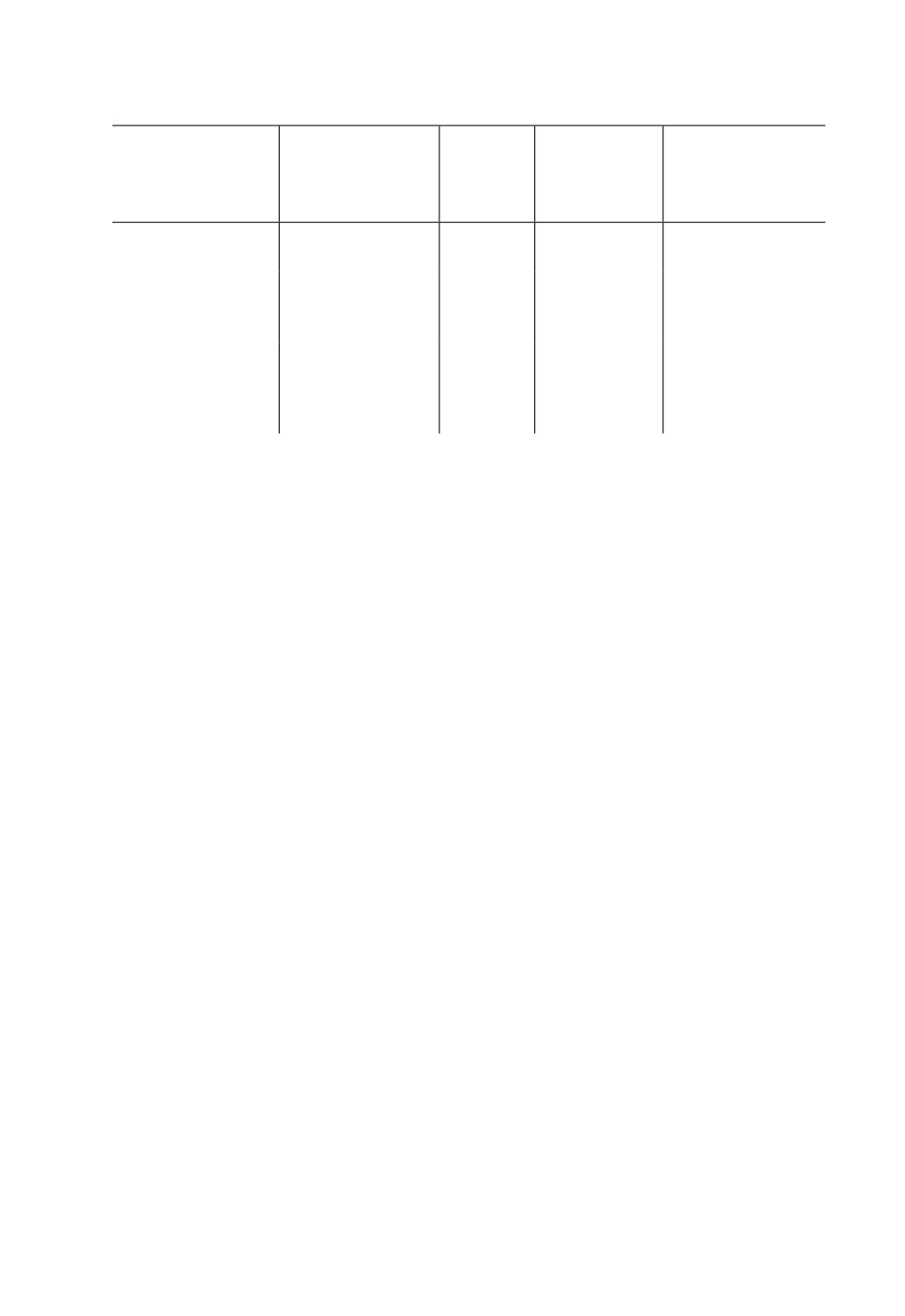
Таблица 4. Параметры модели линейной регрессии изменения средних температур (T) в Ленин-
градской обл.
Уравнение линейной
Коэф-
регрессии
Коэффициент
Среднеквадра-
Температуры
фициент
(T - температура,
детерминации
тическое отклонение
корреляции
Y - год)
Средние за июнь-
T = 0.045 × Y + 16.5
0.36
0.13
1.45
июль
Средние за вегета-
T = 0.048 × Y + 14.1
0.56
0.32
0.87
ционный сезон
(май-сентябрь)
Эффективные
T = 6.16 × Y + 719
0.53
0.28
124
за вегетационный
сезон (май-сен-
тябрь)
тить, что средняя и эффективная температуры за вегетационный сезон растут быстрее
(градиент оцененной регрессии составляет 0.32 и 0.38 соответственно) и существенно
более постоянны (коэффициенты корреляции больше 0.5), чем средняя температура
двух летних месяцев (см. табл. 4). В 2010 г., когда температурные показатели были
близки к показателям 2021 г. (см. рис. 3-5), началась предыдущая вспышка размно-
жения типографа (Селиховкин и др., 2016, 2018), однако появления значительной чис-
ленности второго поколения не наблюдалась. Возможно, что высокая температура
именно июня и июля обеспечивает появление многочисленного второго поколения.
ЗАКЛЮЧЕНИЕ
В Ленинградской области в 2021 г. началась новая и потенциально сильная вспышка
массового размножения короеда-типографа. Перманентное присутствие значитель-
ного количества короеда-типографа в ельниках Карельского перешейка стало предпо-
сылкой быстрого увеличения численности этого вида.
Развитие вспышки размножения было спровоцировано резким повышением темпе-
ратуры в июне-июле 2021 г., за счет которого успешно развилось два сестринских и
два основных поколения. Массовое появление двух генераций короеда-типографа
в ельниках Карельского перешейка отмечено впервые. По бивольтинному типу разви-
валась часть популяции, примерно 56 % особей, остальная ее часть развивалась по
моновольтинному типу.
Короедный запас, обеспечивающий быстрое нарастание численности короеда-типо-
графа в условиях Карельского перешейка, можно оценить в 140-150 тыс. особей/га,
или 10-12 заселенных типографом елей на гектар. В описываемой нами вспышке
короедный запас был выше - 187 тыс. особей/га, что обеспечило появление потомства
в количестве около 3 млн особей/га, т. е. очень высокой численности, способной обес-
печить стремительное нарастание вспышки.
Быстрый и устойчивый тренд роста температуры и наличие большого объема кор-
мовой базы для короеда-типографа в Ленинградской обл. и в юго-западной части Ка-
релии за счет проведения санитарных рубок и других факторов (Селиховкин, 2021,
2022) и, соответственно, сохранение значительного короедного запаса позволяют
249
предположить, что данная вспышка массового размножения короеда-типографа будет
стремительно развиваться, а в ближайшем будущем можно ожидать ее повторения.
ФИНАНСИРОВАНИЕ
Исследование выполнено за счет гранта Российского научного фонда № 21-16-
СПИСОК ЛИТЕРАТУРЫ
Катаев О. А. 1948. Краткая история Лисинского учебно-опытного лесхоза в лесоэнтомологическом отноше-
нии. Рукопись. СПб.: Лесотехническая академия, 56 с.
Катаев О. А. 1952. Вторичные вредители хвойных древостоев Ленинградской области и меры борьбы с ни-
ми. Диссертация на соискание ученой степени кандидата биологических наук. СПб.: Лесотехническая
академия, 257 с.
Катаев О. А. 1956. Обзор санитарного состояния Лисинского лесного массива за 1787-1955 годы. Труды
Лесотехнической академии. № 73. Л., с. 49-58.
Маслов А. Д. 2010. Короед-типограф и усыхание еловых лесов. Пушкино: ВНИИЛМ, 138 с.
Методика организации и проведения работ по мониторингу лесов европейской части России по программе
ICP-Forest (методика ЕЭК ООН). Федеральная служба лесного хозяйства России, Инструкция от
21 февраля 1995 г.
Мозолевская Е. Г., Катаев О. А., Соколова Э. С. 1984. Методы лесопатологического обследования очагов
стволовых вредителей и болезней леса. М.: Лесная промышленность, 152 с.
Осетров А. В., Селиховкин А. В. 1998. Видовое разнообразие и динамика плотности популяций короедов
в хвойных древостоях, поврежденных сильными ветрами. Известия Санкт-Петербургской лесотехни-
ческой академии 6 (164): 27-34.
Селиховкин А. В. 2017. Эффективность санитарно-оздоровительных мероприятий в современных условиях
на примере Ленинградской области. Известия Санкт-Петербургской лесотехнической академии 221:
35-51.
Селиховкин А. В. 2022. Вспышкам массового размножения короедов в лесах России быть! В кн.: Материалы
Третьей Всероссийской конференции с международным участием «Мониторинг и биологические ме-
тоды контроля вредителей и патогенов древесных растений: от теории к практике». 11-15 апреля
2022 года. Москва (в печати).
Селиховкин А. В., Ахматович Н. А., Варенцова Е. Ю., Поповичев Б. Г. 2018. Размножение короеда типографа
и других дендропатогенных организмов на Карельском перешейке. Лесоведение (6): 426-433.
Селиховкин А. В., Варенцова Е. Ю., Поповичев Б. Г. 2017. Сплошные санитарные рубки как метод контроля
плотности популяций стволовых вредителей и распространения дендропатогенных организмов в со-
временных условиях на примере Ленинградской области. Известия Санкт-Петербургской лесотехни-
ческой академии 220: 186-199.
Селиховкин А. В., Глебов Р. Н., Магдеев Н. Г., Ахматович Н. А., Поповичев Б. Г. 2016. Оценка роли насеко-
мых и дендропатогенных организмов в усыхании древостоев Ленинградской области и Республики
Татарстан. Лесоведение 2: 83-95.
Шевырёв И. Я. 1896. Опустошительное размножение короедов в средней России с 1882 г. по 1894 г. и попыт-
ки борьбы с ними. Сельское хозяйство и лесоводство 10: 523-545.
Шиперович В. Я. 1931. Роль энтомофауны в отмирании деревьев в сосново-еловых насаждениях Лисинского
учебно-опытного леспромхоза. Труды Лесотехнической академии. Лесоводственный цикл. М.; Л.:
Государственное научно-техническое издательство, с. 202-240.
Alekseev A. S. 2018. Assessment and inventory of forest ecosystems biodiversity: Case study for Karelian Isthmus
of Leningrad Region, Russia. Open Journal of Ecology 8 (5): 305-323.
Jönsson A. M., Appelberg G., Harding S., Bärring L. 2009. Spatio-temporal impact of climate change on the activity
and voltinism of the spruce bark beetle, Ips typographus. Global Change Biology 15: 486-499.
Jönsson A. M., Harding S., Bärring L., Ravn H. P. 2007. Impact of climate change on the population dynamics of
Ips typographus in southern Sweden. Agricultural and Forest Meteorology 146: 70-81.
Jurc M., Perko M., Džeroski S., Demsar D., Hrasovec B. 2006. Spruce bark beetles (Ips typographus, Pityogenes
chalcographus, Col.: Scolytidae) in the Dinaric mountain forests of Slovenia: Monitoring and modeling.
Ecological Modeling 194: 219-226.
250
Lange H., Økland B., Krokene P. 2010. To be or twice to be? The life cycle development of the spruce bark beetle
under climate change. In: A. A. Minai, D. Braha, Y. Bar-Yam (eds). Unifying Themes in Complex Systems,
Pt. 2. Berlin: Springer Verlag, p. 251-258. (Proceedings of the 6th International Conference on Complex
Systems).
Öhrn P. 2012. The Spruce Bark Beetle Ips typographus in a Changing Climate - Effects of Weather Conditions on
the Biology of Ips typographus. Introductory Research Essay No 18. Uppsala: Department of Ecology, SLU,
27 p.
Schebeck M., Dobart N., Ragland G. J., Schopf A., Staufer C. 2021. Facultative and obligate diapause phenotypes
in populations of the European spruce bark beetle Ips typographus. Journal of Pest Science 95: 889-899.
Štefková K., Okrouhlík J., Doležal P. 2017. Development and survival of the spruce bark beetle, Ips typographus
(Coleoptera: Curculionidae: Scolytinae) at low temperatures in the laboratory and the field. European Journal
of Entomology 114: 1-6.
Wermelinger B. 2004. Ecology and management of the spruce bark beetle Ips typographus—a review of recent
research. Forest Ecology and Management 202: 67-82.
A NEW OUTBREAK OF THE EUROPEAN SPRUCE BARK BEETLE,
IPS TYPOGRAPHUS (L.) (COLEOPTERA, CURCULIONIDAE)
IN LENINGRAD PROVINCE
A. V. Selikhovkin, N. A. Mamaev, M. B. Martirova, S. A. Merkuriev, B. G. Popovichev
Key words: the European spruce bark beetle, Ips typographus, outbreak, generations number.
SUMMARY
A new and potentially strong outbreak of the European spruce bark beetle Ips typographus has begun
in the Leningrad region in 2021. The permanent presence of a significant quantity of the species in the
spruce forests of the Karelian Isthmus (Leningrad Province) created a basis for a rapid increase of the
abundance of this species.
The development of a breeding outbreak was provoked by a sharp increasing in temperature in
June-July 2021. Due to the increase in temperature, two sister and two main generations successfully
developed. The mass appearance of two generations of the European spruce bark beetle in the forests
of the Karelian Isthmus was revealed for the first time. A part of the population, approximately 56%
of individuals, developed according to the bivoltine type (two generations), while the rest developed
according to the univoltine type (one generation). This feature of development provides the European
spruce bark beetle with the resistance of populations to adverse weather conditions. The part of the
second generation that started to develop earlier had a high reproduction energy (the ratio of the number
of offspring beetles to the number of parent beetles - 4.7), close to the reproduction energy of the
first generation (6.0). The next group of the second generation, apparently, was mainly represented
by the offspring of the sister generation. Development indicators in this group were significantly
worse (reproduction energy - 1.3). A significant part of the individuals did not have time to complete
development.
The bark beetle stock (number of individuals per hectare), which ensures a rapid increase in the
number of spruce bark beetle in the conditions of the Karelian Isthmus, can be estimated at 140-
150 thousand beetles per hectare (b/ha). In the case under consideration, the bark beetle stock was
higher - 187 thousand b/ha, which ensured the appearance of offspring in the amount of about 3 million
b/ha, i.e. a very large number capable of providing a rapidly growing outbreak.
The rapid and steady trend of temperature growth and the presence of a large amount of food supply
for the European spruce bark beetle in Leningrad Province and in the southwestern part of Karelia, and
the preservation of the bark beetle stock suggest that this outbreak will develop rapidly, and in the near
future we can expect its recurrence.
251