МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
2019 год, том 31, номер 4, стр. 131-144
МОДЕЛЬ ДИНАМИКИ ЧИСЛЕННОСТИ ПАУКООБРАЗНЫХ
В СПЕКТРЕ ИХ МЕЖВИДОВЫХ КОНКУРЕНТНЫХ ОТНОШЕНИЙ
2019 г. Э.Ф. Юсифов, А.А. Мамедов, Н.Э. Новрузов, В.С. Халилова
Институт зоологии НАН Азербайджана, Баку, Азербайджан
pralimamedov@rambler.ru
DOI: 10.1134/S0234087919040087
В статье рассмотрены вопросы построения и исследования математической модели
для изучения динамики численности паукообразных герпетобионтов в спектре их
трофических конкурентных отношений. Обсуждаются вопросы определения необ-
ходимых переменных и расчетных коэффициентов для построения и исследования
модели применительно к различным трофическим ситуациям. Базисом для созда-
ния модели послужили нелинейные дифференциальные уравнения Лотки-Вольтер-
ра. Исследования, проведенные с помощью построенной модели, показали, что ре-
акция системы на любое возмущение носит колебательный характер. Характер ре-
шения зависит от начального возмущения. Решения отличаются величиной ампли-
туды и периода колебаний. Установившиеся решения математической модели яв-
ляются многопериодичными колебаниями, которые характерны для биологических
систем. Приведены численные и графически представленные результаты исследо-
вания предложенной модели.
Ключевые слова: паукообразные, динамика численности, трофические ситуации,
конкурентные отношения, математическая модель.
THE MODEL OF THE DYNAMICS OF NUMBER OF ARACHNIDS
IN THE SPECTRUM
OF THEIR INTERSPECIES COMPETITIVE RELATIONSHIPS
E.F. Yusifov, A.A. Mamedov, N.E. Novruzov, V.S. Khalilova
Institute of Zoology of the National Academy of Sciences of Azerbaijan, Baku, Azerbaijan
The article deals with the construction and research of a mathematical model for studying
the dynamics of the number of arachnids herpetobionts in the spectrum of their trophic
competitive relations. The problems of determining the necessary variables and calcula-
tion coefficients for constructing and studying the model in relation to various trophic
situations are discussed. The basis for the model was the nonlinear differential equation
of Lotka-Volterra. The studies carried out with the aid of the constructed model have
shown that the response of the system to any perturbation is of an oscillatory nature. The
nature of the solutions depends on the initial perturbation, and differ in magnitude of the
132
Э.Ф. Юсифов, А.А. Мамедов, Н.Э. Новрузов, В.С. Халилова
amplitude and period of the oscillations. The steady-state solutions of the mathematical
model are multi-period oscillations, which are characteristic for biological systems. Nu-
merical and graphically presented data of the research results of the proposed model are
presented.
Key words: arachnids, number dynamics, trophic situations, competitive relations, mathe-
matical model.
1. Введение
Представители трёх отрядов класса паукообразных (Arachnida), такие
как - скорпионы (Scorpiones), сольпуги (Solifugae) и пауки (Aranei) - неотъ-
емлемые компоненты трофических цепей в семиаридных биоценозах. По-
лифагия и широкое распространение этих хищников позволяют предпола-
гать их важную роль в трофических отношениях с представителями других
классов хищных членистоногих (Chilopoda, Insecta) и в регуляции числен-
ности беспозвоночных фито- и сапрофагов. Межвидовые отношения этих
паукообразных между собой и другими хищными членистоногими герпето-
бионтами, объединёнными общей территорией, динамика их численности в
зависимости от изменения численности жертв и потенциальных конкурен-
тов - пока еще мало исследованная область синэкологии.
Существует также много нерешенных вопросов, связанных с особен-
ностями динамики численности и структуры популяций этих хищников, с
установлением механизмов их регуляции и путей адекватного оптимально-
го управления ими.
Своеобразная уникальность и сложность структуры природных экоси-
стем не всегда позволяет проводить полевые и стационарные исследования
в необходимом объёме. Существует вероятность и того, что последствия от
проведения таких исследований могут в целом негативно сказаться на их
экологическом балансе. Главной же трудностью, с которой обычно сталки-
ваются исследователи при изучении природных экосистем, является необ-
ходимость учета большого количества биотических и абиотических факто-
ров. Сложные взаимоотношения между самими живыми объектами также
затрудняют сбор первичных данных и их дальнейший анализ в рамках клас-
сических полевых, стационарных и камеральных методов исследования.
Кроме того, получение количественных экологических показателей тради-
ционными средствами всегда отнимает слишком много времени, в течение
которого сама исследуемая экосистема может претерпевать определенные
качественные и количественные изменения. Исходя из всего выше отме-
ченного, становится очевидным, что при осуществлении глубокого анализа
и систематизации количественных данных о тех или иных компонентах
Модель динамики численности паукообразных в спектре ...
133
биоценоза требуется привлечение других, более оперативных методов про-
ведения исследований. Среди такого рода средств наиболее эффективным
является математическое моделирование [1].
Изучение трофических связей паукообразных герпетобионтов, постро-
ение на основе полученных фактических данных математической модели
позволит дать еще более объективную оценку их роли в биоценозах, с боль-
шей эффективностью проводить экологический мониторинг природных тер-
риторий, прогнозировать их изменение в будущем. Построение математи-
ческих моделей, описывающих динамику численности популяций, исследо-
вание их динамических режимов является одним из необходимых условий
для решения важных прикладных задач оптимального управления популя-
циями этих животных [2].
В большинстве работ, посвященных математическому моделированию
динамики численности популяций животных, для исследования изменения
их плотности, как правило, рассматриваются основные демографические па-
раметры (рождаемость, смертность, миграция и др.) [3-16]. При этом тро-
фический фактор обычно или совсем не учитывается или рассматривается
только для каждого из видов в отдельности, а не в спектре межвидовой
конкуренции с разной интенсивностью за одни и те же пищевые ресурсы.
Однако, как известно, в природных популяциях такие конкурентные ситуа-
ции наблюдаются достаточно часто [17, 18] и, безусловно, требуют подроб-
ного натурного и модельного исследования.
Разработке и исследованию математической модели, получению отве-
тов на некоторые из перечисленных выше вопросов и посвящено данное ис-
следование.
Объектами для изучения были выбраны три крупных представителя
класса паукообразных герпетобионтов: пестрый скорпион Mesobuthus eu-
peus (C.L.Koch, 1839), обыкновенная сольпуга Galeodes araneoides (Pallas,
1772), паук-тарантул Lycosa praegrandis (C.L.Koch, 1836), исходя из их ши-
рокого распространения, совместного обитания на семиаридных территори-
ях, схожести сезонного и суточного ритма активности, а также возможности
использования для их изучения идентичных, широко используемых в поле-
вых условиях методов сбора материала.
2. Материал и методы сбора данных
Сбор фактического материала для получения базовых данных прово-
дился в полевых и стационарных условиях на территории междуречья Пир-
сагат-Джейранкечмез (Гобустан, восточная часть Азербайджана). Для дос-
тижения поставленной цели решался ряд следующих задач:
1) установление относительной численности каждого из трех видов па-
134
Э.Ф. Юсифов, А.А. Мамедов, Н.Э. Новрузов, В.С. Халилова
укообразных традиционными методами [19,20];
2) изучение суточной активности исследуемых видов путем фиксации
активных особей в разные временные интервалы;
3) изучение их спектра питания путем сбора остатков пищи в постоян-
ных укрытиях и визуальной фиксации жертв на хелицерах хищников;
4) изучение состава фауны основных групп беспозвоночных в биотопе
с использованием классических методов применяемых в энтомологии [21];
5) выяснение основных трофических связей паукообразных;
6) определение необходимых переменных для создания модели;
7) расчет коэффициентов для построения модели;
8) построение математической модели типа "хищник-жертва";
9) проверка работоспособности модели путем решения ряда тактиче-
ских задач.
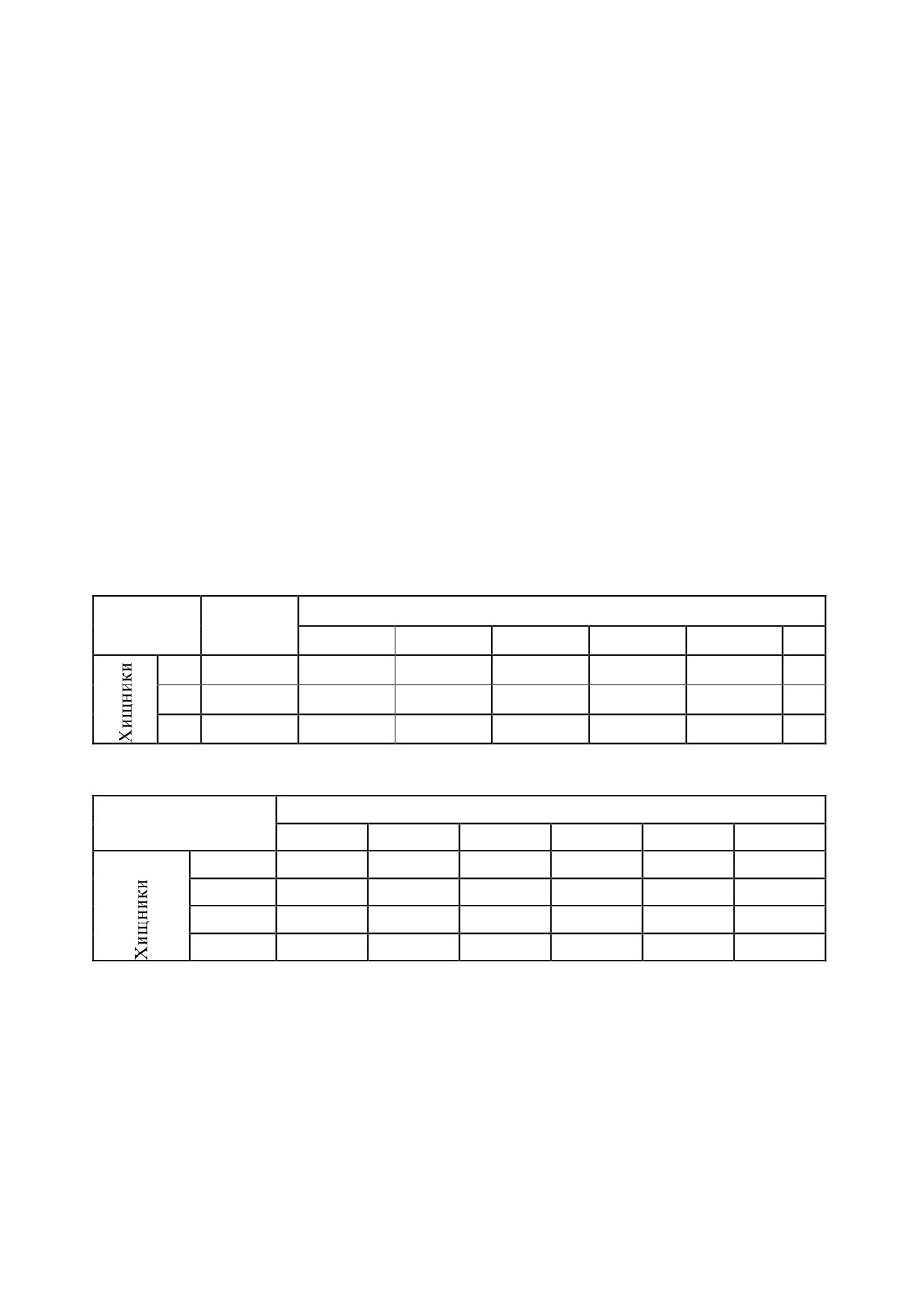
В представленной ниже таблице приведены основные характеристики
объектов: стратегии охоты хищников и способы передвижения их жертв
(табл.1).
Таблица 1. Краткая характеристика объектов рассматриваемого сообщества.
ЖЕРТВЫ
ХИЩНИКИ
СПОСОБ ОХОТЫ
СПОСОБ ПЕРЕДВИЖЕНИЯ
Неподвижные
Ожидание в засаде
Ползающие
в укрытии
Ходящие
СКОРПИОН
Ожидание в засаде
Ползающие
на поверхности
Ходящие
Ходящие
Активный поиск
Бегающие
Летающие
Активный поиск
Прыгающие
Ползающие
СОЛЬПУГА
Преследование
Бегающие
Ходящие
Нападение
Бегающие
Нападение
Ползающие
Ожидание в засаде
Ходящие
ПАУК-ТАРАНТУЛ
в укрытии
Бегающие
Бегающие
Активный поиск
Прыгающие
Примечание. Как бы дополняя табличные данные, следует указать, что
в процессе охоты хищники могут использовать все охотничьи стратегии. Но
Модель динамики численности паукообразных в спектре ...
135
при построении модели мы намеренно упростили ситуацию, рассмотрев на-
иболее характерные стратегии для каждого из хищников. Разделение по спо-
собу передвижения также условно, оно характеризует основные типы жертв.
По совокупности данных полевых и стационарных наблюдений соот-
ношение жертв в питании хищников варьировало, находясь в определенной
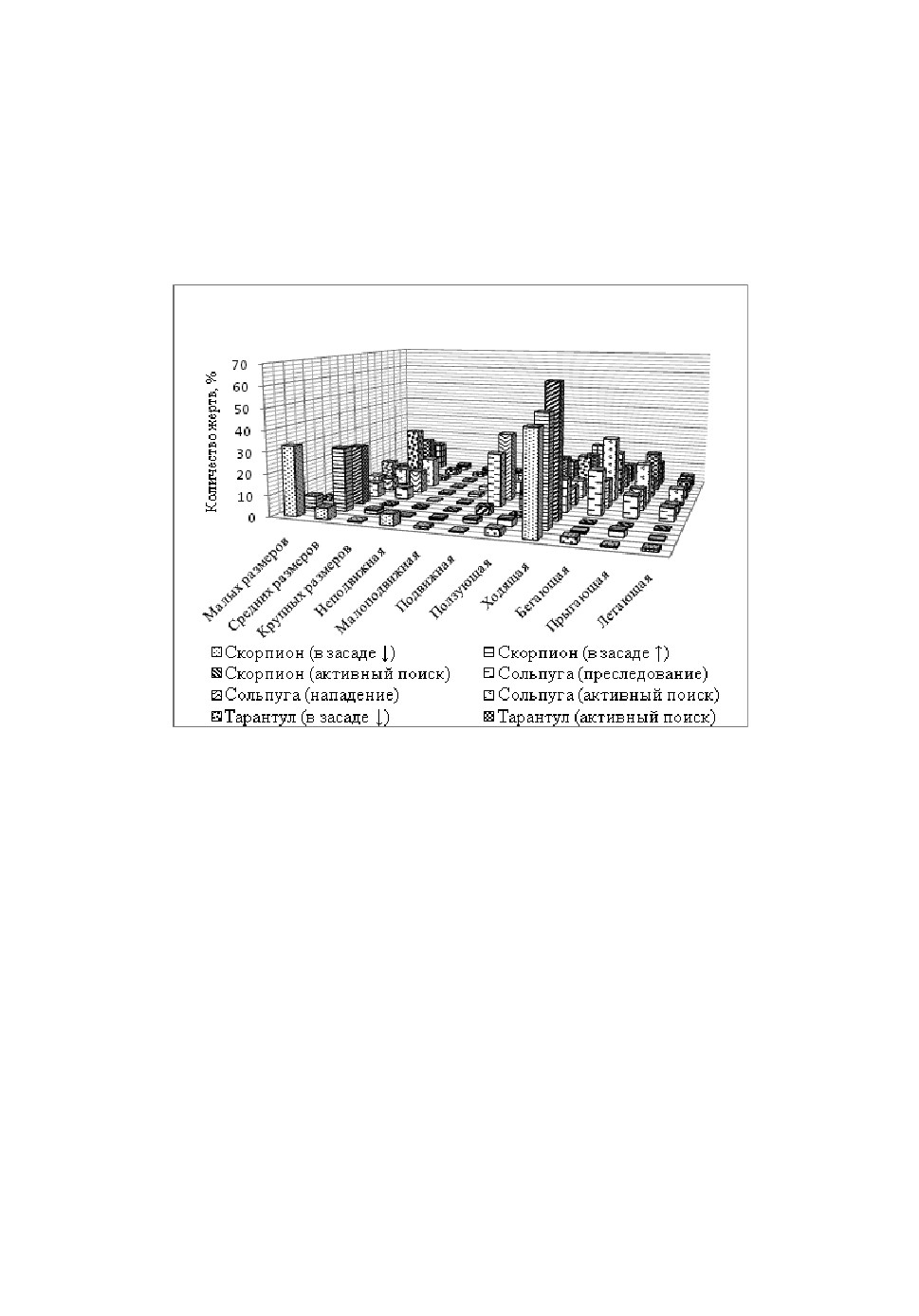
зависимости от используемой ими охотничьей стратегии (рис. 1).
Рис.1. Соотношение жертв в составе добычи паукообразных при разных охотничьих
стратегиях.
3. Построение математической модели
Цель создания математической модели заключалась в определении вли-
яния конкурентного питания на численность трех видов паукообразных.
При создании математической модели мы исходили из следующих предпо-
ложений:
1) у каждого из хищников имеется потребность в объектах питания c
разным способом передвижения;
2) хищники используют одну стратегию охоты;
3) все процессы - рождаемость, смертность - обладают интенсивно-
стью, пропорциональной численности объектов;
4) вследствие напряженных пищевых взаимоотношений внутри систе-
мы, предполагается, что у хищников присутствуют качественные предпоч-
тения в выборе пищи;
5) особенности пространственного распределения особей не учитыва-
ются;
136
Э.Ф. Юсифов, А.А. Мамедов, Н.Э. Новрузов, В.С. Халилова
6) модельное время условно равно одному месяцу.
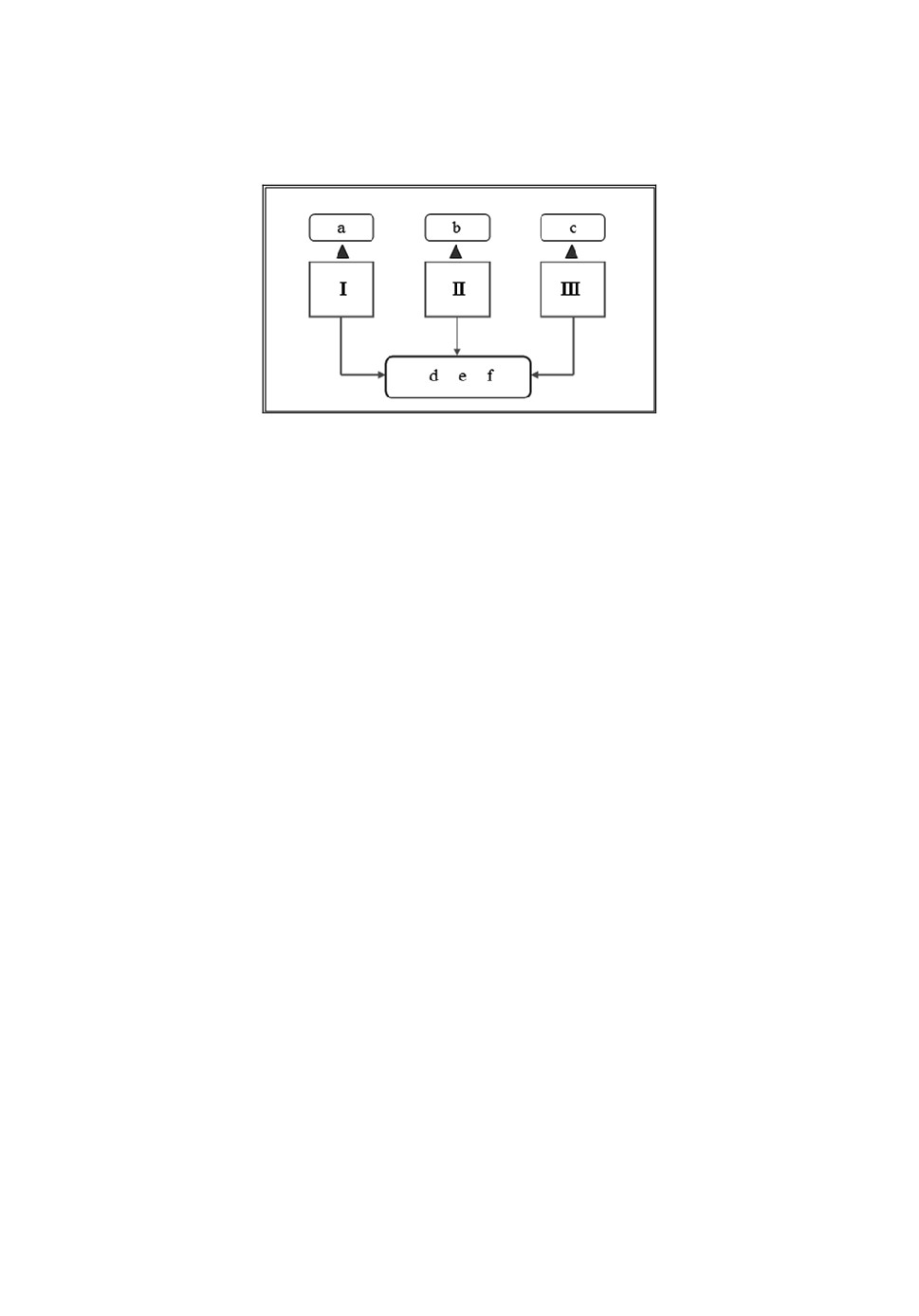
Приведем принципиальную блок-схему математической модели иссле-
дуемого сообщества (рис. 2).
Рис.2. Блок-схема математической модели двухуровневых трофических связей паукооб-
разных (хищники: I - скорпион (x1), II - сольпуга (x2), III - паук-тарантул (x3);
типы жертв по форме передвижения: a - неподвижные (y1), b - летающие (y2),
c - прыгающие (y3), d - ползающие (y4), e - ходящие (y5), f - бегающие (y6).
Как видно из представленной на рис.2 блок-схемы, рассматриваемое со-
общество, подобно любой биологической системе, обладает характерными
особенностями модели "хищник-жертва" и может быть описано уравнения-
ми Лотки [22] и Вольтерра [23]. Такие уравнения, как известно, можно ис-
пользовать для моделирования систем "хищник-жертва", "паразит-хозяин",
конкуренции и других видов взаимодействия между видами [17]. Таким об-
разом, мы располагаем 3 уравнениями, описывающими изменение численно-
сти хищников, и 6 уравнениями, описывающими изменение численности
жертв.
4. Описание модели
Рассматривается ограниченная по площади, обособленная по структуре
ландшафта территория, на которой обитают две группы моделируемых объ-
ектов - "хищники" и "жертвы". При условии отсутствия объектов охоты
хищники не питаются и обречены на гибель, следовательно, уравнение для
численности хищников (без учета численности жертв) принимает вид
1
2
3
dy
/
dt k
y ,
dy
/ dt k
y
,
dy
/ dt k
y
,
(1)
1
0 1
2
0
2
3
0
3
i
где
k
- коэффициент убыли i-го хищника, yi - величина популяции хищ-
0
ников,
dy
/
dt - скорость прироста популяции хищников.
i
Предполагается, что еда для жертв имеется в избытке. Тогда уравнение
изменения количества жертв (без учета хищников) принимает вид
dx
/
dt A
x ,
dx
/
dt A
x
,
dx
/
dt A
x ,
1
1 1
2
2
2
3
3 3
Модель динамики численности паукообразных в спектре ...
137
dx
/
dt A
x
,
dx
/
dt A
x
,
dx
/
dt A
x
,
(2)
4
4
4
5
5
5
6
6
6
где Aj - скорость размножения популяции j-й жертвы, xi - величина попу-
ляции j-й жертвы,
/
dx
dt - скорость прироста популяции j-й жертвы.
j
При встречах хищников и жертв, частота которых прямо пропорцио-
нальна величине
i
k
yx
,
(i=1÷3), (j=1÷6),
(3)
j i j
происходит поедание жертв с коэффициентомkij , сытые хищники способ-
ны к воспроизводству
i i
k
yx
,
(i=1÷3), (j=1÷6),
(4)
j j i j
- коэффициент переработки i-м хищником биомассы j-й жертвы в
собственную биомассу [24].
С учетом изложенных выше механизмов (1)-(4) и конкурентных жертв
система уравнений модели в общем виде будет выглядеть следующим обра-
зом:
1 1
1 1
1 1
1 1
1
dy
/ dt (k
x k
x
k
x k
x
k
)y
,
1
1 11
2
2
2
3 3 3
4
4
4
0
1
2
2
2
2
2
2
2
2
2
dy
/ dt (k
x
k
x
k
x
k
x
k
)y
,
2
2
2
2
3
3
3
4
4
4
6
6
6
0
2
3 3
3 3
3 3
3 3
3
dy
/ dt (k
x
k
x k
x
k
x k
)y
,
3
2
2
2
3
3 3
4
4
4
5
5 5
0
3
1
1
2
3
dx
/ dt (A
k
y
)x
,
dx
/ dt (A
k
y
k
y
k
y
)x
,
1
1
1
1
1
2
2
2
1
2
2
2
3
2
1
2
3
1
2
3
dx
/ dt (A
k
y k
y
k
y
)x
,
dx
/dt(A
k
y
k
y
k
y
)x
,
3
3
3 1
3
2
3
3
3
4
4
4
1
4
2
4
3
4
3
2
dx
/ dt (A
k
y
)x
,
dx
/ dt (A
k
y
)x
5
5
5
3
5
6
6
6
2
6
Базисом для подбора коэффициентов, параметров модели и начальных
данных переменных стали ранее проведенные полевые, стационарные и ла-
1
2
3
бораторные исследования. Значение коэффициентов
k
, k
,k
, а также па-
0
0
0
раметров A1, A2, A3, A4, A5, A6 вычислялось из условия устойчивости реше-
ния модели. Таким образом, мы также могли убрать лишние возмущения,
возможные при определении других коэффициентов и параметров модели.
Формулы для вычисления выделенных коэффициентов и параметров будут
выглядеть следующим образом:
138
Э.Ф. Юсифов, А.А. Мамедов, Н.Э. Новрузов, В.С. Халилова
1
1 1
1 1
1 1
1 1
к
k
x k
x
k
x k
x
0
1 11
2
2
2
3 3 3
4
4
4,
2
2
2
2
2
2
2
2
2
k
k
x
k
x
k
x
k
x
,
0
2
2
2
3
3
3
4
4
4
6
6
6
3
3 3
3
3
3
3
3 3
k
k
x
k
x k
x
k
x
,
0
2
2
2
3
3 3
4
4
4
5
5
5
1
1
2
3
1
2
3
A
k
y
,
A
k
y
k
y
k
y
,
A
k
y k
y
k
y
,
1
1
1
2
2
1
2
2
2
3
3
3 1
3
2
3
3
1
2
3
3
2
A
k
y
k
y
k
y
,
A
k
y
,
A
k
y
4
4
1
4
2
4
3
5
5
3
6
6
2
При определении численных значений коэффициентов математической мо-
дели также учитывалось условие их положительности.
Надо отметить, что в созданной модели количество хищников (i=1÷3) и
количество жертв (j=1÷6) было выбрано из числа характерных хищников и
жертв, присущих рассматриваемому биоценозу.
i
Значения коэффициентов
k
,
A
и параметровij приведены в табл.2,3.
j
j
Таблица 2. Значение коэффициентовkij .
Жертвы
0
1
2
3
4
5
6
1
9.34
0.012
0.043
0.029
0.017
0
2
27.75
0
0.065
0.074
0.077
0
3
13.23
0
0.038
0.036
0.034
0.007
0
i
Таблица 3. Значение параметров
,
A
j
j
Жертвы
1
2
3
4
5
6
1
0.87
0.87
0.87
0.87
0
0
2
0
0.82
0.82
0.82
0
0.82
3
0
0.85
0.85
0.85
0.85
0
А =
0.552
3.415
2.832
1.275
0.133
0.099
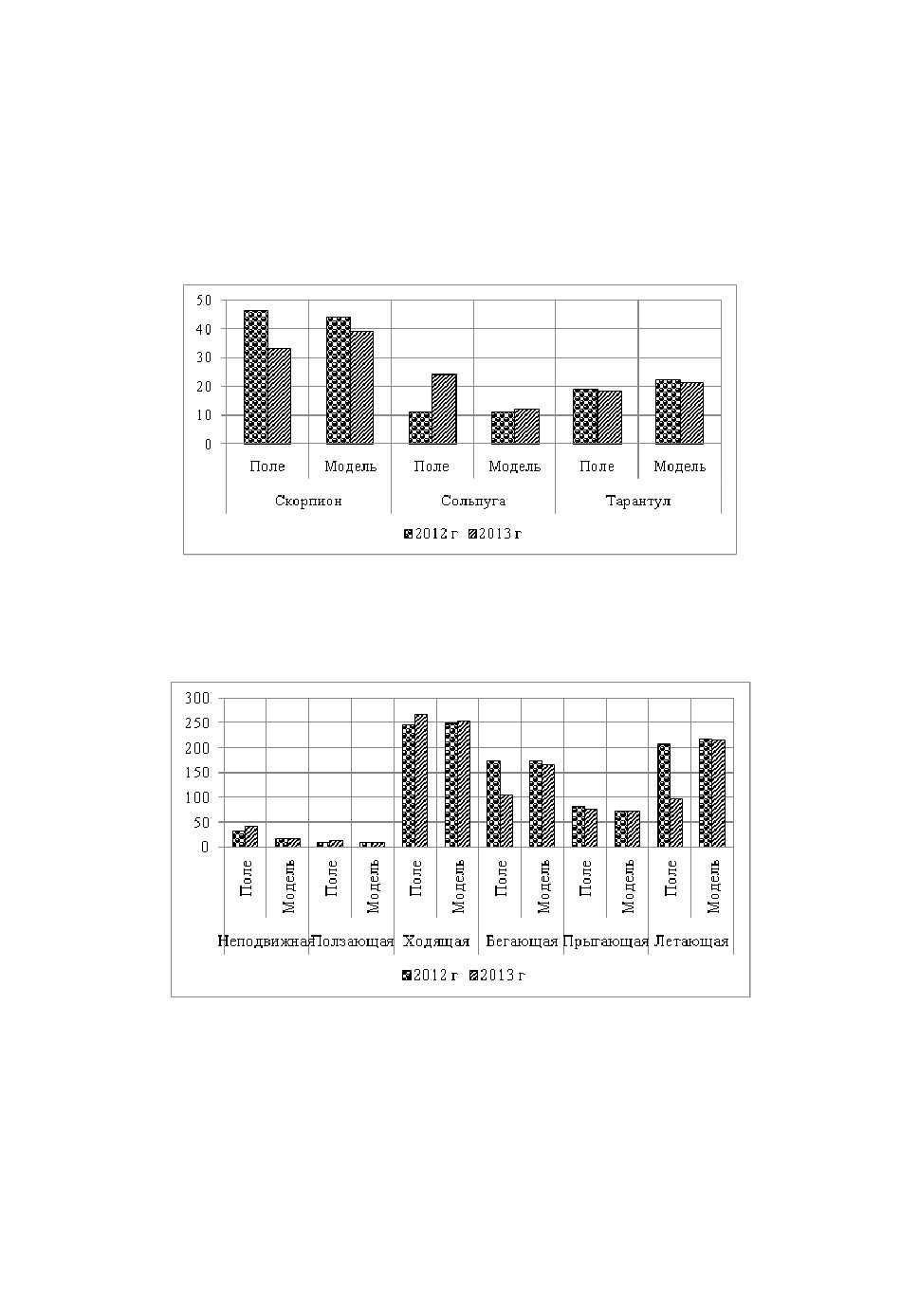
Для показания качественной адекватности математической модели бы-
ли взяты результаты полевых исследований, проведенных в 2012-2013 гг. на
территории междуречья Пирсагат-Джейранкечмез (Гобустан, восточная
часть Азербайджана).
Задав начальные данные для уравнений математической модели, соот-
ветствующие результатам 2012 года, мы естественно получаем стабильное
решение модели. Поэтому задаем возмущение системе путем изменения на-
чального значения одного из переменных. Интегрируем систему до момен-
Модель динамики численности паукообразных в спектре ...
139
та установления колебательного решения и сравниваем решения двух по-
следующих годов с результатами полевых исследований.
Качественное совпадение по динамике двух последующих лет встреча-
лось несколько раз в независимости от условий возмущения системы, т.е.
изменениям начальных данных для разных переменных.
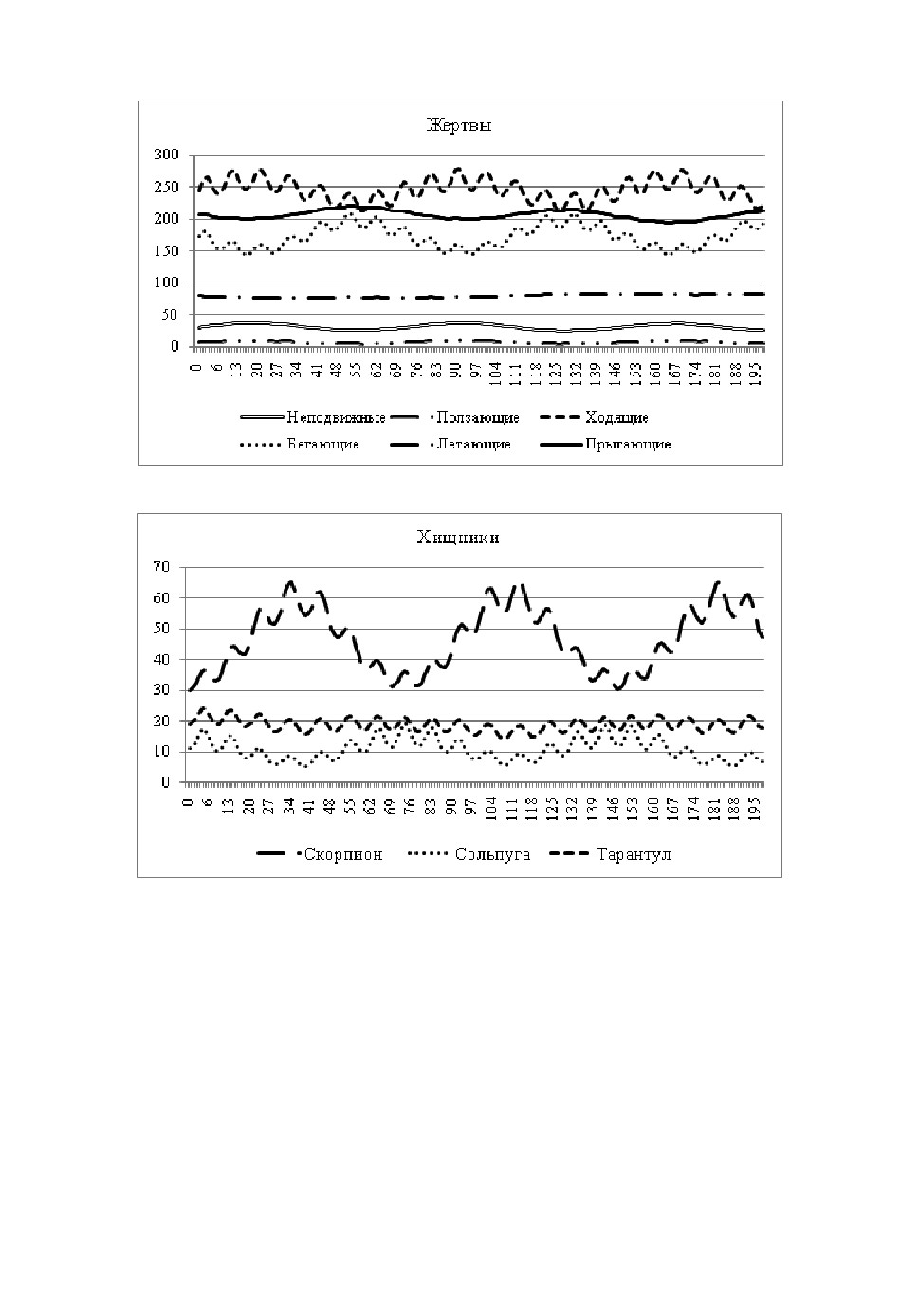
Результаты исследований приведены на рис.3,4.
Рис.3. Численность хищников.
По результатам, приведённым в диаграммах (рис.2,3), можно утверж-
дать, что математическая модель качественно адекватна её биологическому
прообразу.
Рис.4. Численность жертв.
5. Проведение экспериментов с моделью
Для эффективного проведения вычислительных экспериментов на ма-
тематической модели нами была разработана специальная программа с по-
мощью редактора DELPHI-7, которая реализует численное решение данной
модели методом Рунге-Кутты 4-го порядка с погрешностью решения не бо-
лее 0.00001 [25] и создаёт интерфейс для обозрения полученных результатов.
140
Э.Ф. Юсифов, А.А. Мамедов, Н.Э. Новрузов, В.С. Халилова
Эксперименты на математической модели проводились с целью опре-
деления динамики численности каждого из объектов сообщества членисто-
ногих. Для проведения исследований математическая модель запускалась с
различными начальными данными (а - начальное значение плюс 30%, б -
начальное значение минус 30%) переменных, причем каждый раз для от-
дельно взятой переменной. Все полученные решения носили колебательный
характер. Для численного сравнения полученных результатов по каждой из
переменных мы вычислили их среднее арифметическое за время t. Отме-
тим, что при длительном интегрировании среднее арифметическое для каж-
дой переменной восстанавливалось на уровне начальных данных до возму-
щения.
Значение возмущений и процентное соотношение между полученным
результатом и начальным значением для каждого элемента сообщества чле-
нистоногих приведены в табл.4.
Важно отметить, что все решения модели после возмущения являются
колебательными. Характерное поведение динамического решения приведе-
но на рис.5,6.
Таблица 4. Соотношение между полученным результатом и начальным значением
для хищников и жертв (%).
Хищники
Жертвы
Y1=60
17.39
-9.09
-5.26
-6.45
-14.29
-2.86
4.02
1.25
0.97
Y1=30
-26.09
18.18
10.53
9.68
14.29
4.08
-4.6
-1.25
-1.45
Y2=15
-4.35
18.18
-5.26
3.23
0
0.41
-1.72
0
-0.97
Y2=7
4.35
-18.18
10.53
-3.23
0
-0.82
2.3
-1.25
0.97
Y3=25
-4.35
-9.09
26.32
0
0
0
-0.57
-1.25
0.48
Y3=13
4.35
9.09
-26.32
-3.23
0
0.41
0.57
2.5
-0.97
X1=41
4.35
0
0
32.26
0
-0.82
0.57
0
0
X1=21
-4.35
0
0
-32.26
0
0.41
-0.57
0
0
X2=9
2.17
0
0
0
28.57
-0.41
0
0
0
X2=5
-2.17
0
0
0
-28.57
0.41
0
0
0
X3=320
10.87
0
0
-3.23
-14.29
11.43
-11.49
0
-0.48
X3=170
-17.39
9.09
-5.26
6.45
28.57
-15.1
14.94
1.25
0
X4=122
15.22
-27.27
-5.26
-3.23
0
10.2
-20.69
1.25
1.45
X5=104
-2.17
-9.09
15.79
0
0
0
-0.57
27.5
0.48
X5=56
2.17
9.09
-10.53
0
0
0
0.57
-28.75
-0.48
X6=269
-6.52
18.18
-15.79
3.23
14.29
0.41
-4.02
1.25
27.54
X6=145
6.52
-18.18
15.79
-3.23
0
-0.82
3.45
-1.25
-28.5
Модель динамики численности паукообразных в спектре ...
141
Рис.5. Характерная динамика изменения численности жертв.
Рис.6. Характерная динамика изменения численности хищников.
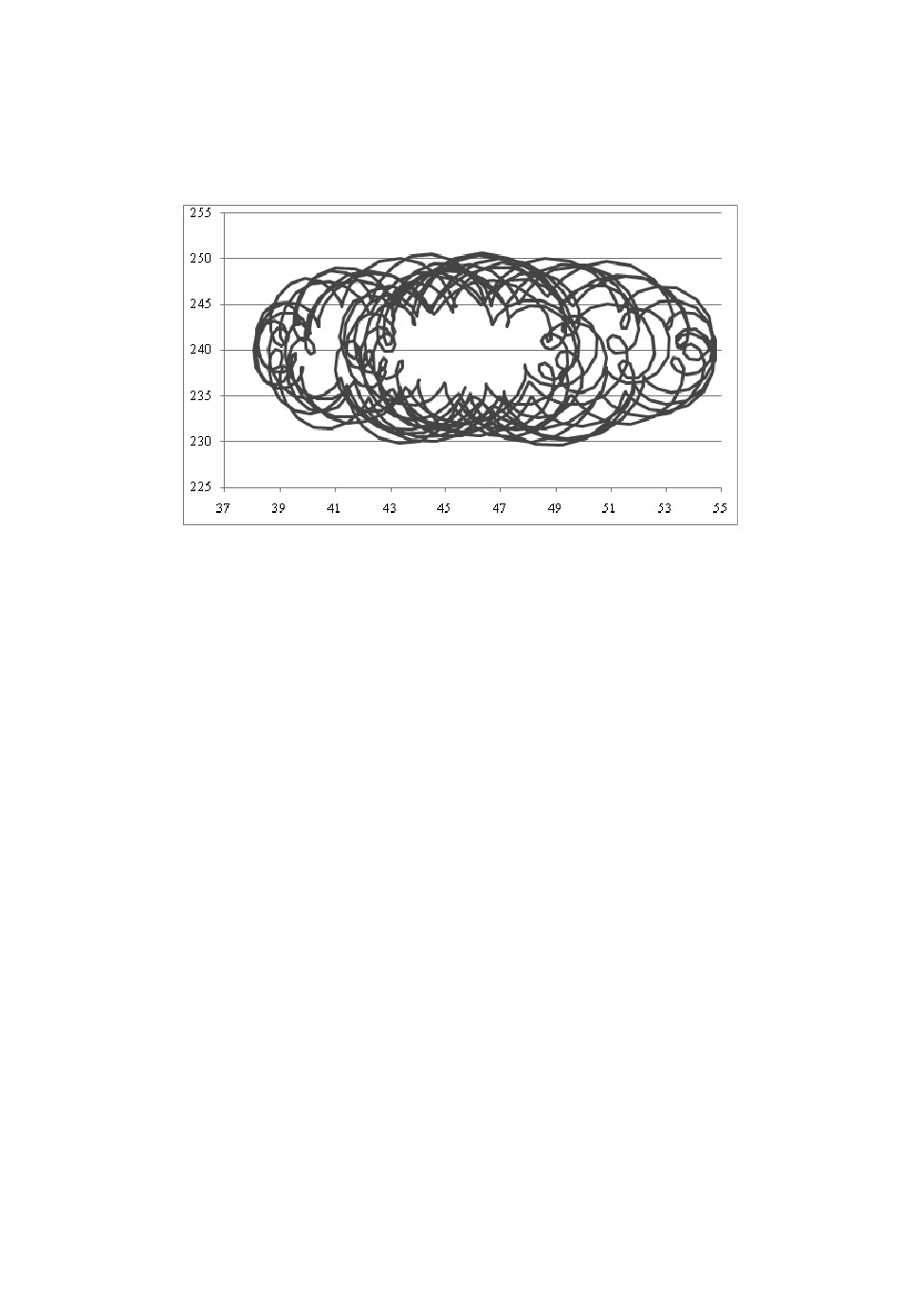
Установившиеся решения математической модели представляли собой
аттракторы (рис.7).
6. Заключение
В статье рассмотрены вопросы построения и исследования математи-
ческой модели для описания динамики численности паукообразных при воз-
никновении межвидовых конкурентных отношений. Обсуждались вопросы
определения переменных и коэффициентов модели. Коэффициенты опреде-
лялись путем выделения характеристик, присущих модельным объектам. В
качестве базовой модели использовалась система "хищник-жертва". Иссле-
дования, проведенные с помощью построенной модели, показали, что реак-
142
Э.Ф. Юсифов, А.А. Мамедов, Н.Э. Новрузов, В.С. Халилова
ция системы на любое возмущение носит колебательный характер. Харак-
тер решений зависит от начального возмущения и отличается величиной
амплитуды и периода колебаний.
Рис.7. Характерный двухмерный график установившегося движения.
Установившиеся решения математической модели представляют собой
аттракторы, которые характерны для биологических систем. Приведенные
численные и графические результаты, полученные с использованием пред-
ложенной модели, убедительно демонстрируют, что любое изменение на-
чальных данных оказывает влияние на динамику численности паукообраз-
ных. Предложенная модель может использоваться для прогнозирования
численности герпетобионтных паукообразных в отдельно взятых семиарид-
ных биотопах. Использование данной модели в перспективе можно сущест-
венно расширить, привлекая большее количество переменных и соответст-
вующих коэффициентов.
СПИСОК ЛИТЕРАТУРЫ
1. Д.М. Смит. Модели в экологии. - М.: Мир, 1976, 184 с.;
D.M. Smith. Models in ecology. - Cambrige University Press, 1974.
2. А.А. Шаров. Моделирование динамики численности популяций насекомых // Итоги
науки и техники. Серия "Энтомология". - М.: ВИНИТИ, 1986, т.6, с.3-115;
A.A. Sharov. Modelirovanie dinamiki chislennosti populiatsii nasekomykh // Itogi nauki i
tekhniki. Seriia "Entomologiia". - M.: VINITI, 1986, t.6, s.3-115.
3. А.Н. Фролов. Динамика численности и прогноз массовых размножений вредных на-
секомых: исторический экскурс и пути развития. Аналитический обзор / Вестник
защиты растений 4(94), 2017, с.5-21;
A.N. Frolov. Dinamika chislennosti i prognoz massovykh razmnozhenii vrednykh naseko-
Модель динамики численности паукообразных в спектре ...
143
mykh: istoricheskii ekskurs i puti razvitiia. Analiticheskii obzor / Vestnik zashchity rastenii
4(94), 2017, s.5-21.
4.
А.П. Шапиро. Математические модели конкуренции // Управление и информация.
Владивосток: ДВНЦ АН СССР, 1974, вып.10, с.5-75;
A.P. Shapiro. Matematicheskie modeli konkurentsii // Upravlenie i informatsiia. Vladi-
vostok: DVNTs AN SSSR, 1974, vyp.10, s.5-75.
5.
Ю.М. Свирижев, Е.Я. Елизаров. Математическое моделирование биологических со-
обществ. - М.: Наука, 1972, 150 с.;
Yu.M. Svirizhev, E.Ya. Elizarov. Matematicheskoe modelirovanie biologicheskikh soob-
shchestv. - M.: Nauka, 1972, 150 s.
6.
Г.Ю. Ризниченко. Лекции по математическим моделям в биологии. - М.: Ижевск:
Институт компьютерных исследований, НИЦ "Регулярная и хаотическая динамика",
2010, 560 с.;
G.Yu. Riznichenko. Lektsii po matematicheskim modeliam v biologii. - M.: Izhevsk: Insti-
tut kompiuternykh issledovanii, NITs "Reguliarnaia i khaoticheskaia dinamika", 2010, 560 s.
7.
Ю.С. Малышев. К вопросу об индикации конкурентных отношений между система-
тически и экологически близкими видами животных / Материалы II Международной
конференции Современные проблемы биологической эволюции, 2014, с.345-349;
Yu.S. Malyshev. K voprosu ob indikatsii konkurentnykh otnoshenii mezhdu sistematicheski
i ekologicheski blizkimi vidami zhivotnykh / Materialy II Mezhdunarodnoi konferentsii
Sovremennye problemy biologicheskoi evoliutsii, 2014, s.345-349.
8.
И.К. Яковлев, Ж.И. Резникова. Межвидовые взаимодействия как арена «быстрой эво-
люции»: модели и перспективы исследований / Материалы II Международной конфе-
ренции Современные проблемы биологической эволюции. - М.: 2014, с.54-56;
I.K. Yakovlev, Zh.I. Reznikova. Mezhvidovye vzaimodeistviia kak arena «bystroi evoliut-
sii»: modeli i perspektivy issledovanii / Materialy II Mezhdunarodnoi konferentsii Sovre-
mennye problemy biologicheskoi evoliutsii. - M.: 2014, s.54-56.
9.
G. Dai, M. Tang. Coexistence Region and Global Dynamics of a Harvesting Predator -
Prey Systems // SIAM J. Appl. Math., 1998, v.58, №1, p.193-210.
10. Yu.V. Utyupin, L.V. Nedorezov. About a continuous-discrete model of Predator-Prey sys-
tem dynamics // Population Dynamics: Analysis, Modelling, 2014, Forecast 3(2), р 55-63.
11. Ю.М. Апонин, Е.А. Апонина. Математическая модель сообщества хищник - жертва с
нижним порогом численности жертвы // Компьютерные исследования и моделиро-
вание, 2009, т.1, №1, с.51-56;
Yu.M. Aponin, E.A. Aponina. Matematicheskaia model soobshchestva khishchnik - zhertva
s nizhnim porogom chislennosti zhertvy // Kompiuternye issledovaniia i modelirovanie,
2009, t.1, №1, s.51-56.
12. D.D. Murray. Mathematical biology. - New York: Springer, 2002, 551 p.
13. Н.А. Гасратова, М.В. Столбовая, Е.Г. Неверова, А.С. Бербер. Математическая мо-
дель «ресурс-потребитель» // Молодой ученый, 2014, №10, c.5-13;
N.A. Gasratova, M.V. Stolbovaya, E.G. Neverova, A.S. Berber. Matematicheskaia model
«resurs-potrebitel» // Molodoi uchenyi, 2014, №10, c.5-13.
14. А. Братусь, А. Новожилов, А. Платонов. Динамические системы и модели в биоло-
гии // Litres, 2017, 400 с.;
144
Э.Ф. Юсифов, А.А. Мамедов, Н.Э. Новрузов, В.С. Халилова
A. Bratus, A. Novozhilov, A. Platonov. Dinamicheskie sistemy i modeli v biologii // Litres,
2017, 400 s.
15. А.С. Исаев, Е.Н. Пальникова, В.Г. Суховольский, О.В. Тарасова. Динамика численно-
сти лесных насекомых-филлофагов: модели и прогнозы. - М.: Товарищество научн.
изд. КМК, 2015, 262 с.;
A.S. Isaev, E.N. Palnikova, V.G. Sukhovolsky, O.V. Tarasova. Dinamika chislennosti les-
nykh nasekomykh-fillofagov: modeli i prognozy. - M.: Tovarishchestvo nauchn. izd. KMK,
2015, 262 s.
16. Е.Я. Фрисман, Е.И. Скалецкая. Странные аттракторы в простейших моделях дина-
мики численности биологических популяций // Обозрение прикладной и промыш-
ленной математики, 1994, т.1, №6, с.988-1008;
E.Ya. Frisman, E.I. Skaletsky. Strannye attraktory v prosteishikh modeliakh dinamiki chis-
lennosti biologicheskikh populiatsii // Obozrenie prikladnoi i promyshlennoi matematiki,
1994, t.1, №6, s.988-1008.
17. Ю. Одум. Основы экологии. - М.: Мир, 1975, 740 с.;
J. Odum. Fundamentals of ecology. - Philadelfia: 1953, 384 p.
18. Э. Пианка. Эволюционная экология. - М.: Мир, 1981. 400 с.;
E. Pianca. Evolutionary ecology. - New York: Harper and Row, 1978.
19. М.С. Гиляров. Методы почвенно-зоологических исследований. - М.: Наука, 1975,
280 с.;
M.S. Gilyarov. Metody pochvenno-zoologicheskikh issledovanii. - M.: Nauka, 1975, 280 s.
20. Ю.А. Песенко. Принципы и методы количественного анализа в фаунистических ис-
следованиях. - М.: Наука, 1982, 288 с.;
Yu.A. Pesenko. Printsipy i metody kolichestvennogo analiza v faunisticheskikh issledova-
niiakh. - M.: Nauka, 1982, 288 s.
21. К.К. Фасулати. Полевое изучение наземных беспозвоночных. - М.: Высшая школа,
1971, 424 с.;
K.K. Fasoulati. Polevoe izuchenie nazemnykh bespozvonochnykh. - M.: Vysshaia shkola,
1971, 424 s.
22. A.J. Lotka. Elements of physical biology. - Baltimore: Williams and Wilkins, 1925, 495 p.;
23. В. Вольтерра. Математическая теория борьбы за существование. - М.: Наука, 1976,
288 с.;
V.Volterra. Lecóns sur la théorie mathematique de la lutte pour la vie. P.: Gauthiers-Vil-
lars, 1931.
24. А.Д. Базыкин. Математическая биофизика взаимодействующих популяций. - М.:
Наука, 1985, 181 с.;
A.D. Bazykin. Matematicheskaia biofizika vzaimodeistvuiushchikh populiatsii. - M.: Nau-
ka, 1985, 181 s.
25. Н.С. Бахвалов, Н.П. Жидков, Г.М. Кобельков. Численные методы. - М.: Лаборатория
Базовых Знаний, 2001. 630 с.
N.S. Bakhvalov, N.P. Zhidkov, G.M. Kobelkov. Chislennye metody. - M.: Laboratoriia Ba-
zovykh Znanii, 2001. 630 s.
Поступила в редакцию 07.06.18
После доработки 07.06.18
Принята к публикации 10.09.18