ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 101, 2, 2022
УДК 595.745:591.499
СТРУКТУРА, РАЗНООБРАЗИЕ И РАСПРЕДЕЛЕНИЕ
АНТЕННАЛЬНЫХ СЕНСИЛЛ РУЧЕЙНИКОВ НАДСЕМ.
PSYCHOMYIOIDEA (TRICHOPTERA, ANNULIPALPIA)
© 2022 г. К. Т. Абу Дийак, * М. Ю. Валуйский, **
С. И. Мельницкий, *** В. Д. Иванов ****
Кафедра энтомологии, Санкт-Петербургский государственный университет
Университетская наб., 7/9, С.-Петербург, 199034 Россия
* e-mail: kdiyak@gmail.com, ** e-mail: sphingonaepiopsis@gmail.com,
*** e-mail: simelnitsky@gmail.com, ****e-mail: v--ivanov@yandex.ru
Поступила в редакцию 21.02.2022 г.
После доработки 4.03.2022 г.
Принята к публикации 4.03.2022 г.
Впервые выполнено сравнительно-морфологическое исследование строения и локализации
сенсилл на антеннах ручейников надсем. Psychomyioidea. Методами электронной микроско-
пии была изучена морфология антеннальных сенсилл у 21 вида из семейств Psychomyiidae (6),
Xiphocentronidae (1), Dipseudopsidae (3), Polycentropodidae (9) и Pseudoneureclipsidae (2). На
антеннах исследованных видов было обнаружено 7 различных типов сенсилл: длинные три-
хоидные, изогнутые трихоидные, хетоидные, базиконические, коронарные, стилоконические и
псевдоплакоидные сенсиллы. Распределение различных типов сенсилл у изученных семейств
в основном соответствует исходному плану строения сенсорной поверхности Trichoptera.
Длинные трихоидные и псевдоплакоидные сенсиллы обычно имеют неспецифическое распре-
деление, хетоидные - фиксированное, базиконические, коронарные и стилоконические сенсил-
лы - специфическое. Сенсорные поля изогнутых трихоидных сенсилл расположены на ба-
зальных сегментах антенны и всегда находятся на вентролатеральной стороне флагелломера.
У изученных видов найдены три варианта формы сенсорных полей. Сенсорная поверхность ан-
тенн представителей надсем. Psychomyioidea в основном имеет консервативное строение, при
этом у отдельных эволюционных линий наблюдаются модификации в строении и локализации
сенсилл.
Ключевые слова: сенсиллы, сенсорные поля, антенны, Trichoptera, ручейники, Annulipalpia,
Psychomyioidea, эволюция.
DOI: 10.31857/S0367144522020113, EDN: HKVTWY
Ручейники (Trichoptera) - отряд насекомых с полным превращением, насчитыва-
ющий около 17 000 видов (Morse, 2022), вместе с чешуекрылыми (Lepidoptera) и тара-
хоптерами (Tarachoptera) они объединяются в надотряд Amphiesmenoptera. Отряд
Trichoptera подразделяется на три подотряда: два современных - кольчатощупиковые
(Annulipalpia) и цельнощупиковые (Integripalpia) - и ископаемый подотряд Proto-
meropina (Ivanov, Sukacheva, 2002; Kopylov et al., 2020). Внутри кольчатощупиковых
333
ручейников выделяются три эволюционные линии, которые рассматриваются в ранге
надсемейств: Philopotamoidea, Hydropsychoidea и Psychomyioidea (Holzenthal et al.,
2011). Надсем. Psychomyioidea включает 7 современных семейств: Psychomyiidae
Walker, 1852; Xiphocentronidae Ross, 1949; Ecnomidae Ulmer, 1903; Dipseudopsidae
Ulmer, 1904; Polycentropodidae Ulmer, 1903; Kambaitipsychidae Malicky, 1992 и
Pseudoneureclipsidae Ulmer, 1951, а также два ископаемых семейства - Electralbertidae
Botosaneanu et Wichard, 1983 и Protobaikalopsychidae Melnitsky et Ivanov, 2020, из-
вестные по единичным находкам из меловых местонахождений Канады и России.
В состав надсемейства входит более 2350 видов из 70 родов. Монофилия Psycho-
myioidea была подтверждена молекулярно-генетическими и морфологическими дан-
ными (Kjer et al., 2016; Thomas et al., 2020).
Предыдущие исследования выявили на антеннах ручейников восемь типов сенсилл:
длинные трихоидные, изогнутые трихоидные, хетоидные, псевдоплакоидные, базико-
нические, коронарные, целоконические и стилоконические (Ivanov, Melnitsky, 2011,
2016; Melnitsky, Ivanov, 2011, 2016; Valuyskiy et al., 2017, 2019, 2020a, 2020b; Melnitsky
et al., 2018). При этом максимальное количество подтипов было обнаружено у псевдо-
плакоидных сенсилл: грибовидные, рожковидные, звездчатые, зубчатые, листовидные,
вильчатые, мультивильчатые, двулопастные, рассеченные, копьевидные, ушковидные,
Т-образные и гребневидные (Ivanov, Melnitsky, 2011, 2016; Melnitsky, Ivanov, 2011;
Valuyskiy et al., 2017, 2020b; Abu Diiak et al., 2021). В пределах одного семейства
обычно отмечается от 7 до 10 типов и подтипов сенсилл (Ivanov, Melnitsky, 2011, 2016;
Melnitsky, Ivanov, 2011, 2016; Valuyskiy et al., 2019, 2020a). Однако в сем. Rhyacophilidae
встречаются сенсиллы 13 подтипов, подобное разнообразие достигается путем струк-
турной дифференциации псевдоплакоидных сенсилл (Valuyskiy et al., 2017). Исходное
состояние сенсорной поверхности антенн у ручейников предположительно включает
шесть типов сенсилл: длинные трихоидные, изогнутые трихоидные, хетоидные, псев-
доплакоидные, коронарные и стилоконические (Valuyskiy et al., 2020a). Высокое
структурное разнообразие сенсилл на антеннах ручейников может быть связано со
сложной и разнообразной феромонной коммуникацией у этих насекомых (Ivanov,
1993; Ivanov, Melnitsky, 2014).
На антеннах ручейников выявлены четыре типа распределения сенсилл: неспецифи-
ческое, специфическое, фиксированное и групповое (сенсорные поля и сенсорные
скопления) (Valuyskiy et al., 2017). Неспецифическое распределение обычно харак-
терно для длинных трихоидных и псевдоплакоидных сенсилл разных подтипов: эти
сенсиллы более или менее равномерно покрывают всю поверхность сегмента ан-
тенны. Специфическое распределение свойственно базиконическим, коронарным и
стилоконическим сенсиллам, они встречаются только в определенной области сег-
мента. Хетоидные сенсиллы характеризуются фиксированным распределением, по-
стоянным положением на сегментах антенны. Изогнутые трихоидные сенсиллы со-
браны в сенсорные поля различной формы на вентральной стороне сегментов.
Количество сенсилл всех типов на сегмент уменьшается к вершине антенны (Valuyskiy
et al., 2017, 2019, 2020a, 2020b; Melnitsky et al., 2018; Abu Diiak et al., 2021).
Сенсорная поверхность антенн подробно исследована у 10 семейств ручейников:
в подотряде Annulipalpia-Philopotamidae (Kubiak et al., 2015; Melnitsky et al., 2018),
Ecnomidae (Valuyskiy et al., 2019 и Hydropsychidae (Abu Diiak et al., 2021); а в под-
отряде Integripalpia - Rhyacophilidae (Valuyskiy et al., 2017), Glossosomatidae (Valuyskiy
334
et al., 2020a), Hydroptilidae (Wells, 1984), Apataniidae (Valuyskiy et al., 2020b),
Limnephilidae (Ivanov, Melnitsky, 2011), Phryganeidae (Ivanov, Melnitsky, 2011) и
Molannidae (Melnitsky, Ivanov, 2016). В приведенных выше работах сенсорную по-
верхность антенн изучали в основном у самцов, так как они во многих группах ручей-
ников коммуникационно активны и используют сенсиллы антенн для поиска полового
партнера (Ivanov, Melnitsky, 1999, 2014). У исследованных семейств подотряда Annuli-
palpia было обнаружено более низкое разнообразие сенсилл и сенсорных полей, чем у
Integripalpia (Melnitsky, Ivanov, 2011; Ivanov, Melnitsky, 2016). Мы предполагаем, что
Annulipalpia в целом характеризуются меньшим разнообразием сенсилл и сенсорных
полей, чем Integripalpia, но в отдельных таксонах кольчатощупиковых ручейников
может наблюдаться высокая вариабельность сенсорных структур (Abu Diiak
et al., 2021). Для проверки этого предположения мы провели сравнительный анализ с
привлечением ранее не исследованных семейств подотряда Annulipalpia.
Новые сравнительно-морфологические данные по различным семействам подот-
ряда Annulipalpia могут быть использованы в комплексном филогенетическом ана-
лизе, что позволит установить основные направления эволюции поверхности антенн у
Annulipalpia и ручейников в целом. Эти данные также будут в дальнейшем сопостав-
лены со сведениями о сенсорных комплексах антенн и сенсилл на ротовых придатках,
изучение которых запланировано в ближайшей перспективе. Особенности функциони-
рования сенсорных комплексов антенн у насекомых не могут быть изучены без пред-
варительного анализа их строения. В конечном итоге сравнительные данные об антен-
нальных сенсиллах могут внести ясность в понимание эволюции коммуникационных
систем ручейников и всего класса насекомых.
МАТЕРИАЛ И МЕТОДИКА
Сенсорная поверхность антенн исследована у самцов 21 вида ручейников из пяти семейств
подотряда Annulipalpia: Neureclipsis bimaculata (Linnaeus, 1758) (материал из России, Ленин-
градская обл.); Polycentropus flavomaculatus (Pictet, 1834) (Швеция); Plectrocnemia conspersa
(Curtis, 1834) (Швеция); Cyrnus fennicus Klingstedt, 1937 (Россия, Ленинградская обл.);
C. trimaculatus (Curtis, 1834) (Россия, Ленинградская обл.); C. flavidus McLachlan, 1864 (Россия,
Ленинградская обл.); Holocentropus picicornis (Stephens, 1836) (Россия, Ленинградская обл.);
H. insignis Martynov, 1924 (Россия, Ленинградская обл.); Pahamunaya taleban Malicky et Chan-
taramongkol, 1993 (Таиланд) (Polycentropodidae); Psychomyia minima (Martynov, 1910) (Монго-
лия); Ps. flavida Hagen, 1861 (Россия, Дальний Восток); Ps. pusilla (Fabricius, 1781) (Россия,
Северный Кавказ); Lype phaeopa (Stephens, 1836) (Россия, Ленинградская обл.); Tinodes turanicus
Martynov,
1927 (Узбекистан); T. waeneri (Linnaeus,
1758) (Россия, Ленинградская обл.)
(Psychomyiidae); Xiphocentron mnesteus Schmid, 1982 (Колумбия) (Xiphocentronidae); Dipseudopsis
indica McLachlan, 1875 (Индия); D. varians Ulmer, 1929 (Малайзия); Hyalopsyche sachalinica
Martynov, 1910 (Россия, Дальний Восток) (Dipseudopsidae); Pseudoneureclipsis proxima Martynov,
1934 (Россия, Дальний Восток); Ps. chrysippus Malicky et Sompong,
2000 (Малайзия)
(Pseudoneureclipsidae).
Исследование было выполнено с применением сканирующей электронной микроскопии
(СЭМ). Был использован материал из коллекций кафедры энтомологии СПбГУ и Зоологическо-
го института РАН, а также собственные сборы. Образцы хранились в 70-96%-ном этаноле или в
сухом виде. Антенны были удалены под диссекционным стереомикроскопом с помощью пинце-
та, высушены на воздухе и прикреплены на столики для электронной микроскопии на токопро-
водящий скотч вентральной стороной кверху. После этого образцы покрывались слоем золота
толщиной 20 нм с помощью напылительной установки Leica EM SCD500. Цифровые фотогра-
335
фии образцов были получены с использованием сканирующих электронных микроскопов JEOL
NeoScope JCM-5000 и Tescan MIRA3 в ресурсном центре СПбГУ «Развитие молекулярных и
клеточных технологий».
Для подсчета и измерения сенсилл использовалась программа ImageJ 1.52a: с помощью ин-
струмента «Multi-point tool» были подсчитаны сенсиллы в сенсорных полях на видимой поверх-
ности сегментов. Пересчет числа сенсилл в поле зрения на всю поверхность сегментов был
выполнен в зависимости от особенностей расположения каждого типа сенсилл. Для удобства
визуализации средние значения количества сенсилл и ошибки среднего вычислялись для групп
из 5 флагелломеров (1-5, 6-10 и т. д.). Для ручной калибровки масштаба и измерения размеров
сенсилл были применены инструменты модулей Edit и Analyze программы ImageJ: функции
«Straight line» / «Segmented line», а также команды «Measure» и «Set scale». У грибовидных псев-
доплакоидных сенсилл в зависимости от формы проводились измерения диаметра (у округлых)
или длины (у вытянутых), у других типов сенсилл была измерена длина. Средние размеры сен-
силл и соответствующие ошибки среднего вычислены на основе 10 образцов данного типа сен-
силл с разных сегментов или с одного и того же сегмента. Все вычисления выполнены с помощью
программы Excel из пакета Microsoft Office 2016.
РЕЗУЛЬТАТЫ
Структурное разнообразие сенсилл
На антеннах исследованных видов подотряда Annulipalpia было обнаружено 9 раз-
личных типов сенсилл. Длинные трихоидные, хетоидные, изогнутые трихоидные и
грибовидные псевдоплакоидные сенсиллы были найдены у всех изученных видов
(табл. 1).
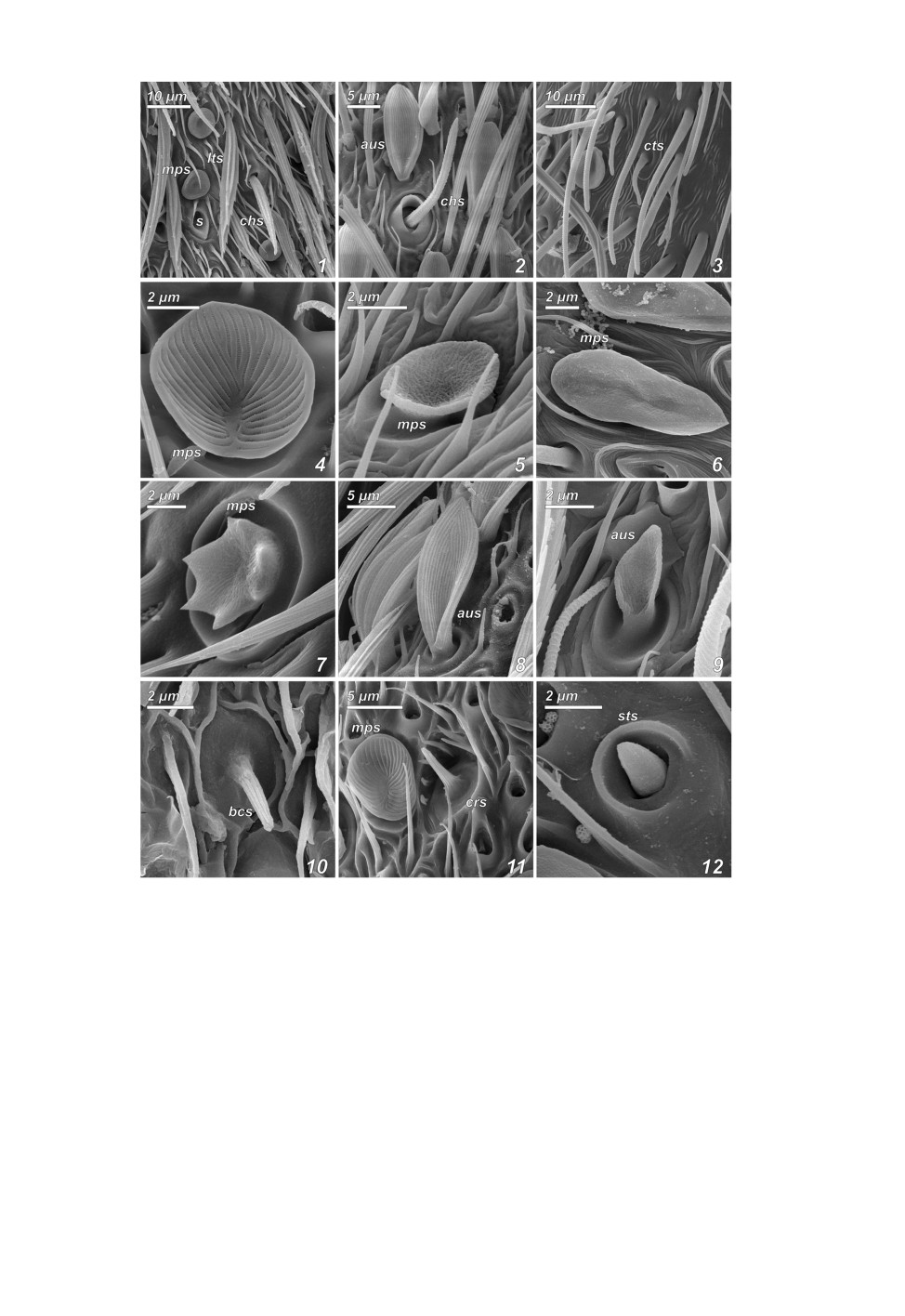
Дл инн ые т р и хо идн ы е с е нс и лл ы (рис. 1, 1: lts) - удлиненные, сильно на-
клоненные к поверхности антенны и уплощенные структуры, имеющие суженное ос-
нование и удлиненные каплевидные теки. У всех исследованных видов сенсиллы
этого типа ребристые, с продольным расположением зазубренных ребер. Они всегда
наклонены в дистальном направлении и покрывают всю поверхность сегмента, кроме
участков, занятых другими сенсиллами. Размер длинных трихоидных сенсилл у изу-
ченных видов находится в пределах от 29 ± 1.2 (Tinodes waeneri) до 57.2 ± 3.5 μm
(Dipseudopsis indica) (рис. 2, 1).
Х ето ид ны е с е нс и л лы (рис. 1, 1, 2: chs) имеют волосковидную форму, про-
дольную исчерченность и круглое поперечное сечение. Эти сенсиллы расположены
почти под прямым углом к поверхности кутикулы и у большинства исследованных
видов имеют полусферические теки (рис. 1, 1). У Pseudoneureclipsis chrysippus и
Ps. proxima (Pseudoneureclipsidae) края тек приподняты и уплощены (рис. 1, 2). Размер
этих структур варьирует в пределах от 17.1 ± 0.7 (Tinodes turanicus) до 35.2 ± 0.6 μm
(Dipseudopsis varians) (рис. 2, 2).
И зо гну т ые т р и хо идн ы е с е н с ил лы (рис. 1, 3: cts; 7, 10-13) короче и
тоньше, чем длинные трихоидные сенсиллы, и обычно имеют кольчатую исчерчен-
ность. У многих исследованных видов эти сенсиллы собраны в сенсорные поля на
вентролатеральной стороне флагелломеров (см. табл. 1). У большинства видов эти
сенсиллы на антенне одного размера, и лишь у двух семейств были обнаружены раз-
личающиеся размерами и, иногда, характером поверхности подтипы этих сенсилл.
Изогнутые трихоидные сенсиллы имеют длину от 13.1 ± 0.7 до 39.7 ± 1 μm (два под-
типа изогнутых трихоидных сенсилл у Hyalopsyche sachalinica) (рис. 2, 3).
336
Таблица 1. Наличие сенсорных полей и разных типов сенсилл у изученных видов из группы
Psychomyioidea
Виды
sf
lts
chs
cts
mps
aus
tps
bcs
crs
sts
Сем. POLYCENTROPODIDAE
Neureclipsis bimaculata (Linnaeus, 1758)
+
+
+
+
+
-
+
+
-
+
Polycentropus flavomaculatus (Pictet, 1834)
+
+
+
+
+
-
-
+
-
-
Plectrocnemia conspersa (Curtis, 1834)
-
+
+
+
+
-
-
+
+
+
Cyrnus fennicus Klingstedt, 1937
-
+
+
+
+
+
-
+
-
+
C. trimaculatus (Curtis, 1834)
-
+
+
+
+
-
-
+
-
+
C. flavidus McLachlan, 1864
+
+
+
+
+
-
-
+
-
+
Holocentropus picicornis (Stephens, 1836)
+
+
+
+
+
-
-
+
-
+
H. insignis Martynov, 1924
+
+
+
+
+
-
-
+
-
+
Pahamunaya taleban Malicky et Chantaramongkol, 1993
-
+
+
+
+
-
-
+
-
-
Сем. PSYCHOMYIIDAE
Psychomyia minima (Martynov, 1910)
+
+
+
+
+
-
-
-
-
+
Ps. flavida Hagen, 1861
+
+
+
+
+
-
-
-
+
+
Ps. pusilla (Fabricius, 1781)
+
+
+
+
+
-
-
-
+
+
Lype phaeopa (Stephens, 1836)
+
+
+
+
+
-
-
+
-
+
Tinodes turanicus Martynov, 1927
+
+
+
+
+
-
-
-
+
+
T. waeneri (Linnaeus, 1758)
-
+
+
+
+
-
-
+
-
+
Сем. XIPHOCENTRONIDAE
Xiphocentron mnesteus Schmid, 1982
-
+
+
+
+
-
-
+
-
+
Сем. DIPSEUDOPSIDAE
Dipseudopsis indica McLachlan, 1875
-
+
+
+
+
-
-
-
-
+
D. varians Ulmer, 1929
-
+
+
+
+
-
-
+
-
+
Hyalopsyche sachalinica Martynov, 1910
-
+
+
+
+
-
-
+
-
+
Сем. PSEUDONEURECLIPSIDAE
Pseudoneureclipsis proxima Martynov, 1934
-
+
+
+
+
+
-
-
+
+
Ps. chrysippus Malicky et Sompong, 2000
-
+
+
+
+
+
-
-
+
+
П р и м е ч а н и е. sf - сенсорные поля, lts - длинные трихоидные сенсиллы, chs - хетоидные сенсиллы,
cts - изогнутые трихоидные сенсиллы, mps - грибовидные псевдоплакоидные сенсиллы, aus - ушковидные
псевдоплакоидные сенсиллы, tps - Т-образные псевдоплакоидные сенсиллы, bcs - базиконические
сенсиллы, crs - коронарные сенсиллы, sts - стилоконические сенсиллы.
Для всех изученных Dipseudopsidae и Pseudoneureclipsidae характерно наличие двух
подтипов изогнутых трихоидных сенсилл. На антеннах Dipseudopsidae есть более ко-
роткие (13.7 ± 0.4 μm у Dipseudopsis indica, 19.6 ± 0.4 μm у D. varians и 13.1 ± 0.7 μm
у H. sachalinica) и более длинные (21.4 ± 0.7 μm у D. indica, 28.4 ± 0.7 μm у D. varians,
и 39.7 ± 1 μm у H. sachalinica) изогнутые трихоидные сенсиллы, оба подтипа обла-
337
Рис. 1. Сенсиллы на антеннах ручейников надсем. Psychomyioidea.
1 - длинная трихоидная и хетоидная сенсиллы Holocentropus picicornis (Stephens, 1836); 2 - хетоидная
сенсилла Pseudoneureclipsis proxima Martynov, 1934; 3 - изогнутые трихоидные сенсиллы Hyalopsyche
sachalinica Martynov, 1910; 4 - грибовидная псевдоплакоидная сенсилла Psychomyia flavida Hagen,
1861; 5 - грибовидная псевдоплакоидная сенсилла Cyrnus fennicus Klingstedt, 1937; 6 - грибовидная
псевдоплакоидная сенсилла Pahamunaya taleban Malicky et Chantaramongkol, 1993; 7 - грибовидная
псевдоплакоидная сенсилла Dipseudopsis varians Ulmer, 1929; 8 - ушковидная псевдоплакоидная сенсилла
Pseudoneureclipsis proxima; 9 - ушковидная псевдоплакоидная сенсилла C. fennicus; 10 - базиконическая
сенсилла Xiphocentron mnesteus Schmid, 1982; 11 - грибовидная псевдоплакоидная и коронарная сенсиллы
Psychomyia flavida; 12 - стилоконическая сенсилла Tinodes waeneri (Linnaeus, 1758).
aus - ушковидная псевдоплакоидная сенсилла, bcs - базиконическая сенсилла, chs - хетоидная сенсилла,
crs - коронарная сенсилла, cts - изогнутая трихоидная сенсилла, lts - длинная трихоидная сенсилла,
mps - грибовидная псевдоплакоидная сенсилла, s - пустой сокет длинной трихоидной сенсиллы,
sts - стилоконическая сенсилла.
338
Рис. 2. Размеры сенсилл на антеннах Psychomyioidea, μm.
1 - длинные трихоидные сенсиллы, 2 - хетоидные сенсиллы, 3 - различные подтипы изогнутых
трихоидных сенсилл (an - кольчатые изогнутые трихоидные сенсиллы, ct - изогнутые трихоидные
сенсиллы без модификаций, l - более длинные изогнутые трихоидные сенсиллы, sh - укороченные
изогнутые трихоидные сенсиллы, sm - гладкие изогнутые трихоидные сенсиллы), 4 - базиконические
сенсиллы, 5 - грибовидные псевдоплакоидные сенсиллы, 6 - стилоконические сенсиллы разных подтипов
(hs - стилоконические сенсиллы с полусферической текой, st - стилоконические
сенсиллы без модификаций).
Планки погрешностей - ошибки среднего.
339
дают слабой кольчатой исчерченностью (рис. 1, 3; 7, 10, 13). У Pseudoneureclipsidae
изогнутые трихоидные сенсиллы гладкие (иногда слабо кольчатые, рис. 7, 10) либо
кольчатые (с глубокой кольчатой исчерченностью, рис. 7, 12). Кольчатые изогнутые
трихоидные сенсиллы длиннее гладких: их длина 33.2 ± 2.5 (Pseudoneureclipsis
proxima) и 24.9 ± 0.8 μm (Ps. chrysippus), а длина гладких изогнутых трихоидных сен-
силл составляет 18 ± 1 (Ps. proxima) и 19.3 ± 1.3 μm (Ps. chrysippus) (рис. 2, 3). Для
Psychomyiidae (кроме Tinodes waeneri) характерна модификация изогнутых три-
хоидных сенсилл: у них исчерченность продольная вместо кольчатой (рис. 7, 11).
Гр и бо в ид ны е п с е вд о пл а ко и дн ы е с е н си л лы (рис. 1, 1, 4-7, 11: mps;
7, 1-6) имеют расширенную апикальную часть, сидящую на короткой толстой ножке и
обычно покрытую сетью бороздок с порами. У большинства изученных видов апи-
кальная часть грибовидных псевдоплакоидных сенсилл круглая и более или менее во-
гнутая (рис. 1, 1, 4, 5, 11). Размер грибовидных псевдоплакоидных сенсилл варьирует
от 4.5 ± 0.2 (Dipseudopsis indica) до 11.5 ± 0.4 μm (Xiphocentron mnesteus) (рис. 2, 5).
У Pahamunaya taleban (Polycentropodidae) и X. mnesteus (Xiphocentronidae) эти сен-
силлы имеют форму эллипса, вытянутого параллельно оси антенны (рис. 1, 6; 7, 2, 6).
Для X. mnesteus характерна продольная исчерченность грибовидных псевдоплако-
идных сенсилл (рис. 7, 6). Апикальная поверхность грибовидных псевдоплакоидных
сенсилл Psychomyiidae (кроме Tinodes waeneri) несет расходящиеся от центра, слабо
ветвящиеся борозды, в которых видны поры (рис. 1, 4, 11; 7, 3, 4). Грибовидные псев-
доплакоидные сенсиллы у видов рода Psychomyia слегка вытянуты в дистальном на-
правлении (рис. 1, 4, 11; 7, 4). У Dipseudopsis varians апикальная часть грибовидных
псевдоплакоидных сенсилл разделена на несколько коротких заостренных лопастей
(рис. 1, 7; 7, 5).
Уш ко в ид ны е пс евдо п ла ко ид ны е с е н с и лл ы (рис. 1, 2, 8, 9: aus;
7, 7, 8) представляют собой видоизмененные грибовидные псевдоплакоидные сен-
силлы. Они были найдены у Cyrnus fennicus (Polycentropodidae), Pseudoneureclipsis
chrysippus и Ps. proxima (Pseudoneureclipsidae). Это вытянутые структуры, плоские в
поперечном сечении. У Ps. chrysippus и Ps. proxima поверхность этих сенсилл покрыта
продольными неветвящимися бороздами, в которых есть поры (рис. 1, 2, 8; 7, 8).
Длина ушковидных псевдоплакоидных сенсилл составляет 4.6 ± 0.1 μm у Cyrnus
fennicus, 11.6 ± 0.2 μm у Ps. chrysippus, и 15.2 ± 0.3 μm у Ps. proxima.
Т -о бр а зны е пс е вд о пл а ко и дн ы е с ен с и ллы (рис. 7, 9) обнаружены
только у Neureclipsis bimaculata. Отросток у этих сенсилл плоский в поперечном се-
чении и T-образный, расположен параллельно продольной оси сегмента. Основание
сенсиллы овальное, располагается ниже уровня окружающей кутикулы. Длина Т-об-
разных псевдоплакоидных сенсилл составляет 9.5 ± 0.6 μm.
Ба зико ниче с ки е с е н си л лы (рис. 1, 10: bcs) - небольшие волосковидные
структуры с продольной исчерченностью, сидящие на округлом основании. Этот тип
сенсилл найден у всех Polycentropodidae, Lype phaeopa, Tinodes waeneri (Psycho-
myiidae), Xiphocentron mnesteus (Xiphocentronidae), Dipseudopsis varians и Hyalopsyche
sachalinica (Dipseudopsidae). Длина отростка базиконических сенсилл составляет от
3.1 ± 0.2 (Plectrocnemia conspersa) до 6.6 ± 0.2 μm (D. varians) (рис. 2, 4).
Коронарные с енсиллы (рис. 1, 11: crs) отличаются от базиконических нали-
чием модифицированных микротрихий вокруг основания сенсиллы. Эти сенсиллы
340
были обнаружены у Plectrocnemia conspersa (Polycentropodidae), Psychomyia flavida,
Ps. pusilla, Tinodes turanicus (Psychomyiidae), Ps. chrysippus и Ps. proxima (Pseudo-
neureclipsidae). У изученных видов микротрихии окружают сенсиллу частично:
у Plectrocnemia conspersa, Psychomyia pusilla и Ps. flavida микротрихии расположены в
дистальной части теки, а у Polycentropus flavomaculatus, T. turanicus, Pseudoneureclipsis
chrysippus и Ps. proxima немногочисленные микротрихии (обычно 3-5) расположены
более или менее равномерно вокруг сенсиллы. Дистальный край основания коро-
нарных сенсилл обычно слегка погружен ниже уровня окружающей кутикулы (кроме
Ps. proxima). У Ps. proxima основание этих сенсилл находится на уровне окружающей
кутикулы. Длина отростка коронарных сенсилл составляет от
3.2
±
0.1 μm
(Plectrocnemia conspersa) до 5.3 ± 0.2 μm (Psychomyia flavida).
Ст ило ко н иче с к ие с е н с ил л ы (рис. 1, 12: sts) - очень короткие структуры
конусовидной формы, не имеющие видимых борозд и пор. Эти сенсиллы обнаружены
у всех изученных видов, кроме Pahamunaya taleban и Polycentropus flavomaculatus
(Polycentropodidae). Длина отростка стилоконических сенсилл варьирует от 1.6 ± 0.1
(Neureclipsis bimaculata) до 5 ± 0.2 μm (Dipseudopsis indica) (рис. 2, 6).
На флагелломерах D. indica (Dipseudopsidae) обнаружены два подтипа стилокониче-
ских сенсилл: первый имеет вогнутую теку с приподнятым краем, а у второго подтипа
тека выпуклая, полусферическая. На антеннах D. varians все стилоконические сен-
силлы с текой выпуклой полусферической формы.
Количество и распределение сенсилл
У изученных видов найдены четыре типа распределения сенсилл.
Ф икс ир о ван но е распределение характерно для хетоидных сенсилл у всех
изученных видов: у них более или менее постоянное положение на протяжении всей
антенны. У видов сем. Polycentropodidae два кольца хетоидных сенсилл - на прокси-
мальной и дистальной частях сегмента. У представителей Psychomyiidae (кроме
Tinodes waeneri) и Xiphocentron mnesteus (Xiphocentronidae) флагелломеры с одним
полным кольцом хетоидных сенсилл у дистального края, на остальной поверхности
сегмента могут встречаться одиночные хетоидные сенсиллы. Для T. waeneri
(Psychomyiidae), Dipseudopsidae и Pseudoneureclipsidae характерно наличие одного
кольца хетоидных сенсилл примерно посередине каждого сегмента флагеллума.
Н е с пец ифи че с ко е распределение имеют длинные трихоидные, изогнутые
трихоидные (Plectrocnemia conspersa, Cyrnus fennicus, C. trimaculatus, Pahamunaya
taleban, Dipseudopsis indica, D. varians, Hyalopsyche sachalinica и Tinodes waeneri),
грибовидные псевдоплакоидные, ушковидные псевдоплакоидные (C. fennicus, Pseudo-
neureclipsis chrysippus и Ps. proxima) и базиконические сенсиллы (Neureclipsis bi-
maculata, Pahamunaya taleban и Xiphocentron mnesteus). При данном типе распреде-
ления сенсиллы покрывают всю поверхность сегмента.
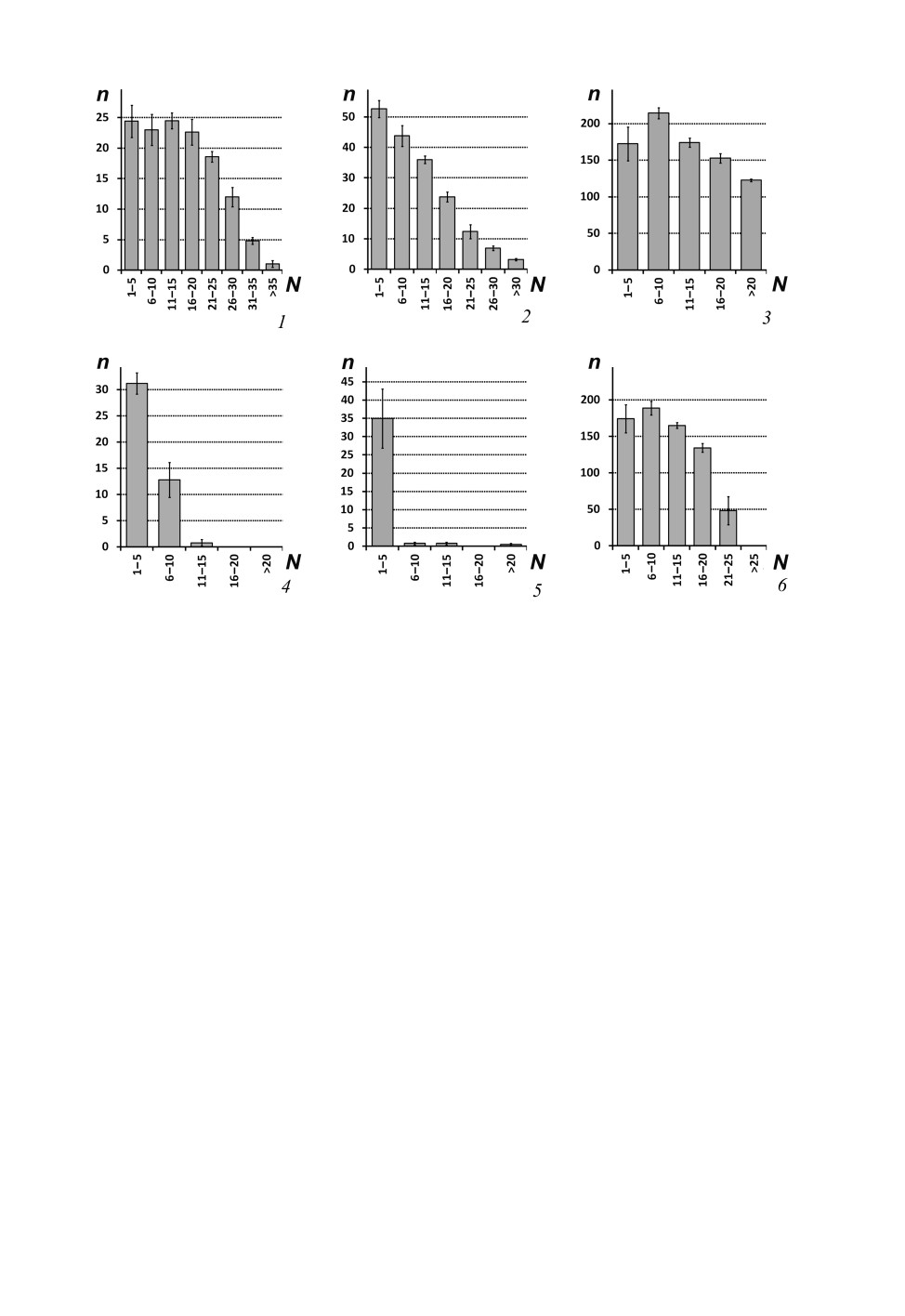
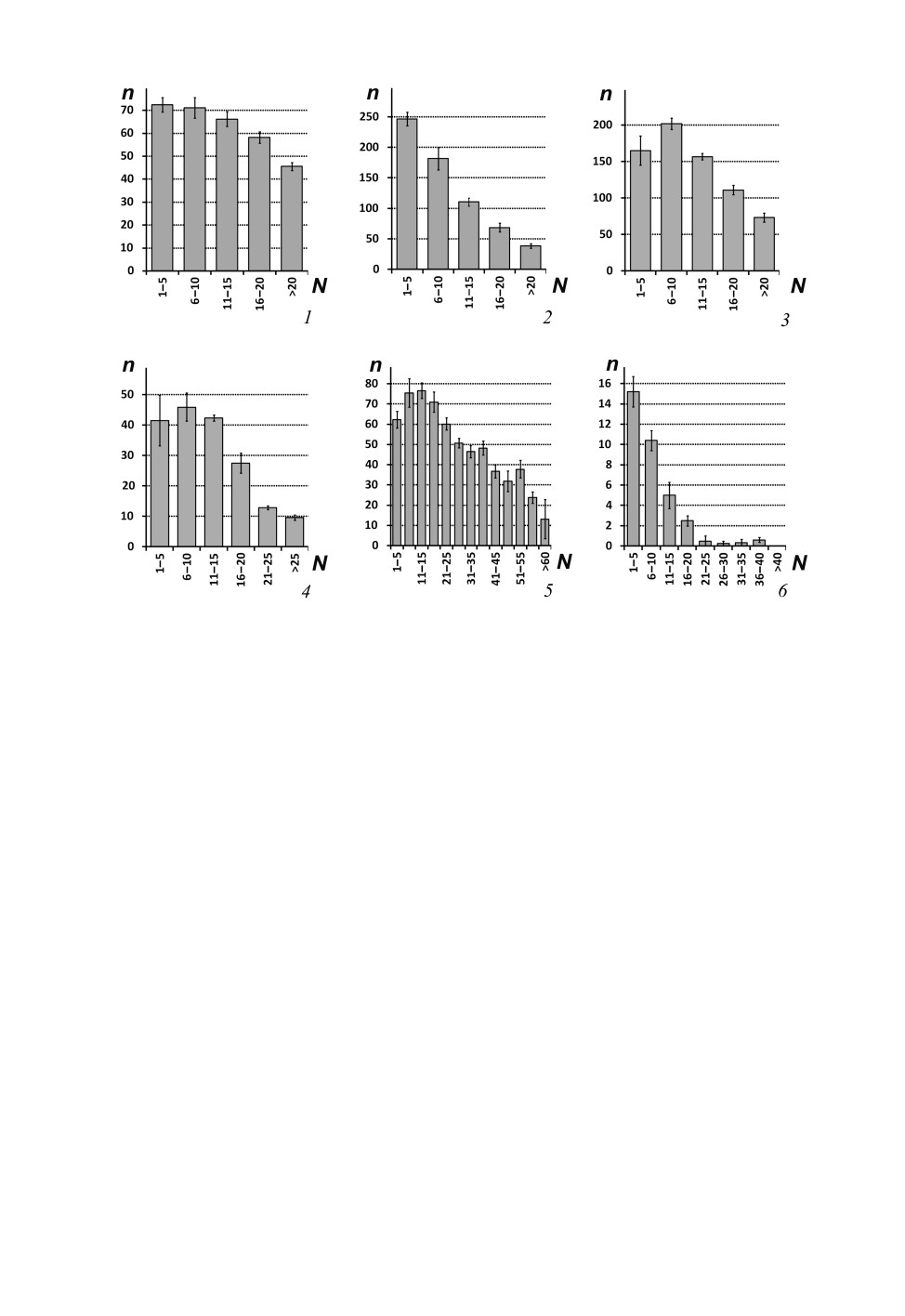
Количество грибовидных псевдоплакоидных сенсилл максимально на прокси-
мальных сегментах флагеллума и снижается на более дистальных флагелломерах
(рис. 6, 1-5). Максимальное число грибовидных псевдоплакоидных сенсилл на фла-
гелломер составляет от 32 (Cyrnus fennicus) до 243 (C. flavidus) у Polycentropodidae; от
30 (Psychomyia minima) до 268 (Ps. flavida) у Psychomyiidae; у Dipseudopsis indica,
341
D. varians и Hyalopsyche sachalinica (Dipseudopsidae) - 93, 186 и 11 соответственно;
у Pseudoneureclipsis proxima и Ps. chrysippus (Pseudoneureclipsidae) - 7 и 5 соответ-
ственно. Для Psychomyia minima характерно резкое уменьшение числа грибовидных
псевдоплакоидных сенсилл до полного отсутствия на 8-м и последующих флагелло-
мерах.
Максимальное число неспецифически распределенных ушковидных псевдоплако-
идных сенсилл на сегмент сильно различается у Cyrnus fennicus, Pseudoneureclipsis
proxima и Ps. chrysippus - 12, 256 и 109 соответственно. Число неспецифически рас-
пределенных базиконических сенсилл на флагелломер достигает 12 у Neureclipsis
bimaculata, 33 у Pahamunaya taleban, и 82 у Xiphocentron mnesteus.
Изогнутые трихоидные сенсиллы многочисленны и демонстрируют неспецифиче-
ское распределение по всей поверхности сегмента у некоторых Polycentropodidae
(Plectrocnemia conspersa, Cyrnus fennicus, C. trimaculatus и Pahamunaya taleban), а
также у Dipseudopsidae и Tinodes waeneri (Psychomyiidae) (рис. 3, 2, 3, 5). Макси-
мальное количество изогнутых трихоидных сенсилл на сегмент при таком типе рас-
пределения варьирует от 236 (Pahamunaya taleban) до 362 (Plectrocnemia conspersa)
у Polycentropodidae, и от 195 (Dipseudopsis varians) до 576 (Hyalopsyche sachalinica)
у Dipseudopsidae. У Tinodes waeneri предельное число изогнутых трихоидных сенсилл
на сегмент составляет 214. Количество изогнутых трихоидных сенсилл на антеннах
Plectrocnemia conspersa, Cyrnus fennicus, C. trimaculatus, Pahamunaya taleban,
Hyalopsyche sachalinica, Dipseudopsis varians и D. indica убывает очень медленно, и
дистальные сегменты флагеллума несут много этих сенсилл (рис. 5, 3). При этом для
Dipseudopsis varians и D. indica характерно наличие двух подтипов изогнутых трихо-
идных сенсилл: более короткие многочисленны и равномерно распределены по вен-
тролатеральной стороне сегментов, а более длинные - одиночные и находятся у дис-
тального края сегмента. У Tinodes waeneri число изогнутых трихоидных сенсилл резко
уменьшается после 24-го флагелломера, вплоть до нуля на субапикальных сегментах.
С пе циф иче ско е распределение имеют изогнутые трихоидные (Xiphocentron
mnesteus, Pseudoneureclipsis chrysippus и Ps. proxima), Т-образные (Neureclipsis
bimaculata), базиконические (кроме N. bimaculata, Pahamunaya taleban и Xiphocentron
mnesteus), коронарные и стилоконические сенсиллы. Сенсиллы с таким типом распре-
деления располагаются на определенных частях поверхности флагелломеров.
Изогнутых трихоидных сенсилл на антеннах X. mnesteus, Pseudoneureclipsis
chrysippus и Ps. proxima немного, и сенсорные поля не формируются (рис. 3, 6).
У X. mnesteus одиночные изогнутые трихоидные сенсиллы (не более 12 на сегмент)
распределены хотя и случайным образом, но лишь по вентролатеральной стороне фла-
гелломеров. Антенны Ps. chrysippus и Ps. proxima несут изогнутые трихоидные сен-
силлы двух подтипов (0-5 сенсилл на сегмент), которые расположены только у дис-
тального края сегментов.
Т-образные псевдоплакоидные сенсиллы у Neureclipsis bimaculata располагаются
либо недалеко от сенсорных полей, либо между изогнутыми трихоидными сенсил-
лами в сенсорных полях; их не бывает больше 18 на одном сегменте.
Специфически распределенные базиконические, коронарные и стилоконические
сенсиллы располагаются поодиночке (обычно не более 5 сенсилл на сегмент) на вен-
тролатеральной стороне флагелломеров. При этом стилоконические сенсиллы нахо-
342
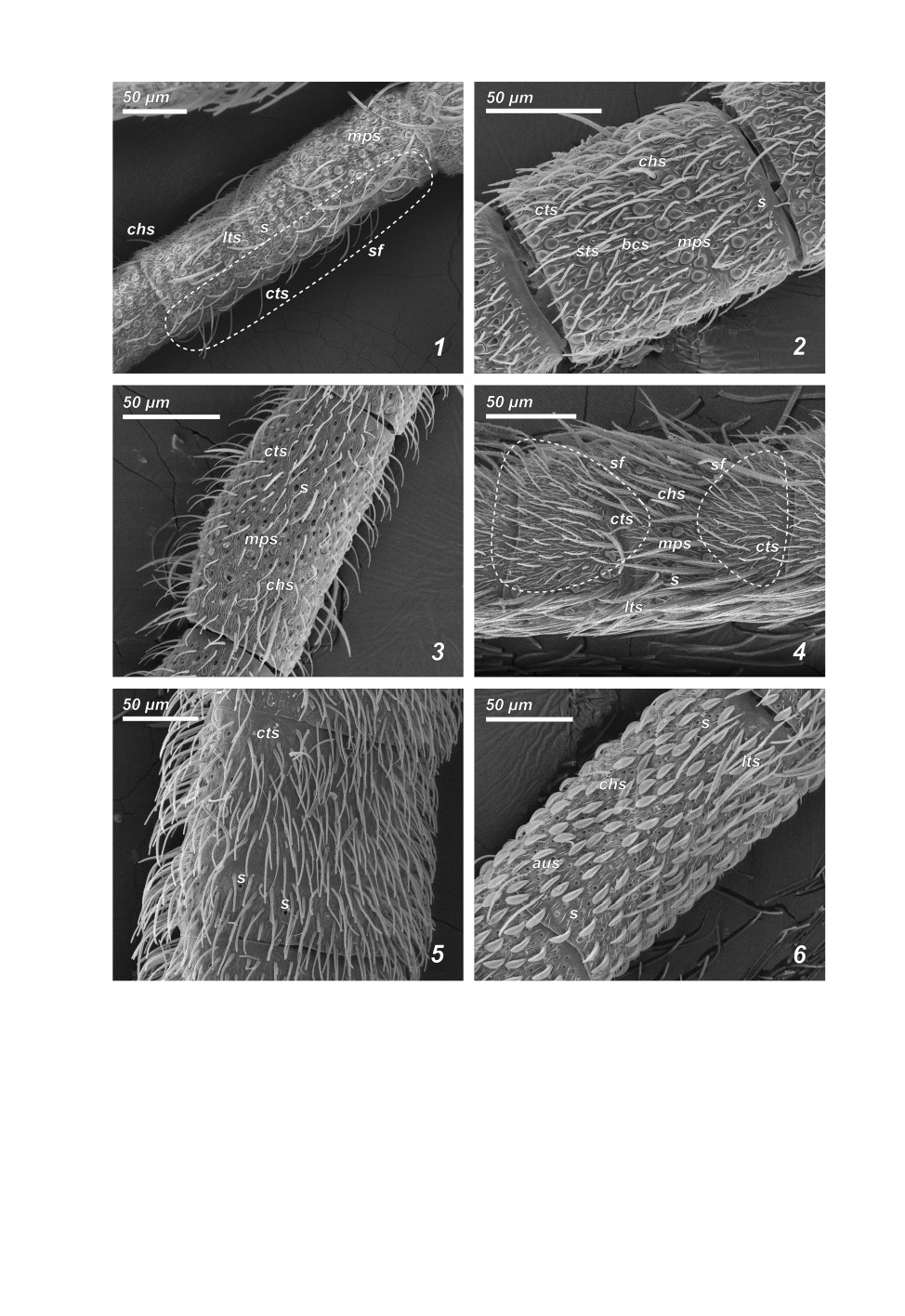
Рис. 3. Флагелломеры ручейников надсем. Psychomyioidea.
1 - 1-й флагелломер Psychomyia flavida Hagen, 1861; 2 - 4-й флагелломер Tinodes waeneri (Linnaeus, 1758);
3 - 13-й флагелломер Cyrnus fennicus Klingstedt, 1937; 4 - 6-й флагелломер Holocentropus picicornis
(Stephens, 1836); 5 - 7-й флагелломер Hyalopsyche sachalinica Martynov, 1910; 6 - 8-й флагелломер
Pseudoneureclipsis proxima Martynov, 1934.
sf - сенсорные поля. Остальные обозначения как на рис. 1.
Пунктирные линии обозначают примерные границы сенсорных полей.
343
дятся у дистального края сегмента, а базиконические и коронарные могут быть и на
дистальной части флагелломера, и на проксимальной.
С е нс о р ны е поля, образованные скоплением изогнутых трихоидных сенсилл,
обычно расположены на базальных сегментах антенны и всегда находятся на вентро-
латеральной стороне флагелломера. В промежутках между изогнутыми трихоидными
могут присутствовать другие типы сенсилл. Количество изогнутых трихоидных сен-
силл в сенсорных полях всегда максимально на базальных флагелломерах и снижа-
ется по направлению к вершине антенны (рис. 5, 1, 2, 4, 5). Наиболее дистальные сег-
менты несут одиночные изогнутые трихоидные сенсиллы, а сенсорные поля на этих
сегментах отсутствуют. У изученных видов найдено три варианта формы сенсорных
полей.
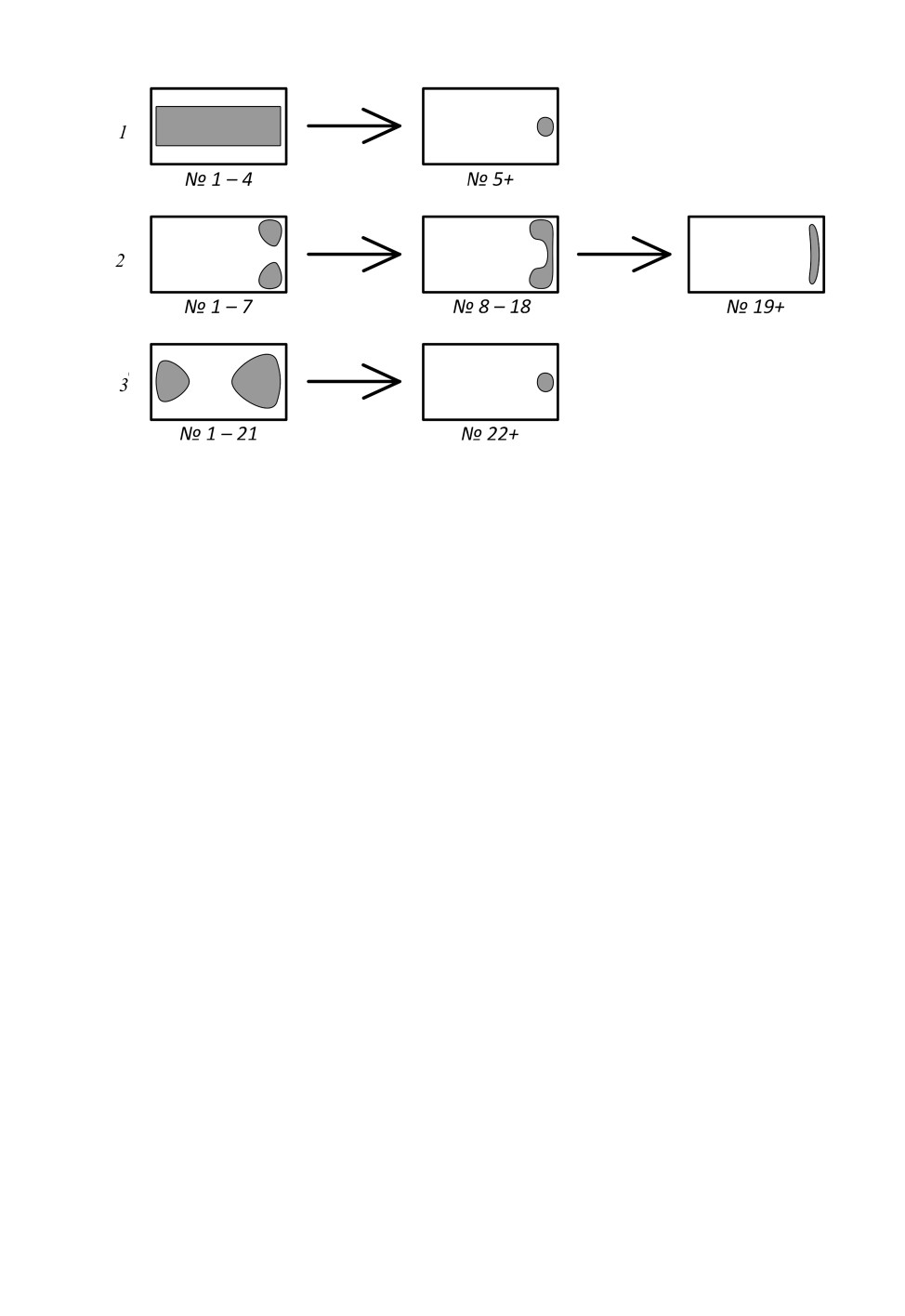
Узкие и вытянутые лентовидные сенсорные поля, тянущиеся от проксимального
края сегмента к дистальному (рис. 3, 1: sf; рис. 4, 1), найдены у большинства
Psychomyiidae (кроме Tinodes waeneri). Эти поля на базальных сегментах включают от
32 (Psychomyia minima) до 51 (Ps. flavida) изогнутых трихоидных сенсилл. К середине
антенны сенсорные поля становятся менее отчетливыми, а к вершине они сохраня-
ются только на дистальной части сегментов, приобретая при этом округлую форму
(рис. 4, 1; рис. 5, 4, 5).
Парные сенсорные поля в дистальной части сегмента (рис. 4, 2) характерны для не-
которых Polycentropodidae (Neureclipsis bimaculata, Polycentropus flavomaculatus,
Cyrnus flavidus). Базальные сегменты антенн несут два сенсорных поля, в каждом из
которых находится от 15 (P. flavomaculatus) до 50 (N. bimaculata) сенсилл. Флагелло-
меры в середине и в дистальной части антенны несут только одно поле, по-видимому,
образованное объединением двух сенсорных полей. На дистальных сегментах эти
сенсорные поля редуцированы до двух или трех изогнутых трихоидных сенсилл.
Поперечно разделенные парные сенсорные поля (рис. 3, 4: sf; рис. 4, 3) (большее по
площади дистальное и меньшее проксимальное) характерны для Holocentropus insignis
и H. picicornis (Polycentropodidae). Количество изогнутых трихоидных сенсилл в сен-
сорных полях у этих видов максимально на первых пяти флагелломерах (60 у
H. insignis и 75 у H. picicornis) и плавно убывает на более дистальных сегментах.
Проксимально расположенное сенсорное поле полностью редуцируется после 22-го
сегмента (рис. 4, 3). Субапикальные сегменты антенны имеют не более 5 изогнутых
трихоидных сенсилл, расположенных дистально.
ОБСУЖДЕНИЕ
Изученные семейства демонстрируют сопоставимое с другими эволюционными ли-
ниями Annulipalpia (Melnitsky et al., 2018; Abu Diiak et al., 2021) морфологическое раз-
нообразие сенсилл (9 типов сенсилл), уникальных типов сенсилл не найдено. Струк-
турные модификации наблюдаются в основном у псевдоплакоидных и изогнутых
трихоидных сенсилл. У отдельных видов грибовидные псевдоплакоидные сенсиллы
преобразованы в ушковидные (Cyrnus fennicus, Pseudoneureclipsis chrysippus и
Ps. proxima), Т-образные (Neureclipsis bimaculata), приобретают вытянутую
(Pahamunaya taleban и Xiphocentron mnesteus) или звездчатую (Dipseudopsis varians)
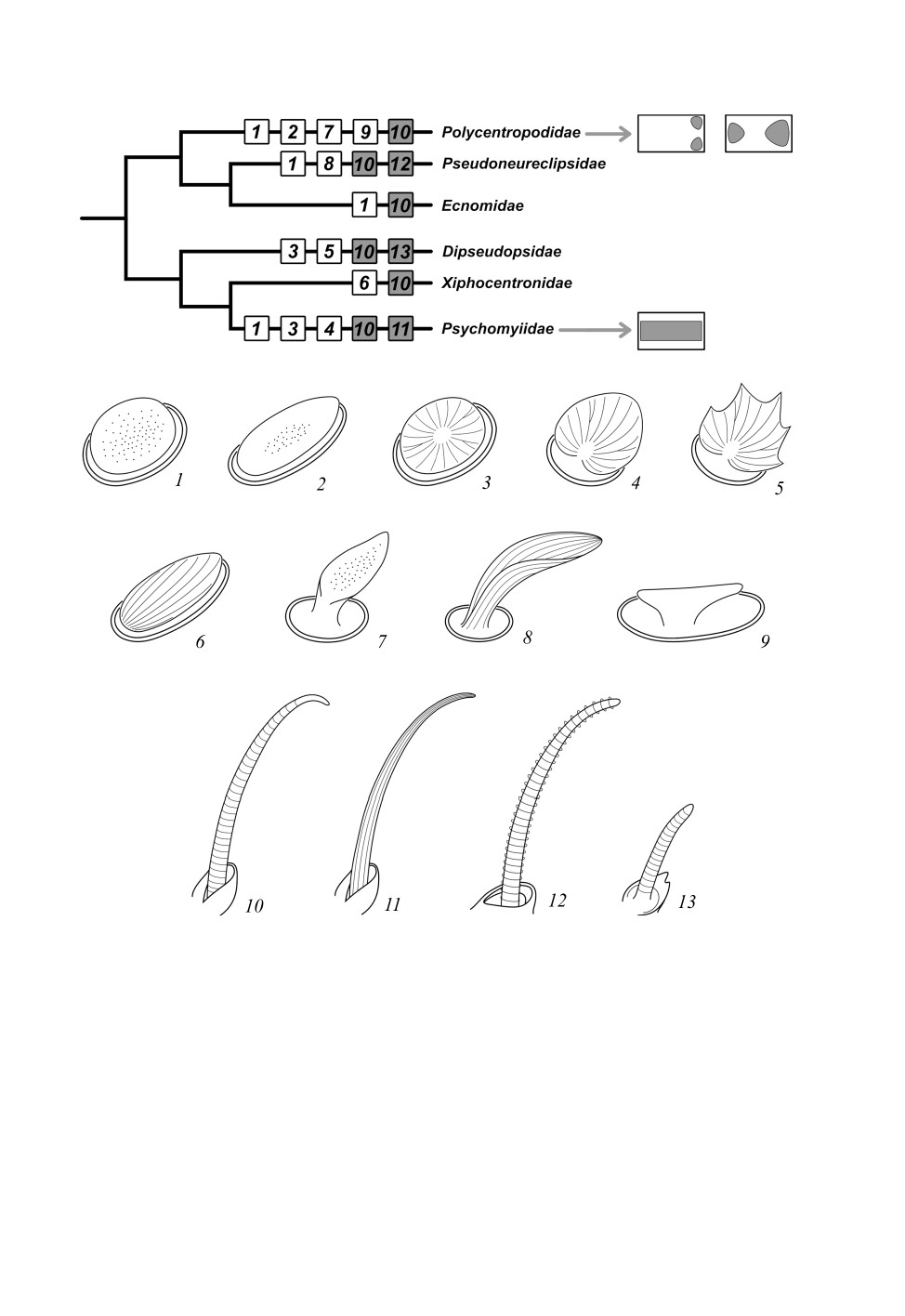
форму (рис. 7, 1-9). Возможно, вытянутые грибовидные псевдоплакоидные сенсиллы
представляют собой переходное состояние к ушковидным. На антеннах у видов, име-
344
Рис. 4. Изменение формы сенсорных полей ручейников надсем. Psychomyioidea
на протяжении антенны.
1 - Psychomyia flavida Hagen, 1861; 2 - Neureclipsis bimaculata (Linnaeus, 1758);
3 - Holocentropus insignis Martynov, 1924.
ющих ушковидные или Т-образные псевдоплакоидные сенсиллы, также присутствуют
и грибовидные псевдоплакоидные сенсиллы. Преобразование грибовидных псевдо-
плакоидных сенсилл в ушковидные и Т-образные ранее было обнаружено у предста-
вителей семейств Rhyacophilidae (Valuyskiy et al., 2017) и Hydropsychidae (подсем.
Macronematinae) (Abu Diiak et al., 2021). Изогнутые трихоидные сенсиллы могут при-
обретать разные модификации исчерченности (Lype phaeopa, Tinodes turanicus,
Pseudoneureclipsis chrysippus и Ps. proxima) или иметь два размерных подтипа на
одной антенне (Dipseudopsis indica и D. varians) (см. рис. 2, 3; рис. 7, 10-13). Для
других типов сенсилл характерно консервативное строение.
Размер изогнутых трихоидных сенсилл очень однообразен в сем. Polycentropodidae
и составляет примерно 20 μm (рис. 2, 3). Представители сем. Psychomyiidae в целом
имеют сходный размер изогнутых трихоидных сенсилл, но у Tinodes waeneri длина их
примерно в 1.5-2 раза меньше, чем у других Psychomyiidae (рис. 2, 3). Изогнутые три-
хоидные сенсиллы у H. sachalinica минимум в 2 раза длиннее, чем у других
Dipseudopsidae. Размеры грибовидных псевдоплакоидных сенсилл более или менее
однообразны у представителей изученных семейств, но вытянутые сенсиллы этого
типа (Pahamunaya taleban и Xiphocentron mnesteus) имеют ожидаемо большие раз-
меры, чем округлые сенсиллы (рис. 2, 5). Длина отростка стилоконических сенсилл
также в целом стабильна в пределах исследованных семейств, но у Dipseudopsis indica
они в 2-2.5 раза крупнее, чем у других видов (рис. 2, 6). Таким образом, размеры сен-
силл одного типа в пределах одного семейства чаще всего варьируют слабо, но у неко-
торых видов их размеры изменчивы.
Распределение различных типов сенсилл у изученных семейств соответствует
закономерностям, наблюдаемым у других семейств и, вероятно, относящихся к исход-
345
Рис. 5. Количественное распределение изогнутых трихоидных сенсилл на флагеллуме
Psychomyioidea.
1 - Polycentropus flavomaculatus (Pictet, 1834); 2 - Holocentropus insignis Martynov, 1924; 3 - Pahamunaya
taleban Malicky et Chantaramongkol, 1993; 4 - Lype phaeopa (Stephens, 1836); 5 - Psychomyia flavida
Hagen, 1861; 6 - Tinodes waeneri (Linnaeus, 1758).
Планки погрешностей - ошибки среднего.
По горизонтальной оси - номера сегментов (N), по вертикальной оси - среднее число сенсилл
на 1 сегмент (n).
ному плану строения сенсорной поверхности Trichoptera (Valuyskiy et al., 2020a):
длинные трихоидные и псевдоплакоидные сенсиллы обычно имеют неспецифическое
распределение, хетоидные - фиксированное, базиконические, коронарные и стилоко-
нические сенсиллы - специфическое. В распределении Т-образных псевдоплакоидных
сенсилл у Neureclipsis bimaculata наблюдается модификация: эти сенсиллы располо-
жены рядом с сенсорными полями (специфическое распределение). Базиконические
сенсиллы у N. bimaculata, Pahamunaya taleban и Xiphocentron mnesteus распределены
неспецифически, что, вероятно, связано с увеличением их числа.
Изогнутые трихоидные сенсиллы собраны в сенсорные поля у некоторых Poly-
centropodidae и большинства Psychomyiidae (рис. 7), при этом виды без сенсорных
полей (с неспецифическим распределением изогнутых трихоидных сенсилл или с
одиночными изогнутыми трихоидными сенсиллами) есть во всех изученных семей-
ствах. Сенсорные поля в пределах надсемейства имеют три варианта формы: узкие
лентовидные сенсорные поля, парные сенсорные поля на дистальной части сегмента,
346
Рис. 6. Количественное распределение псевдоплакоидных сенсилл на флагеллуме
Psychomyioidea.
1 - грибовидные псевдоплакоидные сенсиллы Pahamunaya taleban Malicky et Chantaramongkol, 1993;
2 - грибовидные псевдоплакоидные сенсиллы Psychomyia flavida Hagen, 1861; 3 - грибовидные
псевдоплакоидные сенсиллы Lype phaeopa (Stephens, 1836); 4 - грибовидные псевдоплакоидные сенсиллы
Xiphocentron mnesteus Schmid, 1982; 5 - грибовидные псевдоплакоидные сенсиллы Dipseudopsis indica
McLachlan, 1875; 6 - Т-образные псевдоплакоидные сенсиллы Neureclipsis bimaculata (Linnaeus, 1758).
Планки погрешностей - ошибки среднего.
По горизонтальной оси - номера сегментов (N), по вертикальной оси - среднее число сенсилл
на 1 сегмент (n).
и поперечно разделенные парные сенсорные поля. Количество изогнутых трихоидных
сенсилл в этих сенсорных полях слабо различается (от 30 у Polycentropus flavomaculatus
до 100 у Neureclipsis bimaculata). Сенсорные поля всегда расположены на вентролате-
ральной стороне сегментов. Это может быть связано с тем, что ручейники летают с
поднятыми антеннами, и потоки воздуха при полете обдувают антенны именно с вен-
тролатеральной стороны (Ivanov, Melnitsky, 2011).
Плотность расположения изогнутых трихоидных сенсилл всегда убывает по направ-
лению к вершине антенны. Возможно, это объясняется тем, что дистальные части ан-
тенны сильнее базальных подвержены инерции и действию силы тяжести, поэтому
сенсиллы, расположенные на базальных флагелломерах, могут давать более точную
информацию для ориентации в пространстве (Ivanov, Melnitsky, 2011). Другой при-
чиной убывания густоты сенсилл к концу антенны может быть затрудненный доступ
гемолимфы к дистальным сегментам и худшее снабжение питательными веществами.
Кроме того, каждая сенсилла имеет тонкую проницаемую кутикулу, через которую ис-
347
Рис. 7. Распределение типов сенсорных полей и псевдоплакоидных сенсилл на
филогенетической схеме подсем. Psychomyioidea.
1-6 - грибовидные псевдоплакоидные сенсиллы; 7, 8 - ушковидные псевдоплакоидные сенсиллы;
9 - Т-образная псевдоплакоидная сенсилла; 10 - изогнутая трихоидная сенсилла с кольчатой
исчерченностью; 11 - изогнутая трихоидная сенсилла с продольной исчерченностью;
12 - изогнутая трихоидная сенсилла с глубокой кольчатой исчерченностью;
13 - укороченная изогнутая трихоидная сенсилла.
Реконструкция филогении построена на основании данных из статей: Kjer et al., 2016; Thomas et al., 2020.
Сведения о наличии типов сенсилл у Ecnomidae по: Valuyskiy et al., 2019.
348
паряется вода; в условиях затрудненной циркуляции гемолимфы в дистальных частях
антенны растет риск иссушения, для минимизации которого желательно уменьшение
числа сенсилл.
В пределах сем. Polycentropodidae было найдено два типа парных сенсорных полей.
Парные сенсорные поля в дистальной части сегмента характерны для Neureclipsis
bimaculata, Polycentropus flavomaculatus и Cyrnus flavidus. Похожая форма сенсорных
полей встречается в разных группах Annulipalpia: у некоторых Philopotamidae (виды
рода Philopotamus) и Hydropsychidae (Arctopsychinae, некоторые Hydropsychinae -
Hydropsyche newae и Hydronema persica) (Melnitsky et al., 2018; Abu Diiak et al., 2021).
Виды рода Holocentropus имеют поперечно разделенные парные сенсорные поля, что
также характерно для Leptonema viridianum из сем. Hydropsychidae (Abu Diiak et al.,
2021). У части представителей сем. Polycentropodidae (Plectrocnemia conspersa, Cyrnus
fennicus, C. trimaculatus и Pahamunaya taleban) сенсорных полей нет, а флагелломеры
несут много неспецифически распределенных изогнутых трихоидных сенсилл. Такой
же способ распределения этих сенсилл найден у Tinodes waeneri (Psychomyiidae),
Hyalopsyche sachalinica, Dipseudopsis indica и D. varians (Dipseudopsidae), а также
у видов рода Ecnomus из сем. Ecnomidae (Valuyskiy et al., 2019).
У большинства Psychomyiidae (кроме T. waeneri) форма сенсорных полей сходна,
что может говорить о низкой скорости эволюции сенсорного оснащения поверхности
антенн в пределах этого семейства. У этих видов сенсорные поля тянутся от прокси-
мального к дистальному краю флагелломера, относительно узкие и не покрывают всю
его вентролатеральную сторону; число изогнутых трихоидных сенсилл максимально
на 1-5-м флагелломерах и может убывать постепенно либо резко на 6-10-м сегментах
флагеллума. Распределение изогнутых трихоидных сенсилл претерпевает модифи-
кацию у T. waeneri: сенсорные поля отсутствуют, а изогнутые трихоидные сенсиллы
имеют неспецифическое распределение; их количество максимально на 6-10-м фла-
гелломерах и составляет более 200 на сегмент. Такая модификация может быть свя-
зана с тем, что T. waeneri обитает в озерах, в то время как большинство Psychomyiidae
являются обитателями ручьев. Популяции обитающих в озерах видов занимают
большие площади, чем занимаемые речными видами, и особи в них разделены боль-
шими дистанциями, что осложняет феромонную коммуникацию. Вероятно, увели-
чение количества изогнутых трихоидных сенсилл на антеннах T. waeneri повышает
чувствительность к половым феромонам.
У Xiphocentron mnesteus одиночные изогнутые трихоидные сенсиллы не собраны в
сенсорные поля и расположены на вентральной стороне сегментов. Сенсорные поля
представителей семейства Psychomyiidae, таксономически близкого к Xiphocentronidae
(Holzenthal et al., 2007) расположены сходно, они покрывают вентральную сторону
флагелломеров. Вероятно, исчезновение сенсорных полей у X. mnesteus связано с ре-
дукцией числа изогнутых трихоидных сенсилл без изменения их расположения на сег-
ментах флагеллума. Случаи редукции сенсорных полей до одиночных изогнутых три-
хоидных сенсилл также известны у Pseudoneureclipsis proxima, Ps. chrysippus
(Pseudoneureclipsidae) и Chimarrhodella sp. (Philopotamidae) (Melnitsky et al., 2018).
Сенсорная поверхность антенн у видов рода Ecnomus (Ecnomidae) (Valuyskiy et al.,
2019) демонстрирует значительное сходство с другими представителями Psycho-
myioidea, исследованными в данной работе. Для ранее изученных Ecnomidae также
характерно низкое морфологическое разнообразие сенсилл на антеннах. Размеры
349
сенсилл Ecnomidae близки к размерам сенсилл у изученных видов. Распределение
сенсилл у видов рода Ecnomus сходно со многими представителями изученных
семейств: сенсорные поля отсутствуют, длинные трихоидные, грибовидные псевдо-
плакоидные и изогнутые трихоидные сенсиллы имеют неспецифическое распреде-
ление, хетоидные - фиксированное, одиночные базиконические и стилоконические
сенсиллы - специфическое. Плотность расположения сенсилл всех типов у Ecnomidae
снижается к концу антенны, как и у представителей изученных семейств. Сходство
сенсорных поверхностей антенн свидетельствует, в дополнение к другим данным,
в пользу таксономического единства Psychomyioidea.
Таким образом, в сравнении с другими ранее исследованными кольчатощупиковыми
строение сенсорной поверхности антенн в надсем. Psychomyioidea консервативно.
Лишь у отдельных видов независимо возникают модификации в структуре сенсилл,
размерах или в паттернах их распределения. Подобные уникальные отклонения най-
дены в каждом из семейств, что указывает на возможность использования признаков
строения сенсорной поверхности флагеллумов антенн при решении сравнитель-
но-морфологических задач для тасонов низкого ранга. Таксономическая значимость
структурных особенностей сенсорной поверхности антенн у семейств из надсем.
Psychomyioidea может быть показана только при более подробном сравнительном ана-
лизе с нанесением найденных признаков на филогенетические схемы с высоким раз-
решением.
ФИНАНСИРОВАНИЕ
Исследование выполнено при финансовой поддержке Российского научного
фонда (проект № 22-24-00259) в рамках проектов № 109-16530 и № 109-24431 Ресурс-
ного центра СПбГУ «Развитие молекулярных и клеточных технологий».
СПИСОК ЛИТЕРАТУРЫ
Иванов В. Д., Мельницкий С. И. 1999. Строение стернальных феромонных желез ручейников (Trichoptera).
Энтомологическое обозрение 78 (3): 505-526. [Ivanov V. D., Melnitsky S. I. 1999. Structure of the sternal
pheromone glands in caddisflies (Trichoptera). Entomological Review 79 (8): 926-942].
Иванов В. Д., Мельницкий С. И. 2014. Проблемы молекулярной эволюции феромонов ручейников и прими-
тивных чешуекрылых (Insecta: Trichoptera, Lepidoptera). Энтомологическое обозрение 93 (2): 311-327.
[Ivanov V. D., Melnitsky S. I. 2014. Questions of molecular evolution of pheromone communication in
caddisflies and lower moths (Insecta: Trichoptera, Lepidoptera). Entomological Review 94 (7): 930-942.
Валуйский М. Ю., Мельницкий С. И., Иванов В. Д. 2017. Строение антеннальных сенсилл ручейников рода
Rhyacophila Pictet (Trichoptera, Rhyacophilidae). Энтомологическое обозрение 96 (3): 401-417.
Валуйский М. Ю., Мельницкий С. И., Иванов В. Д. 2019. Сравнительный анализ поверхности антенн у има-
го ручейников рода Ecnomus McLachlan (Trichoptera, Ecnomidae). Энтомологическое обозрение 98 (2):
315-326.
Abu Diiak K. T., Valuyskiy M. Yu., Melnitsky S. I., Ivanov V. D. 2021. Diversity and distribution of antennal sensilla
in Hydropsychidae (Insecta, Trichoptera). Biological Communications 66 (4): 302-315.
Holzenthal R. W., Blahnik R. J., Prather A. L., Kjer K. M. 2007. Order Trichoptera Kirby, 1813 (Insecta), Caddisflies.
Zootaxa 1668: 639-698.
Holzenthal R. W., Morse J. C., Kjer K. M. 2011. Order Trichoptera Kirby, 1813. In: Z.-Q. Zhang (ed.). Animal
biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa 3148 (1):
209-211.
350
Ivanov V. D. 1993. Principles of the sexual communication in caddisflies (Insecta, Trichoptera). In: K. Wiese et al.
(eds). Sensory Systems of Arthropods. Basel: Birkhuser Verlag, p. 609-626.
Ivanov V. D., Melnitsky S. I. 2011. Structure and morphological types of the antennal olfactory sensilla in
Phryganeidae and Limnephilidae (Insecta: Trichoptera). Zoosymposia 5: 210-234.
Ivanov V. D., Melnitsky S. I. 2016. Diversity of the olfactory sensilla in caddisflies (Trichoptera). Zoosymposia 10:
224-233.
Ivanov V. D., Sukatsheva I. D. 2002. Trichoptera (Phryganeida). In: A. P. Rasnitsyn, L. J. Quicke (eds). History of
Insects. Dordrecht, Boston, London: Kluwer Academic Publishers, p. 199-220.
Kjer K. M., Thomas J. A., Zhou X., Frandsen P. B., Prendini E., Holzenthal R. W. 2016. Progress on the phylogeny
of caddisflies (Trichoptera). Zoosymposia 10: 248-256.
Kopylov D. S., Rasnitsyn A. P., Aristov D. S., Bashkuev A. S., Bazhenova N. V., Dmitriev V. Yu, Gorochov A. V.,
Ignatov M. S., Ivanov V. D., Khramov A. V., Legalov A. A., Lukashevich E. D., Mamontov Yu. S.,
Melnitsky S. I., Ogłaza B., Ponomarenko A. G., Prokin A. A., Ryzhkova O. V., Shmakov A. S.,
Sinitshenkova N. D., Solodovnikov A. Yu, Strelnikova O. D., Sukacheva I. D., Uliakhin A. V.,
Vasilenko D. V., Wegierek P., Yan E. V., Zmarzły M. 2020. The Khasurty Fossil Insect Lagerstätte.
Paleontological Journal 54 (11): 1221-1394.
Kubiak M., Beckmann F., Friedrich F. 2015. The adult head of the annulipalpian caddisfly Philopotamus ludificatus
Mclachlan, 1878 (Philopotamidae), mouthpart homologies, and implications on the ground plan of
Trichoptera. Arthropod Systematics and Phylogeny 73 (3): 351-384.
Melnitsky S. I., Ivanov V. D. 2011. Structure and localization of sensilla on antennae of caddisflies (Insecta:
Trichoptera). Journal of Evolutionary Biochemistry and Physiology 47 (6): 593-602.
Melnitsky S. I., Ivanov V. D. 2016. Structure of the antennal sensilla in the genera Molanna and Molannodes
(Trichoptera, Molannidae). Zoosymposia 10 (1): 292-300.
Melnitsky S. I., Ivanov V. D., Valuyskiy M. Yu., Zueva L.V., Zhukovskaya M. I. 2018. Comparison of sensory
structures on the antenna of different species of Philopotamidae (Insecta: Trichoptera). Arthropod Structure
and Development 47: 45-55.
database/trichopt/index.htm] (дата обращения: 10.01.2022).
Thomas J. A., Frandsen P. B., Prendini E., Zhou X., Holzenthal R. W. 2020. A multigene phylogeny and timeline for
Trichoptera (Insecta). Systematic Entomology 45 (3): 670-686.
Valuyskiy M. Yu., Melnitsky S. I., Ivanov V. D. 2017. Structure of antennal sensilla in the caddisfly genus
Rhyacophila Pictet (Trichoptera, Rhyacophilidae). Entomological Review 97 (6): 703-722.
Valuyskiy M. Yu., Melnitsky S. I., Ivanov V. D. 2019. Comparative analysis of antennal surfaces in adult caddisflies
of the genus Ecnomus McLachlan (Trichoptera, Ecnomidae). Entomological Review 99 (3): 302-309.
Valuyskiy M. Yu., Melnitsky S. I., Ivanov V. D. 2020a. Diversity and distribution of antennal sensilla in
Glossosomatidae in comparison with other basal families of caddisflies (Insecta: Trichoptera). Zoosymposia
18: 179-190.
Valuyskiy M. Yu., Melnitsky S. I., Ivanov V. D. 2020b. Structure and evolution of antennal sensory surface in
endemic caddisfly tribes Baicalinini and Thamastini (Trichoptera: Apataniidae) from Lake Baikal. Journal of
Evolutionary Biochemistry and Physiology 56: 318-332.
Wells A. 1984. Comparative studies of antennal features of adult Hydroptilidae (Trichoptera). In: J. C. Morse (ed.).
Proceedings of the 4th International Symposium on Trichoptera, 11-16 July 1983, Clemson (Series
Entomologica: 30). The Hague, The Netherlands: Dr. W. Junk Publishers, p. 423-440.
351
STRUCTURE, DIVERSITY, AND DISTRIBUTION OF THE ANTENNAL
SENSILLA IN THE CADDISFLY SUPERFAMILY PSYCHOMYIOIDEA
(TRICHOPTERA, ANNULIPALPIA)
K. T. Abu Diiak, M. Yu. Valuyskiy, S. I. Melnitsky, V. D. Ivanov
Key words: sensilla, sensory fields, antennae, Trichoptera, caddisflies, Annulipalpia,
Psychomyioidea, evolution.
SUMMARY
Comparative morphological study of structure and localization of antennal sensilla in the caddisfly
superfamily Psychomyioidea was carried out for the first time. The morphology of antennal
sensilla in 21 species from the families Psychomyiidae (6), Xiphocentronidae (1), Dipseudopsidae
(3), Polycentropodidae (9) and Pseudoneureclipsidae (2) was studied by using scanning electron
microscopy. Seven types of sensilla were revealed on the antennae of studied species: the long trichoid,
curved trichoid, chaetoid, basiconic, coronary, styloconic and pseudoplacoid sensilla. The distribution
of different types of sensilla in the studied families mainly corresponds to the ground plan of antennal
sensory surface structure in Trichoptera. Long trichoid and pseudoplacoid sensilla usually have non-
specific distribution, chaetoid - fixed, basiconic, coronary and styloconic sensilla - specific distribution.
Sensory fields of curved trichoid sensilla are located on basal antennal segments and are always positioned
on ventrolateral surfaces of flagellomeres. Three types of the sensory fields shape were revealed in the
studied species. Antennal sensory surface in representatives of superfamily Psychomyioidea mainly
has conservative structure, but particular evolutionary lineages show modifications in structure and
localization of sensilla.
352