ВВЕДЕНИЕ
Баклажан (Solanum melongena L.) является экономически значимой и популярной овощной культурой. В последнее время плоды баклажана рассматриваются как функциональный продукт питания, так как они обогащены широким спектром фенольных соединений: мякоть является источником фенольных кислот, а кожица – антоцианов. Кроме того, листья содержат большое количество кемпферола [1, 2]. Мощные антиоксидантные свойства плодовых экстрактов баклажана делают их применение перспективным для снижения риска различных заболеваний человека [2, 3].
Кожица плода S. melongena характеризуется белой, зеленой или фиолетовой окраской [2, 4]. Оттенки окраски кожицы от бледно-лилового до черно-фиолетового обусловлены синтезом и накоплением разного количества антоцианов с преобладанием гликозидов дельфинидина (93–98% от общей суммы) и минорными включениями производных других антоцианидинов [2, 4]. Другие известные виды баклажана (Solanum aethiopicum L., Solanum macrocarpon L. и др.), широко возделываемые в Африке и Юго-Восточной Азии, формируют плоды желтой, оранжевой или красной окраски за счет накопления каротиноидов. Содержание антоцианов в таких плодах, также как в плодах баклажана с белой и зеленой кожицей, минимально [4].
Путь биосинтеза антоцианов в растениях высоко консервативен и является ветвью пути биосинтеза флавоноидов. В результате последовательных ферментативных реакций, катализируемых халконсинтазой (CHS), халконизомеразой (CHI), флаванон-3-гидроксилазой (F3H), флавоноид-3'-гидроксилазой (F3'H), флавоноид-3'5'-гидроксилазой (F3'5'H), дигидрофлавонол-4-редуктазой (DFR) и антоцианидинсинтазой (ANS), образуются окрашенные антоцианидины – пеларгонидин, цианидин и дельфинидин. Дальнейшая модификация антоцианидинов с участием UDP-глюкозофлавоноид-3-O-глюкозилтрансферазы (UFGT) приводит к образованию стабильных, водорастворимых пигментов. Антоцианидины пеонидин, петунидин и мальвидин образуются в результате метилирования цианидина и дельфинидина под действием O-метилтрансферазы (OMT) [5–7].
В растительных клетках флавоноиды, в том числе антоцианы, синтезируются в цитозоле, а затем транспортируются и накапливаются в вакуолях [8]. Транспорт флавоноидов в вакуоли осуществляется специфической глутатион-S-трансферазой класса phi (кодируется геном GSTF12) [9]. Для многих видов растений показана высокая корреляция уровня транскриптов транспортера GSTF12 с накоплением антоцианов в тканях [9–11].
Экспрессия генов пути биосинтеза антоцианов координируется MBW-комплексом, включающим транскрипционные факторы (ТФ) семейств R2R3-MYB, bHLH и WD-repeat [12, 13]. Различия в составе участников MBW-комплекса определяют пространственно-временную регуляцию синтеза антоцианов в растении [14, 15]. Показано, что ТФ семейства R2R3-MYB также регулируют транскрипцию гена транспортера антоцианов GSTF12 [11].
В транскриптомах S. melongena, по гомологии с известными последовательностями томата и картофеля, идентифицированы основные структурные гены пути биосинтеза антоцианов и определены профили их экспрессии в сопоставлении с содержанием антоцианов [16–18]. К изученным регуляторным генам пути биосинтеза антоцианов у баклажана относятся отдельные представители семейств R2R3-MYB (SmMYB1/SmMYB113 (KF727476.1), SmMYB2 (KF727477.1)), bHLH (SmbHLH1/SmGL3 (GLABRA3; KF727475.1), SmTT8 (TRANSPARENT TESTA8; KP006503.1)) и WD40-repeat (SmAN11 (KF727473)) [16, 19]. У родственных баклажану видов, томата (Solanum lycopersicum L.) и перца (Capsicum annuum L.), идентифицированы новые ТФ семейств R2R3-MYB и bHLH, ассоциированные с биосинтезом антоцианов [15, 20]. Так, показано, что накопление антоцианов в плодах томата связано с активностью гена SlMYB75; сверхэкспрессия данного гена приводит к существенному росту содержания антоцианов, а также общего содержания фенолов, флавоноидов, растворимых сухих и ароматических летучих веществ [20]. Анализ транскриптомов кожицы плода перца, приобретающей пурпурную окраску в условиях ультрафиолетового излучения, выявил факторы транскрипции CabHLH137 и CabHLH143, которые связываются с промоторами структурных генов пути биосинтеза антоцианов, активируя их транскрипцию [15].
Цель работы – поиск в геноме S. melongena последовательностей структурных и регуляторных генов пути биосинтеза антоцианов, включая генов-гомологов SlMYB75, CabHLH137, CabHLH143 и GSTF12; изучение профилей их экспрессии в различных органах трех сортов баклажана и определение взаимосвязи уровней транскрипции с содержанием антоцианов.
МАТЕРИАЛЫ И МЕТОДЫ
Для проведения работы в 2022 г. образцы баклажана S. melongena сортов Снежный, Влас и Агат, различающиеся окраской спелого плода, были выращены до стадии плодоношения в пленочной теплице Федерального научного центра овощеводства (Московская обл.). Все три сорта формируют зеленые листья и цветки с фиолетовыми лепестками. Плоды сорта Снежный имеют белую/светло-желтую окраску в технической/биологической спелости. Плоды сортов Влас и Агат темно-фиолетовые и почти черные в технической спелости и светлеют к стадии биологической спелости (рис. 1).
Рис. 1.
Листья, цветки и плоды (1 – незрелый плод (техническая спелость), 2 – спелый плод (биологическая спелость)) сортов баклажана Снежный (а), Влас (б) и Агат (в).

В августе с растений были собраны листья, цветки, незрелые плоды (стадия технической спелости) и спелые плоды (стадия биологической спелости). У плодов отделяли экзокарп (кожицу), у цветков – лепестки, у листьев – фрагменты ткани без жилок. Собранный растительный материал растирали в жидком азоте и использовали для биохимического анализа и анализа экспрессии генов. Содержание суммы антоцианов (в пересчете на дельфинидин-3-рутинозид) определяли спектрофотометрически в хлороформ-метанольных экстрактах [21] в двух биологических и трех технических повторностях. Суммарную РНК выделяли и очищали от примесей ДНК с помощью наборов RNeasy Plant Mini Kit и RNase free DNasy set (“QIAGEN”, Германия); кДНК синтезировали с использованием набора GoScriptтм Reverse Transcription System (“Promega”, США).
Поиск структурных и регуляторных генов пути биосинтеза антоцианов проводили в геномных (GCA_000787875.1) [22] и транскриптомных данных баклажана S. melongena, доступных в GenBank NCBI (https://www.ncbi.nlm.nih.gov/). Для поиска по гомологии использовали последовательности известных генов баклажана (SmMYB1 (KF727476.1), SmMYB2 (KF727477.1), SmTT8 (KP006503.1), SmGL3 (KP006502.1)), томата S. lycopersicum (CHS1 (NM_001247104.2), CHS2 (NM_001247107.2), CHI (NM_001320711.1), F3H (NM_001329483.1), F3'5'H (NM_001247911.2), DFR (Z18277.1), ANS (NM_001374394.1), UFGT (XM_004249399.4), GSTF12 (XM_004232621.4), SlMYB75 (NM_001279063.2)) и перца C. annuum (bHLH137 (XM_016712236.2), bHLH143 (XM_016695337.2)). Выравнивание найденных последовательностей проводили в программе MEGA 7.0 (https://www.megasoftware.net/). Молекулярную массу (MW) и изоэлектрическую точку (pI) белков определяли с помощью Expasy ProtParam (https://www.web.expasy.org/protparam).
Профиль экспрессии структурных (CHS1, CHI, F3H, F3´5´H, DFR, ANS, UFGT и GSTF12) и регуляторных (GL3, TT8(MYC), bHLH137, bHLH143, MYB1, MYB2 и MYB75) генов пути биосинтеза антоцианов определяли методом количественной ПЦР в реальном времени (ПЦР-РВ) в трех технических повторностях. Праймеры разрабатывали на основе найденных в базе данных NCBI последовательностей (Дополнительные материалы, табл. 1). Относительный уровень экспрессии генов оценивали, нормализуя по экспрессии референсного гена GAPDH [16]. ПЦР-РВ проводили с помощью набора “Реакционная смесь для проведения ПЦР-РВ в присутствии SYBR GreenI и ROX” (ООО “Синтол”, Россия) и амплификатора CFX96 Real-Time PCR Detection System (“Bio-Rad Laboratories”, США) в следующих условиях: 95°C – 5 мин.; 40 циклов (95°C – 15 с, 62°C – 50 с). Визуализацию данных ПЦР-РВ, а также регрессионный анализ (поиск корреляции между уровнем транскриптов гена и содержанием антоцианов) осуществляли в программе GraphPad Prism v. 7.02 (https://www.graphpad.com).
Таблица 1.
Характеристика структурных и регуляторных генов пути биосинтеза антоцианов баклажана S. melongena
| Ген | Контиг/ID* | Ген, пн | Кол-во экзонов | Кодирующая часть, пн | Белок, ао | MW, кДа | pI |
|---|---|---|---|---|---|---|---|
| Структурные гены пути биосинтеза антоцианов | |||||||
| CHS1 | Sme2.5_13923.1/ Sme2.5_13923.1_g00001.1 | 1254 | 2 | 1170 | 389 | 42.5 | 7.17 |
| CHS2 | Sme2.5_02154.1/ Sme2.5_02154.1_g00001.1 | 1344 | 2 | 1170 | 389 | 42.7 | 5.98 |
| CHI | Sme2.5_01193.1/ Sme2.5_01193.1_g00009.1 | 1811 | 4 | 705 | 234 | 25.2 | 5.38 |
| F3H | Sme2.5_00015.1/ Sme2.5_00015.1_g00016.1 | 2613 | 3 | 1101 | 366 | 41.1 | 5.39 |
| F3'5'H | Sme2.5_04313.1/ Sme2.5_04313.1_g00001.1 | 3064 | 3 | 1542 | 513 | 57.7 | 8.88 |
| DFR | Sme2.5_01401.1/ Sme2.5_01401.1_g00004.1 | 1638 | 6 | 1149 | 382 | 42.6 | 5.47 |
| ANS | Sme2.5_01638.1/ Sme2.5_01638.1_g00005.1 | 1556 | 2 | 1242 | 413 | 46.7 | 5.34 |
| UFGT | Sme2.5_00228.1/ Sme2.5_00228.1_g00013.1 | 1329 | 1 | 1329 | 442 | 49.2 | 5.92 |
| Глутатион-S-трансфераза GSTF12 | |||||||
| GSTF12 | Sme2.5_00172.1/ Sme2.5_00172.1_g00024.1 | 1735 | 3 | 693 | 230 | 26.3 | 5.88 |
| ТФ семейства bHLH | |||||||
| TT8 | Sme2.5_00592.1/ Sme2.5_00592.1_g00005.1 | 8531 | 9 | 1896 | 631 | 70.1 | 5.17 |
| GL3 | Sme2.5_12406.1/ Sme2.5_12406.1_g00003.1 | 4556 | 7 | 1887 | 628 | 70.8 | 5.61 |
| bHLH137 | Sme2.5_06479.1/ Sme2.5_06479.1_g00003.1 | 2443 | 7 | 1032 | 343 | 38.2 | 6.27 |
| bHLH143 | Sme2.5_06288.1/ Sme2.5_06288.1_g00003.1 | 1092 | 1 | 1092 | 363 | 39.9 | 4.72 |
| ТФ семейства R2R3-MYB | |||||||
| MYB1 | Sme2.5_05099.1/ Sme2.5_05099.1_g00002.1 | 1230 | 3 | 771 | 256 | 29.6 | 6.17 |
| MYB2 | Sme2.5_00538.1/ Sme2.5_00538.1_g00005.1 | 3727 | 3 | 837 | 278 | 32.1 | 8.48 |
| MYB75 | Sme2.5_13100.1/– | 1051 | 3 | 822 | 273 | 31.8 | 8.87 |
* Согласно [22].
РЕЗУЛЬТАТЫ
Структурные и регуляторные гены пути биосинтеза антоцианов в геноме баклажана. В геноме баклажана S. melongena (https://www.ncbi.nlm.nih.gov/ d-ata-hub/genome/GCA_000787875.1/) был проведен поиск по гомологии структурных и регуляторных генов биосинтеза антоцианов, включая SlMYB75, CabHLH137, CabHLH143 и GSTF12. В результате были найдены и охарактеризованы предположительные ортологи всех известных структурных (CHS1, CHS2, CHI, F3H, F3'5'H, DFR, ANS и UFGT) и регуляторных (ТФ семейств bHLH (TT8, GL3, bHLH137 и bHLH143) и R2R3-MYB (MYB1, MYB2 и MYB75)) генов, а также гена транспортера антоцианидинов – глутатион-S-трансферазы GSTF12. Основные характеристики генов S. melongena и кодируемых ими белков приведены в табл. 1.
Органоспецифичные профили экспрессии структурных генов пути биосинтеза антоцианов. Профили экспрессии структурных генов биосинтеза антоцианов и транспортера антоцианидинов SmGSTF12 были определены в листьях, лепестках цветка, кожице незрелого и спелого плода сортов баклажана Снежный, Влас и Агат (рис. 2).
Рис. 2.
Профили экспрессии структурных генов флавоноидного пути (CHS1, CHS2, CHI, F3H, F3'5'H, DFR, ANS, UFGT) и транспортера антоцианидинов (GSTF12) в листьях (1), лепестках цветка (2), кожице незрелого (3) и спелого (4) плодов сортов баклажана Снежный, Влас и Агат.
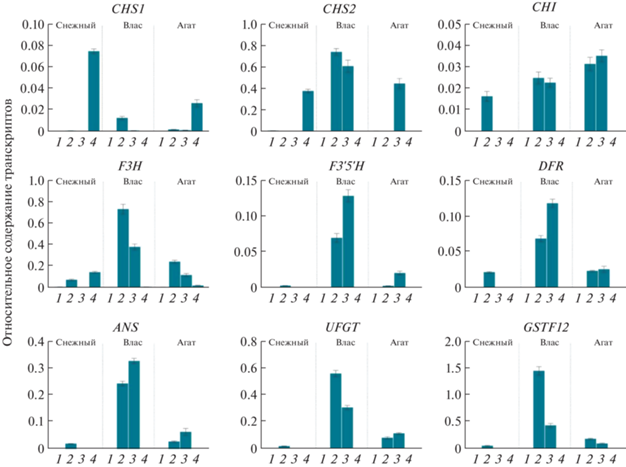
У сорта Снежный в листьях экспрессии генов биосинтеза антоцианов не выявлено, за исключением следовых количеств транскриптов SmCHS2 и SmF3H; в лепестках, имеющих фиолетовую окраску, присутствовали транскрипты всех генов (за исключением SmCHS2) и SmGSTF12. В белой кожице незрелого плода сорта Снежный транскрипты генов не обнаружены, тогда как в светло-желтой кожице спелого плода зафиксированы транскрипты ранних генов флавоноидного пути (SmCHS1, SmCHS2 и SmF3H) (рис. 2).
У сортов Влас и Агат, имеющих фиолетовую окраску лепестков и кожицы плода, показана активность генов в лепестках (за исключением SmCHS2 у сорта Агат) и кожице незрелого плода; в кожице спелого плода выявлены транскрипты SmF3H (оба сорта) и SmCHS1 (сорт Агат). В листьях найдены следовые количества транскриптов SmCHS2 и SmF3H. Транскрипты GSTF12 обнаружены только в лепестках и кожице незрелого плода (рис. 2).
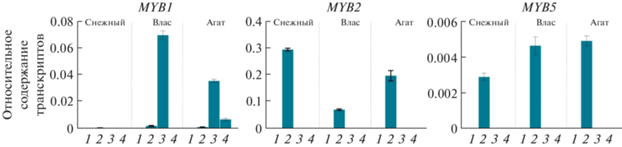
Органоспецифичные профили экспрессии генов ТФ семейств bHLH и R2R3-MYB. В тех же органах, где были определены профили экспрессии структурных генов, проанализированы профили экспрессии генов ТФ семейств bHLH и R2R3-MYB, ассоциированных с биосинтезом антоцианов (рис. 3, 4).
Рис. 3.
Профили экспрессии генов ТФ семейства bHLH в листьях (1), лепестках цветка (2), кожице незрелого (3) и спелого (4) плодов сортов баклажана Снежный, Влас и Агат.
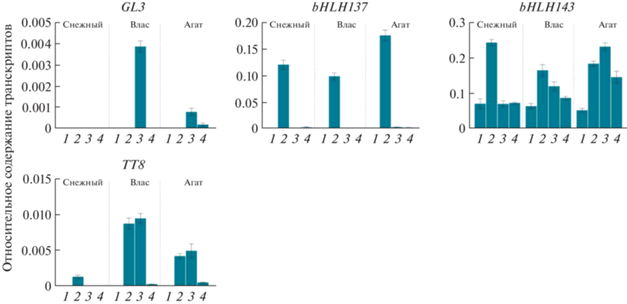
Рис. 4.
Профили экспрессии генов ТФ семейства R2R3-MYB в листьях (1), лепестках цветка (2), кожице незрелого (3) и спелого (4) плодов сортов баклажана Снежный, Влас и Агат.
В результате транскрипты гена SmGL3 выявлены только в фиолетовой кожице плода у сортов Влас (стадия технической спелости) и Агат (обе стадии спелости). Ген SmbHLH137 высоко экспрессировался в лепестках всех трех сортов, небольшие количества транскриптов гена выявлены в кожице плодов. Экспрессия гена SmbHLH143 обнаружена во всех анализируемых органах с максимумом в лепестках цветка (сорта Снежный и Влас) или кожице незрелого плода (сорт Агат). Транскрипты SmTT8 детектированы в лепестках и кожице плодов (сорта Влас и Агат) или только в лепестках (сорт Снежный) (рис. 3).
Экспрессия гена SmMYB1 у анализируемых сортов баклажана выявлена в лепестках и в кожице плодов сортов Влас (незрелый плод) и Агат (незрелый и спелый плод). Транскрипты SmMYB2 и SmMYB75 обнаружены только в лепестках всех трех сортов (рис. 4).
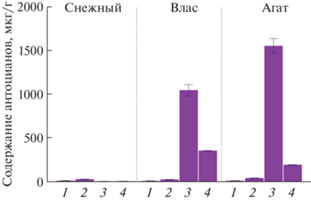
Органоспецифичное содержание суммы антоцианов и его связь с уровнем транскриптов структурных и регуляторных генов биосинтеза антоцианов. В листьях, лепестках цветка и кожице незрелого и спелого плодов сортов баклажана Снежный, Влас и Агат была определена сумма антоцианов. Показано, что листья всех сортов, имеющих зеленую окраску, содержали ~25 мкг/г антоцианов. В фиолетовых лепестках цветка у сортов Снежный и Влас содержание антоцианов было сходным (~40 мкг/г), у а сорта Агат – выше в 1.4 раза (~55 мкг/г). В кожице незрелого плода у сортов Влас и Агат сумма антоцианов составила 1054 и 1555 мкг/г, а в кожице спелого плода – 367 и 206 мкг/г, соответственно; это визуально соответствует изменению окраски плода по мере созревания с темно-фиолетовой/черной на более светлую, коричнево-фиолетовую (рис. 1, 5). У сорта Снежный содержание антоцианов в кожице плодов составило менее 20 мкг/г (рис. 5).
Рис. 5.
Содержание антоцианов в листьях (1), лепестках цветка (2), кожице незрелого (3) и спелого (4) плодов сортов баклажана Снежный, Влас и Агат; мкг/г сырого веса.
Содержание антоцианов было сопоставлено с уровнями экспрессии структурных и регуляторных генов флавоноидного пути. Для анализируемых образцов баклажана была выявлена значимая корреляция содержания антоцианов с уровнями транскриптов генов SmCHI, SmGL3 и SmMYB1 (рис. 6). Отдельно были определены значения коэффициентов корреляции содержания антоцианов в листьях, лепестках цветка и кожице плодов с уровнями транскриптов генов. В листьях содержание антоцианов коррелировало с уровнями транскриптов SmCHS2 и SmF3H; в лепестках цветка – SmCHI и SmbHLH137; в кожице плодов – всех анализируемых генов, кроме SmCHS1, SmF3H и SmbHLH137 (табл. 2).
Рис. 6.
Линейная регрессия содержания антоцианов совокупно во всех анализируемых органах баклажана с экспрессией структурных и регуляторных генов флавоноидного пути. R2 – коэффициент множественной корреляции, r – коэффициент корреляции Пирсона. Звездочкой отмечены значения с P ≤ 0.05.
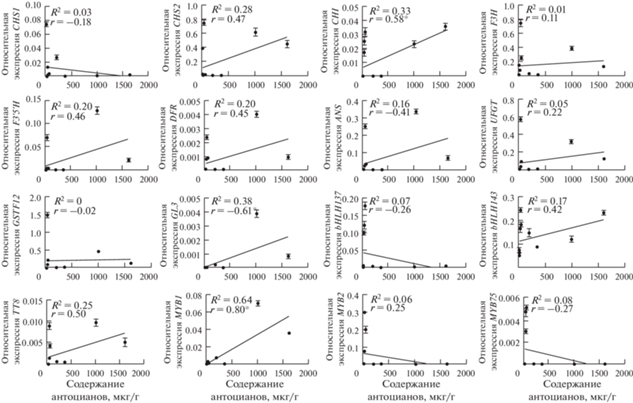
Таблица 2.
Значения коэффициентов корреляции Пирсона (r) между уровнем транскриптов анализируемых генов и содержанием суммы антоцианов отдельно в листьях, лепестках цветка и кожице плода баклажана (r ≥ 0.50 свидетельствует о наличии корреляции).
| CHS1 | CHS2 | CHI | F3H | F3’5’H | DFR | ANS | UFGT | GSTF12 | GL3 | bHLH137 | bHLH143 | TT8 | MYB1 | MYB2 | MYB75 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Листья | – | 0.81 | – | 0.71 | – | – | – | – | – | – | – | –0.27 | – | – | – | – |
| Лепестки цветка | –0.37 | –0.45 | 0.85 | –0.22 | –0.46 | –0.42 | –0.35 | –0.36 | –0.38 | – | 0.94 | –0.33 | –0.12 | –0.07 | 0.02 | 0.63 |
| Кожица плода | –0.47 | 0.65 | 0.98 | 0.48 | 0.50 | 0.55 | 0.54 | 0.67 | 0.58 | 0.53 | 0.40 | 0.85 | 0.76 | 0.75 | – | – |
ОБСУЖДЕНИЕ
Растение баклажана S. melongena является богатым источником различных фенольных соединений, благодаря чему в настоящее время плоды баклажана считаются функциональным продуктом питания [2, 3]. За счет высокого содержания антоцианов (преимущественно производных дельфинидина) кожица плода баклажана имеет фиолетовую окраску с палитрой оттенков от бледно розового до черного [2, 4, 16].
В настоящем исследовании с помощью поиска в геноме S. melongena нами были определены гомологи всех основных известных структурных и регуляторных генов пути биосинтеза антоцианов, а также гена транспортера антоцианидинов GSTF12 (табл. 1). С целью охарактеризовать совокупную роль данных генов в определении содержания антоцианов в различных органах баклажана был определен профиль их экспрессии у образцов трех сортов S. melongena, формирующих зеленые листья, цветки с фиолетовыми лепестками и плоды, кожица которых имеет окраску, различающуюся между сортами (белую у сорта Снежный и оттенки фиолетового у сортов Влас и Агат) (рис. 1).
В целом, для пигментированных тканей (лепестков цветка всех сортов и кожицы плода сортов Влас и Агат) было характерно присутствие транскриптов всех исследуемых генов (кроме SmMYB2 и SmMYB75 в плодах) (рис. 2–4), что подтверждает их участие в биосинтезе антоцианов. В то же время профили экспрессии генов у фиолетовоплодных сортов совпадали, и различие состояло лишь в уровнях транскриптов (и оттенках фиолетовой окраски, соответственно).
Заметим, что в белых и светло-желтых тканях кожицы технически и биологически спелого плода сорта Снежный были обнаружены транскрипты только ранних структурных генов флавоноидного пути (CHS1, CHS2 и F3H) (рис. 2). Это может свидетельствовать о синтезе бесцветных фенольных соединений и о предположительном наличии блоков в пути биосинтеза окрашенных антоцианидинов в плодах. Отсутствие в кожице транскриптов большинства регуляторных генов (рис. 3, 4) говорит о том, что в плодах сорта Снежный не происходит образования регуляторного комплекса MBW в связи с дефицитом двух главных его составляющих – ТФ семейств bHLH и R2R3-MYB. Соответственно, не выполняется активация поздних структурных генов пути биосинтеза антоцианов и гена транспортера антоцианидинов (рис. 2), так как для этого необходим функциональный MBW-комплекс [11].
Интересно, что подобных блоков нет в лепестках цветка сорта Снежный, судя по профилю экспрессии анализируемых генов (рис. 2–4), что предполагает органоспецифичные нарушения в регуляции транскрипции генов bHLH и MYB MBW-комплекса в плодах, характерные для данного белоплодного сорта. При этом транскрипты гена SmMYB1 практически отсутствуют как в кожице плода, так и в лепестках цветка, что, вероятно, связано с обнаруженной ранее у сорта Снежный делецией 26 пн (конец интрона 1 – начало экзона 2), из-за которой в последовательности зрелой мРНК полностью отсутствует второй экзон и сбивается рамка считывания [23]. Праймеры, разработанные нами для анализа экспрессии гена SmMYB1, попадают на транскрибируемый участок, поэтому наличие следовых количеств транскриптов гена в лепестках и их отсутствие в кожице плода косвенно подтверждают предположение о нарушении регуляции транскрипции генов MYB MBW-комплекса в плоде сорта. К примеру, это может происходить благодаря повышенной наработке белков SmBIC (blue light inhibitors of cryptochromes), которые взаимодействуют с фотовозбужденным SmCRY2, ингибируя его активность и высвобождая убиквитиновую активность COP1, тем самым подавляя экспрессию генов биосинтеза антоцианов (в частности, SmMYB1, SmTT8 и SmHY5) [24].
Подчеркнем, что в кожице плода фиолетовоплодных сортов Влас и Агат из трех генов ТФ R2R3-MYB были обнаружены транскрипты только SmMYB1 (рис. 4). Это свидетельствует об участии именно данного ТФ (но не SmMYB2 и SmMYB75) в образовании MBW-комплекса в кожице плода. В то же время в лепестках цветка этих сортов низкие уровни транскриптов SmMYB1 и одновременно высокие SmMYB2 и SmMYB75 (рис. 4) предполагают преимущественное участие в MBW-комплексе ТФ SmMYB2 и SmMYB75. В этой связи высокий уровень транскриптов SmMYB2 и SmMYB75 в лепестках цветка сорта Снежный (рис. 4) может свидетельствовать о компенсаторном (заменяющем SmMYB1) участии данных белков в регуляции активности структурных генов биосинтеза антоцианов.
Таким образом, полученные данные по экспрессии SmMYB1 (также известного в литературе как SmMYB113) и его близких гомологов SmMYB2 и SmMYB75 в кожице плода (рис. 4) согласуются с ключевой ролью гена SmMYB113 в определении разнообразия композиции антоцианов в кожице баклажана и ее окраски [4, 16]. Последние опубликованные данные предлагают в качестве участников индуцированного светом MBW-комплекса баклажана еще три ТФ семейства R2R3-MYB: SmMYB35, SmMYB44 и SmMYB86 [14, 25, 26]. Тем не менее, наши данные по экспрессии SmMYB-1, SmMYB2 и SmMYB75 (рис. 4) исчерпывающе отвечают на вопрос о различии в окраске кожицы баклажана между белоплодными и фиолетовоплодными сортами. Возможно, SmMYB35, SmMYB44 и SmMYB86 выполняют дублирующие (по отношению к основным ТФ R2R3-MYB) роли. Более того, имеются сведения о SmMYB86 как о негативном регуляторе пути биосинтеза антоцианов [26].
Другие, не менее важные участники MBW-комплекса – ТФ семейства bHLH, также способны определять различия в составе антоцианов и окраске лепестков/кожицы плода у баклажанов. Анализ экспрессии четырех генов bHLH, гомологи которых ассоциированы с биосинтезом антоцианов, показал, что два из них – SmGL3 и SmTT8 – имеют существенные различия в уровне транскриптов в кожице плода между белоплодными и фиолетовоплодными сортами (рис. 3). Это является подтверждением ранее предложенной роли для этих двух генов в регуляции накопления антоцианов в растительной ткани [27].
Профиль экспрессии других двух генов bHLH показывает, что SmbHLH137 активен преимущественно в лепестках цветка S. melongena. В то же время повсеместное присутствие транскриптов SmbHLH143 не коррелирует с разницей в окраске кожице плода (рис. 3). Хотя связывание с промоторами структурных генов биосинтеза антоцианов выявлено для гомологов обоих ТФ [15], нужно учитывать, что они могут быть задействованы и в других процессах развития или стрессовых ответах растения [28–30].
Характеристикой роли генов в пути биосинтеза антоцианов может служить также корреляционный анализ зависимости между суммой антоцианов и уровнем транскриптов генов. Ожидаемо, в лепестках цветка присутствовала взаимосвязь наличия транскриптов анализируемых генов с наличием антоцианов (есть транскрипты – есть антоцианы) (рис. 2–5), однако корреляция с количеством антоцианов была выявлена только для генов CHI и bHLH137 (рис. 6; табл. 2). Тем не менее, в кожице плодов содержание антоцианов коррелировало с экспрессией всех анализируемых генов, за исключением CHS1, F3H и bHLH137 (табл. 2).
Таким образом, в данной работе была охарактеризована органоспецифичная функциональная активность структурных и регуляторных генов пути биосинтеза антоцианов в сравнении с содержанием суммы антоцианов и окраской. Было показано, что фиолетовоплодные сорта баклажана отличаются от белоплодного сорта профилем экспрессии анализируемых генов в кожице плода, что соответствует ее окраске. Присутствие антоцианов в фиолетовых лепестках цветка и их отсутствие в белых/светло-желтых плодах у сорта Снежный, а также различия в уровне транскриптов анализируемых генов между данными органами свидетельствуют о существовании блоков пути биосинтеза антоцианидинов в кожице плода данного сорта и особенностей органоспецифичной регуляции транскрипции генов ТФ комплекса MBW. На основе полученных результатов можно предположить, что в регуляции экспрессии структурных генов биосинтеза антоцианов в лепестках цветка S. melongena участвуют транскрипционные факторы SmbHLH137, SmTT8, SmMYB2 и SmMYB75, а в кожице плода – SmGL3, SmTT8 и SmMYB1.
Работа выполнена при финансовой поддержке гранта Российского Научного Фонда (№ 19-16-00016) и Министерства науки и высшего образования РФ.
Настоящая статья не содержит каких-либо исследований с использованием в качестве объекта людей и животных.
Авторы заявляют об отсутствии конфликта интересов.