Развитие аномалий позвоночника у рыб чаще всего изучают в связи с их отрицательным влиянием как на жизнеспособность рыб, так и на качество продукции в аквакультуре (Boglione et al., 2001, 2013; Koumoundouros, 2010; Cavrois-Rogacki et al., 2021; Fang, Xu, 2022). В природных популяциях частота встречаемости аномалий заметно ниже и её нередко рассматривают как показатель уровня загрязнения водоёмов (Boglione et al., 2006; Yershov, 2008; Bogutskaya et al., 2011; Kessabi et al., 2013; Есин, 2015; Яблоков, 2018).
Многие позвонковые нарушения у рыб закладываются во время раннего эмбрионального развития и реализуются в процессе остеогенеза (Ванюшина, 1990; Павлов, 2007; Boglione et al., 2013). Отклонения от нормы в строении позвонков могут развиться и в оптимальных условиях, однако при стрессовых воздействиях число аномалий возрастает. При этом разные причины могут спровоцировать развитие одних и тех же нарушений в строении позвоночника, а один и тот же фактор в зависимости от вида рыбы, стадии развития, времени воздействия и других факторов может вызывать разные аномалии (Von Westernhagen, 1988; Boglione et al., 2006). Наши исследования выявили многочисленные позвонковые аномалии у сеголеток плотвы Rutilus rutilus (Linnaeus, 1758) после воздействия ряда физических (магнитное поле, повышенная температура) и химических (хлорофос, фенол, N-метил-N’-нитро-N-нитрозогуанидин) факторов на эмбрионы (Касьянов и др., 2001; Чеботарева, 2009; Чеботарева и др., 2016).
Помимо нарушений в строении позвонков в наших экспериментах отмечали изменения их среднего числа у экспериментальной молоди. Изменение числа метамерных элементов в осевом скелете рыб относительно естественной нормы – также один из ответов на воздействие факторов различной природы в раннем онтогенезе (Lindsey, 1988; Павлов, 2007). Связь между количественными (число счётных элементов) и качественными (наличие, типы и число аномалий) показателями позвоночника прослеживается не всегда. В экспериментальных выборках у рыб с нарушениями развития позвоночника число позвонков обычно повышено (Касьянов и др., 2001; Чеботарева и др., 2016), тогда как в природных популяциях, напротив, понижено (Bogutskaya et al., 2011; Чеботарева, Изюмов, 2021).
Среди многообразия факторов окружающей среды, способных повлиять на развитие позвонков у рыб, особый интерес представляют низкочастотные магнитные поля. В зависимости от частоты и амплитуды воздействия регистрируемые эффекты могут существенно различаться (Krylov et al., 2016). Интерес также представляет возможная модификация эффектов магнитного поля другими физическими и химическими факторами (Крылов и др., 2010; Чеботарева и др., 2016). В настоящей работе в качестве дополнительного фактора использован хлорофос – широко применяемый инсектицид с нейротоксическими и мутагенными свойствами (Щербаков, 1983).
Ранее было показано, что совместное и/или раздельное воздействие магнитного поля 500 Гц, 150 мкТл и хлорофоса в концентрации 0.01 мг/л приводило к снижению выживаемости эмбрионов, изменению линейных размеров сеголеток, изменению активности пищеварительных ферментов в кишечнике молоди, увеличению числа позвонков, снижению числа отверстий в преоперкуло-мандибулярном канале на зубной кости (Голованова и др., 2006; Krylov et al., 2016). Однако мы не смогли найти информации о влиянии этих факторов на строение позвонков и позвонковые фенотипы.
Исходя из этого цель работы – детально проанализировать аномалии строения позвоночника и позвонковые фенотипы сеголеток плотвы после раздельного и совместного воздействия на эмбрионы магнитного поля и хлорофоса.
МАТЕРИАЛ И МЕТОДИКА
Эксперимент проводили в мае–сентябре 2005 г. Половые продукты были получены от производителей плотвы, отловленных неводом на нерестилище Рыбинского водохранилища: семь самок (стандартная длина 17.6–19.5 см, возраст 5–6 лет) и шесть самцов (17.6–19.5 см, одному 4, остальным – 6 лет).
Через 10 мин после осеменения сухим способом икру разделили на четыре группы и поместили в кристаллизаторы с речной водой (~2500–3000 икринок в каждом). В двух кристаллизаторах после приклеивания и набухания икринок воду заменили раствором хлорофоса с концентрацией 0.01 мг/л. Предыдущие исследования показали относительно высокую выживаемость эмбрионов плотвы, подвергшихся воздействию этого пестицида в указанной концентрации, но при этом наблюдались выраженные морфологические реакции у молоди (Чеботарева, 2009). Два кристаллизатора (один с речной водой, другой с раствором хлорофоса) были помещены внутрь колец Гельмгольца, подключённых к генератору сигналов ГЗ-102. Внутри колец генерировалось синусоидальное магнитное поле с частотой 500 Гц и величиной индукции 150 мкТл. Эти параметры были выбраны исходя из того, что поля с такой частотой чрезвычайно редко встречаются в естественной окружающей среде, в то время как величина индукции соответствует значению, которое может быть у полей, генерируемых различными электрическими устройствами (Havas, 2004). Такой подход позволил исключить вероятность адаптации и оценить влияние магнитного поля, которое не встречается в естественной среде, в отличие от полей с промышленными частотами 50 и 60 Гц, часто используемых в подобных исследованиях. Кроме этого, в предыдущих экспериментах влияние на эмбрионы плотвы магнитного поля с такими же параметрами приводило к значимым изменениям позвонковых фенотипов у сеголеток (Чеботарева и др., 2009). Другие два кристаллизатора (один с речной водой, другой с раствором хлорофоса) оставались в естественном геомагнитном поле. Таким образом, рассматривали четыре группы рыб (варианты), развитие которых проходило в разных условиях: К (контроль) – в речной воде в естественном геомагнитном поле; М – в речной воде в магнитном поле 500 Гц, 150 мкТл; Х – в растворе хлорофоса с концентрацией 0.01 мг/л в речной воде в естественном геомагнитном поле; МХ – в растворе хлорофоса с концентрацией 0.01 мг/л в речной воде в магнитном поле 500 Гц, 150 мкТл. Воду и раствор хлорофоса в кристаллизаторах меняли два раза в сутки. Продолжительность воздействия экспериментальных факторов составила 48 ч (от оплодотворения до начала органогенеза у зародышей), после чего все группы эмбрионов развивались в контрольных условиях (речная вода, естественное геомагнитное поле). Температура воды во время эмбрионального развития варьировала от 14 до 18°С.
После рассасывания желточного мешка и заполнения плавательного пузыря воздухом по 400 личинок из каждого варианта выпустили в пруды с естественной кормовой базой. Молодь отлавливали в 4-месячном возрасте.
Препарирование осевого скелета проводили по методике Яковлева с соавторами (1981). Определяли число позвонков в туловищном (vert.a.), переходном (vert.i.) и хвостовом (vert.c.) отделах позвоночника, а также их сумму (vert.), включая Веберовы и преуральные. Позвонковые фенотипы сеголеток представлены в виде формулы: vert.a.–vert.i.–vert.c. (Касьянов, 1990). Подсчитывали число рыб с определённым фенотипом и их долю (%).
В позвоночнике каждой рыбы определяли наличие аномалий. Таковыми считали деформации тел и дуг позвонков; сращения позвонков (входящие в сращение позвонки обычно укорочены, деформированы, чёткая граница между ними отсутствует); сращения невральных и гемальных дуг соседних позвонков; незамкнутые невральные или гемальные дуги; несращение дуги с телом позвонка; перемещение основания невральной или гемальной дуги на соседний позвонок (на одном из двух соседних позвонков ветвь невральной или гемальной дуги с одной стороны отсутствует, на другом – с той же стороны размещаются основания двух дуг); отсутствие ветвей невральных или гемальных дуг; наличие дополнительных ветвей невральной или гемальной дуги (внешне напоминает асимметричное сращение, но тело позвонка практически нормальной формы, признаки сращения позвонков со стороны дополнительной дуги отсутствуют) (Чеботарева, 2009). Для выборок рыб с аномалиями рассчитывали средние значения числа (встречаемость) каждого вида аномалий, позвонков с нарушениями по отделам позвоночника и определяли общее число позвонков.
Отдельно для рыб со сращениями позвонков проводили их оценку и сравнение, используя средневыборочные значения числа сращений. Число позвонков в одном сращении рассчитывали путём деления числа всех позвонков в составе сращений на общее число сращений позвонков у сеголеток из каждого варианта.
У плотвы встречаются симметричные (с обеих сторон позвоночника одно и то же число сросшихся тел позвонков) и асимметричные (с одной стороны больше тел позвонков, чем с другой) сращения позвонков (Чеботарева и др., 2016) (рис. 1). Подсчитывали долю (%) асимметричных сращений в общем числе сращений позвонков у сеголеток каждого варианта.
Рис. 1.
Типы сращений позвонков у плотвы Rutilus rutilus (вид слева (в центре – схема) и справа): а – нормальные позвонки, б – симметричное сращение, в – асимметричное сращение, г – встречные асимметричные сращения, д – последовательные асимметричные сращения (по: Чеботарева и др., 2016).
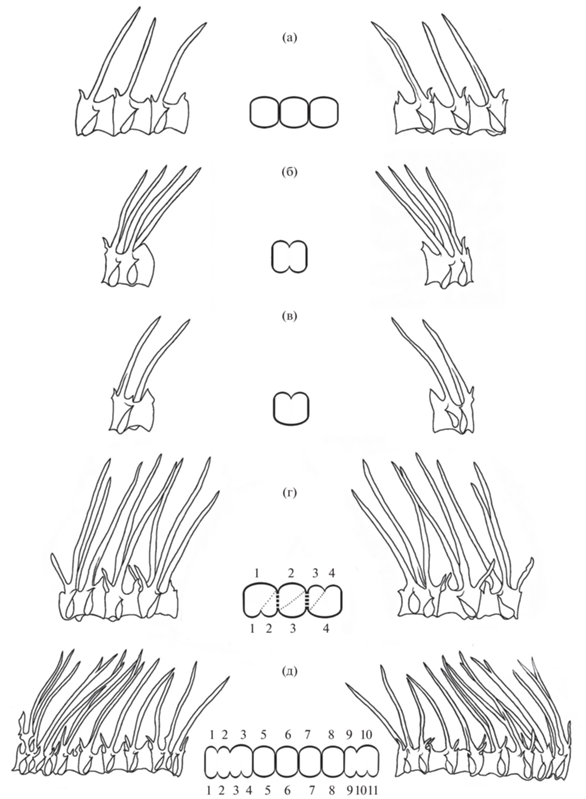
По взаимному расположению множественные асимметричные сращения позвонков у плотвы могут быть последовательными или встречными (Чеботарева и др., 2016). Дополнительные полупозвонки (непарные метамерные элементы позвоночника, развитые в асимметричных сращениях позвонков только слева или только справа) во встречных сращениях расположены с разных сторон позвоночника, а в последовательных – с одной стороны (рис. 1). Для характеристики разных видов сращений позвонков использовали их долю (%) в числе рыб со сращениями.
У рыб с асимметричными сращениями просчитывали позвонки отдельно слева и справа. У рыб с различиями в числе тел позвонков с разных сторон позвоночника (рыб с асимметричным позвоночником) в каждом варианте определяли средневыборочные значения числа позвонков слева и справа, а также разницы между левой и правой сторонами позвоночника.
Статистическую обработку материала проводили в программе Statistica 6.0. Для сравнения частот встречаемости рыб с аномалиями позвоночника и позвонковых фенотипов использовали критерий Животовского (1991).
РЕЗУЛЬТАТЫ
Выживаемость эмбрионов до вылупления в контроле составила 56.01%. В экспериментальных вариантах она достоверно, но незначительно снизилась: 41.14 (М), 44.25 (МХ) и 52.76% (Х). Кривые выживаемости эмбрионов и значения других показателей, не связанных с аномалиями строения позвоночника и позвонковыми фенотипами, для рыб из исследованных групп можно найти в нашей предыдущей статье (Krylov et al., 2016).
Аномалии позвоночника. Доля рыб с отклонениями строения позвонков была существенной во всех группах: в контроле она составила 42.68%, в других группах была более чем в 1.5 раза выше контрольной. Из всех перечисленных аномалий развития позвонков чаще отмечали незамкнутые невральные или гемальные дуги. Также достаточно часто встречались деформированные позвонки и сращения позвонков. К аномалиям со средней встречаемостью относились сращения дуг соседних позвонков, деформации и отсутствие невральных или гемальных дуг. Шесть перечисленных типов нарушений в строении позвонков отмечали во всех выборках сеголеток плотвы (табл. 1, 2).
Таблица 1.
Характеристика аномалий позвоночника сеголеток плотвы Rutilus rutilus в контрольной и экспериментальных выборках
| Показатель | Вариант | |||
|---|---|---|---|---|
| К | М | МХ | Х | |
| Число рыб, экз.: | ||||
| – общее | 82 | 112 | 138 | 130 |
| – с аномалиями | 35 | 76 | 100 | 86 |
| Доля рыб с аномалиями, % | 42.68м, мх, х | 67.86к | 72.46к | 66.15к |
| Число позвонков с аномалиями* | ||||
| Весь позвоночник | 4.20 ± 0.55м, мх | 8.16 ± 0.75к | 6.44 ± 0.61к | 6.35 ± 0.56 |
| Отделы: | ||||
| – туловищный | 2.86 ± 0.53м | 5.13 ± 0.51к | 4.15 ± 0.42 | 4.33 ± 0.42 |
| – переходный | 0.49 ± 0.14 | 0.89 ± 0.14 | 0.58 ± 0.11 | 0.56 ± 0.11 |
| – хвостовой | 0.86 ± 0.20м | 2.13 ± 0.36к | 1.71 ± 0.25 | 1.47 ± 0.30 |
| Число разных видов аномалий* | ||||
| Незамкнутые невральные или гемальные дуги | 3.09 ± 0.50м | 5.95 ± 0.66к, мх, х | 4.33 ± 0.44м | 4.24 ± 0.44м |
| Деформированные позвонки | 0.49 ± 0.14м, мх | 1.45 ± 0.27к | 1.49 ± 0.19к | 1.00 ± 0.17 |
| Сращения дуг соседних позвонков | 0.17 ± 0.08м | 0.79 ± 0.18к, мх, х | 0.44 ± 0.09м | 0.37 ± 0.11м |
| Деформированные невральные или гемальные дуги | 0.23 ± 0.07 | 0.30 ± 0.08 | 0.30 ± 0.10 | 0.29 ± 0.07 |
| Отсутствие невральных или гемальных дуг | 0.11 ± 0.06 | 0.22 ± 0.06 | 0.27 ± 0.06х | 0.10 ± 0.04мх |
| Перемещение основания дуги на соседний позвонок | 0 | 0.013 ± 0.013 | 0.020 ± 0.014 | 0.012 ± 0.012 |
| Несращение дуги с телом позвонка | 0 | 0.053 ± 0.032 | 0.010 ± 0.010 | 0.023 ± 0.016 |
| Дополнительные невральные или гемальные дуги | 0.029 ± 0.029 | 0.053 ± 0.026 | 0.090 ± 0.029 | 0.081 ± 0.030 |
Примечание. Здесь и в табл. 2–4: К – контроль, М – воздействие магнитного поля, МХ – совместное воздействие магнитного поля и хлорофоса, Х – воздействие хлорофоса; к, м, мх, х значение достоверно (p < 0.05) отличается от соответствующего варианта; * среднее значение и его ошибка в расчёте на рыб с аномалиями.
Таблица 2.
Характеристика сращений позвонков у сеголеток плотвы Rutilus rutilus в контрольной и экспериментальных выборках
| Показатель | Вариант | |||
|---|---|---|---|---|
| К | М | МХ | Х | |
| Число* сращений | 1.07 ± 0.07м, мх, х | 1.88 ± 0.17к | 1.71 ± 0.14к | 1.65 ± 0.11к |
| Число* позвонков в сращениях: | ||||
| весь позвоночник | 2.57 ± 0.29м, мх, х | 4.69 ± 0.41к | 4.25 ± 0.38к | 4.15 ± 0.30к |
| отделы: | ||||
| – туловищный | 2.07 ± 0.32 | 3.21 ± 0.38 | 3.08 ± 0.29 | 3.15 ± 0.29 |
| – переходный | 0.21 ± 0.21 | 0.63 ± 0.12 | 0.40 ± 0.12 | 0.47 ± 0.12 |
| – хвостовой | 0.29 ± 0.19 | 0.85 ± 0.20 | 0.76 ± 0.21 | 0.53 ± 0.18 |
| Число позвонков в одном сращении | 2.40 | 2.49 | 2.49 | 2.51 |
| Доля* асимметричных сращений, % | 93.33 | 81.19 | 83.91 | 84.38 |
| Доля* рыб с асимметричными сращениями, %: | ||||
| – последовательными | 7.14 | 26.92 | 26.00 | 32.73 |
| – встречными | 0 | 17.31 | 16.00 | 7.27 |
| – с Веберовыми позвонками | 0 | 5.77 | 12.00 | 9.09 |
| Рыбы с асимметричным позвоночником: | ||||
| – число рыб, экз | 13 | 40 | 41 | 51 |
| – доля*, % | 92.86 | 76.92 | 82.00 | 92.73 |
| Разница в числе позвонков слева и справа: | ||||
| M ± m** | 1.08 ± 0.08 | 1.58 ± 0.15 | 1.46 ± 0.13 | 1.49 ± 0.11 |
| min–max | 1–2 | 1–5 | 1–4 | 1–4 |
| Общее число позвонков, включая Веберовы и преуральные**: | ||||
| – слева | 41.46 ± 0.24мх | 41.93 ± 0.23 | 42.27 ± 0.18к | 41.78 ± 0.14 |
| – справа | 41.92 ± 0.33 | 42.20 ± 0.20мх | 41.63 ± 0.19м | 41.88 ± 0.18 |
Примечание. Средние значения и их ошибка (M ± m), а также доли приведены в расчёте на рыб: * со сращениями позвонков, ** с асимметричным позвоночником. min–max – пределы варьирования показателя. Полужирным шрифтом выделены достоверно различающиеся (p < 0.05) значения числа позвонков слева и справа у рыб с асимметричным позвоночником.
Такие аномалии, как дополнительные невральные или гемальные дуги, перемещение основания дуги на соседний позвонок и несращение дуг с телом позвонка, в исследованных группах молоди встречались единично. Различия между исследованными выборками плотвы по этим трём типам аномалий, а также деформациям невральных или гемальных дуг, не выявлены (табл. 1).
Остальные нарушения в строении позвоночника у контрольных рыб встречались реже, чем у экспериментальных. При этом ближе всего к контролю вариант Х. В варианте М нарушения строения позвоночника были максимальными; по двум показателям (количеству незамкнутых невральных и гемальных дуг и числу сращений дуг соседних позвонков) рыбы с аномалиями из варианта М значимо отличались от сеголеток из групп МХ и Х (табл. 1).
Во всех группах в туловищном отделе позвоночника позвонки с аномалиями отмечались в два–три раза чаще, чем в хвостовом (табл. 1). На рис. 2а приведены схемы, отражающие долю рыб с аномалиями определённых позвонков скелета. Зона позвоночника с 12-го туловищного по 3-й хвостовой позвонок была охвачена нарушениями строения примерно с одинаковой интенсивностью во всех группах. В средних позвонках туловищного отдела в экспериментальных вариантах аномалии встречались чаще, чем в контроле. Заметно выделяется вариант М, где 35–40% рыб имели отклонения развития 7–12-го позвонков. В контрольной группе зарегистрировано меньше аномалий хвостового отдела по сравнению с вариантами М, МХ и Х, причём на 6, 8–11 и 13-м хвостовых позвонках нарушения не отмечены.
Рис. 2.
Доля в контрольной и экспериментальных выборках сеголеток плотвы Rutilus rutilus с аномалиями определённых позвонков в составе позвоночника: а – все аномалии (относительно
рыб с аномалиями), б – сращения (относительно рыб со сращениями); К – контроль, М
– воздействие магнитного поля, МХ – совместное воздействие магнитного поля и хлорофоса,
Х – воздействие хлорофоса. Доля, %: (◻) – 0, ( ) – 0.01–10.00, (
) – 0.01–10.00, ( ) – 10.01–20.00, (
) – 10.01–20.00, ( ) – 20.01–30.00, (◼) – 30.01–40.00. Цифрами обозначены номера позвонков. В рисунок не включены редкие
позвонки – 18–21-й туловищные, 5-й переходный и 17–18-й хвостовые.
) – 20.01–30.00, (◼) – 30.01–40.00. Цифрами обозначены номера позвонков. В рисунок не включены редкие
позвонки – 18–21-й туловищные, 5-й переходный и 17–18-й хвостовые.
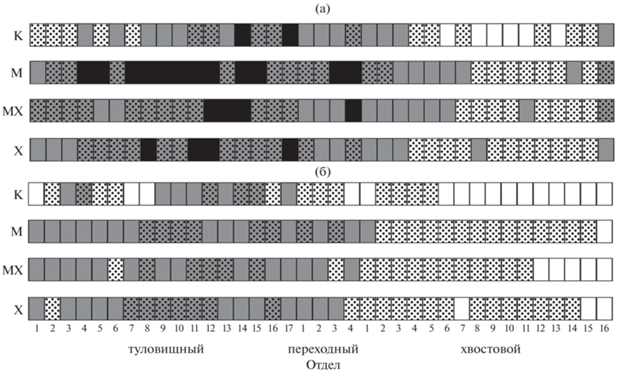
Сращения позвонков также чаще отмечались в туловищном отделе позвоночника (рис. 2б, табл. 2). Во всех исследованных группах сращения в хвостовом отделе позвоночника встречаются в основном не более чем у 10% рыб. В контроле в сращения входили хвостовые позвонки только со второго по пятый. В остальных группах рыб сращения позвонков также отмечались чаще в передней части хвостового отдела позвоночника. Сращения каждого из позвонков переходного отдела в контроле встречались у 5–10% рыб, в экспериментальных выборках доля рыб с такими нарушениями была выше. Каждый из туловищных позвонков (за исключением 6-го в варианте МХ и 2-го в варианте Х) наблюдался в составе сращений более чем у 10% рыб из групп М, Х и МХ. Наиболее часто в сращения входили 7–16-й туловищные позвонки. В контроле наибольшее число сращений приходилось на 4-й и 9–15-й туловищные позвонки.
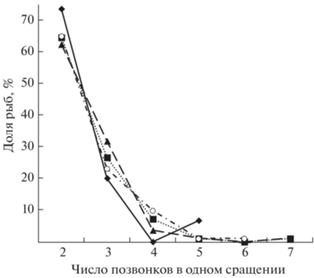
60–70% всех сращений во всех исследованных выборках – двухпозвонковые, около 20–30% – трёхпозвонковые (рис. 3). Сращения, в состав которых входит четыре и больше позвонков, относительно редки. В контрольной группе к таковым относится одно 5-позвонковое сращение. В выборке Х максимальное число позвонков в одном сращении – шесть, в группах М и МХ – семь.
Рис. 3.
Доля сеголеток плотвы Rutilus rutilus с разным числом позвонков в одном сращении: (─◆─) – контроль, (··◼··) – воздействие магнитного поля, (‒ –▲– –) – совместное воздействие магнитного поля и хлорофоса, (· –⚪– ·) – воздействие хлорофоса.
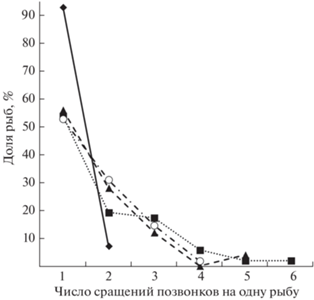
В экспериментальных выборках доля рыб со сращениями позвонков была в два и более раз выше, чем в контроле (табл. 3). Среднее число сращений и позвонков в их составе у рыб со сращениями позвонков в группах М, МХ и Х было значимо выше, чем в группе К (табл. 2). Поражение позвоночника у рыб со сращениями позвонков в экспериментальных вариантах было более выраженным: если в контроле из 14 рыб со сращениями у 13 регистрировали по одному сращению (92.5%), то в группах М, МХ и Х доля таких особей значительно ниже – 53–56%, а максимальное число сращений на одну особь достигает четырёх, пяти и шести в группах соответственно Х, МХ и М (рис. 4).
Таблица 3.
Сравнение морфологических показателей сеголеток плотвы Rutilus rutilus, сгруппированных по состоянию позвоночника
| Показатель | Группа | Вариант | |||
|---|---|---|---|---|---|
| К | М | МХ | Х | ||
| Доля рыб, % | Н | 57.32м, мх, х | 32.14к | 27.54к | 33.85к |
| А | 25.61 | 21.43мх | 36.23м, х | 23.85мх | |
| С | 17.07м, мх, х | 46.43к | 36.23к | 42.31к | |
| vert.a. | Все особи | 16.24 ± 0.07мх | 16.29 ± 0.08 | 16.46 ± 0.07к | 16.29 ± 0.06 |
| Н | 16.06 ± 0.07мх | 15.92 ± 0.07мх | 16.34 ± 0.10к, м, х | 16.02 ± 0.08мх | |
| А | 16.29 ± 0.12 | 16.17 ± 0.14 | 16.14 ± 0.08 | 16.00 ± 0.08 | |
| С | 16.79 ± 0.19 | 16.62 ± 0.15 | 16.86 ± 0.13 | 16.67 ± 0.11 | |
| vert.i. | Все особи | 2.95 ± 0.05м | 3.19 ± 0.06к, х | 3.06 ± 0.05 | 3.04 ± 0.05м |
| Н | 2.94 ± 0.07 | 3.08 ± 0.07 | 2.89 ± 0.08 | 2.89 ± 0.07 | |
| А | 2.86 ± 0.08 | 3.00 ± 0.10 | 3.08 ± 0.07 | 2.97 ± 0.10 | |
| С | 3.14 ± 0.18 | 3.35 ± 0.09 | 3.16 ± 0.10 | 3.20 ± 0.09 | |
| vert.c. | Все особи | 15.43 ± 0.06м | 15.75 ± 0.06к, мх, х | 15.55 ± 0.05м | 15.47 ± 0.06м |
| Н | 15.49 ± 0.09 | 15.67 ± 0.10х | 15.45 ± 0.09 | 15.34 ± 0.07м | |
| А | 15.38 ± 0.13м | 15.88 ± 0.09к, х | 15.68 ± 0.09 | 15.45 ± 0.11м | |
| С | 15.29 ± 0.13м | 15.75 ± 0.11к | 15.50 ± 0.10 | 15.58 ± 0.10 | |
| vert. | Все особи | 41.62 ± 0.08м, мх | 42.23 ± 0.10к, х | 42.07 ± 0.08к, х | 41.80 ± 0.08м, мх |
| Н | 41.49 ± 0.09 | 41.67 ± 0.11х | 41.68 ± 0.09х | 41.25 ± 0.09м, мх | |
| А | 41.52 ± 0.16м | 42.04 ± 0.14к, х | 41.90 ± 0.12х | 41.42 ± 0.11м, мх | |
| С | 42.21 ± 0.24 | 42.71 ± 0.18 | 42.52 ± 0.14 | 42.45 ± 0.13 | |
| Доля типичных фенотипов, % | Все особи | 90.24м, мх, х | 61.61к, мх, х | 75.36к, м | 80.77к, м |
| Н | 97.87 | 94.44 | 94.74 | 100.00 | |
| А | 90.48 | 75.00х | 80.00х | 96.77м, мх | |
| С | 64.29м | 32.69к, мх, х | 56.00м | 56.36м | |
Примечание. Группы сеголеток: Н – нормальные (без аномалий); А, С – с аномалиями соответственно без сращений и со сращениями позвонков; vert.a., vert.i., vert.c., vert. – число позвонков (среднее значение и его ошибка) в туловищном, переходном, хвостовом отделах и их общее число, включая Веберовы и преуральные. Полужирным шрифтом выделены достоверные (p < 0.05) отличия по числу позвонков группы С от Н или А.
Рис. 4.
Число сращений позвонков на одну рыбу среди сеголеток плотвы Rutilus rutilus со сращениями позвонков. Обозначения см. на рис. 3.
Во всех выборках > 80% сращений позвонков были асимметричными (табл. 2). В контроле была зарегистрирована только одна рыба с двумя последовательными сращениями, встречные сращения в этой группе не отмечены. В других вариантах рыбы с последовательными сращениями встречались чаще, чем со встречными, наибольшая разница отмечена в группе Х. В экспериментальных вариантах наблюдали ещё один вариант асимметричного строения позвоночника – сращение первого туловищного полупозвонка с последним Веберовым позвонком. Количество рыб с такой аномалией максимально в группе МХ.
У рыб с асимметричными сращениями различалось число позвонков при подсчёте слева и справа. Средние значения этой разницы представлены в табл. 2. Значимые различия между группами по этому показателю не выявлены. Однако следует обратить внимание, что доля рыб с разницей более чем в один позвонок в группе К составляла 7.69% (одна рыба из 13), а в группах М, МХ и Х – соответственно 35.00, 31.71 и 35.29%. В трёх последних группах максимальная разница между сторонами позвоночника составляла 4–5 позвонков.
По числу позвонков слева значимо различались варианты К и МХ, справа – М и МХ. У рыб из варианта МХ наблюдалась направленная асимметрия позвоночника – число позвонков слева на 0.64 позвонка в среднем больше, чем справа (табл. 2).
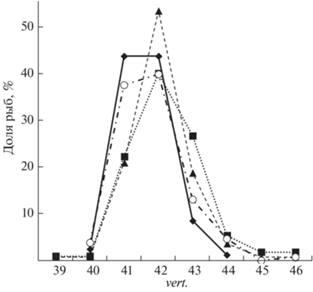
Число позвонков. Общее число позвонков у рыб в вариантах К и Х было значимо меньше, чем в группах М и МХ (табл. 3). Варианты К и Х близки также по распределению общего числа позвонков среди рыб в выборке, в то время как в группах МХ и М была повышена доля рыб соответственно с 42 и 43 позвонками. В экспериментальных вариантах единично отмечались редкие фенотипы – 39, 45 и 46 позвонков (рис. 5). Увеличение общего числа позвонков произошло в группе МХ за счёт туловищного отдела позвоночника, а в группе М за счёт переходного и хвостового отделов.
Рис. 5.
Распределение сеголеток плотвы Rutilus rutilus по общему числу позвонков (vert.). Обозначения см. на рис. 3.
Чтобы выяснить возможную взаимосвязь между нарушениями развития позвоночника и числом позвонков, выборки разделили на три подгруппы – рыбы без аномалий (нормальные), рыбы с аномалиями позвонков, но без сращений (аномальные) и рыбы со сращениями. Значимые различия между нормальными и аномальными рыбами из одной и той же экспериментальной группы не обнаружены. Однако у рыб со сращениями оказалось больше общее число позвонков и число позвонков в туловищном отделе позвоночника. Переходный отдел у рыб со сращениями содержал в среднем больше позвонков по сравнению с рыбами без сращений, но статистически значимые различия отмечены только для некоторых групп (табл. 3).
Позвонковые фенотипы. Во всех экспериментальных группах было зарегистрировано повышение разнообразия позвонковых фенотипов, оценённого с помощью показателя Шеннона, причём наибольший эффект отмечался в варианте М (Krylov et al., 2016).
Типичный для плотвы позвонковый фенотип, на основании наших данных, включает 15–17 туловищных, 2–4 переходных и 14–16 хвостовых позвонков при общем их числе в позвоночнике 40–42 (с учётом Веберовых и преуральных). В естественных популяциях доля таких рыб составляет >95% (Чеботарева, Изюмов, 2021). В данном эксперименте максимальная доля типичных фенотипов отмечалась в контрольной группе. Аномалии развития позвоночника влияют на позвонковый фенотип плотвы. В подгруппах нормальных рыб из всех вариантов воздействия доля типичных фенотипов составляла > 94%. Этот показатель был немного ниже у аномальных сеголеток, а у молоди со сращениями позвонков таких фенотипов значительно меньше – 32–64%. Среди нормальных и аномальных рыб максимальное количество типичных позвонковых фенотипов зарегистрировано в группе Х, а среди сеголеток со сращениями максимальное количество редких фенотипов встречалось в варианте М (табл. 3).
Встречаемость отдельных позвонковых фенотипов в исследуемых группах различна и в ряде случаев отличается от таковой у молоди из прибрежья Рыбинского водохранилища (Чеботарева, Изюмов, 2021) (табл. 4). В вариантах К и Х преобладает фенотип 16–3–15 (самый распространённый у природных сеголеток плотвы). Фенотип 16–3–16 встречался у наибольшего количества рыб из вариантов М и МХ; в группах К и Х он отмечался с близкой частотой; при этом для природной плотвы он менее характерен. Распространённый в природных выборках фенотип 17–2–15 относительно редок у молоди из нашего эксперимента. В число распространённых сочетаний позвонков в экспериментальных группах входит фенотип 17–3–16; в группе М выделяется фенотип 16–4–16. Эти варианты не входят в число типичных, а в природных популяциях встречаются редко и единично. Фенотип 17–4–15, отмеченный во всех экспериментальных выборках, у молоди Рыбинского водохранилища обнаружен не был. В то же время фенотип 16–3–14, нередко встречавшийся у природной молоди, отмечен единично только в варианте М.
Таблица 4.
Встречаемость самых распространённых позвонковых фенотипов сеголеток плотвы Rutilus rutilus, %
| Фенотип (общее число позвонков*) |
Рыбинское вдхр. | Вариант | |||||||
|---|---|---|---|---|---|---|---|---|---|
| К | М | МХ | Х | ||||||
| все особи | С | все особи | С | все особи | С | все особи | С | ||
| 16–3–15 (41) | 40.00 | 34.151 | 21.431 | 10.713 | 5.77 | 11.592 | 2.00 | 26.151 | 9.09 |
| 17–2–15 (41) | 10.62 | 2.44 | 0.89 | 1.92 | 2.90 | 2.00 | 2.31 | 1.82 | |
| 16–3–16 (42) | 9.69 | 24.392 | 26.791 | 7.693 | 26.091 | 10.002 | 20.002 | 14.551 | |
| 17–3–15 (42) | 6.31 | 6.10 | 21.431 | 2.68 | 5.77 | 11.592 | 18.001 | 6.92 | 10.913 |
| 17–2–16 (42) | 5.38 | 7.323 | 7.143 | 4.46 | 3.85 | 7.253 | 10.002 | 3.85 | 3.64 |
| 16–3–14 (40) | 4.92 | 0.89 | |||||||
| 17–3–14 (41) | 3.54 | 2.44 | 0.89 | 1.92 | 0.72 | 0.77 | |||
| 16–4–15 (42) | 2.92 | 3.66 | 14.292 | 3.57 | 1.92 | 7.253 | 8.003 | 7.693 | 10.91 |
| 16–4–16 (43) | 0.31 | 6.25 | 11.542 | 1.45 | 4.00 | 2.31 | 5.45 | ||
| 17–3–16 (43) | 0.46 | 3.66 | 7.143 | 12.502 | 19.231 | 7.253 | 8.003 | 5.38 | 12.732 |
| 17–4–15 (43) | 1.22 | 7.143 | 3.57 | 7.693 | 2.17 | 6.00 | 1.54 | 3.64 | |
Примечание. * Включая Веберовы и преуральные. С – сеголетки со сращениями позвонков; 1, 2, 3 соответственно первые, вторые и третьи по численности фенотипы в группах. Полужирным шрифтом выделены позвонковые фенотипы, распространённые у сеголеток из прибрежья Рыбинского водохранилища (по: Чеботарева, Изюмов, 2021).
Встречаемость позвонковых фенотипов среди рыб со сращениями отличается от той, что наблюдается в полных выборках. В группе К преобладают фенотипы 16–3–15 и 17–3–15, в М – 17–3–16, в МХ – 17–3–15, в Х – 16–3–16 (табл. 4).
ОБСУЖДЕНИЕ
У исследованных сеголеток плотвы отмечены аномалии позвоночника, аналогичные ранее описанным для этого и других видов рыб (Татарко, 1977; Boglione et al., 2001, 2006, 2013; Yershov, 2008; Lewis-McCrea, Lall, 2010; Bogutskaya et al., 2011; Есин, 2015; Яблоков, 2018). В состоянии позвоночника молоди плотвы во всех вариантах эксперимента (включая контроль) выявлены общие тенденции: 1) наибольшее число аномалий позвонков локализовано в туловищном отделе позвоночника; 2) самые распространённые аномалии – незамкнутые невральные или гемальные дуги, деформации тел позвонков, а также сращения тел позвонков; 3) большинство сращений позвонков – асимметричные; 4) у рыб со сращениями позвонков наблюдается увеличение общего числа позвонков, а также их числа в туловищном отделе позвоночника; 5) число позвонков в хвостовом отделе позвоночника не зависит от наличия аномалий как в самом хвостовом отделе, так и в позвоночнике в целом; 6) позвонковые фенотипы рыб без аномалий позвоночника в основном были типичными для плотвы и широко распространёнными также в природных популяциях.
Отмеченные особенности аномалий позвоночника у плотвы в основном аналогичны ранее описанным (Чеботарева, 2009; Чеботарева и др., 2016), в том числе и большее число нарушений развития позвонков туловищного отдела позвоночника по сравнению с хвостовым. При этом наибольшие различия наблюдаются у рыб в контроле. У природных сеголеток ситуация другая: при общей невысокой частоте аномалий они чаще отмечаются в хвостовом отделе позвоночника. Аномалии осевого скелета часто отражают общую несбалансированность эмбриогенеза. Формирование туловищного отдела осевого скелета взаимосвязано с бóльшим числом других жизненно важных структур, чем хвостового отдела, в частности, с плавательным пузырём и кишечником. Бóльшая сложность морфогенеза туловищного отдела позвоночника, вероятно, приводит к относительно большему числу ошибок. При этом дефекты плавательного пузыря и пищеварительной системы являются одними из главных причин гибели личинок при переходе к активному образу жизни и внешнему питанию (Владимиров, Семенов, 1959). У личинок с нелетальными нарушениями жизненно важных систем организма больше шансов выжить в щадящих условиях эксперимента, особенно в контроле, поэтому, как мы предполагаем, экспериментальные рыбы отличаются относительно высоким числом аномалий именно туловищного отдела позвоночника.
Молодь из экспериментальной контрольной группы отличается от природной плотвы не только локализацией, но и количеством аномалий позвоночника. У сеголеток из Рыбинского водохранилища дефекты развития позвонков встречаются намного реже, особенно существенна разница по частоте сращений позвонков: среди природной молоди доля рыб со сращениями не превышает 5% (Чеботарева, Изюмов, 2021), тогда как у контрольной плотвы в нашей работе она составляет 17%. В то же время молодь из контрольной группы по многим исследованным показателям позвоночника была ближе к рыбам из природных популяций, чем к сеголеткам из экспериментальных групп. В первую очередь это касается частоты и разнообразия позвонковых фенотипов (Чеботарева и др., 2016).
Во всех исследованных экспериментальных группах увеличивалось количество и тяжесть аномалий позвоночника по сравнению с контролем. В двух вариантах воздействия магнитного поля наблюдалось также увеличение общего числа позвонков. Максимальные отдалённые эффекты воздействия на эмбриональное развитие плотвы выявлены для магнитного поля, действующего отдельно. Сочетание магнитного поля и хлорофоса вызвало менее выраженные изменения позвоночника сеголеток плотвы. Снижение отрицательных эффектов в случае совместного действия двух факторов может быть связано с замедленным темпом роста молоди в варианте МХ. Стандартная длина тела сеголеток в группе МХ (67.8 ± 0.2 мм) была значимо меньше, чем в контроле (71.8 ± 0.3 мм), а также в группах М и Х (соответственно 78.0 ± 0.3 и 73.9 ± 0.3 мм) (Krylov et al., 2016). Разнонаправленные ответы молоди из разных вариантов на стресс могут быть проявлением двух стратегий защиты организма (Михайленко, 2002): активной – повышение интенсивности метаболизма, направленного на репарацию повреждений и восстановление гомеостаза (варианты М и Х), и пассивной – ограничение реакции организма на экстремальные воздействия, снижение скорости возникновения и распространения повреждений (вариант МХ). Похожий эффект неблагоприятных воздействий – меньшее число нарушений развития позвоночника при сниженном темпе роста плотвы – мы наблюдали ранее после воздействия на эмбрионы хлорофоса и комбинации магнитного поля с повышенной температурой (23°С) (Чеботарева, 2009; Чеботарева и др., 2016).
Формирование осевого скелета плотвы на ранних этапах её онтогенеза очень чувствительно к воздействиям факторов различной природы. При этом, как показала наша работа, низкочастотное магнитное поле может вызывать более значительные нарушения развития позвоночника, чем токсиканты, что подчёркивает важность не только отсутствия химического загрязнения, но и ненарушенного магнитного фона для нормального эмбриогенеза рыб.