ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 101, 2, 2022
УДК 595.722
НОВЫЕ ДАННЫЕ О ТАРАКАНАХ РОДОВ PLACOBLATTA
BEY-BIENKO, 1969 И ANISOLAMPRA BEY-BIENKO, 1969
(DICTYOPTERA, BLABERIDAE: EPILAMPRINAE)
© 2022 г. Л. Н. Анисюткин
Зоологический институт РАН
Университетская наб., 1, С.-Петербург, 199034 Россия
e-mail: Leonid.Anisyutkin@zin.ru, leonid.dictyoptera@gmail.com
Поступила в редакцию 19 марта 2022 г.
После доработки 23 марта 2022 г.
Принята к публикации 23 марта 2022 г.
Работа посвящена систематике и распространению короткокрылых тараканов родов Placoblatta
и Anisolampra. Описаны новый вид Placoblatta kabakovi sp. n. из Вьетнама и ранее неизвестная
самка P. minor. Приводятся дополнительные данные о морфологии и географическом распро-
странении P. rugosa и A. panfilovi, в том числе впервые описывается строение яйцеклада и при-
лежащих структур. Предлагается номенклатура модификаций строения надкрылий и крыльев
тараканов.
Ключевые слова: тараканы, морфология, новый вид, f. macroptera, f. brachyptera, f. microptera,
f. aptera.
DOI: 10.31857/S0367144522020150, EDN: HLOPNZ
Различные варианты укорочения или исчезновение надкрылий и крыльев широко
распространены и встречаются во всех семействах тараканов, возникая параллельно и
независимо. Не является исключением и подсем. Epilamprinae сем. Blaberidae. В этой
статье приводятся новые данные о представителях морфологически сходных и, воз-
можно, филогенетически близких родов Placoblatta Bey-Bienko, 1969 и Anisolampra
Bey-Bienko, 1969, описывается новый вид рода Placoblatta, а также общие замечания
об особенностях морфологии тараканов с укороченными надкрыльями и крыльями.
МАТЕРИАЛ И МЕТОДИКА
Изученный материал был как сухим, наколотым на булавки, так и фиксированным в 70%-ном
этаноле. Автор следует ранее описанной методике работы с материалом (Анисюткин, 2012).
Номенклатура вооружения передненижнего края передних бедер описывается по схеме
Г. Я. Бей-Биенко (1950) и Л. Рота (Roth, 2003a), склеритов гениталий самцов - по К.-Д. Классу
(Klass, 1997). Номенклатура структур гениталий самок дается по Ф. А. МакКиттрик (McKittrick,
1964) и К.-Д. Классу (Klass, 1998).
390
Весь изученный материал, включая голотип нового вида, хранится в коллекции Зоологическо-
го института РАН в С.-Петербурге (ЗИН).
Варианты строения надкрылий и крыльев тараканов
Степень развития и строение органов полета у тараканов могут существенно варьировать - от
полностью развитых надкрылий и крыльев до их полного отсутствия и от тонких мембранозных
крыловых пластинок до жестких элитр с практически полностью утраченным жилкованием. Эти
органы имеют большое биологическое и таксономическое значение, поэтому необходима их не-
противоречивая номенклатура с четко определенными терминами. Отсутствие строгой термино-
логии затрудняет сравнение, биологическую и филогенетическую интерпретацию признаков.
В настоящее время в литературе о тараканах используются не имеющие строгого определения
термины, например, «боковые чешуйки» (Бей-Биенко, 1950) или «macropterous», «brachypterous»,
«apterous», «reduced tegminal pads» (Roth, 2003a). Даже авторы, пытавшиеся использовать при-
знаки развития органов полета для построения классификации (Chopard, 1925; Rehn, 1932), ис-
пользовали описательные термины, не давая им определений, например, «élytres et ailes très
courts» (Chopard, 1925) или «tegmina and wings very short» (Rehn, 1932).
Предложенная ниже номенклатура наиболее близка к использованной Л. Ротом для Panesthiinae
(Roth, 1977), отличаясь большей строгостью определений, и к номенклатуре С. Ю. Стороженко
и Дж.-Ч. Пайка (Storozhenko, Paik, 2007) для прямокрылых.
Forma macroptera - полнокрылая форма (= euptera): надкрылья и крылья полностью развитые,
лётные, по длине превышают брюшко, их задние края перекрываются;
forma brachyptera - короткокрылая форма: надкрылья или крылья укорочены, равны по длине
брюшку или короче его, перекрываются или соприкасаются по заднему краю;
forma microptera - микроптерная форма [= крыловые чешуйки (wing-scales)]: надкрылья или
крылья сильно укорочены, значительно короче брюшка, не соприкасаются по заднему краю,
обычно сильно склеротизованы, в значительной степени утрачивают жилкование;
forma aptera - бескрылая форма: надкрылья или крылья отсутствуют.
Неравномерность укорочения надкрылий и крыльев часто встречается у тараканов, надкры-
лья, как правило, более развиты, поэтому целесообразно выделять форму по максимально разви-
той структуре, т. е. в случае укороченных надкрылий и отсутствия крыльев форму следует
называть короткокрылой (f. brachyptera).
Строение надкрылий и крыльев обычно стабильно в пределах вида, исключения редки: измен-
чивость от f. euptera до f. brachyptera известна, например, у Hebardina concinna (Blattidae) (Yue
et al., 2014) и некоторых Panesthiinae (Blaberidae) (Roth, 1977).
Укорочение органов полета часто подвержено половому диморфизму; как правило, у самцов
они более развиты, что может объясняться их большей подвижностью, которая связана с поис-
ком самок.
По степени склеротизации надкрылий можно выделить три основные группы.
1. Мембранозные надкрылья - крыловая пластинка тонкая и прозрачная, жилкование не
уплотненное. Редко встречающийся вариант, известен у Cardacopsis и Nocticola (Nocticolidae)
(Karny, 1924; Roth, 2003b); надкрылья частично мембранозные у Holocompsa (Corydiidae) (Qiu
et al., 2019, 2020).
2. Кожистые надкрылья (tegmina) - крыловая пластинка утолщенная, умеренно склеротизо-
ванная, почти непрозрачная, жилкование обычно уплотненное. Наиболее часто встречающийся
вариант «тараканьих» надкрылий.
391
3. Элитры (elytra) - крыловая пластинка жесткая и непрозрачная, жилкование обычно не раз-
личимо при осмотре сверху. Спорадически встречающийся в разных таксонах вариант, напри-
мер, у Euthyrrhapha (Corydiidae) (Anisyutkin, 2008) и Diploptera (Blaberidae) (Li et al., 2017).
Крылья практически всегда менее склеротизованы, чем надкрылья.
Различные варианты склеротизации надкрылий могут свободно комбинироваться с варианта-
ми их укорочения.
РЕЗУЛЬТАТЫ
Подсем. EPILAMPRINAE Brunner von Wattenwyl, 1865
Род PLACOBLATTA Bey-Bienko, 1969
Ти по во й в ид: Placoblatta rugosa Bey-Bienko, 1969, по монотипии.
Род был описан как монотипичный из Северного Вьетнама (Бей-Биенко, 1969).
В дальнейшем представители рода отмечались из южного Вьетнама (Анисюткин,
2018), Шри-Ланки (Anisyutkin, Yushkova, 2017) и с о. Сулавеси (Anisyutkin, 2020).
Описанный с о. Сулавеси Placoblatta semialata Anisyutkin, 2020 занимает обособ-
ленное положение в роде: его самцы принадлежат к f. brachyptera, тогда как у
остальных видов рода они принадлежат к f. microptera или f. aptera.
С о ст а в. Placoblatta rugosa, P. minor Anisyutkin, 2018, P. beybienkoi Anisyutkin, 2017
(в: Anisyutkin, Yushkova, 2017), P. semialata и P. kabakovi sp. n.
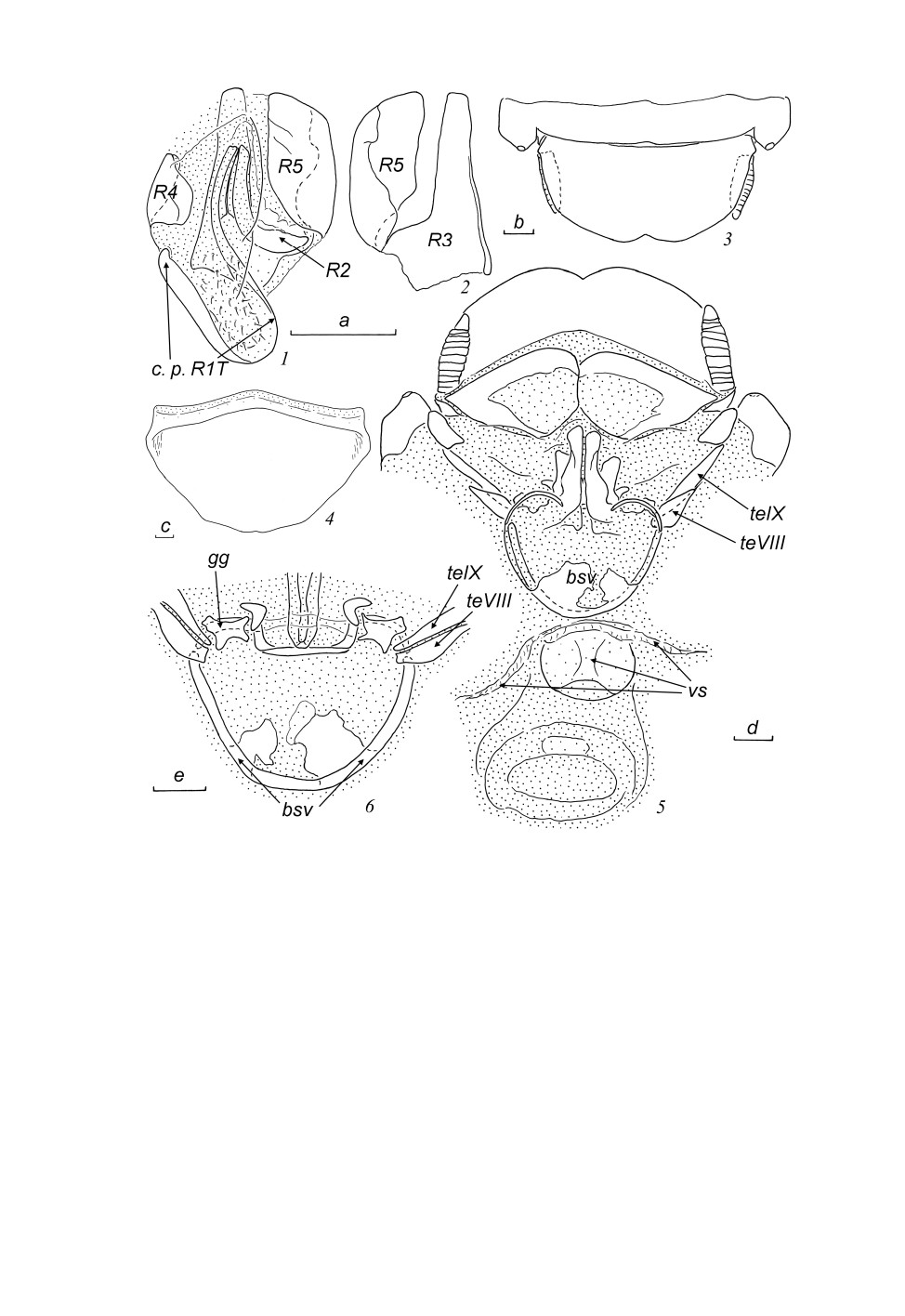
Placoblatta rugosa Bey-Bienko, 1969 (рис. 1, 1-6).
Мат ер иа л. Типовая серия. Вьетнам. «Д. Р. Вьетнам Шон-Зунг О. Кабаков 19-28.II.962»,
1 ♀ (голотип), 1 личинка (паратип); «Д. Р. Вьетнам Там-Дао О. Кабаков 2.XI.962», 1 ♀ (паратип);
горы Там-Дао, 24-26.X.1961 (О. Н. Кабаков), 1 личинка (паратип); Там-Дао у Дао-чу,
16-18.XI.1961 (О. Н. Кабаков), 1 личинка (паратип); 50 км NO Тай-Нгуен, h = 300-400 м,
8.II.1963 (О. Н. Кабаков), 1 личинка (паратип); те же данные, но h = 300 м, 10.V.1963 (О. Н. Ка-
баков), 1 личинка (паратип).
Вьетнам. Провинция Виньфу, Тамдао (Tam Dao), h = 800-900 м, 17-31.V.1995 (А. В. Горохов),
3 ♂, 3 ♀. Провинция Туйен Куанг (Tuyen Quang), заповедник Na Hang, h = 1000 m, VI.2013
(Н. Л. Орлов), 2 ♂, 2 ♀.
Вид был описан по самкам из северного Вьетнама (хр. Тамдао) (Бей-Биенко, 1969),
позднее из той же или близкой точки (национальный парк Тамдао) были описаны
самцы (Анисюткин, 1999). В 2013 г. представители вида были собраны в провинции
Туйен Кванг, граничащей с провинцией Виньфу. Национальный парк Тамдао заходит
в провинцию Туйен Кванг, но заповедник На Ханг расположен на карстовом массиве,
не связанном с хребтом Тамдао (Н. Л. Орлов, личное сообщение).
Описания вида (Бей-Биенко, 1969; Анисюткин, 1999) могут быть дополнены следу-
ющими данными.
С ам ец (экземпляр из Тамдао). Правая фалломера (склериты R + N) (рис. 1, 1, 2) с каудальной
частью склерита R1T удлиненно-прямоугольной формы (рис. 1, 1, c. p. R1T), густо покрытой
крупными щетинками, краниальная часть R1T удлиненная и слабо изогнутая; склерит R2 мас-
сивный, равномерно изогнутый; краниальные части R1T и R2 оттянуты в слабо склеротизован-
ный вырост; R3 широкий, но слабо склеротизованный; R4 в виде широкой пластинки, изогнутой
392
Рис. 1. Placoblatta rugosa Bey-Bienko, 1969, самец (1, 2) и самка (3-6) из Тамдао.
1 - правая фалломера сверху; 2 - склериты R3 и R5 правой фалломеры снизу; 3 - вершина брюшка сверху;
4 - генитальная пластинка снизу; 5 - вершина брюшка снизу, генитальная пластинка удалена;
6 - основание яйцеклада сверху. Пунктировкой показаны мембранозные участки; мембранозные участки не
показаны на створках яйцеклада. Обозначения морфологических структур здесь и далее см. в тексте.
Масштабная линейка - 1 мм: a - к рис. 1, 2; b - к рис. 3; c - к рис. 4; d - к рис. 5; e - к рис. 6.
и частично заходящей на вентральную сторону фалломеры; R5 большой и хорошо склеротизо-
ванный, соединен со склеритом R3.
Са мка (экземпляр из Тамдао). Вершина брюшка (рис. 1, 3-6).
Анальная пластинка (последний, X тергит) широкая (рис. 1, 3), каудально закругленная, с не-
глубоким медиальным вырезом. Церки короткие, уплощенные, с плотно соединенными сегмен-
тами (рис. 1, 3, 5). Парапрокты хорошо склеротизованные (рис. 1, 5), симметричные, без
393
вооружения, но со срединным мембранозным участком. Генитальная пластинка (рис. 1, 4) широ-
кая, со слабым вырезом по заднему краю.
Яйцеклад и прилежащие структуры (рис. 1, 5, 6). Тергальные выросты VIII тергита брюшка не
достигают его паратергитов, короткие и широкие (рис. 1, 5, 6, teVIII); тергальные выросты IX
тергита полностью развиты (рис. 1, 5, 6, teIX). Гонангулюмы крупные и хорошо склеротизован-
ные, их вентральная часть в виде тонкой дугообразной пластинки (рис. 1, 6, gg). Створки яй-
цеклада частично мембранозные; первая пара самая крупная, с рядами щетинок по внутреннему
краю; вторая пара удлиненная, полностью скрыта под первой; створки третьей пары (гоноплаки)
расширены и укорочены; основания второй и третьей пар как на рис. 1, 6. Базивальвула цельная,
широкая и округленная, слегка асимметричная (рис. 1, 5, 6, bsv). Вестибулярная структура слабо
склеротизована, с боковыми ветвями и срединной папиллой (рис. 1, 5, vs). Выводковая сумка
без склеротизованных структур, только с небольшим уплотненным участком, прилежащим к
папилле.
Размеры - см. табл. 1.
За меча ние. В первоначальном описании P. rugosa типовая местность указана как
«Шонзуонг, хребет Тамдао, 28 II 1962» (Бей-Биенко, 1969, с. 843).
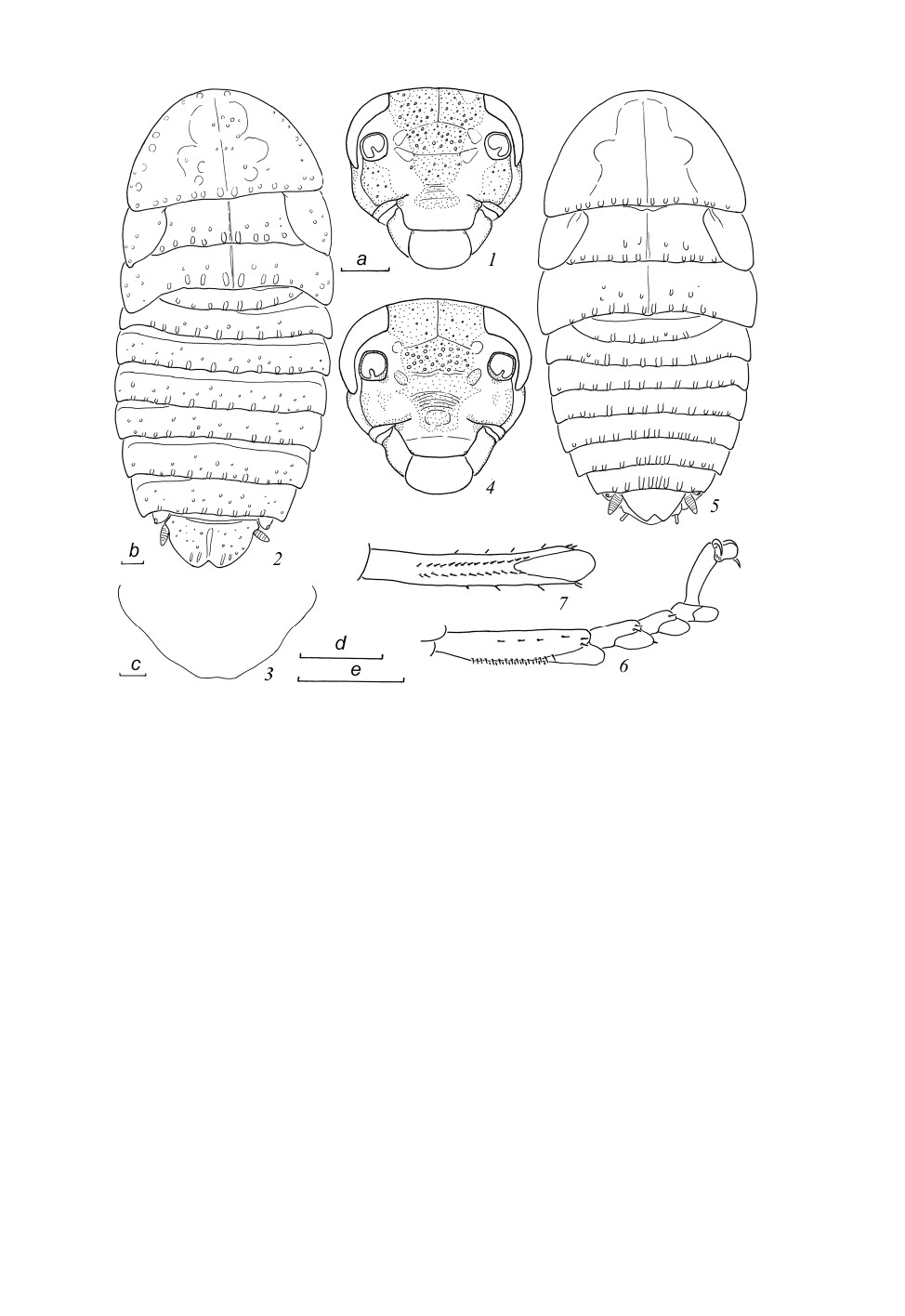
Placoblatta minor Anisyutkin, 2018 (рис. 2, 1-3).
Мат ер иа л. Вьетнам. Провинция Ламдонг (Lam Dong), р-он Lac Duong, заповедник Би
Дуп - Нуй Ба (Bi Dup - Nui Ba Nat. Res.), гора Бидуп (Bidoup), 12°06ʹ33.1ʺ N, 108°39ʹ 30.8ʺ E,
h = 1660-2010 м, 16.VI.2018 (Л. Н. Анисюткин), экспедиция российско-вьетнамского Тропиче-
ского Центра, 1 ♀.
С ам ка (nov.). Сходна с самцом (см. Анисюткин, 2018), но более крупная. Окраска как у сам-
ца, желто-коричневая, местами с более темными, почти черными пятнами; лицевая часть головы
контрастно окрашена (рис. 2, 1). Покровы блестящие, с многочисленными мелкими бугорками
(рис. 2, 2). Голова немного длиннее своей ширины (рис. 2, 1); промеры структур головы - см.
табл. 1; отношение длин 3 апикальных члеников максиллярных щупиков (от 3-го к 5-му) пример-
но 1.3 : 1.0 : 1.2; последний членик максиллярных щупиков не увеличен. Надкрылья в виде бо-
ковых чешуек, крылья отсутствуют (f. microptera - рис. 2, 2). Передненижние края передних
бедер вооружены по типу B, с 6 шипами, включая 1 вершинный. Строение задних лапок сходно
с таковым самцов. Вершина брюшка как на рис. 2, 2. Генитальная пластинка поперечная, по зад-
нему краю слабо вырезана (рис. 2, 3).
Размеры - см. табл. 1.
Placoblatta kabakovi Anisyutkin, sp. n. (рис. 2, 4-7; 3, 1-10).
urn:lsid:zoobank.org:act:0F4861F9-D37B-4781-ACEE-1981C67DF095
Мат ер иа л. Вьетнам. Провинция Зялай (Gia Lai), 38 км С г. K’Bang, национальный парк
Контюранг (Kon Chu Rang), 14°29ʹ14.4ʺ N, 108°34ʹ16.7ʺ E, h = 1000 m, 27.III-6.IV.2018 (А. В. Абра-
мов) (экспедиция российско-вьетнамского Тропического Центра), 2 ♂ (голотип и паратип). Про-
винция Фуйен (Phu Yen), район Сонг Хинь (Song Hinh), 20 км СВ Хай Риенг (Hai Rieng),
12°49ʹ37.7ʺ N, 108°59ʹ58.3ʺ E, h = 200 м, 10-15.VI.2021 (А. В. Абрамов), экспедиция российско-
вьетнамского Тропического Центра, 1 ♂ (паратип).
С ам ец (голотип). Общая окраска коричнево-черная, местами желтая; тергиты груди и
брюшка, за исключением VIII, IX и анальной пластинки, коричневые, с тонким узором из мел-
ких желтых пятнышек; боковые части пронотума, заднегруди и надкрылья желтые, густо покры-
ты коричневыми пятнышками; VIII, IX тергиты брюшка, анальная пластинка и церки
светло-желтые, с коричневыми пятнышками; лицевая часть головы с большим черным пятном,
боковые части - желтые, с черными пятнами (рис. 2, 4); глаза черные; антенны, за исключением
394
Таблица 1. Размеры (мм) и пропорции частей тела Placoblatta spp. и Anisolampra panfilovi
Bey-Bienko
1
2
3
4
5
6
7
8
Placoblatta rugosa
6.5-7.5
6.4-6.8
1.0-1.2
1.8-2.0
10.8-11.9
16.8-17.9
6.8-8.1
4.0-4.5
Bey-Bienko, ♀,
(7.2)
(6.5)
(1.1)
(1.8)
(11.4)
(17.6)
(8.1)
(4.5)
Tam Dao
P. rugosa, ♂,
6.0-6.5
5.5-5.8
1.0-1.1
1.7-1.8
8.9-10.0
13.8-15.5
5.2-6.2
3.4-3.9
Tam Dao
P. rugosa, ♂, Tuyen
5.3-5.9
4.9-5.3
1.1-1.2
1.5
8.0-8.5
13.1-13.5
5.2-5.4
3.6-3.7
Quang
P. rugosa, ♀, Tuyen
6.6-6.8
6.1-6.2
1.2-1.3
1.8-1.9
9.8-10.0
15.6-16.2
6.0-6.5
4.0
Quang
P. minor
3.7
3.5
1.2
2.5
5.6
8.5
3.3
2.2
Anisyutkin, ♀
P. tuberculata sp. n.,
4.0-4.5
3.8-4.6
0.9-1.0
1.7-1.8
5.9-6.5
9.6-10.6
3.5-4.6
2.4-2.5
♂, Gia Lai
(4.0)
(3.8)
(1.0)
(1.8)
(5.9)
(9.6)
(3.5)
(2.5)
P. tuberculata sp. n.,
4.3
3.9
1.1
1.6
6.6
10.0
4.6
2.6
♂, Phu Yen
Anisolampra
6.2-6.3
5.4
(5.4)
0.8-0.9
1.6
(1.6)
9.3-9.4
12.7-12.8
9.1
(9.1)
5.8-6.0
panfilovi, ♀,
(6.3)
(0.9)
(9.4)
(12.8)
(6.0)
Юньнань
A. panfilovi, ♂,
5.2
4.7
0.7
1.4
9.2
11.9
<34*
10.0
Юньнань
A. panfilovi, ♂,
6.4
5.4
0.7
1.4
10.9
14.8
37.0
<10.5**
Вьетнам
A. panfilovi, ♂,
6.0
5.2
0.6
1.2
10.3
13.8
38.6
11.8
Лаос
A. panfilovi, ♀,
6.6-7.4
5.6-6.3
0.8-0.9
1.4-1.7
10.4-11.4
14.9-16.0
9.5-10.9
6.5-7.7
Лаос
П р и м е ч а н и я. 1 - длина головы, 2 - ширина головы, 3 - отношение расстояния между глазами к длине
глаза, 4 - отношение расстояния между антеннальными впадинами к длине скапуса, 5 - длина пронотума,
6 - ширина пронотума, 7 - длина надкрылья, 8 - ширина надкрылья. В скобках указаны параметры голотипа.
*Вершины надкрылий обломаны.
**У сухого экземпляра надкрылья деформированы.
частично затемненного скапуса, антеклипеус, верхняя губа, лабиальные и максиллярные щупи-
ки белесые; тазики, за исключением темно-желтых дистальных участков, черные, вертлуги,
бедра и голени грязно-желтые, бедра и голени с продольными темными (коричневыми) полоса-
ми; лапки светло-желтые, почти белесые; плейриты груди по большей части черные, стерниты
брюшка черные; гипандрий светло-желтый, с коричневыми пятнышками.
Покровы гладкие и блестящие; тергиты груди, брюшка и надкрылья покрыты мелкими бугор-
ками, более крупные бугорки располагаются рядами вдоль задних краев тергитов груди и брюш-
ка (рис. 2, 5); пунктировка не выражена, лицевая часть головы со слабой поперечной
морщинистостью (рис. 2, 4).
395
Рис. 2. Placoblatta minor Anisyutkin, 2018, самка (1-3) и P. kabakovi sp. n., самец, голотип (4-7).
1, 4 - лицевая часть головы; 2, 5 - общий вид сверху; 3 - генитальная пластинка, очертания заднего края;
6 - левая задняя лапка снизу (спереди); 7 - метатарзус (1-й членик левой задней лапки сзади (снизу).
Пунктировкой показаны участки с темной окраской.
Масштабная линейка - 1 мм: a - к рис. 1, 4; b - к рис. 2, 5; c - к рис. 3; d - к рис. 6; e - к рис. 7.
Голова широкая, примерно равной длины и ширины (рис. 2, 4); эпикраниальные швы слабые;
глазковые пятна хорошо заметны; размеры и пропорции структур головы - см. табл. 1; отноше-
ние длин 3 апикальных члеников максиллярных щупиков (от 3-го к 5-му) примерно 1.2 : 1.0 : 1.6;
последний членик максиллярных щупиков увеличен.
Пронотум колоколовидный, с широко округленными передним и боковыми краями, почти
прямым задним и слабо каудально оттянутыми заднебоковыми краями (рис. 2, 5). Про-, мезо- и
метанотум со слабыми продольными швами (рис. 2, 5). Надкрылья в виде боковых чешуек
(f. microptera), крылья отсутствуют (рис. 2, 5).
Передненижние края передних бедер вооружены по типу B, с 3-5 шипами, включая 1 вершин-
ный. Передние голени не утолщены; вооружение голеней и бедер хорошо развито. Строение за-
дних лапок (рис. 2, 6, 7): метатарзус немного короче остальных члеников вместе взятых;
метатарзальная эуплантула по длине составляет примерно треть длины метатарзуса; два при-
мерно равных ряда шипиков располагаются по нижнему краю метатарзуса; эуплантулы 1-3-го
396
члеников с одним или двумя дополнительными шипиками с внешней и внутренней сторон; аро-
лиум крупный, коготки простые и симметричные.
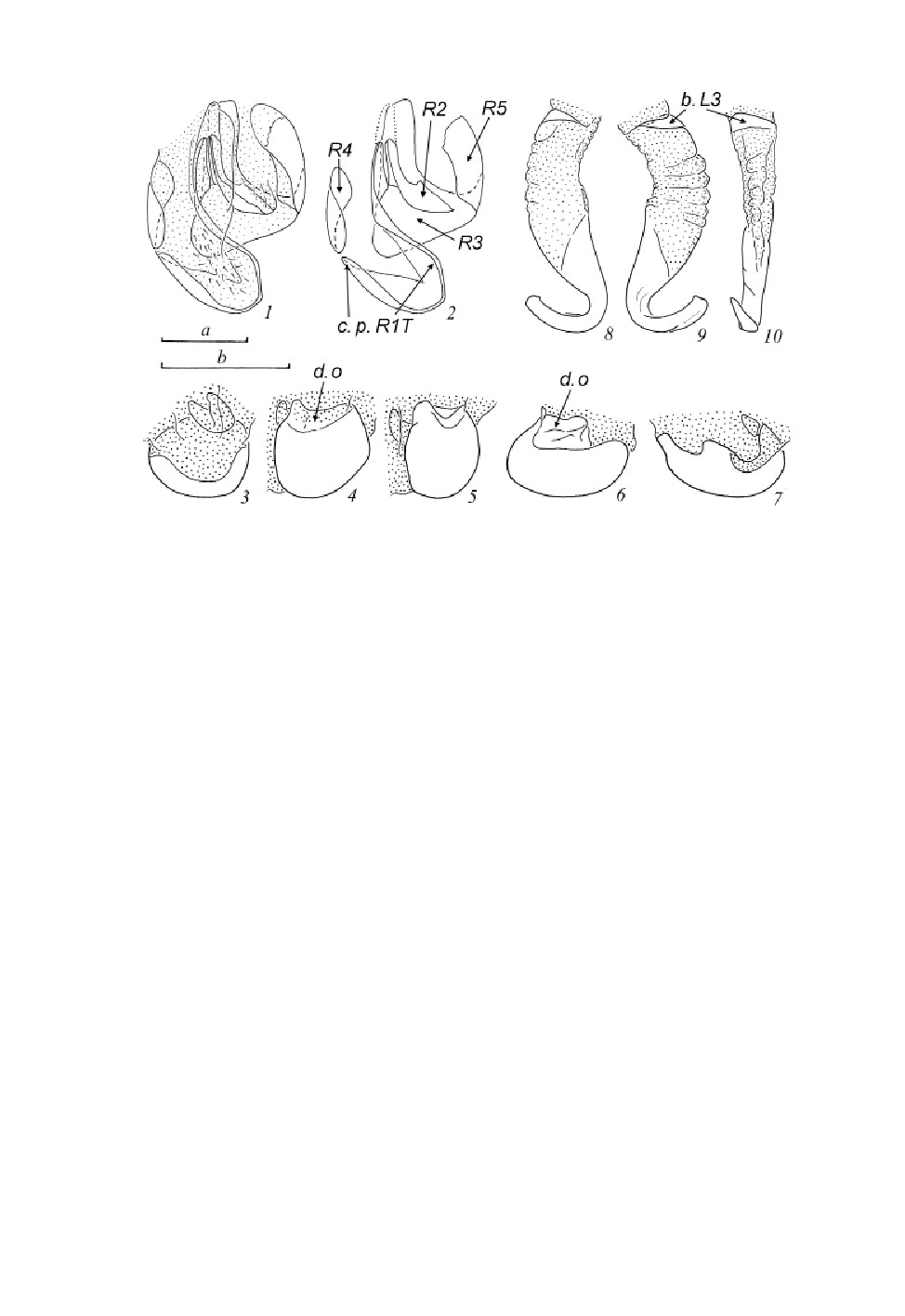
Тергиты брюшка без видимых железистых структур (рис. 2, 5); заднебоковые углы почти не
оттянуты; VIII тергит с развитыми боковыми выростами, несущими дыхальца (рис. 3, 1). Аналь-
ная пластинка (X, последний тергит брюшка) почти треугольная, с широким вырезом по заднему
краю (рис. 3, 1); церки короткие и уплощенные, с плотно соединенными члениками. Парапрокты
блаберидного типа (рис. 3, 2), склериты Pv тонкие и удлиненные. Гипандрий почти симметрич-
ный, округленный по заднему краю; стилусы цилиндрические, почти симметричные (рис. 3, 3).
Гениталии самца (рис. 3, 4-10). Правая фалломера (склериты R + N) (рис. 3, 4): каудальная
часть склерита R1T примерно трапециевидная, выпуклая по каудальному краю (рис. 3, 4,
c. p. R1T), густо покрыта крупными щетинками, краниальная часть R1T удлиненная и изогнутая;
склерит R2 массивный, равномерно изогнутый; краниальные части R1T и R2 оттянуты в слабо
склеротизованный вырост; R3 широкий, слабо склеротизованный; R4 в виде изогнутой пластин-
ки; R5 небольшой, соединен со склеритом R3. Склерит L2D (L1) разделен на апикальный и ба-
зальный субсклериты (рис. 3, 5); апикальный склерит шапочковидный, густо покрыт прижатыми
щетинками (рис. 3, 6, 7); дорсальный вырост хорошо развит (рис. 3, 5-7, d. o). Склерит L3 (L2d)
относительно короткий (рис. 3, 8, 9), базальный субсклерит присутствует (рис. 3, 9, b. L3); склад-
чатая структура слабо выражена, без щетинок; вершина L3 без гребней, с хорошо выраженным
апикальным зубчиком; борозда hge отсутствует (рис. 3, 9, 10). Склерит L4U (L3d) удлиненный
(рис. 3, 8).
В ар и аци и. Паратип из Контюранга крупнее и темнее, лицевая часть головы
большей частью черная, желтые только участки вокруг глазковых пятен и ротовые ор-
ганы; передненижние края передних бедер с 7 шипами, включая 2 вершинных; мета-
тарзальная эуплантула короче, длиной примерно в четверть длины метатарзуса.
С а мка неизвестна.
Размеры - см. табл. 1.
С р ав не ние. Новый вид наиболее сходен с P. rugosa, но хорошо отличается от
него меньшими размерами (см. табл. 1), деталями окраски, а именно светло-желтыми,
за исключением тазиков, ногами с продольными коричневыми полосками (более или
менее однотонно кофейно-коричневые у P. rugosa) и почти черными тазиками и стер-
нитами брюшка (кофейно-коричневые у P. rugosa), слабой скульптурированностью
покровов лицевой части головы, пронотума, тергитов груди и брюшка, а также формой
склеритов L2D и L3 гениталий самцов (ср. рис. 3, 5-10 и рис. 65, 66, 68, 69
в: Анисюткин, 1999).
От P. minor новый вид отличается большими размерами (см. табл. 1) и более темной
окраской;
от P. beybienkoi - более короткими надкрыльями, не превышающими по длине мета-
нотум (достигают II тергита брюшка у P. beybienkoi), строением метатарзуса, гипан-
дрия и склеритов L2D и L3 гениталий самцов (ср. рис. 3, 5-10 и Fig. 7C, F, J, K
в: Anisyutkin, Yushkova, 2017);
от P. semialata - микроптерными надкрыльями самца (короткие у P. semialata:
ср. рис. 2, 5 и fig. 2 в: Anisyutkin, 2020).
Э тим оло гия. Вид назван именем сборщика - энтомолога и геолога О. Н. Каба-
кова.
397
Рис. 3. Placoblatta kabakovi sp. n., самец, голотип.
1 - вершина брюшка сверху; 2 - парапрокты снизу; 3 - гипандрий и VIII стернит брюшка снизу; 4 - правая
фалломера сверху; 5 - склерит L2D сверху; 6, 7 - вершина склерита L2D сверху (6) и сбоку (слева) (7);
8 - склериты L3 и L4U; 9 - склерит L3; 10 - вершина склерита L3. Пунктировкой показаны мембранозные
участки.
Масштабная линейка - 1 мм: a - к рис. 1, 2; b - к рис. 3; c - к рис. 4; d - к рис. 5; e - к рис. 6-10.
Род ANISOLAMPRA Bey-Bienko, 1969
Ти по во й в ид: Anisolampra panfilovi Bey-Bienko, 1969, по монотипии.
Единственный вид рода был описан Г. Я. Бей-Биенко (1969) из Юньнани, типовая
серия была собрана в ходе советско-китайских зоолого-ботанических экспедиций
1955-1959 гг. в двух точках: «лес Маньянгуан, близ Дамонлун» и «Ганланба» (Бей-
Биенко, 1969, с. 841). Мне не удалось выяснить точное местонахождение «Дамон-
луна» и «Ганланбы». Информация от участника экспедиций О. Л. Крыжановского по-
зволила уточнить местонахождение этих точек, они расположены в южной части
провинции Юньнань: «Damonlung (30 km SW of Cheli); Ganlanba (30 km SO of Cheli,
on Mekong, near the frontier)». Необходимо также учитывать, что названия даны в
старой китайской транскрипции.
Типовая серия состоит из двух самок, включая голотип, одного самца и четырех ли-
чинок. Позднее один самец был указан из северного Вьетнама, окрестностей г. Винь
398
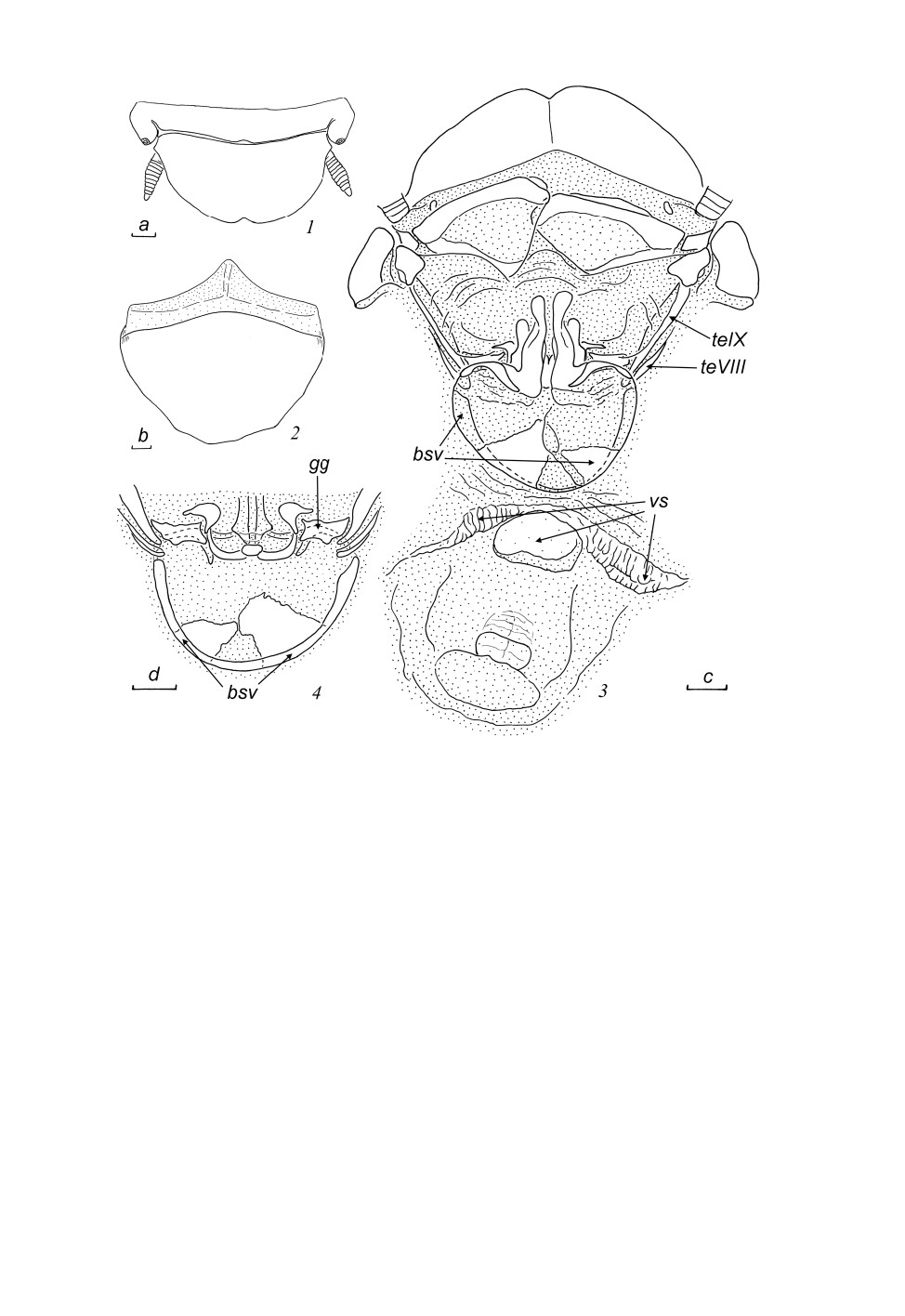
Рис. 4. Anisolampra panfilovi Bey-Bienko, 1969, гениталии самцов, экземпляр из провинции
Нгеан (1, 2, 6-10) и паратип (3-5).
1, 2 - правая фалломера сверху, общий вид (1) и очертания склеритов (2); 3-7 - вершина склерита
L2D снизу (3, 7), сверху (4, 6) и сверху под углом (5); 8-10 - склерит L3. Пунктировкой обозначены
мембранозные участки; щетинки на склеритах L2D не показаны.
Масштабная линейка - 1 мм: a - к рис. 1, 2, 8-10; b - к рис. 3-7.
(Vinh) в провинции Нгеан (Nghe An) (Анисюткин, 1999). Ниже приводится новый ма-
териал из Лаоса и дополняется морфологическое описание этого вида.
Со с т а в. Только типовой вид.
Anisolampra panfilovi Bey-Bienko, 1969 (рис. 4, 1-10; 5, 1-4).
М ате риа л. Китай. Провинция Юньнань, Дамонлун, 50 км ЮЗ Чэли, h = 700 м, 12.IV.1957,
Д. Панфилов, 1 ♀ (голотип), 1 самец (паратип); «Юньнань, Дамонлун, ю. Чэли, 700 м. 12.IV.1957.
Д. Панфилов 54 [возможно, “57”]», 1 ♀ (паратип); Дамонлун, Ю Чэли, h = 700 м, 12.IV.1957
(Пу Фу-ди и др.), 2 личинки (паратипы); Ганланба, 540 м, 19.IV.1957 (Лю Да-хуа), 1 личинка
(паратип); те же данные, h = 560 (Хун Гуан-ди), 1 личинка (паратип).
Вьетнам. Провинция Нгеан (Nghe An), окр. г. Винь (Vinh), ЮЗ Куи-Тяу, верховья р. Кон (Con),
15.02.1964 (О. Н. Кабаков), 1 ♂. Лаос. Провинция Вьентьян (Vientiane), национальный парк Ban
Vangheua, 18°21.097ʹ N, 102°48.897ʹ E, h = 800 м, тропический лес, dung trap, 31.V-01.X.2009
(С. Тарасов), 1 ♂, 11 ♀ (включая экземпляр № 260514/02), 2 личинки.
Описания вида (Бей-Биенко, 1969; Анисюткин, 1999) могут быть дополнены следу-
ющими данными.
Ге нит а л ии с а мц а (рис. 4, 1-10). Правая фалломера (склериты R + N) (рис. 4, 1, 2): кау-
дальная часть склерита R1T примерно трапециевидная, слабо выпуклая по каудальному краю,
399
Рис. 5. Anisolampra panfilovi Bey-Bienko, 1969, самка, Лаос (экземпляр № 260514/02).
1 - вершина брюшка сверху; 2 - генитальная пластинка снизу; 3 - вершина брюшка снизу, генитальная
пластинка удалена; 4 - основание яйцеклада сверху. Пунктировкой обозначены мембранозные участки (не
показаны на створках яйцеклада).
Масштабная линейка - 1 мм: a - к рис. 1, b - к рис. 2, c - к рис. 3, d - к рис. 4.
густо покрыта крупными щетинками, краниальная часть R1T удлиненная и плавно изогнутая;
склерит R2 массивный и короткий; краниальные части R1T и R2 оттянуты в слабо склеротизо-
ванный вырост (показан точками на рис. 4, 2); R3 широкий, слабо склеротизованный; R4 в виде
удлиненной изогнутой пластинки; R5 большой и удлиненный, соединен со склеритом R3. Скле-
рит L2D (L1) разделен на апикальный и базальный субсклериты; апикальный субсклерит шапоч-
ковидный, густо покрыт прижатыми щетинками, его форма варьирует у разных популяций: он
более или менее округленный при осмотре сверху у самца из типовой серии (Юньнань)
(рис. 4, 3-5), но отчетливо поперечный у самцов из Вьетнама (рис. 4, 6, 7) и Лаоса; дорсальный
склерит плоский (рис. 4, 4, 6, d. o), дорсальный вырост не развит. Склерит L3 (L2d) удлиненный
(рис. 4, 8-10), базальный субсклерит присутствует (рис. 4, 8-10, b. L3); складчатая структура
хорошо выражена, щетинки слабые, трудно различимые; вершина L3 без гребней, с хорошо вы-
раженным апикальным зубчиком; борозда hge отсутствует. Склерит L4U (L3d) широкий.
Самка (экземпляр из Лаоса). Вершина брюшка (рис. 5, 1-4).
400
Анальная пластинка (последний, X тергит) широкая (рис. 5, 1), каудально широко закруглен-
ная со слабо выраженным медиальным вырезом. Церки короткие, уплощенные, с плотно соеди-
ненными сегментами (рис. 5, 1). Парапрокты склеротизованные (рис. 5, 3), симметричные, без
вооружения, но со срединным мембранозным участком. Генитальная пластинка (рис. 5, 2) широ-
кая, без выреза по заднему краю.
Яйцеклад и прилежащие структуры (рис. 5, 3, 4). Тергальные выросты VIII тергита брюшка не
достигают его паратергитов, маленькие (рис. 5, 3, 4, teVIII); тергальные выросты IX тергита
полностью развиты (рис. 5, 3, 4, teIX). Гонангулюмы крупные и хорошо склеротизованные, их
вентральная часть в виде изогнутых склеротизованных пластинок (рис. 5, 3, 4, gg). Створки
яйцеклада частично мембранозные; первая пара самая крупная, с рядами щетинок по внутренне-
му краю; вторая пара удлиненная, полностью скрыта под первой; створки третьей пары (гоно-
плаки) расширены и укорочены; основания второй и третьей пар - рис. 5, 4. Базивальвула
цельная, широкая и округленная, слегка асимметричная (рис. 5, 3, 4, bsv). Вестибулярная струк-
тура слабо склеротизована, с боковыми ветвями и срединной папиллой (рис. 5, 3, vs). Выводко-
вая сумка без склеротизованных структур, только с уплотненным участком, прилежащим к
папилле.
Размеры - см. табл. 1.
ОБСУЖДЕНИЕ
Принадлежность родов Placoblatta и Anisolampra к трибе Morphnini McKittrick,
1964 следует из характерного строения правой фалломеры (Анисюткин, 2017;
Anisyutkin, 2020). Сложное строение этой структуры (см. рис. 1, 1, 2; 3, 4; 4, 1, 2) де-
лает маловероятным ее независимое возникновение в разных группах. Строение яй-
цеклада и прилежащих структур у Placoblatta и Anisolampra также в основных чертах
сходно с таковым у рода Morphna Shelford, 1910 (Anisyutkin, 2018).
Представители родов Placoblatta и Anisolampra сходны по следующим признакам:
покровы сильно склеротизованные, скульптурированные или бугорчатые, способ-
ность к полету утрачена как минимум у самок; сходно строение яйцеклада и приле-
жащих структур, особенно характерно строение вентральных частей гонангулюмов в
виде изогнутых склеротизованных пластинок. Можно предположить их близкое род-
ство, но сходство по признакам внешнего строения может объясняться параллельной
реверсией к личиночному состоянию (педоморфозом), а строение яйцеклада и приле-
жащих структур известно далеко не у всех эпиламприн. Думается, это как раз тот
случай, когда помочь в решении вопроса могут молекулярно-генетические исследо-
вания, которые не проводились для этих родов.
Утрата полета у обоих полов (Placoblatta) или только у самок (Anisolampra) и уси-
ление склеротизации покровов могут быть интерпретированы как сдвиг от фитофилии
к геофилии. В пользу геофильности может свидетельствовать и массовое попадание
A. panfilovi в почвенную ловушку в Лаосе (см. Материал). С другой стороны, хорошо
развитые эуплантулы и аролиумы являются адаптациями к перемещению по гладким
поверхностям (Roth, Willis, 1952), в природных условиях - обычно по листве. Можно
предположить, что несмотря на утрату полета представители Placoblatta и Anisolampra
сохранили черты фитофильности. К сожалению, образ жизни видов этих родов неиз-
вестен.
401
БЛАГОДАРНОСТИ
Автор искренне признателен всем сборщикам материала, на котором основана
данная работа. Особенно благодарен автор Н. Л. Орлову (ЗИН) и П. В. Озерскому
(Всероссийский институт растениеводства им. Н. И. Вавилова, С.-Петербург) за кон-
сультации и обсуждение некоторых разделов рукописи.
ФИНАНСИРОВАНИЕ
Исследование выполнено в рамках темы государственного задания «Систематика,
морфология, экофизиология и эволюция насекомых», № 122031100272-3.
СПИСОК ЛИТЕРАТУРЫ
Анисюткин Л. Н. 1999. Тараканы подсем. Epilamprinae (Dictyoptera, Blaberidae) Индокитайского полуостро-
ва. Энтомологическое обозрение 78 (3): 565-588. [Anisyutkin L. N. 1999. Cockroaches of the subfamily
Epilamprinae (Dictyoptera, Blaberidae) from the Indochina Peninsula. Entomological Review 79 (4):
434-454)].
Анисюткин Л. Н. 2012. Описание нового вида тараканов рода Prosoplecta Saussure, 1864 (Dictyoptera,
Ectobiidae) из Южного Вьетнама. Энтомологическое обозрение 91 (4): 742-756. [Anisyutkin L. N. 2013.
A description of a new species of the cockroach genus Prosoplecta Saussure, 1864 (Dictyoptera, Ectobiidae)
from South Vietnam. Entomological Review 93 (2): 182-193).
Анисюткин Л. Н. 2017. О таксономическом положении рода Stictomorphna Bruijning, 1848 (Dictyoptera,
Blaberidae). Энтомологическое обозрение 96 (4): 825-832. [Anisyutkin L. N. 2017. On the taxonomic
position of the genus Stictomorphna Bruijning, 1848 (Dictyoptera, Blaberidae). Entomological Review 97
Анисюткин Л. Н. 2018. Новый вид рода Placoblatta Bey-Bienko (Dictyoptera, Blaberidae: Epilamprinae) из
южного Вьетнама. Энтомологическое обозрение 97 (2): 287-292. [Anisyutkin L. N. 2018. A new species
of the genus Placoblatta Bey-Bienko (Dictyoptera, Blaberidae: Epilamprinae) from Southern Vietnam.
Entomological Review 98 (3): 352-356).
Бей-Биенко Г. Я. 1950. Насекомые таракановые. Фауна СССР, новая серия № 40. М.; Л.: Издательство Акаде-
мии наук СССР, 343 с.
Бей-Биенко Г. Я. 1969. Новые роды и виды таракановых (Blattoptera) из тропической и субтропической Азии.
Энтомологическое обозрение 48 (4): 831-862.
Anisyutkin L. N. 2008. Paraeuthyrrhapha groehni gen. et sp. nov., a new genus of the family Polyphagidae
(Dictyoptera) from Baltic amber and its phylogenetical position. Alavesia 2: 77-85.
Anisyutkin L. N. 2018. Little known Epilamprinae (Dictyoptera: Blaberidae) from the collections of the Muséum
d’histoire naturelle de Genève. Part 3. Revue Suisse de Zoologie 125 (1): 1-16.
Anisyutkin L. N. 2020. New and little known Epilamprinae (Dictyoptera: Blaberidae) from the collections of the
Muséum d’histoire naturelle de Genève and the Zoological Institute of Saint Petersburg. Part 4. Revue Suisse
Anisyutkin L. N., Yushkova O. V. 2017. New data on cockroaches of the subfamily Epilamprinae (Dictyoptera:
Blaberidae) from India and Sri Lanka, with descriptions of new species and the genital complex of Aptera
fusca (Thunberg, 1784). Zootaxa 4236 (1): 41-64.
Chopard L. 1925. Le distribution géographique des Blattinae apteres au subapteres (Orth. Blattidae). Association
française pour l’avancement des sciences, Congres Liege, 1924, p. 975-977.
Karny H. H. 1924. Beiträge zur malayischen Orthopterenfauna. Treubia 5: 1-234.
Klass K.-D. 1997. The External Male Genitalia and the Phylogeny of Blattaria and Mantodea. Bonner Zoologische
Monographien, vol. 42. Bonn: Zoologisches Forschungsinstitut und Museum Alexander Koenig, 341 p.
Klass K.-D. 1998. The Ovipositor of Dictyoptera (Insecta): Homology and Ground-Plan of the Main Elements.
Zoologischer Anzeiger 236: 69-101.
Li X.-R., Li M., Wang Z.-Q. 2017. Preliminary molecular phylogeny of beetle cockroaches (Diploptera) and notes
on male and female genitalia (Blattodea: Blaberidae: Diplopterinae). Zootaxa 4320 (3): 523-534.
402
McKittrick F. A. 1964. Evolutionary Studies of Cockroaches. Cornell University Agricultural Experiment Station
Memoir, vol. 389. Ithaca: Cornell University Agricultural Experiment Station; New York: State College of
Agriculture, 197 p.
Qiu L., Yang Z.-B., Wang Z.-Q., Che Y.-L. 2019. Notes on some corydiid species from China, with the description
of a new genus (Blattodea: Corydiidae: Euthyrrhaphinae). Annales de la Société Entomologique de France
Qiu L., Wang Z.-Q., Che Y.-L. 2020. Discovery of the second Asian Holocompsa species from China, and
supplemental description of the male of H. debilis (Blattodea: Corydiidae: Euthyrrhaphinae). Annales de la
Société Entomologique de France (N. S.) 56 (6): 481-487.
Rehn J. A. G. 1932. On apterism and subapterism in the Blattinae (Orthoptera: Blattidae). Entomological News 43:
201-206.
Roth L. M. 1977. A taxonomic revision of the Panesthiinae of the world. I. The Panesthiinae of Australia (Dictyoptera:
Blattaria: Blaberidae). Australian Journal of Zoology, Supplementary Series 48: 1-112.
Roth L. M. 2003a. Systematics and phylogeny of cockroaches (Dictyoptera: Blattaria). Oriental Insects 37: 1-186.
Roth L. M. 2003b. Some cockroaches from Africa and islands of the Indian Ocean, with descriptions of three new
species (Blattaria). Transactions of the American Entomological Society 129 (1): 167-186.
Roth L. M., Willis E. R. 1952. Tarsal structure and climbing ability of cockroaches. Journal of Experimental Zoology
119 (3): 483-517.
Storozhenko S. Yu., Paik J.-Ch. 2007. Orthoptera of Korea. Vladivostok: Dalnauka, 231 p.
Yue Q., Wu K., Qiu D., Hu J., Liu D., Wei X., Chen J., Cook C. E. 2014. A formal re-description of the cockroach
Hebardina concinna anchored on DNA barcodes confirms wing polymorphism and identifies morphological
characters for field identification. PLoS ONE 9 (9): e106789.
NEW DATA ON THE SYSTEMATICS AND DISTRIBUTION OF THE
COCKROACH GENERA PLACOBLATTA BEY-BIENKO, 1969
AND ANISOLAMPRA BEY-BIENKO, 1969
(DICTYOPTERA, BLABERIDAE: EPILAMPRINAE)
L. N. Anisyutkin
Key words: cockroaches, morphology, new species, f. macroptera, f. brachyptera,
f. microptera, f. aptera.
SUMMARY
The paper deals with cockroaches of the genera Placoblatta and Anisolampra. A new species
Placoblatta kabakovi sp. n. is described from Vietnam, and female of P. minor is described for the first
time. Additional data on the morphology and geographical distribution of P. rugosa and A. panfilovi
are presented. The ovipositor and adjacent structures of these species are described for the first time.
A nomenclature of modifications of the structure of fore and hind wings of cockroaches is proposed.
403