Кунаширский пролив расположен между восточным побережьем о-ва Хоккайдо (Япония) и о-вом Кунашир (Россия). Вместе с проливом Измены Кунаширский пролив формирует общую акваторию, которую в Японии называют прол. Немуро – самый южный пролив, соединяющий Охотское море и Тихий океан. Северная часть пролива наиболее глубоководная, глубины здесь достигают 2000 м, тогда как южная его часть у п-ова Немуро относительно мелководна, с глубинами <100 м и преобладанием песчаных и илистых отложений. Наиболее узкая и мелководная часть этого пролива расположена между Хоккайдо (п-ов Ноцуке) и южной частью о-ва Кунашир. Расстояние между островами на этом участке составляет ~16 км, а глубины <20 м (Shinohara et al., 2012). Большое влияние на гидрологический режим Кунаширского пролива оказывает тёплое течение Соя, проходящее вдоль северного побережья Хоккайдо и являющееся ветвью Цусимского течения. У северной оконечности п-ова Сиретоко (северо-восток Хоккайдо) течение Соя разделяется на три ветви, первая из которых проникает в Кунаширский пролив (Анцулевич, Бобков, 1992). В зимний период этот пролив почти полностью покрывается дрейфующим из Охотского моря льдом. Максимальное ледовое покрытие наблюдается в марте, но в апреле лёд активно разрушается (Yoshida, 1989; Ким Сен Ток, Бирюков, 2009). В течение года температура воды на глубине 50 м около п-ова Сиретоко может изменяться от −1.7 до 20.5°С (Nobetsu et al., 1998).
В современный период в Кунаширском проливе осуществляют многолетний промысел таких коммерчески важных видов, как минтай Theragra chalcogramma, треска Gadus macrocephalus, камбалы (семейство Pleuronectidae), южный однопёрый терпуг Pleurogrammus azonus, тихоокеанские лососи (род Oncorhynchus), а также других морских рыб и гидробионтов (Yoshida, 1989; Mizushima, Torisawa, 2005; Буслов и др., 2013).
В биогеографическом плане южная часть Охотского моря, прилегающая к побережью Сахалина, Хоккайдо и южным Курильским островам, включая Кунаширский пролив, относится к Японской низкобореальной подобласти (Шунтов, 1985). Контрастные условия морской среды этого пролива, географические и океанологические, способствуют формированию высокого видового богатства рыб, встречающихся в пределах рассматриваемой акватории. Первые отечественные сведения о видовом составе рыб из промысловых уловов у Южных Курил, включая Кунаширский пролив, появились ещё в конце 1940-х гг. (Веденский, 1949). Однако до сих пор морская ихтиофауна района исследований остаётся слабоизученной. Лишь недавно опубликован предварительный список морских видов рыб пролива Немуро (Shinohara et al., 2012).
Значительно больше внимания уделяли исследованиям видового состава различных сообществ рыб, встречающихся в бо́льших по площади соседних акваториях. В частности, опубликована работа, обобщающая многолетние исследования эпипелагического сообщества рыб и головоногих моллюсков прикурильских вод Тихого океана по данным комплексных экспедиций ТИНРО-центра (Иванов, 1998). Дополнительные сведения о составе нектонных сообществ и межгодовой изменчивости миграций этих рыб и кальмаров в тихоокеанских водах южных Курильских о-вов были получены в исследованиях с применением дрифтерных сетей (Савиных, 1998; Савиных и др., 2003). Опубликован также видовой состав рыб у южных Курильских о-вов по данным донных траловых съёмок 1980–2000-х гг. с разделением на охотоморский и океанский районы (Ким Сен Ток, Бирюков, 2009). Определённые сведения о распространении и встречаемости отдельных видов рыб в проливе Немуро можно найти в монографии Мицусимы и Торисавы (Mizushima, Torisawa, 2005), в аннотированном списке рыб дальневосточных морей (Борец, 2000) и в других публикациях.
В умеренной зоне дальневосточных морей как в целом, так и в её отдельных районах наибольшее видовое богатство характерно для демерсальной ихтиофауны (Линдберг, 1959; Лапко, 1996; Борец, 1997, 2000; Дылдин и др., 2020). Не являются исключением в этом отношении Кунаширский пролив и пролив Немуро в целом. В частности, в предварительном списке рыб, указанных для этого района (Shinohara et al., 2012), подавляющее большинство видов (>86%) относится к демерсальной группе. Поэтому повышенное внимание к исследованиям состава донных и придонных видов рыб рассматриваемого района вполне оправданно.
Изучение ихтиофауны различных водоёмов и акваторий важно по многим причинам, в том числе для оценки состояния водных экосистем. Биологическое разнообразие широко признано в качестве одного из краеугольных камней для оценки здоровья экосистем (Worm et al., 2006). Статусные оценки биоразнообразия позволяют проводить сравнение в пространственных или временны́х масштабах, часто в ассоциации с некоторыми внешними факторами, такими как физические или биогеохимические условия (Cochrane et al., 2016). Усиливающаяся антропогенная нагрузка на акватории южных Курильских о-вов – рыболовство (с использованием донных тралов, снюрреводов, сетей), а также судоходство и так далее (Буслов и др., 2013) – обусловливает особую актуальность исследования сообществ демерсальных рыб в Кунаширском проливе.
Цель нашего исследования – изучить видовой состав рыб из промысловых уловов донных сетей в 1998–2014 гг. в этом районе и сравнить результаты с данными по прилегающим акваториям, сопоставимым по площади и глубинам.
МАТЕРИАЛ И МЕТОДИКА
В основу исследований положены материалы по видовому составу рыб, собранные при пробном промысле минтая и южного однопёрого терпуга ставными донными сетями на японских рыболовных судах в российской (восточной) зоне Кунаширского пролива в 1998–2014 гг. Промысел этих двух видов в указанном районе проводится на основании Соглашения между правительствами Российской Федерации и Японии от 1998 г.
В соответствии с Соглашением в начальный период лова каждого из этих объектов в течение двух недель осуществляли так называемый “пробный” промысел. В этот период специалисты СахНИРО по приглашению японской стороны ежегодно проводили наблюдения и сбор материалов по промыслу минтая и терпуга, их биологическим показателям. Одной из важнейших задач этих наблюдений являлась оценка видового состава объектов прилова. Специалисты СахНИРО выполняли функции наблюдателей, находясь на флагманском судне как при промысле минтая, так и терпуга. Сбор материалов проводил один наблюдатель непосредственно на борту судна во время подъёма сетей на борт, при сортировке и раскладке улова в тару.
Пробный лов минтая осуществляли во второй половине января, а терпуга в основном в конце сентября–первой половине октября, иногда до начала ноября. В целом для сетного промысла этих двух видов характерны заметные различия не только по сезонам лова, но и по границам районов добычи, глубинам выставления сетей, числу сетей, параметрам и размерам ячеи сетного полотна, времени застоя сетей. В частности, границы промысла терпуга были немного севернее и восточнее, глубины выставления сетей меньше (75–220 против 70–315 м), а время застоя сетей обычно было существенно меньше, чем при добыче минтая (4.0–8.5 против 5.0–95.0 часов), так же, как и размеры ячеи (35 × 35 против 48 × 48 мм). Все приведённые различия обусловлены особенностями биологии этих видов рыб в указанные сезоны года. На рис. 1 приведены схемы районов постановки донных сетей в восточной части Кунаширского пролива при промысле минтая и терпуга.
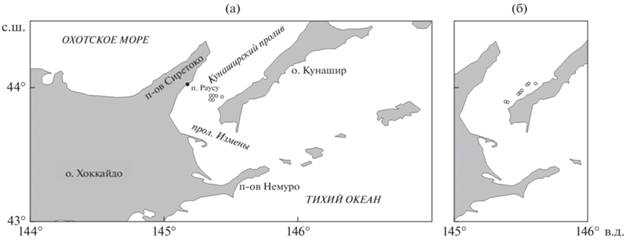
Идентификацию видов рыб осуществляли в соответствии с определительными ключами, изложенными в отечественных и японских источниках (Линдберг, Красюкова, 1975, 1987; Masuda et al., 1984). Латинские видовые названия рыб в основном приведены в соответствии с последними таксономическими ревизиями (Fricke et al., 2022). Однако название минтая мы оставили в традиционном биномене Theragra chalcogramma, которого придерживаются российские учёные в связи с преждевременностью изменения родового названия этого вида (Булатов, 2014; Stroganov, 2015; Дылдин и др., 2020). Названия семейств и порядок их расположения приведены по Нельсону (2009).
Общую статистическую обработку материалов выполнили с использованием компьютерной программной среды “Excel”. Схемы районов лова японскими судами в российской зоне Кунаширского пролива построили в программе “Surfer”.
Для сравнения видового состава рыб из разных районов использовали индекс сходства Чекановского–Сёренсена (Ics) в модификации для качественных данных (Песенко, 1982): Ics = 2а/(b + с), где b и с – число видов в двух сравниваемых фаунистических списках, а – число общих видов в этих списках.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
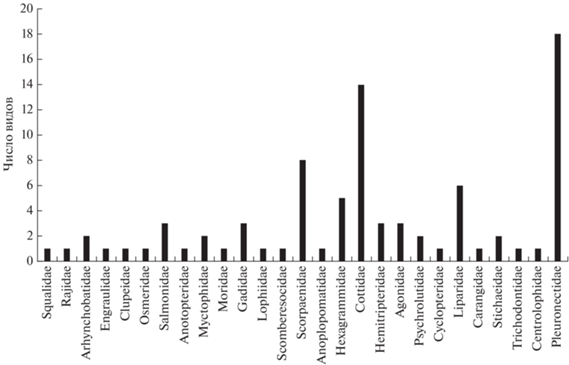
В 1998–2014 гг. в Кунаширском проливе в уловах донных сетей выявлено 85 видов рыб, принадлежащих 56 родам, 27 семействам и 13 отрядам (таблица). Соотношение числа видов в составе разных семейств в районе наблюдений показано на рис. 2. Наиболее разнообразными по числу видов оказались семейства Pleuronectidae (18 видов) и Cottidae (14). Заметными в этом отношении были семейства Scorpaenidae (8), Hexagrammidae (5) и Liparidae (6). Остальные семейства были представлены одним–тремя видами.
Видовой состав рыб в уловах донных сетей в российской (восточной) части Кунаширского пролива (объединённые данные за 1998–2014 гг.)
| Вид | Род | Семейство | Отряд |
|---|---|---|---|
| Squalus suckleyi (Girard, 1855)* | Squalus | Squalidae | Squaliformes |
| Beringraja pulchra (Liu, 1932) | Beringraja | Rajidae | Rajiformes |
| Bathyraja aleutica (Gilbert, 1896)* | Bathyraja | Arhynchobatidae | То же |
| Bathyraja parmifera (Bean, 1881)* | То же | То же | » |
| Engraulis japonicus Temminck et Schlegel, 1846 | Engraulis | Engraulidae | Clupeiformes |
| Clupea pallasii Valenciennes, 1847 | Clupea | Clupeidae | То же |
| Osmerus dentex Steindachner et Kner, 1870 | Osmerus | Osmeridae | Osmeriformes |
| Oncorhynchus keta (Walbaum, 1792)* | Oncorhynchus | Salmonidae | Salmoniformes |
| Oncorhynchus masou (Brevoort, 1856)* | То же | То же | То же |
| Oncorhynchus tshawytscha (Walbaum, 1792)* | » | » | » |
| Anotopterus nikparini Kukuev, 1998* | Anotopterus | Anotopteridae | Aulopiformes |
| Lampanyctus jordani Gilbert, 1913* | Lampanyctus | Myctophidae | Myctophiformes |
| Lampanyctus regalis (Gilbert, 1892)* | То же | То же | То же |
| Physiculus japonicus Hilgendorf, 1879* | Physiculus | Moridae | Gadiformes |
| Gadus macrocephalus Tilesius, 1810* | Gadus | Gadidae | То же |
| Eleginus gracilis (Tilesius, 1810) | Eleginus | То же | » |
| Theragra chalcogramma (Pallas, 1814) | Theragra | » | » |
| Lophiomus setigerus (Vahl, 1797)* | Lophiomus | Lophiidae | Lophiiformes |
| Cololabis saira (Brevoort, 1856) | Cololabis | Scomberesocidae | Beloniformes |
| Sebastes alutus (Gilbert, 1890)* | Sebastes | Scorpaenidae | Scorpaeniformes |
| Sebastes glaucus Hilgendorf, 1880 | То же | То же | То же |
| Sebastes itinus (Jordan et Starks, 1904)* | » | » | » |
| Sebastes owstoni (Jordan et Thompson, 1914)* | » | » | » |
| Sebastes schlegelii Hilgendorf, 1880 | » | » | » |
| Sebastes steindachneri Hilgendorf, 1880* | » | » | » |
| Sebastes taczanowskii Steindachner, 1880 | » | » | » |
| Sebastes trivittatus Hilgendorf, 1880 | » | » | » |
| Anoplopoma fimbria (Pallas, 1814) | Anoplopoma | Anoplopomatidae | » |
| Hexagrammos lagocephalus (Pallas, 1810) | Hexagrammos | Hexagrammidae | » |
| Hexagrammos otakii Jordan et Starks, 1895 | То же | То же | » |
| Hexagrammos stelleri Tilesius, 1810 | » | » | » |
| Pleurogrammus azonus Jordan et Metz, 1913 | Pleurogrammus | » | » |
| Pleurogrammus monopterygius (Pallas, 1810) | То же | » | » |
| Alcichthys elongatus (Steindachner, 1881)* | Alcichthys | Cottidae | » |
| Enophrys diceraus (Pallas, 1787) | Enophrys | То же | » |
| Gymnocanthus detrisus Gilbert et Burke, 1912 | Gymnocanthus | » | » |
| Gymnocanthus herzensteini Jordan et Starks, 1904 | То же | » | » |
| Gymnocanthus pistilliger (Pallas, 1814)* | » | » | » |
| Hemilepidotus gilberti Jordan et Starks, 1904 | Hemilepidotus | » | » |
| Hemilepidotus papilio (Bean, 1880)* | То же | » | » |
| Icelus cataphractus (Pavlenko, 1910) | Icelus | » | » |
| Myoxocephalus brandtii (Steindachner, 1867) | Myoxocephalus | » | » |
| Myoxocephalus jaok (Cuvier, 1829) | То же | » | » |
| Myoxocephalus polyacanthocephalus (Pallas, 1814) | » | » | » |
| Myoxocephalus stelleri Tilesius, 1811 | » | » | » |
| Taurocottus bergii Soldatov et Pavlenko, 1915 | Taurocottus | » | » |
| Triglops jordani (Jordan et Starks, 1904)* | Triglops | » | » |
| Blepsias bilobus Cuvier, 1829 | Blepsias | Hemitripteridae | » |
| Blepsias cirrhosus (Pallas, 1814) | То же | То же | » |
| Hemitripterus villosus (Pallas, 1814) | Hemitripterus | » | » |
| Podothecus sachi (Jordan et Snyder, 1901) | Podothecus | Agonidae | » |
| Podothecus sturioides (Guichenot, 1869) | То же | То же | » |
| Tilesina gibbosa Schmidt, 1904 | Tilesina | » | » |
| Dasycottus setiger Bean, 1890 | Dasycottus | Psychrolutidae | » |
| Eurymen gyrinus Gilbert et Burke, 1912 | Eurymen | То же | » |
| Aptocyclus ventricosus (Pallas, 1769) | Aptocyclus | Cyclopteridae | » |
| Careproctus colletti Gilbert, 1896* | Careproctus | Liparidae | » |
| Careproctus rastrinus Gilbert et Burke, 1912* | То же | То же | » |
| Careproctus roseofuscus Gilbert et Burke, 1912* | » | » | » |
| Crystallias matsushimae Jordan et Snyder, 1902* | Crystallias | » | » |
| Crystallichthys mirabilis Jordan et Gilbert, 1898* | Crystallichthys | » | » |
| Liparis ochotensis Schmidt, 1904* | Liparis | » | » |
| Seriola quinqueradiata Temminck et Schlegel, 1845* | Seriola | Carangidae | Perciformes |
| Stichaeus grigorjewi Herzenstein, 1890 | Stichaeus | Stichaeidae | То же |
| Stichaeus nozawae Jordan et Snyder, 1902 | То же | То же | » |
| Arctoscopus japonicus (Steindachner, 1881) | Arctoscopus | Trichodontidae | » |
| Hyperoglyphe japonica (Döderlein, 1884)* | Hyperoglyphe | Centrolophidae | » |
| Atheresthes evermanni Jordan et Starks, 1904 | Atheresthes | Pleuronectidae | Pleuronectiformes |
| Glyptocephalus stelleri (Schmidt, 1904) | Glyptocephalus | То же | То же |
| Microstomus achne (Jordan et Starks, 1904)* | Microstomus | » | » |
| Hippoglossus stenolepis Schmidt, 1904* | Hippoglossus | » | » |
| Reinhardtius hippoglossoides (Walbaum, 1792) | Reinhardtius | » | » |
| Verasper moseri Jordan et Gilbert, 1898 | Verasper | » | » |
| Acanthopsetta nadeshnyi Schmidt, 1904 | Acanthopsetta | » | » |
| Cleisthenes pinetorum Jordan et Starks, 1904* | Cleisthenes | » | » |
| Eopsetta grigorjewi (Herzenstein, 1890)* | Eopsetta | » | » |
| Hippoglossoides dubius Schmidt, 1904 | Hippoglossoides | » | » |
| Hippoglossoides elassodon Jordan et Gilbert, 1880* | То же | » | » |
| Hippoglossoides robustus Gill et Townsend, 1897* | » | » | » |
| Lepidopsetta polyxystra Orr et Matarese, 2000* | Lepidopsetta | » | » |
| Lepidopsetta mochigarei Snyder, 1911 | То же | » | » |
| Limanda punctatissima (Steindachner, 1879) | Limanda | » | » |
| Platichthys stellatus (Pallas, 1787) | Platichthys | » | » |
| Pseudopleuronectes herzensteini (Jordan et Snyder, 1901) | Pseudopleuronectes | » | » |
| Pseudopleuronectes schrenki (Schmidt, 1904) | То же | » | » |
| Всего: 85 | 56 | 27 | 13 |
Рис. 2.
Число видов рыб разных семейств по уловам донных сетей в российской части Кунаширского пролива в 1998–2014 гг.
Эпипелагические виды, например, такие как японский анчоус Engraulis japonicus, кета O. keta и некоторые другие, в уловах отмечали не ежегодно. Вероятно, они попадали в донные сети при их постановке или подъёме. Однако мы не исключили эти виды из общего списка, тем более что диапазон глубин их встречаемости (Ohshimo, Hamatsu, 1997; Ishida et al., 2001; Великанов и др., 2016) частично совпадает с глубинами лова донными сетями в районе наблюдений.
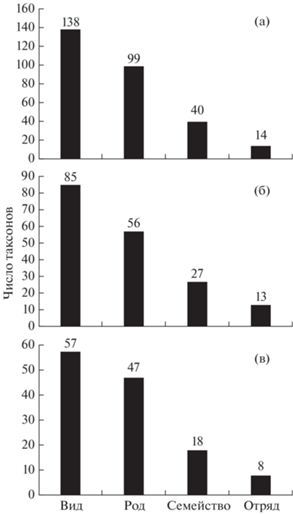
Сравнение числа видов из уловов рыб донными сетями в российской зоне Кунаширского пролива со сведениями из литературы показало следующее (рис. 3). По данным Синохары с соавторами (Shinohara et al., 2012), предварительный список морских рыб в проливе Немуро представлен 138 видами, принадлежащими 99 родам, 40 семействам и 14 отрядам. Список рыб для российской части Кунаширского пролива на 53 вида меньше. В то же время для обоих списков характерно определённое сходство: индекс Чекановского–Сёренсена между ними составил 0.45.
Рис. 3.
Число видов, родов, семейств и отрядов рыб: а – прол. Немуро (по: Shinohara et al., 2012), б – российская зона Кунаширского пролива, в – Юго-Восточный Хоккайдо, район Дото (по: Yamamura, 2003).
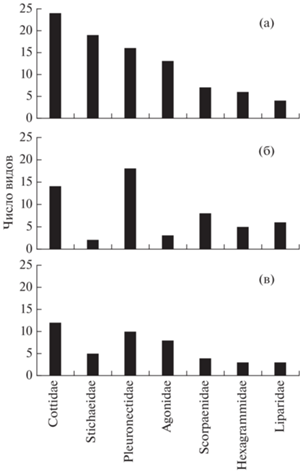
В предварительном списке, как и в нашем, для большинства семейств характерно наличие одного–шести видов, а доминирующее положение по числу видов занимали Cottidae (24 вида), Stichaeidae (19), Pleuronectidae (16) и Agonidae (13) (рис. 4). В районе наших исследований отмечено меньше видов в сумме, меньше видов в отдельных семействах, а также меньше доминирующих семейств – только два. Это, вероятно, обусловлено рядом ограничивающих факторов – использование только одного типа орудий лова (донные сети), применение двух сравнительно крупных размеров ячеи сетного полотна, лов в определённом диапазоне глубин и другие. В частности, в российской зоне Кунаширского пролива донные сети в период наших наблюдений использовали только на глубинах от 70–75 до 315 м. Следовательно, верхняя часть шельфа этого района, как и более глубоководные участки свала, остались не обследованными. В то же время зональное распределение видовых сообществ демерсальных рыб в зависимости от глубины ранее было установлено для многих шельфовых районов и присваловых участков морей (Horikawa, Toiyama, 1985; Bianchi, 1992; Yamamura et al., 1993; Fujita et al., 1995; Колпаков, 2004; Ким Сен Ток, 2005).
Рис. 4.
Число видов рыб в доминирующих по видовому разнообразию семействах в проливе Немуро (а), российской зоне Кунаширского пролива (б) и у Юго-Восточного Хоккайдо, район Дото (в).
Тем не менее в нашем списке представлено 35 видов рыб, которые отсутствуют в предварительном списке для пролива Немуро. Это представители семейств Squalidae (1 вид), Arhynchobatidae (2), Salmonidae (3), Anotopteridae (1), Myctophidae (2), Moridae (1), Gadidae (1), Lophiidae (1), Scorpaenidae (4), Cottidae (4), Liparidae (6), Carangidae (1), Centrolophidae (1), Pleuronectidae (7). Отметим, что проникновение и нахождение всех 35 видов рыб, зафиксированных в таблице, не является случайным в восточной части Кунаширского пролива. Обзор данных литературы выявил, что все эти виды имеют широкое распространение и регулярно встречаются во всех морских акваториях, сопредельных с районом наших исследований – южной части Охотского моря, прибрежье Северного Хоккайдо, тихоокеанских водах у Хоккайдо и южных Курильских о-вов (Борец, 2000; Mizushima, Torisawa, 2005; Ким Сен Ток, Бирюков, 2009; Великанов и др., 2016). В связи с этим можно обоснованно предположить, что все неописанные (новые) виды для предварительного списка рыб пролива Немуро либо постоянно обитают в районе лова сетями (преимущественно демерсальные рыбы), либо совершают в район наблюдений сезонные миграции (прежде всего эпипелагические). Соответственно, они могли регулярно появляться в проливе, в том числе в его восточной части, что и подтверждают результаты многолетней практики ловов донными сетями.
Таким образом, в целом список видов рыб, встречающихся в Кунаширском проливе и проливе Немуро, может быть увеличен до 173. Однако и эта цифра не является окончательной, так как немало видов рыб, распространённых в сопредельных с Кунаширским проливом акваториях, рано или поздно могут быть обнаружены в рассматриваемом районе, в том числе дальневосточная мойва Mallotus catervarius, большая корифена Coryphaena hippurus, горбуша O. gorbuscha, дальневосточная сардина Sardinops melanostictus и другие (Великанов, 1986, 2010; Борец, 2000; Yamamura, 2003; Mizushima, Torisawa, 2005; Ким Сен Ток, Бирюков, 2009; Буслов и др., 2013; Филатов, 2015).
В частности, по данным Ямамуры (Yamamura, 2003), на верхней части шельфа (глубины 33–116 м) Юго-Восточного Хоккайдо, от м. Эримо до м. Носаппу (район Дото), при облове донными сетями (ячея 21–121 мм) выявлено 57 видов рыб, принадлежащих 47 родам и 18 семействам (рис. 3). В этом районе в уловах преобладали демерсальные виды рыб, хотя встречались и эпипелагические (Clupea pallasii, S. melanostictus, E. japonicus и другие). При этом доминирующими по числу видов были семейства Cottidae (12), Pleuronectidae (10) и Agonidae (8) (рис. 4). Видовой состав рыб и доминирующих семейств в районе Дото также имеет определённое сходство с предварительным списком для пролива Немуро – индекс Чекановского–Сёренсена между ними составил 0.41, т.е. был несколько меньше, чем при сравнении с восточной частью Кунаширского пролива. В то же время между районом наших исследований и районом Дото индекс сходства был заметно выше и составил 0.56, что, видимо, обусловлено применением сходных орудий лова при сборе материала. В последнем районе при сравнении также выявлено 17 видов (при пяти неидентифицированных), которые отсутствуют в списке для пролива Немуро. Следовательно, проникновение в пролив Немуро (включая Кунаширский пролив) со стороны Тихого океана из района Дото вблизи м. Носаппу ряда новых, по сравнению с предварительным списком, видов вполне вероятно и ожидаемо в связи с отсутствием каких-либо географических препятствий.
Представляет интерес сравнение рассмотренных выше данных с опубликованными материалами по прибрежной ихтиофауне вод у Северного Хоккайдо, которые были собраны у г. Момбецу донными сетями на глубинах ≤70–80 м (Suda et al., 2003, 2004). Хотя в этом районе сбор проб проводили в основном на верхних участках шельфа, т.е. на глубинах существенно меньших, чем в восточной части Кунаширского пролива, тем не менее по числу видов в прибрежных водах Момбецу преобладали такие семейства, как Pleuronectidae (7), Cottidae (6), Stichaeidae (3), Hexagrammidae (3) и Agonidae (3). Очевидно, что и у Северного Хоккайдо доминирующими по числу видов оказались те же семейства, которые преобладали во всех трёх указанных выше районах (пролив Немуро, восточная часть Кунаширского пролива, район Дото), за исключением морских окуней и липаровых (рис. 4), предпочитающих в основном бо́льшие глубины. В соответствии с вертикальным распределением представителей последних двух семейств относят преимущественно к таким батиметрическим группировкам, как мезобентальная и верхнебатиальная, реже – элиторальная и сублиторальная (Линдберг, Красюкова, 1987; Снытко, 2001). При изучении видового состава рыб в каком-либо районе имеет значение также селективность используемых орудий лова. При добыче минтая и терпуга донными сетями в районе наблюдений применяли сетное полотно с различной величиной ячеи, учитывая высоту тела половозрелых рыб – 48 мм для минтая и 35 мм для терпуга. Размеры ячеи донных сетей, как и дрифтерных пелагических, конечно, оказывают определённое влияние на видовой состав уловов и видовое разнообразие рыб в пробах из этих орудий лова (Gulland, Harding, 1961; Савиных, 1998; Савиных и др., 2003; Yamamura, 2003). Примеры влияния размеров ячеи сетей на вылов тех или иных видов рыб можно видеть и на материалах многолетней выборки в восточной части Кунаширского пролива. В частности, обращает на себя внимание то, что в предварительных списках рыб пролива Немуро и района Дото семейство Agonidae входит в число наиболее широко представленных видами семейств, тогда как в наших пробах из восточной части Кунаширского пролива видов этой группы было отмечено минимальное число. Для представителей этого семейства в основном характерны небольшие длина (≤20 см) и высота тела (Линдберг, Красюкова, 1987; Борец, 2000). Возможно, по указанным выше причинам в большинстве своём они были недоступны для облова донными сетями с крупными размерами ячеи. Поэтому в нашем списке отмечено лишь три вида этой группы рыб – Podothecus sachi, P. sturioides и Tilesina gibbosa. Все они достигают довольно большой длины тела по сравнению с другими агонидами – соответственно 50, 29 и 36 см (Борец, 2000), а также и большей высоты тела. С другой стороны, более высокотелых морских окуней (Линдберг, Красюкова, 1987) регистрировали в наших пробах гораздо чаще. На наш взгляд, именно эти морфологические особенности в наибольшей мере способствовали тому, что в уловах донных сетей, применяемых в российской части Кунаширского пролива, рыбы семейства Scorpaenidae по числу видов (8) вышли на третье место.
ЗАКЛЮЧЕНИЕ
Таким образом, в рассматриваемом районе выявлено 85 видов рыб из 27 семейств. По числу видов доминировали семейства Pleuronectidae (18) и Cottidae (14). При этом 35 видов ранее не были отмечены в публикациях о видовом составе Кунаширского пролива и пролива Немуро в целом. Это свидетельствует о высоком видовом богатстве рыб в проливе, разделяющем южные Курильские о-ва и о. Хоккайдо, в котором число видов в настоящее время уже составило 173. Имеется немало оснований полагать, что приведённая цифра не является окончательной, а дальнейшие исследования позволят существенно увеличить этот показатель.